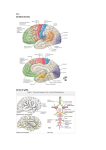
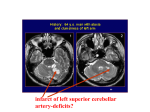
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Cognitive neuroscience of music wikipedia , lookup
Aging brain wikipedia , lookup
Executive functions wikipedia , lookup
Apical dendrite wikipedia , lookup
Cortical cooling wikipedia , lookup
Neuroesthetics wikipedia , lookup
Neuroeconomics wikipedia , lookup
Nervous system network models wikipedia , lookup
Subventricular zone wikipedia , lookup
Psychophysics wikipedia , lookup
Metastability in the brain wikipedia , lookup
Neuroanatomy wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Neuroanatomy of memory wikipedia , lookup
Development of the nervous system wikipedia , lookup
Anatomy of the cerebellum wikipedia , lookup
Emotional lateralization wikipedia , lookup
Synaptic gating wikipedia , lookup
Neural coding wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Time perception wikipedia , lookup
C1 and P1 (neuroscience) wikipedia , lookup
Optogenetics wikipedia , lookup
Cerebral cortex wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
Neural correlates of consciousness wikipedia , lookup
Eyeblink conditioning wikipedia , lookup
Channelrhodopsin wikipedia , lookup
Proc. Natl. Acad. Sci. USA Vol. 93, pp. 739–743, January 1996 Neurobiology Formation of mnemonic neuronal responses to visual paired associates in inferotemporal cortex is impaired by perirhinal and entorhinal lesions (memoryymedial temporal lobeybackward connectionylearningyprimate) SEI-ICHI HIGUCHI AND YASUSHI MIYASHITA* Department of Physiology, The University of Tokyo School of Medicine, 7-3-1, Hongo, Bunkyo-ku, Tokyo 113, Japan Communicated by Larry R. Squire, University of California, San Diego, CA, July 18, 1995 preparation so that individual IT neurons became devoid of the backward signals but could receive forward visual signals when visual stimuli were presented. We found selective loss of the ability of IT neurons to represent associations between picture pairs, whereas the ability of the neurons to respond to particular visual stimuli was left intact. ABSTRACT Functional roles of the cortical backward signal in long-term memory formation were studied in monkeys performing a visual pair-association task. Before the monkeys learned the task, the anterior commissure was transected, disconnecting the anterior temporal cortex of each hemisphere. After training with 12 pairs of pictures, single units were recorded from the inferotemporal cortex of the monkeys as the control. By injecting a grid of ibotenic acid, we unilaterally lesioned the entorhinal and perirhinal cortex, which provides massive direct and indirect backward projections ipsilaterally to the inferotemporal cortex. After the lesion, the monkeys fixated the cue stimulus normally, relearned the preoperatively learned set (set A), and learned a new set (set B) of paired associates. Then, single units were recorded from the same area as for the prelesion control. We found that (i) in spite of the lesion, the sampled neurons responded strongly and selectively to both the set A and set B patterns and (ii) the paired associates elicited significantly correlated responses in the control neurons before the lesion but not in the cells tested after the lesion, either for set A or set B stimuli. We conclude that the ability of inferotemporal neurons to represent association between picture pairs was lost after the lesion of entorhinal and perirhinal cortex, most likely through disruption of backward neural signals to the inferotemporal neurons, while the ability of the neurons to respond to a particular visual stimulus was left intact. METHODS The cerebral cortex is organized so that hierarchically different subareas communicate by forward and backward neuronal projections (1); however, no functional role of the backward connections has so far been experimentally demonstrated at the single-neuron level. Neuropsychological studies have proposed a hypothesis for the role of the backward connections from limbic cortex to neocortex in the formation of long-term memory for facts and events (declarative memory) (2–5). Behaviorally the declarative memory has often been tested in humans by learning of a series of paired words or figures (2–3, 5–8). Previous work with monkeys identified a group of neurons which acquired associative mnemonic codes for pictures through paired-associate learning in the