Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Management of acute coronary syndrome wikipedia , lookup
Aortic stenosis wikipedia , lookup
Coronary artery disease wikipedia , lookup
Cardiac surgery wikipedia , lookup
Artificial heart valve wikipedia , lookup
Lutembacher's syndrome wikipedia , lookup
Mitral insufficiency wikipedia , lookup
Quantium Medical Cardiac Output wikipedia , lookup
Hypertrophic cardiomyopathy wikipedia , lookup
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
Atrial septal defect wikipedia , lookup
Dextro-Transposition of the great arteries wikipedia , lookup
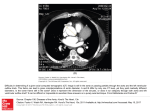
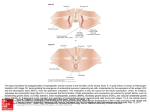
Sox4-Deficiency Syndrome in Mice Is an Animal Model for Common Trunk Jing Ya, Marco W. Schilham, Piet A.J. de Boer, Antoon F.M. Moorman, Hans Clevers, Wouter H. Lamers Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 Abstract—Embryonic mice lacking functional Sox4 transcription factor die from cardiac failure at embryonic day (ED) 14. Heart morphogenesis in these embryos was analyzed in hematoxylin-azophlochsin or immunohistochemically stained, 3-dimensionally reconstructed serial sections between ED12 and ED14. Although Sox4 is expressed in the endocardially derived tissue of both the outflow tract and atrioventricular canal, Sox4-deficient hearts only suffer from defective transformation of the endocardial ridges into semilunar valves and from lack of fusion of these ridges, usually resulting in common trunk, although the least affected hearts should be classified as having a large infundibular septal defect. The more serious cases are, in addition, characterized by an abnormal number and position of the semilunar valve-leaflet anlagen, a configuration of the ridges typical for transposition of the great arteries (with linear rather than spiral course of both ridges and posterior position of the pulmonary trunk at the level of the valve), and variable size of the aorta relative to the pulmonary trunk. The coronary arteries always originated from the aorta, irrespective of its position relative to the pulmonary trunk. The restriction of the malformations to the arterial pole implies that the interaction between the endocardially derived tissue of the outflow tract and the neural crest– derived myofibroblasts determines proper development of the arterial pole. (Circ Res. 1998;83:986-994.) Key Words: mouse n semilunar valve n transposition n common trunk n outflow tract development of the semilunar valves.4 Homozygous Sox4deficient embryos could not be distinguished externally from their littermates until the onset of valvular morphogenesis in the arterial pole of the heart (ED12 to ED13). Then, they rapidly developed generalized edema to die at ED14. Dying mutant embryos typically displayed an oscillatory, nearly futile movement of blood in the arterial pole of the heart, showing that the valvular function of the outflow tract was affected. A preliminary histological examination revealed that the embryos suffered from common arterial trunk and defective semilunar valve development.4 We now show that Sox4 deficiency causes a syndrome with, in addition to impaired semilunar valve development, malformations ranging from infundibular septum defect to complete transposition of the great arteries. Common trunk is a still poorly understood malformation of the ventriculo-arterial junction. Due to the lack of an established concept of ventriculo-arterial development and malformations, some opt for purely descriptive definitions and use the term “common trunk” to mean “single arterial trunk that leaves the heart by way of a single arterial valve and gives rise directly to the coronary, systemic, and one or both pulmonary arteries.”8,9 Others take a more ontogenetic view and define common trunk as the most severe form of a C ongenital malformations of the heart are the most common birth defects in man, accounting for 0.5% to 1% of live births, 10% of stillbirths, and possibly up to 20% of spontaneous abortions.1–3 Thus far, the systematic analysis of these defects has rested almost entirely on findings in human neonates with these structural anomalies. As a consequence, little direct information is available about the etiology and pathogenesis of the malformations. In particular, it is usually not known to what extent a particular malformation can be traced to a single morphogenetic event or process. However, recent progress in molecular and developmental biology has changed the general approach for the study of cardiovascular diseases profoundly. In particular, the technique to genetically modify the expression of genes in vivo has proven to be a powerful approach to identify genes that are involved in the development of specific cardiac malformations. A case in point is the Sox4-null mutant.4 The Sox4 gene is a member of the Sox (for Sry-box) family of transcription factors, of which more than a dozen members have been identified in the mouse.5,6 The Sox4 protein, which binds to the sequence motif AACAAAG,7 has been shown to be a transcriptional activator in immature B- and T-lymphocytes in adult mice.7 However, gene disruption in mice resulted in cardiac failure because of the defective Received June 1, 1998; accepted August 20, 1998. From the Department of Anatomy and Embryology (J.Y., P.A.J.d.B., A.F.M.M., W.H.L.), Academic Medical Center, University of Amsterdam, and the Department of Immunology (M.W.S., H.C.), University of Utrecht, The Netherlands. Correspondence to Wouter H. Lamers, Department of Anatomy and Embryology, Academic Medical Center, University of Amsterdam, Meibergdreef 15, 1105 AZ, Amsterdam, The Netherlands. E-mail [email protected] © 1998 American Heart Association, Inc. Circulation Research is available at http://www.circresaha.org 986 Ya et al Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 “trunco-conal septal defect,”10 whereas others have been intrigued by its similarity with tetralogy of Fallot and claim that