Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
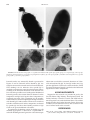
IMMUNOLOGY AND MOLECULAR BIOLOGY In Vitro Studies of Chicken Egg Yolk Antibody (IgY) Against Salmonella enteritidis and Salmonella typhimurium E. N. Lee,* H. H. Sunwoo,* K. Menninen,† and J. S. Sim*1 *Department of Agricultural, Food and Nutritional Science, University of Alberta; and †AAFRD, Animal Health Lab Branch, Edmonton, Alberta, Canada, T6G 2P5 medium. The growth rate of S. enteritidis incubated with S. enteritidis-specific IgY was fourfold less than that of the control group during a 4-to-6-h incubation. Cell counts of S. typhimurium incubated with S. typhimurium-specific IgY were reduced by 1.6 log cfu/mL in comparison to that of the control group after 6 h of incubation. The specific binding activity of IgY was further evaluated by using immunofluorescence and immunoelectron microscopy. It was found that Salmonella-specific IgY could bind to the antigens expressed on the Salmonella surface, resulting in structural alterations of the bacterial surface. ABSTRACT Chicken egg yolk antibody (IgY) raised against Salmonella enteritidis or Salmonella typhimurium was found in highly specific activity levels by ELISA. S. enteritidis- and S. typhimurium-specific IgY powder, prepared by freeze-drying the egg yolk water-soluble fraction, contained 15.5 and 10.0% of specific IgY, respectively. Anti-S. enteritidis IgY cross-reacted 55.3% with S. typhimurium. The cross-reactivity of anti-S. typhimurium IgY with S. enteritidis was 42.4%. Salmonella-specific IgY was demonstrated to inhibit Salmonella growth in liquid (Key words: immunoglobulin Y, Salmonella enteritidis, Salmonella typhimurium, antibacterial activity, immunofluorescence and immunoelectron microscopy) 2002 Poultry Science 81:632–641 binding to bacteria may be essential to promote their antibacterial properties. In this regard, it is necessary that IgY binding activity and its effect on bacteria be investigated in more detail to provide basic information on the antibacterial properties of IgY. Two Salmonella serovars, Salmonella enteritidis and Salmonella typhimurium, were selected as bacterial antigens to raise IgY because they have been the main cause of salmonellosis outbreaks in humans and infections in chickens (O’Brien, 1988; CDC, 1990; Roberts et al., 1996; Khakhria et al., 1997). In vivo studies have shown the preventive effect of IgY on Salmonella infection (Peralta et al., 1994; Yokoyama et al., 1998a,b). However, the immunological properties of IgY on the binding activity against Salmonella have not been reported previously. Therefore, the objective of this study was to investigate the antibacterial properties of IgY against S. enteritidis and S. typhimurium for its specificity and activity by using various in vitro methods, such as ELISA, growth inhibition assay, and microscopy. INTRODUCTION Chicken egg yolk antibody, referred to as Immunoglobulin Y (Leslie and Clem, 1969), differs from mammalian IgG in terms of structural and immunological properties (Higgins, 1975; Kobayashi and Hirai, 1980). It also has advantageous characteristics over mammalian IgG, including stability under various physico-chemical conditions and its suitability as an immunological tool (BarJoseph and Malkinson, 1980; Vieira et al., 1984; Losch et al., 1986; Shimizu et al., 1988, 1992, 1994; Yolken et al., 1988; Otani et al., 1991). IgY can be produced in large quantities from the yolk of chicken eggs by simple methods, and without distress to the animals (Kwan et al., 1991; Akita and Nakai, 1993). Therefore, IgY has received much attention due to its potential as an alternative to conventional mammalian antibodies. Antibacterial properties of IgY have been one of the main interests in IgY studies. Many reports have shown that IgY demonstrated an immune function in preventing bacterial shedding or infection in vivo (Ikemori et al., 1992; Peralta et al., 1994; Imberechts et al., 1997; Yokoyama et al., 1998 a,b; Marquardt et al., 1999). Antibody MATERIALS AND METHODS Bacteria and Culture Conditions Bacteria were cultured to use for the immunization of chickens. Salmonella enteritidis (ESO 9325-92) and S. 2002 Poultry Science Association, Inc. Received for publication December 1, 2000. Accepted for publication November 30, 2001. 1 To whom correspondence should be addressed: jsim@afns. ualberta.ca. Abbreviation Key: TSB = tryptic soy broth; WSF = water-soluble fraction. 