Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
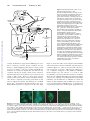
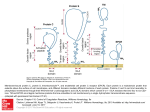
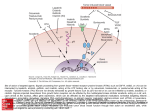
See related article, pages 255–262 Endothelial Cell Protein C Receptor Role Beyond Endothelium? Meenakshisundaram Thiyagarajan, Tong Cheng, Berislav V. Zlokovic E Downloaded from http://circres.ahajournals.org/ by guest on August 3, 2017 ndothelial cell protein C receptor (EPCR) is a type 1 transmembrane glycoprotein with homology to the major histocompatibility complex (MHC)-class 1/CD1 family of molecules.1 EPCR was isolated and cloned as an endothelial cell-specific, high selectivity and high affinity binding protein for protein C (PC) and activated PC (APC).1 EPCR binds PC on the endothelial surface and presents it to the thrombin:thrombomodulin (TM) complex for activation (Figure 1). Thrombin bound to TM proteolytically activates PC and generates APC which exerts independent anticoagulant and cellular activities.2 The APC anticoagulant pathway is mediated by proteolytic inactivation of blood coagulation Factors Va and VIIIa with the contribution of several cofactors, including protein S, high density lipoproteins, phosphatidylserine, cardiolipin, and glucosylceramide to name a few. The APC cellular pathway on endothelium requires EPCR and the protease activated receptor-1 (PAR-1)3 which mediates APC’s cytoprotective effects, including alterations in gene expression profiles, antiapoptotic activity, antiinflammatory activity and protection of endothelial barriers4 –10 (Figure 1). EPCR also regulates endogenous physiologic activation of PC by thrombin which is linked to PAR-1-dependent APC protective autocrine signaling in endothelium.11 EPCR signaling can decrease inflammation. APC binding to EPCR rescues baboons from E. coli sepsis.12 EPCR has also cardioprotective role in lipopolyscharide-induced endotoxemia in mice.14 In addition to the cell-surface EPCR, soluble EPCR lacking the transmembrane helix of native EPCR interacts with the integrin CD11b/CD18 (Mac-1) (␣M2) (CR3) on leukocytes (Figure 1), suggesting that binding of soluble EPCR to Mac-1 might regulate leukocytes adhesion.15 Proteinase-3 (PR3), a serine protease with elastase-like properties stored in granules of neutrophils, binds both Mac-1 and soluble EPCR which may be implicated in APC mediated signaling and activation of PC on leukocytes, because soluble EPCR:PR3 complexes bind both APC and PC.15 Recent evidence indicates that EPCR is expressed on different cells beyond aortic endothelial cells. For example, EPCR is expressed on the surface of monocytes, CD56⫹ natural killer cells, neutrophils and eosinophils16 –18, immature hematopoietic stem cells,19 brain capillary endothelial cells.13,20 EPCR is also expressed by embryonic giant trophoblast cells, and EPCR is critical for embryo development, because EPCR null mice die in mid-gestation.21 The study by Bretschneider et al, in the present issue of Circulation Research demonstrates functionally active EPCR in systemic vascular smooth muscle cells (SMCs).22 The data show that EPCR mediates APC signaling in SMCs, resulting in increases in intracellular [Ca2⫹]i, phosphorylation of extracellular signal-regulated kinases 1 and 2 (ERK-1/2) and DNA synthesis (Figure 1). Along with EPCR, PAR-1 is also necessary to transduce APC signaling in SMCs. These findings are consistent with previous work demonstrating that both EPCR and PAR-1 are required for APCmediated activation of ERK1/2 and [Ca2⫹]i signaling in systemic and cerebral capillary endothelial cells.5,20 These findings open a new chapter in biology of EPCR by demonstrating that EPCR can be expressed in cells which have different biological functions from endothelial and hematopoetic cells. In this context, it is of note that the microarray gene expression profiling of human cerebral SMCs and astrocytes have indicated the presence of EPCR mRNA and showed that the levels of EPCR transcripts in these brain cells were not altered by normal aging in humans or aging Alzheimer’s