Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Inclusive fitness in humans wikipedia , lookup
Homo floresiensis wikipedia , lookup
Criticism of evolutionary psychology wikipedia , lookup
Recent African origin of modern humans wikipedia , lookup
The Descent of Man, and Selection in Relation to Sex wikipedia , lookup
Darwinian literary studies wikipedia , lookup
Sociobiology wikipedia , lookup
Evolutionary psychology wikipedia , lookup
Before the Dawn (book) wikipedia , lookup
Anatomically modern human wikipedia , lookup
Behavioral modernity wikipedia , lookup
Adaptive evolution in the human genome wikipedia , lookup
History of anthropometry wikipedia , lookup
Discovery of human antiquity wikipedia , lookup
Craniometry wikipedia , lookup
Origins of society wikipedia , lookup
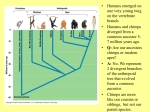
LENScience Senior Biology Seminar Series Walking Upright: The cost of human evolution Jacquie Bay, Alan Beedle, Tatjana Buklijas, Peter Gluckman Human evolution is the process by which over time, humans have changed to optimise the fit between the individual members of a population and their environment. In contrast to the modern human desire to live a long life, evolution is directed primarily by the biological drive to reproduce and pass genes on to successive generations. Only lineages that can successfully reproduce will survive. The process of evolution is driven by the interaction of humans with their environment. The sum of the interactions between the individual and all biotic and abiotic components of their environment will determine the success of the individual in passing on genes and therefore contributing to the survival of the population and features of future generations. These interactions are vast and complex. They present significant variables, including the complexity of intra and inter‐specific interactions playing out in response to biotic and abiotic factors, which when combined with the variation present within the individuals within the population, determine survival. In biological terms, humans sit with the great apes within the hominoid group but are distinguished within this group by specific physical and behavioural traits including bipedalism, brain size, hairlessness, tool making, language development, abstract thought, social organisation, and culture. Current molecular evidence suggests that the pathway to modern humans can be traced back along the hominoid lineage with the first split being the Orang‐utan lineage around 15 million years ago, with the last branch that leads to Homo sapiens splitting from apes 5‐6 million years ago (Fig 1). These dates are based on evidence from the molecular clock which assumes a consistent rate of mutational change over time. The evolution of Hominins (humans and their direct ancestors) has been noted for the development of large brains (leading to intelligence) and relatively long lives. Bipedalism is one of several factors selected for that have resulted collectively in the development of intelligence and the success of the hominins. However, the adaptive advantages that these selected features bring are accompanied by adaptive costs. Collectively adaptive advantage must outweigh adaptive cost for the evolutionary success of the species. However evolutionary fitness (and the adaptations selected) does not necessarily match health and longevity. Fig 1: Evolutionary relationship between the Hominoids. Dates are in millions of years before the present. 1 Box 1: The concept of FITNESS is central to understanding evolution. Evolutionary fitness is a measure of the match between an individual and its environment to best enable successful reproduction. It is important to note that fitness is measured as the ability to reproduce and live long enough to support offspring to the point where they reach reproductive age. Selection is based on adaptations that contribute to the success of reproduction. Health and longevity beyond reproductive years are not generally drivers in evolutionary selection processes. Adaptive Radiation Adaptive radiation is an evolutionary pattern whereby (relatively) rapid diversification within a lineage can be traced from a common ancestral origin, resulting in the existence of multiple related genera and species which have evolved to occupy multiple niches present within an environment. Both the primates and the hominins display this pattern of evolutionary development. While exact relationships continue to be debated within the hominin group, it is clear that this group existed from around 6 million years ago, and included multiple related species, some of which may have co‐existed (Fig 2). Fig 2: Hominin Evolution Adaptive radiation leading to speciation is driven either by species entering new ecosystems, or changes in environment which create different selection pressures and therefore the potential to alter the phenotype that is selected for and survives. Each species thrives in a niche for which it is best adapted, with divergence reducing competition between species. It is likely that the adaptive radiation seen in the hominins species was driven by environmental change, perhaps reflecting climate change. Fig 3: Evidence suggests that climate change events around 4‐5 Million Years Ago led to a change in habitat in Eastern Africa, from Wooded Forest to Wooded Savannah. This change created more open spaces, changing potential food sources and potentially creating a driver for bipedalism. 