Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
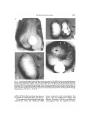
AMER. ZOOL., 24:589-603 (1984) The Cellular Basis of Gastrulation in Xenopus laevis: Active, Postinvolution Convergence and Extension by Mediolateral Interdigitation R. E. KELLER Department of Zoology, University of California, Berkeley, California 94720 SYNOPSIS. Time-lapse videomicrographic and SEM analyses of normal and microsurgically altered gastrulation show that the morphogenetic movements of the dorsal marginal zone (DMZ)—extension, convergence, and involution—all result from behavior that occurs after the marginal zone has involuted. Before its involution, the DMZ shows no detectable capacity for autonomous convergence or extension. If its involution is prevented, the DMZ will show convergence and extension but only at developmental stages at or beyond the stage at which it normally would have involuted. Thus autonomous convergence and extension, which have been ascribed to the DMZ are, in fact, properties of the dorsal mesodermal mantle (DMM) and the archenteron roof. SEM analysis of cell shape and packing patterns, suggest that cells of the DMM merge (interdigitate) mediolaterally, between one another, beginning just beyond the point of involution. This behavior is thought to reduce the width and increase the length (postinvolution convergence and extension) of the DMM. The decrease in circumference (width) at the vegetal-most part of the newly involuted DMM forms a constriction ring just inside the blastopore. Constriction and concurrent elongation of the DMM act in concert to move the blastoporal lip vegetally. The DMZ is passively pulled vegetally and over the blastoporal lip as deep cells are recruited for participation in mediolateral interdigitation at the vegetal end of the DMM. REVIEW OF GASTRULATION IN XENOPUS LAEVIS Gastrulation in the anuran amphibian, Xenopus laevis, occurs by a rolling over the blastoporal lip (involution) of the superficial layer of the marginal zone to form the endodermal roof of the archenteron, and the concurrent involution of the deep region of the marginal zone to form the mesodermal mantle (Keller, 1975, 1976). Xenopus has no mesoderm on the surface (see Nieuwkoop and Florschutz, 1950; Keller, 1975, 1976), in contrast to other anurans and urodeles that have been studied (see Vogt, 1929; Pasteels, 1942). As the marginal zone involutes, the animal region undergoes an increase in area, or epiboly (see Vogt, 1929; Keller, 1978). During epiboly, the dorsal marginal zone (DMZ) lengthens greatly (extension) and narrows (convergence). Extension supplies more 1 From the Symposium on Gastrulation presented at the Annual Meeting of the American Society of Zoologists, 27-30 December 1982, at Louisville, Kenlucky. 589 material to the blastoporal lip than is involuted; thus the blastoporal lip moves vegetally across the yolk plug (see Keller, 1981). Convergence of the DMZ results in concurrent constriction of the blastopore. Because most convergence and extension occur in the dorsal sector, the dorsal and dorsolateral lips move much farther across the yolk plug than the lateral and ventral lips, and therefore the blastopore closes eccentrically over the ventral region of the yolk plug (see Keller, 1975). Concurrent with the onset of involution of the deep region of the DMZ, bottle cells form by apical constriction and apical-basal elongation of superficial, epithelial cells in the dorsal region (see Holtfreter, 1943a, b; Baker, 1965; Perry and Waddington, 1966). These changes in cell shape result in formation of the blastoporal groove by invagination—the bending of a cell sheet (see Lewis, 1947). Further deepening of the groove to form the archenteron results from vegetal extension and involution of the blastoporal lips rather than invagination (see Keller, 1981). The roof of the 590 R. E. KELLER archenteron is attached tightly to the deep mesodermal cells. These mesodermal cells appear to migrate toward the animal pole (Nakatsuji, 1975, 1976; Keller and Schoenwolf, 1977) and carry the archenteron roof with them (Keller, 1981). More accurately, the involuted mesoderm pulls the preinvolution material vegetally, over itself. In urodeles, bottle cells were thought to migrate into the interior of the gastrula and pull everything inside (Holtfreter, 1943a, b). However, there are several arguments against this mechanism operating in anurans (see Keller, 1981) and urodeles (Daniel and Yarwood, 1939). In Xenopus, an anuran, bottle cell removal does not prevent extension, convergence, or involution (Cooke, 1975; Keller, 1981). Spreading of the superficial layer occurs by increase in area, flattening, and division of superficial cells (Keller, 1978, 1980). Deep cells do not move out into the superficial layer as they do in the urodele (see Holtfreter, 1943a). The deep region spreads by interdigitation of several layers of deep cells to form fewer layers of greater area. This interdigitation may be an active, force-producing process in epiboly or a passive response to forces