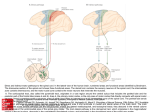
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 1. Overview: The Nervous System Anatomical planes The brain can be dissected for study in several ways: A medial cut or section divides the brain into right and left halves of equal size, separating the right and left hemispheres from one another. A sagittal cut runs parallel to the medial cut, but divides the brain into right and left portions of unequal size. A medial section may be considered to be a type of sagittal cut. However, a sagittal section is not a type of medial cut. A coronal cut runs from ear to ear, separating the brain into front and back portions. Horizontal (equal halves) or transverse cuts are perpendicular to coronal, medial, and sagittal cuts. They divide the brain into upper and lower sections. When describing the nervous system, anatomists use the terms anterior and posterior to indicate front and back. Superior and inferior are used to refer to the upper and lower parts of the nervous system. Cranial and cephalic may be used as synonyms for superior. Rostral, which literally means "toward the beak," is also sometimes substituted for superior. The antonym for rostral is caudal, a term that means "toward the tail," and may be used to replace inferior in descriptions of the brain and spinal cord. Ventral means "toward the belly" and dorsal means "toward the back." Structures in the lower part of the brain may be described as ventral. Medial means toward the center while the term lateral signifies toward the sides. The neuraxis or central nervous system consists of the brain and spinal cord The Brain is made up of the cerebral cortex , sub cortical structures, brain stem and cerebellum The spinal cord consists of grey and white matter surrounded by meninges in which cerebro-spinal fluid circulates. It runs from just below the medulla to small of back. Below that the cauda equina consisting of projections from the spinal cord goes down to the coccygeal area. Peripheral Nervous System Cranial Nerves There are twelve pairs of cranial nerves, Ten of them have their cell bodies in the brain stem. Some are motor; some are sensory, and some are both motor and sensory. Six of them are involved in speech and swallowing. Spinal Nerves They connect the central nervous system to the body. There are thirty-one pairs each of which is both sensory and motor. Autonomic Nervous System Distribution Involved in control of all automatic and glandular functions, it is controlled by the hypothalamus It works with the the endochrine system for control of hormonal secretion. Sympathetic and Parasympathetic Divisions Sympathetic prepares the body for flight or fight. Parasympathetic helps, among other things, to bring the body back to normal. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 2. General Description of the Central Nervous System The Cortex The two hemispheres of the brain are covered by a layer of cells called the cortex. (Cortex means "bark" in Latin.) The surface of the cortex is ridged, as it is made up of gyri and sulci. A gyrus is a raised fold of tissue. A sulcus is a groove lying between two gyri. A particularly deep sulcus may be called a fissure. A convolution includes both gyri and sulci. Because it is convoluted, a large amount of cortical tissue fits into a relatively small area. The Subcortical Structures The Basal Ganglia The basal ganglia are made up of two structures, the caudate nucleus and the lenticular nucleus. The caudate nucleus is bounded on one side by the lateral ventricle and is divided into a head, body and tail. The lenticular nucleus are lens-shaped structures that have two components, the globus pallidus and the putamen. The putamen is the more lateral of the two. The Internal Capsule The internal capsule lies between the lenticular and caudate nuclei. It is a group of myelinated ascending and descending fiber tracts including the pyramidal tract that connect the cortex to other parts of the central nervous system. It begins as a corona radiata (radiating crown) from motor cells in the premotor, primary motor, and primary sensory areas of the cortex and converges into the internal capsule. The capsule itself ends within the cerebrum, but the axons that pass through it continue down to the brain stem, and spinal cord. Because so many axons join together to pass through this area, the internal capsule is sometimes referred to as a bottleneck of fibers. Despite its close proximity to the caudate nucleus and lenticular nucleus, the internal capsule is not part of the basal ganglia. The internal capsule and the basal ganglia are collectively referred to as the corpus striatum. The Limbic System This is the most ancient and primitive part of the brain. It is also called the rhinencephalon as the term "rhino" means nose in Latin and much of this area is dedicated to the processing of olfactory stimuli. The limbic system is involved, among other things, with emotion and memory. The Thalamus This subcortical structure sits within the brain at the level of the temporal lobe. It is well protected in this location. The thalamus is made up of three parts, including two thalamic bodies and the tissue that connects them which is called the massa intermedia, or the interthalamic adhesion. The thalamic bodies are separated by the third ventricle, one of the spaces in the brain that is filled with cerebral spinal fluid. The massa intermedia lies within the ventricle. The thalamus receives and organizes sensory information from the periphery. Messages from all sensory modalities with the exception of smell pass through the thalamus on their way to cortical centers and other structures for further processing. (Information about smell travels directly to the temporal lobe.) Sensory information, including touch and kinesthesia passes from the thalamus to the parietal lobe. Auditory information comes into the thalamus from the inferior colliculi of the midbrain. It is processed by the medial geniculate bodies of the thalamus before being sent to the temporal lobe. In the temporal lobe, it first arrives at Heschl's gyrus, which is the primary auditory area. From there it is sent to association areas for further processing. Visual information comes into the thalamus from the superior colliculi of the midbrain. It is processed by the lateral geniculate bodies of the thalamus and sent to the primary visual area in the occipital lobe. The visual association areas, also in the occipital lobe, further analyze the information. Lesions in the thalamus can cause a type of aphasia. The Hypothalamus The hypothalamus is located immediately below the thalamus. Part of it is also slightly anterior to the thalamus. The hypothalamus regulates the functioning of the pituitary gland, so it controls basic biological functions like appetite, body temperature, sex drive, etc. The hypothalamus is the part of the brain that makes you shiver when you are cold and sweat when you are hot. A part of the hypothalamus monitors the level of glucose in the blood and, when it notices a significant decrease, it sends messages to the stomach producing sensations of hunger. Diabetes Insipidus, the most serious type of diabetes, is caused by lesions in the hypothalamus, or between the hypothalamus and the pituitary gland. While the thalamus is an input structure, sending messages to higher brain areas, the hypothalamus is an output structure, sending messages to glands and other parts of the body. The Brain Stem The Midbrain (Mesencephalon) This is the most superior part of the brain stem. The corpora quadrigemina, the red nucleus, the substantia nigra, the cerebral peduncles, and the cell bodies of two cranial nerves are located in the midbrain. The corpora quadrigemina consists of the tectum which is the roof of the brain stem, and of four protrusions located on the tectum which are called colliculi. The two superior colliculi are involved in vision. They relay information to the lateral geniculate bodies of the thalamus. The two inferior colliculi are involved in hearing. They relay information to the medial geniculate bodies of the thalamus. The red nucleus is part of the extrapyramidal tract and connects the cerebellum to the thalamus and spinal cord. The substantia nigra is a group of dark colored cell bodies which produce dopamine. It is also part of the extrapyramidal tract. The cerebral peduncles connect the pons to the cerebrum. The nuclei of cranial nerve III, the oculomotor cranial nerve, and of cranial nerve IV, the trochlear cranial nerve which both provide innervation for eye movement are also located in the midbrain. The Pons The word "pons" is Latin for "bridge." Fibers found there connect the brain stem to the cerebellum. The cell bodies for cranial nerves V and VI, the trigeminal and abducens, as well as nuclei of cranial nerve VII, the facial nerve, are located there. The Medulla Oblongata This structure, which is the most inferior part of the brain stem, sits on top of the superior end of the spinal cord. Because it has a rounded shape, it was once called "the bulb." (The term "bulbar" refers to the brain stem.) It is involved in circulation and respiration and has several important landmarks. The pyramids, which mark the decussation of the pyramidal tract, lie on either side of the median fissure. The olivary nuclei are posterior to the pyramids. They are involved in the processing and relay of auditory information. The cell bodies of cranial nerves VIII-XII are located here. Some of the nuclei of CN VII are also found in the medulla. The Cerebellum The word "cerebellum" means "little brain" in Latin. This structure has two hemispheres, each of which is divided into lobes and is covered by the cortex. It is one of the newer parts of the brain and is very important for the production of speech. It organizes muscle activity and plays a role in the coordination of fine motor movements and also in balance. The cerebellum receives both motor and sensory input, and so is the center of a feedback loop. All motor messages that leave the brain also go to the cerebellum, including information about the strength of the impulses. The cerebellum integrates motor output so that movements are smooth and coordinated. Muscle spindles, joints and tendons send information about movement back to this area. The cerebellum then relays these messages to the cortex, completing the feedback loop. Lesions here will cause cerebellar or ataxic dysarthria which involves jerky, uncoordinated movements of the speech musculature. The cerebellum is connected to the brain stem by three pairs of tracts called the cerebellar peduncles. The Spinal Cord The spinal cord contains the cell bodies of the spinal nerves as well as their afferent and efferent fibers. It begins below a large opening in the base of the skull called the magnum foramen and extends downward, surrounded and protected by the vertebral column. It does not continue through the whole length of the column, terminating instead slightly above the level of the waist. The part of the vertebral column that lies below the spinal chord is called the cauda equina, which is Latin for horse's tail. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 3. The Meninges and Cerebrospinal Fluid The Meninges The meninges are three layers of protective tissue called the dura mater, arachnoid mater, and the pia mater that surround the neuraxis. The meninges of the brain and spinal cord are continuous, being linked through the magnum foramen. Dura Mater The dura mater is the most superior of the meningeal layers. Its name means "hard mother" in Latin and it is tough and inflexible. This tissue forms several structures that separate the cranial cavity into compartments and protect the brain from displacement. The falx cerebri separates the hemispheres of the cerebrum from one another. The falx cerebelli separates the lobes of the cerebellum from one another. The tentorium cerebelli separates the cerebrum from the cerebellum. The dura mater also forms several vein-like sinuses that carry blood which has already given its supply of oxygen and nutrients to the brain back toward the heart. The superior sagittal sinus runs across the top of the brain in an anterior-posterior direction. Other sinuses include the straight sinus, the inferior sinus, and the transverse sinus. The epidural space is a potential space that may exist between the dura mater and the skull. If there is hemorrhaging in the brain, blood may collect here. Adults tend to bleed here as a result of closed head injury. The subdural space is another potential space that may exist between the dura mater and the medial layer of the meninges, the arachnoid mater. When bleeding occurs in the cranium, blood may collect here and push down on the lower layers of the meninges. If bleeding continues, brain damage will result from this pressure. Children are especially likely to have bleeding in the subdural space in cases of head injury. Arachnoid Mater The arachnoid or arachnoid mater is the middle layer of the meninges. In some areas, it projects into the sinuses formed by the dura mater. These projections are the arachnoid granulations or villi . They transfer cerebrospinal fluid, the fluid found in the ventricles, back into the bloodstream. The subarchanoid space lies between the arachnoid and pia mater. It is filled with cerebrospinal fluid. All blood vessels entering the brain, as well as cranial nerves pass through this space. The term arachnoid refers to the spider web like appearance of the blood vessels within the space. Pia Mater The pia mater is the innermost layer of the meninges. Unlike the other layers, this tissue adheres closely to the brain, running down into the sulci and fissures of the cortex. It fuses with the ependyma, the membranous lining of the ventricles to form structures called the choroid plexes which produce cerebrospinal fluid. Cerebrospinal Fluid Purpose Cerebrospinal fluid is a clear liquid produced within spaces in the brain called ventricles. It is also found inside the subarachnoid space of the meninges which surrounds both the brain and the spinal chord. In addition, a space inside the spinal chord called the central canal also contains cerebrospinal fluid. It acts as a cushion for the neuraxis, also bringing nutrients to the brain and spinal cord and removing waste from the system. Choroid Plexus All of the ventricles contain choroid plexuses which produce cerebrospinal fluid by allowing certain components of blood to enter the ventricles. The choroid plexuses are formed by the fusion of the pia mater, the most internal layer of the meninges and the ependyma, the lining of the ventricles. The Ventricles These four spaces are filled with cerebrospinal fluid and protect the brain by cushioning it and supporting its weight. The two lateral ventricles