pulmonary atresia plus partial or complete absence of the aorticopulmonary septum are its landmark features.11,12 Irrespective of all these uncertainties, common trunk in genetically defined mouse models becomes manifest after ED12,4,13,14 that is, at the onset of the period in which the structural transformation of the embryonic outflow tract into the ventriculo-arterial junction occurs. The outflow tract of the preseptation embryonic heart is a long, slowly conducting myocardial structure that functions as a sphincter at the arterial pole of the heart prior to the emergence of the 1-way semilunar valves.15,16 Extensive remodeling of the tissues of the outflow tract, as well as immigrating cells of the neural crest are responsible for connecting the right ventricle with the pulmonary trunk and the left ventricle with the aorta, that is, for the developmental formation of the definitive ventriculo-arterial junction. The precise mechanisms underlying this important architectural adaptation are still debated.17–23 Based on our findings in the Sox4-deficient mouse model and reports in the literature, we will argue that the tissue interactions between the endocardially derived cells of the outflow tract and cells derived from the neural crest determine the developmental fate of the embryonic outflow tract. Materials and Methods Animals Details concerning the method used to disrupt the Sox4 gene have been described elsewhere.4 Homozygous Sox4-disrupted embryos were identified by Southern blot analysis and accounted for 18% to 20% of the litter at embryonic day (ED) 12, 13, and 14 (not shown). Embryos were fixed either in a mixture of ice-cold methanol: acetone:water (2:2:1) or in 4% formaldehyde diluted in PBS (150 mmol/L NaCl; 10 mmol/L Na-phosphate; pH 7.4). After overnight fixation, the embryos were dehydrated in a graded series of ethanol, embedded in Paraplast plus, serially sectioned (7 mm thick), and mounted on poly-L-lysine– or 3-aminopropyltriethoxysilane– coated slides for immunohistochemistry or in situ hybridization staining, respectively. The sections were stored at 4°C until use. Immunohistochemistry The detailed procedure for the histological processing of the tissues has been described.24 Briefly, the serial sections were incubated overnight with alternately monoclonal antibodies against a-myosin heavy chain (a-MHC; 1:1025), b-myosin heavy chain (b–MHC; 1:1026), a-smooth-muscle actin (a-SMA; 1:1000; Sigma IMMH-2), desmin (1:50; Monosan, Mon 3001, clone 33), and a polyclonal antibody against fibronectin (1:6000; AB 1942, Brunschwig Chemie). After incubation with the primary antibodies, the sections incubated with monoclonal antibodies were stained with rabbit anti-mouse IgG (1:7500; noncommercial) followed by incubation with goat anti-rabbit IgG (1:250; noncommercial), whereas the sections incubated with the polyclonal antifibronectin antiserum were stained directly with goat anti-rabbit IgG. Antibody binding was demonstrated with rabbit peroxidase-antiperoxidase complex (1:750; Nordic, Tilburg, The Netherlands) and 3,3-diaminobenzidinetetrahydrochloride and H2O2 as substrates. In Situ Hybridization In situ hybridization was performed on formaldehyde-fixed sections to visualize the expression of Sox4, Nfatc, and SERCA 2 (sarcoendoplasmic reticulum calcium ATPase 2) as described in detail previously.27 The cRNA probes were transcribed from linearized November 16, 1998 987 Figure 1. The arterial pole of the heart in ED 12 and 13 mouse embryos that are heterozygous for Sox4 deficiency (A through G), homozygous (H), or wild-type (I through L). Panels A and C are stained immunohistochemically for the presence of desmin, and panels B and D are stained for the presence of fibronectin, whereas panels F, J, and K are stained by in situ hybridization for the presence of Sox4 mRNA, panels E, I, and K for the presence of SERCA2, and panels G and H for the presence of Nfatc. SERCA 2 mRNA is expressed in the cardiomyocytes, whereas Sox4 mRNA is expressed exclusively in the endocardially derived tissue of the outflow tract and the atrioventricular canal. Expression of Sox4 is also found in the mesenchyme surrounding the branchial arch arteries, trachea, and bronchi. Nfatc expression is concentrated in the endocardium overlying the endocardial ridges and cushions. ao indicates aorta; la, left atrium; lv, left ventricle; os, outlet septum; pt, pulmonary trunk; pv, pulmonary valve; ra, right atrium; rv, right ventricle; tr, trachea. Bar5100 mm. cDNA in pBluescript using T3 RNA polymerase and 35S-UTP. The Sox4 probe (EcoRI-Sacl fragment; 300 bp4) contained unique, untranslated sequences from the 59 end of the mRNA. The Nfatc3 probe (SacII-XhoI fragment28) was provided kindly by Dr G.R. Crabtree (Stanford, Calif). Selected hearts of Sox4-mutant mice were reconstructed 3-dimensionally as described29 and rendered with the help of a medical artist. Results Development of the Arterial Pole in Wild-Type and Heterozygous Littermates of Sox4-Deficient Mice No differences in morphological development were observed between wild-type and heterozygous littermates of Sox4deficient mice (Figure 1A through 1D). In ED12 mice, the aorta and pulmonary trunk have become separate vessels at and above the distal rim of the myocardium of the outflow tract. The fusion of the ridges continues toward the ventricle during ED13. Before fusion, the distal ends of the parietal and septal endocardial ridges each contain a central rod of condensed, a-SMA–positive mesenchymal cells of neural crest origin. When both ridges fuse, their central rods of condensed mesenchyme do so, too, by forming a central mass 988 Sox4-Deficiency Syndrome in Mice of a-SMA–positive cells. At ED14, the a-SMA–positive mesenchyme is still recognizable at the level of the valves. Further upstream, however, the presence of many apoptotic cells reveals that, soon after fusion, it is being eliminated and replaced by myocardial cells that can be seen to migrate into the outlet