Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
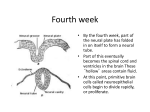
/ . Embryol exp. Morph. Vol. 53, pp. 237-243, 1979 Printed in Great Britain © Company of Biologists Limited 1979 237 The use of avian double monsters in studies on induction of the nervous system By R. J. ULSHAFER 1 AND A. CLAVERT 1 From the Laboratoire d'Embryologie, Faculte de Medecine, Universite Louis Pasteur, Strasbourg, France SUMMARY Ninety-eight duck double monsters (siamese twins) were studied between 48 and 72 h of incubation in whole-mount or serial-sectioned preparations. Each case was classified into one of four categories depending on the orientations of the two embryonic axes. In all classes, defects in primary neural induction of various parts of the embryonic nervous system were observed which depended to a large extent on the distance separating the anterior tips of the two notochords or on the relative positions of the notochords. Our observations suggest that: when the same inducing field of one embryo overlaps to a critical extent with that of the twin, the two inducers act as one; induction of forebrain and optic vesicles is purely prechordal; the notochord exerts an inhibition on induction of forebrain by prechordal mesoderm; and the presence of rhombencephalic structures depends on both prechordal and chordamesodermal interactions. INTRODUCTION The study of naturally occurring or mechanically induced double monsters (siamese twins) offers excellent model systems for problems concerning organizing phenomena, symmetry of body plan, teratogenesis/?er se and also induction and regulation. The phenomenon occurs spontaneously quite frequently in birds, especially in ducks where about 2 % of all fertilized eggs form double monsters; the effect is believed to be caused by orientation changes of the egg within the uterus during the critical period of symmetrization which results in the formation of two (or more) organizing centres (Vintemberger & Clavert, 1956; Clavert, 1962). The phenomenon can also be produced experimentally with up to 80 % success by removing eggs from the uterus prior to the critical period and subjecting them to series of rotations in a mechanical apparatus (Vintemberger & Clavert, 1960). From an experimental point of view, this anomaly corresponds to a heterotopic graft of the primary organizer. The relative positions of the two. organizing centres determine the orientations of the embryonic axes and location of inducing tissue which may therefore have consequences on induction of embryonic structures. The purpose of the present study was to determine if 1 Authors' address: Laboratoire d'Embryologie, Faculte de Medecine, Universite Louis Pasteur, 11 Rue Humann, 67085 Strasbourg Cedex, France. 16 EMB 53 238 R. J. ULSHAFER A N D A. CLAVERT differences exist in brain structure between the twins as a result of interference with induction caused by orientation and location of the embryonic axes of the double monsters. Since, in our study, the effect occurs spontaneously in ovo without the intervention of microsurgery, it presents the advantage of eliminating artifacts that may be introduced during experimental manipulation. MATERIALS AND METHODS Ninety-eight spontaneous double monsters were obtained from about 5000 duck eggs between 48 and 72 h incubation, used in previous or concurrent studies. The blastoderms were excised, photographed using reflected light and then fixed in Bouin's fixative. In about half the cases permanent whole-mount preparations were made, while in the others the embryos were embedded in paraffin, sectioned at 6 jtcm and stained with haematoxylin and eosin. Sections were analysed by counting the number of forebrains, optic vesicles or cups, hypophyses, hindbrains and otic vesicles. In relevant cases the distance separating the anterior extremities of the notochords of the twins was measured using a micrometer having 5 /*m per division under a Leitz optical microscope. RESULTS We observed four different groups of double monsters, as distinguished by the relative positions of the embryonic axes and the number of differences in brain structures noted between the two embryos. Group I. 13 cases. The notochords approached each other medially. The twins shared a common cephalic extremity, having one well-formed prosencephalon with one set of optic rudiments and one or two hypophyses (Fig. 1). Group II. 41 cases. The notochords approached each other medially. Each embryo had its own forebrain which had either both or only one lateral optic rudiment (Fig. 2). F I G U R E S 1-4 Fig. 1. Group I double monster at about 72 h incubation. The twins share a common cephalic extremity. Serial sections of this embryo revealed one forebrain, one set of optic cups, one hypophysis, two hindbrains and two sets of otic vesicles. Fig. 2. Group II double monster at about 60 h incubation. Two separate heads are visible, each having a pair of optic cups, one hypophysis, a normal hindbrain and a pair of otic vesicles. Fig. 3. Group ITI double