2 Selection for evolutionary success The evolutionary success of the hominin group can be linked to selection that led to the development of intelligence. This is not the result of selection relating to one characteristic. Rather it is the selection for a combination of characteristics which collectively contribute to the development of intelligence, and the accompanying evolutionary success. Bipedalism, tool making, increased brain size, and the development of culture are key contributors to the success of the human species. Bipedalism Evidence from the fossil record suggests that some bipedal tendencies were present as far back as Orrorin tugenesis (6MYA). There is some possibility that O. tugenesis was not in fact in the hominin line, suggesting either that bipedalism evolved in more than one taxa or that bipedalism had started to develop before the split between the last common ancestor of apes and humans. The well known female Australopithecus afarensis skeleton, “Lucy” (Fig 4) is dated 3 million years ago and is believed to have been habitually bipedal, adapted for bipedal walking, but not able to run efficiently. A. afarensis existed from 4‐3MYA so Lucy was a relatively late example of this species. Evidence from earlier fossils of A. afarensis show that some arboreal behaviours were also part of the niche of A. afarensis, suggesting that this was the period of gradual evolution from quadrupedal to bipedal locomotion. Selection for bipedalism in this period must have offered survival advantages to the species, which are likely to have resulted from changes in the environment which made this form of locomotion preferential. Evidence suggests that selection for bipedalism was driven by environmental change seen in eastern Africa around 4MYA in which near continuous forest changed to wooded savannah, creating more open habitats in which food resources became more dispersed (Fig 3). In such habitats, bipedal movement is energetically more efficient than quadripedal movement, particularly at walking speeds. Calculations of the net costs of transport for different gaits (Fig 5) show that human walking is more efficient than most other forms of locomotion in animals (Sockol et al 2007). The paleoanthropologist Robert Foley has calculated that, provided the early hominid spent the majority of its time in terrestrial (as opposed to arboreal) foraging, then bipedalism would be adaptive in such environments. Thus the evolution of bipedalism can be seen to have been progressively adaptive as the hominin line evolved, driven by habitat change. Fig 4: Australopithecus afarensis, cast from Museum national d'histoire naturelle, Paris. Image source Wikicommons; Creative Commons license. Fig 5: Comparison of the net cost of transport (ml of O2 kg−1 m−1) for chimpanzee quadrupedal walking (blue), chimpanzee bipedal walking (red), and human walking (yellow). Dashed lines indicate trendlines for running and walking in birds and mammals. The running trendline is for 65 terrestrial species. Walking data (open symbols) were collected from the literature. Sockol M. D et al PNAS 2007; 104: 12265—12269 3 Other arguments that have been put forward for the origin of bipedalism include : improved thermoregulatory efficiency, leading to the ability to forage through the heat of the day improved ability to see and therefore avoid predators improved ability to carry food, tools, and infants during their period of dependence These adaptations are likely to be secondary to the drive for selection based on energetic efficiency, and could have arisen from the initial adaptation of standing upright and adopting a bipedal gait. While bipedalism was selected for and therefore had adaptive advantage, it also had adaptive costs that resulted from the structural changes that are associated with the upright posture and bipedal gait, as shown in the table 1 below. Table 1: Adaptations for Bipedalism—Advantage and Cost Adaptation Advantage The head is positioned over the top of Foramen Magnum has moved from the rear to the base of the skull the spine, balanced for upright stance. The neck muscles are able to be reduced in size as they have less of a role in holding the head upright. The development of an S‐shaped spine with a forward curve in the cervical and lumbar regions and backward curve in the thoracic region Cost Back pain, particularly in The S‐shaped spine places the centre the lumbar region; slipped of gravity directly over the feet which discs. provide the supporting platform. This Hernia—resulting from the fact that the gut is no improves stability and balance. longer hanging, as in the quadrupeds. The pelvis shape has changed to become Greater stability for upright stance. A reduced birth canal short, broad, & bowl shaped. leading to problems in child The weight of the body is transmitted Gluteal muscles birth. This problem is directly to the legs. are strengthened, compounded by the providing support increase in brain size & preventing tilt associated with human when walking. evolution. The femur is angled inwards, keeping the knee and foot directly under the centre of gravity. The outer condyle of the knee (where the femur meets the knee) is increased in size to cope with this angle change and stop sideways deflection of the muscles The lower limbs have lengthened, enabling a faster, longer stride. The ability to extend the knee joint to Increased risk of fracture of The foot forms a platform. The big toe is aligned (non‐ opposable). The foot is arched. The ability to grasp with the foot is lost. The aligned big toe provides the a straight position for upright stance. neck of the femur This also allows the leg to swing (particularly in old age as through, enabling a heel to toe strike bone density decreases). Varicose veins resulting action of the foot. from the increase difficulty The knee is brought in line with the in return flow of blood head of the femur and centre of the foot when walking—improving weight from the lower limbs brought about by upright bearing capacity. stance. ability to walk with a forward thrust. The arch offers shock absorption. 