632 BINDING ACTIVITY OF IGY AGAINST SALMONELLA typhimurium (ATCC 14028) were obtained from the Animal Health Laboratories Branch, Alberta Agriculture, Food and Rural Development, Edmonton, Alberta, Canada. Bacteria were cultured in tryptic soy broth (TSB)2 and harvested by centrifugation. The formalin-inactivated cells were lyophilized and stored at −20 C until used. Immunization of Chickens Chickens were immunized to obtain Salmonella-specific IgY loaded eggs. All chickens were cared for in accordance with the Canadian Council on Animal Care guidelines of animal welfare. Immunization of hens was carried out as described (Sunwoo et al., 1996). Lyophilized S. enteritidis (500 µg of cell/mL; 3 µg of protein/mL) and S. typhimurium (500 µg of cell/mL; 4.5 µg of protein/mL) whole cells were emulsified with an equal volume of Freund’s incomplete adjuvant3 to immunize eight 23-wkold Single Comb White Leghorn chickens intramuscularly. Booster immunizations were given at 2 wk and 4 wk after the initial immunization in the same manner. Eggs were collected daily and stored at 4 C until used. Isolation of Water-Soluble Fraction Containing IgY from Egg Yolk The water-soluble fraction (WSF) was isolated from egg yolk to utilize for the analysis of IgY properties. The WSF was prepared using the water dilution method developed by Akita and Nakai (1992). The egg yolk separated from egg white was first mixed gently with eight volumes of cold distilled water (acidified with 0.1 N HCl to a pH of 4.0) and then with cold, acidified, distilled water (pH 2.0) to make a final dilution of 1:10. After mixing well, the mixture was adjusted to pH 5.0∼5.2 and incubated at 4 C for 12 h. The WSF was obtained by centrifugation at 3,125 × g at 4 C for 20 min and then stored at −20 C until analyzed. The WSF containing specific IgY with high activity levels, determined by the following ELISA or non-specific IgY, was neutralized with 0.1 N NaOH and lyophilized. The IgY powder was analyzed for concentrations of protein, total IgY, and specific IgY. ELISA The prepared WSF and IgY powder were assayed by an ELISA procedure as described by Sunwoo et al. (1996, 2000) with modifications. Specific Activity of IgY. The specific binding activity of IgY against whole bacterial cells was tested as follows. A microtiter plate4 was coated with 150 µL of lyophilized S. enteritidis (1.67 mg of cells/mL; 10 µg of protein/mL) 2 Difco Laboratories, Detroit, MI 48232. Sigma Chemical Co., St. Louis, MO 63103. 4 COSTAR威, Corning Incorporated, Corning, NY 14831. 5 Vmax威, Molecular Devices Co., Sunnyvale, CA 94089-1136. 3 633 or S. typhimurium (1.11 mg of cells/mL; 10 µg of protein/ mL) whole cells in carbonate-bicarbonate buffer (0.05 M, pH 9.6). The WSF (diluted 1:1,000 in PBS, 150 µL per well) containing specific egg yolk antibodies (IgY) or nonspecific IgY as a control was reacted with coated antigens. The same volume (150 µL) of rabbit anti-chicken IgG conjugated with horseradish peroxidase3 (diluted 1:1,000 in PBS) and freshly prepared substrate solution, 2-2′azino-bis (3-ethylbenzthiazoline-6-sulfonic acid)3 in 0.05 M phosphate citrate buffer (pH 5.0) containing 30% hydrogen peroxide were used for secondary antibody and substrate, respectively. Absorbance of the mixture was read at 405 nm by a kinetic microplate reader.5 Cross-Reactivity of IgY. The cross-reactivity of IgY was determined by using the above ELISA and the following bacterial cells: S. enteritidis, S. typhimurium, Escherichia coli O157:H7 and E. coli 987P. Wells of the microtiter plate were coated with 150 µL of lyophilized whole cells in carbonate-bicarbonate buffer at the following concentrations: S. enteritidis, 1.67 mg/mL; S. typhimurium, 1.67 mg/ mL; E. coli O157:H7, 0.31 mg/mL; and E. coli 987P, 0.5 mg/mL. IgY powder specific for S. enteritidis (40 to 10 µg/mL) or S. typhimurium (64 to 16 µg/mL) was serially diluted with PBS and added (150 µL per well) to react with coated antigens. The cross-reactivity of anti-S. enteritidis or anti-S. typhimurium IgY against selected bacteria was determined by comparing activities against those bacteria with activity against S. enteritidis or S. typhimurium, respectively. Total IgY Concentration. To measure the total IgY concentration of the WSF or IgY powder, ELISA was performed as described above, except the microtiter plate was coated with 150 µL of rabbit anti-chicken IgG3 at a final concentration of 3.75 µg/mL. Samples of the WSF were diluted 1:90,000 with PBS. Specific or nonspecific IgY powder was reconstituted and serially diluted with PBS (2 to 0.125 µg/mL). Twofold serial dilutions of purified chicken IgG3 (1 mg/mL) in PBS (0.5 to 0.031 µg/ mL) were used as the reference antibodies to prepare a standard curve. The standard curves were then compared to provide a relative measurement of total IgY concentration. Specific IgY Concentration. The concentrations of S. enteritidis- or S. typhimurium-specific IgY were measured by ELISA as described by Sunwoo et al. (2000). Wells of a microtiter plate were coated with 150 µL of rabbit antichicken IgG3 (3.75 µg/mL) and lyophilized S. enteritidis (1.67 mg/mL) or S. typhimurium (1.67 mg/mL) whole cells in carbonate-bicarbonate buffer (0.05 M, pH 9.6). Twofold serial dilutions of reconstituted specific (12.5 to 1.56 µg/mL) and nonspecific (4.5 to 0.28 mg/mL) IgY