Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Bioluminescence wikipedia , lookup
Proteolysis wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Cyanobacteria wikipedia , lookup
Community fingerprinting wikipedia , lookup
Butyric acid wikipedia , lookup
Biochemistry wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
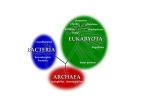
Chapter 2 Microbiology of the Rumen T.G. Nagaraja The Ruminal Ecosystem The rumen, or more appropriately the reticulo-rumen, is a large chamber (50–100 l capacity in adult cattle) in which the ingested feed is first subjected to microbial digestion. The rumen is an ideal microbial habitat because the conditions that exist are conducive for the survival and growth of microorganisms. The temperature remains relatively constant (36–40°). Water the animal drinks and the only exocrine secretion that rumen receives, saliva, provide a moist environment required for microbial growth. Ingested food provides the energy and other nutrients needed for microbial growth and activity. Normal reticulo-ruminal motility (peristalsis and antiperistalsis) helps mix the contents, which brings microbes into contact with fresh substrate. The end products of fermentation are removed by absorption (acids) into the blood or eructation (gases). Absorption coupled with the buffering effect provided by salivary secretions help regulate ruminal pH. The rumen ecosystem, often referred to as a continuous culture system, operates as a biological fermentation unit, like a ‘chemostat’, under well-defined conditions that are extremely important regulators of the types, number and biochemical activities of the microorganisms (Fig. 2.1). There is more or less continuous availability of substrate (at least in grazing cattle), removal of end products (by absorption, eructation or passage), and passage of undigested and waste products. The contents in the rumen are heterogeneous, consisting of complex mass of digesta, which may float (forage) or sediment at the bottom (grain) depending on the density of the feed, liquid fraction with microbial cells and fine feed particles, and a free gas cap in the dorsal sac. T.G. Nagaraja (*) Department of Diagnostic Medicine/Pathobiology, College of Veterinary Medicine, Kansas State University, Manhattan, KS 66506-5606, USA e-mail: [email protected] © Springer International Publishing Switzerland 2016 D.D. Millen et al. (eds.), Rumenology, DOI 10.1007/978-3-319-30533-2_2 39 40 T.G. Nagaraja Fig. 2.1 Rumen as a continuous culture system Anaerobiosis and Its Consequences The ruminal environment is anaerobic with gas phase composed mostly of carbon dioxide (~65 %) and methane (~35 %), although small amounts of other gases (H2, N2, O2, etc) are present. Carbon dioxide is produced by microbial activities and by neutralization of acids by bicarbonate that enters the rumen from saliva and blood. The trace amounts of O2 and N2 are contaminants from air that enters via feed and water. The absence of oxygen makes the rumen highly anaerobic with a redox potential of 150 to −350 mV. Therefore the type of metabolism that takes place in the rumen is fermentation where the final electron acceptor is organic (major) or inorganic (minor) compounds, and not the oxygen. The anaerobic condition imposes two major restrictions. One is the energy yield from substrates (moles of ATP/mole of glucose) is considerably lower (no electron transport phosphorylation) than aerobic metabolism (4 vs. 38). Because of the limited ATP yield there is rapid throughput of substrates to sustain microbial growth. Secondly, the substrates are only partially oxidized, which allows the animal to absorb the products (acetate, propionate and butyrate) as energy source and the reducing equivalents generated during fermentation are disposed by producing reduced products like lactate and propionate, and taken up by methanogens to produce methane. Interactions Between the Host and the Microbes The microorganisms in the rumen and the ruminant animal live in symbiotic relationship. In addition to providing nutrients to the microbes, the host also contributes to the maintenance of physical and chemical conditions for optimal microbial fermentation. In return, microorganisms provide energy, protein, and vitamins to the host (Table 2.1). 2 Microbiology of the Rumen 41 Table 2.1 Symbiotic relationship between the ruminal microorganisms and the host animal Host contributions and their outcome to the microbes Mastication and rumination of the feed breakdown large feed particles into smaller particles (comminution), which increase surface area for microbial attachment and digestion Salivary input to the rumen provides aqueous environment necessary for microbial growth, nutrients (nitrogen), but more importantly supplies the major buffering compounds (bicarbonates and phosphates) essential for regulation of ruminal pH Ruminal contractions (peristalsis and antiperistalsis) help mix the digesta, which brings microbes into contact with fresh substrate, and facilitates passage of digesta to make room for additional feed Removal of fermentation products by eructation (gases) and absorption (acids) are critical for maintaining optimal conditions (pH) for microbial growth Microbial contributions and their outcome to the host Host is absolutely dependent on microbes to digest fiber. Only microbes have fibrolytic enzymes to degrade cellulose and hemicelluloses Microbes can use nonprotein nitrogen (urea, nitrate, nucleic acids) as source of ammonia and synthesize amino acids and protein Production of fermentation products, particularly VFA, which serve as the major source of energy to the host Production of microbial cells which in the lower tract (abomasums and small intestine) serve as the major source of protein and vitamins The Ruminal Microorganisms The rumen is inhabited by a multitude of microorganisms. Many of these organisms colonize and grow and are considered indigenous, and hence are termed “normal flora”, also called autochthonous microbiota. These microbes, for the most part, live in harmony with the host. Additionally, the ruminal flora includes microbiota, referred to as allochthonous, that do not get established (colonization and growth), are dormant and in transit. These are derived largely from ingested food and water and to a small extent from swallowed air or from another habitat of the host (e.g., skin, respiratory tract, or reproductive tract). The nonindigenous microbes also include a variety of gastrointestinal pathogens that may colonize and grow to establish infections. Also, some members of the normal flora could assume