Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
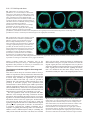
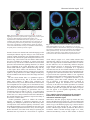
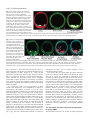
5113 Development 127, 5113-5122 (2000) Printed in Great Britain © The Company of Biologists Limited 2000 DEV5407 A micromere induction signal is activated by β-catenin and acts through Notch to initiate specification of secondary mesenchyme cells in the sea urchin embryo David R. McClay*, Robert E. Peterson, Ryan C. Range, Anne M. Winter-Vann and Michael J. Ferkowicz Department of Biology, DCMB Group, Box 91000, Duke University, Durham, NC 27708, USA *Author for correspondence (e-mail: [email protected]) Accepted 25 September; published on WWW 2 November 2000 SUMMARY At fourth cleavage of sea urchin embryos four micromeres at the vegetal pole separate from four macromeres just above them in an unequal cleavage. The micromeres have the capacity to induce a second axis if transplanted to the animal pole and the absence of micromeres at the vegetal pole results in the failure of macromere progeny to specify secondary mesenchyme cells (SMCs). This suggests that micromeres have the capacity to induce SMCs. We demonstrate that micromeres require nuclear βcatenin to exhibit SMC induction activity. Transplantation studies show that much of the vegetal hemisphere is competent to receive the induction signal. The micromeres induce SMCs, most likely through direct contact with macromere progeny, or at most a cell diameter away. The induction is quantitative in that more SMCs are induced by four micromeres than by one. Temporal studies show that the induction signal is passed from the micromeres to macromere progeny between the eighth and tenth cleavage. If micromeres are removed from hosts at the fourth cleavage, SMC induction in hosts is rescued if they later receive transplanted micromeres between the eighth and tenth cleavage. After the tenth cleavage addition of induction-competent micromeres to micromereless embryos fails to specify SMCs. For macromere progeny to be competent to receive the micromere induction signal, β-catenin must enter macromere nuclei. The macromere progeny receive the micromere induction signal through the Notch receptor. Signaling-competent micromeres fail to induce SMCs if macromeres express dominant-negative Notch. Expression of an activated Notch construct in macromeres rescues SMC specification in the absence of induction-competent micromeres. These data are consistent with a model whereby βcatenin enters the nuclei of micromeres and, as a consequence, the micromeres produce an inductive ligand. Between the eighth and tenth cleavage micromeres induce SMCs through Notch. In order to be receptive to the micromere inductive signal the macromeres first must transport β-catenin to their nuclei, and as one consequence the Notch pathway becomes competent to receive the micromere induction signal, and to transduce that signal. As Notch is maternally expressed in macromeres, additional components must be downstream of nuclear βcatenin in macromeres for these cells to receive and transduce the micromere induction signal. INTRODUCTION Balzer, 1967, pp. 111-112; based upon Boveri, 1901). Boveri, and later Horstadius (Horstadius, 1928), isolated fragments of eggs using the subequatorial pigmented band on Paracentrotus eggs as a landmark. They found that fertilized fragments of eggs containing the vegetal-most cytoplasm were capable of forming skeleton, but no other part of the egg had that capability. Boveri and Horstadius further studied the ‘regulatory activity’ of micromeres. Micromere regulatory activity was demonstrated in a classic experiment by Horstadius. Micromeres recombined with animal halves of 8-cell stage embryos induced the animal halves to produce endoderm. In the absence of micromeres the animal halves developed into dauer blastulae with no mesoderm or endodermal structures The earliest known specification event in sea urchin embryos occurs in micromeres. As a result of an unequal fourth cleavage, micromeres arise at the vegetal pole below their large sister macromeres. Experiments at the turn of the last century demonstrated that cytoplasmic determinants localized to the vegetal pole were the earliest potential cause of this specification event. In 1901 Boveri recognized that “the area nearest the vegetative pole possesses the greatest potentiality to bring development completely to the pluteus stage. It is the ‘priority region’ where differentiation begins. And when differentiation has begun, ‘from this center all other regions are determined in their role by a regulatory action” (cited in Key words: Notch, β-catenin, Induction, Specification, Sea urchin 5114 D. R. McClay and others (Horstadius, 1939). Those experiments were interpreted to suggest that one of the early functions of micromeres was to provide an inductive signal to the macromere cells above them. Horstadius further showed that a secondary axis could be induced if he transplanted micromeres to the animal pole. This experiment dramatically demonstrated the inductive capacity of micromeres. For more than 75 years since that experiment there has been interest in the mechanism of micromere induction. Recent experiments returned to the micromere inductive capabilities at a molecular level. Micromeres were transplanted to the animal pole, as Horstadius had done, and the induced, ectopic, secondary gut expressed lineage markers just like those expressed in the endogenous gut (Ransick and Davidson, 1993). Furthermore, Ransick also found that micromeres were necessary for normal gut specification, at least for the early expression of endoderm markers (Ransick and Davidson, 1995). In other recent experiments absence of micromeres led to the absence of secondary mesenchyme cells (SMCs) in host embryos (Sweet et al., 1999). This led Sweet et al. to propose that an endogenous micromere signal induces SMCs. The mechanism of that induction was not clear, though from experiments by others (Sherwood and McClay, 1999), signaling through the Notch pathway was suggested by Sweet et al. as an obvious possibility. Maternal Notch is distributed on every blastomere in early cleavage. When SMCs are specified Notch disappears from cells of the presumptive SMC lineage (Sherwood and McClay, 1997). This loss of Notch is the earliest known lineage marker for the SMCs. Manipulation of Notch alters the number of SMCs: if embryos express constitutively active Notch, extra SMCs develop; conversely if one expresses dominant-negative Notch in the embryo, SMCs are