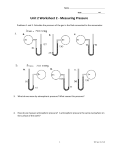
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Heart failure wikipedia , lookup
Electrocardiography wikipedia , lookup
Coronary artery disease wikipedia , lookup
Jatene procedure wikipedia , lookup
Myocardial infarction wikipedia , lookup
Cardiac surgery wikipedia , lookup
Antihypertensive drug wikipedia , lookup
Dextro-Transposition of the great arteries wikipedia , lookup
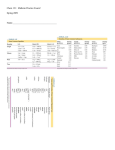
Clinical Science (2001) 100, 303–309 (Printed in Great Britain) Attenuated cardiac baroreflex in men with presyncope evoked by lower body negative pressure Duminda N. WIJEYSUNDERA, Gary C. BUTLER, Shin-ichi ANDO, Michael J. POLLARD, Peter PICTON and John S. FLORAS Division of Cardiology of the Toronto General and Mount Sinai Hospitals and the University of Toronto, Suite 1614, 600 University Avenue, Toronto, Ontario, Canada M5G 1X5 A B S T R A C T Mechanisms responsible for presyncope during lower body negative pressure (LBNP) in otherwise healthy subjects are poorly understood. Muscle sympathetic nerve activity (MSNA), blood pressure, heart rate (HR), HR power spectra, central venous pressure (CVP) and stroke volume were determined in 14 healthy men subjected to incremental LBNP. Of these, seven experienced presyncope at LBNP k15 mmHg. Subjects who tolerated LBNP k15 mmHg had significantly lower CVP (2n6p1n0 versus 7n2p1n2 mmHg ; meanspS.E.M., P 0n02), HR (59p2 versus 66p3 beats/min, P 0n05) and MSNA burst frequency (29n0p2n4 versus 39n0p3n5 bursts/min, P 0n05) during supine rest. LBNP at k15 mmHg had no effect on blood pressure, but caused similar and significant reductions in stroke volume and cardiac output in both groups. Subjects who tolerated LBNP had significant reflex increases in HR, MSNA burst frequency and burst amplitude with LBNP of k15 mmHg. These responses were absent in those who experienced presyncope. The gain of the cardiac baroreflex regulation of MSNA was markedly attenuated in pre-syncopal subjects (1n2p0n6 versus 8n8p1n4 bursts/100 heart beats per mmHg ; P 0n001). Healthy subjects who experience presyncope in response to LBNP appear more dependent, when supine, upon MSNA to maintain preload, and less able to increase sympathetic vasoconstrictor discharge to skeletal muscle reflexively in response to orthostatic stimuli. INTRODUCTION The cause of vasodepressor syncope in otherwise healthy adults remains obscure. Much attention has been directed at neural events, often sudden, occurring at the time blood pressure begins to fall [1–4] and at physical or pharmacological manoeuvres that might unmask any underlying predisposition to faint [5,6]. A common theme in such experiments has been the selection of patients with a prior history of recurrent syncope, presumably due to a neurally mediated mechanism. During the course of our investigation of sympathetic nervous system responses to lower body negative pressure (LBNP) in healthy, middle-aged men, symptoms or signs of presyncope at levels greater than k15 mmHg LBNP resulted in premature termination of this stimulus in 50 % of these subjects. Because none of these individuals had a history of prior syncope, and several were engaged in occupations requiring a high degree of alertness, this finding was unanticipated. Our purpose in conducting the present analysis was to compare the haemodynamic and sympathoneural characteristics of those who became pre-syncopal, as opposed to those who did not, under two conditions : at rest, prior to the application of LBNP ; and in response to LBNP at k15 mmHg, which was the stimulus that all subjects were able to tolerate without developing symptoms. Differences between these two groups, both at rest and in Key words : baroreceptor reflex, central venous pressure, heart rate variability, microneurography, sympathetic nervous system. Abbreviations : LBNP, lower body negative pressure ; MSNA, muscle sympathetic nerve activity ; CVP, central venous pressure ; HR, heart rate. Correspondence : Dr John S. Floras (e-mail john.floras!utoronto.ca). # 2001 The Biochemical Society and the Medical Research Society 303 304 D. N. Wijeysundera and others response to this simulated orthostatic stimulus, raise hypotheses concerning neural aspects of circulatory regulation of individuals at risk of