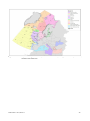
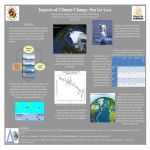
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
ICES ADVICE 2013 AVIS DU CIEM Books 1- 10 Report of the ICES Advisory Committee 2013 Book 1 Introduction, Overviews and Special Requests International Council for the Exploration of the Sea Conseil International pour l’Exploration de la Mer H.C. Andersens Boulevard 44-46 DK-1553 Copenhagen V Denmark Telephone (+45) 33 38 67 00 Telefax (+45) 33 93 42 15 www.ices.dk [email protected] Report of the ICES Advisory Committee 2013. Books 1 - 10 December 2013 Recommended format for purposes of citation: ICES. 2013. Report of the ICES Advisory Committee 2013. ICES Advice, 2013. Books 1-11, 2095 pp. ICES. 2013. Report of the ICES Advisory Committee 2013. ICES Advice, 2013. Book 1. 348 pp. For permission to reproduce material from this publication, please apply to the General Secretary. ISBN 978-87-7482-119-9 TABLE OF CONTENTS ICES ADVICE 2013 BOOK 1 Section 1 Page INTRODUCTION, OVERVIEW AND SPECIAL REQUESTS .......................................................................... 1 1.1 About ICES ................................................................................................................................................... 1 1.2 1.3 1.4 1.5 General context of ICES advice .................................................................................................................... 3 Technical basis for the advice ..................................................................................................................... 25 Structure of the Report ................................................................................................................................ 28 Answers to non-Ecoregion specific Special Requests................................................................................. 30 1.5.1 EU DG Mare .............................................................................................................................. 30 1.5.1.1 Request from EU concerning monitoring of bycatch of cetaceans and other protected species ........................................................................................................................ 30 1.5.1.2 New information regarding the impact of fisheries on other components of the ecosystem ................................................................................................................... 34 1.5.1.3 Request from EU concerning monitoring of bycatch of seabirds ............................... 39 1.5.2 EU DGENV ............................................................................................................................... 48 1.5.2.1 Request from EU for scientific advice on data collection issues ................................ 48 1.5.2.2 Request from EU for scientific advice on data collection issues – part 2 ................... 59 1.5.3 1.5.4 1.5.5 HELCOM................................................................................................................................... 72 NASCO ................................................................................................................................... 72 NEAFC ................................................................................................................................... 73 1.5.5.1 1.5.5.2 1.5.5.3 1.5.5.4 1.5.5.5 1.5.5.6 1.5.5.7 1.5.6 OSPAR ................................................................................................................................. 195 1.5.6.1 Ecological quality objective for seabird populations in OSPAR Region III (Celtic Seas) ......................................................................................................................... 195 Data collection and storage to implement the OSPAR seabird recommendations .. 205 OSPAR special request on review of the technical specification and application of common indicators under D1, D2, D4, and D6 ........................................................ 208 OSPAR special request on maximizing the use of available sources of data for monitoring of biodiversity ........................................................................................ 222 OSPAR/NEAFC special request on review of results of the Joint OSPAR/NEAFC/ CBD Workshop on EBSAs (same as 1.5.5.5)........................................................... 232 OSPAR/NEAFC special request on Existing and potential new management measures for EBSAs (same as 1.5.5.6)..................................................................................... 270 OSPAR/NEAFC special request on review and reformulation of four EBSA Proformas (same as 1.5.5.7) ..................................................................................... 277 OSPAR request on spatial design of a regional monitoring programme for contaminants in sediments ........................................................................................ 327 1.5.6.2 1.5.6.3 1.5.6.4 1.5.6.5 1.5.6.6 1.5.6.7 1.5.6.8 1.5.7 1.6 1.7 1.8 1.9 Vulnerable deep-water habitats in the NEAFC Regulatory Area ............................... 73 Evaluation of the appropriateness of buffer zones...................................................... 83 Assessment of the list of VME indicator species and elements .................................. 85 Advice on threshold levels for long line fishing ......................................................... 98 OSPAR/NEAFC special request on review of results of the Joint OSPAR/NEAFC/ CBD Workshop on EBSAs (same as 1.5.6.5)............................................................. 99 OSPAR/NEAFC special request on Existing and potential new management measures for EBSAs (same as 1.5.6.6)..................................................................................... 137 OSPAR/NEAFC special request on review and reformulation of four EBSA Proformas (same as 1.5.6.7) ..................................................................................... 144 Member States ......................................................................................................................... 334 General trends in the Northeast Atlantic ................................................................................................... 335 Acronyms and terminology ....................................................................................................................... 341 Maps ................................................................................................................................................. 345 List of Reviewers ...................................................................................................................................... 348 ICES Advice 2013, Book 1 i BOOK 2 Section 2 Page ICELAND AND EAST GREENLAND ................................................................................................................... 1 2.1 Ecosystem Overview ...................................................................................................................................... 1 2.2 The status of fish stocks and fisheries ............................................................................................................ 1 2.3 Ecosystem Assessments and Advice .............................................................................................................. 5 2.3.1 2.3.2 2.3.3 Assessments and advice regarding protection of biota and habitats............................................... 5 Assessment and advice regarding fisheries .................................................................................... 5 Special Requests ............................................................................................................................ 6 2.3.3.1 Request from Iceland to ICES to evaluate longterm management plan and harvest control rule for Icelandic haddock ................................................................................................ 6 2.3.3.2 Request from Iceland to ICES to evaluate longterm management plan and harvest control rule for Icelandic saithe .................................................................................................. 16 2.4 Stock summaries 2.4.1 2.4.2 2.4.3 2.4.4 2.4.5 2.4.6 2.4.7 2.4.8 2.4.9 2.4.10 2.4.11 2.4.12 2.4.13 2.4.14 ii ....................................................................................................................................... 29 Capelin in Subareas V and XIV and Division IIa west of 5°W (Iceland–East Greenland–Jan Mayen area) 29 Inshore Cod in NAFO Subarea 1 (Greenland Cod)...................................................................... 35 Offshore Cod in ICES Subarea XIV (Greenland cod) ................................................................. 41 Cod in Division Va (Icelandic cod).............................................................................................. 43 Greenland halibut in Subareas V, VI, XII and XIV ..................................................................... 51 Haddock in Division Va (Icelandic haddock) .............................................................................. 64 Herring in Division Va (Icelandic summer-spawning herring) .................................................... 71 Introduction to the redfish complex in Subareas V, VI, XII, XIV ............................................... 77 Beaked Redfish (Sebastes mentella) in Division Va and Subarea XIV (Icelandic Slope stock)...91 Beaked Redfish (Sebastes mentella) in Subareas V, XII, XIV and NAFO Subareas 1+2 (Shallow Pelagic stock <500 m) .................................................................................................. 99 Beaked Redfish (Sebastes mentella) in Subareas V, XII, XIV and NAFO Subareas 1+2 (Deep Pelagic stock >500 m)............................................................................... 108 Beaked Redfish (Sebastes mentella) in subarea XIVb (Demersal) ............................................ 117 Golden Redfish (Sebastes marinus) in Subareas V, VI, XII and XIV ....................................... 118 Saithe in Division Va (Icelandic saithe) ..................................................................................... 120 ICES Advice 2013, Book 1 BOOK 3 Section 3 Page THE BARENTS SEA AND THE NORWEGIAN SEA ......................................................................................... 1 3.1 Ecosystem overview ....................................................................................................................................... 1 3.2 The status of fish stocks and fisheries ............................................................................................................ 1 3.3 Ecosystem Assessments and Advice .............................................................................................................. 5 3.4 3.3.1 Assessments and advice regarding protection of biota and habitats............................................... 