type (R. Zidovetski, N. Chow, B. Zlokovic, unpublished data, 2007). The exact role of EPCR and how its expression is regulated in nonendothelial cell types including systemic SMCs, primitive hematopoetic stem cells or cerebral SMCs, remain to be elucidated. A regulatory element 5.5 kb upstream to the human EPCR gene (hEPCR) translation start site is essential for driving hEPCR expression in endothelium and primitive hematopoetic cells which share a common precursor.23 It would be interesting to find out whether the same site in hEPCR is responsible for driving hEPCR expression in human SMCs.22 The present findings by Bretschneider et al22 raise not only a possibility that EPCR could play an important role in regulating the biology of vascular SMCs, but also suggest that its expression in SMCs might have implications for the pathogenesis of cardiovascular diseases such as atherosclerosis, and by extension, cerebral amyloid angiopathy in Alzheimer’s disease. EPCR may be a potential therapeutic target: novel EPCR agonists might modify cytoprotective APC/PC cellular pathways in injured or inflamed blood vessels. We still have much to learn about the role of EPCR beyond the endothelium, and perhaps in the endothelium itself. For The opinions expressed in this editorial are not necessarily those of the editors or of the American Heart Association. From the Frank P. Smith Laboratory for Neuroscience and Neurosurgical Research (M.T., T.C., B.V.Z.), Department of Neurosurgery, University of Rochester Medical Center, NY. Correspondence to Berislav V. Zlokovic, MD, PhD, University of Rochester Medical Center, 601 Elmwood Avenue, Box 645, Rochester, New York 14642. E-mail [email protected] (Circ Res. 2007;100:155-157.) © 2007 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000258167.48227.84 155 156 Circulation Research February 2, 2007 EPCR Leukocyte PAR-1 sEPCR Mac1 Va & VIIIa Vi & VIIIi PR3 Inflammatory Mediators APC Thrombin Y PC APC EPCR PAR-1 PLC IP3 EPCR TM Gi Gq NF-kB Pro-Inflammatory genes 2+ p53 Ca Bcl-2 Bax 2+ Ca regulated Kinases Downloaded from http://circres.ahajournals.org/ by guest on August 3, 2017 Apoptosis activation of initiator caspases Mitochondria ERK1/2 Nucleus pERK1/2 DNA Synthesis Death Receptors example, Dr Esmon has suggested that EPCR appears to be able to translocate from the plasma membrane into the nucleus, a cellular trafficking pattern that is not observed with its Cys to Ser mutant.24 One may speculate that EPCR may internalize its endogenous ligands PC and APC into the cytoplasm and into the nucleus, which in turn could have important implications for regulation of gene expression and control of APC activity. Figure 2a shows dense punctate staining of APC on the surface of brain endothelial normoxic cells, which is completely removed by pretreatment of cells with an antibody that blocks APC binding site on EPCR (Figure 2b),25 demonstrating that APC binding to the cell is highly EPCR-dependent, as suggested.1 On the other hand, A B Figure 1. EPCR binds protein C (PC) on endothelium and presents it to TM (thrombomodulin)-thrombin complex for activation by thrombin. Activated PC (APC) dissociates from EPCR and TM-thrombin complexes and inactivates coagulation factors Va and VIIIa into inactive forms Vi and VIIIi promoting anticoagulation. APC bound to EPCR activates the protease activated receptor-1 (PAR-1) which binds the Gq and Gi proteins to activate phosophlipase(s) (PLC) in SMCs and endothelium resulting in IP3 (inositol triphosphate)-mediated release of intracellular [Ca2⫹]i, activation of mitogen-activated kinases ERK1/2 and stimulation of DNA synthesis.22 In injured endothelium, EPCR-APCmediated PAR-1 activation blocks mitochondria-mediated apoptotic pathway (eg, inhibition of p53 and Bax, activation of antiapoptotic Bcl-2 gene, blockade of caspase-9), and death receptor-mediated pathway through inhibition of caspase-8. Whether the same antiapoptotic signaling exists in injured SMCs is unknown. Soluble form of EPCR (sEPCR) binds APC/PC and forms complexes with proteinase-3 (PR3) which