powder in PBS were added to wells (150 µL per well) coated with S. enteritidis or S. typhimurium whole cells. Wells coated with rabbit anti-chicken IgG were filled with two-fold serial dilutions of purified chicken IgG3 (1 mg/ mL) in PBS (0.5 to 0.008 µg/mL). Secondary antibody, substrate and the measurement of absorbance, were identical as described in a previous section, Specific Activity of IgY. The optical density at 405 nm was converted to 634 LEE ET AL. micrograms of specific IgY per milligram of IgY powder by using a quantitative standard curve determined by the titration between rabbit anti-chicken IgG and purified chicken IgG. Protein Assay The Bio-Rad protein assay6 (Microtiter Plate Protocol), based on the method of Bradford (1976), was performed using purified chicken IgG3 (1 mg of protein/mL) as the reference protein. Growth Inhibition Assay This assay was conducted to investigate whether the binding activity of anti-Salmonella IgY could inhibit Salmonella growth in a liquid medium. The same strain of S. enteritidis or S. typhimurium used as an antigen for immunizing chickens was subcultured on blood agar plates (5% defibrinated sheep blood in Columbia Agar)7 and suspended in TSB. The suspension was adjusted to an optical density of 0.05 at 600 nm, corresponding to a cell density of approximately 2.7 × 107 cfu/mL. The same volume of 20% (vol/vol) glycerol in TSB was added to the suspension, which was then stored at −70 C until used. Two milliliters of prepared bacterial culture were mixed with 2 mL of TSB and incubated at 37 C with shaking. The turbidity of the culture (optical density at 600 nm) was measured by a spectrophotometer8 at 1-h intervals. The growth curve was plotted until the stationary phase was reached. Specific or nonspecific IgY powder was reconstituted to 90, 180, and 360 mg/mL with TSB. IgY solution was then centrifuged at 1,500 × g at 4 C for 20 min. The supernatant was taken and sterilized by using a 0.22-µm membrane filter.9 Two milliliters of specific or nonspecific IgY solution were then added to the same volume of prepared S. enteritidis or S. typhimurium culture. The bacteria and IgY mixtures were incubated at 37 C with shaking. Aliquots of samples (100 µL) were taken at 0, 2, 4, and 6 h of incubation. Plate counts were performed by the spread plate method on TSB agar plates2 in duplicate. The inoculated plates were incubated at 37 C overnight. The number of colony-forming units per plate was counted to determine the total number of bacteria colony-forming units per mL of sample. Microscopic Analyses The specific binding activity of Salmonella-specific IgY against Salmonella was further evaluated by microscopic observation. Immunofluorescence and immunoelectron microscopy were carried out to visualize Salmonella bound by IgY. Immunofluorescence Microscopy. One hundred microliters of S. enteritidis (3.8 × 102 cfu/mL) or S. typhimurium (1.5 × 104 cfu/mL) cells suspended in PBS were incubated with the same volume of specific IgY or nonspecific IgY (100 µg of IgY powder/mL PBS) or without IgY at 37 C for 1 h. After washing with PBS two times, fluorescein isothiocyanate-conjugated rabbit anti-chicken IgG3 diluted 1:250 in PBS was added and then incubated at 37 C for 1 h. The samples were washed as before and resuspended in 50 µL of PBS. Cell suspension (10 µL) was smeared on the microscope slide, which was then air-dried and a coverslip was mounted by using a drop of mounting buffer (Glycerol-PBS, pH 7.2). Immunofluorescent staining of specimens was detected using a 2001 confocal laser scanning microscope.10 Immunoelectron Microscopy. One milliliter of S. enteritidis (3.8 × 102 cfu/mL) or S. typhimurium (1.5 × 104 cfu/mL) cells suspended in PBS was centrifuged at 12,800 × g for 10 min. One milliliter of specific IgY or nonspecific IgY (100 µg of IgY powder/mL of 1% BSA in PBS) was added to the cell pellets. After incubation at 37 C for 1 h, samples were washed with 1% BSA in PBS, two times, before adding 100 µL of rabbit anti-chicken IgG3 (diluted 1:14 in 1% BSA in PBS), followed by incubation at 37 C for 1 h. After being washed, samples were incubated with 300 µl of goat anti-rabbit IgG gold conjugate3 (diluted 1:25 with 1% BSA in PBS). The suspended cells were used for negative staining and ultrathin sectioning. For negative staining, bacterial cells were washed with distilled water two times and subsequently mounted on a 300 mesh copper grid. Grid-mounted samples were stained with 2% uranyl acetate. After being washed and dried, specimens were observed with a transmission electron microscope.11 For the ultrathin section method, bacteria treated with antibodies were washed with 1% BSA in PBS two times, fixed with 2.5% glutaraldehyde for 1 h, and postfixed with 1% osmium tetroxide for 1 h. The fixed samples were dehydrated in a graded series of ethanol and embedded in Spurr’s medium (Spurr, 1969). After infiltration with Spurr’s medium, polymerization was accomplished at 70 C for 12 h. The specimens were then thin sectioned with an ultramicrotome.12 Ultrathin sections were mounted on a 200-mesh copper grid and stained with 2% uranyl acetate and then with lead citrate. The specimens were examined with a transmission electron microscope.11 Statistical Analyses 6 Bio-Rad Laboratories, Hercules, CA 94547. Oxoid, Basingstoke, Hampshire, RG248PW UK. 8 Spectronic 20, Bausch and Lomb, Rochester, NY 14604-2701. 