pathogenic roles (opportunistic pathogens) when the ecosystem is perturbed in some way or when a breach occurs in the integrity of the ruminal wall. The microbial population in the rumen includes members that belong to all three domains, Eubacteria (Bacteria), Archaea (Methanogens), and Eukarya (Protozoa and Fungi). The rumen ecosystem contains the following types of microorganisms: Bacteria, Protozoa, Fungi, and Bacteriophages (Table 2.2). Ruminal Bacteria Rumen has a dense population of bacteria with numbers ranging from 108–1011 per g of contents. The number is reflective of the digestibility of the feed, which is why bacterial counts in grain-based diets are 10- to 100-fold higher than the forage based T.G. Nagaraja 42 Table 2.2 Types of ruminal microorganisms Types Bacteria Methanogens Protozoa Flagellates Ciliates Fungi Bacteriophages Domain Eubacteria Archaea Eukarya Eukarya – Number (per ml or g of ruminal contents) 109–1011 105–108 102–103 104–106 – 1011–1012 Percentage of total microbial mass 40–90 2–4 0–60 10 – diets. Majority of the bacteria is obligately anaerobic although facultative bacteria do exist. Anaerobic bacterial counts are typically about 1000-fold higher than f acultative bacterial counts. Most of the facultative bacteria in the rumen are nonindigenous and transient population carried into the rumen through feed and water. Ruminal bacteria are predominantly gram-negative accounting for 80–90 % of the population. Certain species of ruminal bacteria (Butyrivibrio fibrisolvens) have typical gram-positive cell wall structure (bilayer with thick peptidoglycan) but stain gram negative. The proportion of gram-positive bacteria increases in grain-fed animals (20–30 % of the total). Morphologically, ruminal bacteria are rod-, spherical- or spiral- shaped organisms, and majority is rod shaped organisms. Spiral shaped organisms, belonging to the genus Treponema, constitute only a small fraction (<1 %) of the bacterial population. Based on the distribution of bacteria in the rumen, they can be categorized into bacteria that float freely in rumen fluid and bacteria that are adherent to feed particles (loosely or tightly), protozoal cells, fungal sporangia, or epithelial cells (Fig. 2.2). The free-floating bacteria constitute a minor component (~30 %) of the total bacterial population. Bacteria associated with feed particles are the major component (70 %) of the total bacterial population. Bacteria attached to ruminal epithelial cells, called epimural bacteria, constitute a small fraction of the total bacterial population in the rumen. Bacteria that float freely or attached to feed particles participate actively in the digestion of feedstuffs. However, epimural bacteria do not make any significant contribution to the ruminal digestion. Many of the epimural bacteria are facultatively anaerobic and produce urease enzyme. Because ruminal wall is highly oxygenated and allow diffusion of urea from the blood into the rumen, it is speculated that the role of epimural bacteria is to maintain anaerobiosis by scavanging any oxygen that otherwise would have diffused into the rumen, and hydrolyze urea. Also, epimural bacteria may digest the cells that get sloughed off from the wall. Historically, much of the information on numbers and kinds of ruminal bacteria has been obtained by cultivation-based methods (Fig. 2.3). Isolation of bacteria in pure culture has enabled identification of the biochemical activities and fermentation products produced. Recently, applications of cultivation-independent techniques, particularly based on 16S rRNA gene sequence analyses, have indicated that the number of bacterial species in the rumen is vastly underestimated. It is generally believed that culture-based procedures have only identified about 10 % of bacterial species present in the rumen. 2 Microbiology of the Rumen 43 Fig. 2.2 Diagrammatic representation of the distribution of bacteria in the rumen Methanogens Methanogens are members of the Archaea domain and phylogenetically differ from bacteria protozoa and fungi. They lack peptidoglycan and have unusual lipid structures. Methanogens constitute about 2–4 % of the bacterial population in the rumen. Methanogens that have been cultured from ruminal contents belong to only few genera and species, although methanogens outside the rumen habitat have been classified into as many as 23 genera and hundreds of species. Methanogens in the rumen that have been cultivated belong to five genera and seven species: Methanobacterium formicium, Methanobacterium bryantii, Methanobrevibacter olleyae, Methanobrevibacter millerae, Methanobrevibacter ruminantium, Methanomicrobium mobile, Methanoculleues olentangyi, and Methanosarcina barkeri. Cultivation-independent methods that have assessed archaeal population in ruminal contents have revealed abundance of a variety of methanogenic genera and species. Based on cultivation independent method (16S rRNA gene sequence analyses), majority of ruminal methanogen belongs to three genus-level groups. They are Methanobrevibacter, Methanomicrobium, and a large group of uncultured methanogens. Ruminal ciliated protozoa harbor methanogens on the outside surface (ectosymbionts) and inside the cell (endosymbionts) based on characteristic fluorescence of cofactors (F350 and F420). Based on 16S rRNA genes analyses, majority of the methanogens associated with ciliated protozoa belong to the same three groups described in ruminal contents. The most prevalent and best characterized methanogen in the rumen is Methanobrevibacter ruminantium. 44 T.G. Nagaraja Fig. 2.3 Classification of ruminal bacteria based on morphology and oxygen relationship Most species of methanogens can grow using H2 and formate as their energy sources and use the electrons derived from H2 and formate to reduce CO2 to form methane (Table 2.3). Some species can use methyl groups from methanol, methylamines, or acetate (Methanosarcina barkeri) to produce methane. However, the amount of acetate used by methanogens is insignificant because the acetate-utilizing methanogens do not survive in the rumen because their growth rates are slower than the rate of passage of contents. 