generated elsewhere (Keller, 1980). Active interdigitation could presumably result in autonomous spreading and perhaps extension of the DMZ (Keller, 1980). Isolation experiments on other amphibians (Holtfreter, 1939; Townes and Holtfreter, 1955; Ikushima and Maruyama, 1971) and rearrangement experiments by Schechtman (1942) and others (see Spemann, 1938), suggest that the extension of the DMZ is an active, autonomous process. But in all these works it is not clear whether the observed extension occurred during the gastrula stages or represented extension of the notochord during neurula stages. Changes in cell morphology, arrangement, and probably behavior, occur at the point of involution, particularly in the deep region (Keller and Schoenwolf, 1977; Keller, 1981). Only the deep cells of the marginal zone have the capacity to complete these changes and involute (Keller, 1981). It is not clear what these changes are, or how thev function in involution. There are changes in the extracellular matrix (Kosher andSearls, 1973;Johnson, 1977a-d, 1984; also see Lee et al., 1982) and cell surface charge (Schaeffer et al., 1973) that may be associated with the involuting cells. The task at hand is to identify a population or populations of cells and their corresponding behavior patterns that bring about the movements of gastrulation. Previously, the cellular basis of epiboly was modeled in terms of two alternative mechanisms—active or passive interdigitation of deep cells (Keller, 1980). Other work strongly suggested that the deep region of the marginal zone is the critical element in the process of involution (Keller, 1981). The goal of the present work was to determine what region of the gastrula can undergo autonomous convergence and extension and what cell behavior is involved. Microsurgical alteration of the DMZ and analysis of the resulting gastrulation by time-lapse videomicroscopy (TLV) and scanning electron microscopy (SEM), strongly suggests that the force-generating process that brings about all these movements—convergence, extension, and involution—does not occur in the preinvolution DMZ but in the postinvolution dorsal mesodermal mantle (DMM), beginningjust inside the point of involution. It is proposed that extension, convergence, constriction of the blastopore, and involution form a complex of processes that all result from a common postinvolution process— active, mediolateral interdigitation of deep mesodermal cells. The principal elements of this notion of gastrulation and the key experiments supporting them will be set forth here. Further description, more detailed analyses, and tests of the major hypotheses of this model will follow in subsequent publications. EVIDENCE THAT ACTIVE EXTENSION AND CONVERGENCE ARE POSTINVOLUTION PROCESSES Does active, autonomous convergence and extension occur? If so, when and where does it occur? In the classical literature (see Spemann, 1938), extension and convergence were associated with the "marginal XENOPUS GASTRULATION 591 FIG. 1. Microsurgical manipulations of the early gastrula are shown diagrammatically in sagittal (a, b, d) and vegetal (c, e) views. Dorsal marginal zone (DMZ) was grafted to the animal pole (a). Patches of animal pole were grafted to the dorsolateral marginal zone (b, c). Patches of the dorsal marginal zone were rotated 90 degrees clockwise (d, e). The superficial layer is shaded heavily and the deeper layer is shaded lightly. MM, mesodermal mantle: BC, bottle cells; B, blastocoel. zone," which was defined as the region that is marginal to and above the blastopore. But with time the marginal zone involutes to form the mesodermal mantle and archenteron roof (Vogt, 1929; Keller, 1975, 1976). The lateness of the autonomous extension and convergence shown in classical works (see Spemann, 1938; Schechtman, 1942) suggests that these processes normally occur after involution, and are properties of the DMM or the archenteron roof rather than the marginal zone. The following experiments support this contention. does not show this behavior. When grafted to this site in an early gastrula (Fig. la), it shows little or no extension (Fig. 2a) until after it reaches the midgastrula stage. Then it begins to elongate, arch above the surface of the host, and form a notochord (Fig. 2b-e). Usually, the end that begins extension and arches above the surface of the host is the former vegetal end. The important point is that this extension occurs at developmental ages equivalent to the midgastrula and beyond, when normally the vegetal end of this region would have involuted. Grafts of DMZ to the animal pole Bilateral blockage of involution If extension of the preinvolution DMZ is active and autonomous, it would be expected to occur when the DMZ is grafted to the animal pole, a region that ordinarily Grafts of animal pole deep cells to the marginal zone will block involution in the region of the graft (see Keller, 1981). If bilateral grafts of deep and superficial layer 592 R. E. KELLER FIC. 2. A graft of early gastrula (stage 10) DMZ to the