reduced in number or eliminated altogether (Sherwood and McClay, 1999). These data suggest that Notch signaling is involved in the SMC specification. Another early molecular event in vegetal specification involves components of the Wnt pathway. Late in the fourth cleavage β-catenin enters the nuclei of micromeres (Logan et al., 1999). If nuclear entry of β-catenin is prevented, the micromeres fail to become specified toward their normal skeletogenic fate (Logan et al., 1999) (Wikramanayake et al., 1998) (Emily-Fenouil et al., 1998), and in the absence of nuclear β-catenin, markers show that micromeres become specified as ectoderm (Logan et al., 1999). Micromeres without nuclear β-catenin do not have the ability to induce a secondary axis when transplanted to the animal pole of host embryos (Logan et al., 1999). Thus β-catenin appears to be required for micromere signaling competence. Furthermore, macromeres lacking nuclear β-catenin fail to specify endoderm and macromeres expressing excess β-catenin are vegetalized. Given that both β-catenin and Notch appear to be involved in early specification of vegetal pole tissues, and that the micromere induction signal is required for endoderm and mesoderm specification, we designed a series of experiments to determine the nature of the micromere induction signal pathway. The results of that effort show that β-catenin is upstream of a micromere induction signal, and β-catenin is also upstream of the macromere’s response to that induction signal. We demonstrate that the micromere induction signal induces SMCs at the eighth to tenth cleavage through activation of Notch. If micromeres are eliminated, SMC specification is rescued by expression of activated Notch signal in macromeres. The possible existence of an earlier endoderminducing activity of micromeres is discussed. MATERIALS AND METHODS Animals Adult Lytechinus variegatus were obtained from Jennifer Keller (Beaufort, NC, USA) and from Susan Decker (Hollywood, FL, USA). Gametes were harvested and fertilized in artificial sea water (ASW) as described (Hardin et al., 1992). Embryos were cultured at 21-23°C. mRNA preparation and injection into zygotes All LvNotch, β-catenin and cadherin DNA constructs used in this study have been previously described (Miller and McClay, 1997; Sherwood and McClay, 1999) and were used as templates to generate in vitro transcribed 5′ capped mRNAs using the T3 mMessage mMachine kit (Ambion). The concentrations of mRNAs were determined and the mRNAs were mixed with glycerol (40% v/v, containing FITC-dextran 40 kD, 10%) for injection as described (Sherwood and McClay, 1999). Eggs were passed (3-5×) through 102 µm Nitex cloth to remove the jelly coat, and were aligned on 60 mm Petri dish covers treated with 1% protamine sulfate, fertilized and injected as described (Sherwood and McClay, 1999). Embryos to be dissected later were fertilized in 5 mM p-aminobenzoic acid (PABA) to prevent fertilization membrane hardening. Micromere transplants Embryos to be dissected were transferred by mouth pipet to calciumfree SW. They were then inserted into Kiehart chambers (Kiehart, 1982) that were inverted relative to the original protocol. A joystick micromanipulator was used to manipulate a glass needle as the dissection tool. The needle was broken to make a blunt tip with an inside diameter just smaller than the diameter of the cells to be transplanted. Micromeres were removed by suction using a Gilmont 2 ml microsyringe containing silicon oil attached to the needle to provide a suction action. For micromere transplantations, the four micromeres were first removed from host embryos then donor micromeres were harvested from either red (RITC-stained, as in Logan et al., 1999) or green embryos (FITC coinjected with RNA) and transplanted into position on the hosts. The color of the PMCs later in development unambiguously identified the origin of the transplanted micromere (thus preventing misinterpretation due to inadvertent failure to remove all micromeres from the host embryo). The embryo was slightly compressed between two coverslips for several minutes to hold the donor cell(s) in place. The chamber was then flooded gradually with ASW and the embryos were then transferred to 96-well plates for culture in ASW. Immunolocalization and image analysis Late mesenchyme blastula embryos were fixed in glass depression slides with cold methanol for 10 minutes. After fixation, the embryos were washed 2× with PBS, blocked for 10 minutes in PBS/5% normal goat serum (NGS) (Gibco/BRL), and incubated in primary antibody for 2-3 hours at room temperature or overnight at 4°C, again with the antibody diluted in PBS/5% NGS. Embryos were then washed 3× with PBS/5% NGS, incubated in secondary antibody (Cy5-, Cy2- or CY3-conjugated; Jackson Immunoresearch Laboratories) for 2 hours, washed 3× with PBS, and mounted in 7:3 (v/v) glycerol:PBS. βcatenin was localized with a guinea pig α-β-catenin pAb as previously described (Logan et al., 1999; Miller and McClay, 1997). Notch was identified with an antibody that stains the extracellular EGF-repeat region of the molecule (Sherwood and McClay, 1997), and SMCs were stained with SMC-1, a marker that specifically stains Micromere induction signal 5115 this lineage shortly after it is first specified (Sweet et al., 1999). Images were obtained by sequential confocal sections of embryos using a Zeiss 410 laser-scanning microscope. All images were collected at 2 µm intervals. To calculate the area of SMC territory the length of SMC specified tissue was measured in each section of a complete stack. Given that each image was 2 µm apart the area was then calculated. In the images below, blue was the assigned pseudocolor of cells labeled with fluorescein dextran or stained with Cy2 secondary antibody; red was the assigned pseudocolor for cells labeled with rhodamine or stained with Cy3 secondary antibody; green was the assigned pseudocolor for cells stained with Cy5 secondary antibody. RESULTS Micromeres are necessary for induction of SMCs If one removes micromeres no SMCs are specified (Sweet et al., 1999). Fig. 1 confirms this and shows further that if one removes micromeres and then transplants donor micromeres from another embryo, the ability to make SMCs is rescued. Donor micromeres were labeled with rhodamine isothiocyanate (Logan and McClay, 1997). Host embryos were unlabeled. Host micromeres were removed at the 16-cell stage and replaced with red donor micromeres (Fig. 1C), or not replaced (Fig. 1B). Fig. 1A shows a control embryo with a normal area of SMC specification