5 3.3.2 Assessment and advice regarding fisheries .................................................................................... 5 3.3.3 Special Requests ............................................................................................................................ 6 3.3.3.1 Status and harvest potential of the harp seal stocks in the Greenland Sea and the White Sea/Barents Sea, and of the hooded seal stock in the Greenland Sea ..................................... 6 Stock summaries ....................................................................................................................................... 21 3.4.1 3.4.2 3.4.3 3.4.4 3.4.5 3.4.6 3.4.7 3.4.8 3.4.9 Capelin in Subareas I and II, excluding Division IIa west of 5°W (Barents Sea capelin) ........... 21 Cod in Subareas I and II (Northeast Arctic cod) .......................................................................... 32 Cod in Subareas I and II (Norwegian coastal waters cod) ........................................................... 43 Haddock in Subareas I and II (Northeast Arctic) ......................................................................... 50 Greenland halibut in Subareas I and II ........................................................................................ 60 Beaked redfish (Sebastes mentella) in Subareas I and II ............................................................. 66 Golden redfish (Sebastes marinus) in Subareas I and II .............................................................. 74 Saithe in Subareas I and II (Northeast Arctic) ............................................................................. 81 Northern shrimp (Pandalus borealis) in Subareas I and II (Barents Sea) .................................... 89 ICES Advice 2013, Book 1 iii BOOK 4 Section 4 Page THE FAROE PLATEAU ECOSYSTEM ............................................................................................................... 1 4.1 Ecosystem Overview ...................................................................................................................................... 1 4.2 The status of fish stocks and fisheries ............................................................................................................ 1 4.3 Ecosystem Assessments and Advice ............................................................................................................. 6 4.3.1 4.3.2 4.4 iv Assessments and advice regarding protection of biota and habitats............................................... 6 Assessment and advice regarding fisheries .................................................................................... 6 Stock summaries ......................................................................................................................................... 7 4.4.1 Cod in Subdivision Vb1 (Faroe Plateau) ........................................................................................ 7 4.4.2 Cod in Subdivision Vb2 (Faroe Bank).......................................................................................... 15 4.4.3 Haddock in Division Vb .............................................................................................................. 20 4.4.4 Saithe in Division Vb ................................................................................................................... 29 ICES Advice 2013, Book 1 BOOK 5 Section 5 Page CELTIC SEA AND WEST OF SCOTLAND ......................................................................................................... 1 5.1 5.2 5.3 5.4 Ecosystem Overview ...................................................................................................................................... 1 The status of fish stocks and fisheries ............................................................................................................ 1 Ecosystem Assessments and Advice ........................................................................................................... 13 5.3.1 Assessments and advice regarding protection of biota and habitats............................................. 13 5.3.2 Assessment and advice regarding fisheries .................................................................................. 16 5.3.3 Special requests ............................................................................................................................ 17 5.3.3.1 Request from the European Commission on distribution of the stock of megrims in Subarea IV and Division VIa .......................................................................................................................... 17 5.3.3.2 Request from NEAFC to evaluate the proposals for the harvest control components of the management plan for Rockall haddock fisheries.......................................................................... 20 5.3.3.3 Request from NEAFC on the closure area and additional measures for the protection of juvenile haddock on Rockall Bank ............................................................................................................ 28 Fish stock advice ....................................................................................................................................... 34 5.4.1 Anglerfish (Lophius piscatorius and L. budegassa) in Divisions IIIa, and Subareas IV and VI .. 34 5.4.2 Anglerfish (Lophius piscatorius and L. budegassa) in Divisions VIIb-k and VIIIa,b,d .............. 51 5.4.3 Cod in Division VIa (West of Scotland) ...................................................................................... 59 5.4.4 Cod in Division VIb (Rockall) ..................................................................................................... 72 5.4.5 Cod in Division VIIa (Irish Sea) .................................................................................................. 74 5.4.6 Cod in Divisions VIIe-k (Celtic Sea cod) .................................................................................... 76 5.4.7 Grey gurnard in Subarea VI and Divisions VIIa-c and e-k (Celtic Sea and West of Scotland) ... 85 5.4.8 Haddock in Division VIa (West of Scotland)............................................................................... 87 5.4.9 Haddock in Division VIb (Rockall) ............................................................................................. 98 5.4.10 Haddock in Division VIIa (Irish Sea)......................................................................................... 108 5.4.11 Haddock in Divisions VIIb-k ..................................................................................................... 117 5.4.12 Herring in Division VIa North ................................................................................................... 124 5.4.13 Herring in Divisions VIa (South) and VIIb,c ............................................................................. 135 5.4.14 Herring in Division VIIa North of 52° 30´ N (Irish Sea) ........................................................... 143 5.4.15 Herring in Divisions VIIa South of 52° 30´ N and VIIg,h,j,k (Celtic Sea and South of Ireland) ......................................................................................................................... 151 5.4.16 Lesser-spotted dogfish (Scyliorhinus canicula) in Subdivisions VI, VIIa-c, e-j ........................ 161 5.4.17 Megrim (Lepidorhombus spp.) in Divisions IV (North Sea) and Division Via (West of Scotland) ...................................................................................................................... 162 5.4.18 Megrim (Lepidorhombus spp) inDivision VIb (Rockall) ........................................................... 172 5.4.19 Megrim (Lepidorhombus whiffiagonis) in Divisions VIIb-k and VIIIa,b,d ............................... 178 5.4.20 Nephrops in Division VIa .......................................................................................................... 181 5.4.20.1 Nephrops in North Minch (FU 11) ............................................................................. 189 5.4.20.2 Nephrops in the South Minch (FU 12) ....................................................................... 198 5.4.20.3 Nephrops in the Firth of Clyde + Sound of Jura (FU 13) ........................................... 207 5.4.21 Nephrops in Subarea VII ............................................................................................................ 220 5.4.21.1 Nephrops in Irish Sea East (FU14) ............................................................................. 229 5.4.21.2 Nephrops in Irish Sea West (FU 15) .......................................................................... 236 5.4.21.3 Nephrops on Porcupine Bank (FU 16) ....................................................................... 246 5.4.21.4 Nephrops on Aran Grounds (FU 17) ........................................................................... 255 5.4.21.5 Nephrops off the south-eastern and south-western coasts of Ireland (FU 19) ............. 264 5.4.21.6 Nephrops in the FU 20 (Labadie, Baltimore and Galley), FU 21 (Jones and Cockburn)................................................................................................................................... 273 5.4.21.7 Nephrops in the Smalls (FU 22) .................................................................................. 277 5.4.22 Norway pout in Division VIa (West of Scotland) ...................................................................... 284 5.4.23 Plaice in Division VIIa (Irish Sea) ............................................................................................. 286 5.4.24 Plaice in Division VIIb,c (West of Ireland)................................................................................ 294 5.4.25 Plaice in Division VIIe (Western Channel) ................................................................................ 296 5.4.26 Plaice in Divisions VIIf,g (Celtic Sea) ....................................................................................... 305 5.4.27 Plaice in Divisions VIIh-k (Southwest of Ireland) ..................................................................... 313 5.4.28 Pollack in SubareasVI and VII (Celtic Sea and West of Scotland) ........................................... 319 5.4.29 Rays and Skates in the Celtic Seas ecoregion ............................................................................ 321 5.4.29.1 Rays and Skates in the Celtic Seas ecoregion ............................................................. 323 ICES Advice 2013, Book 1 v 5.4.30 5.4.31 5.4.32 5.4.33 5.4.34 5.4.35 5.4.36 5.4.37 5.4.38 5.4.39 5.4.40 5.4.41 5.4.42 5.4.43 5.4.44 vi 5.4.29.2 Blonde ray (Raja brachyura) in Subarea VI ............................................................... 325 5.4.29.3 Blonde ray (Raja brachyura) in Divisions VIIa,f,g..................................................... 325 5.4.29.4 Blonde ray (Raja brachyura) in Division VIIe ........................................................... 