monster at about 60 h incubation. The twins approach each other frontally, neither one having forebrain or optic rudiments. Serial sections of this monster revealed two hindbrains, but only the one on the left had otic rudiments. Fig. 4. Group IV double monster at about 60 h incubation. The embryonic axes intersect each other almost perpendicularly at the level of the hindbrain. The embryo on the left lacked forebrain and optic vesicles but had normal hindbrain and otic rudiments. The embryo on the right had a normal nervous system. Induction in avian double monsters 239 16-2 240 R. J. ULSHAFER AND A. CLAVERT Table 1. Distance separating the anterior extremities of the notochords and the resulting number of forebrain structures in double monsters which approached each other medially Number of cases Distance separating the Number anterior notochord of (mm) forebrains 6 3 9 0-20-0-40 0-40-0-50 0-45-0-80 Number of optic vesicles Number of hypophyses Class 2 2 4 1 or 2 2 2 I II II 1 2 2 Table 2. Distance separating the anterior extremities of the notochords and the resulting number of hindbrain structures in the frontal approach of the notochords (Group III) Number of cases 4 1 1 9 Distance separating the notochords (mm) Number of hindbrains Number of otic rudiments 0 1 2 2 0 1 2 4 000 (fused) 0-30 0-40 0-48-0-70 Table 3. Level where secondary embryo intersects the first and the resulting presence of neural structures in the secondary embryo of Group 111 monsters Level where secondary Number of embryo meets first cases Fore-/midbrain Hindbrain Spinal cord 6 5 4 Presence of forebrain Presence of hindbrain Presence of spinal cord + 0 0 + + + + + + Group III. 27 cases. The notochords approached each other frontally. Both embryos lacked all forebrain structures. In some cases rhombencephalic structures of both embryos were normal while frequently one or both twins lacked otic rudiments and/or hindbrain entirely (Fig. 3). Group IV. 17 cases. The secondary embryo approached the first somewhat perpendicularly. The primary embryo was normal while the twin lacked optic vesicles and frequently other brain structures (Fig. 4). Within groups I and II a correlation existed between the distance separating the anterior tips of the notochords and the lack of forebrain. structures (Table 1). When the two notochords were separated by a distance less than 0-40 mm (mean 0-30 mm), only one forebrain was induced. When a mean distance of 0-45 mm separated the anterior notochords, two forebrains resulted but only Induction in avion double monsters 241 one optic rudiment per head formed, and always on the lateral side. Only when a distance of at least 0-50 mm (with one exception at 0-45 mm) separated the two notochords, did two normal forebrains occur, each with its own pair of optic vesicles. In group III the presence of hindbrain structures also depended on a critical distance separating the anterior tips of the notochords, which in this case approached each other head-on (Table 2). When the two notochords were separated by a distance of at least 0-48 mm, both embryos developed normal rhombencephalic structures. When the two notochords were fused, no hindbrains or otic rudiments were induced. Two intermediate cases were seen, one having one hindbrain and one otic vesicle and the other having two hindbrains and two otic vesicles. Prosencephalons and optic rudiments never formed in this group. In group IV the axis of one embryo intersected that of the twin almost perpendicularly at the level of the brain or spinal cord. In this group, the embryo whose anterior notochord was located near the trunk of the second developed little or no forebrain and no optic rudiment at all. When the embryos met near the anterior neural tube somewhat more development of the brain occurred than when they met more posteriorly, although optic vesicles never developed in this group (Table 3). DISCUSSION Symmetrization in birds is related to direction of rotation of the egg within the uterus. Eggs of species which have unusually large and elastic uteri, such as the duck, frequently undergo rolling movements which reverse the orientation of the egg in the uterus and thus change the direction of rotation. If this reversal occurs during the critical period of symmetrization (within a span of a few hours) a new centre of symmetrization and therefore a new embryonic axis forms — the result being a double monster. That one of the resulting embryos is always slightly older than the other also indicates that symmetrization, and subsequently gastrulation, of the two embryos do not coincide in time. Due ultimately to the telolecithal nature of the bird egg, a certain degree of continuity of germ layers exists between the two embryos. Even though the neural tubes are frequently discrete, the twins almost always share structures derived from all germ layers. Consequently, identical twins are very rare in birds while Siamese twins are relatively more common. Although the orientation of the embryonic axes of the two embryos ultimately