septum; that is, the septum forms between the subaortic and subpulmonary outlets on fusion of the septal and parietal endocardial ridges. The scallops of the semilunar valve leaflets in the aorta and pulmonary trunk, and the coronary arteries become discernible at ED13. The normal morphogenesis of the arterial pole therefore is virtually complete at ED14, which is comparable to 7 weeks of development (Carnegie stage 19 to 20) in man. Expression of Sox4 mRNA in Wild-Type and Heterozygous Littermates of Sox4-Deficient Mice Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 Sox4 mRNA is expressed abundantly in the nervous system, thymus, mesenchyme surrounding the branchial arch arteries, trachea, and esophagus, but not in the tissue of the aorticopulmonary septum (Figure 1E, 1F, and 1I through 1L). Sox4 expression levels in the endocardially derived tissues of the outflow tract and atrioventricular canal are similar. Expression of a-MHC, b-MHC, a-SMA, Desmin, Fibronectin, and Nfatc in Heterozygous and Homozygous Sox4-Deficient Embryos We used a- and b-MHC expression as a parameter to monitor myocardial development, fibronectin expression to selectively delineate the endocardially derived tissues, and, finally, a-SMA and desmin expression to follow both the maturation of the myocardium and the population of the endocardial tissue– derived cells with neural crest– derived cells.24,30 We observed no conspicuous differences in the spatio-temporal pattern of expression of these genes between deficient embryos and their wild-type or hemizygous littermates, suggesting that Sox4-deficiency does not primarily affect myocardial or endocardial development. Because Sox4-deficient mice resemble Nfatc-deficient mice with respect to the (failing) development of the semilunar valves, Nfatc expression in the endocardium was investigated and found to be normal (cf Figure 1G and 1H). Malformations of the Arterial Pole of the Heart in Sox4-Deficient Mice A total of 21 Sox4-deficient embryos were studied histologically, 4 of which were from ED12, 8 from ED13, and 9 from ED14. Cardiac morphogenesis was found to become progressively retarded in these embryos but did not stop. For example, in only 25% of Sox4-deficient ED13 hearts, coronary arteries were seen to develop, whereas this was the case in all hearts of wild-type littermates and in all ED14 Sox4deficient embryos. Furthermore, 2 dorsal aortae were still present in 7 of 8 ED13 Sox4-mutant embryos, whereas the right dorsal aorta had disappeared in all wild-type littermates. In affected ED14 embryos, the right dorsal aorta, if present, had diminished greatly in diameter. At ED12, nearly normal amounts of endocardially derived tissue was found in the outflow tract of Sox4-deficient hearts (Figure 2), suggesting that the morphogenesis of the endocardial Figure 2. Embryos Nos. 92 and 13. ED12 embryo No. 92 (A through C) belongs to the least-affected Sox4-deficient embryos in the series, whereas ED12 embryo No. 13 (D through F) is among the most seriously affected specimens. The embryos are stained immunohistochemically for the presence of b-MHC (A and D) and a-SMA (B, C, E, and F). Note that in the leastaffected embryo, the developing aorticopulmonary septum and the distal portion of the endocardial ridges are positioned between the lumen of the aortic and subpulmonary outlets (A and B), whereas in the seriously affected specimen, the endocardial ridges and the rudimentary aorticopulmonary septum are positioned anteriorly to the lumen of the outflow tract (D and E). As a consequence, the subpulmonary (o) and aortic outlets occupy anterior and right-lateral positions in embryo 92 (A), whereas the single arterial outlet (o) occupies the right lateral position in embryo 13 (D). Note also that there is no myocardium covering the aortic outlet at the level of the origin of the 6th arch arteries in the least affected specimen (A), whereas this area contains myocardium in the seriously affected specimen (D). Further upstream, both endocardial ridges occupy more lateral positions in the outflow tract in both embryos (C and F). Panel F further shows that these ridges contain, in comparison with the less-affected specimen (C), only few a-SMA–positive cells. A reconstruction of the heart of embryo No. 13 is shown in Figure 5D. ao indicates aorta; aps, aorticopulmonary septum; pt, pulmonary trunk; and r, endocardial ridge. *Condensed mesenchyme. Bar5100 mm. ridges rather than the formation of endocardially derived tissue is primarily affected. In ED13 and ED14 hearts, the developing septal and parietal endocardial ridges could be distinguished from the so-called intercalated endocardial ridges18 by the presence of a rod of condensed, a-SMA–positive mesenchyme (Figures 3 and 4). Except in the most seriously affected embryos, the 2 endocardial ridges were always present, but the number and position of intercalated ridges between them differed considerably. Although we noted no correlation between the number of intercalated ridges and the severity of the malformation, a normal position of the intercalated ridges was seen only in the least affected hearts. Sox4-Deficiency Syndrome In the Table, the embryos are arranged in order of increasing severity of their malformation at the arterial pole. Features Ya et al Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 Figure 3. Embryos No. 324 (A through C) and 101 (D through F). These ED13 embryos are stained with hematoxylinazophlochsin (A through C) or immunohistochemically for the presence of a-SMA (D and E) or b–MHC (F). Both embryos demonstrate typical malformations in the arterial pole of Sox4deficient embryos, viz anterior position of the aorta (A and D; note origin of coronary arteries), 1 (E) or 2 (B) intercalated ridges that are positioned anterior to the adjacent septal and parietal endocardial ridges. The septal and parietal ridges, which contain rods of condensed mesenchyme, gradually converge onto the inner curvature (C and F). A reconstruction