anterior inferotemporal (IT) cortex (9), a visual memory storehouse in primates (2–5, 8). There is strong evidence that IT cortex receives direct and indirect backward projections from the perirhinal and entorhinal cortex (10–13). The hypothesis for the role of the backward connections predicts that the lesion of the perirhinal and entorhinal cortex would impair the formation of the associative code for pictures in the IT cells. The present study tested this prediction. We combined single-unit recording techniques with a surgically manipulated chronic monkey Animals and Surgical Procedures. Two adult monkeys (Macaca fuscata) were used in accordance with the University of Tokyo Animal Care Committee. Surgery on the anterior commissure (AC) was carried out under aseptic conditions and anesthesia with sodium pentobarbital (25–30 mgykg). The right hemisphere was retracted from the falx with a brain spoon. An aspirator was used to make a sagittal incision #5 mm in length in the corpus callosum, entering the lateral ventricle at the level of the interventricular foramen. The AC was sectioned at the floor of the lateral ventricle lateral to the fornix. Antibiotics were delivered to prevent infection, and the monkeys were given sufficient rest for recovery for at least 2 weeks. The lesions of the entorhinal and perirhinal cortex were made by injecting a small volume of the neurotoxin ibotenic acid (Ibo). Ibo has been reported to destroy cell bodies selectively while leaving fibers of passage in the underlying white matter undamaged (14, 15). With the help of electrophysiological mapping, we inserted a Hamilton syringe needle into the anteroventral temporal lobe. A grid of 1-ml injections of Ibo (15 mgyml, 0.03 mlymin) was made with '2 mm between tracks. One monkey (M) received injections at 22 tracks, and the other monkey (N) at 19 tracks. Behavioral Task. The pair-association task was described previously (9). Colored fractal patterns were used as the visual stimuli (see Fig. 3A). We generated the patterns according to the algorithm described previously (16) and arranged them into 12 pairs. In the task, following a fixation stimulus, a cue stimulus was presented in the center of the video monitor for 0.5 s. After a delay period, the paired associate of the cue stimulus and one stimulus from a different pair were shown. The monkey obtained fruit juice as a reward for touching the correct paired associate. Eye position was monitored with a magnetic search coil. The criterion for acquisition of preoperative learning (the set A stimuli) was two consecutive days of 26 correct responses in 30 trials. Monkey M and monkey N took 270 and 383 trials per picture to reach this criterion, respectively. The criterion after the Ibo lesion was made slightly easier (two consecutive days of 23 correct responses in 30 trials). In relearning the set A paired associates after the lesion, 10 and 50 trials per picture were necessary for monkeys The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked ‘‘advertisement’’ in accordance with 18 U.S.C. §1734 solely to indicate this fact. Abbreviations: AC, anterior commissure; IT, inferotemporal; CC, correlation coefficient; PI, pair index; Ibo, ibotenic acid. *To whom reprint requests should be addressed. 739 740 Proc. Natl. Acad. Sci. USA 93 (1996) Neurobiology: Higuchi and Miyashita M and N, respectively. In learning the new set (set B) after the lesion, one monkey (M) took 101 trials and the other (N) took 93 trials per picture to reach the criterion. Electrophysiology and Data Analysis. Recording and analysis of extracellular spike discharges of single neurons have been described (9, 17). At the beginning of unit recording before the lesion, locations of responsive cells were mapped in the anterior IT cortex and an area (about 2 mm 3 3 mm, hatched in Fig. 2) with a high density of responsive cells was identified. All neuronal recordings were done from this cortical area both before and after the lesion. Each location of the electrode track was measured by x-ray imaging and verified histologically at the end of the experiments. Evaluation of cue responses was done by collecting number of spikes over 80–480 ms after the presentation of each picture cue, and for 400 ms immediately preceding the appearance of the fixation stimulus. A neuron was regarded as responsive to the picture if the two distributions were significantly different (paired t test, P , 0.05) and as selective if the response to a stimulus was significantly different from others by analysis of variance (ANOVA, P , 0.05). Two indices were used to analyze pair-coding responses of these cells: the standard correlation coefficient (CC) and the pair index (PI) (9). The CC was calculated from xi and x9i (i 5 1–12) for each cell, where xi denotes a mean discharge rate during the cue period for the ith picture (the ith and i9th pictures belong to a pair): O 12 CC 5 HF O 12 i51 i51 @~xi 2 x!