9 Millipore Co., Bedford, MA 01730. 10 Molecular Dynamics, Sunnyvale, CA 94085-4520. 11 Hitachi H-7,000 TEM, Tokyo, 105-8717Japan. 12 Ultracut E model, Reichert-Jung, Austria. 7 The results were analyzed by analysis of variance. When testing the significance of the differences between the experimental and control group, Student’s t-test was used. Data are presented as means ± standard deviation. A probability level of P < 0.05 was considered statistically significant. 635 BINDING ACTIVITY OF IGY AGAINST SALMONELLA FIGURE 2. The change of specific activity of IgY in the egg yolk from chickens immunized with Salmonella enteritidis whole cells or Salmonella typhimurium whole cells during the immunization period. The level of specific activity in a 1,000-fold dilution of the water-soluble fraction was measured by ELISA using bacterial whole cells as an antigen and expressed as an ELISA value [optical density (OD) at 405 nm]. Values are the mean of quadruple samples. Vertical bars indicate the standard deviation. Arrows indicate the week of immunization. In this study, the WSF containing anti-S. enteritidis IgY or anti-S. typhimurium IgY was obtained with a purity (total IgY in protein) of 21.5 or 21.4%, respectively. The recovery of IgY might exist in the range of 92 to 96%, based on results of Akita and Nakai (1993), who used the same conditions (10-fold dilution to a final pH of 5.0 to 5.2) to obtain the WSF. Consequently, IgY present in the WSF could be produced in large quantities by using the water dilution method characterized as simple and economical. FIGURE 1. The concentrations of protein and total IgY in the watersoluble fraction obtained from chickens immunized with (a) Salmonella enteritidis; (b) Salmonella typhimurium whole cells during the immunization period. Values are the mean of quadruple samples. Vertical bars indicate the standard deviation. RESULTS AND DISCUSSION Concentrations of Protein and Total IgY in the WSF The total IgY concentration (Figure 1) was relatively constant (P > 0.05) during the immunization period, and the total average IgY concentration was 7.08 ± 1.24 and 7.09 ± 1.40 mg/mL for anti-S. enteritidis IgY and anti-S. typhimurium IgY, respectively. There was no difference between those two values (P > 0.05), which indicates that the total IgY concentration in the egg yolk is independent of the type of antigens used to raise antibodies as shown by Sunwoo et al. (1996). Specific Activities of IgY Against Salmonella Antigens As shown in Figure 2, the level of specific activities of IgY increased 1 wk after the initial immunization and then rose constantly. The lag time of 1 wk can be explained by the time it takes for specific antibodies produced in chicken serum to be transferred and accumulated in egg yolk as reported by Li et al. (1998). The activity of anti-S. enteritidis IgY against S. enteritidis reached a peak (optical TABLE 1. Concentrations of protein, total IgY and specific IgY in IgY powder prepared from the water-soluble fraction containing Salmonella-specific or nonspecific IgY. Values are the mean of quadruple samples ± SD Concentration (mg/g) IgY powder Anti-S. enteritidis Specific IgY Anti-S. typhimurium Specific IgY Nonspecific IgY Protein Total IgY 590 ± 38 129 ± 10 605 ± 41 468 ± 62 140 ± 35 93 ± 21 Specific IgY 20 ± 7 14 ± 6 0.14 ± 0.037 636 LEE ET AL. FIGURE 3. The cross-reactivity of (a) anti-S. enteritidis IgY; (b) antiS. typhimurium IgY with other members of enterobacteriaceae, including Escherichia coli O157:H7 and E. coli 987P. Values are the mean of triplicate samples. Vertical bars indicate the standard deviation. Asterisks denote values that differ (P < 0.05) from E. coli O157:H7 and E. coli 987P. density of 1.10) at 8 wk and declined thereafter. AntiS. typhimurium IgY activity against S. typhimurium also showed the same pattern as that of anti-S. enteritidis IgY. The level of activity increased to a maximum optical density of 0.62 at 7 wk and then decreased. The overall activity of anti-S. typhimurium IgY was lower (P < 0.05) than that of anti-S. enteritidis IgY. This difference may be attributed to the prior immune status of chickens or inactivated S. typhimurium as a weak immunogen. Chickens immunized in this study were not tested for previous exposure to S. typhimurium or to bacteria carrying cross-reacting antigens with S. typhimurium. Another reason for