326 5.4.29.5 Thornback Ray (Raja clavata) in Sybarea VI ............................................................. 327 5.4.29.6 Thornback Ray (Raja clavata) in Divisions VIIa,f,g .................................................. 328 5.4.29.7 Thornback ray (Rajaclavata) in Division VIIe............................................................ 329 5.4.29.8 Small-eyed ray (Raja microocellata) in Divisions VIIfg ............................................ 330 5.4.29.9 Small-eye ray (Raja microcellata) in Division VIIe ................................................... 331 5.4.29.10 Spotted ray (Raja montagui) in Subarea VI ............................................................... 332 5.4.29.11 Spotted ray (Raja montagui) in Divisions VIIa,f,g .................................................... 333 5.4.29.12 Raja undulata in Division VIIj ................................................................................... 334 5.4.29.13 Sandy ray (Leucoraja circularis) in the Celtic Seas.................................................. 335 5.4.29.14 Shagreen ray (Leucoraja fullonica) in the Celtic Seas ............................................... 336 5.4.29.15 Cuckoo ray, Leucoraja naevus in Subarea VI and Division VIIa-c, e-j .................... 337 5.4.29.16 Common skate Dipturus batis complex (flapper skate Dipturus cf. flossada and blue skate Dipturus cf. intermedia) in Subarea VI, and Divisions VIIa-c, e-j ................... 338 5.4.29.17 Other ray and skate species in Subarea VI, and Divisions VIIa-c, e-j ....................... 338 Saithe in Subarea VI (West of Scotland and Rockall) ............................................................... 339 Sandeel in Division VIa (West of Scotland) .............................................................................. 340 European seabass in Divisions IVbc, Via and VIId-h (Irish Sea, English Channel and southern North Sea) .................................................................................................................................. 341 European seabass in Divisions Via, VIIb and VIIj (West of Scotland and Ireland) ................... 349 Sole in Division VIIa (Irish Sea) ................................................................................................ 355 Sole in Division VIIb,c (West of Ireland) .................................................................................. 362 Sole in Division VIIe (Western Channel) .................................................................................. 364 Sole in Divisions VIIf,g (Celtic Sea) ......................................................................................... 372 Sole in Division VIIh-k (Southwest of Ireland) ......................................................................... 380 Sprat in Subarea VI and Divisions VIIa-c and f-k (Celtic Sea and West of Scotland)............... 386 Sprat in Divisions VIId,e (Channel) ........................................................................................... 388 Whiting in Division VIa (West of Scotland) .............................................................................. 395 Whiting in Division VIb (Rockall)............................................................................................. 403 Whiting in Division VIIa (Irish Sea) .......................................................................................... 405 Whiting in Divisions VIIe-k (Celtic Sea whiting) ...................................................................... 407 ICES Advice 2013, Book 1 BOOK 6 Section 6 Page NORTH SEA ......................................................................................................................................... 1 6.1 Ecosystem Overview ...................................................................................................................................... 1 6.2 The status of fish stocks and fisheries ............................................................................................................ 1 6.3 Ecosystem Assessments and Advice ............................................................................................................ 15 14 6.3.1 6.3.2 6.3.3 6.3.4 6.3.5 6.4 Multispecies considerations for the North Sea stocks (WGSAM) .............................................. 15 Mixed-fisheries advice for Subarea IV (North Sea) and Divisions IIIa North (Skagerrak) and VIId (Eastern Channel) (WGMIXFISH) ..................................................................................... 24 Assessments and advice regarding protection of biota and habitats............................................. 34 Assessment and advice regarding fisheries .................................................................................. 34 Special requests ............................................................................................................................ 35 6.3.5.1 EU request on changing the TAC year for Norway pout in the North Sea .................... 35 6.3.5.2 Joint EU-Norway request to evaluate the long-term management plan for whiting in the North Sea ....................................................................................................................... 42 6.3.5.3 EU request on inter-annual quota flexibility plaice in the North Sea ............................ 49 6.3.5.4 EU request on inter-annual quota flexibility saithe in the North Sea............................. 52 6.3.5.5 Joint EU-Norway request on TAC setting options for cod in the North Sea and Skagerrak ....................................................................................................................... 55 Fish stock advice 6.4.1 6.4.2 6.4.3 6.4.4 6.4.5 6.4.6 6.4.7 6.4.8 6.4.9 6.4.10 6.4.11 6.4.12 6.4.13 6.4.14 6.4.15 6.4.16a 6.4.16b 6.4.17 6.4.18 6.4.19 6.4.20 6.4.21 6.4.22 Brill in Subarea IV and Divisions IIIa and VIId,e ....................................................................... 63 Cod in Division IIIa East (Kattegat) ............................................................................................ 69 Cod in Subarea IV (North Sea) and Divisions VIId (Eastern Channel) and IIIa West (Skagerrak) 71 Dab in Subarea IV and Division IIIa ............................................................................................ 91 Flounder in Subarea IV and Division IIIa .................................................................................... 98 Grey gurnard in Subarea IV (North Sea) and Divisions VIId (Eastern Channel) and IIIa (Skagerrak - Kattegat)…………………………………………………………………104 Haddock in Subarea IV (North Sea) and Division IIIa West (Skagerrak).................................. 105 Herring in Division IIIa and Subdivisions 22–24 (Western Baltic spring spawners)................. 119 Herring in Subarea IV and Divisions IIIa and VIId (North Sea autumn spawners) ................... 130 Horse mackerel (Trachurus trachurus) in Divisions IIIa, IVb,c and VIId (North Sea stock) .... 148 Lemon sole in Subarea IV, and Divisions IIIa and VIId ………………………………………155 Lesser-spotted dogfish (Scyliorhinus canicula) in IIIa, IV and VIId ........................................ 160 Striped red mullet in Subarea IV (North Sea) and Divisions VIId (Eastern English Channel) and IIIa (Skagerrak–Kattegat) .......................................................................................................... 161 Nephrops in Division IIIa........................................................................................................... 169 Nephrops in Division IV (North Sea) ....................................................................................... 176 6.4.15.1 Nephrops in Botney Gut – Silver Pit (FU 5) .............................................................. 184 6.4.15.2 Nephrops in Farn Deeps (FU 6) .................................................................................. 185 6.4.15.3 Nephrops in Fladen Ground (FU 7) ........................................................................... 192 6.4.15.4 Nephrops in Firth of Forth (FU 8) ............................................................................... 199 6.4.15.5 Nephrops in Moray Firth (FU 9) ................................................................................. 207 6.4.15.6 Nephrops in Noup (FU 10).......................................................................................... 214 6.4.15.7 Nephrops in Norwegian Deeps (FU 32) ...................................................................... 215 6.4.15.8 Nephrops off Horn’s Reef (FU 33) ............................................................................ 216 6.4.15.9 Nephrops in Devil’s Hole (FU 34) ............................................................................... 217 Norway pout in Subarea IV (North Sea) and Division IIIa (Skagerrak – Kattegat) June advice 217 Norway pout in Subarea IV (North Sea) and Division IIIa (Skagerrak – Kattegat) October advice.......................................................................................................................................... 228 Plaice in Division IIIa West (Skagerrak) ................................................................................... 237 Plaice in Subarea IV (North Sea) ............................................................................................... 248 Plaice in Division VIId (Eastern Channel) ................................................................................. 261 Pollack in Subarea IV and Division IIIa ................................................................................... 267 Saithe in Subarea IV (North Sea) Division IIIa (Skagerrak) and Subarea VI (West of Scotland and Rockall) ................................................................................................................. 268 Sandeel in Division IIIa and Subarea IV .......................................................................................... 280 6.4.22.1 Sandeel in the Doggerbank area (SA 1)....................................................................... 287 ICES Advice 2013, Book 1 vii 6.4.22.2 6.4.22.3 6.4.22.4 6.4.22.5 6.4.22.6 6.4.22.7 6.4.23 6.4.24 6.4.25 6.4.26 6.4.27 6.4.28 6.4.29 6.4.30 6.4.31 6.4.32 6.4.33 6.4.34 6.4.35 viii Sandeel in the South Eastern North Sea (SA 2) ........................................................... 292 Sandeel in the Central Eastern North Sea (SA 3) ........................................................ 297 Sandeel in the Central Western North Sea (SA 4) ....................................................... 303 Sandeel in the Viking and Bergen Bank area (SA 5) ................................................... 307 Sandeel in Division IIIa East (Kattegat, SA6) ............................................................. 309 Sandeel in the Shetland area (SA 7) ............................................................................ 