extend across a large area of the brain. The anterior horns of these structures are located in the frontal lobes. They extend posteriorly into the parietal lobes and their inferior horns are found in the temporal lobes. The third ventricle lies between the two thalamic bodies. The massa intermedia passes through it and the hypothalamus forms its floor and part of its lateral walls. The fourth ventricle is located between the cerebellum and the pons. The four ventricles are connected to one another. The two foramina of Munro, which are also know as the interventricular foramina, link the lateral ventricles to the third ventricle. The Aqueduct of Sylvius which is also called the cerebral aqueduct connects the third and fourth ventricles. The fourth ventricle is connected to the subarachnoid space via two lateral foramina of Luschka and by one medial foramen of Magendie. Subarachnoid Space Although cerebrospinal fluid is manufactured in all of the ventricles, it circulates through the system in a specific pattern, moving from the lateral ventricle to the third, and then from the third to the fourth. From the fourth ventricle, the cerebrospinal fluid passes into the subarachnoid space where it circulates around the outside of the brain and spinal cord and eventually makes its way to the superior sagittal sinus via the arachnoid granulations or arachnoid villi. In the superior sagittal sinus, the cerebrospinal fluid is reabsorbed into the blood stream. The cerebrospinal fluid of the neuraxis is regenerated several times every twenty-four hours. Endolymph and perilymph, the fluids of the inner ear, are derived from cerebrospinal fluid. Currently, there is no consensus regarding the manner in which cerebrospinal fluid enters the inner ear. Osmosis may be involved. A condition called hydrocephalus occurs when, for some reason, too much cerebrospinal fluid is produced and the ventricles swell, putting pressure on the tissue of the brain. Tumors are one potential cause of an over-production of cerebrospinal fluid. Hydrocephalus should not be confused with hydroencephali. The term hydroencephali literally means "water brain" and refers to a rare birth defect in which the cerebrum is absent and the space where it should be is entirely filled with cerebrospinal fluid. In the past, before CT and MRI technology existed, a technique involving cerebrospinal fluid called pneumoencephalography was used to view the brain. A small amount of cerebrospinal fluid was removed from the ventricular system and replaced with air or some other inert gas. This allowed the examiner to view the ventricles in a scan and make inferences about brain pathology. Tumors and hemorrhages could sometimes be located by examining the shapes and sizes of the ventricles. Because space within the cranium is limited, growths or coagulated blood (hematoma) will displace white and gray matter, pushing them into the ventricular system. Cerebrospinal fluid can be analyzed to make judgements about a person's general health. A sample is taken from the spinal cord via a lumbar puncture which is also known as a spinal tap. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 4. Cerebral Lobes, Cerebral Cortex, and Brodmann's Areas The Cerebral Lobes Each cerebral hemisphere is divided into four lobes; the frontal, parietal, temporal, and the occipital. The Frontal Lobe is the most anterior lobe of the brain. Its posterior boundary is the fissure of Rolando, or central sulcus, which separates it from the parietal lobe. Inferiorly, it is divided from the temporal lobe by the fissure of Sylvius which is also called the lateral fissure. This lobe is associated with higher level cognitive functions like reasoning and judgement. Most importantly for speech pathologists, the frontal lobe contains several cortical areas involved in the control of voluntary muscle movement, including those necessary for the production of speech and swallowing. Broca's Area is found on the inferior third frontal gyrus in the hemisphere that is dominant for language. This area is involved in the coordination or programming of motor movements for the production of speech sounds. While it is essential for the execution of the motor movements involved in speech it does not directly cause movement to occur. The firing of neurons here does not generate impulses for motor movement; that is the function of neurons in the motor strip. The neurons in Broca's area generate motor programming patterns when they fire. This area is also involved in syntax which involves the ordering of words in speech. Injuries to Broca's area may cause apraxia or Broca's aphasia. The precentral gyrus, which may also be called the primary motor area or, most commonly, the motor strip is immediately anterior to the central sulcus. It controls the voluntary movements of skeletal muscles; cell bodies of the pyramidal tract are found on this gyrus. The amount of tissue on the precentral gyrus that is dedicated to the innervation of a particular part of the body is proportional to the amount of motor control needed by that area, not just its size. For example, much more of the motor strip is dedicated to the control of the articulators than to the legs. The premotor area or supplemental motor area is immediately anterior to the motor strip. It is responsible for the programming for motor movements. It does not, however program the motor commands for speech as these are generated in Broca's area which is also located in the frontal lobe. The most anterior part of the frontal lobe is involved in complex cognitive processes like reasoning and judgment. Collectively, these processes may be called biological intelligence. A component of biological intelligence is executive function. According to Denckla, 1996, executive function regulates and directs cognitive processes. Decision making, problem solving, learning, reasoning and strategic thinking are all part of executive functioning. Some characteristics of right hemisphere syndrome are considered problems of the executive function. They include left side neglect where there is a lack of awareness of the left side of the body. The Parietal Lobe is immediately posterior to the central sulcus. It is anterior to the occipital lobe, from which it is not separated by any natural boundary. Its inferior boundary is the posterior portion of the lateral fissure which divides it from the temporal lobe. The parietal lobe is associated with sensation, including the sense of touch, kinesthesia, perception of warmth and cold, and of vibration. It is also involved in writing and in some aspects of reading. The postcentral gyrus which is also called the primary sensory area or the sensory strip is immediately posterior to the central sulcus. This area receives sensory feedback from joints and tendons in the body and is organized in the same manner as the motor strip. Like the motor strip, the sensory strip continues down into the longitudinal cerebral fissure and so has both a lateral and a medial aspect. The presensory, secondary sensory, or sensory association areas are located behind the postcentral gyrus. These areas are capable of more detailed discrimination and analysis than is the primary sensory area. They might, for example, be involved in sensing how hot or cold something is rather than simply identifying it as hot or cold. Information is first processed in the primary sensory area and is then sent to the secondary sensory areas. The angular gyrus lies near the superior edge of the temporal lobe, immediately posterior to the supramarginal gyrus. It is involved in the recognition of visual symbols. Geschwind described this area as "the most important cortical areas of speech and language" and the "association cortex for association cortices". He also claims that the angular gyrus is not found in non-human species. Fibers of many different types travel through the angular gyrus, including axons associated with hearing, vision, and meaning. The arcuate fasciculus, the groups of fibers connecting Broca's area to Wernicke's area in the temporal lobe connects to this area. The following disorders may result from damage to the angular gyrus in the hemisphere that is dominant for speech and language: anomia, alexia with agraphia, left-right disorientation, finger agnosia, and acalcula. Anomia is a difficulty with word-finding or naming. Someone suffering from anomia can list the functions of an object and explain it meaning, but cannot recall its name. Alexia with Agraphia refers to difficulties with reading and writing. Left-right disorientation is an inability to distinguish right from left. Finger agnosia or tactile agnosia is the lack of sensory perceptual ability to identify by touch. Acalcula refers to difficulties with arithmetic. The Temporal Lobe is inferior to the lateral fissure and anterior to the occipital lobe. It is separated from the occipital lobe by an imaginary line rather than by any natural boundary. The temporal lobe is associated with auditory processing and olfaction. It is also involved in semantics, or word meaning, as Wernicke's area is located there. Wernicke's Area is located on the posterior portion of the superior temporal gyrus. In the hemisphere that is dominant for language, this area plays a critical role in the ability to understand and produce meaningful speech. A lesion here will case Wernicke's aphasia. Heschl's Gyrus, which is also known as the anterior transverse temporal gyrus, is the primary auditory area. There are two secondary auditory or auditory association areas which make important contributions to the comprehension of speech. They are not completely responsible for this ability, however, as many areas, including Wernicke's area, are involved in this process. The Occipital Lobe, which is the most posterior lobe, has no natural boundaries. It is involved in vision. The primary visual area receives input from the optic tract via the thalamus. The secondary visual areas integrate visual information, giving meaning to what is seen by relating the current stimulus to past experiences and knowledge. A lot of memory is stored here. These areas are superior to the primary visual cortex. Damage to the primary visual area causes blind spots in the visual field, or total blindness, depending on the extent of the injury. Damage to the secondary visual areas could cause visual agnosia. People with this condition can see visual stimuli, but cannot associate them with any meaning or identify their function. This represents a problem with meaning, as compared to anomia, which involves a problem with naming, or wordrecall. The Island of Reil or Insula is a cortical area which lies below the fissure of Sylvius and is considered by some anatomists to be the fifth lobe of the cerebrum. It can only be seen by splitting the lateral fissure. Little is known about the connections of this area, but it may be linked to the viscera. Drunkers, 1996 feels that it may be involved in programming for speech for speech sounds. It is important to remember that, while some functions can be localized to very specific parts of the brain, others cannot be classified in this way because many areas are involved in their performance. Word-finding, for example, is associated with several different areas. Also, we cannot say that all higher level cognitive functioning is associated with the frontal lobe; the processing of word meaning carried out by Wernicke's certainly involves a sophisticated type of cognition. Also, right hemisphere lesions often result in cognitive/perceptual problems. The Cerebral Cortex The cortex is about four millimeters thick and is composed of six layers. Listed from most superior to most inferior, these layers are; the molecular layer, the external granular layer, the internal pyramidal layer, the internal granular layer, the ganglionic layer, and the fusiform or multiform layer. The molecular layer is the most superior layer of the cortex. It contains the cell bodies of neuroglial cells. The external granular layer is very dense and contains small granular cells and small pyramidal cells. The external or medial pyramidal layer contains pyramidal cells arranged in row formation. The cell bodies of some association fibers are found here. The internal granular layer is thin, but its cell structure is the same as that of the external granular layer. The ganglionic layer contains small granular cells, large pyramidal cells as well as the cell bodies of some association fibers. The association fibers that originatehere form two large tracts: The Bands of Baillarger and Kaes Bechterew. The fusiform layer is also known as the multiform layer; its axons enter white matter, it function is unknown. All layers are present in all parts of the cortex. However, they do not have the same relative density in all areas. Depending upon the function of a particular area, some of these layers will be thicker than others in that location. The cortex wraps around the brain, covering its inferior surface and lining the gap between the right and left cerebral hemispheres, which is called the longitudinal or interhemispheric fissure. The part of the cortex covering the sides of the hemispheres is called lateral cortex while the part covering the sides of the hemispheres that lie within the longitudinal cerebral fissure are called medial cortex. Brodmann's Classification System Studies done by Brodmann in the early part of the twentieth century generated a map of the cortex covering the lobes of each hemisphere. These studies involved electrical probing of the cortices of epileptic patients during surgery. Brodmann numbered the areas that he studies in each lobe and recorded the psychological and behavioral events that accompanied their stimulation. The Frontal Lobe contains areas that Brodmann identified as involved in cognitive functioning and in speech and language. Area 4 corresponds to the precentral gyrus or primary motor area. Area 6 is the premotor or supplemental motor area. Area 8 is anterior of the premotor cortex. It facilitates eye movements and is involved in visual reflexes as well as pupil dilation and constriction. Areas 9, 10, and 11 are anterior to area 8. They are involved in cognitive processes like reasoning and judgement which may be collectively called biological intelligence. Area 44 is Broca's area. Areas in the Parietal Lobe play a role in somatosensory processes. Areas 3, 2, and 1 are located on the primary sensory strip, with area 3 being superior to the other two. These are somastosthetic areas, meaning that they are the primary sensory areas for touch and kinesthesia. Areas 5, 7, and 40 are found posterior to the primary sensory strip and correspond to the presensory to sensory association areas. Area 39 is the angular gyrus. Areas involved in the processing of auditory information and semantics as well as the appreciation of smell are found in the Temporal Lobe. Area 41 is Heschl's gyrus, or the primary auditory area. Area 42 immediately inferior to area 41 and is also involved in the detection and recognition of speech. The processing done in this area of the cortex provides a more detailed analysis than that done in area 41. Areas 21 and 22 are the auditory association areas. Both areas are divided into two parts; one half of each area lies on either side of area 42. Area 37 is found on the posterior-inferior part of the temporal lobe. Lesions here will cause anomia. The Occipital Lobe contains areas that process visual stimuli. Area 17 is the primary visual area. Areas 18 and 19 are the secondary visual areas. The Homunculus A pedagogical device called the homunculus, which literally means "little man," is often used to explain the organization of the motor strip and to demonstrate that specific areas of this gyrus are responsible for sending commands to specific parts of the body. The body is represented on the motor strip in an upside-down fashion. The lower parts of the body, like the feet and the legs, receive motor movement commands from the superior part of the precentral gyrus. Parts of the face, on the other hand are innervated by the inferior part of the motor strip. The motor strip extends down some distance into the longitudinal cerebral fissure. The portion inside this fissure is its medial aspect. The part on the lateral surface of the hemisphere is called its lateral aspect. The medial cortex controls the movements of the body from the hips on down while the lateral aspect sends commands to the upper body including the larynx, face, hands, shoulders, and trunk. The medial and lateral aspects of the motor strip have different blood supplies. Blood comes to the medial area from the anterior cerebral artery while the lateral portion is supported by the middle cerebral artery. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 5. The Corpus Striatum, Rhinencephalon, Connecting Fibers, and Diencephalon The Corpus Striatum The corpus striatum, or "striped body" consists of the basal ganglia and the internal capsule. The basal ganglia is made up of nuclei, so it is gray matter. The internal capsule is a group of tracts containing myelinated axons, so it is white. Because the internal capsule runs between the caudate and lenticular nucleus of the basal ganglia, the group of structures forms a stripe. The Basal Ganglia The basal ganglia, which is the largest subcortical structure in the brain, is located at the level of the thalamus. Its components are the caudate nucleus and the lenticular nucleus, which consists of the putamen and the globus pallidus. The caudate nucleus is bounded on one side by the lateral ventricle and is divided into a head, body and tail. It contains endorphins, chemicals that, among other things, produce a positive emotional state. The lenticular nucleus is also know as the lentiform nucleus. (Lentiform means lensshaped in Latin). It is located between the caudate nucleus and the Island of Reil with its anterior aspect being attached to the head of the caudate nucleus. The putamen is the most lateral part of the structure. When the globus pallidus, the more medial part of the lenticular nucleus, is probed, sensations of thirst are produced. Some anatomists consider the claustrum to be part of the basal ganglia. The amygdala, which is involved in emotion, was once classified as part of the basal ganglia, but is no longer categorized in this way. It is still considered to be a part of the limbic system. It is attached to the tail of the caudate nucleus. The subthalamic nuclei and the substantia nigra are both functionally related to the basal ganglia, but are not considered to be part of that structure. The Internal Capsule The internal capsule lies between the lenticular and caudate nuclei. It is a group of myelinated ascending and descending fiber tracts including the pyramidal tract that connect the cortex to other parts of the central nervous system. The capsule itself ends within the cerebrum, but the axons that pass through it continue down to the brain stem and spinal cord. They descend through the midbrain within two large bundles called the cerebral peduncles or cruz cerebri. Because so many axons join together to pass through this area, the internal capsule is sometimes referred to as a bottleneck of fibers. This makes it a very bad place to get a lesion. The striata, a branch of the middle cerebral artery, brings blood to the internal capsule. The striata is called the "artery of stroke" because it is prone to hemorrhage and supports so many important nerve fibers. If there is a problem with the blood supply in this area, many efferent and afferent tracts can be damaged. Despite its close proximity to the caudate nucleus and lenticular nucleus, the internal capsule is not part of the basal ganglia. As stated previously it forms part of the corpus striatum along with the caudate and lenticular nuclei. The Rhinencephalon The term "rhinencephalon," which literally means "smell-brain," is used by many anatomists to refer to the limbic system. Others make a distinction between these two terms considering the olfactory tract and olfactory bulbs to be part of the rhinencephalon but not of the limbic system. The limbic system is sometimes called the limbic lobe. (This term is only descriptive; it does not imply that this group of structures is comparable to the lobes of the cerebral hemispheres which are neo-cortex. The limbic structures are much older.) The limbic system consists of both cortical and subcortical structures which are located on the medial, inferior surfaces of the cerebral hemispheres. Its components are phylogenetically related, being some of the most ancient parts of the brain. The cortical areas classified as part of the limbic system include the hippocampus, the cingulate gyrus, and the subcallosal gyrus. The hippocampus, is a gyrus found on the medial edge of the temporal lobe. It is named for its shape, as hippocampus literally means "sea horse." The hippocampi are very close to the basal ganglia and to the lateral ventricles. The cingulate gyrus is immediately superior to the corpus callosum. The subcallosal gyrus is immediately inferior to the corpus callosum. The subcortical components of the rhinencephalon include the olfactory pathways, the amygdaloid bodies, the mamillary bodies, the dorsal-medial and ventral-anterior nuclear groups of the thalamus, parts of the reticular formation, and the septal region. The olfactory pathway originates in the nasal area. As it passes posteriorly to enter the temporal lobe at the hippocampal gyrus the olfactory tract is immediately superior to the optic tract. The mamillary bodies are also known as the mamillary nucleus. They are connected to the hippocampus, the thalamus and the fornix. The septal region includes both the septum pellucidum, which is a double walled structure located between the corpus callosum and the fornix, and the septal nuclei. The fornix is a group of fibers that arises from each hippocampus, and project to the contralateral hippocampus. It links the rhinencephalon to both the thalamus and the hypothalamus. It is connected to the septal nuclei, the mamillary bodies, and anterior nucleus of the thalamus. The limbic system is involved in recent memory, emotion and in motivation and reinforcement. According to Love and Webb (1992) responses mediated by the limbic system include pleasure, satiety, guilt, punishment, inhibition, wakefulness, alertness, excitement, and autonomic activity. Based on the behavioral correlates of lesions to the limbic system, it may also be involved in cortical speech and language behavior. The nature of this involvement is not known at the present. Lesions to the limbic system can also cause a variety of behaviors, including aggression, extreme fearfulness, altered sexual behavior, and changes in motivation. Damage specifically to the hippocampus can affect recent memory and emotion. Lesions on the olfactory pathways can cause anosmia, which is a loss of the sense of smell. An uncinate fit is an epileptic seizure that is receded by an olfactory hallucination. Connecting Fibers There are three major types of axons, or nerve fibers, in the brain. Efferent fibers take messages from the brain to the peripheral nervous system. They are almost always motor fibers. Afferent fibers take messages from the periphery back to the brain. They are almost always sensory fibers. Interconnecting fibers connect structures within the brain. There are two types of interconnecting fibers: commissural fibers, and association fibers. Commissural fibers connect the hemispheres of the brain. The corpus callosum, the anterior commissure, and the posterior commissure are all composed of commissural fibers. Some sources consider them to be association fibers. The corpus callosum, which is Latin for "large body" is the major group of commissural fibers. It is located some distance down inside the longitudinal cerebral fissure, the split that separates the hemispheres. It contains at least 200 million axons. Most of these fibers connect mirror image sites on the two hemispheres; axons might connect Brodmann's area 3 in the parietal lobe of the right hemisphere to area 3 in the parietal lobe of the left hemisphere. Not all of the connections follow this pattern, however. For example, area 17 in the occipital lobe is connected to areas 18 and 19 of the other hemisphere rather than to area 17. One treatment for severe epileptic seizures is commissurectomy, an operation that severs the corpus callosum. Both Sperry and Gazzaniga conducted experiments on split-brain patients and noted differences between the functions of the left and right cerebral hemispheres. The right hemisphere appears to be involved in the intuitive, holistic processing of information and in spatial reasoning. The left hemisphere, on the other hand, seems to be more adapted for logic and analytical reasoning. Most importantly for speech pathologists, it was found that the left hemisphere plays a dominant role in the speech and language abilities of most people. Of course, both hemispheres are apparently involved to some extent in any cognitive process. The other two groups of commissural fibers are called the anterior commissure and the posterior commissure. Both are connected to the corpus callosum. Many of the commissural fibers that connect the two temporal lobes pass through the anterior commissure. The anterior commissure also connects the temporal lobe to the amygdala and to the occipital lobe in the other hemisphere. The anterior commissure is used by neurosurgeons to locate the circle of Willis. Almost all parts of the cortex receive commissural fibers. The "hand area" of the primary sensory strip is one of the few areas that does not. Association fibers connect areas within the same hemisphere. The cell bodies of association fibers are the most prevalent type of neuron found in the cortex. Long association fibers connect areas that are located in different lobes of the brain. For example, the arcuate fasciculus, which connects Broca's area in the frontal lobe with Wernicke's area in the temporal lobe, is composed of long association fibers. The term arcuate fasciculus means "arching bundle" in Latin. Lesions to this particular bundle of long association fibers will cause conduction aphasia. Short association fibers connect areas that are located in the same lobe. For example, the fibers which connect Heschl's gyrus with the auditory association areas are short association fibers. The Diencephalon Location and Description The diencephalon consists of the thalamus,epithalamus (includes pineal gland), subthalamus and hypothalamus. Some sources classify the diencephalon as part of the brain stem. This is not the view of the majority and, for the purposes of this class, the diencephalon should be considered part of the cerebrum. Both the thalamus and hypothalamus are located in the center of the brain at the level of the temporal lobe. They are very well protected in this area. The thalamus is located below the caudate nucleus and the fornix and is medial to the lenticular nucleus. It is composed of two bodies which are separated from one another by the third ventricle, with one lying in each hemisphere. The two thalamic bodies are connected to one another by another part of the thalamus, the massa intermedia or thalamic adhesion, which makes up part of the ventricle. The epithalamus includes the pineal gland and is involved autonomic functions. The subthalamus is located ventral to the thalamus and is important for motor movement. It has connections to the basal ganglia, thalamus and brainstem. The hypothalamus is a solid structure that is located immediately inferior to the thalamus. Part of it is also anterior to the thalamus. It forms the floor and part of the lateral walls of the third ventricle. The Thalamus The thalamus has been described as the switchboard for the cortex. It receives information from the cerebellum, the basal ganglia and from all sensory pathways with the exception of the olfactory tract; it integrates the messages and sends them on to the cortex for further processing. The thalamic bodies are composed of several different nuclei which are divided from one another by lamina or thin walls of tissue. The thalamic nuclei can be divided into four groups based on their functions; the specific relay nuclei, the association nuclei, the non-specific nuclei, and a subcortical nucleus. Five areas of the thalamus are classified as the ventral nucleus complex. These include the lateral geniculate body, the medial geniculate body, the ventral posterolateral nucleus, the ventral posteromedial nucleus, and the ventral lateral/ventral anterior nuclei. All of the above receive sensory information and are considered specific relay nuclei. The ventral lateral and ventral anterior are motor relay nuclei The lateral geniculate body is part of the visual information pathway. It receives information from the superior colliculus of the midbrain and then relays it to the visual areas of the cortex in the occipital lobe. The medial geniculate body processes auditory information. It receives messages from the inferior colliculus of the midbrain and transfers them to the auditory areas of the cortex in the temporal lobe. The ventral posterolateral nucleus or VPL is involved in the processing of somatosensory information. Messages come in from the spinothalamic tract and the medial lemniscus and are passed on to the somato-sensory cortex found in the parietal lobe. This nucleus mediates sensations of pain and temperature as well as proprioception. The ventral posteromedial nucleus or VPM also handles sensory information. It receives input from the trigeminothalamic