comparatively low activity of anti-S. typhimurium IgY could possibly be suggested from findings that inactivated S. typhimurium is less immunogenic than live S. typhimurium and thus inactivated S. typhimurium immunogens do not stimulate a sufficient immune response to eliminate those bacteria in chickens (Germanier, 1972; Barrow et al., 1990; Hassan et al., 1991). However, the value of anti-S. typhimurium IgY activity may not be low, considering S. typhimurium whole cells as crude antigens in comparison to the degree of activity of IgY against purified S. typhimurium lipopolysaccharide (LPS) shown in Sunwoo et al. (1996). FIGURE 4. The effect of IgY on the growth of Salmonella in a liquid medium. Bacteria (approximately 1 × 107 cfu/mL) were grown in tryptic soy broth mixed with Salmonella-specific or nonspecific IgY powder at 37 C with shaking: (a) growth curves of S. enteritidis or S. typhimurium; (b) growth of S. enteritidis incubated with 0.9 mg of specific IgY powder/ mL; (c) growth of S. typhimurium incubated with 0.63 mg of specific IgY powder/mL. Viable cells were counted by the plate count method. Values are the mean of triplicate samples. Vertical bars indicate the standard deviation. Asterisks denote values that differ (P < 0.05) from nonspecific control IgY. It is thus likely that both Salmonella-antigens used in the form of inactivated bacterial whole cells are immunogenic enough to induce an immune response and produce Salmonella-specific antibodies in chickens. The immunogenicity of an antigen is influenced by several factors, including the species or strain being immunized, antigen properties and dosage, or the route of administration and BINDING ACTIVITY OF IGY AGAINST SALMONELLA 637 adjuvant (Kuby, 1997). In this study, the production of Salmonella-specific antibody could be efficiently elicited in Single Comb White Leghorn chickens immunized intramuscularly with S. enteritidis or S. typhimurium whole cells emulsified with Freund’s incomplete adjuvant. As a consequence, IgY in the WSF was assessed to possess specific binding activity against Salmonella whole cells and could be used for further study. Properties of IgY Powder The specific IgY concentration of total IgY was 16% and 10% for S. enteritidis-specific IgY powder and S. typhimurium-specific IgY powder, respectively (Table 1). The specific IgY concentration in nonspecific IgY powder, which was prepared as a control from nonimmunized chicken egg yolk, was significantly lower (P < 0.05) than that of specific-IgY powder as expected. The purity (total IgY in protein) of S. enteritidis-specific IgY powder and S. typhimurium-specific IgY powder was 21.9 and 23.1%, respectively. Cross-Reactivity of IgY The anti-S. enteritidis IgY cross-reacted 55.3% with S. typhimurium in contrast to significantly low cross-reactivity (P < 0.05) of anti-S. enteritidis IgY with E. coli O157:H7 or E. coli 987P as shown in Figure 3a. Anti-S. typhimurium IgY also showed low cross-reactivity with both E. coli strains (P < 0.05), although it did cross-react 42.4% with S. enteritidis (Figure 3b). The high cross-reactivity of anti-Salmonella IgY between S. enteritidis and S. typhimurium compared to E. coli strains can be explained by the fact that both Salmonella spp. share somatic antigens (O:1 and O:12) and common epitopes on the different flagellin (Le Minor, 1984; van Zijderveld et al., 1992). Anti-Salmonella IgY used in this study was a polyclonal antibody against bacterial whole cells, and, thus, it may be possible to raise antibodies against crossreacting antigens. The cross-reactivity of IgY can add more value to antibacterial properties of IgY in that IgY might have an antibacterial effect on bacteria with crossreacting antigen as well as target bacteria. Growth Inhibitory Effect of Anti-Salmonella IgY The growth curves of S. enteritidis and S. typhimurium showed similar patterns that consisted of lag (0 to 2 h of incubation time), exponential (2 to 6 h of incubation time), and stationary phase (Figure 4a). Accordingly, S. enteritidis or S. typhimurium was incubated with IgY for 6 h during which samples were taken at 2-h intervals to perform the growth inhibition assay. In this assay, different concentrations of specific or nonspecific IgY were used while considering the importance of antibody quantities to effectively interact with antigens. The growth of S. enteritidis incubated with specific IgY showed a significant reduction in bacterial growth after FIGURE 5. Immunofluorescence micrographs of S. enteritidis incubated with (a) specific IgY; (b) nonspecific IgY; and (c) without IgY (magnification 400 ×). 638 LEE ET AL. FIGURE 6. Immunofluorescence micrographs of S. typhimurium incubated with (a) specific IgY; (b) nonspecific IgY; and (c) without IgY (magnification 400 ×). 