of the heart of embryo No. 101 is shown in Figure 5C. ao indicates aorta; aps, aorticopulmonary septum; ca, coronary artery; cm, condensed mesenchyme; ct, common trunk; is, intercalated ridge; la, left atrium; pt, pulmonary trunk; r, ridge; ra, right atrium; and rv, right ventricle. Bar550 mm. associated with increasing severity of the malformation were fusion of the endocardial ridges on the posterior wall of the outflow tract near the lesser curvature, fusion of the condensed mesenchymal rods on the posterior wall of the outflow tract, side-by-side or posterior position of the pulmonary trunk, difference in size of the pulmonary trunk and aorta, abnormal number or position of the intercalated ridges, and, finally, the presence of esophago-tracheal fistula. Based on the position of the ascending aorta relative to the pulmonary trunk, we distinguished 4 groups. The least severe grade of malformation (Figures 2A through 2C, 5A, and 6; group 1 of the Table; '25% of cases) was characterized by a complete separation of aorta and pulmonary trunk, 2 separate semilunar valve anlagen, and a remnant of the developing outlet septum below the valvular rings. Usually, all 6 endocardial swellings could be identified at the level of the developing valves. Although the endocardial ridges had fused at and just below the valves, they had failed to do so further toward the embryonic right ventricle. As a result, a large infundibular ventricular septal defect existed in all cases. The aortic arch and arterial duct were always on the left side. Furthermore, the diameter of the aortic and pulmonary roots were usually comparable, with the pulmonary root located anterior to that of the aorta. In more serious malformations (Figures 5B and 7; group 2 of the Table; '25% of cases), the ascending aorta and November 16, 1998 989 Figure 4. Embryo No. 114. This ED14 Sox4-deficient embryo is stained immunohistochemically for the presence of a-SMA (A through C), and b-MHC (D). The pulmonary and ascending aortic trunks are partially and unequally separated by an underdeveloped aorticopulmonary septum (A and B), whereas the outflow tract is a single tube directly connected to the right ventricle (C and D). The pulmonary arteries arise from the posteriorly located large pulmonary trunk (A). Note the a-SMA–positive condensed mesenchyme (C) in the underdeveloped outflow tract ridges, which converge onto the inner curvature at the ventricular end of the outflow tract (D). ao indicates aorta; aps, aorticopulmonary septum; cm, condensed mesenchyme; ct, common trunk; pa, pulmonary artery; pt, pulmonary trunk; and rv, right ventricle. Bar550 mm. pulmonary trunk were separate vessels to the extent that the pulmonary arteries arose from the pulmonary trunk and coronary arteries arose from the aorta. However, at and below the level of the semilunar valve anlagen, only a single lumen was present. Furthermore, the root of the pulmonary trunk and aorta typically occupied side-by-side positions. Nevertheless, the morphological presentation of the endocardial ridges was still rather normal with the septal and parietal ridges each following their characteristic, spiral downstream course in the outflow tract. As the severity of the malformation increased further, posterior position of the pulmonary trunk relative to the aorta was found invariably (Figures 3, 4, and 5C; group 3 of the Table; 35% of cases). At the level of the semilunar valve anlagen, only 3 or 4 endocardial swellings were found. The swellings containing the condensed mesenchymal rods typically occupied a posterior, adjacent position, that is, without intercalated swelling between (Figure 3B, 3C, and 3E). Further toward the ventricle, the septal and parietal ridges were fused into a single ridge that was positioned on the posterior wall of the outflow tract, that is, near the lesser curvature (Figures 3C, 3F, and 4D). In 4 embryos, this fused 990 Sox4-Deficiency Syndrome in Mice Features Characterizing the Sox4 Deficiency Syndrome Post Fusion Embryo No. 92 Pulmonary Trunk ED Ridges CM Valve Position Size ETF 12 2 2 ND Ant PT5A 2 302 13 2 2 Ant PT5A 2 18 14 2 2 Ant PT5A 2 91 14 2 2 N1 14 2 2 92 14 2 2 36 12 2 2 101 14 2 2 ND ND Ant PT5A 2 Ant PT5A 2 Ant PT.A 2 Lat? PT5A 2 Lat PT5A 2 103 14 1 2 Lat PT.A 2 331 13 2 2 Lat PT5A 1 307 13 1 2 Lat PT5A 2 Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 321 13 2 2 Post PT5A 2 313 13 2 2 Post PT5A 2 N2 14 1 2 324 13 1 1 ND Post* PT5A 2 Post PT,A 2 1 101 13 1 1 Post PT,A 201 13 1 1 Post PT,,A 1 114 14 1 1 Post PT.A 2 93 12 1 Abs ND Single vessel 2 13 12 1 Abs ND Single vessel 2 72 14 1 Abs Single vessel 1 The Sox4-deficiency syndrome was subdivided into 4 groups with increasing severity of the abnormalities. Criteria were whether or not the endocardial ridges had fused, whether or not condensed mesenchyme (CM) was recognizable in 2 of the ridges, and whether or not these rods fused on the posterior wall of the outflow tract (Post Fusion). Abs indicates absent. The configuration of the endocardial ridges at the valve level is described by a pictogram, in which dots represent ridges containing condensed mesenchyme (Valve). The position of the pulmonary trunk (PT) relative to that of the aorta (A) is indicated as Ant (PT anterior), Lat (PT and A side-by-side), or Post (PT posterior), whereas differences in diameter are indicated by ,, 5, or .. The presence of an esophagotracheal fistula (ETF) as indicated by 1. ND indicates not determined because of age (ED12) or plane of sectioning (Nos. N1 and N2); and ?, the position of the ascending aorta relative to the pulmonary trunk cannot be determined with certainty yet. *This case was described by Schilham et al4 as having its ascending aorta in the anterior position; however, reinspection of the limited number of sections that were available indicated that the aorta of this specimen also occupied an anterior position. posterior portion of the ridges occupied more than one third of the length of the outflow