at the vegetal plate. Notch disappears from presumptive SMCs between the eighth and tenth cleavage, making it a convenient early marker of SMC specification (Sherwood and McClay, 1997; Sherwood and McClay, 1999) (Fig. 1A,C). The Notch disappearance involves endocytosis of the extracellular region, presumably as part of the SMC specification process (Sherwood and McClay, 1999). Apical Notch staining at this time is observed lateral to the SMCs in a ring around the vegetal plate. Lineage marking shows these cells to be presumptive endoderm (Sherwood and McClay, 1997; Sherwood and McClay, 1999). If micromeres are removed at the 16-cell stage, the embryo fails to activate Notch signaling, no SMCs are specified at the vegetal pole, and instead the cells express apical Notch at that location, indicating that endoderm has been specified in the absence of SMC induction (Fig. 1B). The requirement for micromeres is indicated in Figs 1A,C in that if present, or returned to micromereless embryos, (red donor cells in Fig. 1C), SMCs are specified. Thus micromeres are necessary for induction of SMCs. Activation of β-catenin and the presence of micromeres induce SMCs Loss of nuclear β-catenin (Logan et al., 1999) or a block in Notch signaling with a dominant-negative form of Notch, results in few, if any, SMCs (Sherwood and McClay, 1999). We concluded that both β-catenin and Notch are necessary for SMC specification. These earlier experiments did not indicate which cells require β-catenin signaling and which cells receive the Notch signal, since the constructs are expressed ubiquitously. To sort out the requirements for β-catenin and Notch we undertook transplant experiments where a donor and host combination permitted us to learn where β-catenin and Notch were required for the induction sequence. In the first set of experiments, if one vegetalizes embryos by ubiquitous expression of the stable form of β-catenin, embryos specify excess endoderm, and in the absence of micromeres, no SMCs (Fig. 2A). If micromeres are present, excess expression of β-catenin results in overproduction of endoderm plus excess numbers of SMCs (Fig. 2B). In these embryos the entire surface of the embryo expresses apical Notch except for the presumptive SMC region. Micromeres induce SMCs no matter where they are placed in such embryos. For example, in Fig. 2C, red control micromeres were placed at the animal pole of activated β-catenin-expressing hosts and a second site of SMC specification was set up there. Absence of β-catenin in the nucleus has the reciprocal phenotype since no mesoderm or endoderm are specified in such cases (Logan et al., 1999). We conclude that β-catenin is necessary for specification of vegetal tissues and micromeres are necessary for specification of SMCs in those vegetal tissues. The difficulty with these data is that they reveal little about how β-catenin might be involved in SMC and endoderm specification other than being required. To establish details about how β-catenin and Notch work in SMC and endoderm specification we undertook a series of experiments designed to learn where and when these molecules are required in the induction process. The following set of experiments sequentially examines the induction sequence and the roles of β-catenin and Notch. Micromeres require nuclear β-catenin to attain competence as inducing cells The animal pole transplant experiment is the classic way to demonstrate inducing activity of micromeres (Ransick and Davidson, 1993; Horstadius, 1939). Earlier we demonstrated that nuclear β-catenin is necessary for that inductive capacity (Logan et al., 1999). Unfortunately that experiment does not allow one to conclude that the activity exhibited at the animal pole is the same activity normally used by micromeres at the vegetal pole. Accordingly we asked whether β-catenin is required by micromeres for their normal inductive activity at the vegetal pole. Fig. 3 shows the result of a micromere swap experiment to test the hypothesis that β-catenin is required for endogenous micromere inductive competence. Because βcatenin is present in all cells and used by other cells in the embryo for signaling (see below), it was important to alter β-catenin just in the micromeres. This was possible with the swap experiment. Two populations of embryos were cultured. Control micromere donor embryos were labeled red at the 8-cell stage and micromeres removed from these embryos at the 16-cell stage. Micromeres from embryos injected with truncated cadherin are labeled blue in Fig. 3. The expression of truncated cadherin in the plasma membrane eliminates the nuclear signaling capacity of endogenous β-catenin by preventing it from entering the nucleus (Logan et al., 1999). Host embryos had their micromeres removed at the 16-cell stage. Immediately thereafter they received either red control donor micromeres, or blue micromeres that were unable to signal through nuclear β-catenin. The red control micromeres induced SMCs as expected (Fig. 3A) (n=79/79). Blue micromeres lacking nuclear β-catenin failed to induce SMCs (Fig. 3B) (n=29/31) and also failed to ingress into the blastocoel, since without nuclear β-catenin they were unable to follow the PMC lineage. Thus transplantation of micromeres lacking 5116 D. R. McClay and others Fig. 1. Micromeres are necessary for SMC specification. (A) A control embryo is stained with Notch (green) and a marker for PMCs (blue). At the vegetal pole the central region is unstained due to an earlier consumption of Notch, delineating SMC specification (between the arrowheads). Lateral to the vegetal pole Notch is strongly expressed apically on cells that are fated to be endoderm, thereby providing a marker of endodermal specification. (B) Micromeres were removed at the 16-cell stage (confirmed by the absence of PMCs in the blastocoel). The entire vegetal plate stains apically, indicating endoderm specification and no SMC specification. (C) Red donor micromeres were transplanted to the vegetal pole of micromereless embryos at the 16-cell stage. SMC specification occurred, as revealed by the unstained region in the vegetal plate (arrowheads). Fig. 2. Augmentation of β-catenin signaling expands the endoderm and accommodates expanded SMC induction. Zygotes were injected with 0.03 pg of RNA encoding stabilized β-catenin. (A) Apical Notch surrounds a micromereless embryo, indicating a strongly vegetalized phenotype, yet no SMCs are specified. (B) The same amount of β-catenin was injected, and this time micromeres were present. The resulting embryo was vegetalized and had a vastly