2 Microbiology of the Rumen 45 Table 2.3 Substrates for methanogenesis in the rumen Substrates Major H2 and CO Formate Minor Methanol Methylamine Dimethylamine Trimethylamine Acetate Reactions 4H2 + CO2 4HCOOH CH4 + 2H2O CH4 + 3CO2 + 2H2O 4CH3OH 4CH3NH2Cl + 2H2O 2(CH3)2NHCl + 2H2O 4(CH3)3NHCl + 2H2O CH3COOH 3CH4 + CO2 + 2H2O 3CH4 + CO2 + 4NH4Cl 3CH4 + CO2 + 2NH4Cl 3CH4 + CO2 + 2NH4Cl CH4 + CO2 Ruminal Protozoa Protozoa were the first rumen microorganisms to be discovered (by Gruby and Delafond in 1843), which is not surprising because of their conspicuous cell size (up to 200 μM in length) and active motility. Like bacteria, protozoa in the rumen are anaerobic. Certain species of ciliated protozoa, similar to other anaerobic protozoa like Trichomonads, possess an organelle, called hydrogenosomes, which are implicated in oxygen utilization in the rumen. These structures make protozoa somewhat aerotolerant and are also responsible for the role ciliated protozoa play in scavenging of oxygen to maintain anaerobiosis. Protozoa in the rumen are broadly classified into flagellates and ciliates, depending on whether they have flagella or cilia. Flagella are larger in length and thickness compared to cilia and function in motility. Cilia are small and thin, more numerous and function in motility and also aid in ingestion of food. Flagellates are smaller in size (3–12 μM), fewer in number (102–103 per ml), and utilize only soluble nutrients; therefore, their contribution to overall ruminal fermentation is insignificant. Ciliated Protozoa. Ciliated protozoa make up the majority of protozoa in the rumen and range in size from 10 × 20 to 120 × 200 μM. Ciliated protozoa are highly specialized, single cells and have structures and features similar to animals. A single cell is bounded by a pellicle or skin and has internal structures that can be described as digestive tract (mouth, cytopharynx, anus, etc.), urinary tract (contractile vacuoles), skeletal structures (skeletal plates), etc. They are binucleated with one being large (macronucleus) and the other being small (micronucleus). Based on morphological features (ciliary arrangement, location of the macronucleus, and absence, presence, and size and number of skeletal plates), ciliates are grouped under different genera and species. There are two broad groups of ciliated protozoa that differ in morphological features (Fig. 2.4a and b) and functional roles (Table 2.4). The ‘holotrichs’, belonging to the order Trichostomatida, have cilia covering the entire or almost the entire surface. The group contains two of the moist common genera, Isotricha and Dasytricha. The entodiniomorphid ciliates, belonging to the order Entodiniomorphida, have cilia at the anterior end and may or may T.G. Nagaraja 46 Fig. 2.4 (a) A diagrammatic representation of a holotrichid ciliated protozoon. (b) A diagrammatic representation of an entodiniomorphid ciliated protozoon Table 2.4 Comparison between holotrichid and entodiniomorphid ciliates Characteristics Morphology Ciliary arrangement Ectoplasm Nucleus Skeletal plates Number Proportion Diurnal variation Function Substrates fermented Genera (common) Holotrichs Entodiniomorphs Distributed on the entire (or almost the entire) surface of the cell Thin Located in the endoplasm Spherical or oval shaped macronucleus Absent Restricted ciliary zones 1–10 × 104 10–25 % Two- to fourfold increase in number in 1–2 h after feeding Do not hydrolyze structural polysaccharides (Cellulose, hemicellulose) Starch, pectin, soluble sugars, proteins Isotricha Dasytricha Charonina Thick Located in the ectoplasm Rod shaped with or without lobes. Useful in species identification Present. Number and location is useful in generic identification 1–10 × 105 75–90 % Numbers generally decrease following feeding Do hydrolyze structural polysaccharides Cellulose, hemicellulose, starch, pectin, soluble sugars, proteins Entodinium Diplodinium Eudiplodinium Ostracodinium Metadinium Polyplastron Ophryoscolex Epidinium 2 Microbiology of the Rumen 47 not have a zone or rows of cilia at the dorsal end of the cell. The group contains a several genera, including the most prevalent genus, Entodinium. Holotrichid protozoa are much more motile than entodiniomorphid ciliates. The entodiniomorphs are more numerous in the rumen compared to holotrichs. The diurnal variations in population size of the holotrichs and entodiniomorphs in the rumen differ. Generally, holotrichid population increases 1–2 h following feeding and then decline to the prefeeding concentration. The explanation is large numbers of holotrichid ciliates are sequestered on the reticular wall and or settled at the bottom of the rumen and migrate to the rumen in response to some sort of chemical stimulation (nutrient) provided by the feed that enters the rumen. In contrast, entodiniomorphs decrease following feeding, attributed to dilution of ruminal contents, and then increase gradually to reach the prefeeding concentration. Several genera and hundreds of species of ciliated protozoa have been identified in the rumen. Generally, description of the ciliated protozoal population in the rumen is done at the generic level. Ciliated protozoa generally reproduce asexually by binary fission. In some instances protozoa may exchange genetic material by conjugation and then undergo division. All ciliates actively ingest bacteria and digest the cells. In fact, bacteria are the major source of protein for ciliates. Contribution to Ruminal Digestion. Ciliated protozoa constitute an important fraction of the total microbial mass in the rumen and they actively participate in ruminal digestion. They possess full complement of hydrolytic enzymes to ferment the major components of the feedstuffs. However, our knowledge of the biochemical activities of ruminal protozoa is somewhat limited because of the difficulty in cultivating them in vitro. The impact of ciliated protozoa on ruminal digestion and fermentation products depends on their concentration and generic composition of the population. The estimates of generic composition based on numbers may not be meaningful because of large differences in cell size or volume. For example, the genus Entodinium based on the number accounts for 60–80 % of the total protozoa. However, based on volume, the Entodinium accounts for 10–40 % of the total. It is possible that the voluminous ciliated protozoa (Isotricha, Metadinium, Polyplastron, Ophryoscolex, and Epidinium) contribute to a greater extent to the metabolic activity of the total protozoa. Holotrichid ciliates are primary users of soluble sugars, while entodiniomorphs use a large variety of substrates. Most entodiniomorphs, with the exception of small entodinia, are able to ingest small plant particles and use cell wall carbohydrates. All entodiniomorphid ciliates have high amylase activity to digest engulfed starch granules. Some of the engulfed starch may be stored as granules or in the skeletal plate as amylopectin. Digestion of the engulfed protein occurs within the cells. Compared to bacteria, ciliated protozoa are less