animal pole retains its initial shape until the midgastrula (stage 10.5) (a), at which time it begins to extend at its vegetal end through the late gastrula (stage 11.5, b; stage 12, c) and neurula (stage 15, d; stage 17, e). FIG. 3. Grafts of patches of early gastrula animal pole (shaded) to the dorsolateral marginal zone block involution on both sides of the DMZ. The DMZ extends beyond the blocking patches and begins to narrow from the midgastrula stage (a) onward through the late gastrula (c). From the early neurula (d) through the middle (e, f) and late neurula stages (g), the extension of the DMZ is dramatic. During the neurula stages, the lateral and ventral lips of the blastopore expand and secondarily uncover the yolk plug (YP). This extension of the DMZ (notochord) stretches the grafted patches (d-g). XEMOPUS GASTRULATION 593 FIG. 4. Photographs of neurulae derived from early gastrula that received grafts of animal pole material to the dorsolateral marginal zone. The grafted patches (pointers in a) can be seen on both sides of the DMZ (notochord) which has extended between them. The DMZ invariably skews off to the right or left and extends most of the distance across the newly exposed yolk plug. The yolk plug is re-exposed by extension (thick arrows, b) of the lateral and ventral lips of the blastopore (thin arrows, b). of the animal pole are grafted to the dorsolateral marginal zone (Fig. lb, c), the DMZ between them is isolated in situ by regions on either side that will not involute (Fig. 3). The blastopore continues to constrict but the patches of animal material refuse to move inside. The isolated dorsal sector begins to narrow, arch above the host, and extend beyond the lip of the blastopore (Fig. 3a-c). In the late gastrula (Fig. 3d) and neurula stages (Fig. 3e-g), the dorsal sector extends greatly. Histological sections (not shown) show these extensions to consist of notochord. The extensions stretch the grafted patches along their flanks as they elongate (Fig. 4a, b). The lateral and ventral lips begin to extend circumferentially in the late gastrula and neurula stages and dilate the blastopore (Fig. 3d-g; see arrows in Fig. 4b). Histological sections of the resulting "ring embryo" (see Schechtman, 1942) shows that the extending lateral and ventral sectors contain somites. The secondary dilation of the blastopore and the associated extension of the somitic mesoderm will be dealt with in detail in a later publication. The important point here is that the isolated, dorsal sector constricts and extends from the midgastrula or late midgastrula stage onward. 594 R. E. KELLER FIG. 5. A square patch of DMZ rotated 90 degrees clockwise in the early gastrula has begun to elongate to the left at its vegetal end by the midgastrula stage (a). It fails to involute but continues to elongate around the left side of the blastopore during the second half of gastrulation (b-c) and through the middle (d) and late (e) neurula stages. Note the progressively greater extension at the former vegetal end (marker dots, ac). NP, neural plate; YP, yolk plug. Ninety degree rotation of the DMZ If a small patch of DMZ is rotated clockwise 90 degrees (Fig. Id, e), it fails to involute, and it fails to extend or spread significantly during early gastrulation. In the midgastrula stage, it begins to extend (Fig. 5a), and by the late midgastrula stage, it has extended in the proper direction, with respect to its own axis, and around the left side of the blastopore, but it fails to involute (Fig. 5b, c). Note that it is the former vegetal end that begins to extend first (see markers, Fig. 5). Extension continues into the neurula stages and the redirected DMZ forms notochord around the inside of the enlarged left lip of the unclosed blastopore (Figs. 5d, e, 6a). Rotation of the deep region alone has similar effects (Fig. 6b), whereas rotation of the superficial layer alone usually, but not always, has little effect (Fig. 6c). The effect of 90 degree rotation is dependent on the size of the rotated patch. Small patches have less effect than large patches. If a large patch is rotated clockwise 90 degrees, it is not deflected to the left around the blastopore but behaves independently. It elongates parallel to its own animal-vegetal axis but perpendicular to that of the host (Fig. 6d). The former vegetal end (to the left, Fig. 6d) extends above the surface of the host and narrows in the process, whereas the opposite end remains broad (pointers, Fig. 6d). The extending DMZ often projects far from the XENOPUS GASTRULATION 595 Fie. 6. A midneurula (stage 15) after 90 degree clockwise rotation of the DMZ (a) shows incomplete blastopore closure and truncated neural plate (NP). The rotated DMZ has extended into the thickened, left lateral lip. Removal and replacement without rotation has little effect (left in b). Rotation of the deep region alone has nearly the effect of rotating both layers (right in b), whereas rotating the superficial layer alone usually has little effect (c). A large patch of DMZ rotated clockwise 90 degrees narrows and extends to the left (arrow), from its former vegetal end whereas the opposite end remains spread (pointers) by the late gastrula stage (g). Magnifications: a, 52; b, 21; c, 37; d, 62. surface of the embryo and sections show it to consist of notochord (data not shown). Three important facts emerge from these experiments. First, the redirected DMZ shows extension and convergence but again, only at midgastrula stages and beyond. Secondly, it shows extension and convergence from the vegetal end first. 