presyncope, which merit prospective testing in future investigations. METHODS Subjects Healthy, active, middle-aged men (n l 14, 44n5p1n7 years ; meanpS.E.M.) with no prior history of syncope were studied. Their mean weight was 81n4p3n5 kg and their height 180n4p1n4 cm. None were taking any medication. All subjects provided informed written consent as approved by our Institutional Human Subjects Review Committee, in accordance with the Declaration of Helsinki. Procedures Subjects emptied their bladder, then were placed supine into a chamber custom-built for recording muscle sympathetic nerve activity (MSNA) from the right peroneal nerve during LBNP [7]. An airtight seal was obtained by a neoprene kayak skirt fitted at the iliac crests. The right leg was held in position by form-fitting supports at the thigh, knee and ankle, and caudal displacement of the subject was prevented by bilateral foot plates. Before any additional recording instruments were applied, a brief trial of LBNP was performed so as to familiarize subjects with the sensation of lower body suction and thereby minimize any inadvertent contraction of leg muscles during the protocol. A volume-clamp cuff (Finapres model 2300 ; Ohmeda, Rexdale, Canada) was placed on the left-middle finger for continuous non-invasive beat-by-beat recording of blood pressure. After local anaesthesia, a central venous catheter was introduced into an antecubital vein of the right arm and advanced to an intrathoracic position [7]. A respiratory belt was secured around the abdomen. Central venous pressure (CVP) and respiratory excursions were measured continuously by Statham P23ID pressure transducers (Gould Inc., Cleveland, OH, U.S.A.) and recorded simultaneously with lead II of the electrocardiogram, heart rate (HR) and the MSNA neurogram by an ink recorder on to paper. Post-ganglionic MSNA was recorded from the right peroneal nerve [7,8]. Stroke volume was calculated over approx. 10 consecutive cardiac cycles from two-dimensional echocardiographic and continuous-wave Doppler recordings, using methods reported previously by our group [7,9]. Cardiac output was determined from HR and stroke volume. Protocol Supine bed rest (20 min) was followed by a 10 min baseline period. LBNP was then applied at k5, k15 k30 and k40 mmHg. If tolerated, each level of LBNP # 2001 The Biochemical Society and the Medical Research Society was sustained for 10 min. Blood pressure, HR, CVP and MSNA were recorded continuously. Stroke volume was derived over the last 2 min of baseline, each level of LBNP and the last 2 min of the recovery period. Subjects were monitored for any sensation or indication of presyncope, such as pallor, tachypnea, yawning, a drop in systolic blood pressure 30 mmHg, or heart rate 20 beats\min. If these arose, LBNP was stopped and subjects were instructed to begin mild leg muscle contractions to facilitate venous return. Data analysis Muscle sympathetic nerve activity Pulse synchronous bursts of MSNA were identified by inspection of the mean voltage neurogram [7] by two trained, but blinded, assessors. Any disagreement with respect to burst identification was resolved by the senior investigator. MSNA was expressed as burst frequency (bursts\min), incidence (bursts\100 cardiac cycles) and units of integrated nerve activity (the product of burst frequency and mean burst amplitude in mm). The latter serves as a quantitative representation of the strength of each burst of post-ganglionic discharge. Spectral analysis of heart rate variability A total of 7 min of the ECG (from the second to the eighth minute of each level of LBNP) for each experimental condition was analysed to determine HR variability, using coarse-graining spectral analysis. The specific details of this technique have been reported previously [10–12]. Total spectral power (PT) was divided into its fractal power (PF), low-frequency harmonic (0n0– 0n15 Hz ; PL) and high-frequency harmonic (0n15– 0n50 Hz ; PH) components, with total harmonic power (PHP) comprising the sum of PL and PH. The ratio PH\PT was used to estimate the contribution of parasympathetic modulation of HR to total spectral power [13–16], and ratio PL\PH was applied to estimate the contribution of sympathetic nervous system modulation of HR, or the ‘ sympathovagal balance ’, to