4 h incubation (P < 0.05). However, control IgY had no effect on bacterial growth, which maintained a lag phase and exponential phase from 0 to 2 h and 2 to 6 h of incubation, respectively (Figure 4b). Cell counts of the specific and control groups increased by 0.3 log cfu/mL and 1.2 log cfu/mL, respectively, during 4 to 6 h of incubation, indicating that bacteria in the specific treatment group proliferated four times less than the control group. The difference in bacterial growth between the two groups implied that specific IgY has an inhibitory effect on the growth of S. enteritidis. The effective concentration of S. enteritidis-specific IgY in the inhibition of bacterial growth was 0.9 mg/mL, which was determined by specific IgY concentration of IgY powder (20 mg/g) without considering loss of specific IgY during the preparation of IgY solution. The growth patterns of S. typhimurium incubated with S. typhimurium-specific or nonspecific IgY (control) were similar to those observed in the experiment of S. enteritidis with IgY as shown in Figure 4c. There was no difference in bacterial growth between the two groups during 4 h of incubation (P > 0.05). However, the degree of bacterial growth when incubated with specific IgY decreased 16fold compared to that of the control group. Consequently, cell counts of the specific group were reduced by 1.6 log cfu/mL in comparison to those of the control group at 6 h of incubation time. The concentration of specific IgY that showed a growth-inhibitory effect was 0.63 mg/mL. As a result, both Salmonella-specific IgY were found to inhibit the growth of homologous Salmonella in a liquid medium. The mechanism by which antibodies can suppress bacterial growth is not clearly understood. The agglutination of bacterial cells cross-linked by antibodies, which lead to bacterial cells with less motility and opportunity to take nutrients and proliferate than a free-motile single bacterial cell, may not be one of the causes of bacterial growth inhibition. There have been reports that bacterial growth is inhibited by antibodies in solid medium where bacteria cannot aggregate, demonstrating that the reduction of bacterial colony counts mediated by antibodies is not attributed to the agglutination reaction (Feldmann et al., 1992; Sadziene et al., 1992). In addition, Kubo et al. (1973) reported that the agglutinating property of IgY can be displayed only at raised salt concentrations or low pH conditions due to the steric hindrance caused by so closely aligned Fab arms of IgY. Therefore, there may be other reactions between IgY and bacteria to cause the growth inhibition of bacteria than the agglutination reaction. The specific binding of IgY to bacteria appears to be involved in bacterial growth inhibition. Particular components expressed on the bacterial surface, which are crucial factors for the bacterial growth, may be recognized and bound by related polyclonal antibody, IgY. This binding may block or impair the function of growth-related bacterial components and lead to bacterial growth inhibition. Outer membrane protein, lipopolysaccharide, fimbriae (or pili), and flagella may be included in these bacterial surface components (Sim et al., 2000). Therefore, the specific binding activity BINDING ACTIVITY OF IGY AGAINST SALMONELLA 639 FIGURE 7. Immunoelectron micrographs of S. enteritidis incubated with specific IgY (specific group) or nonspecific IgY (control group): negatively stained specimen of (a) specific group (magnification 6,000 ×), (b) control group (6,300 ×); ultrathin sectioned specimen of (c) specific group (9,000 ×), (d) control group (5,100 ×). of IgY, as a potential candidate for the major antibacterial property, requires more intensive studies to uncover the mechanism of inhibition. Microscopic Observation of IgY Binding to Salmonella Figure 5 shows immunofluorescence micrographs, in which fluorescence was observed in S. enteritidis incubated with S. enteritidis-specific IgY; however, there was no fluorescence in the control groups. The presence of fluorescence implies that IgY binds specifically to bacteria, demonstrating the specific binding activity of IgY against the bacteria. A positive result was also obtained from the experiment of S. typhimurium with IgY. S. typhimurium-specific IgY was shown to possess specific binding activity against S. typhimurium as indicated by fluorescence in Figure 6a. The immunofluorescence microscopy performed could be a preliminary test to confirm the specific binding activity of IgY and to further evaluate its activity through immunoelectron microscopy. Figure 7 shows immunoelectron micrographs of S. enteritidis incubated with S. enteritidis-specific IgY or nonspecific IgY. Immunogold particles were observed around bacteria incubated with specific IgY (Figures 7a and 7c), in contrast to the observation of bacteria incubated with nonspecific IgY (Figures 7b and 7d). Bacteria labeled with immunogold indicated that S. enteritidis were bound by specific