able to transport amino acids into the cell. One of the commonly used approaches to study the role of ciliated protozoa in ruminal digestion is to eliminate them from the rumen, a process called defaunation. Because ciliated protozoa are preferentially retained in the rumen, their contribution to the postruminal microbial protein supply is not as important as bacteria. In fact, their overall value to the host is a subject of some debate. The effect of the presence or absence of ciliated protozoa on the ruminant host may depend greatly on the diet and 48 T.G. Nagaraja numbers and kinds of protozoa. In animals fed low-protein diet, elimination of protozoa may increase protein supply. An increase in microbial protein synthesis because of increased bacterial numbers (lack of predation) and decrease in protein degradation result in increased flow of protein into the small intestine. Also, in ciliatefree animals, the cell wall and organic matter digestibilities are lowered resulting in lower absorbed energy than faunated animals. In animals fed high-grain diets, presence of ciliated protozoa may be beneficial by controlling the bacterial numbers and slowing down the rate of starch fermentation. The presence of ciliated protozoa results in a more stable ruminal fermentation Ruminal Fungi Anaerobic fungi, as a member of the ruminal microbial population, were discovered in 1975 by Colin Orpin. Before the discovery, fungal structures (spores) were mistaken to be flagellated protozoa. Since the discovery in the rumen, anaerobic fungi have been isolated from the gut contents of a number of animal species. Types and Morphology. Based on morphological features, fungi are classified into two broad groups: Yeasts and Molds. Yeasts are single celled organisms and molds are multicellular forming a network of filaments called hyphae. Hyphae are tubular structures and are collectively called mycelium. Functionally, fungi in the rumen are broadly classified into two groups (Fig. 2.5). The group 1 consists of fungi that are facultatively anaerobic or aerobic, are transient and do not contribute to ruminal digestion. The second group consists of obligately anaerobic fungi that are indigenous to the rumen and contribute to ruminal digestion. Yeasts or yeast-like cells in the rumen range in concentration from 102 to 103 per g of contents and are generally believed to be transient population and make no significant contribution to the ruminal fermentation. However, the number of yeasts increases in the rumen of animal that is acutely acidotic with lactic acid accumulation. The increase is probably because of availability of highly fermentable sugars. The functionally important fungi are referred to as Chytridomycete because they are placed in a phylum, Chytridomycota, which includes fungi that reproduce with motile zoospores. So far five genera and several species have been identified in the rumen and other gut regions of cattle and other animals. The genera are recognized based on the type of sporangium and thallus development, type of rhizoid development, and the number of flagella on the zoospores. Life Cycle. The life cycle of chytridomycete fungi in the rumen consists of two stages (Fig. 2.6); a motile, flagellated zoospore stage in ruminal fluid, and a nonmotile, mycelial stage, referred to as thalli, which is associated with solid feed particles. The zoospores that float freely in ruminal fluid reach plant fragments, possibly mediated by chemotactic response to soluble nutrients, attach and encyst (looses flagella). The encysted zoospore germinates by first forming a germ tube, which then grows and branches into a rhizoidal structure or thallus in Anaeromyces, 2 Microbiology of the Rumen Fig. 2.5 Morphological and functional classification of ruminal fungi Fig. 2.6 Life cycle of ruminal fungi 49 50 T.G. Nagaraja Orpinomyces, Neocallimastix, and Piromyces or into a spherical body called holdfast in Caecomyces. At the same time, the encysted spore grows into a sporangium, a sac like structure. The nucleus in the sporangium undergoes mitotic division and each nucleus encloses cytoplasm to develop into a flagellated zoospore. The sporangium then ruptures to release the spores and the cycle is then repeated. Based on the type of sporangium development, ruminal fungi can be monocentric (single sporangium per thallus) or polycentric (multiple sporangia per thallus). In monocentric fungus (Neocallimastix, Piromyces), the nucleus is retained in the encysted zoospore which enlarges into a sporangium, which is called endogenous sporangium development or exogenous sporangial development, where the nucleus migrates out of the zoospore and the sporangium develops in the germ tube or sporangiophore. In monocentric fungus, the mycelium is anucleated. In polycentric fungus (Anaeromyces, Caecomyces, Orpinomyces), the nucleus from the encysted zoospore migrates into the mycelium where it undergoes mitotic division and as a result the mycelium is polynucleated. The sporangia are formed on sporangiophore singly or in groups of up to six. Contribution to Ruminal Digestion. The contribution of fungi to microbial mass is difficult to assess because of the two-stage life cycle and the extensive growth of the thalli within plant fragments. The counts of zoospores (103–104 per ml) and the sporangia on feed particles do not reflect fungal mass. Using chitin as a marker, it has been estimated that fungi account for up to 10 % of the total microbial mass in the rumen. The relative contribution of fungi to ruminal digestion is not known. Experiments in which ruminal fungi were eliminated from the rumen or greatly reduced have provided evidence for the contribution of fungi to the digestion and overall fermentation of carbohydrates in the rumen. Ruminal fungi produce the hydrolytic enzymes required to break down the major components of plant biomass. The enzymes include cellulases, hemicellulases, pectin lyases, amylases, and proteases. Although fungi can break down pectin, they are unable to use the degradation products. Also, ruminal fungi produce phenolic esterases (p-coumaroyl and feruloyl) that can break cross linkages between hemicelluloses and lignin, which allow the fungus to have increased access to hemicelluloses. The rhizoidal development of the thalli is able to penetrate plant tissue better than bacteria and protozoa, which may lead to greater degradation of forage. Ruminal fungi are able to use a wide range of di or monosaccharides. All ruminal fungal species can utilize glucose, cellobiose, and lactose but are unable to use arabinose. Very few fungi are able to utilize maltose, galactose, mannose, ribose, rhamnose, trehalose, and melezitose. It is surprising that many fungi are unable to utilize sugars like arabinose, galactose, mannose, and ribose, which are common constituents of plant carbohydrates. Based on pure culture studies, ruminal fungi, like bacteria, have a mixed acid fermentation. They can metabolize hexoses or pentoses to produce acetate, formate, lactate, ethanol, CO2, and H2. Because fungi produce the major precursors of methane (Formate and H2); the fermentation profile is altered (less ethanol, lactate, and H2) in the presence of methanogens. The shift in fermentation products, as a result of interspecies H2 transfer, leads to an increased ATP production, which increases fungal biomass, enzyme production and rate and extent of substrate utilization. 