power spectral density [8,13,15–20]. The non-harmonic component of spectral power was characterized further by plotting the log of spectral power as a function of the log of frequency (a 1\fβ plot), with β the slope of this linear regression [10,11,21]. Statistical analysis Mean valuespS.E.M. are reported throughout. Subjects were divided on the basis of their response to LBNP into pre-syncopal and non-pre-syncopal groups. The principal comparison in this analysis was between baseline measurements and responses to k15 mmHg LBNP, which was the highest level tolerated by all subjects. Within-group responses were assessed, applying Stu- Attenuated cardiac reflex preceding presyncope dent’s paired t test to normally distributed data and the Wilcoxon rank sum test for non-parametric data, whereas Student’s unpaired t test and the Mann Whitney rank sum test were used for between-group comparisons. Statistical significance was accepted if P 0n05. RESULTS All subjects completed the k5 and k15 mmHg LBNP segments of the protocol, but three developed presyncope at k30 mmHg and four at k40 mmHg. Characteristics, at baseline, of subjects who completed the full protocol and subjects who developed presyncope appear in Table 1. A continuous measure of CVP was obtained in 12 subjects, six in each group. There was no significant difference in mean age, blood pressure, stroke volume or cardiac output between Table 1 subjects who completed the full protocol and those who experienced presyncope. However, the latter group was characterized by significantly higher sympathetic burst frequency, heart rate and CVP (Table 1). A comparison of baseline power spectral analysis data did not detect any difference in the initial indices for heart rate variability in these two groups (Table 2). Five of these subjects had participated in previous research studies and were familiar with these procedures. Of these, three developed presyncope and two did not. Responses to k15 mmHg LBNP This level of LBNP did not significantly affect systolic or diastolic blood pressure in either group. As anticipated, there were significant reductions in CVP, stroke volume and cardiac output in both groups. LBNP at k15 mmHg Haemodynamics and muscle sympathetic nerve activity in subjects without and with presyncope n l 7 for each group, an l 6 for each group ; meanspS.E.M. *P §P 0n005 for within-group differences. 0n05 ; **P 0n02 for differences between groups at baseline. †P Without presyncope 0n05 ; ‡P 0n01 ; With presyncope Variable 0 mmHg k15 mmHg 0 mmHg k15 mmHg Systolic blood pressure (mmHg) Diastolic blood pressure (mmHg) Pulse pressure (mmHg) CVP (mmHg)a Stroke volume (ml) Heart rate (beats/min) Cardiac output (L/min) MSNA (bursts/min) MSNA (bursts/100 heart beats) MSNA burst amplitude (mm) MSNA (units/min) MSNA (units/100 heart beats) 125p6 80p4 45p3 2n6p1n0 71p6 59p2 4n1p0n4 29n0p2n4 47n7p3n2 6n8p0n4 201p19 328p36 121p5 76p4 46p3 0n3p0n2† 60p4‡ 64p3‡ 3n8p0n3† 41n5p1n1§ 62n2p3n4§ 10n5p1n4† 441p70† 651p89‡ 119p3 76p2 42p2 7n2p1n2** 67p7 66p3* 4n4p0n5 39n0p3n5* 58n1p5n4 7n7p0n5 302p38* 456p62 115p4 76p2 39p3 3n0p1n2§ 54p5† 69p3 3n8p0n3† 42n0p3n4 65n2p5n6† 8n6p1n2 405p84 589p124 Table 2 R–R interval and heart rate variability in subjects without and with presyncope n l 7 for each group ; meanspS.E.M. *P 0n05, **P 0n01 for within-group comparisons. Without presyncope With presyncope Variable 0 mmHg k15 mmHg 0 mmHg k15 mmHg R–R interval (ms) PT (ms2) PF (ms2) PF/PT PHP (ms2) PH (ms2) PH/PT PL (ms2) PL/PH β 987p67 1493p459 828p243 0n62p0n07 664p278 241p121 0n11p0n03 423p160 3n8p1n3 1n15p0n10 912p51* 1208p188 870p151 0n71p0n03 337p57 72p28 0n06p0n02 266p42 18n7p8n0 1n46p0n10** 896p38 825p163 591p118 0n74p0n05 234p64 61p20 0n07p0n02 175p56 7n0p4n1 1n33p0n10 868p41* 1724p793 1258p566 0n74p0n04 481p228 52p21 0n04p0n01 422p207 22n3p10n8 1n53p0n16 # 2001 The Biochemical Society and the Medical Research Society 305 306 D. N. Wijeysundera and others heart beat increase in MSNA for every 1 mmHg decrease in CVP and a 17n0p19n2 units\100 heart beat increase in MSNA for each mmHg decrease in CVP (P 0n001 and P l 0n027 respectively). DISCUSSION Figure 1 Gain of cardiac reflex control of MSNA Means of ratios between increases in MSNA (response) and decreases in CVP (stimulus) in response to k15 mmHg LBNP in