311 Northern shrimp (Pandalus borealis) in Divisions IIIa West and IVa East (Skagerrak and Norwegian Deeps)........................................................................................................ 313 Northern shrimp (Pandalus borealis) in Division IVa (Fladen Ground) ................................... 324 Rays and skates in Divisions and Subareas IIIa, IV, and VIId, e (Kattegat, Skagerrak, North Sea, and English Channel) ................................................................................................................. 325 6.4.25.1 Blonde ray (Raja brachyuran) in Divisions IVc and VIId, e (Southern North Sea and English Channel) ......................................................................................................... 327 6.4.25.2 Thornback ray (Raja clavata) in Subarea IV and in Divisions IIIa and VIId, e (North Sea, Skagerrak, Kattegat, and English Channel) ......................................................... 328 6.4.25.3 Small-eyed ray (Raja microocellata) in Divisions VIId, e (English Channel) ............ 329 6.4.25.4 Spotted ray (Raja montagui) in Subarea IV and in Divisions IIIa and VIId (North Sea, Skagerrak, Kattegat, and eastern English Channel) ..................................................... 330 6.4.25.5 Undulate ray (Raja undulata) in Divisions VIId, e (English Channel) ........................ 331 6.4.25.6 Cuckoo ray (Leucoraja naevus) in Subarea IV and in Divisions IIIa and VIId (North Sea, Skagerrak, Kattegat, and eastern English Channel) ............................................. 332 6.4.25.7 Common skate (Dipturus batis) complex (Dipturus cf. flossada and Dipturus cf. intermedia) in Subarea IV and in Divisions IIIa and VIId (North Sea, Skagerrak, Kattegat, and eastern English Channel) ....................................................................... 333 6.4.25.8 Starry ray (Amblyraja radiata) in Subarea IV and in Divisions IIIa and VIId (North Sea Skagerrak, Kattegat, and eastern English Channel) ...................................................... 334 6.4.25.9 Other ray and skate species in Subarea IV and in Divisions IIIa and VIId (North Sea, Skagerrak, Kattegat, and eastern English Channel) .................................................... 335 Sole in Division IIIa and Subdivisions 22–24 (Skagerrak, Kattegat, and the Belts) .................. 336 Sole in Subarea IV (North Sea) .................................................................................................. 346 Sole in Division VIId (Eastern Channel) ................................................................................... 361 Sprat in Division IIIa (Skagerrak – Kattegat)............................................................................. 371 Sprat in the Subarea IV (North Sea)........................................................................................... 377 Turbot in Division IIIa ............................................................................................................... 387 Turbot in Subarea IV (North Sea) .............................................................................................. 393 Whiting in Division IIIa (Skagerrak – Kattegat) ........................................................................ 395 Whiting in Subarea IV (North Sea) and Division VIId (Eastern Channel) ................................ 401 Witch in Subarea IV, and Divisions IIIa and VIId ..................................................................... 416 ICES Advice 2013, Book 1 BOOK 7 Section 7 Page BAY OF BISCAY AND ATLANTIC IBERIAN WATERS ................................................................................. 1 7.1 Ecosystem Overview ...................................................................................................................................... 1 7.2 The status of fish stocks and fisheries ............................................................................................................ 1 7.3 Ecosystem Assessments and Advice .............................................................................................................. 7 7.3.1 Multispecies advice ........................................................................................................................ 7 7.3.2 Mixed fisheries advice for the Bay of Biscay and Atlantic Iberian Waters ................................... 7 7.3.3 Assessment and advice regarding protection of biota and habitats ............................................... 7 7.3.4 Assessments and advice regarding fisheries .................................................................................. 7 7.3.5 Special requests ............................................................................................................................. 8 7.3.5.1 EU request to evaluate the management plan for Iberian sardine ..................................... 8 7.3.5.2 EU request for the evaluation of the harvest control rule for sole in the Bay of Biscay ................................................................................................................... 16 7.4 Fish stock advice .......................................................................................................................................... 21 7.4.1 Anchovy in Subarea VIII (Bay of Biscay) .................................................................................. 21 7.4.2 Anchovy in Division IXa (West of Galicia, Portuguese coast and Golf of Cadiz) ..................... 30 7.4.3 Black-bellied anglerfish (Lophius budegassa) in Divisions VIIIc and IXa (Atlantic Iberian waters) 37 7.4.4 White anglerfish (Lophius piscatorius) in Divisions VIIIc and IXa (Atlantic Iberian waters).... 44 7.4.5 Blue Jack mackerel (Trachurus picturatus) in Subdivision Xa2 (Azores) .................................. 52 7.4.6 Grey gurnard in Subarea VIII and Division IXa (Bay of Biscay and Atlantic Iberian waters) ... 54 7.4.7 Hake in Division VIIIc and IXa (Southern stock) ....................................................................... 55 7.4.8 Horse mackerel (Trachurus trachurus) in Division IXa (Southern stock) .................................. 67 7.4.9 Four spot megrim (Lepidorhombus boscii) in Divisions VIIIc and IXa (Bay of Biscay and Atlantic Iberian waters) ............................................................................................................................. 73 7.4.10 Lesser-spotted dogfish (Scyliorhinus canicula) in VIII and IX ................................................... 81 7.4.11 Lesser-spotted dogfish (Scyliorhinus canicula) in VIIIc and IXa ............................................... 82 7.4.12 Megrim (Lepidorhombus whiffiagonis) in Divisions VIIIc and IXa (Bay of Biscay and Atlantic Iberian waters) ............................................................................................................................. 83 7.4.13 Nephrops in Divisions VIIIa,b (Bay of Biscay, FU 23–24) ........................................................ 90 7.4.14 Nephrops in Division VIIIc (North Galicia and Cantabrian Sea, FU 25 and 31) ........................ 92 7.4.14.1 Nephrops in North Galicia (FU 25) .............................................................................. 95 7.4.14.2 Nephrops in the Cantabrian Sea (FU 31) ....................................................................... 97 7.4.15 Nephrops in Divisions IXa (West of Galicia, Portuguese coast and Golf of Cadiz). .................. 99 7.4.15.1 Nephrops in West Galicia and North Portugal (FU 26–27) ........................................ 103 7.4.15.2 Nephrops in South-West and South Portugal (FU 28–29) .......................................... 104 7.4.15.3 Nephrops in Gulf of Cadiz (FU 30) ............................................................................ 106 7.4.16 Plaice in Subarea VIII and Division IXa (Bay of Biscay and Atlantic Iberian waters) ............. 107 7.4.17 Pollack in Subarea VIII and Division IXa (Bay of Biscay and Atlantic Iberian waters) .......... 109 7.4.18 Rays and skates in Subareas VIII and IX (Bay of Biscay and Atlantic Iberian waters) ............ 111 7.4.18.1 Thornback ray (Raja clavata) in Subarea VIII (Bay of Biscay and Cantabrian Sea) .... 113 7.4.18.2 Cuckoo ray (Leucoraja naevus) in Subarea VIII (Bay of Biscay and Cantabrian Sea) .. 114 7.4.18.3 Spotted ray (Raja montagui) in Subarea VIII (Bay of Biscay and Cantabrian Sea) ....... 115 7.4.18.4 Spotted ray (Raja montagui) in Division IXa (west of Galicia, Portugal, and Gulf of Cadiz)................................................................................................................................. 116 7.4.18.5 Cuckoo ray (Leucoraja naevus) in Division IXa (west of Galicia, Portugal, and Gulf of Cadiz)................................................................................................................................. 117 7.4.18.6 Thornback ray (Raja clavata) in Division IXa (west of Galicia, Portugal, and Gulf of Cadiz)................................................................................................................................. 118 7.4.18.7 Blonde ray (Raja brachyuran) in Division IXa (west of Galicia, Portugal, and Gulf of Cadiz)................................................................................................................................ 119 7.4.18.8 Common skate (Dipturus batis) complex (flapper skate (Dipturus cf. flossada) and blue skate (Dipturus cf. intermedia)) in Subarea VIII and Division IXa (Bay of Biscay and Atlantic Iberian waters) .............................................................................................. 120 7.4.18.9 Other skates and rays in Subarea VIII and Division IXa (Bay of Biscay and Atlantic Iberian waters) ................................................................................................ 121 7.4.19 7.4.20 7.4.21 Sardine in Division VIIIc and IXa (Bay of Biscay and Atlantic Iberian waters) ...................... 122 Sardine in Divisions VIIIa,b,d and Subarea VII ........................................................................ 131 Sole in Divisions VIIIa,b (Bay of Biscay) ................................................................................. 137 ICES Advice 2013, Book 1 ix 7.4.22 7.4.23 7.4.24 7.4.25 7.4.26 x Sole in Divisions VIIIc and IXa (Atlantic Iberian waters) ........................................................ 147 Whiting in Subarea VIII and Division IXa (Bay of Biscay and Atlantic Iberian waters).......... 149 No advice................................................................................................................................... 151 European sea bass in Divisions VIIIa,b (Bay of Biscay) ........................................................... 151 European sea bass in Divisions VIIIc and IXa (Atlantic Iberian waters) .................................. 156 ICES Advice 2013, Book 1 BOOK 8 Section 8 Page THE BALTIC SEA 8.1 8.2 8.3 ......................................................................................................................................... 