~x9i 2 x9!# ~xi 2 x!2 GF O 12 i51 ~x9i 2 x9!2 GJ 1y2 , where x and x9 are the means of xi and x9i (i 5 1–12), respectively. The PI was defined (9) as the difference of two correlation indices, CIp and CIr, where for paired associates, CIp 5 1 Np OO i j ~ i, j ! ~ j5i9 ! ~xi 2 b!~xj 2 b! 3 100, ~xbest 2 b!~x2nd-best 2 b! and for other combinations, CIr 5 1 Nr OO j i ~ i,j) (jÞi9) ~xi 2 b!~xj 2 b! 3 100, ~xbest 2 b!~x2nd-best 2 b! where b is a spontaneous discharge rate, xbest is the mean discharge rates for the stimulus that produced best responses in each cell, and x2nd-best is the mean discharge rate for the second-best cue-optimal pictures. Np and Nr are the total number of combinations for two cases. The CC and PI were calculated for each cell. Histology. At the end of the experiment, the monkey was deeply anesthetized with an overdose of sodium pentobarbital (45 mgykg, i.m.) and then perfused through the aorta with 0.9% NaCl followed by 10% formalin in phosphate buffer (pH 7.4). The brain was sectioned coronally at 40 mm. Serial sections were stained with cresyl violet and with the modified Gallyas method (18). Seventeen (monkey M) or 18 (monkey N) sections were sampled at 760- to 960-mm separation, and a two-dimensional unfolded map was constructed (19). The shrinkage due to histological procedures was estimated and adjusted by a correction factor, but a possible local atrophy due to the Ibo lesion (see Results) was not corrected. RESULTS Two macaque monkeys were trained in a pair-association task (9). We sorted a set of 24 geometrically distinct pictures into 12 pairs, so that the monkeys learned the combinations of the paired associates and retrieved the other pair member associated with each cue picture. Before the monkeys learned the task, the AC was surgically transected (Fig. 1 A and B) just to the right of the midline (arrow in Fig. 1B). The septal nuclei, the fornix, and the underlying diencephalic structures were spared. This surgery disconnected the anterior temporal cortex of each hemisphere from the other, since most commissural fibers of this area traverse the AC rather than the corpus callosum (20–22). After recovery, the monkey was trained with a set (set A) of the paired associates to the criterion performance level, and then extracellular spike discharges of single neurons were recorded as a prelesion control in the anterior IT cortex (9, 23, 24). We deprived IT neurons of backward neural information by unilateral Ibo lesions of the entorhinal and perirhinal cortex. Fig. 1 C–E show photographs of a cross section from the anterior temporal cortex of a lesioned monkey. The lesion covered both the medial and lateral banks of the rhinal sulcus completely, and most of the entorhinal and perirhinal cortex. The lesioned cortex suffered atrophy, but fibers in the underlying white matter were left undamaged. Higher magnification (Fig. 1E) of the area framed in Fig. 1D indicates the border FIG. 1. Unilateral Ibo lesion of the perirhinal and entorhinal cortex. (A) Lateral (Left) and ventral (Right) view of a monkey brain, indicating the locations of sections in B (no. 1), C–E (no. 2), and Fig. 2 (nos. 3 and 4). (B) Fiber-stained coronal section through the AC. The AC was cut (arrow) just to the right of the midline (M). The fornix (Fx) was left intact. (Bar 5 2 mm.) (C) Cresyl violet-stained section showing the left (intact) entorhinal and perirhinal cortex. (Bar 5 2 mm.) (D) Same section as C, but showing the right (lesioned) cortex. (E) Higher magnification of the area framed in D. Double arrowheads indicate the border between the intact area (to the right) and incompletely lesioned area. Arrowheads indicate the border between the completely (to the left) and incompletely lesioned area. H, hippocampus. amts, anterior middle temporal sulcus. sts, superior temporal sulcus. rs, rhinal sulcus. Neurobiology: Higuchi and Miyashita FIG. 