subjects with or without presyncope above k15 mmHg LBNP (n l 7 for each group). Note log scale. lowered CVP in subjects who did not tolerate hypotensive LBNP to the initial level observed during supine rest in the other group (P 0n05 for comparison between groups). At these similar levels of CVP, stroke volume tended to be lower in the pre-syncopal group (55p5 ml versus 71p6 ml ; P l 0n053). Subjects who tolerated hypotensive LBNP (i.e. k15 mmHg) responded to k15 mmHg LBNP, with a significant increase in heart rate, sympathetic burst frequency, incidence, amplitude and integrated nerve activity. In contrast, in those who subsequently experienced presyncope, there were no significant reflexive increases in heart rate or these several representations of MSNA, with the exception of sympathetic burst incidence (see Table 1). LBNP at k15 mmHg did not affect harmonic power spectra in either group. However, there was a significant increase in the slope of the log power–log frequency relationship in the non-presyncopal group (Table 2). Cardiac reflex control of MSNA Because LBNP at k15 mmHg did not have any significant effect on arterial blood pressure, or on PH\PT, the principal representation of parasympathetic withdrawal in response to arterial baroreceptor unloading, the primary analysis of reflex control of MSNA focused on the gain of the cardiac reflex. To estimate this, a ratio relating changes in MSNA to changes in CVP was constructed. Regardless of the representation of MSNA selected, there was a marked attenuation in the calculated gain of this reflex in those subjects who experienced presyncope at higher levels of LBNP (Figure 1). In the group who tolerated LBNP without presyncope, there was an 8n8p1n4 unit increase in burst incidence for every mmHg decrease in CVP and a 248p87 unit incidence increase for every mmHg decrease in CVP. In contrast, in the pre-syncopal group, there was a 1n2p0n6 burst\100 # 2001 The Biochemical Society and the Medical Research Society Our objective in this analysis was to compare the haemodynamic and sympathoneural characteristics of healthy, middle-aged subjects, with no prior history of unexplained syncope, who did or did not experience presyncope in response to graded LBNP. The two groups differed significantly in the following three respects. Subjects who became pre-syncopal : (1) had higher initial CVP, heart rate and MSNA burst frequency, prior to the application of LBNP ; (2) did not increase heart rate, MSNA burst frequency, amplitude, or integrated MSNA in response to k15 mmHg LBNP ; and (3) displayed marked attenuation of the gain of the cardiac baroreflex control of MSNA in response to this stimulus. For similar levels of CVP, stroke volume was substantially lower in subjects who subsequently experienced presyncope. These observations suggest that subjects with a predisposition to presyncope in response to an orthostatic stimulus are more dependent on central sympathetic outflow to skeletal muscle, to maintain preload, and on sympathetic nerve traffic to the sino-atrial node to maintain cardiac output. Venous pooling and withdrawal of preload by graded LBNP would therefore predispose these subjects to syncope unless countered by reflex sympathetic activation. However, these individuals were unable to mount the appropriate additional increase in sympathetic vasoconstrictor discharge in response to this stimulus, presumably due to impairment of their cardiac baroreflex control of MSNA. Right atrial pressure remained higher in the pre-syncopal group throughout LBNP, and there was no abrupt cut-out of MSNA during the course of the experiment, which argues against the ‘ collapse-firing ’ hypothesis of neurogenic syncope. Young subjects with borderline or mild essential hypertension also have increased MSNA burst frequency and incidence, often in the setting of higher central blood volume, heart rate, cardiac output and decreased venous distensibility [22–25]. However, in contrast to the present findings, their reflex forearm vasoconstrictor and MSNA responses to non-hypotensive LBNP are either augmented, or unchanged, when compared with agematched normotensive counterparts [26,27]. Because all were healthy, active individuals with no prior history of fainting, the pre-syncopal response to hypotensive LBNP was