IgY, which substantiates the specific binding property of IgY against S. enteritidis. The experiment of S. typhimurium with IgY also resulted in the presence of immunogold particles around bacteria incubated with S. typhimurium-specific IgY as shown in Figures 8a and 8c. S. typhimurium-specific IgY was verified to have specific binding activity against S. typhimurium as well. Furthermore, immunoelectron microscopic observation revealed the distribution of immunogold particles around the bacterial surface. It was also found that the 640 LEE ET AL. FIGURE 8. Immunoelectron micrographs of S. typhimurium incubated with specific IgY (specific group) or non-specific IgY (control group): Negatively stained specimen of (a) specific group (magnification 9,000 ×), (b) control group (9,000 ×); Ultrathin sectioned specimen of (c) specific group (6,000 ×), (d) control group (6,000 ×). bacterial surface was structurally altered as presented in Figures 7c and 8c. Bacterial surface bound by IgY was rough in contrast to the smooth surface of control bacteria. These findings can be indicative that specific IgY is attached to components exposed on the bacterial surface, resulting in the structural alterations of the bacterial surface. One of the possible causes leading to the inhibition of bacterial growth is the reaction between bacterial surface components and IgY raised against those components as explained previously. This assumption may be corroborated on the basis of this morphological change of bacteria with bound IgY. Further studies remain to be carried out to define which bacterial surface components are bound by specific IgY and how those binding activities of IgY result in the growth inhibition of bacteria. In conclusion, the binding activity of chicken egg yolk antibody (IgY) against S. enteritidis or S. typhimurium resulted in inhibiting bacterial growth in vitro. Microscopic observation revealed the structural alterations of Salmonella surface bound by IgY. In vitro IgY studies may suggest that IgY binds to Salmonella surface molecules, which are critical for bacterial growth, and leads to the functional impairment of those components. ACKNOWLEDGMENTS Support for this research was provided in part by the Canada-Alberta Hog Industry Development Fund, the Poultry Industry Council of Canada and NLRI international cooperative research fund from Korea. The authors thank all staff in the Poultry Research Center, University of Alberta, for the technical services and assistance in the care of animal used in this research. REFERENCES Akita, E. M., and S. Nakai. 1992. Immunoglobulins from egg yolk: Isolation and purification. J. Food Sci. 57:629–634. BINDING ACTIVITY OF IGY AGAINST SALMONELLA Akita, E. M., and S. Nakai. 1993. Comparison of four purification methods for the production of immunoglobulins from eggs laid by hens immunized with an enterotoxigenic E. coli strain. J. Immunol. Methods 160:207–214. Bar-Joseph, M., and M. Malkinson. 1980. Hen egg yolk as a source of antiviral antibodies in the enzyme linked immunosorbent assay (ELISA): A comparison of two plant viruses. J. Virol. Methods 1:179–183. Barrow, P. A., T. O. Hassan, and A. Berchieri. 1990. Reduction in fecal excretion of Salmonella typhimurium strain F98 in chickens vaccinated with live and killed S. typhimurium organisms. Epidemiol. Infect. 194:413–426. Bradford, M. M. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248–254. Centers for Disease Control 1990. Salmonella surveillance. Annual Summary 1988. CDC, Atlanta, GA. Feldmann, R. C., B. Henrich, V. Kolb-Bachofen, and U. Hadding. 1992. Decreased metabolism and viability of Mycoplasma hominis induced by monoclonal antibody-mediated agglutination. Infect. Immun. 60:166-174. Germanier, R. 1972. Immunity in experimental salmonellosis. III. Comparative immunization with viable and heat-inactivated cells of Salmonella typhimurium. Infect. Immun. 5:792–797. Hassan, J. O., A. P. A Mockett, D. Catty, and P. A. Barrow. 1991. Infection and reinfection of chickens with Salmonella typhimurium: Bacteriology and immune responses. Avian Dis. 35:809–819. Higgins, D. A. 1975. Physical and chemical properties of fowl immunoglobulins. Vet. Bull. 45:139–154. Ikemori,Y., M. Kuroki, R. C. Peralta, H. Yokoyama, and Y. Kodama. 1992. Protection of neonatal calves against fatal enteric colibacillosis by administration of egg yolk powder from hens immunized with K99-piliated enterotoxigenic Escherichia coli. Am. J. Vet. Res. 53: 2005–2008. Imberechts, H., P. Deprez, E. Van Driessche, and P. Pohl. 1997. Chicken egg yolk antibodies against F18ab fimbriae of Escherichia coli inhibit shedding of F18 positive E. coli by experimentally infected pigs. Vet. Microbiol. 54:329–341. Khakhria, R., D. W. Woodward, M. Johnson, and C. Poppe. 1997. Salmonella isolated from humans, animals and other sources in Canada. 1983–92. Epidemiol. Infect. 