tract so that, in addition, the pillars of condensed mesenchyme had become fused. Because the condensed mesenchyme in the endocardial ridges is continuous with the strongly a-SMA–positive mesenchyme of the aorticopulmonary septum, a ring of a-SMA–positive mesenchyme encircles the pulmonary outlet in this group (Figure 5C). The relative diameters of pulmonary trunk and aorta varied substantially, the most severe cases presenting with stenosis of the pulmonary trunk (Figures 3 and 5C). Narrowing of the pulmonary trunk always was accompanied by absence of the arterial duct, whereas the aortic arch was left-sided and normal in diameter. On the other hand, when the diameter of the aortic root was smaller than that of the pulmonary trunk (Figure 4), the embryo also had an enlarged diameter of the arterial duct. In all these cases, both the aortic arch and the arterial duct were situated on the left side. The coronary artery anlagen always were associated with the aorta (Figure 3A and 3D). A single arterial trunk, which was a direct continuation of the outflow tract and from which both the coronary arteries and the pulmonary arteries originated, was found in 15% of cases (Figures 2D through 2F, 5D, 5E, and 8; group 4 of the Table). In the ED14 embryo that is illustrated in Figure 8, the arterial duct was absent, whereas the aortic arch was on the right side. Tissue with the same staining properties as those of the endocardial ridges surrounded the lumen of the outflow tract as a cuff, but rods of condensed mesenchyme were absent. Two ED12 specimens revealed that this condition arose as a result of merging of the distal portion of the endocardial ridges on the anterior side of the lumen of the outflow tract (Figure 2D through 2F). As a result, the aortic Ya et al Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 Figure 5. The spectrum of cardiac malformations in Sox4deficient embryos. Panels A through E show 3– dimensional reconstructions of hearts of Sox4-deficient embryos with increasing degrees of severity of the malformation. A, Grade 1 malformation (ED14 embryo No. 18; see also Figure 6). B, Grade 2 malformation (ED14 embryo No. 101; see also Figure 7). C, Grade 3 embryo (ED13 embryo No. 101; see also Figure 3D through 3F). D and E, Grade 4 embryos (ED12 embryo No. 13, see also Figure 2D through 2F; and ED14 embryo No. 72, see also Figure 8, respectively). For a classification of groups 1 to 4, see the Table. Dashed lines in panels A and D indicate the upper boundary of the myocardium. ao indicates aorta; ca, coronary arteries; ct, common trunk; et, endocardially derived tissue; pa, pulmonary artery; pt, pulmonary trunk; r, ridge; rv, right ventricle; and te, common part of trachea and esophagus. sac is not divided between ascending aorta and pulmonary trunk. The ED12 embryos further resembled the ED14 specimen in that relatively few a-SMA–positive cells were seen in the endocardial tissue and that the right-sided fourth branchial arch artery was better developed than the left, whereas both sixth-arch arteries were reduced in size. Two embryos of this group suffered, in addition, from a hypoplastic left ventricle. Associated Malformations in Sox4-Deficient Mice November 16, 1998 991 Figure 6. Embryo No. 18. This ED14 Sox4-deficient embryo is among the least affected in the series. The embryo is stained immunohistochemically for the presence of b-MHC (A and D), a-SMA (B), and fibronectin (C). A, Above the semilunar valves, the outflow tract is separated into aorta and pulmonary trunk by the aorticopulmonary septum. However, near the junction with the right ventricle, the endocardial ridges do not fuse (compare panels B and C with panel D). Note further that the semilunar valve leaflets in the pulmonary trunk (A) and aorta (D) are rudimentarily developed. A reconstruction of this heart is shown in Figure 5A. ao indicates aorta; aps, aorticopulmonary septum; av, aortic valve; cm, condensed mesenchyme; os, outlet septum; pt, pulmonary trunk; pv, pulmonary valve; r, ridges; and rv, right ventricle. Bar5100 mm. pulmonary trunk (type IV according to Angelini and Leachman10), via a common origin of aorta and pulmonary trunk at the level of the valve anlagen (type 1 according to Collett and Edwards,31 type A1 according to van Praagh and van Praagh,11 and type II according to Angelini and Leachman10), to 2 pulmonary artery orifices in the dorso-caudal truncal wall (type 2 according to Collett and Edwards,31 type A2 according to van Praagh and van Praagh,11 and type I according to Angelini and Leachman10). Additionally, we did observe embryos in which the diameter of the pulmonary trunk was much smaller than that of the aorta (van Praagh and van Praagh’s type A311) and embryos in which the diameter of the Branchial arch anomalies in ED13 and ED14 embryos, such as absence (4 cases) or increased size (2 cases) of the arterial duct, were correlated with the size of the pulmonary trunk relative to that of the aorta. An esophagotracheal fistula was found in 4 of the more serious cases (Figure 8). These malformations probably represent an independent feature of Sox4 deficiency, because the transcription factor also is expressed strongly in the mesenchyme of the branchial arches and on the dorsal surface of the trachea. Discussion Even though only a single gene (Sox4) was inactivated, we observed a range of malformations of the arterial pole of the heart. The anomalies ranged from a solitary defect in the proximal (conal) portion of the outlet septum, that is, with separate valve anlagen supporting the roots of the aorta and Figure 7. Embryo No. 101. This Sox4-deficient ED14 embryo is stained immunohistochemically for the presence of a-SMA (A) and b-MHC (B). At the semilunar valve level, the aorta and pulmonary channels occupy side-by-side positions (A and B). A reconstruction of this heart is shown in Figure 5B. ao indicates aorta; cm, condensed mesenchyme; la, left atrium; pa, pulmonary artery; pv, pulmonary valve; r, endocardial