596 R. E. KELLER Lastly, the DMZ can not involute when presented to the blastopore at right angles to its normal orientation. Conclusions lution (Keller, 1981). Grafts containing animal pole deep cells will block involution. Animal pole deep cells will actually pass through the zone of involution but then stop on the other side where they would be expected to leave the involution zone and join the involuted material (see Fig. 8b, Keller, 1981). This fact suggests that involution, like extension and convergence of the outer (preinvolution) DMZ, is a passive process, dependent on postinvolution events. Firstly, the preinvolution DMZ shows no detectable capacity to extend at developmental ages prior to the time at which it normally would have involuted. If a capacity to extend autonomously exists prior to the normal time of involution, it is too weak to distort the surrounding, uncooperative tissues in these experiments. This is ANALYSIS OF POSTINVOLUTION EVENTS unlikely. DMZs isolated in culture, without adjacent tissues to resist their extension, What postinvolution events drive the appear to show little extension until at least convergence and extension of the dorsal the late midgastrula stage (Holtfreter, sector of the gastrula? Do they occur in the 1939; Schechtman, 1942; Townes and deep region (the DMM) or the superficial Holtfreter, 1955; Ikushima and Maru- endodermal epithelium (the archenteron yama, 1971). The fact that the preinvolu- roof)? The answers are not clear. Grafting tion DMZ will not extend autonomously experiments show that the processes of until a developmental age when it normally convergence, extension, and involution all would have involuted suggests that the appear to be less sensitive to perturbations extension of the dorsal side of the embryo in the superficial layer than in the deep is normally due to a property of the DMZ region (Keller, 1981). Thus, for the after it has involuted to form the DMM moment, I will focus on the morphologies and the archenteron roof. and packing-patterns of the deep mesoSecondly, involution is not required for dermal cells that comprise the DMM as they extension, since the DMZ will extend on relate to postinvolution convergence and the outside of the embryo without extension. Earlier work focused on the fact undergoing the process of bending over that as they leave the involution zone, the the lip, either when grafted to the surface DMM cells appear to form a "stream" of of the host, as was the case in the present cells in which individuals migrate toward experiments, or in organ culture (see Holt- the animal pole on the inner surface of the freter, 1939, 19436; Schechtman, 1942; preinvolution material (Nakatsuji, 1975, Townes and Holtfreter, 1955). Thirdly, 1976; Keller and Schoenwolf, 1977). This extension occurs first and most extensively work emphasizes the interaction of deep from the vegetal end of explanted or redi- mesodermal cells with the inner surface of the gastrular wall. I will return to this idea rected patches of DMZ. Fourthly, it is important to note that the and suggest that it be modified somewhat. lateral side of a patch of DMZ rotated 90 Now I wish to examine the possibility that degrees will not involute, even though it there are interactions between these lies within the material of the marginal zone mesodermal cells that bring about postincapable of involution (see Keller, 1976, volution convergence and extension. 1981). This suggests that the material of the marginal zone must be presented to the Morphology and pattern of DMM cells involution zone with the proper axial oriThe overlying preinvolution material was entation in order for involution to occur. dissected off, after fixation, and the outer Lastly, involution itself appears to be surface of the involuted DMM was examdependent on events that occur after cells ined with stereo SEM by methods similar have passed through the involution zone. to those described in detail elsewhere (KelThe type of deep cell is critical for invo- ler and Schoenwolf, 1977). Patterns of cell XENOPUS GASTRULATION arrangement and shape that may be related to the processes of dorsal convergence and extension were seen. At the midgastrula stage, cells in the DMM appear to be more tightly packed with smaller intercellular spaces than those lateral to the DMM (Fig. 7a, b). High magnification (Fig. 7c) shows that cells in this region have less relief and are more tightly packed than those located laterally. Many are elongated and aligned transversely to the animal-vegetal