119:15–23. Kobayashi, K., and H. Hirai. 1980. Studies on subunit components of chicken polymeric immunoglobulins. J. Immunol. 124:1695–1704. Kubo, R. T, B. Zimmerman, and H. M. Grey. 1973. Phylogeny of immunoglobulins. Pages 417–477 in The Antigens. Vol.1 M. Sela, ed. Academic Press, San Diego, CA. Kuby, J. 1997. Immunology. 3rd ed. J. Kuby, ed. W. H. Freeman and Company, New York. Kwan, L., E. Li-Chan, N. Helbig, and S. Nakai. 1991. Fractionation of water-soluble and -insoluble components from egg yolk with minimum use of organic solvents. J. Food Sci. 56:1537–1541. Le Minor, L. 1984. Bergery’s Manual of Systematic Bacteriology. 1st ed. Williams and Wilkins, Baltimore, MD. Lesile, G.A., and L. W. Clem. 1969. Phylogeny of immunoglobulin structure and function III. Immunoglobulins of the chicken. J. Exp. Med. 130:1337–1352. Li, X., T. Nakano, H. H. Sunwoo, B. H. Paek, H. S. Chae, and J. S. Sim. 1998. Effects of egg and yolk weights on yolk antibody (IgY) production in laying chickens. Poult. Sci. 77:266–270. Losch, Y., I. Schranner, R. Wanke, and L. Jurgens. 1986. The chicken egg, an antibody source. J. Vet. Med. B33:609–619. Marquardt R. R., L. Z. Jin, J. W. Kim, L. Fang, A. A. Frohlich, and S. K. Baidoo. 1999. Passive protective effect of egg-yolk antibodies against enterotoxigenic Escherichia coli K88+ infec- 641 tion in neonatal and early-weaned piglets. FEMS Immunol. Med. Microbiol. 23:283–288. O’Brien, J. D. P. 1988. Salmonella enteritidis infection in broiler chickens. Vet. Rec. 122:214. Otani, H., K. Matsumoto, A. Saeki, and A. Hosono. 1991. Comparative studies on properties of hen egg yolk IgY and rabbit serum IgG antibodies. Lebensm. Wiss. Techonol. 24:152–158. Peralta, R.C., H. Yokoyama, Y. Ikemori, M. Kuroki, and Y. Kodama. 1994. Passive immunization against experimental salmonellosis in mice by orally administered hen egg yolk antibodies specific for 14-kDa fimbriae of Salmonella enteritidis. J. Med. Microbiol. 41:29–35. Roberts, T. A., A. C. Baird-Parker, and R. B. Tompkin. 1996. Salmonellae. Pages 217–264 in Microorganisms in Foods 5. Characteristics of microbial pathogens. Blackie Academic and Professional, London. Sadziene, A., P. A. Rosa, P. A. Thompson, D. M. Hogan, and A. G. Barbour. 1992. Antibody-resistant mutants of Borrelia burgdorferi: In vitro selection and characterization. J. Exp. Med. 176:799–809. SAS Institute. 1991. SAS威 User’s Guide: Statistics. Version 6.04 Edition. SAS Institute, Cary, NC. Shimizu, M., H. Nagashima, K. Hashimoto, and T. Suzuki. 1994. Egg yolk antibody (IgY) stability in aqueous solution with high sugar concentrations. J. Food Sci. 59:763–765. Shimizu, M., H. Nagashima, K. Sano, K. Hashimoto, M. Ozeki, K. Tsuda, and H. Hatta. 1992. Molecular stability of chicken and rabbit immunoglobulin G. Biosci. Biotech. Biochem. 56:270–274. Shimizu, M., R. C. Fitzsimmons, and S. Nakai. 1988. Anti-E. coli immunoglobulin Y isolated from egg yolk of immunized chickens as a potential food ingredient. J. Food Sci. 53:1360–1366. Sim, J. S., H. H. Sunwoo, and E. N. Lee. 2000. Ovoglobulin IgY. Pages 227–252 in Natural Food Antimicrobial Systems. A. S. Naidu, ed. CRC Press, Boca Raton, FL. Spurr, A. R. 1969. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastructure Res. 26:31–43. Sunwoo, H. H., X. Li, E. N. Lee, Y. K. Kim, and J. S. Sim. 2000. Preparation of antigen-specific IgY for food application. Pages 311–322 in Egg Nutrition and Biotechnology. J. S. Sim, S. Nakai., and W. Guenter, ed. CAB International, New York. Sunwoo, H.H., T. Nakano, W. T. Dixon, and J. S. Sim. 1996. Immune responses in chickens against lipopolysaccharide of Escherichia coli and Salmonella typhimurium. Poult. Sci. 75:342–345. van Zijderveld, F. G., A. M. van Zijderveld, and J. Anakotta. 1992. Comparison of four different enzyme-linked immunosorbent assays for the serological diagnosis of Salmonella enteritidis infections in chickens. J. Clin. Microbiol. 30:2560– 2566. Vieira, J. G. H., M. A. D. Oliveira, E. M. K. Russo, R. M. B. Maciel, and A. B. Pereira. 1984. Egg yolk as a source of antibodies for human parathyroid hormone (hPTH) radioimmunoassay. J. Immunoassay 5:121–129. Yokoyama, H., K. Umeda., R. C. Peralta., T. Hashi., F. C. Icatlo., M. Kuroki., Y. Ikemori., and Y. Kodama. 1998a. Oral passive immunization against experimental salmonellosis in mice using chicken egg yolk antibodies specific for Salmonella enteritidis and S. typhimurium. Vaccine16:388–393. Yokoyama, H., R. C. Peralta, K. Umeda, T. Hashi, F. C. Icatlo, M. Kuroki, Y. Ikemori, and Y. Kodama. 1998b. Prevention of fatal salmonellosis in neonatal calves, using orally administered chicken egg yolk Salmonella-specific antibodies. Am. J. Vet. Res. 59:416–420. Yolken, R.H., F. Leister, S. E. Wee, R. Miskuff, and S. Vonderfecht. 1988. Antibodies to rotaviruses in chickens’eggs: A potential source of antiviral immunoglobulins suitable for human consumption. Pediatrics 81:291–295.