ridges; ra, right atrium; and rv, right ventricle. Bar5100 mm. 992 Sox4-Deficiency Syndrome in Mice Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 Figure 8. Embryo No. 72. This ED14 Sox4-deficient embryo is stained with hematoxylin-azophlochsin. In this embryo, only a single ascending trunk arises from the embryonic right ventricle (C), from which the coronary arteries (B) and the pulmonary arteries (A) arise. No condensed mesenchyme can be seen in the endocardial tissue. Note the common part of the esophagus and trachea in panel A. A reconstruction of this heart is shown in Figure 5E. ca indicates coronary artery; ct, common trunk; et, endocardially derived tissue; la, left atrium; pa, pulmonary artery; r, ridge; ra, right atrium; rv, right ventricle; and te, common part of trachea and esophagus. A and B, Bar550 mm. C, Bar5100 mm. aorta was much smaller than that of the pulmonary trunk (van Praagh and van Praagh’s type A411). Such a wide spectrum in the phenotypic presentation of a single gene deficiency is probably characteristic of genes that are either components of a signal-transduction pathway or transcription factors. Most likely, the downstream members of these signal-transduction pathways or the target genes of the transcription factors continue to be active, but as a consequence of a lack of regulation, the level of these downstream activities is no longer (tightly) controlled, thus allowing the emergence of large interindividual differences in phenotype. Our finding that Nfatc is expressed in Sox4-deficient embryos and vice versa,32 therefore does not prove the presence of independent regulatory pathways. On the other hand, accurate descriptions of single gene– deficiency syndromes are necessary to decide which congenital malformations may share a common deficiency and to obtain a sufficiently large catalogue of syndromes to deduce which regulatory pathways are involved in a particular morphogenetic process. Both Sox4 and Nfatc32,33 deficiency are characterized by death due to heart failure on day 14 of embryonic development. The observed regurgitation of blood in embryos of this age shows that the lack of transformation of the distal portion of the endocardial ridges into one-way semilunar valves is, in all likelihood, the direct cause of the heart failure in both the mildly and seriously affected embryos (compare with References 34 to 36). The timing of the onset of this functional failure strongly supports our hypothesis that it is the remodeling of endocardial tissue into valvular structures rather than the formation of the endocardial tissue itself that is primarily affected. Before the development of the semilunar valves on ED13 and ED14, an adequate amount of endocardially derived stuffer tissue in the outflow tract suffices to allow the outflow tract to function as a myocardial sphincter that prevents regurgitation of blood.16 Although population of the endocardial cushions of the atrioventricular canal by neural crest cells is quantitatively insignificant compared with that of the endocardial ridges of the outflow tract,21,37 Sox4 expression levels in both endocardial tissue– derived structures are comparable (Figure 1F, 1J, and 1L). Sox4 expression therefore is localized almost certainly in endocardially derived cells. Because the malformations in Sox4-deficient embryos are largely restricted to the arterial pole, whereas the development of the atrioventricular cushions is not visibly affected, our study suggests that it is the interaction between the neural crest– derived myofibroblasts30 and the endocardially derived original cells of the outflow tract, that determines proper development of the arterial pole. The least affected Sox4-deficient hearts, strictly speaking,8,9 do not suffer from common trunk but should be classified as having a large infundibular ventricular septal defect. In the more serious cases, the following are seen: a linear rather than a spiral configuration of the ridges and, consequently, a posterior position of the pulmonary trunk at the level of the valve (the configuration present in transposition of the great arteries), an abnormal number and position of the intercalated ridges relative to the parietal and septal endocardial ridges, lack of fusion of the ridges, and variable size of the aorta relative to the pulmonary trunk. In the most serious cases, even the formation of the endocardial ridges failed, so that a cuff of endocardial tissue, in which the neural crest– derived rod(s) of condensed mesenchyme could not even be identified as such, continued to surround the lumen of the outflow tract. Because the emergence of spiraling septal and parietal endocardial ridges coincides temporally with the ingrowth of a-SMA–positive neural crest cells30 and because the position of these 2 endocardial ridges greatly differs between individual affected mice, it appears that the migration of the neural crest cells into the endocardial tissue of the outflow tract is poorly guided in Sox4-deficient mice. The association of an abnormal position of the endocardial ridges and an abnormal number and position of the intercalated ridges with the presence of a single arterial outlet and often with a difference in diameter of aorta and pulmonary trunk, implies a correspondingly abnormal position of the aorticopulmonary septum. We know of only 2 earlier descriptions of common trunk in embryos.38,39 For that reason, the morphogenetic events underlying the development of common trunk are deduced mostly from observations in human neonates. The conventional explanation of common trunk is a complete or partial failure of down-growth of the aorticopulmonary septum, Ya et al Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 together with incomplete development of the endocardial ridges of the outflow tract (eg, References 8 to 10, 38, and 40). Some consider the abnormal development of the ridges to be secondary to faulty development of the aorticopulmonary septum (eg, Reference 9). In