2. Location and extent of the ibotenic acid lesion in monkey M (A) and monkey N (B), illustrated on flat maps of the temporal lobe. Anterior (A), posterior (P), medial (M), and lateral (L) directions are indicated. Dot-dash lines represent cut lines of map; thick lines indicate the lips of sulci. Completely (coarsely stippled) and incompletely (finely stippled) lesioned areas are shown. In the flat map of monkey M, the location of sections 1 and 2 of Fig. 1 A is illustrated with thin line, and the dot-dash lines correspond to sections 3 and 4 of Fig. 1 A. Locations of recorded neurons are also indicated on the maps (hatched area). PA, periamygdaloid cortex. PreS, presubiculum. amts, anterior middle temporal sulcus. ots, occipito-temporal sulcus. sts, superior temporal sulcus. rs, rhinal sulcus. between the intact and lesioned area. In the intact area (to the right) the laminar organization of the normal neocortex remained, while in the lesioned area (to the left) no neurons were found (only glial cells). Fig. 2 shows the location and extent of the lesions on flat maps of the temporal lobe for each monkey. The lesion was nearly complete along most of the anteroposterior extent of the rhinal sulcus. However, the most anterior portion of the rhinal sulcus on the temporal pole is not shown in the flat maps, and the cortex in this area was not damaged in either monkey. The most lateral part of the perirhinal cortex in one monkey (N) was spared. After the lesion, extracellular spike discharges of single neurons were recorded from the same area (hatched in Fig. 2) as in the prelesion control. Proc. Natl. Acad. Sci. USA 93 (1996) 741 Average performance for set A stimuli during neural recording was 81% (before lesion) and 80% (after the lesion), and for set B stimuli, 72% after the lesion. In spite of the lesion of the entorhinal and perirhinal cortex, the sampled neurons responded strongly to both the set A and set B patterns during the cue period of the task. The responses of one cell are shown in Fig. 3 B and C. This cell was activated selectively by a few stimuli of set B (Fig. 3D) and very weakly by the stimuli of set A (Fig. 3E). The lesion did not affect the monkey’s fixation to the cue stimulus as shown in the eye position traces of Fig. 3 B and C. In contrast to these unaffected properties, the strongest and the second-strongest responses of the cell were not ascribed to a particular pair of the set (Fig. 3D), and thus the cell did not encode the pair association. We recorded from 256 neurons prior to the Ibo lesion and tested the responses to the set A stimuli. One hundred nine of them were unresponsive. Since weak responses could not be ascribed to the optimal stimulus (25), we examined 92 cells whose maximum discharge rates were distributed beyond 20 spikes per second (the leftmost saddle point in the distribution of all responsive cells in the control and lesioned cortices). Of the 92 cells, 10 cells were nonselective to the stimuli. We analyzed pair-coding responses of these cells by calculating the CC and the PI for each neuron. Fig. 4A (●) shows the cumulative frequency distribution of CC values in the 92 cells. The distribution of CC was shifted toward the positive values and showed that the paired associates elicited significantly correlated responses in the prelesion control (P , 0.01; Wilcoxon signed-rank test, n 5 92). Fig. 4B (●) similarly shows the cumulative frequency distribution of PI values in the 92 cells. The curve also shows that paired associates elicited significantly correlated responses in the control (P , 0.01, n 5 92). These results confirmed the conclusion of a previous report (9). We recorded from 363 neurons after the lesion, and tested the responses to set A stimuli for 341 cells and the responses to set B stimuli for 332 cells. Of those cells tested with the set A stimuli, 202 were unresponsive and 67 gave weak responses. FIG. 3. Responses of anterior IT neurons in the pair-association task. (A) Examples of pairs of fractal patterns. (B) Single-neuron response after the Ibo acid lesion, shown by rastergrams (Top), spike-density histograms (Middle), and eye position traces (Bottom; H, horizontal. V, vertical). Bin width, 40 ms. The green square for fixation or the cue stimulus was presented during the time period denoted as f or c. These trials were collected for a cue picture 7 in the stimulus set B. (C) Same as B, but showing