4 The weight of the body is resting on the feet and can cause fallen arches, distorted bones & pain on walking. Tools, brain expansion & social development Bipedalism provided the potential for a number of significant behavioural adaptations and innovations that contributed to the evolution of modern Homo sapiens and the relative success of the species. Tool culture, brain expansion and social development are amongst these adaptations. The earliest evidence of the use of tools to modify or manipulate objects and elements of the environment is seen in Homo habilis. This is the same time that we observe an increase in brain size in both absolute terms and relative to body size. Australopithecines had absolute and relative brain sizes not greatly different from those of the modern apes. In comparison, the brain size of the Homo lineage showed an exponential increase from H.habilis onwards (Fig 6). Log Brain Volume (cm3) Brain Volume Relative to Body Size H. sapiens Hominids H. erectus H. habilis Australopithecines A.boisei A.afarensis Gorilla Orang Great Apes Chimp Log body weight (kg) Fig 6. Change in brain volume relative to body size, calculated from fossil skull dimensions. Redrawn from Bonner J.T., Why Size Matters. Princeton University Press 2006 Brain size is not inherently linked to evolutionary success. The Neanderthal brain had a larger volume than the modern human brain, yet the species was not as successful. Indeed, brains are energetically expensive and there are many large animals with highly successful evolutionary histories yet relatively small brains (witness the dinosaur prior to the asteroid‐induced extinction). Thus the question of why hominins evolved large brains and Homo sapiens is endowed with its unique set of capacities is not self‐evident. There are several inter‐related theories on the evolutionary origin of brain expansion. Effectively they either place emphasis on Homo species finding adaptive advantage in social interactions within their group, or in planning their affairs for hunting and tool making. The weight of evidence now favours the former. The Expensive Brain Humans have a relatively large brain compared to body mass. This increase in brain size evolved rapidly from the Homo erectus onwards. Evolution of the large brain came at a high metabolic cost. The brain requires a higher level of energy to sustain activity than any other tissue in the body. Humans expend around 20% of their energy intake on the brain compared to 8‐10% in most other primates and 3‐5% in non‐primate mammals. Humans have evolved to eat an energy and nutrient rich diet which supports the high level of metabolic need of the brain. Fossil evidence suggests that major changes in brain size seen in Homo erectus coincides with the evolution of hunting and gathering communities. These communities consumed a higher proportion of animal foods than ever seen before among primates, had home bases to which they transported foods and were in the habit of sharing food within a community group. These improvements in diet quality are thought to be important in the development of the large brain in relation to body size (Leonard et al 2007). 5 Adaptive advantage from social interactions There is a close relationship between social group size, brain size and levels of intentionality which can be achieved (Box 1). Essentially, fitness is advanced by maintaining stable social alliances that promote potential mating opportunities. But if this strategy is to be successful, then an ‘evolutionary ratchet’ may be created where societal pressures build, requiring even greater intellectual complexity to maintain reproductive opportunity. Thus a series of feed‐forward loops may be created in which rising social complexity requires greater intelligence. In turn this allows development of more sophisticated technology, more complex ways of living, and more sophisticated social structures. These in turn generate the need for even greater higher cognitive function. Communication becomes a critical component of such a feed‐forward system, and the development of language was clearly permitted and expedited by this increasing complexity of the brain. Individuals living in a group must have the capacity to consider the implications of their actions on the group or members of the group, and to interpret the intentions of others. The development of language and the communication of abstract thoughts allowed this ability to develop. This possibly gave rise to art, music, science and religion. Box 2 Theory of mind ‐ Orders of Intentionality This theory describes how members of a species interact when interpreting each others’ behaviour and thoughts. The basis of theory of mind is a hierarchy of orders of intentionality. 