expanded area of SMC specification in addition (arrowheads). (C) Red micromeres were added to the animal pole of vegetalized embryos inducing an SMC territory at the animal pole. Blue endogenous PMCs induced the vegetal SMC territory (lower arrowheads) and the red PMCs were responsible for inducing the animal SMC territory (upper arrowheads). nuclear β-catenin mimics the phenotype seen in the micromereless embryos as in Fig. 1. These data support the hypothesis that β-catenin is necessary in micromeres for production of the micromere inductive signal. The micromere induction signal is short-range and quantitative Now that we had an assay for micromere inductive activity a number of experiments could be performed to characterize the properties of the signal. First, we asked whether the signal acts at a short range. Normally the vegetalmost veg 2 cells are specified as SMCs while other veg 2 cells are specified as endoderm. The cells specified as SMCs normally are in close proximity to micromere progeny. From experiments with activated Notch we knew that the region of SMC specification could be expanded to include most, if not all, veg 2 progeny (Sherwood and McClay, 1999). First, we designed experiments to ask whether micromere position dictated the induction, or if induction always occurred in the vegetalmost veg 2 cells. The first hypothesis tested was the prediction that SMC induction occurs through short-range interactions between micromeres and responding macromere progeny. To test this hypothesis we needed to distinguish the location of the added micromeres relative to the true vegetal pole. All micromeres were removed from hosts at the 16-cell stage. A single red signaling-incompetent micromere was added at the true vegetal pole to provide a spatial point of reference. As shown above, a micromere expressing the cadherin cytoplasmic tail is incapable of signaling and inducing SMCs. To confirm that, identical control cadherin-injected embryos (including the embryo from which the micromere was taken) were cultured and in all cases these controls developed as ectoderm-only dauer blastulae. This gave us a point of reference to locate the original vegetal pole later in development. Next, we placed a single blue signaling-competent micromere at a position lateral to the vegetal pole. To label the blue signaling micromere, FITC was introduced into that donor embryo shortly after fertilization (actually the ‘blue’ is the assigned pseudo-color of Fig. 3. β-catenin is required in micromeres for the micromeres to induce SMCs. (A) Control red micromeres were transplanted to the vegetal pole of micromereless embryos at the 16-cell stage. Those cells became PMCs (red) and induced the formation of SMCs at the vegetal pole. (B) Blue micromeres from embryos expressing truncated cadherin were transplanted to the vegetal pole of control micromereless embryos. No SMCs were induced as shown by the expression of Notch throughout the vegetal pole region, and the blue micromeres remained in the blastoderm rather than ingress as PMCs. Micromere induction signal 5117 Fig. 4. The micromere inductive signal is short-range. A red nonsignaling micromere was placed at the vegetal pole to provide a point of reference. Blue signaling-competent micromeres were transplanted at several distances from the vegetal pole. (A,B) Two examples where SMCs were induced lateral to the vegetal pole. In (A) the inducing micromere was placed closer to the vegetal pole than in (B). Arrowheads in A and B mark the vegetal pole and the center of the induced SMC territory. The endoderm is delineated by the apical pattern of Notch expression in green. the FITC fluorescence obtained with confocal imaging). At the mesenchyme blastula stage the embryos were fixed and the position of SMC specification was determined relative to the position of the inserted red signal-incompetent micromere. As shown in Fig. 4, the reference red cells failed to induce SMCs and were surrounded by endoderm, as stained with apical Notch. In every case the site of SMC induction occurred at the position of the inserted blue signaling micromere (we looked only at transplants in the vegetal hemisphere). We conclude that the SMC inductive signal is indeed short-range and may in fact require direct contact with micromeres. Further, we conclude that cells lateral to the true vegetal pole are perfectly capable of being induced to be SMCs, though normally they become endoderm in the absence of the short-range micromere signal. We next asked whether there is a quantitative aspect to micromere induction activity, that is, do more micromeres induce larger numbers of SMCs? This experiment was performed in two ways. First, different numbers of micromeres were transplanted to the vegetal pole of micromereless hosts. After induction the area of SMC specification was measured. Fig. 5 shows the results of that experiment. The area of SMC specification at the beginning of gastrulation shows an incremental increase in embryos with 0, 1, 2 or 4 micromeres (0, 644, 1408 and 2335 µm2, respectively, of SMC induction surface area (n=6+ embryos measured with each combination). We conclude that there is a quantitative aspect to the ability of micromeres to induce SMCs. Another quantitative question is whether the induction response is amplified if vegetalized micromeres are the inducing cells. Ideally this question will be addressed by direct overexpression of the inductive ligand by micromeres. Since the inducing ligand has yet to be identified our only choice was to ask whether vegetalized micromeres convey an extra signal. Donor micromeres were taken from embryos expressing the stabilized form of β-catenin at a level known to vegetalize Fig. 5. The number of micromeres correlates with the size of the SMC induction field. 