3 Ecosystem Overview ...................................................................................................................................... 3 The status of fish stocks and fisheries ............................................................................................................ 3 Ecosystem Assessments and Advice .............................................................................................................. 8 8.3.1 Assessments and advice regarding protection of biota and habitats............................................... 8 8.3.2 Assessments and advice regarding fisheries................................................................................... 8 8.3.3 Multispecies considerations for the central Baltic stocks: cod in Subdivisions 25–32, herring in Subdivisions 25–29 and 32, and sprat in Subdivisions 22–32 ....................................................... 9 8.3.4 Special Requests .......................................................................................................................... 15 8.3.4.1 Review of HELCOM draft Red List assessment of cod (Gadus morhua) ..................... 15 8.4 Fish stock advice ....................................................................................................................................... 19 8.4.1 8.4.2 8.4.3 8.4.4 8.4.5 8.4.6 8.4.7 8.4.8 8.4.9 8.4.10 8.4.11 8.4.12 8.4.13 8.4.14 8.4.15 8.4.16 8.4.17 Brill in Subdivisions 22–32 (Baltic Sea) ...................................................................................... 19 Cod in Subdivisions 22–24 (Western Baltic Sea) ........................................................................ 25 Cod in Subdivisions 25–32(Eastern Baltic Sea) ........................................................................... 37 Dab in Subdivisions 22–32 (Baltic Sea)....................................................................................... 48 Flounder in Subdivisions 22–32 (Baltic Sea) ............................................................................... 55 Herring in Division IIIa and Subdivisions 22–24 (Western Baltic spring spawners) .................. 68 Herring in Subdivisions 25–29 and 32 (excluding Gulf of Riga herring) .................................... 69 Herring in Subdivision 28.1 (Gulf of Riga) ................................................................................. 79 Herring in Subdivision 30 (Bothnian Sea) ................................................................................... 88 Herring in Subdivision 31 (Bothnian Bay)................................................................................... 95 Plaice in Subdivisions 21–23 (Kattegat, Belts and Sound) ........................................................ 103 Plaice in Subdivisions 24–32 (Baltic Sea) ................................................................................. 110 Salmon in Subdivisions 22–31 (Main Basin and Gulf of Bothnia) ............................................ 118 Salmon in Subdivision 32 (Gulf of Finland) .............................................................................. 142 Sprat in Subdivisions 22–32 (Baltic Sea) ................................................................................... 154 Sea Trout in Subdivisions 22– 32 (Baltic Sea)........................................................................... 166 Turbot in Subdivisions 22–32 (Baltic Sea) ................................................................................ 168 ICES Advice 2013, Book 1 xi BOOK 9 Section 9 WIDELY DISTRIBUTED AND MIGRATORY STOCKS .................................................................................. 1 9.1 Ecosystem overview ....................................................................................................................................... 1 9.2 The status of fish stocks and fisheries ............................................................................................................ 1 9.3 Assessments and advice ................................................................................................................................. 5 9.3.1 Assessments and advice regarding protection of biota and habitats............................................... 5 9.3.2 Assessments and advice regarding fisheries................................................................................... 5 9.3.3 Special requests .............................................................................................................................. 6 9.3.3.1 NEAFC request to ICES to evaluate the Harvest Control Rule element of the long-term management plan for blue whiting .................................................................................. 6 9.3.3.2 NEAFC request to ICES to evaluate possible modifications of the long-term management arrangement for the Norwegian Spring-Spawning herring stock ................................... 20 9.3.3.3 EU request on Technical Evaluation of Eel Management Plan Progress ....................... 34 9.3.3.4 EC request to ICES to evaluate possible modifications of the long-term management arrangement for the Western horse mackerel stock ....................................................... 50 9.3.3.5 NEAFC request on effects on assessments of historical unaccounted landings for mackerel and the utility of new and existing surveys..................................................... 57 9.3.3.6 EC request to ICES to evaluate the proposed long-term management plan for boarfish and possible in-year revision of the TAC for 2013 ........................................................ 59 9.3.3.7 NEAFC request to ICES to evaluate the extra harvest control rule options for the longterm management plan for blue whiting......................................................................... 61 9.4 xii Page Fish stock advice ....................................................................................................................................... 69 9.4.1 Alfonsinos/Golden eye perch (Beryx spp.) in the Northeast Atlantic ......................................... 69 9.4.2 Angel shark (Squatina squatina) in the North East Atlantic ........................................................ 70 9.4.3 Basking shark (Cetorhinus maximus) in the Northeast Atlantic .................................................. 71 9.4.4 Black scabbard fish (Aphanopus carbo) in the Northeast Atlantic .............................................. 72 9.4.5 Blue whiting in Subareas I-IX, XII and XIV (Combined stock) .................................................. 73 9.4.6 Boarfish in the Northeast Atlantic ................................................................................................ 87 9.4.7 European Eel ................................................................................................................................ 98 9.4.8 Greater forkbeard (Phycis blennoides) in the Northeast Atlantic ............................................... 105 9.4.9 Greater Silver Smelt (Argentina silus) in the Northeast Atlantic ............................................... 106 9.4.10 Hake in Division IIIa, Subareas IV, VI and VII and Divisions VIIIa,b,d (Northern stock) ....... 107 9.4.11 Herring in Subareas I, II, V and Divisions IVa and XIVa (Norwegian spring-spawning herring) …………………………………………………….... 118 9.4.12 Horse mackerel (Trachurus trachurus) in Divisions IIa, IVa, Vb, VIa, VIIa-c, e-k, and Subarea ... VIII (Western stock) .................................................................................................................. 130 9.4.13 Kitefin shark (Dalatias licha) in the Northeast Atlantic ............................................................ 149 9.4.14 Leafscale gulper shark (Centrophorus squamosus) in the Northeast Atlantic ........................... 150 9.4.15 Ling (Molva molva) in the Northeast Atlantic ........................................................................... 151 9.4.16 Blue ling (Molva dypterygia) in the Northeast Atlantic ............................................................. 152 9.4.17 Mackerel in the Northeast Atlantic (combined Southern, Western and North Sea spawning components) ............................................................................................................................... 153 9.4.18 Orange roughy (Hoplostethus atlanticus) in the Northeast Atlantic .......................................... 171 9.4.19 Porbeagle (Lamna nasus) in the Northeast Atlantic ................................................................... 172 9.4.20 Portuguese dogfish (Centroscymnus coelolepis) and leafscale gulper shark (Centrophorus squamosus) in the Northeast Atlantic ................................................................ 173 9.4.21 Rays and skates (mainly thornback ray) in the Azores and Mid-Atlantic Ridge (ICES Divisions X, XII, XIV).................................................................................................... 174 9.4.22 Red (=blackspot) seabream (Pagellus bogaraveo) in the Northeast Atlandic............................ 175 9.4.23 Roundnose grenadier (Coryphaenoides rupenstris) in the Northeast Atlantic ........................... 176 9.4.24 Smooth-hounds (Mustelus spp.) in the Northeast Atlantic ......................................................... 178 9.4.25 Red gurnard in the Northeast Atlantic ........................................................................................ 179 9.4.26 Spurdog (Squalus acanthias) in the Northeast Atlantic ............................................................. 180 9.4.27 Striped red mullet in the Northeast Atlantic ............................................................................... 181 9.4.28 Tope,(Galeorhinus galeus) in the Northeast Atlantic ................................................................ 182 9.4.29 Tusk (Brosme brosme) in the Northeast Atlantic ....................................................................... 183 ICES Advice 2013, Book 1 BOOK 10 Section Page 10 NORTH ATLANTIC SALMON STOCKS ............................................................................................................ 1 10.1 Introduction 10.1.1 10.1.2 10.1.3 10.1.4 10.1.5 Main Tasks ..................................................................................................................................... 1 Management framework for salmon in the North Atlantic ............................................................ 2 Management objectives .................................................................................................................. 3 Reference points and application of precaution ............................................................................. 3 Catch of North Atlantic Salmon ..................................................................................................... 