1st order: 2nd order: 3rd order: 4th order: 5th order: I am aware of my own thoughts I think I know what you are thinking I think I know what you are thinking about me I think I know what you are thinking about what I am thinking ....and I think I know what will happen if you do not respond in the way I would like you to respond..... And this can continue, serving as the structure of adult human interactions. The more political our intent, the higher the levels of intentionality that we need to employ. Most adults can reach 5‐6 orders of intentionality before we become confused. Observational evidence suggests that some non‐human primates operate to the 3rd order of intentionality, practising deception in relation to sexual matters and sometimes food access. Humans today interact socially with multiple individuals within multiple communities. Just think about the number of groups you interact with on a daily basis; family, school, work, friends from sports clubs, church groups, cultural groups, beebo, facebook, twitter friends. Technology means we now have multiple interacts with people who we do not see face to face. Fig 7: Anthropologist Robin Dunbar from the University of Oxford has suggested that living in larger groups had an evolutionary advantage for our ancestors. The larger group size offered security of food supply and increased protection, Dunbar has also shown that living in large groups requires intelligence. Fig 7 shows that group size in primates is correlated positively to brain size. In particular the noecortex, a part of the brain associated with higher order functions such as sensory perception, motor control, spatial reasoning, conscious thought and in humans, language is larger in comparison to the rest of the brain in species that live in larger groups. Based on correlations with other primates such as chimpanzees who live in groups of 50, Dunbar predicted that humans have evolved to live in groups of about 150 individuals . 6 The reproductive cost of brain expansion The development of the large brain had other consequences for hominins. Most significant are the problems associated with giving birth to a baby with a large head through the narrow and rigid birth canal that arose as a result of the pelvic changes associated with bipedalism. If the human infant was born at the same stage of maturity as other primates, then pregnancy would last about 21 months: this would require a pelvic canal so wide that it would be impractical for efficient bipedal locomotion. The compromise has been that humans give birth to an infant at a stage when the head can fit through the birth canal, but this means that the human infant is entirely dependent on its mother for many months after birth. This need to give birth to a totally dependent infant has determined human social structure—if the mother is to support the infant she in turn needs to be confident of support from the father. Thus while early hominids showed great sexual dimorphism, with the males much larger than the females, implying a harem type mating system with fighting between males for mating rights, Homo sapiens has a much lesser degree of sexual dimorphism, suggesting that in general females were able to have continued support from one male. The potential impact of life history on evolution and our understanding of evolution Life history is the name given to the patterns of growth, development, reproduction and mortality typically seen within a species. An organism should be viewed as having a strategy of growth, reproduction and longevity which has been adaptively shaped by natural selection. Understanding human life history is essential in understanding the evolution of the human life course. Human life history is characterised by typically a singleton pregnancy, relatively high levels of fat at birth (Box 3), a long phase of postnatal nutritional dependency, a prolonged juvenile (childhood) period, delayed sexual maturity, only modest sexual dimorphism, and a long life span. Females lose the ability for reproduction before the end of their intrinsic lifespan and males are capable of reproduction into old age. Humans produce very few offspring, who benefit from high parental investment, ensuring a comparatively high rate of survival to adulthood. The slow rate of senescence (Box 4) in humans means that life spans in excess of 70 years are not exceptional. Some of these human life history traits are unusual, most notably the long childhood phase, the pubertal growth spurt, and menopause. Box 3: Why are human babies so fat? Human babies are born with higher levels of fat than any other species. Traditionally this was explained in relation to our hairlessness. It was assumed that natural selection compensated for our loss of fur with a layer of insulative blubber. A newer perspective notes that this excess adipose tissue is well‐suited to serve as a backup energy supply for another distinctive human trait – our large brains. Brains have among the highest metabolic rates of any tissue or organ in the body, and they are quickly damaged in the event of even temporary disruption in energy supply. Humans have exceptionally large brains, especially early in life. Roughly 70‐80% of the body’s metabolism is devoted to this costly organ in the newborn. The developmental pattern of high fat levels seen in humans is likely to have arisen to cope with the very high demands of the proportionally large infant brain. Fig 8: Proportion of resting metabolic rate (RMR) allocated to the brain during human growth and development. Brain metabolism represents >60% of RMR in infancy, and about 20% in adulthood (from Leonard WR, et al. Comp Biochem Physiol A Mol Integr Physiol 2003;136:5‐15). Box 4: Aging vs Senescence Aging is the simple fact of time – getting older. Senescence is a biological process (or processes) in which, following a phase of development and attainment of maximal reproductive potential, there is an age‐associated physiological degeneration which adversely affects vitality and function and leads to an increased risk of death. Senescence is a universal phenomenon of sexually reproductive species and is associated with a number of biochemical and cellular changes. 