2 Microbiology of the Rumen 51 It is generally recognized that with forage-based diets, particularly of low quality plant biomass, ruminal fluid has a lot of zoospores and a substantial portion of plant fragments in the rumen are also colonized with fungi. These observations have led to the suggestion that fungi may contribute to a greater extent to the digestion of fibrous plant material. In vitro fermentation studies with pure cultures of ruminal fungal species, depending on the feedstuffs, up to 75–90 % of cell wall degradations have been observed. Bacteriophages Bacteriophages are viruses that infect bacteria. They are composed of nucleic acids (DNA or RNA, either single or double stranded) and protein. When a phage attacks a bacterial cell, it first attaches to a receptor on the bacterial surface and then injects the nucleic acid into the cell. Once the DNA enters the cell, one of two processes happens, lysis or lysogeny. In the lytic process, viruses replicate (synthesis and assembly) and result in lysis and release of phages. In the lysogenic process, the phage DNA gets incorporated into the host cell DNA. The phage remains latent and does not cause lysis. Such a state is called lysogeny. The inserted phage DNA is called prophage and is passed on to the daughter cells when the host cell divides. Prophage may confer new properties to the host cell or may get excised from the bacterial chromosome and enter a lytic phase. Based on electron microscopic observation, a large and diverse population of phages has been described in the rumen. The number of phage particles in the rumen could range from 107 to 1011 per ml or g of ruminal contents. Both lytic and lysogenic phages and phages specific to several bacterial species have been identified. No known function has been identified of any temperate phages and it is possible that lytic phages contribute to nutrient recycling and have some influence on the species or strain composition of the bacterial population in the rumen. Microbiology of Ruminal Digestion The major components of ruminant diets are polymers and include carbohydrates, nitrogenous (protein and nonprotein) substances, lipids, and lignins. The polymers, except for lignins, are hydrolyzed to monomers, which are then metabolized to various fermentation products, chiefly acids and gases, depending on the microbial species. The extent to which the polymers are degraded in the rumen depends on the feedstuff and length of retention in the rumen. Lignins are polymer of phenolic compounds and are virtually indigestible feed component. Because lignins are located in the plant cell wall and are covalently linked to hemicelluloses, there is a negative relationship between lignin content and the extent of fiber degradation in the rumen. In bacteria and fungi, plant polymers (cellulose, hemicelluloses, pectin, starch, protein, and lipids) are hydrolyzed extracellularly to small oligomers (<6-monomer length) and monomers, which are then transported into the cell for 52 T.G. Nagaraja Fig. 2.7 Carbohydrate fermentation in the rumen further metabolism into fermentation products. In ciliated protozoa, the degradation of the polymer takes place inside the cell because feed particles are ingested and then digested in a food vacuole within the endoplasm. Carbohydrate Fermentation. Feed carbohydrates include polysaccharides and simple sugars. Polysaccharides may be structural (cellulose, hemicelluloses, and pectin) or non-structural (starch) (Fig. 2.7). Bacteria, ciliated protozoa, and fungi produce a variety of glycosyl hydrolases that breakdown the glycosidic bonds to produce oligosaccharides first and then the di- and monosaccharides. The first step in fiber degradation is the attachment of microbes to the feed particles and the attachment is mediated by capsule, and in some cases specific binding protein may be involved. Several species of ruminal bacteria and ciliated protozoa and all species of ruminal fungi possess cellulolytic activities. The three bacterial species that are considered to be most abundant in the rumen are Fibrobacter succinogenes, Ruminococcus albus, and Ruminococcus flavefaciens. Interestingly, all three are not proteolytic. Some of the noncellulolytic bacteria are capable of utilizing cellodextrins produced by cellulolytic bacteria. The major cellulolytic bacteria can also digest hemicelluloses and pectin. Noncellulolytic bacteria that can digest hemicelluloses include Prevotella sp. (albensis, brevis, bryanti, and rumincola), Butyrivibrio fibrisolvens, Pseudobutyrivibrio xylanivorans. The major enzymes involved in hemicelluloses degradations are endoxylanases and several debranching enzymes with arabinofuranosidase being the most important. Although pectin is a structural 2 Microbiology of the Rumen 53 polysaccharide it is completely digested in the rumen. The major pectinolytic bacteria include Prevotella sp., Lachnospira multiparus, Streptococcus bovis, and Trepnema sp. (bryantii and saccharophilum). Although S. bovis is pectinolytic, it does not utilize the products of pectin degradation (d-galacturonic acid). Similarly ciliated protozoa can breakdown pectin but cannot utilize the products. The predominant pectinolytic enzyme is pectin lyase. Starch is rapidly digested in the rumen and the extent of digestion depends on the grain type and the degree of grain processing. The major amylolytic bacteria in the rumen include Ruminobacter amylophilus, Selenomonas ruminantium, Streptococcus bovis and species of Lactobacillus and Bifidobacterium. The major enzymes involved are alpha-amylase and debranching enzyme, pullulanase. Entodiniomorphid protozoa engulf starch granules and ferment it slowly, which in a way contributes to slowing down the rate of starch fermentation in the rumen. Fungi have amylase activity but it is believed to