axis (Fig. 7c). This pattern is more pronounced among cells just inside the zone of involution, at the vegetal extremity of the mesodermal mantle, where most are elongated parallel to the circumference of the blastopore (Fig. 7d). This band of elongate cells extends laterally into the dorsolateral sectors of the posterior mesodermal mantle and is contiguous with the shield of compacted cells extending anteriorly from it (Fig. 7a, b). In late gastrulation, a similar but longer and narrower area is found in the DMM, extending anteriorly from the zone of involution (Fig. 8a, b). Cells in this region are distinguished from those on either side by the presence of large, lamellar protrusions, a tighter packing pattern with fewer intercellular spaces and a much more flattened morphology. These characteristics are shared with the corresponding area of the midgastrula, but their expression is subtle at the midgastrula whereas it is more extreme in the late gastrula. This region is clearly the developing notochord at the late gastrula stage, and it is set off distinctly from the paraxial mesoderm (Fig. 8b; also see Fig. 2, Youn et ai, 1980). In the early neurula, the notochord consists of a more compact, narrow array of cells clearly separated from the prospective somite mesoderm lateral to it (Fig. 8c). There is good evidence that the area of unique cell shape and arrangement in the DMM of the mid (Fig. 7), and late (Fig. 8) gastrula stages are homologous to the notochord (Fig. 8c) in the neurula. These regions in the gastrula and neurula map as prospective notochord (Keller, 1976, and unpublished data). The less-ordered, indistinct packing-pattern and morphology of this region of the midgastrula develops 597 without discontinuity into the highly ordered cell shape and arrangement characteristic of the notochord in the neurula. Convergence and extension by mediolateral interdigitation? There is no direct evidence pertaining to the behavior of the DMM cells described above in SEMs. There is indirect evidence, however, that these cells undergo mediolateral interdigitation and repacking to form an array that is narrower and longer (convergence and extension). The dorsal convergence of the DMM in the gastrula and early neurula stages is extraordinary, and this dramatic movement dorsally is expressed primarily as an increase in length (Keller, 1975, 1976), rather than as an increased thickness, though the developing notochord does thicken slightly (Fig. 8b). These facts imply that DMM cells moving dorsally must repack, mediolaterally, to form a mass of greater anterioposterior extent. Direct evidence on this point is needed. The emergence of the dorsal packingpattern is spatially and temporally coincident with the increasing capacity for autonomous extension in the second half of gastrulation and in the neurula. The packing-pattern and the capacity to extend autonomously are both associated with the prospective or developing notochord. Notochord is invariably found in the protrusions that break away from the surface of the gastrula (see Schechtman, 1942). Thus a major part of the dorsal convergence and extension in the gastrula stage and notochordal extension in the neurula stages may be based on similar cellular morphologies, packing-patterns and behavior. Though the capacity for convergence and extension in the dorsal sector is normally coincident with the notochord anlage, such capacity is not strictly associated with notochord formation. The lateral blastoporal lips of "ring embryos" described above and in Schechtman (1942) contain only somites but participate in constriction of the blastopore and then undergo dramatic extension. Moreover, irradiated embryos that never form a B FIG. 7. A posteriodorsal view of the involuted mesodermal mantle of a midgastrula (stage 10.5) shows the DMM cells (pointers) characterized by low relief, closer packing, and smaller intercellular spaces (a, b). High magnification clearly shows the difference between medial and lateral cells in these respects (c). The vegetal end of the DMM, just inside the mesodermal involution zone (MIZ), shows closely packed mesodermal cells elongated — — 1 AA. U OAA. „ XENOPUS GASTRULATION 599 Fie. 8. A posteriodorsal view of the DMM of a late gastrula (stage 11.5) shows an elongate region of closepacked cells extending anteriorly from the dorsal mesodermal involution zone (MIZ) (a). Higher magnification shows these cells, which are prospective notochord, to be more flattened, closely-packed, and bearing more lamelliform protrusions than the somite mesoderm to either side (b). A dorsal view of the late gastrula (stage 13) shows the contrast between the cells of the notochord (N) and paraxial mesoderm (PM) (c). Magnifications: a, 100; b, 250; c, 500. proper notochord show extension (Malacinski and Youn, 1981). CELLULAR MECHANISMS OF GASTRULATIONS Previous proposals Previously, two alternative mechanisms were proposed to explain spreading or epiboly of the preinvolution region of the gastrula (Keller, 1980, 1981). According to the