support of this view are the extensive population of the ridges of the outflow tract by neural crest cells21,37 and the prevalence of malformations of the arterial pole of the heart in animal models in which genes that are expressed in neural crest derivatives are disrupted13,14,41– 45 (for a review, see Reference 46). Accordingly, common trunk, together with tetralogy of Fallot and interrupted aortic arch, is described often as one of the symptoms identifying a typical neurocristopathy in human neonates,47 but it almost never is associated with a configuration reminiscent of (complete) transposition of the great arteries. In Sox4-deficient embryos, however, we did not observe tetralogy of Fallot or interrupted aortic arch, whereas we did observe the configuration of the endocardial ridges that corresponds with transposition in '50% of cases. These differences in presentation are suggestive of distinctive roles for the endocardially derived cells and the neural crest cells in the regulation of morphogenesis of the outflow tract. In addition, the lack of differentiation of the semilunar valves in Sox4- and Nfatc-deficient mice, and the almost normal development of semilunar valves in common trunk caused by the dysfunction of neural crest cells (eg, resulting from deficiency of nuclear retinoic acid receptors13,44 or the Pax3 transcription factor14) also support a distinct morphogenetic role for endocardially derived and neural crest– derived tissues. Conclusion The Sox4-deficiency syndrome can be ascribed to a defective function of the endocardial tissue of the outflow tract, leading to a lack of development and/or fusion of the endocardial ridges, a lack of development of the semilunar valves, and an arrangement of the ventriculo-arterial connection corresponding with transposition of the great arteries. The Sox4deficiency syndrome is therefore an animal model for abnormal development of the ventriculo-arterial junction because of a defective function of the endocardially derived tissue. The fate of the endocardial tissue– derived structures involved in the formation of the ventriculo-arterial junction could be followed, thanks to the presence of the highly characteristic rods of condensed mesenchyme. The restriction of the malformations to the arterial pole, even though the Sox4 gene is equally expressed in the endocardially derived tissue of the atrioventricular canal, implies that the interaction between the endocardially derived tissue of the outflow tract and the neural crest– derived myofibroblasts30 determines proper development of the arterial pole. The lack of differentiation of the semilunar valves in Sox4- and Nfatc-deficient32,33 mice and their normal development in animal models with neural crest deficiencies implies that the neural crest does not regulate the development of the semilunar valves. References 1. Hoffman JIE, Christianson R. Congenital heart disease in a cohort of 19,502 births with long-term follow-up. Am J Cardiol. 1978;42:641– 647. November 16, 1998 993 2. Hoffman JIE. Incidence of congenital heart disease, II: prenatal incidence. Pediatr Cardiol. 1995;16:155–165. 3. Friedman WF. Congenital heart disease in infancy and childhood, in Braunwald E (ed): Heart Disease: A Textbook of Cardiovascular Medicine, 3rd edition. Philadelphia, Pa: Saunders Press; 1988:895–913. 4. Schilham MW, Oosterwegel MA, Moerer P, Jing Y, de Boer PAJ, Verbeek S, Lamers WH, Kruisbeek AM, Cumano A, Clevers H. Sox-4 gene is required for cardiac outflow tract formation and pro-B lymphocyte expansion. Nature. 1996;380:711–714. 5. van de Wetering M, Clevers H. Sox 15, a novel member of the murine Sox family of HMG box transcription factors. Nucleic Acids Res. 1993; 21:1669. 6. Kanai Y, Kanai-Azuma M, Noce T, Saido TC, Shiroishi T, Hayashi Y, Yazaki K. Identification of two Sox17 messenger RNA isoforms with and without the high mobility group box region, and their differential expression in mouse spermatogenesis. J Cell Biol. 1996;133:667– 681. 7. van de Wetering M, Oosterwegel M, van Norren K, Clevers H. Sox-4, an Sry-like HMG box protein, is a transcriptional activator in lymphocytes. EMBO J. 1993;12:3847–3854. 8. Crupi G, Macartney FJ, Anderson RH. Persistent truncus arteriosus. Am J Cardiol. 1977;40:569 –578. 9. Bartelings MM, Gittenberger-de Groot AC. Morphogenetic considerations on congenital malformations of the outflow tract, 1: common arterial trunk and tetralogy of Fallot. Int J Cardiol. 1991;32:213–230. 10. Angelini P, Leachman RD. Trunco-conal septal defects, an anatomic and embryologic discussion of common truncus and related malformations. Eur J Cardiol. 1974;2:11–22. 11. van Praagh R, van Praagh S. The anatomy of common aortico-pulmonary trunk (truncus arteriosus communis) and its embryologic implications. Am J Cardiol. 1965;16:406 – 425. 12. van Praagh R. Classification of truncus arteriosus communis (TAC). Am Heart J. 1976;92:129 –132. 13. Mendelsohn C, Lohnes D, Decimo D, Lufkin T, LeMeur M, Chambon P, Mark M. Function of the retinoic acid receptors (RARs) during development, II: multiple abnormalities at various stages of organogenesis in RAR double mutants. Development. 1994;120:2749 –2771. 14. Franz T. Persistent truncus arteriosus in the Splotch mutant mouse. Anat Embryol (Berl). 1989;180:457– 464. 15. de Jong F, Opthof T, Wilde AAM, Janse MJ, Charles R, Lamers WH, Moorman AFM. Persisting zones of slow impulse conduction in developing chicken hearts. Circ Res. 1992;71:240 –250. 16. Moorman AFM, Lamers WH. Molecular anatomy of the developing heart. Trends Cardiovasc Med. 1994;4:257–264. 17. Noden DM, Poelmann RE, Gittenberger-de Groot AC. Cell origins and tissue boundaries during outflow tract development. Trends Cardiovasc Med. 1995;5:69 –75. 18. Kramer TC. The partitioning of the truncus and conus and the formation of the membranous portion of the interventricular septum in the human heart. Am J Anat. 1942;71:343–370. 19. de Vries PA, De CH, Saunder JB: Development of the ventricles and spiral outflow tract in the human heart. Contrib Embryol. 