tract which it passes on to the somatosensory cortex of the parietal lobe. Sensory information mediated by the trigeminal nerve is processed in this area. For example, information about toothaches is carried by this tract. The ventral lateral and ventral anterior or VL/VA are motor relay nuclei of the thalamus. They process motor information. They gets input from the cerebellum and the basal ganglia and sends output to the motor and premotor cortex in the frontal lobe. As lesions here will affect motor abilities, knowledge of these nuclei is extremely important for speech-language pathologists. Association nuclei form connections between different areas of the thalamus. They are involved in the integrating and correlating processes that it performs. Three parts of the thalamus are classified as association nuclei. These include the pulvinar, the lateral posterior nucleus, and the dorsal medial nucleus. The pulvinar receives information from other thalamic nuclei and from the superior colliculus. It sends output to association areas in the parietal, occipital and temporal lobes. The medial and lateral geniculate bodies are located on top of the pulvinar but are not a part of it. The lateral posterior nucleus, or the LP, integrates input that it receives from oth The dorsal medial nucleus receives input from the amygdaloid bodies, the hypothalamus, and from other thalamic nuclei. It sends information to the pre-frontal cortex. As this nucleus connects parts of rhinencephalon with one of the motor areas of the frontal lobe, it may be involved in the hypothesized connection between the limbic system and communication. Two parts of the thalamus are categorized as non-specific nuclei. They are the intralaminar nucleus and part of the ventral anterior nucleus (the motor relay). Information comes to the intralaminar nucleus from the basal ganglia, the reticular formation, and other thalamic nuclei. It sends output to many different cortical areas. The motor relay of the ventral anterior nucleus receives input from other structures as well as from within the thalamus and sends it to the pre-motor and pre-frontal cortex of the frontal lobe. The ventral lateral nucleus is part of the motor feedback circuit between the cortex and the cerebellum. One portion of the thalamus, the reticular nucleus, is classified as subcortical. This name does not refer to the location of the structure, as all parts of the thalamus are subcortical according to this definition. Instead, the reticular nucleus is labeled this way because it does not project to the cortex. Its input comes from other thalamic nuclei and its output is passed on to thalamic nuclei. The Hypothalamus The hypothalamus is considered a nodal point in the pathways mediating autonomic, emotional, endocrine, and somatic functions. It is involved in the following specific functions: Release of some hormones from the pituitary gland. Temperature regulation of the body. Intake of food and water. Autonomin nervous system pathways: The hypothalamus is connected to reticular nuclei in the brain stem that relay axons that control autonomic motor functions. They are particulary involved with coughing and vomiting reflexes as well as the reflexes involved in expelling inspirated substances. The larynx spasms violently in response to food (liquid or solid) getting into the laryngeal glottis. Emotion: The hypothalamus is connected to the septum and the amygdaloid bodies, which are part of the limbic system, via a group of fibers called the stria terminalis. Reciprocal connections exist between the hypothalamus and the thalamus, pituitary gland, brain stem, and the temporal lobe. The hypothalamus is composed of several groups of nuclei and regions, including the preoptic area, the supraoptic area, the paraventricular nucleus, the dorsalmedial nucleus, the ventral-medial nucleus, the lateral region, and the posterior region. The preoptic area is located in the anterior portion of the hypothalamus. It is involved in temperature regulation of the body, including the dilation of peripheral blood vessels and sweating. The supraoptic area is inferior to the preoptic area and just above the optic chiasm. It is connected to the pituitary gland and regulates water intake and output via control of the kidneys. The paraventricular nucleus is also involved in water regulation. Both the supraoptic area and the paraventricular nucleus produce ADH or antidiuretic hormone. The dorsal-medial and the ventro-medial nuclei are involved in the control and expression of emotions like rage, fear, and extreme anxiety. The lateral region contains the apostat, which monitors the level of glucose in the blood and sends messages to the stomach provoking hunger when the blood sugar level drops to a certain level. The posterior region is involved in controlling the body's temperature in cold environments. This area produces shivering by causing the muscles of the body to vibrate at a rate of between seven and thirteen Hertz. This area also controls sexual behavior. Lesions in the hypothalamus may cause obesity, loss of the ability to control body temperature, or loss of interest in sex. Damage to the hypothalamus or to connections between the hypothalamus and the pituitary gland may cause diabetes insipidus which is far more serious than sugar diabetes, the more common form of the disease. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 6. The Midbrain, Pons, Medulla, and Reticular Formation The Midbrain (Mesencephalon) The mesencephalon is the most superior part of the brainstem. It is divided into an anterior and a posterior section by the Aqueduct of Sylvius which connects the third and fourth ventricles. Motor tracts, including the fibers of the pyramidal system, pass downward on the midbrain's anterior surface. Sensory axons, including those of the spinothalamic tract also ascend, along the front of the midbrain behind the motor tracts. The corpora quadrigemina, which is located on the posterior surface of the midbrain, is composed of two superior colliculi and two inferior colliculi. The superior colliculi are part of the visual system, relaying input from the optic tract to the lateral geniculate bodies of the thalamus. The inferior colliculi are part of the auditory pathway and send information to the medial geniculate bodies of the thalamus. Several important nuclei are located in the midbrain, including the red nuclei, the substantia nigra, and the nuclei of cranial nerves III and IV. The red nuclei connect the midbrain to the cerebellum. The substantia nigra is a group of dark-colored, dopaminergic cells. CN III is the oculomotor nerve. CN IV is the trochlear nerve. Both of these cranial nerves provide innervation for motor movements of the eyes. The cerebral peduncles (cruz cerebri) are two very large bundles of axons which are a continuation of the efferent projections within the internal capsule. The Pons The pons, which is also part of the brain stem, is inferior to the midbrain and superior to the medulla. Its posterior border is separated from the cerebellum by the Aqueduct of Sylvius, and more inferiorly, by the fourth ventricle. Motor and sensory tracts traverse the anterior surface of the pons. The sensory fibers are located behind the motor fibers. The nuclei of cranial nerves V and VI are located in the pons. CN V, or the trigeminal, sends motor messages to the jaw and receives sensory messages from the teeth, tongue, and parts of the face. CN VI, or the abducens, provides motor innervation to the eye. The motor nucleus of cranial nerve VII, the facial nerve, is also located in the pons. This part of the nerve innervates the muscles of facial expression including the eye lids, forehead and the lips. The Medulla Oblongata The medulla is the most inferior part of the brain stem. The cell bodies of the following cranial nerves are located there: Lower Motor Neuron Cell Bodies CN VIII, the auditory nerve CN IX, the glossopharyngeal nerve CN X, the vagus nerve CN XI, the spinal accessory nerve CN XII, the hypoglossal nerve Because the nuclei of the vagus nerve are found in the medulla, it is considered to be a center for circulation and respiration. It is also quite important to swallowing. It controls muscles of the pharynx, larynx and velum for swallowing. (Note: Most of the cranial nerves important for speech and swallowing are located in the medulla.) The Reticular Formation The reticular formation is a set of interconnected nuclei that are located throughout the brain stem. Its dorsal tegmental nuclei are in the midbrain while its central tegmental nuclei are in the pons and its central and inferior nuclei are found in the medulla. The reticular formation has two components: The ascending reticular formation is also called the reticular activating system. It is responsible for the sleep-wake cycle, thus mediating various levels of alertness. This part of the reticular system projects to the mid-line group of the thalamus, which also plays a role in wakefulness. From there, information is sent to the cortex. The descending reticular formation is involved in autonomic nervous system activity as it receives information from the hypothalamus. The descending reticular formation also plays a role in motor movement. Interneurons of the reticular formation receive some of the cortico-bulbar fibers from the motor cortex. It is those fibers that innervate the three cranial nerves involved in eye movement. Other cortico-bulbar fibers innervate cranial nerves directly. The descending reticular nuclei in the brain are involved in reflexive behavior such as coughing and vomiting. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 7. The Cerebellum The Cerebellum Location and Description The cerebellum is located in the posterior fossa of the skull, dorsal to the pons and medulla from which it is separated by the Aqueduct of Sylvius and the fourth ventricle. Like the cerebrum, the cerebellum is covered by cortex and consists of two hemispheres, each of which is divided into lobes. The hemispheres are separated from one another by a thin structure called the vermis, or "worm." The anterior lobe, or paleocerebellum, is the second oldest part of the cerebellum. It receives proprioceptive input from the spinal cord and controls the anti-gravity muscles of the body, thus regulating posture. The posterior lobe, or neocerebellum, is the newest part of the cerebellum. It is involved in the coordination of muscle movement via the inhibition of involuntary movement. Inhibitory neurotransmitters, especially GABA, are found here. This lobe plays an important role in fine motor coordination. The flocculonodular lobe consists of the flocculi, the most ancient part of the cerebellum, and the nodulus, the narrowest and most inferior part of the vermis. This lobe is involved in the maintenance of equilibrium. Four different nuclei are located deep within each cerebellar hemisphere; the dentate nucleus, the emboliform nucleus, the globose nucleus, and the fastigal nucleus. These deep nuclei have axons that project to the brain stem, sending messages out to be conveyed to other parts of the central nervous system. The deep nuclei are regulated by radish-shaped cells located in the cerebellar cortex called Purkinje cells. The Purkinje cells control the output of the cerebellum by inhibiting the firing of the deep nuclei. The Purkinje cells located in the lateral cortex of the cerebellum project to the dentate nuclei, while those in the intermediate cortex synapse with the emboliform and globose nuclei. The fastigial nuclei receive input from Purkinje cells found in the cerebellar cortical covering of the vermis. Importance of the Cerebellum The cerebellum is involved in a feedback loop for muscle movement. When the cortex sends a message for motor movement to the lower motor neurons in the brain stem and spinal cord, it also sends a copy of this message to the cerebellum. This is conveyed from pyramidal fibers in the cortex on the corticopontinecerebeller tract to the cerebellum. In addition, information gets to the cerebellum from muscle spindles, joints and tendons. This information (proprioception and kinesthesia) lets the cerebellum know about the movements that have been executed, so that it can determine how well motor commands coming from the cortex are being carried out. This has ben called its comparator function. The cerebellum plays a major role in the coordination of muscle activity for the production of smooth movement through its connections with the pyramidal and extrapyramidal systems and the descending reticular formation. Due to its role in the coordination of fine motor movements, the cerebellum makes important contributions to the control of rapid, alternating muscle movements necessary for speech and swallowing. The Cerebellar Peduncles Three fiber bundles called peduncles connect the cerebellum to the brain stem. On these tracts, information runs in both directions, with all messages sent and received by the cerebellum traveling on these fibers. The superior cerebellar peduncle or the superior brachium conjuctivum connects the cerebellum to the midbrain and contains efferent fibers from the dentate, emboliform, and globose nuclei. These are the axons that send feedback to the motor cortex in the frontal lobe via the red nucleus in the midbrain and thalamus. Afferent fibers traveling in this peduncle bring proprioceptive information to the cerebellum from the lower body. This information ascends along the spinal cord in the ventrospinocerebellar tract, before entering the cerebellum. The middle cerebellar peduncle or the middle brachium pontis is the largest of the peduncles and links the cerebellum with the pons. Via this connection, the cerebellum receives a copy of the information for muscle movement that the pyramidal tract is carrying down to lower motor neurons. The inferior cerebellar peduncle or the restiform bodies connects the cerebellum with the vestibular nuclei located in the lower pons and medulla and also with the reticular formation. Proprioceptive information from the upper body, travelling along the dorsospinocerebellar tract enters the cerebellum on the inferior cerebellar peduncle. Feedback Pathways Sensory and motor input to the cerebellum: The cerebellum receives both proprioceptive and kinesthetic information from the periphery. It also gets information about the strength and type of muscle movements occurring. Proprioception, according to Love and Webb, 1992, refers to sensory information about pressure, movement, vibration, position, muscle pain, and equilibrium received from muscles, joints and tendons. Kinesthesia is a more specific term than proprioception. It refers to feedback that comes only from muscle spindles. Kinesthesia is the "ability to detect the range and direction of movements of the