contribute minimally because fungal population decreases in grain-fed animals. Many of the polymer-fermenting bacteria, ciliated protozoa, and fungi can ferment disaccharides and monosaccharides produced from initial hydrolysis. Additionally, rumen has active sugar fermenting bacteria that belong to the genera Streptococcus, Bifidobacterium, Lactobacillus, and Treponema. Of these, S. bovis and Lactobacillus sp. are important because of their propensity for explosive growth and production of lactic acid in situations where rumen is presented with excessive fermentable carbohydrate. Quantitatively, glucose and xylose are the predominant sugars presented to the microbes and Embden-Meyerhof or hexose monophosphate or pentose phosphate pathways metabolize them, with pyruvate as the major intermediate product. The types of fermentation products produced from pyruvate depend on the microorganism and ruminal conditions like pH and dilution rates (Fig. 2.8). The fermentation products produced in the rumen include acetoin, butanediol, acetate, formate, ethanol, lactate, succinate, propionate, butyrate and valerate. Formate is utilized by methanogens to produce methane. Lactate and succinate are intermediate products because lactate-utilizing and succinate-utilizing bacteria further metabolize them, respectively. Succinate is decarboxylated to propionate and the major organism involved is Selenomonas, which is why succinate does not accumulate in the rumen. Lactate is metabolized to acetate, propionate, and butyrate by two major lactate-utilizng bacteria, Megasphaera elsdenii and Selenomonas ruminantium. If rate of lactic acid production exceeds the rate of fermentation, then lactate accumulates in the rumen, a condition called lactic acidosis. Protein and Non-Protein Nitrogen Fermentation. Many of the carbohydrate- fermenting bacteria are also proteolytic. The most active proteolytic bacteria are Prevotella sp., Ruminobacter amylophilus, Butyrivibrio fibrisolvens, Streptococcus bovis, Selenomonas ruminantium, and Megasphaera elsdenii. Proteins are broken down into polypeptides and then to short peptides and amino acids (Fig. 2.9). The rate of peptide breakdown depends on amino acid composition. The peptide fermentation in the rumen is a two-step process. In the first step, dipeptidyl peptidases cleave dipeptides from the n-terminal end of the polypeptides, followed by cleaving of dipeptides into amino acids by dipeptidase. A common bacterial species that has been shown to contain high dipetidyl peptidase activity is Prevotella ruminicola. The dipeptidase 54 T.G. Nagaraja Fig. 2.8 Pyruvate metabolism in ruminal bacteria activity has been detected in many bacterial species and ciliated protozoa. Ruminal fungi also possess aminopeptidase activity but the extent of fungal contribution to peptide fermentation is not known. The rumen has very little free amino acids, which suggests that amino acids are rapidly fermented in the rumen. Deamination is probably the most common mode of amino acid fermentation and almost all proteolytic bacteria are involved in deamination. The rumen also has a specialized group of bacteria called ‘hyperammonia producers’ that do not ferment carbohydrates but can hydrolyze small peptides and deaminate amino acids. So far four species have been identified as hyper ammonia producers: Peptostreptococcus anaerobius, Clostridium sticklandii, Clostridium aminophilum, and Fusobacterium necrophorum. Therefore, amino acid deamination in the rumen can be carried out by bacteria that have low activity but are high in numbers or by bacteria that are low in numbers but have high activity. Ciliated protozoa also have a significant role in deamination. Urea is rapidly fermented in the rumen and the source of urease activity is believed to be epimural bacteria. The microbial species involved in fermentation of nucleic acids or nitrates is poorly understood. Neither ciliated protozoa nor fungi have any ureolytic activity. Most of the rumen bacteria are capable of using ammonia as a nitrogen source. Therefore, hydrolysis of NPN such as urea, nitrates or nucleic acids is beneficial to ruminal fermentation. Glutamic dehydrogenase and glutamine synthetase -glutamine synthase are key enzymes involved in ammonia assimilation by ruminal bacteria. 2 Microbiology of the Rumen Fig. 2.9 Overall scheme of nitrogen fermentation in the rumen 55 56 T.G. Nagaraja Fig. 2.10 Lipid fermentation in the rumen Lipid Fermentation. The predominant lipids in ruminant feeds are galactolipids, triglycerides, and phospholipids. The lipids are rapidly hydrolyzed by lipases or esterases produced by bacteria and ciliated protozoa (Fig. 2.10). Fungal contribution to lipid fermentation is not known. The major lipolytic bacteria in the rumen are Anaerovibrio lipolytica and Butyrivibrio fibrisolvens. Certain strains of Treponema also have lipolytic activity. The products of lipid hydrolysis are glycerol and fatty acids and galactose in case of galactolipids. Glycerol is rapidly fermented (A. lipolytica, B. fibrisolvens, and Selenomonas ruminantium) to acetate, propionate and butyrate. The long chain fatty acids are not degraded in the rumen but may get incorporated into cellular lipids by the microbes. In anaerobic ecosystems other than the gut, oxidation of free fatty acids is carried by sulfate-reducing bacteria or by a specialized group of bacteria called obligate H2-producing (proton-reducing) acetogenic bacteria. In the rumen, if the fatty acids are unsaturated a process called biohydrogenation will hydrogenate them. The two major unsaturated fatty acids in ruminant diets are linoleic (cis-9, cis-12-C18:2) and linolenic acid (cis-9, cis-12, cis-15-C18:3). The prerequisite for biohydrogenation is that the fatty acid has a free carboxyl group. The unsaturated fatty acids are isomerized (cis to trans) before sequential addition of hydrogen to saturate each double bond to produce stearic acid (C18:0). Biohydrogenation leads to some transient intermediates and the two that have received a great deal of attention are conjugated linoleic acid (CLA) and trans10 and 11 fatty acids. A range of positive health benefits have been ascribed to CLA and 2 Microbiology of the Rumen 57 both CLA and trans-fatty acids, particularly trans-10 inhibit lipogenesis in mammary glands and are contributing factors in low milk fat syndrome. Both bacteria and ciliated protozoa are involved in biohydrogenation. However, protozoa may not be a major contributor because defaunation decreases biohydrogenation only slightly. Bacterial species that have been shown to biohydrogenate include Butyrivibrio