7 Life history and “fitness” Life history traits that species “manipulate” to achieve optimal fitness include: the number, size and sex ratio of offspring; size and maturity at birth; pattern of growth; age and size at maturity; parental investment in offspring; and the investment in repair and maintenance affecting intrinsic mortality and lifespan. There is an extraordinary variation in the solutions that different species of mammal have evolved, each providing an adaptive solution to maintaining gene flow within their usual environment(s). Despite this variation, there are trade‐offs that constrain the possible solutions such as number vs quality of offspring; current vs future reproduction; age vs size at maturity; fecundity (total lifetime reproductive performance) vs lifespan. These life history traits are being selected for to improve fitness — that is the ability of the species to reproduce and pass on inherited characteristics. Selection for reproductive success, and therefore evolutionary success, does not equate to selection for health and longevity. There will be evolutionary “trade offs” which may be advantageous to reproductive fitness but not for health and longevity. Humans have evolved to have a small number of offspring with a very high chance of survival. This is a trade off — number of offspring vs likelihood of survival. There is a greater chance that more will survive if there are fewer to care for. The delayed onset of puberty and sexual maturity that has been selected for in humans is a trade off between investing energy in continued growth vs investing energy in reproduction. By delaying reproductive maturity, there is greater opportunity for growth and through this an increased chance of reproductive success. The longevity of an individual will be affected by their environment. Human longevity is greatly influenced by our cultural evolution, as we have developed technologies to deal with many extrinsic causes of mortality, particularly in recent decades through advances in public health and medicine. Thus while our rate of senescence has not changed over the last few millennia, the average lifespan has increased. Human Evolution—trade offs and costs The evolution of humans has involved complex interactions between pre‐existing variation, environmental conditions, life history, and selection. No one factor has dominated the resultant shape of modern Homo sapiens, however we can identify some key factors and tradeoffs. Bipedalism is a significant adaptation that led to selection relating to increased brain size, cultural evolution, and the development of intelligence. However, each of these groups of adaptations has involved costs and tradeoffs. Overall, the collective advantage from the adaptations outweigh the costs to the individual and therefore promote the adaptive success of the species. Humans are still evolving Arguments suggesting that the human species is no longer evolving are centred on the belief that our cultural and technological advances have insulated our biology from the selective pressures of the environment. Those supporting the view that human evolution has slowed would argue that only calamitous events such as a pandemic would create a situation where selection would occur. However the persistence of sickle cell anaemia in African nations where the heterozygous condition incurs protection from Malaria is evidence of the continuing power of selection. Human populations have traditionally been small and isolated. Modern humans exist in very large and mobile human populations, meaning that a larger number of potentially adaptive mutations will occur in each generation, providing an increased level of variation for selection to work on. Biological evolution cannot be separated from cultural evolution. Cultural trends, population demographics and the differing use of technologies in human cultural groups will all play a role in the continuing evolution of the human species. References and further reading: Gluckman, P.D., Beedle, A.S., Hanson, M.A. (2009) Principles of Evolutionary Medicine Oxford University Press Sockol M. D et al Chimpanzee locomotor energetics and the origin of human bipedalism PNAS 2007; 104: 12265—12269 Bonner J.T., Why Size Matters. Princeton University Press 2006 Princeton, NJ. Leonard WR, et al. 2003. Metabolic correlates of hominid brain evolution. Comp Biochem Physiol A Mol Integr Physiol 136: 5‐15. Dunbar, R.I.M (2008) Why humans aren't just great apes British Academy Review 11, 15‐17. Leonard, W. R., Snodgrass, J. J. andRobertson, M. L., (2007) Effects of Brain Evolution on Human Nutrition and Metabolism Annu. Rev. Nutr., 2007, 27, 311–328. For further information please contact: Jacquie Bay [email protected] | Alan Beedle [email protected] | Tatjana Buklijas [email protected] LENScience Bringing Schools and Scientists Together Copyright © Liggins Institute 2009 http://LENS.auckland.ac.nz 8