limbs" (Bhatnager & Andy, 1995, pg. 341). Sensory information ascends to the cerebellum along the spinal cord. The two main tracts that bring information from the periphery to the cerebellum are the ventrospinocerebellar tract and the dorsospinocerebellar tract. The ventrospinocerebellar tract contains proprioceptive fibers from the lower body. (The axons of this tract decussate and travel upward on the contralateral side of the spinal cord for some distance before crossing again and continuing upward ipsilaterally.) The axons of this tract enter the cerebellum on the superior cerebellar peduncle. The dorsospinocerebellar tract takes proprioceptive information from the upper body that reaches the cerebullum on the inferior cerebellar peduncle (restiform body). The reticulocerebellar tract carries messages received by the reticular nuclei in various parts of the brain stem from the cortex, spinal cord, vestibular system and red nucleus. The vestibulocerebellar tract brings information from the semi-circular canals of the inner ear to the cerebellum via the vestibular nucleus located in the lower pons and medulla. These fibers travel to the flocculi on the inferior cerebellar peduncle. Cortical Input The corticopontinecerebellar tract brings motor information to the cerebellum from the frontal lobe. It leaves the precentral gyrus and descends in the internal capsule along with pyramidal tract fibers. Its axons synapse with cells in the pons. These pontine nuclei then send second order neurons to the cerebellum on the middle cerebellar peduncle. The axons of the corticopontine tract bring the cerebellum a copy of the information that the corticobulbar and corticospinal aspects of the pyramidal tract are conveying to the cranial and spinal nerves. Thus, the cerebellum "knows" the details of the messages being sent to lower motor neurons by the upper motor neurons of the pyramidal tract. The messages sent by the corticopontinecerebellar tract includes information about the nature of the motor impulse being sent by the precentral gyrus, its destination, its strength and its speed. This connection is contralateral, meaning that information from the frontal lobe of one hemisphere is sent to the posterior lobe on the opposite side of the brain. The cerebellum acts on the motor messages carried by the corticopontinecerebellar tract. In an unknown fashion, it integrates the information and exerts control over the message through the firing of Purkinje cells. The corticopontinecerebellar tract is considered by many to be part of the extrapyramidal system, although it originates in the precentral and postcentral gyri with pyramidal tract fibers it synapses with cells in the pons rather than with spinal or cranial nerves. The portion of the tract from the cortex to the pons consist of first order neurons; and from the pons to the cerebellum, second order. The first order neurons may be considered by some to be part of the pyramidal system. Output from the cerebellum The tracts discussed below may be considered part of the extrapyramidal system since they carry involuntary, and automatic information and are indirect and multisynaptic. As mentioned previously some sources consider the extrapyramidal tract to consist of the basal ganglia only. Output to the red nucleus The dentate nuclei, which receive inhibitory messages from the Purkinje cells in the lateral cerebellar cortex, and the emboliform and globose nuclei, which are controlled by the Purkinje cells in the intermediate cortical areas all project to the contralateral red nucleus in the midbrain via the superior cerebellar peduncle. Some of the information sent on the superior cerebellar peduncle travels from the red nucleus to the ventrolateral nucleus of the thalamus. From there it passes on to the precentral gyrus of the frontal lobe. Other ascending projections travel to the thalamus directly. These connections allow the cerebellum to perform its comparatot function by providing feedback to the motor cortex regarding the messages that it has received from corticopontinecerebellar fibers as well as feedback from the muscles innervated by lower motor neurons that receive impulses from the pyramidal and extra pyramidal tracts. The Structure of the Feedback Loop for Motor Movement Precentral gyrus --> Pontine nuclei -- > Cerebellum -- > (after input from muscles innervated by lower motor neurons, and comparisons made by the cerebellum between output from the cortex and input from muscles) --> Red nuclei --> Thalamus --> Precentral gyrus where adjustments are made Fibers from the red nucleus also descend to synapse with the spinal nerves, forming the rubrospinal tract. The vestibulospinal tract The fastigal nucleus, which receives inhibitory control from the Purkinje cells of the vermis, sends messages to the vestibular nuclei in the lower pons and medulla. From the vestibular nuclei, information is sent to lower motor neurons in the brain stem and spinal cord. This tract brings information about the body's position in space to the antigravity muscles. The reticulospinal tract This tract carries information related to the functioning of the autonomic nervous system including circulation of the blood, dilation of blood vessels, respiration and visceral activity. (Note that while this tract influences autonomic activity, it is not part of the autonomic nervous system.) Ataxia, which is an incoordination of motor movement, results from cerebellar lesions. The term ataxia is also used to describe the unsteady walk and usual postures seen in patients who have suffered injury to the cerebellum. People sometimes compensate for these problems by walking with their feet far apart in a broad-based gait (Love & Webb,1992). CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1999/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 8. The Spinal Cord, Spinal Nerves, and the Autonomic Nervous System The Spinal Cord Description The spinal cord begins below the medulla and ends just above the small of the back at the conus medularis. Recall that the area within the vertebral column beyond the end of the spinal cord is called the cauda equina. Meninges Recall also that the spinal cord is protected by the vertebrae and the meninges. The dura mater, arachnoid mater and pia mater of the spinal cord are continuous with those of the brain. Cerebrospinal fluid is in the subarachnoid space that lies between the arachnoid and pia mater and in the central canal, a space in the middle of the gray matter of the cord. It provides a hydraulic cushion for the spinal cord. Internal Composition Dorsal (sensory) and ventral (motor) horn cells When the cord is viewed in a cross-section, its gray matter is "H" shaped or, as described by Bhatnagar and Andy,1995, butterfly shaped. It has two ventral and two dorsal horns. The white matter surrounding the cell bodies of the cord is made up of ascending and descending fibers. Motor tracts are found on the ventral and lateral aspects of the cord while sensory tracts run along its dorsal area. Neuronal types Motor neurons These lower motor neurons are located on the ventral aspect of the cord. They are either alpha or gamma cells. Alpha cells are the principle lower motor neurons of the spinal cord and form the main portion of the final common pathway. They conduct rapid motor impulses, with each alpha cell innervating approximately 200 muscle fibers. Gamma neurons are also part of the final common pathway according to some sources but they are only half as numerous as alpha cells. Gamma cells conduct slow motor impulses. Their major function is to stretch muscle spindles. Association neurons Interneurons connect the anterior and posterior horns of the gray matter and are involved in the reflex arc. They work within the same segment of the spinal cord, with a segment being defined as the horizontal section of the cord that gives rise to one pair of spinal nerves. Internuncial Neurons travel between segments, sending projections up to the brain stem and cerebellum. They project in an ascending, not descending manner. These association neurons are found throughout the central nervous system. They are much more numerous than motor neurons; the ratio between the two types of cells is 30:1. The main function of the association neurons in the spinal cord is that of inhibitory control. They also interconnect other cells with one another. Some sources, including Bhatnager and Andy, (1995), do not distinguish between interneurons and internuncial neurons. Even if these two types of association neurons are grouped together, they should definitely be distinguished from the spinal nerves which are lower motor neurons, forming a final common pathway for information descending from the brain. The Spinal Nerves Description There are thirty-one pairs of spinal nerves. These nerves are mixed, having both a sensory and a motor aspect. Their motor fibers begin on the ventral part of the spinal cord at the anterior horns of the gray matter. The roots of their sensory fibers are located on the dorsal side of the spinal cord in the posterior root ganglia. When the motor and sensory fibers exit the spinal column through the intervertebral foramina and pass through the meninges, they join together to form the spinal nerves. Spinal nerves receive only contralateral innervation from first order neurons. Eight pairs of spinal nerves are located in the uppermost, cervical region of the cord: Twelve pairs are found in the thoracic region. Five pairs are in the lumbar area. Five pairs are in the sacral area. One pair is found in the most inferior, coccygeal region. Function These second order lower motor neurons, the spinal nerves, form part of the final common pathway for information traveling from the central nervous system to the periphery. The spinal nerves provide innervation to body areas below the neck while cranial nerves (also second order neurons) carry impulses only to the head and neck, except for the vagus. (You will understand shortly that cranial nerves can be sensory, motor or both). Reflex arc Also, the sensory and motor fibers of the spinal nerves form a reflex arc. This type of reflexive behavior occurs when a message from sensory fibers causes a motor reaction directly, without traveling to the brain. For example, if you touch a hot burner on the stove, sensory information about the temperature of the burner travels along spinal nerves to your spinal cord and are carried directly to their motor nuclei by interneurons; the motor command goes out along the axons of the lower motor neuron causing you to move your hand away from the stove. As messages do not have to travel up to the brain to be processed, reactions mediated by this reflex arc can occur very rapidly. Of course you WILL feel pain shortly thereafter (milliseconds) as the information gets to the parietal lobe via the thalamus The Autonomic Nervous System The autonomic nervous system is involved in the control of the heart, glands and smooth muscles of the body and plays a major role in regulating unconscious, vegetative functions. It works together with the endocrine system to control the secretion of hormones and is itself controlled by the hypothalamus. Because motor fibers make up the bulk of the autonomic system, some anatomists consider it to be purely motoric although it does include some afferent axons that carry information from the viscera. Although the autonomic nervous system is considered to be one of the three main divisions of the human nervous system in its own right, parts of both the central nervous systems and the peripheral nervous systems play a role in its functions. The autonomic nervous system has two components, the sympathetic system and the parasympathetic system. These two aspects have antagonistic functions. Sympathetic System The sympathetic system prepares the body for fight or flight reactions. Action of this system results in accelerated heart rate, increased blood pressure and blood flow away from the periphery and digestive system toward the brain, heart and skeletal muscles. It also causes adrenaline to be released, temporarily increasing physical strength. Parasympathetic System The parasympathetic system brings the body back to a state of equilibrium. It slows heart rate and decreases the release of hormones into the blood stream. The activity of the parasympathetic system causes more localized reactions than does the sympathetic system as much of its output is to specific organs. The autonomic nervous system consists of four chains of nuclei or ganglia, two of which are located on either side of the spinal cord. The outer chains of nuclei form the parasympathetic division of the system while those closest to the spinal cord make up its sympathetic element. Rami communicantes The rami of the autonomic nervous system are the axons of pre-ganglionic and ganglionic cells. Pre-ganglionic cells of the autonomic nervous system are neurons located in some of the cranial nerves of the brain stem and in some of the spinal nerves that project to the ganglionic chains of the autonomic nervous system. The autonomic nervous system is closely connected with the central and peripheral nervous systems due to this arrangement. Ganglionic cells originate within the ganglia. They project to post-ganglionic neurons. Post-ganglionic cells are neurons that are located in the target organs and muscles of the autonomic nervous system. It can be said that the motor pathways of the autonomic nervous system are made up of its pre-ganglionic and ganglionic cells. The fibers and ganglionic chain of the parasympathetic chain are not as welldefined as those of the sympathetic chain. All preganglionic neurons of the sympathetic system synapse with the sympathetic chain. This is not true of the parasympathetic preganglionic cells, however. Some of them synapse with the chain, but others go directly to end organs or muscles. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1999. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 9. The Cranial Nerves (including lower motor neurons involved in swallowing and speech) The cranial nerves innervate the muscles of the jaw, face, tongue, neck, pharynx, and larynx. Some of them are motor, some are sensory and some are mixed nerves, containing both sensory and motor fibers. Six of them are involved in speech and swallowing, and are therefore very important to the speech, language pathologist. The motor nuclei and processes of the cranial nerves are lower motor neurons as they form a final common pathway for information descending from the cerebrum to the periphery. Because the motor roots of the cranial nerves are located in the brain stem, messages from the precentral and postcentral gyri reach the cranial nerves on the corticobulbar portion of the pyramidal tract. With the exception of part of CN VII (the facial nerve) and parts of CN XII the hypoglossal nerve, this innervation is bilateral. Along with the spinal nerves, the cranial nerves make up the peripheral nervous system. There are twelve pairs of cranial nerves. Cranial Nerves Involved in Smell and Vision CN I, the olfactory nerve, is a purely sensory nerve. It has receptors within the mucous membrane of the nose. Information runs posteriorly along the olfactory tract and through the olfactory bulb to the temporal lobe where it is processed. Remember that this is the only sense not mediated by the thalamus. CN II, the optic nerve, is also a sensory nerve. Visual information from the retina is carried back to the superior colliculus of the midbrain on the optic tract which is immediately below the olfactory tract in the more anterior part of the brain. From the superior colliculus, messages are first passed on the lateral geniculate body in the thalamus and then to the cortex of the occipital lobe. CN III is the oculomotor nerve. The nucleus of this motor nerve is located in the midbrain. It mediates movements of the eyeball and constriction and dilation of the pupil. CN IV or the trochlear nerve, is a motor nerve. Its nucleus also lies in the mesencephalon. This nerve also innervates eye movements and damage to it will cause double vision. CN VI is the abducens nerve. The nucleus of this motor nerve is found in the pons. This nerve provides innervation for movements of the eyeball. The Cranial Nerves Involved in Speech, Hearing and Swallowing CN V is the trigeminal nerve. The motor nucleus of this large mixed nerve originates in the pons. It provides motor innervation to the muscles that control the mandible (jaw), the tensor veli palatini muscle of the velum, and the tensor tympani muscle of the middle ear. It mediates sensation from the head, jaw, face, some of the sinuses and tactile sensation from the anterior two thirds of the tongue. CN VII or the facial nerve, is most often classified as a motor nerve, but can also be considered a mixed nerve. Its motor nucleus which is located in the pons innervates all of the muscles of facial expression including those in the forehead, cheeks, and lips, as well as the stapedius muscle of the middle ear. It also sends motor impulses to the rest of the ear; if you can wiggle your ears, this action is mediated by CN VII. The part of the nucleus that sends commands to the upper part of the face receives bilateral (ipsalateral and contralateral) innervation from upper motor neuronal tracts. However, the portion that controls the lower part of the face receives only contralateral (unilateral) innervation. This means that unilateral lesions of the pyramidal tract may have noticeable effects on voluntary movements of the cheeks and lips. Note that involuntary facial expressions of emotion will not be impaired in the case of a pyramidal tract lesion as they are controlled by the extrapyramidal tract. They will be affected by lower motor neuron lesions. The sensory aspect of the facial nerve mediates taste in the anterior two thirds of the tongue. CN VIII is the auditory/vestibular nerve. The two branches of this sensory nerve carry information from the cochlea and from the vestibular end organs in the inner ear. The auditory nerve originates in the medulla. CN IX, or the glossopharyngeal nerve, is a mixed nerve. Its motor aspect contributes to the action of the middle pharyngeal constrictor muscle and innervates the stylopharyngeus muscle. Its sensory aspect carries input from the posterior one third of the tongue, the velum, and the pharynx including the tonsils. CN X is the vagus nerve. This mixed nerve originates in the medulla. One of the motor nuclei of the vagus innervates the majority of the viscera, including the heart, respiratory system, and digestive system. Another motor nucleus sends motor commands to the pharyngeal constrictor muscles and completely controls the intrinsic musculature of the larynx. The superior branch of the vagus innervates the cricothyroid muscle and so is involved in pitch changes. Its recurrent branch innervates all of the other intrinsic laryngeal musculature. The vagus also innervates the glossopalatine and levator veli palatine muscles, making it primarily responsible for palatal functioning. CN XI is the spinal accessory nerve, a motor nerve that originates in the medulla. It innervates the trapezius and sternocleidomastoid muscles of the neck. It also sends some motor messages to the uvula and the levator veli palatine (raises the velum). CN XII, is the hypoglossal nerve, another motor nerve that originates in the medulla. It controls tongue movement, innervating both the intrinsic and extrinsic tongue muscles. The part of the nucleus that innervates the genioglossus, the muscle involved in tongue protrusion, is connected only to contralateral fibers from the pyramidal tract. Thus, unilateral upper motor lesions can affect this type of movement. All other tongue muscles receive bilateral innervation for voluntary movements. If there is damage to the hypoglossal nerve itself, the tongue may reveal the presence of the lower motor neuron lesion by fasiculating (twitching). The Six Cranial Nerves Involved in Speech and Swallowing CN V - - the trigeminal nerve CN VII - - the facial nerve CN IX - - the glossopharyngeal nerve CN X - - the vagus nerve CN XI - - the spinal accessory nerve CN XII - - the hypoglossal nerve Stages of Deglutition (Logemann,1989, 1994. Morrell,1984. In Groher,1984.) In the Oral-preparatory stage, food is moved around the mouth, chewed and tasted. Time needed for this stage is variable. During the oral stage, food is moved to the back of the mouth by the tongue. This stage lasts for about 1 second, ending when the bolus touches the back of the oropharynx. In the first part of the pharyngeal stage, or laryngeal substage, a number of things happen simultaneously. The larynx moves up and forward, the vocal folds approximate and the epiglottis falls over the top of the larynx. During the second part of this stage, the bolus of food is moved down the pharynx by the stripping action of the pharyngeal constrictor muscles. This phase of the pharyngeal stage ends when the cricopharyngus muscle, also known as the p.e. segment, opens, allowing the food to enter the esophagus. It is not known what leads the p.e. segment to open at the right time during the swallowing sequence. Some think that it opens due to stretching caused by the elevation of the larynx. The total time required for the entire pharyngeal phase is 1 second. During the esophageal stage, food travels down the esophagus via a wave-like motion called peristalsis. This phase lasts between 8 and 20 seconds. Note that peristalsis occurs only in the esophagus and colon, not in the pharynx. In some older literature on swallowing, there is confusion about this, and the stripping action of the pharyngeal constrictor muscles is mislabeled as peristalsis. Swallowing is not just a motor reflex; it requires a combination of sensory and motor control. The swallowing center, which is located in the nucleus ambiguous of the medulla, recognizes a pattern that consists of both sensory and motor elements and then triggers the swallowing response. This is why the swallow is considered to be a type of patterned response or pattern recognition system. Sensory input that initiates the swallow response comes from the trigeminal, facial and glossopharyngeal nerves. The trigeminal nerve is involved in the oral preparatory and oral stages. It provides the innervation that controls jaw movement for chewing and it also mediates tactile sensation in the anterior two thirds of the tongue. During the oral preparatory and oral stages, taste is carried from the anterior two thirds of the tongue by the facial nerve. The glossopharyngeal nerve is responsible for taste in the posterior one third of the tongue and for tactile sensation to the posterior part of the oral cavity, including the velum, tonsils, and walls of the oropharynx. It provides the feedback that is most important in the elicitation of the swallow. Feedback from motor movements, especially tongue movements which are mediated by the hypoglossal nerve, also help to trigger the swallow. Input from both the cerebral cortex and the cerebellum is responsible for the coordination and timing of the motor movements involved in swallowing. Mnemonic for the Cranial Nerves On (olfactory) Some (sensory) Old (optic) Say (sensory) Olympus's (oculomotor) Marry (motor) Towering (trochlear) Money (motor) Top (trigeminal) But (both) A (abducens) My (motor) Finn (facial) Mother* (motor) And (auditory) Says (sensory) German (glossopharyngeal) Bad (both) Vended (vagus) Business (both) At (accessory) Marry (motor) Hopps (hypoglossal) Money (motor) The facial nerve could also be classified as both sensory (taste for anterior two thirds of tongue) and motor, in which case the word in this part of the rhyme would change to "brother." It is usually classified as a motor nerve. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 10. Upper Motor Neuronal Tracts In order to reach the muscles of the body, motor commands generated in the central nervous system must travel down upper motor neurons and lower motor neurons. Upper motor neurons are a type of first order neuron. They are unable to leave the central nervous system. The pyramidal tract is a very important upper motor neuron tract. The extrapyramidal tract also consists of upper motor neurons. As upper motor neurons must remain inside the neuraxis, they synapse with neurons of another type called lower motor neurons which can carry messages to the muscles of the rest of the body. Lower motor neurons, or second order neurons are cranial and spinal nerves. The cell bodies of these neurons are located in the neuraxis, but their axons can leave the central nervous system and synapse with the muscles of the body. All lower motor neurons are either spinal or cranial nerves. All spinal nerves have a lower motor neuron component as they are mixed nerves. However, not all cranial nerves have lower motor neuron components. Some of the cranial nerves contain only sensory fibers and therefore cannot be classified as lower motor neurons. For example, CN I, the olfactory nerve, CN II the optic nerve, and CN VIII, the auditory nerve, do not have motor components. The Pyramidal Tract This group of fibers carries messages for voluntary motor movement to the lower motor neurons in the brain stem and spinal cord. Approximately 80% of the cell bodies of the pyramidal tract are located on the precentral gyrus of the frontal lobe, which is also known as the motor strip. Particularly large cells located here whose axons are part of the pyramidal tract are called pyramidal cells. Approximately 20% of the pyramidal tract fibers also originate in the postcentral gyrus of the parietal lobe, in Brodmann's areas 1, 2, and 3. Regardless of the location of their cell bodies, pyramidal tract fibers descend from the cortex inside the internal capsule. This tract is direct and monosynaptic, meaning that the axons of its neurons do not synapse with other cells until they reach their final destination in the brain stem or spinal cord. These direct connections between the cortex and the lower motor neurons allow messages to be transmitted very rapidly from the central nervous system to the periphery. The fibers of the pyramidal tract that synapse with spinal nerves sending information about voluntary movement to the skeletal muscles form the corticospinal tract. These axons are among the longest in the central nervous system, as some of them travel all the way from the cortex to the inferior part of the spinal cord. As they descend through the brain, they form part of the posterior limb of the internal capsule. At the pyramids in the inferior part of the medulla, eighty-five to ninety percent of corticospinal fibers decussate, or cross to the other side of the brain. The remaining ten to fifteen percent continue to descend ipsilaterally. The fibers that decussate are called the lateral corticospinal tract or the lateral pyramidal tract. Because they descend along the sides of the spinal cord, the uncrossed or direct fibers that synapse with spinal nerves on the ipsilateral side of the body are called the direct pyramidal tract. They may also be referred to as the ventral pyramidal tract or the anterior corticospinal tract since they travel down the ventral aspect of the spinal cord. The spinal nerves receive only contralateral innervation from the corticospinal tract. This means that unilateral pyramidal tract lesions above the point of decussation in the pyramids will cause paralysis of the muscles served by the spinal nerves on the opposite side of the body. For example, a lesion on the left pyramidal tract could cause paralysis on the right side of the body. The fibers of the pyramidal tract that synapse with cranial nerves located in the brain stem form the corticobulbar tract. Obviously, this is the part of the pyramidal tract that carries the motor messages that are most important for speech and swallowing. Corticobulbar axons descend from the cortex within the genu or bend of the internal capsule. Almost all of the cranial nerves receive bilateral innervation from the fibers of the pyramidal tract. This means that both the left and right members of a pair of cranial nerves are innervated by the motor strip areas of both the left and right hemispheres. This redundancy is a safety mechanism. If there is a unilateral lesion on the pyramidal tract, both sides of body areas connected to cranial nerves will continue to receive motor messages from the cortex. The message for movement may not be quite as strong as it was previously but paralysis will not occur. The two exceptions to this pattern are the portion of CN XII that provides innervation for tongue protrusion and the part of CN VII that innervates the muscles of the lower face. These only receive contralateral innervation from the pyramidal tract. This means that they get information only from fibers on the opposite side of the brain. Therefore, a unilateral upper motor neuron lesion could cause a unilateral facial droop or problems with tongue protrusion on the opposite side of the body. For example, a lesion on the left pyramidal tract fibers may cause the right side of the lower face to droop and lead to difficulty in protruding the right side of the tongue. The other cranial nerves involved in speech and swallowing would continue to function almost normally as both members of each pair of nuclei still receives messages from the motor strip. Because most cranial nerves receive bilateral innervation, lesions of the upper motor neurons of the pyramidal tract must be bilateral in order to cause a serious speech problem. (The effects of the inability to protrude