trials for cue 1 of set B. (D) Mean discharge rates for each cue presentation from set B (mean 6 SEM) relative to the spontaneous discharge (arrowhead) in the same cell as in B and C. Twelve pairs of stimuli are labeled on the abscissa. The stippled and open bars in pair no. 1 refer to the responses to the picture 1 and 19, respectively. This neuron selectively responded to patterns of set B but did not code pair associations. (E) Mean discharge rates of the same cell as in D, but for pictures of another set (set A). The fractal patterns of pairs 7, 9, and 1 of set B are shown in the top, middle, and bottom rows of A. 742 Neurobiology: Higuchi and Miyashita Proc. Natl. Acad. Sci. USA 93 (1996) correlated responses before the lesion (11 cells) was larger than that after the lesion for set A (5 cells) or set B (1 cell), and the percentages of the correlated cells were statistically significantly different (x2 5 7.06, P , 0.05). The frequency distributions of PI after the lesion (Fig. 4B) also show that the paired associates did not elicit significantly correlated responses either for set A (V, P . 0.3) or for set B (‚, P . 0.4). Fig. 5 compares the responsiveness of the cells in the three groups (two-way ANOVA and t test). The results for the two monkeys were not significantly different (P . 0.25) in the two-way ANOVA in any of the following indices [F(1, 233) 5 0.01 for CC, F(1, 233) 5 1.11 for PI, F(1, 233) 5 0.69 for SPON, F(1, 233) 5 0.00 for MAX] and have been pooled. The CCs were reduced after the lesion compared with that before the lesion (P , 0.05). The CCs in the control were significantly larger than zero (P , 0.01) but those after the lesion were not, either for set A (P . 0.4) or for set B (P . 0.6). The PIs were also reduced after the lesion compared with that before the lesion (P , 0.05). The PIs before the lesion were significantly larger than zero (P , 0.05) but those after the lesion were not, either for set A (P . 0.6) or for set B (P . 0.4). After the lesion, the cells responded to both the set A and set B pictures, even more strongly than the control (MAX, P , 0.05). The spontaneous discharge rates (SPON) were not significantly different among the three groups (P . 0.2). We conclude that lesion of the entorhinal and perirhinal cortex disrupted the associative code of the IT neurons between the paired associates, without impairing the visual response to each stimulus. FIG. 4. Response correlation to paired associates. Analysis by the CC (A) and by the PI (B). The ordinates indicate the cumulative number of cells, which was normalized by the total number of cells in each group. The curves show the cumulative frequency histograms in the three cell groups: neurons tested with the set A stimuli prior to the lesion of the entorhinal and perirhinal cortex (●, n 5 92), those tested after the lesion with the set A stimuli (V, n 5 72), and those tested after the lesion with the set B stimuli (‚, n 5 75). Of those cells tested with the set B stimuli, 195 were unresponsive and 62 gave weak responses. Thus, 72 of the 341 cells for set A and 75 of the 332 cells for set B were analyzed after the lesion. The fraction of responsive cells (72y341 for set A, 75y332 for set B) dropped in the lesion groups (P , 0.01) compared with that before the lesion (92y256), which may reflect some physiological effects of the lesion. Seven of the 72 cells and 8 of the 75 cells were nonselective to the stimuli in the two postlesion groups. Thus, the fraction of visually selective cells after the lesion (65y72 and 67y75) did not differ significantly from that before the lesion (82y92) (P . 0.8). We also compared the response selectivity by calculating the number of effective stimuli (NES) for each cell. The 50%, 10%, and 90% points of the cumulative distributions of the NES were determined. There was a slight tendency that the cells in the lesioned groups were less selective (50%, 10%, and 90% values of NES were 3.0, 1.4, and 12.0 before lesion; 5.0, 1.8, and 17.3 for set A; and 5.0, 1.5, and 15.9 for set B after the lesion). However, the tendency was not statistically significant (P . 0.15; Kolmogorov–Smirnov test) between the control group and the lesioned group for either set A or set B. Fig. 4A shows that the cumulative frequency distributions of CC after the lesion (V and ‚) were significantly different from the distribution before the lesion (●) (P , 0.05; Kolmogorov– Smirnov test) and that the distributions after the lesion did not differ from each other (P . 