unanticipated. This may be a feature unique to this specific protocol, in which, initially, there was greater unloading of low pressure than high pressure mechanoreceptors. In contrast, standing or tilt Attenuated cardiac reflex preceding presyncope unloads simultaneously both low pressure cardiopulmonary and high pressure aortic baroreceptor afferent nerve endings. This redundancy, which provides two independent pathways for reflex increases in efferent sympathetic vasoconstrictor discharge, may be sufficient to protect healthy individuals from presyncope during standing, but not during the stimulus of graded LBNP. For several decades incremental LBNP has been used to isolate reflexes arising from the atria, pulmonary veins and the left ventricle (which sense changes in filling pressures and chamber volumes and elicit reflex changes in sympathetic discharge to skeletal muscle) from reflexes arising from afferent nerve endings situated in the aortic arch and carotid sinus (which sense changes in blood pressure and stroke volume) [28–30]. Both the haemodynamic changes observed in the present protocol, and the absence of any significant drop in parasympathetic heart rate modulation, imply that unloading of these low pressure receptors was the predominant stimulus to sympathetic activation k15 mmHg LBNP. In a retrospective comparison of healthy subjects, Jacobs et al. [31] reported a significant increase in forearm noradrenaline spillover, in response to k15 mmHg in subjects who tolerated k40 mmHg LBNP, whereas there was a non-significant trend to lower forearm noradrenaline spillover in those who experienced presyncope during that stimulus. There were no differences between these two groups, at baseline, in blood pressure, heart rate, plasma beta-endorphin concentrations, arterial catecholamines, total body noradrenaline spillover or forearm noradrenaline spillover into plasma. These authors did not record CVP, and therefore could not quantify cardiac reflex gain. Mosqueda-Garcia et al. [4] performed a detailed study of responses to incremental tilt in three groups of subjects : controls, healthy subjects with no prior history of syncope, but who consistently fainted during tilt, and subjects under investigation for recurrent neurally mediated syncope. They described a progressive reduction, across these three groups, in the gain of the relationship between the absolute change in CVP and the relative (%) increase in MSNA induced by progressive tilting. Reductions in CVP and systemic blood pressure with this stimulus also differed in their time course and magnitude. In those with recurrent neurogenic syncope, they observed attenuation of the MSNA response at low tilt angles, followed by a gradual decline and then an abrupt cut-off. (However, in our experience, it is difficult to discriminate between abrupt reflex cessation of central sympathetic outflow and displacement of the microelectrode by leg movement during negative pressure.) The greatest fall in CVP was observed in the otherwise healthy subjects in whom repeated tilt-testing evoked syncope, i.e. analogous to the response observed in our pre-syncopal subjects. Although, in contrast to the present findings, MSNA increased significantly, in re- sponse to tilt, in this group. However, no statistical comparison between these three calculated gains was performed ; relative, rather than absolute increases in MSNA were plotted on the ordinate ; and the tilt stimulus applied was not specific to cardiac mechanoreceptor unloading. Morillo et al. [2] also examined responses to tilt in subjects with a history of orthostatic syncope. Approx. 40 % of these developed recurrent syncope during this experiment. At baseline, heart rate, blood pressure and MSNA were similar in pre-syncopal and non-syncopal patients, but parasympathetic responses to nitroprussideinduced reductions in blood pressure were attenuated. The arterial baroreflex control of MSNA was not different between these two groups. Since CVP was not determined, these authors could not comment on cardiac reflex gain. Our demonstration of markedly lower cardiac reflex control of MSNA in subjects who experienced presyncope would therefore appear to be a novel finding. This may reflect long-standing differences in the discharge characteristics of