first, autonomous spreading of the DMZ occurs by active, radial interdigitation of several layers of deep cells to form fewer layers of greater area (see Fig. 19, Keller, 1980). This proposal was supported by the evidence from the literature, which appeared to favor the active, autonomous extension of the marginal zone and animal region (see Keller, 1980). However, the tests of these alternatives, presented here, strongly suggest that spreading of the DMZ is passive, and thus favor the second mechanism. According to this mechanism, the superficial epithelium would be stretched by tension generated at the blastopore, and deep cells would rearrange and occupy the additional space made available on the inner surface of the superficial layer (see Fig. 19e,f, Keller, 1980). The movement of the deep cells would not, in itself, force the spreading of the marginal zone, though this is the only sense in which their behavior would be considered passive. Their apposition to the inner surface of the superficial epithelium and their repacking to form fewer layers could be viewed as a rachet mechanism—not actively forcing the expansion of the epithelium but stabilizing it once it was stretched by external forces. A new mechanism: Active, postinvolution convergence and extension by mediolateral interdigitation The above evidence suggests that the external force that stretches the DMZ is 600 R. E. KELLER DMZ FIG. 9. The Postinvolution Convergence and Extension Model is illustrated diagrammatically in a lateral view of a midgastrula with the right dorsal and lateral marginal zone removed (a) to expose the dorsal mesodermal mantle (DMM), and in a gastrula section midsagittally (b). The cells of the DMM (outlined) merge dorsally by mediolateral interdigitation (inset, a), beginning just inside the zone of involution. The DMM narrows (converges) as indicated by the arrows pointing dorsally in the DMM (a). Such narrowing is expressed by lengthening (extension) of the DMM at the middorsal region (dashed arrow, a) and by formation of a constriction zone in the DMM just inside the blastoporal lip. The constriction zone and the extension of the DMM result in the DMM pushing vegetally, on the inside of the blastoporal lip (short arrows, a) and concurrently constricting the blastopore. As the DMM pushes vegetally (short arrow, b) on the inner surface of the superficial epithelium (SE), the overlying dorsal marginal zone (DMZ) is towed vegetally (curved arrows, b). Deep cells of the marginal zone (shaded) are recruited to join the posterior DMM (b). The bottle cells (BC) anchor the superficial epithelium to the mesodermal mantel (see Keller, 1981). most likely generated by postinvolution events in the DMM. The following cellular model of gastrulation, based on the arguments above, is proposed. The object of proposing this model is to stimulate research that will either support or refute its principal elements, which are as follows. Firstly, there are deep mesodermal cells in the DMZ, spread far laterally on both sides of the dorsal midline, that have, as part of their morphogenetic repertoire, the capacity to converge dorsally by active, mediolateral interdigitation (Fig. 9a). Secondly, these cells begin to exercise this part of their morphogenetic repertoire just after they leave the zone of involution and join the vegetal end of the mesodermal mantle (Fig. 9a, b). There they merge between one another and converge on the dorsal midline. This behavior is characteristic of and continues to occur among involuted cells along the length of the DMM as well as at its vegetal end. Thus width of the vegetal end of the DMM decreases and forms a constriction ring in the mesodermal mantle just inside the blastoporal lip (Fig. 9a, b). Because this constriction ring acts below the equator of a spherical embryo, it tends to pull the blastoporal lip vegetally, across the yolk plug (Fig. 9a). Because convergence begins earlier in dorsal and dorsolateral sectors, the blastoporal lips in these regions move greater distances across the yolk plug and close the blastopore eccentrically, which is the case (Keller, 1975). In addition, the mediolateral interdigitation XEXOPUS GASTRULATION of cells toward the dorsal midline would narrow the DMM along its entire length (convergence) and increase its length, causing extension vegetally across the yolk plug in concert with the constriction movement of the blastoporal lips. Lastly, preinvolution (marginal zone) deep cells are continually recruited to join the vegetal edge of the DMM. The requirement for successfully doing so is the adoption of the behavioral rules (or rule) that underlie active, mediolateral interdigitation. Determining the cellular, motile behavior underlying mediolateral interdigitation of DMM cells is essential in our understanding of gastrulation in Xenopus and probably many other anurans as well. Holtfreter (19436) and Townes and Holtfreter (1955) noted that the isolated DMZ would elongate and form notochord if left intact but would form notochord tissue without elongating if dissociated and reaggregated. This fact might suggest that the order of neighbor relationships are required expression of a morphogenetic function (extension) but no histodifferentiation (of notochord tissue). Alternatively, it could reflect a greater sensitivity of the former process to the trauma of dissociation. 