4 10.1.5.1 10.1.5.2 10.1.5.3 10.1.5.4 10.1.6 ......................................................................................................................................... 1 Nominal catch of salmon ................................................................................................. 4 Unreported catches .......................................................................................................... 5 Catch-and-release ............................................................................................................ 5 Farming and sea ranching of Atlantic salmon ................................................................. 6 NASCO has asked ICES to report on significant, new or emerging threats to, or opportunities for, salmon conservation and management ........................................................................................... 6 10.1.6.1 10.1.6.2 10.1.6.3 10.1.6.4 10.1.6.5 Dam Impact Analysis Model for Atlantic Salmon in the Penobscot River, Maine ......... 6 Marine influences on North American Atlantic salmon populations ............................... 6 West Greenland foraging ecology and implications for survival..................................... 7 Tracking and acoustic tagging studies in Greenland and Canada .................................... 7 The impact of artificial night light on Atlantic salmon fry dispersal and the onset of smolt migration ................................................................................................ 10 10.1.6.6 Stock Identification of Salmon caught in the Faroes Fishery ........................................ 10 10.1.6.7 Update on EU project ECOKNOWS ............................................................................. 10 10.1.6.8 Diseases and parasites.................................................................................................... 12 10.1.6.9 Changing biological characteristics of salmon .............................................................. 13 10.1.6.10 New initiatives in relation to management of mixed-stock coastal fisheries in northern Norway ............................................................................................................ 14 10.1.7 10.1.8 10.1.9 10.1.10 10.1.11 10.1.12 10.1.13 NASCO has asked ICES to provide a review of examples of successes and failures in wild salmon restoration and rehabilitation and develop a classification of activities which could be recommended under various conditions or threats to the persistence of populations................... 14 NASCO has asked ICES to advise on the potential threats to Atlantic salmon from exotic salmonids including rainbow trout and brown trout where appropriate ....................................... 15 Reports from ICES Expert Group Reports relevant to North Atlantic salmon ............................ 19 NASCO has asked ICES to provide a compilation of tag releases by country in 2012 ............... 20 NASCO has asked ICES to further develop a risk-based framework for the provision of catch advice for the Faroese salmon fishery reporting on the implications of selecting different numbers of management units...................................................................................................... 20 NASCO has asked ICES to update the Framework of Indicators to identify any significant change in previously provided multi-annual management advice ............................................... 24 NASCO has requested ICES to identify relevant data deficiencies, monitoring needs, and research requirements................................................................................................................... 24 Annex 10.1 Glossary of acronyms ................................................................................................................................... 48 Annex 10.2 References cited ....................................................................................................................................... 52 10.2 10.3 10.4 Stock Summary - Atlantic Salmon from the Northeast Atlantic .................................................................. 58 10.2.1 Supporting information ................................................................................................................ 61 Advice for Atlantic Salmon from North America ........................................................................................ 78 10.3.1 Supporting information ................................................................................................................ 80 Advice for Atlantic Salmon at West Greenland ........................................................................................... 97 10.4.1 Supporting information ................................................................................................................ 99 ICES Advice 2013, Book 1 xiii BOOK 11 Section Page 11 TECHNICAL SERVICES ....................................................................................................................................... 1 xiv 11.1 About ICES Technical services ...................................................................................................................... 1 11.2 Answers to requests ........................................................................................................................................ 2 11.2.1 EC ......................................................................................................................................... 2 11.2.1.1 Opinion on modification to the list of deep-sea sharks .................................................... 2 11.2.1.2 Opinion on the outcome of an in Year revision for Northern Hake ................................. 4 11.2.1.3 Response on special management measures for skates and rays ..................................... 5 11.2.2 HELCOM ..................................................................................................................................... 22 11.2.2.1 ICES-organized external peer review of the BALTFIMPA Generic Tool .................... 22 11.2.2.2 ICES-organized external peer review of the BALTFIMPA Generic Tool - Stage Two 27 ICES Advice 2013, Book 1 Preface This report contains the 2013 ICES advice to the advice recipients regarding marine management issues. The report is produced by the Advisory Committee (ACOM) providing advice on behalf of the Council. The Advisory Committee include one designated scientist from each of the ICES member countries and the committee has an independently elected chair and four vice-chairs. The chair of the Science Committee is member ex-officio. ICES has invited the competent authorities, which are requesting advice from ICES, and stakeholder groups to be present at Advisory Committee meetings (whether physical or virtual) in observer capacity. The advice is developed though a process which is setup to ensure that it is based on the best available science, that it is independent, peer reviewed and that the process is transparent and considered legitimate by authorities and stakeholders. The process starts out with expert groups who compiles the relevant data, makes a scientific analysis and provides a report presenting data, methods and outcomes. These reports are then peer reviewed by designated groups. The review groups are composed of scientists who are not members of the expert group under review and who normally do not originate from countries with a strong interest in the stocks concerned. Some review groups include invited reviewers who are not otherwise involved in the ICES assessment at all and in some cases recruited from institutions not normally involved with ICES advisory work, including research institutes from outside the North East Atlantic area. The Expert Working Group chairs assist the review groups. In the case of analysis which is updated with new annual data, but otherwise following an agreed methodology (typically annual fish stock assessments), this peer review of methods takes place in benchmark workshops which produce a description of the agreed methodology and the data to use. This description is then applied for subsequent years by the expert group, where members of the expert group annually will make an audit of each others work, that methodology described in the benchmark has been properly implemented. The advisory text is developed in advice drafting groups guided by a chair appointed by ACOM. ICES Delegates and ACOM members appoint participants in the advice drafting groups. ACOM then approves the advice. Structure of the report Book 1 explains the conceptual and institutional framework for the assessments and advice. It contains a general introduction to the ICES advice, and includes general and non-regional advice. Books 2 - 8 are regional reports covering the following marine ecoregions (Figure 1): • Book 2: Iceland and East Greenland • Book 3: The Barents Sea and the Norwegian Sea • Book 4: The Faroe Plateau Ecosystem • Book 5: Celtic Sea and West of Scotland • Book 6: North Sea • Book 7: Bay of Biscay and Iberian Seas • Book 8: The Baltic Sea Book 9 is a separate chapter for widely distributed and migratory stocks and Book 10 provides information on the North Atlantic salmon. Book 11 documents the technical services that ICES has provided to its advice recipients throughout the year. ICES Advice 2013, Book 1 xv ACOM members and alternates 2013 ACOM Chair ACOM Vice-chairs SCICOM Chair Belgium Member: Alternates: xvi Canada Member: Alternate: Denmark Member: Alternates: Estonia Member: Alternate: Finland Member: Alternates: France Member: Alternates: Germany Member: Alternates: Iceland Member: Alternates: Jean Jacques Maguire (Canada) Carmen Fernandez (Spain) Han Lindeboom (NL) John SImmonds (UK) Mark Tasker (UK) Manuel Barange (UK) Steven Degraer (Jan–Jul) Els Torreele (Aug–Dec) Kris Cooreman Daan Delbare Kris Hostens Kelle Moreau Hans Polet Els Torreele (Jan–Jul) Ghislain Chouinard Michel Gilbert Joanne Morgan Bernard Sainte-Marie Morten Vinther Jesper Boje Fritz W. Köster Henrik Mosegaard Toomas Saat Henn Ojaveer Georg Martin Tiit Raid Matti Salminen Eero Aro Ari Leskelä Atso Romakkaniemi Alain Biseau Michel Bertignac Mathieu Doray Philipp Hess Joel Knoery Pascal Lorance Paul Marchal Christopher Zimmermann Joachim Gröger Thomas Lang Anne Sell Christian von Dorrien Björn Steinarsson Astthor Gislason Einar Hjörleifsson Sólveig Ólafsdottir ICES Advice 2013, Book 1 Ireland Member: Alternates: Latvia Member: Alternates: Lithuania Netherlands Member: Member: Alternate: Norway Member: Alternates: Poland Member: Alternates: Portugal Member: Alternates: Russia Member: Alternate: Spain Member: Alternates: ICES Advice 2013, Book 1 Maurice Clarke Leonie Dransfeld Ciaran Kelly Colm Lordan Evin McGovern Maris Plikshs (Jan–May) Didzis Ustups (June–Dec) Juris Aigars Janis Birzaks Anda Ikauniece Georgs Kornilovs Didzis Ustups (Jan–May) Tomas Zolubas Tammo Bult (Dec–Jun) Floor Quirijns (Jul–Dec) Lisette Enserink Martin Pastoors (Jul–Dec) Floor Quirijns (Dec–Jun) Reidar Toresen (Jan–Jul) Harald Gjøsæter (Aug–Dec) Bjørn