the tongue and of paralysis of the lower face on speech are negligible.) On the other hand, unilateral lesions of the lower motor neurons may cause paralysis. This occurs because the lower motor neurons are the final common pathway for neural messages traveling to the muscles of the body. At the level of the lower motor neurons, there is no alternative route which will allow messages from the brain to reach the periphery. Muscles on the same side of the body as the lesion will be affected. Lesions on the cranial nerve nuclei located in the brain stem are called bulbar lesions. The paralysis that they produce is called bulbar palsy. Lesions to the axons of the cranial nerves are called peripheral lesions. As cranial nerves are lower motor neurons, both bulbar and peripheral lesions are lesions of the final common pathway. When bilateral lesions of the upper motor neurons of the pyramidal tract occur, they produce a paralysis resembles that which occurs in bulbar palsy. For this reason, the condition is known as pseudo-bulbar palsy. If a lesion occurs in the brain stem and damages both the nucleus of a cranial nerve and one side of the upper motor neurons of the pyramidal tract, a condition known as alternating hemiplegia may result. This involves paralysis of different structures on each side of the body. The lesioning of the nucleus of the cranial nerve will cause a paralysis of the structures served by that nerve on the same side of the body as the injury. Because the pyramidal tract provides only contralateral innervation to the spinal nerves, damage to the upper motor neurons will meanwhile cause a paralysis of different structures on the other side of the body. For example, a lesion that affected the right nucleus of the trigeminal cranial nerve and the right side of the pyramidal tract would cause paralysis of the right side of the jaw and of part of the left side of the body. Both the corticospinal and corticobulbar tracts send some axons to the pontine nuclei in the pons as they descend to synapse with lower motor neurons. These fibers that end in the pons form the corticopontine tract. This pathway carries information about the type and strength of the motor impulses generated in the cortex to the cerebellum. While the corticopontine fibers actually end in the pontine nuclei, second order neurons carry their message to the cerebellum via the middle cerebellar peduncle. This tract may be considered to be a part of the extrapyramidal system rather than a component of the pyramidal tract since it does not synapse directly with lower motor neurons. The Extrapyramidal Tract This system is involved in automatic motor movements, and in gross rather than fine motor movement. It works with the autonomic nervous system to help with posture and muscle tone and has more influence over midline structures than over those in the periphery. Facial expression is one important communicative behavior that is mediated by the extrapyramidal tract. In contrast to pyramidal tract, the extrapyramidal tract is an indirect, multisynaptic tract. Components of the extrapyramidal tract include the basal ganglia, the red nucleus, the substantia nigra, the reticular formation and the cerebellum. All of these structures send information to the lower motor neurons in an indirect fashion. Some sources, including the text by Love and Webb, 1992, consider the basal ganglia to be the sole constituent of the extrapyramidal system, saying that the other structures listed above synapse with the extrapyramidal tract but are not part of it. The basal ganglia acts to inhibit the release phenomenon, or the rapid firing of motor neurons. It is aided in this function by the substantia nigra of the midbrain. The muscles most often affected by this inhibitory functions are those controlling the head, the hands, and the fingers. The neurotransmitters involved in the inhibitory function of the basal ganglia include dopamine, which is produced by the substantia nigra, acetylcholine, and GABA (gamma amino butric acid), which is a glutamate. Dopamine is an especially powerful inhibitor. Extrapyramidal Projections to Lower Motor Neurons The extrapyramidal tract has an important role in motor movement. it has projections that carry autonomic motor impulses to voluntary muscles in the body, including the muscles for speech and swallowing. During speech, muscles are receiving input from both the pyramidal and extrapyramidal systems. it is involved in gross motor movement rather than fine. It is responsible for facial expression such as sadness, irony and happiness. The rubrospinal tract passes through the red nucleus. The cerebellum sends messages to the spinal nerves along this tract. Information flows from the superior cerebellar peduncle to the red nucleus and finally to the spinal nerves. This information is very important for somatic motor, or skeletal muscle control and the regulation of muscle tone for posture. The reticulospinal tract runs from the reticular nuclei of the pons and medulla to the spinal nerves. It is involved in somatic motor control like the rubrospinal tract and also plays an important role in the control of autonomic functions. The tectospinal tract has points of origin throughout the brain stem, but especially in the midbrain area, and ends in the spinal nerves. It is involved in the control of neck muscles and also in visual and auditory reflexes. So, when you jump after hearing a noise or duck when you see something coming toward you, this tract helps to mediate these reactions. The vestibulospinal tract runs from the vestibular nuclei located in the lower pons and medulla to the spinal nerves. It is involved in balance. (Note that all of these tracts receive input from the cerebellum.) Extrapyramidal Diseases and Syndromes Affecting Communication/Swallowing Lesions in the extrapyramidal tract cause various types of diskinesias or disorders of involuntary movement. The problems most commonly affecting the extrapyramidal tract include degenerative diseases, encephalitis, and tumors. Parkinson's Disease, which is a degenerative disease, is probably the most frequently occurring illness that results from extrapyramidal tract lesions. It occurs when the dopaminergic neurons of the substantia nigra are destroyed. Its symptoms include: Tremor Festinating movements, especially a festinating gait. (Festinating movements are movements which become increasingly rapid and uncontrolled). Hypokinetic dysarthria Weak Voice Mask-like facial expression Diseases associated specifically with lesions of the basal ganglia include Huntington's Chorea and Sydenham's Chorea. The term "chorea" comes from the Greek "khoros" which means dance. Both of these diseases are associated with jerky, uncontrolled movements of the limbs. Sydenham's chorea was probably the cause of the malady that was known as St. Vitus' Dance during the middle ages. Huntington's Chorea is an inherited degenerative disease. Sydenham's tends to clear up spontaneously. Essential Tremor Syndrome, which is associated with Spastic Dysphonia may also be the result of basal ganglia lesions. Lesions of the basal ganglia will also cause hyperkinetic dysarthria. Note that not only is the definition of the extrapyramidal system controversial, but also many sources say that it is very difficult to make functional distinctions between the extrapyramidal and pyramidal systems. When upper motor neuron lesions occur, it is often difficult to determine which tract has been damaged. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 11. The Blood Supply The Blood Supply The Blood Supply The Blood Supply Medial View The Blood Supply Lateral View Blood transports oxygen and other nutrients necessary for the health of neurons, so a constant flow of blood to the brain must be maintained. According to Love and Webb,1992, the brain uses approximately twenty percent of the body's blood and needs twenty-five percent of the body's oxygen supply to function optimally. Blood flow in a healthy person is 54 milliliters per 1000 grams of brain weight per minute. There are 740 milliliters of blood circulating in the brain every minute. 3.3 milliliters of oxygen are used per minute by every 1000 grams of brain tissue. This means that approximately 46 milliliters of oxygen are used by the entire brain in one minute. During sleep, blood flow to the brain is increased, but the rate of oxygen consumption remains the same. Subclavian Artery The main artery of the body is called the aorta. It supplies blood to all parts of the body with the exception of the lungs. The aorta ascends from the heart and forms an arch, from which arise two subclavian arteries. Each subclavian has two main branches, the common carotid and the vertebral. Both of these carry blood to the brain. Each common carotid divides into an external carotid artery, which supplies blood to the face and an internal carotid artery which supplies the brain with blood. The external carotid is a fairly straight artery, so it is not prone to blockages due to the build up of cholesterol. Even if a blockage does occur, it would obviously not cause a stroke as this artery does not carry blood to the brain. The Internal Carotid Each internal carotid artery ascends along one side of the neck. They pass behind the ear into the temporal lobe and enter the subarachnoid space. Then, they run posteriorly to the medial end of the fissure of Sylvius where they bifurcate into two main branches, the anterior cerebral artery and the middle cerebral artery. As the internal carotids have many twists and turns, there are many places where plaque can build up, causing a blockage. Such blockages can be identified by sonogram (non-invasive), or by angiograms (invasive). Also, a sound called a bruit can sometimes be heard via stethoscope when a blockage exists. The anterior cerebral artery goes above the optic chiasm to the medial surface of the cerebral hemispheres. It arches around the genu (horn) of the corpus callosum (FitzGerald, 1996). It supplies blood to the medial cortex, including the medial aspect of the motor strip and the sensory strip. This means that damage to the anterior cerebral artery can cause sensory and motor impairment in the lower body. For example, a patient who has had a stroke affecting this artery may be incontinent or have unilateral paralysis from the hips on down. The anterior cerebral artery also delivers blood to some parts of the frontal lobe and corpus striatum. So a blockage in this artery can affect cognition and cause motoric problems due to damage to fibers in the internal capsule or to the basal ganglia. The other main branch of the internal carotids is the middle cerebral artery. This large artery has tree-like branches that bring blood to the entire lateral aspect of each hemisphere. This means that this artery supplies blood to the cortical areas involved in speech, swallowing and language, including the lateral motor strip, lateral sensory strip, Broca's area, Wernicke's area, Heschl's gyrus, and the angular gyrus. In addition, it provides most of the blood supply to the corpus striatum. If a patient has a blockage in the middle cerebral artery, it is probable that s/he will have aphasia. S/ he will probably also have impaired cognition and corticohyposthesia, or numbness, on the opposite side of the body. Problems with hearing and the sense of smell may also result from damage to this artery because it supplies the lateral surface of the temporal lobe. The central branches of the middle cerebral are the medial and lateral striata arteries. The striata supply the basal ganglia, internal capsule, and thalamus (FitzGerald, 1996). Because they are the main blood supply to the internal capsule, they are called by some the arteries of stroke. When something happens to these arteries, the bottleneck of fibers within the internal capsule can be damaged, causing many disabilities. The striata are very thin arteries and blood pressure within it high. For this reason, they are cobsidered by many to be more vulnerable to hemorrhages than to blockages, although FitzGerald says that occlusion of one of these areteries is the major cause of of classical stroke where pyramidal tract damage results in contralateral hemiplegia. Other arteries which arise from the internal carotid arteries include the anterior communicating artery and the posterior communicating arteries. The anterior communicating artery joins the anterior cerebral arteries of each hemisphere together. The posterior communicating arteries join the middle cerebral arteries to the posterior cerebral arteries, which are part of the basilar artery system. The Vertebral Artery Both of the vertebral arteries ascend through the spinal column and enter the brain through the magnum foramen. Once in the brain, they continue to ascend, traveling beside the brain stem. At the lower border of the pons the two vertebral arteries join together to form the basilar artery or vertebrobasilar artery. The vertebral arteries and the basilar are straight arteries and therefore not as subject to blockages due to the build up of cholesterol as are the internal carotids. The posterior inferior cerebellar not only supply the cerebellum but take blood to the lateral medulla. Anterior and posterior spinal arteries the ventral and dorsal medulla, respectively (FitzGerald 1996). The three arteries are branches of the vertebral. The side of the pons and the cerebellum receive blood from the anterior inferior cerebellar artery and the superior cerebellar artery. These arteries are branches of the basilar. The anterior inferior cerebellar artery also has a branch, the labyrinthine artery, that supplies the inner ear. The basilar also gives off about twelve pontine arteries that supply the medial pons (FitzGerald, 1996). At the superior border of the pons, the basilar artery divides to form the two posterior cerebral arteries. Before the basilar artery divides, several other arteries arise from it. These include the anterior, inferior, and posterior cerebellar arteries as well as pontine branches. So, the cerebellum and pons are supplied by branches of the basilar. The posterior cerebral arteries supply the part of the brain found in the posterior fossa of the skull, including the medial area of the occipital lobes and the inferior aspects of the temporal lobes. They also supply yhe midbrain and deliver blood to the thalamus and some other subcortical structures. Blockages in this artery can affect the sense of smell, and cause cranial nerve damage, as well as visual problems, including visual agnosia, hemianopsia and alexia. The choroidal arteries, which arise both from the divisions of the internal carotid arteries and from the basilar system, supply blood to the choriod plexuses and also to the hippocampus. Blockages in these arteries can affect the production