binds to Mac-1 on leukocytes and activates antiinflammatory signaling. EPCR-APC– dependent PAR-1 activation in leukocytes and endothelium blocks the expression of proinflammatory genes by preventing nuclear translocation of nuclear factor-B (NF-B). EPCR translocates itself from the plasma membrane to the nucleus and may internalize APC into injured vascular cells, but the physiological importance of EPCR internalization alone and/or of its ligands remains at present elusive. Figure 2c shows that APC can be transported intracellularly and into the nucleus of brain endothelial cells under hypoxic conditions (Figure 2c), which is associated with cell protection and survival, as reported.13 This transport pattern, however, completely disappears if APC binding to EPCR is blocked (Figure 2d), and cells die as a result of APC unavailability, as shown by faint APC staining on cell debris. Thus, the role of EPCR-APC-PAR-1-dependent intracellular cytoprotective signaling and of EPCR-mediated intracellular trafficking and internalization of APC/PC in normal and injured vascular endothelial cells and SMCs, may open a new era in our understanding of EPCR biology, its physiological and pathological functions, and therapeutic opportunities for C D Figure 2. Human brain endothelial cells were cultured under normoxic and hypoxic (⬍ 1% oxygen, no glucose) conditions, as we described.10,13,20 Human plasma-derived APC (20 nM) was incubated with cells for 4 hour under either normoxic (A and B) or hypoxic (C and D) conditions in the absence (A and C) and presence (B and D) of an anti-EPCR antibody (RCR-252) which blocks APC binding site on EPCR.25 APC binding was detected by an APC/PC-specific antibody (C3), as described,11,13 using confocal microscopy. In normoxic cells, a punctate staining shows binding of APC to the plasma membrane (A) and complete displacement of APC binding by anti-EPCR antibody (B). In hypoxic cells, APC is internalized into the cytoplasm and nucleus which is associated with the cell survival (C). In contrast, anti-EPCR antibody prevents APC binding to hypoxic cells which is associated with loss of protection and cell death (D), as reported.13 Thiyagarajan et al disorders of the cardiovascular system and cerebral circulation. References Downloaded from http://circres.ahajournals.org/ by guest on August 3, 2017 1. Fukudome K, Esmon CT. Identification, cloning, and regulation of a novel endothelial cell protein C/activated protein C receptor. J Biol Chem. 1994;269:26486 –26491. 2. Mosnier LO, Zlokovic BV, Griffin JH The cytoprotective protein C pathway. Blood. 2006. In press. 3. Vu TK, Hung DT, Wheaton VI, Coughlin SR. Molecular cloning of a functional thrombin receptor reveals a novel proteolytic mechanism of receptor activation. Cell. 1991;64:1057–1068. 4. Joyce DE, Gelbert L, Ciaccia A, DeHoff B, Grinnell BW. Gene expression profile of antithrombotic protein C defines new mechanisms modulating inflammation and apoptosis. J Biol Chem. 2001;276: 11199 –11203. 5. Riewald M, Petrovan RJ, Donner A, Mueller BM, Ruf W. Activation of endothelial cell protease activated receptor 1 by the protein C pathway. Science. 2002;296:1880 –1882. 6. Riewald M, Ruf W. Protease-activated receptor-1 signaling by activated protein C in cytokine-perturbed endothelial cells is distinct from thrombin signaling. J Biol Chem. 2005;280:19808 –19814. 7. Nick JA, Coldren CD, Geraci MW, Poch KR, Fouty BW, O’Brien J, Gruber M, Zarini S, Murphy RC, Kuhn K, Richter D, Kast KR, Abraham E. Recombinant human activated protein C reduces human endotoxininduced pulmonary inflammation via inhibition of neutrophil chemotaxis. Blood. 2004;104:3878 –3885. 8. Feistritzer C, Riewald M. Endothelial barrier protection by activated protein C through PAR-1-dependent sphingosine 1-phosphate receptor-1 crossactivation. Blood. 2005;105:3178 –3184. 9. Finigan JH, Dudek SM, Singleton PA, Chiang ET, Jacobson JR, Camp SM, Ye SQ, Garcia JG. Activated protein C mediates novel lung endothelial barrier enhancement: role of sphingosine 1-phosphate receptor transactivation. J Biol Chem. 2005;280:17286 –17293. 