Ådlandsvik (Jan–Jul) Asgeir Aglen (Jan–Jul) Harald Gjøsæter (Jan–Jul) Geir Huse Jarle Klungsøyr Cecilie Kvamme (Jun–Dec) Sigbjørn Mehl (Aug–Dec) Hein Rune Skjoldal Jan Erik Stiansen (Aug–Dec) Einar Svendsen (Jan–Jul) Else Torstensen Jan Horbowy Henryka Dabrowska Piotr Margonski Fátima Borges Ivone Figueiredo Alberto Murta Yuri Efimov Yuri A. Kovalev Dmitry A. Vasilyev Javier Pereiro Pablo Abaunza Pablo Carrera Jose Fumega Elena Guijarro (Sep–Dec) Santiago Lens Francisco Velasco xvii xviii Sweden Member: Alternate: United Kingdom Member: Alternates: USA Member: Alternate: Faroe Islands Greenland Observer: Observer: Massimiliano Cardinale Michele Casini Joachim Hjelm Carl O’Brien Nick Bailey Walter Crozier Alejandro Gallego Emma Hatfield Coby Needle Bill Turrell Gary Shepherd Larry Alade Thomas Noji Jakúp Reinert Jesper Boje ICES Advice 2013, Book 1 1 Introduction, overview and special requests 1.1 About ICES ICES was established in 1902 as an intergovernmental organisation. The ICES Convention from 1964 outlines the fundamental purposes of ICES, which are: to promote and encourage research and investigations for the study of the sea particularly related to the living resources thereof; to draw up programmes required for this purpose and to organise, in agreement with the Contracting Parties, such research and investigations as may appear necessary; to publish or otherwise disseminate the results of research and investigations carried out under its auspices or to encourage the publication thereof. Under the Convention, ICES is concerned with the Atlantic Ocean and adjacent seas, primarily the North Atlantic. Today, ICES provides the scientific underpinning for most of the regulatory commissions concerned with fisheries and the environment in the Northeast Atlantic and the Baltic Sea. ICES has grown from a small body of like-minded researchers to a an organisation involving about 4000 scientists, with 20 Member Countries, including all countries with a coastline to the North Atlantic including the Baltic from the USA in the west to the Strait of Gibraltar in the East. There are also several Countries from outside this area and non-governmental organisation which are observers to ICES. Today the member states and the network of scientists use ICES as a platform to plan and coordinate marine science, to present and discuss outcomes of marine research, to plan and implement the collection of marine data including planning of surveys with research vessels, to store and exchange marine data, as an outlet for publication of marine research results and as an advisory body to provide science-based advice to public authorities with competence regarding marine management such as environmental policy or fisheries management. ICES fulfils these functions through an Annual Science Conference, two committees governing the science and the advisory work (the Science and Advisory Committees), more than 150 working and study groups, several symposia annually, and a wide range of publications. There is a Secretariat located in Copenhagen, which currently has about 50 full-time professional and support staff. It is the scientists who participate in ICES activities who generate ICES products. The main products are scientific information based on research conducted in the Member Countries and scientific advice containing information provided in a format that can be used by policy-makers. The responsibility for the production of scientific advice rests with the Advisory Committee. ICES is requested to provide advice on a range of issues relating to marine policies and management. Requests for advice may be submitted by public authorities with competence for marine management including: • Governments of ICES’ member countries, • European Commission (EC) • Helsinki Commission (HELCOM), • North Atlantic Salmon Commission (NASCO), • North East Atlantic Fisheries Commission (NEAFC) • OSPAR Commission (OSPAR) ICES may also on its own initiative draw the attention of competent authorities to marine matters which may require policy and management attention. ICES Advice 2013, Book 1 1 ICES works with its advice recipients to find the most effective way of delivering integrated advice. The immediate delivery of advice is through the ICES internet site and the annual summary of advice is provided in this document that includes all aspects of ICES’ advice. 2 ICES Advice 2013, Book 1 1.2 Advice basis June 2013 1.2.1 General context of ICES advice ICES advises competent authorities on marine policy and management issues related to the impacts of human activities on marine ecosystems and the sustainable use of living marine resources. Overarching international agreements on exploitation of living marine resources An important part of ICES advice regards the management of the exploitation of living marine resources. The context for this part of ICES advice is set by several international agreements and policies: • United Nations Convention on the Law of the Sea (UN, 1982 (known as UNCLOS)), which includes a call for a maximum sustainable yield (MSY) approach to managing fisheries; • United Nations Conference on Environment and Development (UN, 1992a (known as UNCED)), including Chapter 17 of Agenda 21 which highlights a precautionary approach; • United Nations Straddling Fish Stocks Agreement of 1995 (UN, 1995 (known as the UN Fish Stocks Agreement or UNFSA)) and the FAO Code of Conduct for Responsible Fisheries (FAO, 1995), both of which call for a precautionary approach; • Convention on Biological Diversity (UN, 1992b (known as CBD)), which calls for conservation of biological diversity through an ecosystem approach; • Johannesburg Declaration of the World Summit on Sustainable Development (UN, 2002 (known as WSSD)), which calls for an ecosystem approach and rebuilding fisheries to maximum sustainable yield. In addition, ICES advice responds to the policy and legal needs of ICES member countries and multinational and intergovernmental organizations that use the advice as the scientific basis to manage human activities that affect, and are affected by, marine ecosystems. Some applicable policy and legal instruments are: • The Common Fisheries Policy of the European Union (EC, 2002) • Communication from the European Commission on Implementing Sustainability in EU Fisheries through Maximum Sustainable Yield (EC, 2006) • The Marine Strategy Framework Directive (EC, 2008) • Norwegian Marine Resources Act (Lovdata, 2008 (Lov om forvaltning av viltlevande marine ressursar)), • Russian Federal Law on Fisheries and conservation of biological resources in the waters. N 166-P3 20/12/2004 (Anon., 2004) • Icelandic Fisheries Management Act (No. 38, 15 May 1990) (Anon., 1990) • Faroe Islands Fisheries Management Act (Løgtingslóg nr. 28 um vinnuligan fiskiskap frá 10. mars 1994) (Anon., 1994) ICES provides advice in the context of these agreements. Impacts of human activities on marine ecosystems Almost all ICES member countries have policies that address the impacts of human activities on marine ecosystems which have been developed under the above international agreements. These policies may explicitly be framed as an implementation of an ecosystem approach. An important example is the Marine Strategy Framework Directive (MSFD) of the European Union (EC, 2008), which is a comprehensive framework for achieving good environmental status (GES) for European marine ecosystems. The Directive calls for scientifically-based indicators and standards for eleven descriptors of GES such as Biodiversity, Non-indigenous Species, Commercially Exploited Fish and Shellfish Stocks, Foodwebs, and Sea-floor Integrity. The Regional Seas conventions have a role in ensuring the cohesion of assessments within their regions. Both OSPAR and HELCOM have established specific coordinating platforms for the regional implementation of the MSFD, striving ICES Advice 2013, Book 1 3 for harmonized national marine strategies to achieve good environmental status and implementing their overall agreed commitment to an ecosystem approach. The ICES scientific community and ICES advisory services have played a key role in providing scientific guidance to define GES indicators and standards. The process of developing these indicators and standards at the European level is ongoing and the process is now being continued by revising current monitoring activities and developing programmes of measures. The MSFD is an important challenge for the scientific community, and ICES welcomes the MSFD as an opportunity to apply an ecosystem approach. Marine spatial planning is envisioned as a key mechanism in achieving GES. The idea is to integrate planning and management actions across human activities (e.g. fisheries, renewable and non-renewable energy development, mineral extraction, transportation, tourism, recreation, etc.) to take into account the cumulative impact of all of these activities on ecosystems. This will require more spatially resolved data on more types of activities, and a better understanding of how these activities impact ecosystems. It will also require integrated ecosystem monitoring systems where ICES has special experience to offer in particular on the fisheries surveys side. A draft EU Directive on establishing a framework for maritime spatial planning and integrated costal management has been released in March 2013. An ecosystem approach to marine environmental policy, a precautionary approach and an MSY approach regarding living marine resources are prominent themes of the agreements and policies that set the context for ICES advice. A compilation of acronyms and terminology used in the ICES advice is available at http://www.ices.dk/advice/icesadvice.asp as acronyms_and_terminology.pdf. 1.2.1.1 An ecosystem approach to management of the marine environment An ecosystem approach is intended to contribute to sustainable development. Sustainable development is defined in the Brundtland Report (WCED, 1987) as development that “meets the needs of the present without compromising the ability of future generations to meet their own needs.” An ecosystem approach has been defined in various ways but mainly emphasizes a management regime that maintains the health of the ecosystem alongside appropriate human uses of the environment, for the benefit of current and future generations. For example, the 1992 UN Convention on Biological Diversity (UN, 1992b) defines an ecosystem approach as “ecosystem and natural habitats management” to “meet human requirements to use natural resources, whilst maintaining the biological richness and ecological processes necessary to sustain the composition, structure and function of the habitats or ecosystems concerned.” The Reykjavik Declaration (FAO, 2001) forms the basis for using an ecosystem approach in the management of the marine environment: “… in an effort to reinforce responsible and sustainable fisheries in the marine ecosystem, we will individually and collectively work on incorporating ecosystem considerations into that management to that aim.” The World Summit on Sustainable Development (UN, 2002) indicated that States should: “(30.d) Encourage the application by 2010 of the ecosystem approach, noting the Reykjavik Declaration on Responsible Fisheries in the Marine Ecosystem 1 and decision V/6 of the Conference of Parties to the Convention on Biological Diversity”. An ecosystem approach is expected to contribute to achieving long-term sustainability for the use of marine resources, including the fisheries sector. An ecosystem approach serves multiple objectives, involves strong stakeholder participation, and focuses on human behaviour as the central management dimension. ICES is in the process of regionalizing its advice and building the scientific foundation for regional ecosystem advice. The methods needed to allow ecosystem advice are being developed by working groups on integrated ecosystem assessments and will support summaries of ecosystem state and pressures documented in “ecosystem overviews”. These overviews will focus on ecosystem processes in order to enable ecosystem drivers to be incorporated into traditional fish stock assessments and to enable operational advice to be given regarding possible measures. This process is necessary for 1 “While it is necessary to take immediate action to address particularly urgent problems on the basis of the precautionary approach, it is important to advance the scientific basis for incorporating ecosystem considerations, building on existing and future available scientific knowledge.” Source: Reykjavik Declaration, appendix I, pg. 107. (FAO, 2001). 