hungatei, B. proteoclasticus (formerly Clostridium proteoclasticum) and Propionibacterium acnes. Biohydrogenation may be a detoxification mechanism because unsaturated fatty acids are more toxic than saturated fatty acids to microbes. Microbial Interactions Fermentation of feedstuffs in the rumen is the result of coordinated activities of multitude of microorganisms. Because microbes compete for nutrients and space in the rumen, it is not surprising that there are different types of interactions. A major interaction that exists is cross feeding among microbes that result in a relatively more complete utilization of feedstuffs into the final fermentation products of VFA and methane. The major components of ruminant feedstuffs are cellulose, hemicelluloses, pectin, starch, proteins and lipids. Lignin is also plant polymer but it is not degraded by the microbes and in fact acts as a physical barrier to restrict digestion of cell wall polysaccharides. Based on the type of substrates metabolized, ruminal microbes can be grouped into three metabolic groups (Fig. 2.11). The group 1 includes fermentative or hydrolytic bacteria, protozoa and fungi that breakdown complex polymers (carbohydrate, proteins, and lipids) initially into oligomers and monomers, and finally to acids (mainly acetate, propionate and butyrate), alcohols (ethanol and methanol), and gases (mainly H2 and CO2). Among fermentative microbes, some are polymer fermenters (Group IA) that can convert polymer to the final fermentation products and monomers (Group IB) that cannot breakdown polymer but can utilize monomers to produce acids, alcohols, and gases. The fermentative bacteria include bacteria that ferment carbohydrates, proteins, and lipids. The second group includes the archaeal domain, methanogens (Group II) that convert H2 and CO2 or acetate to methane. The rumen also contains another metabolic group of bacteria, called homoacetogens (Group III), which can convert H2 and CO2 to acetate. Acetogens are not functional in the rumen because of their inability outcompete methanogens for hydrogen. The following are some of the well-documented interactions in the rumen: 1. Interactions between fibrolytic and proteolytic microbes. This interaction is the basis of protein or nonprotein nitrogen supplementation of animals consuming low quality forages. The products of protein degradation, branched chain fatty acids and ammonia, are important growth factors for fibrolytic bacteria and the interaction results in higher degradation of fiber. 2. Interactions among succinate producing fibrolytic or amylolytic bacteria and succinate-utilizing bacteria (Selenomonas ruminantium). This interaction explains why succinate, a product of many bacterial species, does not accumulate in the rumen and instead becomes propionic acid. 58 T.G. Nagaraja Fig. 2.11 Microbial groups and stages in ruminal fermentation 3.Interactions among lactic acid producing microbes (bacteria, protozoa, and fungi) and lactate-utilizing microbes (bacteria and protozoa only). This interaction explains why lactic acid does not accumulate in the rumen unless the rate of production exceeds the rate of utilization, as in lactic acid acidosis. 4. Interaction, referred to as interspecies hydrogen transfer, between hydrogen producing microbes (bacteria, protozoa, and fungi) and hydrogen-utilizing bacteria, mainly methanogens. This interaction results in change in fermentation products (more acetate and less reduced fermentation products like lactate, ethanol, succinate, and propionate) and higher ATP yield (because of more acetate) by fermentative bacteria because of the hydrogen utilization by methanogens. Another major interaction is the predatory role of ciliated protozoa that prey on bacteria and fungal spores. In the absence of bacteria (defaunated rumen) bacterial and fungal spores increase 10- to 100-fold. Ciliated protozoa engulf bacteria, which serves as a major source of nitrogen. Ruminal Microbial Development in the Calf A newborn calf is functionally a monogastric animal, with the abomasum serving as the primary site of digestion. The development of the rumen into a functional organ in the neonate requires a period of growth and development involving transitional changes from the undeveloped organ to the fully functional rumen. The transitional 2 Microbiology of the Rumen 59 development is characterized by a number of changes in the anatomy, physiology, and microbiology of the rumen. The stimulation of ruminal anatomical and physiological development by ruminal fermentation product suggests an interrelationship between the ruminal development and the microbial activity. The length and extent of the changes are affected by several conditions, most importantly the consumption of dry feed. Factors that influence establishment include but are not limited to type of diet, water, dirt, bedding material and proximity of other ruminants. The rumen of a calf at birth is practically a sterile organ. Microbial colonization of the digestive systems of newborn ruminants follows a typical succession in that bacteria proliferate in the fluid phase immediately after birth and colonize the ruminal wall within 36–48 h. These fluid-phase bacteria facilitate the subsequent sequential colonization of the fluid by fungi, followed by protozoa, which result in the establishment of a complex consortium that eventually develops in the fluid, on feed particles and on ruminal epithelial surfaces. Once established, they are generally stable and will change only when the nutrients are changed. Establishment of Ruminal Bacteria. In the first few days of birth, anaerobic bacteria inhabit the rumen predominantly. Facultative bacterial counts are greatest at 1–3 weeks of age and then decrease steadily as the calf ages and the number of anaerobic bacteria increases during the first 3 weeks. Although the neonatal bacterial population is predominantly anaerobic, the predominant genera observed in the neonate are quite different from those in adult ruminants. The sequence of appearance of much of the ruminal bacteria appears to be primarily dependent upon the diet. Even though isolated calves develop ruminal bacterial populations, the source and establishment of many ruminal bacteria are not certain. Early research identified the predominate bacteria isolated from neonates as coliforms, lactobacilli, streptococci, and gram negative, facultative, anaerobes. Cellulolytic bacteria have been detected at relatively high numbers in the rumen of calves at 3 days of age. Ruminal methanogens have also been detected in calves at 3 days of age and in high numbers (108/g) in calves