0.5). The paired associates did not elicit significantly correlated responses after the lesion either for set A (V; P . 0.9; Wilcoxon signed-rank test) or for set B (‚; P . 0.9). We also used the CC to evaluate the number of individual cells which showed significantly correlated responses (at P , 0.05); the number of cells with significantly DISCUSSION The results support the hypothesis that the backward signal from the medial temporal lobe to the IT neurons plays a crucial role in visual pair-association memory in the primate. The hypothesis raised a specific prediction that the lesion of the perirhinal and entorhinal cortex would impair the formation of the associative code for pictures in the IT cells. The present study tested this prediction and confirmed it. FIG. 5. Effects of the lesion of the entorhinal and perirhinal cortex upon neuronal responsiveness in the pair-association task. The CC (A), PI (B), spontaneous discharge rate (SPON) (C), and maximum discharge rate with spontaneous discharge subtracted (MAX) (D) were compared among three groups: neurons that were recorded with the set-A stimuli prior to the lesion (stippled bars, n 5 92), those recorded with the set A stimuli after the lesion (hatched bars, n 5 72), and those recorded with the set B stimuli after the lesion (solid bars, n 5 75). Data are shown as mean 6 SEM. Proc. Natl. Acad. Sci. USA 93 (1996) Neurobiology: Higuchi and Miyashita We combined single-unit recording techniques with surgical manipulations in the monkey. First, the AC of the monkeys was cut to remove influences upon anterior IT neurons from those in the other hemisphere (20–22). Behaviorally, the monkeys fixated the stimuli and learned the pair-association task quite normally, just as in the previous study (9). Neuronal responses of IT cells to complex patterns (control in Fig. 5D) were not affected by the anterior commissurectomy, which confirmed a previous report on anesthetized monkeys (26). The response correlation to the paired associates was also not affected (control in Figs. 4 A and B and 5 A and B). Then, we placed a unilateral Ibo lesion in the entorhinal and perirhinal cortex. The electrophysiological indices for spontaneous discharge rates and maximum firing rates (Fig. 5 C and D) suggested that the cortex of the recording area was not pathological after the limbic lesion, which was also confirmed by normal histology of the cortex at the recording sites. Visual selectivity was another factor which may affect the pair-coding property of a cell. However, the percentage of visually selective cells did not differ significantly before and after the lesion. The number of effective stimuli of each cell exhibited a slight tendency for the control group to be more sharply tuned, but the tendency was not statistically significant. All these data suggest that the major effect of the lesion can be specifically related to the pair-coding properties of IT cells. A previous lesion study demonstrated that monkeys with bilateral removal of the entorhinal and perirhinal cortex neither relearn visual paired associates that were learned preoperatively nor learn new associations among new visual stimuli (27). In the present study monkeys with a unilateral lesion learned both the old and new associations easily. The mild behavioral effects of the unilateral lesion were most likely due to the intact temporal lobe in the contralateral hemisphere. At the level of neuronal responses, the lesion destroyed the associational ability of the IT neurons that had been isolated from the intact temporal lobe in the contralateral hemisphere by the anterior commissurectomy. The finding of retrograde amnesia, in terms of neuronal responses, also agreed with previous behavioral data (27). The pair association that was learned prior to the lesion was lost (Fig. 5 A and B, set A after the lesion). Long and gradual learning of the paired associates prior to the lesion (.3 months) may have produced such longer retrograde amnesia than that found by Zola-Morgan and Squire in an object-discrimination problem (28). To examine implications of the results, we need to consider the limitations of this animal preparation. The Ibo lesion obliterated most directly the backward projection from entorhinal–perirhinal cortex to IT neurons, but eliminated other, indirect