of cerebrospinal fluid and can also cause memory problems. The Circle of Willis The Circle of Willis or the Circulus Arteriosus is the main arterial anastomatic trunk of the brain. According to Bhatnagar and Andy, 1995, anastomosis occurs when blood vessels bring blood to one spot from which it is then redistributed. The Circle of Willis is a point where the blood carried by the two internal carotids and the basilar system comes together and then is redistributed by the anterior, middle, and posterior cerebral arteries. The anterior cerebral arteries of the two hemispheres are joined together by the anterior communicating artery. The middle cerebral arteries are linked to the posterior cerebral arteries by the posterior communicating arteries. This anastamosis or communication between arteries make collateral circulation which Love and Webb, 1995, define as "the flow of blood through an alternate route" (p. 40) possible. This is a safety mechanism, allowing brain areas to continue receiving adequate blood supply even when there is a blockage somewhere in an arterial system. The blood streams of the internal carotid system and the basilar system meet in the posterior communicating arteries. If there are no problems in either system, the pressure of the streams will be equal and they will not mix. However, if there is a blockage in one of them blood will flow from the intact artery to the damaged one, preventing a cerebral vascular accident. As long as the Circle of Willis can maintain blood pressure at fifty percent of normal, no infarction or death of tissue will occur in an area where a blockage exists. If collateral circulation is good, no permanent effects may result from a blockage. Sometimes, an adjustment time is required before collateral circulation can reach a level that supports normal functioning; the communicating arteries will enlarge as blood flow through them increases. In such cases, a transient ischemic attack may occur, meaning that parts of the brain are temporarily deprived of oxygen. Some people lack one of the communicating arteries that form the Circle of Willis. In this case, if a blockage develops, collateral circulation will be impeded and the collateral blood supply will be compromised, causing brain damage to occur. There are some watershed areas in the brain located at the ends of the vascular systems. Problems with blood supply are particularly likely to occur here, especially in those who have hardening of the arteries. Blockages in the water shed areas can cause transcortical aphasia. Extraneural Factors Affecting the Blood Supply to the Brain Low or High Blood Pressure Abnormally low blood pressure can cause brain damage. This may occur as a result of surgical shock which involves blood pressure as low as 70 milliliters per kilogram of tissue. Hypertension, or blood pressure that remains high regardless of activity level, can cause arteries to narrow over time. Cerebrovascular Resistance Cerebrovascular resistance makes collateral circulation more difficult. It can be caused by an arterial spasm (remember that arteries are lined with muscles). Another potential cause of resistance is increased viscosity of the blood, which can result from leukemia or from high levels of tri-glycerides in the blood, or other factors such as increased red blood cells, often seen in chronic obstructive pulmonary disease (COPD). Increased cerebrospinal fluid pressure can also lead to high levels of cerebrovascular resistance. Atherosclerosis This is hardening of the arteries which often occurs with old age but can also happen in young people. An occluded artery may cause a stroke due to one of the extraneural factors listed above which can compromise the overall integrity of the cerebrovascular system. If an individual does not have one of these problems and has a sufficient number of communicating arteries, a blockage may not have a significant effect on blood supply throughout the brain. The Blood-Brain Barrier Many substances present in the blood supply are unable to pass through the meninges into the cells of the central nervous system. The blood brain barrier includes two components, the blood/ cerebrospinal fluid barrier and the arachnoid barrier layer. Cerebrospinal fluid is a filtrate of blood by the choroid plexuses (capillary networks) of the ventricles which are formed by fusion of the pia mater and the ependyma (ventricular lining). In the course of this process, not all components of blood are allowed to enter the brain. According to Webster, 1999, only clear plasma passes through, leaving blood cells behind. The arachnoid barrier layer is a part of the arachnoid meningeal layer. It is formed by tight junctions between the endothelial cells of cerebral capillaries in the arachnoid mater. Glucose diffuses across the blood-brain barrier through a process that is like selective osmosis. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable. The Neuroscience on the Web Series: SPPA 362, Neuroanatomy of Speech, Swallowing and Language CSU, Chico, Patrick McCaffrey, Ph.D Unit 12. Neurochemistry The Nerve Cell Image of a Nerve Cell The nerve cells of the central and peripheral nervous systems are called neurons. Most neurons have three parts; an axon, a cell body or soma, and dendrites. All neurons have one soma and one axon, but while some neurons have many dendrites, others have none. Neurons vary in size; the smallest have a diameter of 5 microns while the largest are approximately 100 microns in width. (A micron is one one-thousandth of a millimeter). The soma of a neuron contains the cell's nucleus and cytoplasm, a jelly-like substance that surrounds the nucleus. Chromosomes, which consist of molecules of DNA (deoxyribonucleic acid) are found in the nucleus of the neuron. RNA (ribonucleic acid) molecules are also located within the nucleus. Nissl Bodies or Nissl Substances, which also contain RNA, and Golgi Apparati are found in the cytoplasm. DNA forms the genetic code that determines the cell's function. As DNA cannot pass through the nuclear membrane, its commands are carried to the cytoplasm by messenger RNA which can travel outside of the nucleus. These RNA molecules link up with the Nissl substances, connecting to them as a key would fit into a lock. This combination causes the cell to work, or use glucose. The waste products generated by this process are removed from the cell by the Golgi apparati. The axon allows the neuron to send messages to other nerve cells. Each neuron has only one axon, but this may have numerous branches which connect the cell to many other. Axons vary in length; the axons of some pyramidal cells in the precentral gyrus are long enough to travel all the way down to the end of the spinal cord but other axons are very short. Cells can be classified based on the length of their axons. Golgi Type I neurons have long axons while those of Golgi Type II cells are short. Axons arise from an area on the cell body of the neuron called the axon hillock. Most axons form many branches as they extend away from the soma. At its end, each axonal branch divides into a number of telodendria. The boutones terminaux or boutones de passage, which contain neurotransmitters, are located on the telodendria. Myelin Many of the axons in the central and peripheral nervous systems are covered at regular intervals with a fatty insulating substance called myelin. The segments of the axon which lie between areas of myelin and are therefore in direct contact with extracellular fluid are called the Nodes of Ranvier. Myelin coating increases the speed with which an axon can transmit messages. The neural impulse travels by a process known as saltatory conduction, jumping from one unmyelinated segment to the next. This means that the impulse does not have to be propagated through the entire area of the axon. Dendrites The dendrites of a neuron receive messages from the axons of other nerve cells. There are two types of dendrites, apical dendrites and basilar dendrites. Apical dendrites have stalks filled with cytoplasm that appear to be part of the soma of the neuron to which they are attached. Most of these dendrites are found in the cerebral cortex. Basilar dendrites do not have a stalk. They are more numerous than apical dendrites. Links with relevant information about nerve cells: Image of a Nerve Cell For more information on Nerves and nerve cells visit the following web sites Neural Definitions: http://psych.hanover.edu/Krantz/neural/neurldef.html A Self-Quiz: http://psych.hanover.edu/Krantz/neural/struct3.html The Transmission of Neural Messages The Action Potential The messages conducted along axons are electrochemical in nature. Four different types of ions, or electrically charged atoms, are involved in the transmission of neural impulses; chloride ions (Cl -), sodium ions (Na+), potassium ions (K+), and organic anions (A-). When a neuron is at rest, there are high concentrations of A- and K+ within the cell, while most Na+ and Cl- ions are located outside its membrane. The resting potential of the neuron is -70 millivolts, meaning that the electrical charge of the cell is slightly negative in comparison to that of the extracellular fluid surrounding it. This arrangement is due to the selective permeability of the neural membrane. Cl- and K+ can pass through the membrane, but Cl- does not enter the cell in great quantities because both it and the interior of the neuron are negatively charged. Organic anions cannot move through the membrane due to their large size. As Na+ is positively charged and the interior of the nerve cell has a negative electrical potential, these ions should pass into the cell. However, sodium cannot readily pass through the membrane and most of the Na+ ions that do enter the cell are extruded by the sodium-potassium pump. This is the name given to a group of molecules located in the cellular membrane which push Na+ out of the cell and draw K+ ions inside. The resting potential of a neuron changes when messages are received from other nerve cells. Inhibitory impulses cause the electrical charge of the neuron to become even more negative, decreasing its ability to fire, or send messages to other nerve cells. When excitatory messages are received, however, the permeability of the cell membrane changes, allowing Na+ ions to enter the neuron. This influx of positively charged ions causes the cell's electrical potential to temporarily become positive, peaking at +40 millivolts. This change of the electrical potential of the cell from negative to positive is the action potential, which ultimately causes the cell to fire. When the charge of the cell reaches its positive peak, K+ ions are forced out of the neuron because they are positively charged. The exit of these ions causes the potential of the cell to become negative again, temporarily dipping below -70 millivolts before returning to resting potential. After the cell fires, it goes through a refractory period during which it will not fire again. The refractory period may be divided into two phases, the absolute refractory period and the relative refractory period. During the absolute refractory period, the neuron will not fire again, no matter how strong the excitatory messages that it receives. The cell will fire during the relative refractory period, but only if it receives a very strong stimulus. Neurotransmitters When a neuron fires, it communicates with other nerve cells through the release of chemicals called neurotransmitters. Neurotransmitters are found in the boutons terminaux located on the teledendria of the axons. They are stored in circular or oval-shaped capsules called synaptic vesicles. When an excitatory impulse of sufficient strength reaches the teledendria, the synaptic vesicles fuse with the axonal membrane and open, spilling the chemicals they contain into the extracellular fluid. The neurotransmitters then travel across a small space called the synaptic cleft to attach to the neuron that will receive their message. After a message has been sent, excess quantities of the neurotransmitter that remain in the synaptic cleft must be cleared away in order to allow further communication between the cells. In some cases, the left-over chemicals are recycled; they are picked up and repackaged in new synaptic vesicles to be used in future transmissions. Other types of neurotransmitters are destroyed by enzymes when they remain in the synaptic cleft. Acetylcholine (ACh) is one neurotransmitter that has been well-studied. It is the major neurotransmitter of the peripheral nervous system and is also present in the central nervous system. It carries messages controlling voluntary muscle movement, as the nerve fibers located in muscles and in the spinal and cranial nerves are acetylcholinergic. After messages have been transmitted, ACh is broken down in the synaptic cleft by an enzyme called acetylcholinesterase. An insufficient supply of acetylcholine, whether due to excess quantities of acetylcholinesterase or resulting from inadequate synthesis of the chemical, causes Myasthenia Gravis. In this disorder, the strength of neural impulses is attenuated, causes the voluntary movement of muscles, including those involved in articulation, voicing and respiration, to be weakened. Myasthenia Gravis is distinguished from other disorders like ALS by the administration of a derivative of curare. This drug will temporarily improve the strength of someone suffering from Myasthenia Gravis, but will have no effect in cases of ALS. Myasthenia Gravis should not be confused with Myasthenia Laryngis, which is a localized weakness in the larynx and is not the result of a neurological condition. Other known neurotransmitters include two groups of chemicals called the monoamines and the peptides. The monoamines, which include dopamine, norepinephrin, and serotonin, are all synthesized from proteins called amino acids. The peptides, including enkephalin, endorphins, and substance P, are large molecules which may be involved in blocking sensations of pain. Consequences of Neuronal Damage When cells in the rest of the body are injured, they can regenerate and repair themselves. Neurons, on the other hand, do not have this capability. If an axon is damaged, the soma of the cell may also degenerate. Also, when a cell body is injured or an axon is severed from the soma, Wallerian Degeneration, or death of the axon occurs. When a neuron dies, it is ingested by one of the support cells in the nervous system. This process is called phagocytosis. CSU Chico | Glossary | References | Neuroscience on the Web | SPPA362 Home | Next Other courses in the Neuroscience on the Web series: SPPA 336, Neuropathologies of Language and Cognition | SPPA 342 (Neuropathologies of Swallowing and Speech) Copyright, 1998/2001. Patrick McCaffrey, Ph.D. This page is freely distributable.