0 (A), 4 (B), 1 remaining (C), or one red micromere added to a micromereless embryo (D) are compared using apical Notch to delineate the extent of endoderm surrounding the SMC-specified territory. The results show the sections with the widest expanse induced to specify SMCs (between arrows). The actual areas of the territories measured are reported in the text. whole embryos (Logan et al., 1999). When untreated host micromereless embryos received one vegetalized micromere there was a slight increase in induced SMCs relative to controls but the difference was not statistically different. Response to the induction, however, is affected by vegetalization. If a single control micromere is tranplanted to a vegetalized micromereless host, the area of SMC induction is twice that of the control response (one control micromere transplanted to the vegetal pole of a control micromereless host). We cannot know at present from this experiment whether or not vegetalized micromeres produce more inductive signal, but even if they do, the inductive influence of a vegetalized micromere is not significantly greater than control induction. The receptive area of sensitivity to the inductive signal is affected by vegetalization though the mechanism for that increased sensitivity is not known. From these experiments we conclude that the area of SMC induction depends on the number of micromeres present at the vegetal pole. The inductive response does not appear to be amplified significantly when the micromeres are vegetalized. The β-catenin signal in macromeres is necessary before these cells are competent to receive the micromere induction signal Macromeres and their progeny import β-catenin into their nuclei at the fifth cleavage (Logan et al., 1999). From then until about the tenth cleavage β-catenin continues to be localized in nuclei of veg 2 cells and their progeny. Veg 1 cells lose nuclear β-catenin at the sixth cleavage but veg 1 progeny fated to become endoderm again localize β-catenin to their nuclei at around the tenth to eleventh cleavage (Logan et al., 1999; Sherwood and McClay, 1999). In earlier studies we showed 5118 D. R. McClay and others Fig. 6. A β-catenin signal in macromeres is necessary for SMC specification. (A) Control embryos at the 16-cell stage had their micromeres removed and replaced with control Signaling-competent micromeres. The embryo in A has PMCs that ingressed normally, and SMCs that were specified at the vegetal plate as normal (arrow points to cells stained with SMC 1 antibody). (B) RNA expressing truncated cadherin makes an embryo specify neither endoderm nor SMCs (no SMC-1 staining), and fails to specify the micromere lineage (no PMCs in the blastocoel). (C) Control micromeres were added to a micromereless embryo that had been injected with RNA to truncated cadherin. Those micromeres became PMCs, had inductive competence, ingressed normally and expressed the PMC marker, but the macromeres, without nuclear β-catenin, were unable to respond to the control micromere induction signal, and failed to specify either endoderm or SMCs. Fig. 7. SMCs are induced through the Notch signal. (A) One red micromere was transplanted onto the vegetal plate of control micromereless embryos. As before, SMCs were specified (between arrowheads). (B) Fertilized eggs received RNA expressing activated Notch. At the 16-cell stage micromeres were removed from these embryos. In the absence of micromeres, but in the presence of activated Notch, SMCs were specified in these embryos (between arrowheads). (C) Red signaling-competent micromeres were added to micromereless embryos that had been injected with RNA expressing dominant-negative Notch RNA. These embryos failed to specify SMCs despite having a competent induction signal. (D) The dominantnegative Notch was expressed only in micromeres. These cells still induced SMCs when transplanted to control micromereless hosts (between arrowheads). Thus, Notch is unnecessary in micromeres for the induction signal, but necessary in macromeres for reception of the signal. that if β-catenin is prevented from entering the nuclei of macromeres these cells develop as ectoderm and fail to specify either endoderm or SMCs (Logan et al., 1999). In that experiment β-catenin also failed to enter micromere nuclei. Thus, it was possible that entry of β-catenin into macromere nuclei required the micromere signal and/or endoderm specification required β-catenin in micromeres. To address these questions we asked if signaling-competent micromeres could rescue endoderm and/or SMCs in macromeres lacking nuclear β-catenin. Fig. 6 shows the results of a swap experiment in which normal (red) micromeres were transplanted to nuclear βcatenin-minus macromeres. As shown in Fig. 6C, when signaling-competent micromeres are provided the β-cateninminus macromeres fail to specify SMCs (nor do they specify endoderm). Embryos in Fig. 6 are stained with two markers, both of which are mouse IgMs. The SMC marker (arrow, Fig. 6A), stains a population of SMCs (Sweet et al., 1999), and the other marker stains PMCs (the same color in this figure). The failure to express the SMC marker in macromeres lacking nuclear β-catenin (Fig. 6C) indicates that β-catenin is necessary in macromeres to receive the micromere induction signal. In Fig. 6C the control micromeres ingress to become PMCs but the remainder of the embryo remains ectoderm-like. Control micromereless embryos failed to specify SMCs as in Fig. 1 (Fig. 6B). We conclude that β-catenin is required in macromeres for these cells to become receptive to the micromere induction signal. From earlier data we suspected the micromere induction signal to act through Notch (McClay and Sherwood, 1999; Sweet et al., 1999). This prompted an examination of the relationship between β-catenin and Notch in macromeres. Notch in macromere progeny is maternal and is expressed on macromeres even in the absence of nuclear β-catenin. Thus, β-catenin is necessary either to augment maternal Notch with zygotic copies of the Notch protein, or to establish other components in the complete Notch signal transduction pathway. Double injections of RNA encoding activated Notch and the truncated cadherin fail to rescue β-catenininhibited macromeres (data not shown). Since activated Notch fails to rescue the β-catenin-deficient macromeres we conclude that β-catenin is independently necessary in both micromeres and macromeres. In micromeres β-catenin is necessary for specification of the PMC lineage and for the pathway leading to production of the micromere inductive signal. In macromeres β-catenin is required for vegetal specification and for competence of the Notch signaling pathway. The micromere induction signal acts through Notch to specify SMCs The experiments of Sweet et al. (1999), our earlier expression Micromere induction signal 5119 data (Sherwood and McClay, 1999), and the experiments above suggested that Notch might be the receptor for the micromere induction signal. If that were true several predictions were testable. First, in the absence of the micromere induction signal, provision of an activated Notch signal to macromere progeny should rescue SMCs. To test this hypothesis micromeres were removed from embryos expressing the constitutively activated form of Notch. Fig. 7B shows that a territory of SMC specification appears in these embryos. As