601 involves lateral and ventral sectors. Thus these processes begin earlier, continue longer, and involve more cells in the dorsal sector than in lateral and ventral sectors (see Keller, 1976). However, Schechtman (1942) suggested that lateral and ventral sectors have properties that are different from the dorsal sector and that these regions must interact in order to produce gastrulation. His results and interpretation are consistent with those presented here in that the local rule of cellular behavior— the postinvolution, mediolateral interdigitation—must operate with the dorsal, lateral and ventral sectors' circumblastoporal continuity in order to be effective in producing involution. However, it is not clear whether differences in dorsal and lateral or ventral behavior are matters of timing, degree, or character of behavior. It is also not clear now convergence and extension of the marginal zone at a given stage is related to the notochordal and somitic mesoderm. It is clear that anterioposteriorly organized events—extension and involution—are intimately coupled to dorsoventrally organized events—dorsal convergence and constriction of the blastopore—because they are results of a common process—postinvolution mediolateral interdigitation. Mediolateral and anterioposterior organization of convergence and extension Postinvolution convergence and transitions in cell behavior The above mechanism does not apply to the initial movement of the blastoporal lip vegetally and the initial involution of the marginal zone. The anterior (early involuting) mesodermal mantle has little capacity for convergence and extension, based on the evidence presented here and by Schechtman (1942) and, in fact, does not converge or extend (Keller, 1976). Studies in progress show that other processes, including bottle cell formation and mesodermal migration, have a greater role in the initial movements; these will be described elsewhere. The degree of convergence and extension increases progressively from anterior to posterior in the mesodermal mantle. Moreover, it begins dorsally at about the midgastrula stage, and progressively The postinvolution convergence and extension model is economical in terms of the cellular behavior required since it accounts for the movements of convergence, extension, and involution with one basic cellular, motile process—active mediolateral interdigitation—acting in a specific set of global geometrical constraints. One transition in cellular behavior is required just after the cells involute. In contrast, the idea that the preinvolution marginal zone actively extends toward the blastopore requires that the deep cells of the marginal zone first interdigitate radially and the entire marginal zone would have to form a stiff plate, capable of pushing toward the involution zone (see Keller, 1980). Then, in the involution zone, the cells must change behavior again, in such 602 R. E. KELLER embryonalen Formbildung. Arch. Exp. Zella way as to allow the stiff plate to bend forsch. besonders Gewebequecht 23:169-209. around the blastoporal lip. Finally, on leavHoltfreter, J. 1943a. Properties and function of the ing the involution zone, they would have surface coat in amphibian embryos. J. Exp. Zool. to change behavior again and migrate on 93:251-323. the roof of the preinvolution material, Holtfreter, J. 1943A. A study of the mechanics of gastrulation. Part I. J. Exp. Zool. 94:261-318. toward the animal pole (see Nakatsuji, Ikushima, N. and S. Maruyama. 1971. Structureand 1975, 1976; Keller and Schoenwolf, 1977). developmental tendency of the dorsal marginal Such mechanical and behavioral complexzone in the early amphibian gastrula. J. Embryol. ity seems unlikely. Exp. Morphol. 25:263-276. Johnson, K. 1977a. Changes in the cell coat at the onset of gastrulation in Xenopus laevis embryos. J. Exp. Zool. 199:137-143. Johnson, K. 19776. Extracellular matrix synthesis in Earlier work suggested that the invoblastula and gastrula stages of normal and hybrid frog embryos. I. Toluidine blue and lanthanum luted deep mesodermal cells were orgastaining. J. Cell Sci. 25:313-322. nized as a stream of cells that actively K. 1977c. Extracellular matrix synthesis in migrate upwards, as individuals, toward the Johnson, blastula and gastrula stages of normal and hybrid animal pole, on the inner surface of the frog embryos. II. Autoradiographic observations preinvolution material (see Nakatsuji, on the sites of synthesis and mode of transport of galactose- and glacosamine-labelled materials. 1975, 1976; Keller and Schoenwolf, 1977). J. Cell Sci. 25:323-334. This is probably true of the early involutK. 