10. Cheng T, Petraglia AL, Li Z, Thiyagarajan M, Zhong Z, Wu Z, Liu D, Maggirwar SB, Deane R, Fernandez JA, LaRue B, Griffin JH, Chopp M, Zlokovic BV. Activated protein C inhibits tissue plasminogen activatorinduced brain hemorrhage. Nat Med. 2006;12:1278 –1285. 11. Feistritzer C, Schuepbach RA, Mosnier LO, Bush LA, Di Cera E, Griffin JH, Riewald M. Protective signaling by activated protein C is mechanistically linked to protein C activation on endothelial cells. J Biol Chem. 2006;281:20077–20084. 12. Taylor FB, Stearns-Kurosawa DJ, Kurosawa S, Ferrell G, Chang ACK, Laszik Z, Kosanke S, Peer G, Esmon CT. The endothelial cell protein C receptor aids in host defense against Escherichia coli sepsis. Blood. 2000;95:1680 –1686. EPCR Beyond Endothelium 157 13. Cheng T, Liu D, Griffin JH, Fernandez JA, Castellino F, Rosen ED, Fukudome K, Zlokovic BV. Activated protein C blocks p53-mediated apoptosis in ischemic human brain endothelium and is neuroprotective. Nat Med. 2003;9:338 –342. 14. Iwaki T, Cruz DT, Martin JA, Castellino FJ. A cardioprotective role for the endothelial protein C receptor in lipopolysaccharide-induced endotoxemia in the mouse. Blood. 2005;105:2364 –2371. 15. Kurosawa S, Esmon CT, Stearns-Kurosawa DJ. The soluble endothelial protein C receptor binds to activated neutrophils: involvement of proteinase-3 and CD11b/CD18. J Immunol. 2000;165:4697– 4703. 16. Stephenson DA, Toltl LJ, Beaudin S, Liaw PC. Modulation of monocyte function by activated protein C, a natural anticoagulant. J Immunol. 2006;177:2115–2122. 17. Galligan L, Livingstone W, Volkov Y, Hokamp K, Murphy C, Lawler M, Fukudome K, Smith O. Characterization of protein C receptor expression in monocytes. Br J Haematol. 2001;115:408 – 414. 18. Sturn DH, Kaneider NC, Feistritzer C, Djanani A, Fukudome K, Wiedermann CJ. Expression and function of the endothelial protein C receptor in human neutrophils. Blood. 2003;102:1499 –1505. 19. Balazs AB, Fabian AJ, Esmon CT, Mulligan RC. Endothelial protein C receptor (CD201) explicitly identifies hematopoietic stem cells in murine bone marrow. Blood. 2006;107:2317–2321. 20. Domotor E, Benzakour O, Griffin JH, Yule D, Fukudome K, Zlokovic BV. Activated protein C alters cytosolic calcium flux in human brain endothelium via binding to endothelial protein C receptor and activation of protease activated receptor-1. Blood. 2003;101:4797– 4801. 21. Gu JM, Crawley JTB, Ferrell G, Zhang FJ, Li WH, Esmon NL, Esmon CT. Disruption of the endothelial cell protein C receptor gene in mice causes placental thrombosis and early embryonic lethality. J Biol Chem. 2002;277:43335– 43343. 22. Bretschneider E, Uzonyi B, Weber AA, Fischer JW, Pape R, Lotzer K, Schror K Human Vascular Smooth Muscle Cells Express Functionally Active Endothelial Cell Protein C Receptor. Circ Res. 2007;100: 255–262. 23. Mollica LR, Crawley JT, Liu K, Rance JB, Cockerill PN, Follows GA, Landry JR, Wells DJ, Lane DA. Role of a 5⬘-enhancer in the transcriptional regulation of the human endothelial cell protein C receptor gene. Blood. 2006;108:1251–1259. 24. Esmon CT. The endothelial cell protein C receptor. Thromb Haemost. 2000;83:639 – 643. 25. Ye X, Fukudome K, Tsuneyoshi N, Satoh T, Tokunaga O, Sugawara K, Mizokami H, Kimoto M. The endothelial cell protein C receptor (EPCR) functions as a primary receptor for protein C activation on endothelial cells in arteries, veins, and capillaries. Biochem Biophys Res Commun. 1999;259:671– 677. KEY WORDS: endothelial protein C receptor 䡲 activated protein C 䡲 protease activated receptor-1 䡲 endothelium 䡲 vascular smooth muscle cells Endothelial Cell Protein C Receptor: Role Beyond Endothelium? Meenakshisundaram Thiyagarajan, Tong Cheng and Berislav V. Zlokovic Downloaded from http://circres.ahajournals.org/ by guest on August 3, 2017 Circ Res. 2007;100:155-157 doi: 10.1161/01.RES.0000258167.48227.84 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 2007 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/100/2/155 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/