the afferent nerve endings, perhaps due to altered compliance of the cardiac chambers or large vessels which house these mechanoreceptors, or differences in the distribution of blood volume [32] between these two groups. If so, this could explain why the significantly higher CVP did not appear to reflexively suppress MSNA under resting conditions. It is also possible that LBNP stimulated the release of sympathoinhibitory peptides, such as vasopressin, atrial natriuretic peptide and endogenous endorphins. Plasma concentrations of these peptides increase markedly prior to syncope, or when orthostatic maneouvers elicit nausea, light-headedness and related symptoms of impending faint [3,31,33]. An inhibitory interaction between such peptides and the sympathetic nervous system, centrally, or at the ganglionic level [7,34–36], could reset the cardiac baroreflex acutely, causing a gradual or abrupt loss of sympathetic vasoconstrictor discharge to skeletal muscle. If these peptidergic responses differed within subjects, from one test stimulus to the next, such variability could explain why subjects differ in their tolerance to repeated orthostatic stress. In a previous report [37], subjects with unexplained syncope who developed recurrent symptoms during tilttesting had a decrease in their PL\PH ratio, whereas this ratio increased in those without symptoms. We therefore explored frequency domain characteristics of heart rate variability. There was no difference in baseline values for this ratio between the two groups, and k15 mmHg LBNP had no effect in either group, supporting our previous conclusion that this proposed power spectral estimate of efferent sympathetic outflow to the sinoatrial node is insensitive to significant between-group differences in the magnitude of sympathetic nervous system activation [8]. # 2001 The Biochemical Society and the Medical Research Society 307 308 D. N. Wijeysundera and others There are several limitations to the present analysis. First, this was a post hoc comparison of subjects who did or did not develop presyncope in response to a protocol of LBNP applied for another purpose, rather than a prospective study of subjects predisposed to recurrent syncope. Secondly, it is not known whether these observations would be reproducible within subjects if this protocol were to be repeated. Thirdly, our observations do not permit us to determine the mechanisms responsible for these baseline differences between groups, their contrasting sympathoneural responses to LBNP and the attenuated cardiac reflex control of MSNA. These may reflect long-standing differences in neurogenic cardiovascular control in such individuals. Alternatively, they may arise from day-to-day variations in sodium balance, and therefore cardiopulmonary and total intravascular blood volume, or reflect an alerting response to the laboratory environment. These differences may also be due to variations between subjects, in the corelease of sympatho-inhibitory peptides and neurotransmitters, or in their actions. Thus the present report should be considered as hypothesis-generating. Future studies of this issue should address questions of reproducibility, predictive value and mechanism. Fourthly, the two groups were well matched with respect to age, height, blood pressure and several other baseline characteristics, but differed with respect to their initial CVPs. However, the higher initial CVP of the pre-syncopal group would suggest that they were operating at the steeper portion of the sigmoid CVP–MSNA stimulus– response relationship, and therefore might be expected to display an augmented, rather than a diminished, sympatho-neural response to the fall in CVP elicited by non-hypotensive LBNP. Fifthly, we did not study concurrently blood volume, or plasma markers of other vasoactive systems involved in salt and water balance. It is possible that these pre-syncopal subjects maintain increased blood volume or redistribute their blood volume centrally as a result of a generalized activation of several neurohormonal mechanisms. Finally, we cannot comment on whether women predisposed to presyncope would display the same baseline differences or attenuated reflex responses to LBNP. In summary, this analysis would suggest that the predisposition to presyncope in