neural connections to IT cells as well: the lesion damaged cells with forward projection from entorhinal– perirhinal cortex to other structures, such as the frontal cortex, the cingulate cortex, or the basal ganglia. Thus, the Ibo lesion might have altered the functional circuitry for an anterior IT neuron in a complicated manner. It remains to be examined whether the indirect functional circuitry could explain the observed changes of associative coding in the IT neurons. In the primate inferior temporal cortex and part of the superior temporal cortex, neurons selectively respond to complex objects such as faces, hands, Fourier descriptors, and fractal patterns (5, 25, 29–31). Some, if not all, of these responses are acquired through learning in adulthood (5, 9, 23, 31–33). The present results show that the backward connection from the medial temporal lobe is necessary for some of these acquired responses, such as pair-association coding, but not for 743 others. Indeed, the fractal patterns introduced after the lesion (set B) activated IT neurons highly selectively. Since this stimulus selectivity is most likely obtained by a tuning mechanism that molds and sharpens neuronal response selectivity to an object through repetitive stimulus presentation (33, 34), our finding suggests at least partial preservation of the tuning mechanism after the lesion of the entorhinal and perirhinal cortex. These results are consistent with the proposal of decomposition of the primate long-term memory system into several different neural mechanisms (2, 8, 33, 34). We thank Dr. K. Sakai for helpful discussions. This research was supported by Grants-in-Aid for Specially Promoted Research from the Japanese Ministry of Education, Science, and Culture (02102008, 07102006) and a grant from Uehara Memorial Foundation. 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. Van Essen, D. C., Anderson, C. H. & Felleman, D. J. (1992) Science 255, 419–423. Squire, L. R. (1987) Memory and Brain (Oxford Univ. Press, New York). Milner, B. (1990) in Vision, Memory and the Temporal Lobe, ed. Iwai, E. & Mishkin, M. (Elsevier, New York), pp. 43–53. Mishkin, M. (1982) Philos. Trans. R. Soc. London B 298, 85–92. Miyashita, Y. (1993) Annu. Rev. Neurosci. 16, 245–263. Wechsler, D. (1987) Wechsler Memory-Scale Revised (Harcourt Brace Jovanovich, San Antonio, TX). Petrides, M. (1985) Neuropsychologia 23, 601–614. Squire, L. R. & Zola-Morgan, S. (1991) Science 253, 1380–1386. Sakai, K. & Miyashita, Y. (1991) Nature (London) 354, 152–155. Van Hoesen, G. W. (1982) Trends Neurosci. 5, 345–353. Shiwa, T. (1987) Arch. Ital. Biol. 125, 139–154. Webster, M. J., Ungerleider, L. G. & Bachevalier, J. (1991) J. Neurosci. 11, 1095–1116. Suzuki, W. A. & Amaral, D. G. (1994) J. Neurosci. 14, 1856–1877. Olney, J. W. (1983) Excitotoxins (Macmillan, London), pp. 82–95. Newsome, W. T., Wurtz, R. H., Dursteler, M. R. & Mikami, A. (1985) Exp. Brain Res. 58, 392–399. Miyashita, Y., Higuchi, S., Sakai, K. & Masui, N. (1991) Neurosci. Res. 12, 307–311. Miyashita, Y., Rolls, T., Cahusac, P. M. B., Niki, H. & Feigenbaum, J. D. (1989) J. Neurophysiol. 61, 669–678. Hess, T. D. & Merker, B. H. (1983) J. Neurosci. Methods 8, 95–97. Van Essen, D. C. & Maunsell, J. H. R. (1980) J. Comp. Neurol. 191, 225–281. Pandya, D. N., Karol, E. A. & Lele, P. P. (1973) Brain Res. 49, 177–180. Zeki, S. M. (1973) J. Comp. Neurol. 148, 167–176. Demeter, S., Rosene, D. L. & Van Hoesen, G. W. (1990) J. Comp. Neurol. 302, 29–53. Miyashita, Y. (1988) Nature (London) 335, 817–820. Miyashita, Y. & Chang, H. S. (1988) Nature (London) 331, 68–70. Gross, C. G. (1972) in Handbook of Sensory Physiology (Springer, Berlin), pp. 451–482. Gross, C. G., Bender, D. B. & Mishkin, M. (1977) Brain Res. 131, 227–239. Murray, E. A., Gaffan, D. & Mishkin, M. (1993) J. Neurosci. 13, 4549–4561. Zola-Morgan, S. & Squire, L. R. (1990) Science 250, 288–290. Desimone, R., Albright, T. D., Gross, C. G. & Bruce, C. (1984) J. Neurosci. 4, 2051–2062. Rolls, E. T. & Baylis, G. C. (1986) Exp. Brain Res. 65, 38–48. Perrett, D. I., Mistlin, A. J. & Chitty, A. J. (1987) Trends Neurosci. 10, 358–364. Miyashita, Y., Date, A. & Okuno, H. (1993) Neuropsychologia 31, 1119–1132. Sakai, K., Naya, Y. & Miyashita, Y. (1994) Learn. Mem. 1, 83–105. Squire, L. R. (1994) in Advances in the Study of Memory and Memory Systems, eds. Schacter, D. & Tulving, E. (MIT Press, Cambridge, MA), pp. 203–231.