1962;37:87–114. 20. van Mierop LHS, Alley RD, Kausel HW, Stranahan A. Pathogenesis of transposition complexes, I: embryology of the ventricles and great arteries. Am J Cardiol. 1963;12:216 –225. 21. Kirby ML, Gale TF, Stewart DE. Neural crest cells contribute to normal aorticopulmonary septation. Science. 1983;220:1059 –1061. 22. Kirby ML, Waldo K. Neural crest and cardiovascular patterning. Circ Res. 1995;77:211–215. 23. Beall AC, Rosenquist TH. Smooth muscle cells of neural crest origin form the aorticopulmonary septum in the avian embryo. Anat Rec. 1990; 226:360 –366. 24. Jing Y, Markman MWM, Wagenaar GTM, Blommaart PJE, Moorman AFM, Lamers WH. Expression of the smooth-muscle proteins alpha smooth-muscle actin and calponin, and of the intermediate filament protein desmin are parameters of cardiomyocyte maturation in the prenatal rat heart. Anat Rec. 1997;249:495–505. 25. de Groot IJM, Lamers WH, Moorman AFM. Isomyosin expression pattern during rat heart morphogenesis: an immunohistochemical study. Anat Rec. 1989;224:365–373. 26. Sant’Ana Pereira JAA, Wessels A, Nytmans L, Moorman AFM, Sargeant AJ. New method for the accurate characterization of single human skeletal muscle fibres demonstrates a relation between mATPase and MyHC expression in pure and hybrid fiber types. J Muscle Res Cell Motil. 1995;16:21–34. 994 Sox4-Deficiency Syndrome in Mice Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 27. Moorman AFM, Vermeulen JLM, Koban MU, Schwartz K, Lamers WH, Boheler KR. Patterns of expression of sarcoplasmic reticulum Ca21ATPase and phospholamban mRNAs during rat heart development. Circ Res. 1995;76:616 – 625. 28. Ho SN, Thomas DJ, Timmerman LA, Li X, Francke U, Crabtree GR. NFATc3, a lymphoid-specific NFATc family member that is calciumregulated and exhibits distinct DNA binding specificity. J Biol Chem. 1995;270:19898 –19907. 29. Verbeek FJ, Huysmans DP, Baeten RJAM, Schoutsen NJC, Lamers WH. Design and implementation of a database and program for 3D-reconstruction from serial sections: a data-driven approach. Microsc Res Tech. 1995;30:496 –512. 30. Jing Y, van den Hoff MJB, de Boer PAJ, Tesink-Taekema S, Franco D, Moorman AFM, Lamers WH. The normal development of the outflow tract in the rat. Circ Res. 1998;82:464 – 472. 31. Collett RW, Edwards JE. Persistent truncus arteriosus: a classification according to anatomic subtypes. Surg Clin North Am. 1949;29:1245–1270. 32. de la Pompa JL, Timmerman LA, Takimoto H, Yoshida H. Role of the NF-ATc transcription factor in morphogenesis of cardiac valves and septum. Nature. 1998;392:182–185. 33. Ranger AM, Grusby, Hodge MR, Gravallese EM. The transcription factor NF-ATc is essential for cardiac valve formation. Nature. 1998;392:186–190. 34. Hartwig NG, Vermeij-Keers C, de Vries HE, Gittenberger-de Groot AC. Aplasia of semilunar valve leaflets: two case reports and developmental aspects. Pediatr Cardiol. 1989;12:114 –117. 35. Miyabara S, Ando M, Yoshida K, Saito N, Sughihara H. Absent aortic and pulmonary valves: investigation of three fetal cases with cystic hygroma and review of the literature. Heart Vessels. 1994;9:49 –55. 36. Rossant J. Mouse mutants and cardiac development: new molecular insights into cardiogenesis. Circ Res. 1996;78:349 –353. 37. Kirby ML. Cellular and molecular contributions of the cardiac neural crest to cardiovascular development. Trends Cardiovasc Med. 1993;3:18–23. 38. van Mierop LHS, Patterson DF, Schnarr WR. Pathogenesis of persistent truncus arteriosus in light of observations made in a dog embryo with the anomaly. Am J Cardiol. 1978;41:755–762. 39. Maron BJ, Hutchins GM. Truncus arteriosus malformation in a human embryo. Am J Anat. 1996;134:167–174. 40. Rosenquist GC, Bharati S, McAllister HA, Lev M. Truncal valve anomalies associated with small conal or truncal defects. Am J Cardiol. 1976;37:410 – 412. 41. Schatteman GC, Motley ST, Effman EL, Bowen-Pope DF. Plateletderived growth factor receptor alpha subunit deleted patch mouse exhibits severe cardiovascular dysmorphogenesis. Teratology. 1995; 51:351–366. 42. Kurihara Y, Kurihara H, Oda H, Maemura K, Nagai R, Ishikawa T, Yazaki Y. Aortic arch malformations and ventricular septal defect in mice deficient in endothelin-1. J Clin Invest. 1995;96:293–300. 43. Donovan MJ, Hahn R, Tessarollo L, Hempstead BL. Identification of an essential nonneural function of neurotrophin 3 in mammalian cardiac development. Nat Genet. 1996;14:210 –213. 44. Kastner P, Grondona JM, Mark M, Gansmuller A, LeMeur M, Decimo D, Vonesch P, Dolle P, Chambon P. Genetic analysis of RXRa developmental function: convergence of RXR and RAR signaling pathways in heart and eye morphogenesis. Cell. 1994;78:987–1003. 45. Sucov HM, Dyson E, Gumeringer CL, Price J, Chien C, Evans RM. RXRa-mutant mice establish a genetic basis for vitamin A signaling in heart morphogenesis. Genes Dev. 1994;8:1007–1018. 46. Jing Y, Schilham MW, Clevers H, Moorman AFM, Lamers WH. Animal models of congenital defects in the ventriculoarterial connection of the heart. J Mol Med. 1997;75:551–566. 47. van Mierop LHS, Kutsche LM. Cardiovascular anomalies in Di George syndrome and importance of neural crest as a possible pathogenetic factor. Am J Cardiol. 1986;58:133–137. Sox4-Deficiency Syndrome in Mice Is an Animal Model for Common Trunk Jing Ya, Marco W. Schilham, Piet A. J. de Boer, Antoon F. M. Moorman, Hans Clevers and Wouter H. Lamers Downloaded from http://circres.ahajournals.org/ by guest on June 14, 2017 Circ Res. 1998;83:986-994 doi: 10.1161/01.RES.83.10.986 Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 1998 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7330. Online ISSN: 1524-4571 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circres.ahajournals.org/content/83/10/986 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in the Permissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation Research is online at: http://circres.ahajournals.org//subscriptions/