responses to graded LBNP is associated with greater dependence on vasoconstriction and venoconstriction in skeletal muscle, to maintain preload under conditions of supine rest, and attenuation of the reflex vasoconstrictor response to preload reduction, subserved by the low pressure cardiopulmonary receptors. This diminished cardiac reflex control of muscle sympathetic nerve activity would then result in an inability to maintain stable blood pressure and cerebral blood flow in the face of sustained displacement of blood volume from the cardiopulmonary circulation. # 2001 The Biochemical Society and the Medical Research Society ACKNOWLEDGMENTS This work was supported by Grants-in-Aid from the Heart and Stroke Foundation of Ontario (T3054 and T4050). J. S. F. holds a Career Investigator Award from the Heart and Stroke Foundation of Ontario. G. C. B. received a Fellowship from the Medical Research Council of Canada. S.A. received a Canadian Hypertension Society\Merck Frosst Fellowship and was supported, in addition, by the George R. Gardiner Foundation (Toronto). We thank Beverley L. Senn, R. N. and Steven C. Brooks, B.Sc. for their technical assistance. REFERENCES 1 Wallin, B. G. and Sundlof, G. (1982) Sympathetic outflow to muscles during vasovagal syncope. J. Auton. Nerv. Syst. 6, 287–291 2 Morillo, C. A., Eckberg, D. L., Ellenbogen, K. A., Beightol, L. A., Hoag, J. B., Tahvanainen, K. U. O., Kuusela, T. A. and Diedrich, A. M. (1997) Vagal and sympathetic mechanisms in patients with orthostatic vasovagal syncope. Circulation 96, 2509–2513 3 Jardine, D. L., Ikram, H., Frampton, C. M., Frethey, R., Bennett, S. I. and Crozier, I. G. (1998) Autonomic control of vasovagal syncope. Am. J. Physiol. 274, H2110–H2115 4 Mosquedo-Garcia, R., Furlan, R., Fernandez-Violante, R., Desai, T., Snell, M., Jarai, Z., Ananthram, V., Robertson, R. M. and Robertson, D. (1999) Sympathetic and baroreceptor reflex function in neurally mediated syncope evoked by tilt. J. Clin. Invest. 99, 2736–2744 5 Waxman, M. B., Yao, L., Cameron, D., Wald, R. W. and Roseman, J. (1989) Isoproterenol induction of vasodepressor-type reaction in vasodepressor-prone persons. Am. J. Cardiol. 63, 58–65 6 Almquist, A., Goldenberg, I. F., Milstein, S., Chen, M. Y., Hansen, R., Gornick, C. C. and Benditt, D. G. (1989) Provocation of bradycardia and hypotension by isoproterenol and upright posture in patients with unexplained syncope. N. Engl. J. Med. 320, 346–351 7 Floras, J. S. (1990) Sympathoinhibitory effects of atrial natriuretic factor in normal humans. Circulation 81, 1860–1873 8 Notarius, C. F., Butler, G. C., Ando, S., Pollard, M. J., Senn, B. L. and Floras, J. S. (1999) Dissociation between microneurographic and heart rate variability estimates of sympathetic tone in normal and heart failure subjects. Clin. Sci. 96, 557–565 9 Naughton, Y., Rahman, M. A., Hara, K., Floras, J. and Bradley, T. D. (1995) Effect of continuous positive airway pressure on intrathoracic and left ventricular transmural pressure in patients with congestive heart failure. Circulation 91, 1725–1731 10 Yamamoto, Y. and Hughson, R. L. (1991) Coarse-graining spectral analysis : new method for studying heart rate variability. J. Appl. Physiol. 71, 1143–1150 11 Butler, G. C., Yamamoto, Y., Xing, H. C., Northey, D. R. and Hughson, R. L. (1993) Heart rate variability and fractal dimension during orthostatic challenges. J. Appl. Physiol. 75, 2602–2612 12 Butler, G. C., Ando, S. and Floras, J. S. (1997) Fractal component of variability of heart rate and systolic blood pressure in congestive heart failure. Clin. Sci. 92, 543–550 13 Pomeranz, B., Macaulay, R. J. B., Caudill, M. A., Kutz, I., Adam, D., Gordon, D., Kilborn, K. M., Barger, A. C., Shannon, D. C., Cohen, R. and Benson, H. (1985) Assessment of autonomic function in humans by heart rate spectral analysis. Am. J. Physiol. 246, H151–H153 Attenuated cardiac reflex preceding presyncope 14 Saul, J. P., Berger, R. D., Chen, M. H. and Cohen, R. J. (1989) Transfer function analysis of autonomic regulation. II. Respiratory sinus arrhythmia. Am. J. Physiol. 