Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
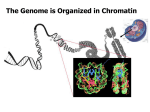
The nucleus as an organized organelle • Individual chromosomes occupy distinct domains within the interphase nucleus without overlapping other chromosomes • The telomeres (ends of chromosomes), and condensed, transcriptionally inactive regions (heterochromatin) are attached to the nuclear envelope (via the lamina) • Ribosomal RNA (rRNA) is synthesized in a site corresponding to the nucleolus • Pre-mRNAs are synthesized at 20-50 sites that appear as speckles under fluorescent microscopy • Rather than diffusing away from their sites of synthesis, pre-mRNAs stream out towards the nuclear pores while they are being processed into mature mRNAs Spliced introns Pre-mRNAs Processed mRNAs (b) The nuclear matrix • Treatment of eukaryotic nuclei with non-ionic detergents and high salt, removes the histones and non-histone proteins from chromatin, leaving a halo of DNA • Digestion of DNA fibers with a nuclease (Dnase) leaves a network of thin, protein containing fibers The nuclear matrix • Newly transcribed pre-mRNAs are associated with matrix fibers, and treatment with an Rnase destroys the matrix. Id nuclear matrix as a dynamic (moving) ribonucleoprotein infrastructure • The matrix is attached to the nuclear envelope, and may represent an internal scaffolding for the nucleus Packaging the genome: Chromatin structure Levels of condensation 1. DNA wraps twice around a core of histones (H2A, H2B, H3, H4), forming nucleosomes 2. Nucleosome filaments (beadson-a-string) coil into “30 nm fibers” 3. Fibers attach at intervals to a protein anchor forming “loops” approx. 50 kbp in length 4. During mitosis, the chromosome condenses to ~1 cm DNA/µm DNA double helix (2 nm in diameter) DNA Nucleosome core particle Histones (8 subunits) Nucleosome filament (10 nm in diameter) 30 nm fiber Looped domains Metaphase chromosome H1 histone Replicating linear DNA: Telomerase • The lagging strand of the telomeres is synthesized by telomerase (reverse transcriptase) • Telomerase activity is very low in most cells. Shortening of telomeres → aging • Some cancer cells are “immortal” (divide indefinitely); contain high telomerase activity → a target for anticancer drugs Telomerase RNA 3' 5' U C 1 CCCCAACCCCAACCC 5' U A A AAA CCCCAAC GGGGTTGGGGTTGGGGTTGGGGTTGGGG 3' Elongation CCCCAACCCCAACCC 5' U A A A AACCCCAAC GGGGTTGGGGTTGGGGTTGGGGTTGGGGTTG Translocation UU C A 3' 3' 5' U C 3 CCCCAACCCCAACCC C A 3' 5' U C 2 UU 5' U A A A AACCCCAAC U U GGGGTTGGGGTTGGGGTTGGGGTTGGGGTTG 3' Elongation 3' 5' U C 4 CCCCAACCCCAACCC 5' U A A U A AA CCCCAAC U GGGGTTGGGGTTGGGGTTGGGGTTGGGGTTGGGGTTG (b) C A 3' C A Control of gene expression in eukaryotes • Occurs at three primary levels: 1. Transcriptional-level control 2. Processing-level control 3. Translational-level control • Transcriptional-level control Steps in mRNA synthesis that are regulated: 1. Initiation: General factors (TFIID, TFIIB, TFIIH, TFIIE, TFIIJ, etc..), accessory factors that bind to enhancer sequences, and RNA polymerases (RNP I, II, and III) Accessory factors bind to upstream (or downstream) enhancers, and either increase (activation) or decrease (repression) the rate for assembly of a PIC (pre-initiation complex) on the promoter (TATA + initiator sequences) Enhancer DNA-binding domains Transcription activating domains TFIIE TFIIJ TATA TFIIB TFIIH TFIID TFIIF Initiator RNAPII Start site DNA Control of gene expression in eukaryotes Many DNA-binding proteins bind only to defined sequences. They contain domains e.g. bHLH, bZIP, zinc-finger, etc.. with amino acids that project into the major or minor grooves, where they hydrogen bond with the DNA bases Often, cooperative binding through multiple contact surfaces e.g. two subunits of a dimer, increases selectivity. 3' 5' 108 Basic region 124 166 Helix 1 Helix 2 137 146 Loop 5' 3' Role of nucleosome positioning in transcription Nucleosome • Nucleosomes may block the access of TAFs to promoters ⇒ repression • Nucleosome structure may also aid cooperative interactions between distant sites ⇒ activation • Histone:TAF interactions also influence TAF binding to promoters Enhancer Start site Nucleosome Transcription factor Promoter (a) Transcription factor Enhancer Promoter Nucleosome (b)