shown earlier, uninjected micromereless embryos fail to specify SMCs. Thus, activated Notch rescues SMCs in the absence of the micromere inductive signal. A second prediction is that provision of signaling-competent micromeres should fail to induce SMCs if dominant-negative Notch is supplied to macromeres. Results in Fig. 7C show that prediction to be supported as well. Signaling-competent micromeres do indeed fail to induce macromeres expressing the dominant-negative form of Notch. Third, if Notch is required in macromeres only, micromeres expressing the dominant-negative form of Notch should be competent to signal and induce SMCs (Fig. 7D). This prediction is correct. SMCs are specified by micromeres expressing dominant-negative Notch. These data are consistent with the model that Notch, in macromere progeny, is the receptor for the micromere inductive signal. The rescue experiment with activated Notch demonstrates a curious property of the induction response. Earlier it was shown that the signal from micromeres provides spatial localization of SMC specification. Fig. 7 shows that in the absence of micromeres the SMCs are specified at the vegetal pole. All cells of the micromereless embryo in Fig. 7B express activated Notch but only vegetal pole cells become specified as SMCs. The number of cells that become specified as SMCs varies with the concentration of RNA to the activated Notch. We conclude that the position of SMC specification is decided both by proximity to micromere-released signal, and by an intrinsic bias in the vegetalmost veg 2 cells. The data support a model whereby the most sensitive area for reception of the micromere induction signal is at the vegetal pole. Lateral to that, induction will occur if the micromeres are placed ectopically over vegetally specified cells, or if activated Notch signal is provided the territory is expanded. The micromere induction signal acts between the eighth and tenth cleavage division, or approximately at the time when the Notch signal is active in veg 2 cells The disappearance of Notch from presumptive SMCs occurs at around 7 hours after fertilization or roughly between the eighth to tenth cleavages (Sherwood and McClay, 1997; Sherwood and McClay, 1999). Based on the phenotypes seen with either activated or dominant-negative Notch, it was proposed that Notch is removed from the plasma membrane of SMCs as it signals or shortly thereafter, thereby creating the vegetal region devoid of Notch. Knowing that Notch leaves the vegetal plate during the eighth to tenth cleavages we asked when, relative to those cleavage divisions, does the micromere induction signal occur. Micromeres were removed from the embryos at the fourth cleavage. The micromereless embryos were then cultured for varying periods of time before receiving blue, signaling- competent micromeres from donor embryos. In the first experiments we transferred micromeres from 16-cell stage donors to older host micromereless embryos. Hosts received the donor micromeres at the fifth-seventh and ninth cleavages, or after being without micromeres for 1-3 or 5 cleavages. After as many as 3 (or 4 cleavages), readdition of micromeres allowed SMCs to be specified in the host embryos (Fig. 8C). However, if the micromereless embryos were missing micromeres for 5 cleavages we saw no SMC territory specified and the embryos displayed apical Notch throughout the vegetal hemisphere as is typical of control micromereless embryos (Fig. 8D). We conclude that micromeres are unnecessary for SMC induction for several cleavages after their appearance and that the inductive signal is necessary at or sometime shortly after the eighth cleavage. It was possible that failure to induce SMCs at the ninth cleavage was because the young micromeres simply were not capable of immediately inducing SMCs when transferred at the ninth cleavage. If this were the case we predicted that older donor micromeres should be able to immediately induce if transplanted to ninth cleavage hosts (assuming this is actually when the induction occurs). Accordingly micromeres were labeled red with RITC and transferred initially to surrogate unlabeled host embryos for varying intervals. At the same time the experimental hosts had their micromeres removed. When the micromereless embryos reached the ninth cleavage the red micromeres were harvested from the surrogate hosts and inserted into the vegetal plate of the micromereless hosts. The older micromeres induced SMCs (Fig. 8E). We conclude that the micromere induction signal is unnecessary for SMC induction prior to the eighth or ninth cleavage. We were unable to rescue induction when micromeres were returned to embryos at tenth cleavage or thereafter. These data further support the model that an immediate consequence of the micromere induction signal is the surface disappearance of Notch in the cells that have just received the induction signal through Notch. DISCUSSION Micromeres are shown to induce SMCs through Notch signaling. The induction occurs between the eighth and tenth cleavages and it relies on close or direct contact between the micromeres and the targeted cells of the veg 2 layer. As a consequence maternal Notch disappears from the cells that will become SMCs during or following reception of the micromere signal. The dynamics of Notch disappearance are incompletely understood but appear to be similar to those seen in other systems using the Notch signaling pathway. By immunofluorescence, the extracellular region of Notch relocates to intracellular vesicles at the eighth to tenth cleavages; coincident with the time that SMCs receive the micromere induction signal (Sherwood and McClay, 1997). Whether endocytosis occurs during or following the signal reception is not known. Either way, Notch is lost from the newly specified SMC population, leaving that population negative for Notch. Of the many uncertainties that remain, the precise time that micromeres become competent to signal is not known since the micromere ligand that activates Notch has not been identified. Based on the timing experiments it is 5120 D. R. McClay and others Fig. 8. Micromeres induce at the eighth to tenth cleavages. Micromeres were removed from host embryos at the 16-cell stage. In (A), a control micromereless embryo expresses Notch throughout the vegetal region. Blue donor micromeres were returned to similarly treated hosts either immediately, one cleavage later (fifth) (B), three cleavages later (seventh) (C), or 5 cleavages later (ninth) (D). The blue micromeres were in each case from fourth cleavage