1977a1. Extracellular matrix synthesis in ing regions when extension and conver- Johnson, blastula and gastrula stages of normal and hybrid gence have no major role, but in the late frog embryos. III. Characterization of galactoseinvoluting regions (posterior mesodermal and glucosamine-labelled materials. J. Cell Sci. 25:335-356. mantle) the coordinated interactions between mesodermal cells that underlie Johnson, K. E. 1984. Glycoconjugate synthesis during gastrulation in Xenopus laevis. Amer. Zool. 24: mediolateral interdigitation are probably 605-614. more important, at least in Xenopus. Such Keller, R. 1975. Vital dye mapping of the gastrula may not be true of the urodeles, such as and neurula of Xenopus laevis. I. Prospective areas and morphogenetic movements of the superficial the Mexican axolotl, which appear to have layer. Develop. Biol. 42:222-241. a less cohesive mesodermal mantle in which R. 1976. Vital dye mapping of the gastrula cells migrate as individuals (see Nakatsuji, Keller, and neurula of Xenopus laevis. II. Prospective areas 1984). and morphogenetic movements of the deep region. Develop. Biol. 51:118-137. Keller, R. 1978. Time-lapse cinemicrographic analACKNOWLEDGMENTS ysis of superficial cell behavior during and prior to gastrulation in Xenopus laevis. J. Morph. 157: I wish to thank Steven D. Black, Robert 223-248. Gimlich, and George Oster for their con- Keller, R. 1980. The cellular basis of epiboly: An tributions in stimulating discussions on gasSEM study of deep cell rearrangement during trulation and Paul Tibbitts and John Shih gastrulation in Xenopus laevis. J. Embryol. Exp. Morphol. 60:201-234. for their technical assistance. This work R. 1981. An experimental analysis of the role was supported by NSF Grant PCM81- Keller, of the bottle cells and the deep marginal zone in 10985 to the author. gastrulation of Xenopus laei'is. J. Exp. Zool. 216: 81-101. Keller, R. and G. Schoenwolf. 1977. An SEM study REFERENCES of cellular morphology, contact, and arrangeBaker, P. 1965. Fine structure and morphogenetic ment as related to gastrulation in Xenopus laei'is. movements in the gastrula of the the treefrog, Wilhelm Roux' Arch. Develop. Biol. 182:165Hyla regilla. J. Cell Biol. 24:95-116. 186. Cooke, J. 1975. Local autonomy of gastrulation Kosher, R. and R. Searles. 1973. Sulfated mucomovements after dorsal lip removal in two anuran polysaccharide synthesis during the development amphibians. J. Embryol. Exp. Morphol. 33:147— of Rana pipiens. Develop. Biol. 32:50-68. 157. Lee, G., R. O. Hynes, and M. Kirschner. 1982. TemDaniel, J. F. and E. Yarwood. 1939. The early poral and spatial expression of fibronectin in early embryology of Triturus torosus. Univ. of Calif. Publ. Xenopus development. J. Cell Biol. 95:135a. Zool. 43:321-356. Lewis, W. H. 1947. Mechanics of invagination. Anat. Holtfreter, J. 1939. Gewebeaffinitat, ein Mittel der Rec. 97:139-156. Mediolateral interdigitation versus migration of mesoderm XENOPUS GASTRULATION Malacinski, G. and B.-W. Youn. 1981. Neural plate morphogenesis and axial stretching in "notochord defective" Xenopus laevis embryos. Develop. Biol. 88:352-357. Nakatsuji, N. 1975. Studies on the gastrulation of amphibian embryos: Cell movement during gastrulation in Xenopus laevis embryos. Wilhelm Roux' Arch. Develop. Biol. 178:1-14. Nakatsuji, N. 1976. Studies on the gastrulation of amphibian embryos: Ultrastructure of the migrating cells of anurans. Wilhelm Roux' Arch. Develop. Biol. 180:229-240. Nakatsuji, N. 1984. Cell locomotion and contact guidance in amphibian gastrulation. Amer. Zool. 24:615-627. Nieuwkoop, P. and P. Florshutz. 1950. Quelquescaracteres speciaux de la gastrulation et de la neurulation de l'oeuf de Xenopus laevis, Daud. et de quelques autres Anoures. 1 ere partie.—Etude descriptive. Arch. Biol. (Liege) 61:113-150. Pasteels, J. 1942. New observations concerning the maps of presumptive areas of the young amphibian gastrula (Amblystoma and Discoglossus). J. Exp. Zool. 89:255-281. Perry, M. and C. H. Waddington. 1966. Ultrastructure of the blastoporal cells in the newt. J. Embryol. Exp. Morphol. 15:317-330. 603 Schaeffer, B., H. Schaeffer, and 1. Brick. 1973. Cell electrophoresis of amphibian blastula and gastrula cells; the relationship of surface charge and morphogenetic movement. Develop. Biol. 34:6676. Schechtman, A. M. 1942. The mechanism of amphibian gastrulation. I. Gastrulation-promoting interactions between various regions of an anuran egg (Hyla regilla). Univer. Calif. Publ. Zool. 51:1-39. Spemann, H. 1938. Embryonic development and induc- tion. Yale University Press. Reprinted 1962, Hafner Publishing Company, Inc., New York. Townes, P. L. and J. Holtfreter. 1955. Directed movements and selective adhesion of embryonic amphibian cells. J. Exp. Zool. 128:53-120. Vogt, W. 1929. Gestaltungsanalyse am Amphibienkeim mit ortlicher Vitalfarbung. II. Teil. Gastrulation und Mesodermbildung bei Urodelen und Anuren. Wilhelm Roux' Arch. Entwicklungsmech. Org. 120:384-706. Youn, B.-W. and G. Malacinski. 1981. Neural plate morphogenesis and axial stretching in "notochord-defective" Xenopus laevis embryos. Develop. Biol. 88:352-357.