at 1 week of age. Therefore, the initial establishment of cellulolytic bacteria in the rumen seems to be independent of the amount of cellulose digested or kind of diet fed. Establishment of Ruminal Protozoa. Flagellated and ciliated protozoa establish themselves at different times and establishment may be influenced by several factors. In general, flagellated protozoa become established more easily than ciliates, but their contribution to the ruminal development is not significant. Ruminal establishment of ciliated protozoa in the young calf requires some form of contact with faunated ruminants, and calves that were isolated from mature ruminants usually do remain defaunated. Ciliates may be established as early as 1 week of age; however, complete establishment of a ruminal protozoal population requires several months. Ciliates probably establish themselves according to their sensitivity to low ruminal pH. Entodinium, being the least sensitive to low pH, is usually established first, followed by Diplodinium and then holotrichs. Establishment of Ruminal Fungi. Ruminal fungi typically appear in the rumen in 7–8 days after birth, which means fungi can survive in the rumen in the absence of plant materials. In the first 2 weeks, rumen will have saliva and sloughed epithelial 60 T.G. Nagaraja cells and some milk that leaked from the reticular groove. Some of the species typically seen in the adult ruminant (Neocallimastic, Piromyces, etc.) have been isolated from the rumen of the newborn. Subsequent development of the fungal population is to a large extent is dependent on consumption of solid feed. It is not certain whether fungal establishment in a newborn requires contact with or in close proximity of adults. Because fungi have been detected in saliva and feces and it is likely that they can be transmitted from the adults to the new born. Ruminal Fermentative Changes. The milk fed neonate has a ruminal pH of approximately 6.0 with little postfeeding diurnal variation. As the calf consumes dry feed and ruminal function develops, the ruminal pH steadily drops to pH 5–5.5. Age at which this decline is observed is diet and intake dependent, however it is usually observed between 4 and 5 weeks of age in calves offered dry feed from birth. Weaned calves consistently have a lower pH than nonweaned calves of the same age. This drop in ruminal pH is generally linked to increased ruminal microbial activity, and corresponds to the subsequent increase of ruminal VFA concentrations. The pH appears to stay low only a week or two and then gradually increases to pH 6.0 or higher at approximately 10 week of age. This increase may be due to increased absorption of VFA as the rumen matures and possibly to increased salivary secretion. The increased dry feed consumption and change in diet from milk to dry feed also causes a shift in the VFA produced. Acetate decreases and propionate increases decreasing the acetate:propionate ratio. This decreased acetate:propionate ratio is probably a result of increased dry feed entering the rumen stimulating increased amylolytic activity, resulting in more propionate production, combined with a decrease in acetate producing facultative bacteria. Molar proportions of butyrate increase with the age of the calf. This increase may be due to the increased production of butyrate from lactate fermentation due to the low ruminal pH of the young calf. In addition, the rate of neonatal absorption of ruminal butyrate is low. Molar proportions of isobutyrate and isovalerate tend to decrease with the age of the calf. These branch-chained fatty acids are growth factors for cellulolytic bacteria, and the decline of these acids may be due to increased cellulolytic activity in the calf. Ruminal lactate concentrations steadily increase until 3–4 weeks of age, then drop to <0.5 mM by 12 week of age. Dry feed consumption changes the microbial composition away from a facultative, lactate producing population towards a lactate utilizing anaerobic population. This population shift is probably responsible for the steady decline in ruminal lactate concentration that often occurs after 4 weeks of age. Ruminal ammonia-N concentrations generally decrease as calves age. Decreased ruminal ammonia-N is most likely a result of increased ruminal bacterial utilization, and increased ruminal absorption. References Bonhomme A. Rumen ciliates: their metabolism and relationships with bacteria and their hosts. Anim Feed Sci Technol. 1990;30:203–66. Chaucheyras-Durand F, Ossa FG. Review: The rumen microbiome: composition, abundance, diversity, and new investigative tools. Prof Anim Sci. 2014;30:1–12. 2 Microbiology of the Rumen 61 Firkins JL, Yu Z, Morrison M. Ruminal nitrogen metabolism: perspectives for integration of microbiology and nutrition for dairy. J Dairy Sci. 2007;90:E1–16. Gruninger RJ, Puniya AK, Callaghan TM, Edwards JE, Youssef N, Dagar SS, et al. Anaerobic fungi (phylum Neocallimastigomycota): advances in understanding their taxonomy, life cycle, ecology, role and biotechnological potential. FEMS Microbiol Rev. 2015;90:1–17. Hespell RB, Aiken DE, Dehority BA. Bacteria, fungi, and protozoa in the rumen. In: Mackie RI, White BA, editors. Gastrointestinal microbiology. New York: Chapman & Hall; 1997. p. 59–141. Hungate RE. The rumen protozoa. In: Kreier JP, editor. Parasitic protozoa, vol. 2. NY: Academic; 1978. p. 655–95. Janseen PH, Kirs M. Structure of the archaeal community of the rumen. Appl Environ Microbiol. 2008;74:3619–25. Jenkins TC. Lipid metabolism in the rumen. J Dairy Sci. 1993;76:3851–63. Krause DO, Denman SE, Mackie RI, Morrison M, Rae AL, Attwood GT, et al. Opportunities to improve fiber degradation in the rumen: microbiology, ecology, and genomics. FEMS Microbiol Rev. 2003;27:663–93. Kim M, Morrison M, Yu Z. Status of the phylogenetic diversity census of ruminal microbiomes. FEMS Microbiol Rev. 2011;76:49–63. Krause DO, Nagaraja TG, Wright ADG, Callaway TR. Board invited review: Rumen microbiology: leading the way in microbial ecology. J Anim Sci. 2013;91:331–41. Lourenco M, Ramos-Morales E, Wallace RJ. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal. 2010;4:1008–23. Martin C, Morgavi DP, Doreau M. Methane mitigation in ruminant: from microbe to the farm scale. Animal. 2010;4:351–65. Mountfort DO, Orpin CG. Anaerobic fungi: biology, ecology, and function. New York: Marcel Dekker, Inc.; 1994. Theodorou MK, Lowe SE. The fermentative characteristics of anaerobic rumen fungi. Biosystems. 1988;21:371–6. Wallace RJ. Ruminal metabolism of peptides and amino acids. J Nutr. 1996;126:S1326–34. http://www.springer.com/978-3-319-30531-8