donors. In (E) ninth cleavage micromereless embryos received red donor micromeres that were also in the 9th cleavage at the time of donation. The embryos were stained with Notch (green) to indicate apical endoderm and the area of SMC specification (cells missing apical Notch at the vegetal pole, between arrowheads). predicted that the induction signal is present on the micromeres as early as the sixth cleavage. The micromere induction signal described herein appears to be a single event that activates Notch. However, these data do not mean that the Notch induction signal is the only signal from the micromeres. Earlier studies (Ransick and Davidson, 1995), suggest that an additional signal from micromeres may be involved in specification of endoderm. They showed that expression of Endo 16, an early endoderm gene, depends upon the presence of micromeres for its full expression at the vegetal plate. Endo 16 appears in macromere progeny in the absence of micromeres, but presence of micromeres between the fourth and sixth cleavage increases the level of Endo 16 expression at the mesenchyme blastula stage significantly (Ransick and Davidson, 1995). Between the fifth and eighth cleavage β-catenin is in the nuclei first of macromeres and then of the veg 2 layer (the layer immediately above micromeres). Our experiments have shown that β-catenin nuclear entry is essential for macromere endodermal specification. It occurs cell-autonomously, even in the absence of micromeres (Logan et al., 1999). The Ransick/Davidson data point to an additional micromereinduced requirement between the fourth and sixth cleavages that somehow amplifies the initial endoderm specification. Curiously, the endoderm induction appears to be independent of SMC specification because, as shown here, SMC specification does not require micromeres to be present during the fourth to seventh cleavages. Removal of micromeres delays invagination of the archenteron. In part that delay could be due to the failure to fully specify endoderm in the absence of micromeres, but another possible explanation is due to the absence of SMCs. Normally, invagination begins at the vegetal pole with bottle cells initiating the inward bending primary phase of archenteron formation (Nakajima and Burke, 1996; Kimberly and Hardin, 1998). Neither of these groups examined the lineage origins of the bottle cells but by location they are likely to be SMCs. When the entire vegetal pole is endoderm, there may be a reduced capacity to initiate archenteron formation. Invagination of the archenteron eventually occurs, but during the delay the embryo also regulates and replaces the SMCs (Sweet et al., 1999). If β-catenin is depleted from cells so that it cannot enter the nucleus, there is no specification of micromeres (Wikramanayake et al., 1998; Emily-Fenouil et al., 1998; Logan et al., 1999). The asymmetric division that forms the micromeres still occurs, but those micromeres are incapable of inducing a second axis at the animal pole (Logan et al., 1999), and they are incapable of inducing SMCs (Fig. 3). Thus, one of the pathways initiated by nuclear β-catenin in micromeres is the synthesis or activation of the ligand that binds to Notch. The Delta or Serrate homologues are the obvious candidates for the micromere signal but these have yet to be identified and tested for this ligand function in the sea urchin. From the data we have so far, fourth cleavage micromeres are incapable of immediately inducing through Notch. By about the sixth to seventh cleavage micromere progeny have the ability to induce eighth to tenth cleavage veg 2 progeny to specify SMCs through Notch. Thus we predict that when the ligand is identified it will be produced zygotically, or activated within one or two cleavage divisions of nuclear β-catenin in micromeres, and within one or two cleavages after ligand appearance, the SMC induction occurs through the Notch signal. Micromeres begin moving β-catenin into their nucleus during the 20 minutes between the fourth and fifth cleavages. In a population of embryos fixed early between the fourth and fifth cleavages we see only a fraction of the embryos with nuclear β-catenin. That fraction increases if embryos are fixed later in the 20 minute period between cleavages, and by fifth cleavage all micromeres have nuclear β-catenin. These data suggest that β-catenin accumulates in micromere nuclei during the fourth interphase. Is there a Wnt signal to activate β-catenin to go to the nucleus? So far the evidence is negative and points to a cell-autonomous mechanism. If we separate blastomeres at each cleavage and constantly culture the isolated cells separately, β-catenin still enters micromere nuclei at the correct time (Logan et al., 1999). This suggests that a cell-autonomous mechanism governs nuclear entry. Further, in separate experiments with wnt 8 we find no Micromere induction signal 5121 evidence of a wnt signal being involved in directing β-catenin to micromere nuclei (A. H. Wikramanayake et al., unpublished). β-catenin is maternally expressed and appears to be expressed equally by each cell. In situ data also indicate that β-catenin RNA is equally distributed throughout the embryo. By the 16-cell stage the protein is associated with adherens junctions all around the embryo (Miller and McClay, 1997), but then the protein goes into the nuclei only in micromeres (and in the lower tier of macromeres a cleavage later). Inhibition of endogenous GSK-3 enhances movement of β-catenin to the nuclei (Emily-Fenouil et al., 1998), but this does not offer any proof of how nuclear entry normally occurs. Thus, the earliest known molecular event of consequence for mesoderm and endoderm specification occurs at the 16-cell stage, though as yet it is not known what triggers the localized nuclear entry of β-catenin. The induction signal is not immediately present on micromeres but appears shortly thereafter in a βcatenin-dependent process When micromeres from the fourth cleavage are transplanted to the vegetal pole of micromereless embryos at the seventh cleavage, the micromeres induce SMCs during the eighth to tenth cleavage. Fourth cleavage micromeres fail to induce if transplanted to ninth cleavage micromereless hosts. If βcatenin is prevented from entering the nucleus of micromeres the cells are unable to induce SMCs. Together the timing experiments suggest that the micromeres are competent to signal within about 2-3 cleavages of the fourth, or after about 80 minutes of elapsed time. This is plenty of time for several steps in a pathway prior to the production or