results from the location of the symmetrization centre as a function of direction of rotation and orientation of the egg within the uterus, the morphological groups of double monsters that we described are also due to interactions between tissues of the twins during gastrulation and induction, The monsters in group I arise because of complete overlapping of two prechordal plates, which has the effect of a single inducing field, thus producing 242 R. J. ULSHAFER AND A. CLAVERT one set of forebrain structures of normal dimensions. As the distance separating the anterior tips of the notochords increases, the inducing effect of each prechordal plate becomes more and more distinct, producing the monsters of group IT. It appears that the size of the induced organ therefore does not depend on the mass of inducing tissue but rather on its own intrinsic capacity to respond to the message within a given area. It should be emphasized, however, that the relative distances noted here do not correspond to the actual distances which separated the tissues during primary induction, due to growth of embryonic organs and interstices. In group III the notochords were always so close as to obliterate totally or inhibit the action of the prechordal plate mesoderm on overlying ectoderm, and therefore no forebrain developed. Prechordal plate mesoderm, however, appears necessary for normal induction of hindbrain structures since, in the monsters in which the two notochords were fused no hindbrains were induced, while in those cases where the two notochords were separated by a space containing prechordal plate, two normal hindbrains formed. In group IV one embryo was always normal while the one approaching it from the side always lacked brain structures when it met the first at the level of the trunk but not when they met more anteriorly. Here, again, the prechordal plate of the approaching embryo overlapped the inducing field of the chorda mesoderm of the primary embryo which apparently inhibited prechordal plate from inducing forebrain. Since prechordal plate was present, however, hindbrain structures in the approaching embryo were normal. The more caudal the location of the approaching embryo, the more drastic was the reduction in brain structures. These observations suggest that induction of forebrain and optic vesicles is purely prechordal, that the notochord exerts an inhibitory effect on induction of forebrain by prechordal plate, and that this effect is stronger in the more caudal regions of the notochord. Naturally occurring multiple monsters resemble those produced experimentally by heterotopic grafting of Hensen's node, as reported by Gallera & Ivanov (1964) and Gallera (1971). All of their monsters would belong to our groups III and IV. That none was induced resembling our groups I and II is probably due to their precise orientations of the grafts with respect to the embryonic axis of the host (anterior end of the graft was always turned toward the centre of the host). Although they did not report any inhibiting effect of chorda mesoderm on forebrain, it appears in their figures that the same phenomenon occurred during their experiments. In amphibians, however, only monsters corresponding to our groups I and II are possible (after Spemann, 1962), primarily due to the ovoid nature of the amphibian egg. Our results therefore correspond to most of the generally accepted theories on induction of the nervous system (reviewed by Saxen & Toivonen, 1962) in that the more anterior parts of the nervous system are induced predominantly by prechordal plate, spinal cord is induced purely by chorda and somitic mesoderm, and intervening brain structures by an interplay of the two. Induction in avian double monsters 243 This work was supported by Grant no. ATP 68.78-100 from the INSERM. Dr Ulshafer is supported by the CNRS/NSF Exchange of Scientists Program. REFERENCES CLAVERT, J. (1962). Symmetrization of the egg of vertebrates. Adv. Morphogenesis 2, 27-60. GALLERA, J. (1971). Primary induction in birds. Adv. Morphogenesis 9, 149-180. GALLERA, J. & IVANOV, L. (1964). La competence neurogene du feuillet externe du blastoderme de Poulet en fonction du facteur 'temps'. J. Embryol. exp. Morph. 12, 693-711. SAXEN, L. & TOIVONEN, S. (1962). Primary Embryonic Induction. London: Logos Press limited. SPEMANN, H. (1962). Embryonic Development and Induction. New York: Hafner Publishing Company. VJNTEMBERGER, P. & CLAVERT, J. (1956). Sur la frequence de la monstruosite double et le degre d'instabilite de I'orientation de l'oeuf dans I'uterus chez diflferents Oiseaux. C. r. hebd. Seanc. Acad. Sci., Paris 243, 2149-2151. VINTEMBERGER, P. & CLAVERT, J. (1960). Sur le determinisme de la symetrisation bilaterale chez les Oiseaux: les facteurs de I'orientation de l'embryon par rapport a l'axe de l'oeuf et la regie de Von Baer a la lumiere de nos experiences d'orientation dirigee sur l'oeuf de Poule extrait de I'uterus. C. r. Seanc. Soc. Bio/. 154, 1072-1076. (Received 8 January 1979, revised 27 April 1979)