256, H153–H161 15 Malliani, A., Pagani, M., Lombardi, F. and Cerutti, S. (1991) Cardiovascular neural regulation explored in the frequency domain. Circulation 84, 482–492 16 Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. (1996) Heart rate variability : standards of measurement, physiological interpretation and clinical use. Circulation 93, 1043–1065 17 Pagani, M., Lombardi, F., Guzetti, S. et al. (1986) Power spectral analysis of heart rate and arterial pressure variabilities as a marker of sympatho-vagal interaction in man and conscious dog. Circ. Res. 59, 178–193 18 Saul, J. P., Rea, R. F., Eckberg, D. L., Berger, R. D. and Cohen, R. J. (1990) Heart rate and muscle sympathetic nerve variability during reflex changes of autonomic activity. Am. J. Physiol. 258, H713–H721 19 Montano, N., Ruscone, T. G., Porta, A., Lombardi, F., Pagani, M. and Malliani, A. (1994) Power spectrum analysis of heart rate variability to assess the changes in sympathovagal balance during graded orthostatic tilt. Circulation 90, 1826–1831 20 van de Borne, P., Montano, N., Zimmerman, B., Pagani, M. and Somers, V. K. (1997) Relationship between repeated measures of hemodynamics, muscle sympathetic nerve activity, and their spectral oscillations. Circulation 96, 4326–4332 21 Nakamura, Y., Yamamoto, Y. and Muraoka, I. (1993) Autonomic control of heart rate during physical exercise and fractal dimension of heart rate variability. J. Appl. Physiol. 74, 875–881 22 Takeshita, A. and Mark, A. L. (1979) Decreased venous distensibility in borderline hypertension. Hypertension 1, 202–206 23 Anderson, E. A., Sinkey, C. A., Lawton, W. J. and Mark, A.L (1989) Elevated sympathetic nerve activity in borderline hypertensive humans : evidence from direct intraneural recordings. Hypertension 14, 177–183 24 Floras, J. S. and Hara, K. (1993) Sympathoneural and haemodynamic characteristics of young subjects with mild essential hypertension. J. Hypertension 11, 647–655 25 Schmeider, R. E., Schobel, H. P. and Messerli, F. H. (1995) Central blood volume : A determinant of early cardiac adaptation in arterial hypertension ? J. Am Coll. Cardiol. 26, 1692–1698 26 Mark, A. L. and Kerber, R. E. (1982) Augmentation of cardiopulmonary baroreflex control of forearm vascular resistance in borderline hypertension. Hypertension 4, 39–46 27 Rea, R. F. and Hamdan, M. (1990) Baroreflex control of muscle sympathetic nerve activity in borderline hypertension. Circulation 82, 856–862 28 Zoller, R. P., Mark, A. L., Abboud, F. M., Schmid, P. G. and Heistad, D. D. (1972) The role of low pressure baroreceptors in reflex vasoconstrictor responses in man. J. Clin. Invest. 51, 2967–2972 29 Johnson, J. M., Rowell, L. B., Niederberger, M. and Eisman, M. M. (1974) Human splanchnic and forearm vasoconstrictor responses to reductions of right atrial and aortic pressures. Circ. Res. 34, 515–524 30 Abboud, F. M., Eckberg, D. L., Johannsen, U. J. and Mark, A. L. (1979) Carotid and cardiopulmonary baroreceptor control of splanchnic and forearm vascular resistance during venous pooling in man. J. Physiol. (Cambridge, U.K.) 286, 173–184 31 Jacobs, M. C., Goldstein, D. S., Willemsen, J. J., Smits, P., Thien, T., Dionne, R. A. and Lenders, J. W. (1995) Neurohumoral antecedents of vasodepressor reactions. Eur. J. Clin. Invest. 25, 754–761 32 Mack, G. W., Convertino, V. A. and Nadel, E. R. (1993) Effect of exercise training on cardiopulmonary baroreflex control of forearm vascular resistance. Med. Sci. Sports Exerc. 25, 722–726 33 Riegger, G. A. and Wagner, A. (1991) Excessive secretion of vasopressin during vasovagal reaction. Am. Heart J. 121, 602–603 34 Floras, J. S., Aylward, P. E., Mark, A. L. and Abboud, F. M. (1987) Inhibition of muscle sympathetic nerve activity in humans by arginine vasopressin. Hypertension 10, 409–416 35 Thoren, P., Floras, J. S., Hoffmann, P. and Seals, D. R. (1990) Endorphins and exercise : Physiological mechanisms and clinical implications. Med. Sci. Sports Exerc. 22, 417–428 36 Floras, J. S. (1995) Inhibitory effect of atrial natriuretic factor on sympathetic ganglionic neurotransmission in humans. Am. J. Physiol. 269, R406–R412 37 Kouakam, C., Lacriox, D., Zghal, N., Logier, R., Klug, D., Le Franc, P., Jarwe, M. and Kacet, S. (1999) Inadequate sympathovagal balance in response to orthostatism in patients with unexplained syncope and a positive head up tilt test. Heart 82, 312–318 Received 3 August 2000/20 October 2000; accepted 24 November 2000 # 2001 The Biochemical Society and the Medical Research Society 309