activation of the Notch ligand, though the synthesis and insertion of the Notch ligand is likely to be slow relative to the production of any transcriptional regulatory factors upstream of that ligand synthesis. Whether the β-catenin-to-ligand production is a direct or an indirect pathway remains to be determined. The micromere induction signal acts at short range No matter where the induction occurs in the vegetal hemisphere, if micromeres are present, SMC induction occurs by direct contact or within at most one cell diameter away from the micromere. Several years ago we showed that micromeres and their PMC progeny extend thin filopodia that extend the apparent range of contact of those cells (Miller et al., 1995). Thus even if the cell body is two or more diameters away from an induced cell it is hard to rule out the possibility that the inducing cell makes direct contact with the induced cell via thin filopodia. These data support the hypothesis that induction requires direct contact or very short distances between the inducing and the receiving cell. Possible exceptions to a direct contact requirement might include cases where the embryo is vegetalized or when the Notch signal is activated at a high level. Even then direct contact might be required in that as a consequence of vegetalization the expansion of the signaling source might be expanded. The inductive signal acts through Notch Notch disappears from the presumptive SMCs at the vegetal pole (Sherwood and McClay, 1999), and our experiments support this disappearance as being a part of the SMC specification sequence. The Notch disappearance appears to be directly related to the Notch signal. Several curious features of this signaling are worth mentioning. When activated Notch specifies SMCs in the absence of micromeres, endogenous Notch disappears from the SMCs as well. Thus one consequence of Notch signaling is removal of Notch from the cell surface, even if that Notch was not involved in the signaling. In any Notch-related induction it is likely that only a small portion of the surface Notch actually is used for the signal, but as shown by the subsequent disappearance, an early consequence of Notch signaling is removal of all remaining Notch receptor from the cell surface (at least in this case). This biology is useful but also complicates future experiments. We are fortunate in these experiments because removal of all remaining Notch provides a convenient marker to indicate that SMC specification has been initiated. The complication will occur in future experiments if only a small amount of Notch actually performs the signaling and the remaining Notch is removed by endocytosis. Western analysis of cells allows one to visualize the fragments of Notch. Unfortunately, the endogenous post-signaling disappearance of Notch makes it difficult to distinguish between fragmentation due to Notch signal transduction, and fragmentation due to post-signaling degradation. A start has been made toward characterization of the micromere signal. There are many gaps to fill, however. We do not know what triggers the β-catenin-induced signaling. We know very little about the pathway between β-catenin and the appearance of the inducing signal. We know little about how Notch transduces the signal and even less about downstream events. Nevertheless, from this study the biological framework is in place for those detailed analyses. REFERENCES Balzer, T. B. (1967). Life and Work of a Great Biologist. Berkeley, CA: University of California Press. Boveri, T. (1901). Uber die Polaritat des Seeigel-Eies. Verh. Phys. Med. Ges. Wurzburg 34, 145-176. Emily-Fenouil, F., Ghiglione, C., Lhomond, G., Lepage, T. and Gache, C. (1998). GSK3beta/shaggy mediates patterning along the animal-vegetal axis of the sea urchin embryo. Development 125, 2489-2498. Hardin, J., Coffman, J. A., Black, S. D. and McClay, D. R. (1992). Commitment along the dorsoventral axis of the sea urchin embryo is altered in response to NiCl2. Development 116, 671-685. Horstadius, S. (1928). Uber die determination des keimes bei echinodermen. Acta Zoologica 9, 1-191. Horstadius, S. (1939). The mechanics of sea urchin development as studied by operative methods. Biol. Rev. 14, 132-179. Kiehart, D. P. (1982). Microinjection of echinoderm eggs: apparatus and procedures. Meth. Cell Biol. 25, 13-31. Kimberly, E. L. and Hardin, J. (1998). Bottle cells are required for the initiation of primary invagination in the sea urchin embryo. Dev. Biol. 204, 235-250. Logan, C. Y. and McClay, D. R. (1997). The allocation of early blastomeres to the ectoderm and endoderm is variable in the sea urchin embryo. Development 124, 2213-2223. Logan, C. Y., Miller, J. R., Ferkowicz, M. J. and McClay, D. R. (1999). Nuclear beta-catenin is required to specify vegetal cell fates in the sea urchin embryo. Development 126, 345-357. McClay, D. R. and Logan, C. Y. (1996). Regulative capacity of the archenteron during gastrulation in the sea urchin. Development 122, 607616. Miller, J., Fraser, S. E. and McClay, D. (1995). Dynamics of thin filopodia during sea urchin gastrulation. Development 121, 2501-2511. Miller, J. and McClay, D. R. (1997). Changes in the pattern of adherens 5122 D. R. McClay and others junction-associated β-catenin accompany morphogenesis in the sea urchin embryo. Dev. Biol. 192, 310-322 Nakajima, Y. and Burke, R. D. (1996). The initial phase of gastrulation in sea urchins is accompanied by the formation of bottle cells. Dev. Biol. 179, 436-446. Ransick, A. and Davidson, E. H. (1993). A complete second gut induced by transplanted micromeres in the sea urchin embryo. Science 259, 1134-1138. Ransick, A. and Davidson, E. H. (1995). Micromeres are required for normal vegetal plate specification in sea urchin embryos. Development 121, 32153222. Sherwood, D. R. and McClay, D. R. (1997). Identification and localization of a sea urchin Notch homologue: insights into vegetal plate regionalization and Notch receptor regulation. Development 124, 3363-3374. Sherwood, D. R. and McClay, D. R. (1999). LvNotch signaling mediates secondary mesenchyme specification in the sea urchin embryo. Development 126, 1703-1713. Sweet, H. C., Hodor, P. G. and Ettensohn, C. A. (1999). The role of micromere signaling in notch activation and mesoderm specification during sea urchin embryogenesis [In Process Citation]. Development 126, 5255-5265. Wikramanayake, A. H., Huang, L. and Klein, W. H. (1998). β-Catenin is essential for patterning the maternally specified animal-vegetal axis in the sea urchin embryo. Proc. Natl. Acad. Sci. USA 95, 9343-9348.