4 ICES Advice 2013, Book 1 ICES to provide robust, contextual and relevant advice on ecosystems. ICES will work closely together with the regional seas commissions (RSCs), European Environment Agency, and EU/DG Environment, to ensure that our efforts are supplementing on-going activities. ICES Advice 2013, Book 1 5 1.2.1.2 A precautionary approach in fisheries management A precautionary approach (PA) is described in the UN Fish Stocks Agreement (UN, 1995) as follows: “States shall be more cautious when information is uncertain, unreliable or inadequate. The absence of adequate scientific information shall not be used as a reason for postponing or failing to take conservation and management measures.” Annex 2 of the UNFSA contains guidelines for applying a precautionary approach within an MSY framework. To quote: “The fishing mortality rate which generates maximum sustainable yield should be regarded as a minimum standard for limit reference points. For stocks which are not overfished, fishery management strategies shall ensure that fishing mortality does not exceed that which corresponds to maximum sustainable yield, and that the biomass does not fall below a predefined threshold. For overfished stocks, the biomass which would produce maximum sustainable yield can serve as a rebuilding target.” In addition, the guidelines indicate: • Precautionary reference points should be used to guide management; • Target reference points are intended to achieve management objectives; • Precautionary reference points should take account of reproductive capacity, the resilience of each stock, and the characteristics of fisheries exploiting the stock, as well as other sources of mortality and major sources of uncertainty; • Management strategies shall seek to maintain stocks at, or restore stocks to, levels consistent with previously agreed precautionary reference points. Such reference points shall be used to trigger pre-agreed conservation and management action. Management strategies shall include measures which can be implemented when precautionary reference points are approached; • Fishery management strategies shall (a) ensure that the risk of exceeding limit reference points is very low, (b) initiate actions to facilitate stock recovery for stocks below precautionary reference points, and (c) ensure that target reference points are not exceeded on average; and • When information for determining reference points for a fishery is poor or absent, provisional reference points shall be set. Although some aspects of the guidelines are not entirely clear (e.g. the relationship between precautionary and limit reference points is unclear) or consistent (the fishing mortality rate to achieve MSY is referenced as both a target reference point and a limit reference point), it is most useful to recognize that MSY and a precautionary approach are complementary, and this is the spirit in which ICES applies these concepts. Populations need to be maintained within safe biological limits according to a precautionary approach to make MSY possible. However, within safe biological limits, an MSY approach is necessary to achieve MSY. Lack of scientific information should not be an excuse for postponing management action to maintain populations within safe biological limits and/or to delay implementing a strategy to attain MSY. In a sense, a precautionary approach is a risk-averse concept intended to avoid unproductive situations while an MSY approach is intended to make the best use of the ecosystem productivity. A precautionary approach (PA) is a necessary, but not a sufficient condition for MSY. The ICES precautionary approach (including the methods for estimating PA reference points) is described in more detail in the introduction of previous volumes of ICES advice (e.g. ICES, 2009b). 1.2.1.3 The maximum sustainable yield concept Maximum sustainable yield has been a widely accepted objective for fisheries management for many decades. The United Nations Convention on the Law of the Sea (UN, 1982) notes “…State(s) must set an allowable catch, based on scientific information, which is designed to maintain or restore species to levels supporting a maximum sustainable yield (MSY).” This policy was reaffirmed by WSSD (UN, 2002) which called on States to 6 ICES Advice 2013, Book 1 “Maintain or restore stocks to levels that can produce the maximum sustainable yield with the aim of achieving these goals for depleted stocks on an urgent basis and where possible not later than 2015”. Maximum sustainable yield is a broad conceptual objective aimed at achieving the highest possible yield over the long term (an infinitely long period of time). It is non-specific with respect to: (a) the biological unit to which it is applied; (b) the models used to provide scientific advice; and (c) the management methods used to achieve MSY. The MSY concept can be applied to an entire ecosystem, an entire fish community, or a single fish stock. The choice of the biological unit to which the MSY concept is applied influences both the sustainable yield that can be achieved and the associated management options. For reasons discussed later, implementation of the MSY concept by ICES will first be applied to individual fish stocks, but even on a single-stock basis MSY can only be reached in a healthy environment. In practice, MSY depends on: • The production of the unit, which describes the relation between productivity and the size of the unit (e.g., population biomass), which in turn depends on the growth rates, natural mortality rates, and reproductive rates of the members of the production unit; • Interactions of members within the production unit and interactions with other production units (intra- and interspecific interactions); • Environmental conditions (e.g., climate, environmental quality), which affect production, and intra- and interspecific interactions; and • Fishing practices and fishery selectivity that determine the size and age composition of the catch (both the landings and the discards). Many of the models (mathematical and conceptual) used to estimate MSY and associated parameters typically assume that factors not explicitly included in the models remain constant or vary around a long term mean. Thus, MSY estimates are generally conditional on current conditions and assumptions. This assumption is reasonable as long as the analysis does not attempt to project changes which are very different from the prevailing conditions, or from conditions which have been observed historically. Marine ecosystems are, however, dynamic and fish stocks will not only change in response to the fisheries directly targeting them but also to changes in fishing patterns and fishing pressures on their prey or their predators. ICES considers MSY estimates as applicable only in the short-term and they should be subject to regular re-estimation. This has implications for the further development of the MSY approach as discussed in the context of incorporation of ecosystem considerations in ICES advice below (Section 1.2.2.5). 1.2.1.4 ICES approach to fisheries advice: Maximum sustainable yield within a precautionary approach The ICES approach to fisheries advice integrates a precautionary approach, maximum sustainable yield, and an ecosystem approach into one advisory framework. The aim is, in accordance with the aggregate of international guidelines, to inform policies for high long-term yields while maintaining productive fish stocks within healthy marine ecosystems. ICES recognises that although the advice is based on stock objectives, the method is through the management of fisheries and changes in stock size are the result of both the changes in the fishery and the environment. ICES provides fisheries advice that is consistent with the broad international policy norms of the precautionary approach, MSY, and an ecosystem approach while also responding to the specific needs of the management bodies requesting advice. A precautionary approach has been recognised as an important basis for fisheries management in all the jurisdictions advised by ICES. ICES notes that in the past the fisheries for which it provides advice have generally not been managed with MSY as an objective. The current European Commission policy (EC, 2006) calls for EC fisheries to be managed according to MSY by 2015. Therefore, the nature of ICES fisheries advice is evolving. The evolution includes options for a transition process to attain full implementation of an MSY approach by 2015. Ecosystem limitations on fisheries have typically not yet been identified in management policies in the ICES area. However, as the EU MSFD is implemented, such limits will be recognized to achieve environmental objectives, especially regarding biodiversity, sea floor integrity, and food webs. Therefore, harvests may be further modified in consideration of potential fishery impacts on marine ecosystems beyond the impact on target fish stocks. ICES considers that a precautionary approach, an ecosystem approach and maximum sustainable yield, are not alternative strategies but are nested boundaries for the harvesting of living marine resources, where the outer boundary is defined by a precautionary approach to maintain fish stock productivity (Figure 1.2.1). Considerations of wider ecosystem impacts and interactions may further modify harvest strategies and not all fishing strategies within these precautionary limits will lead to the largest long-term yields as MSY is constrained by the prevailing ecosystem considerations. ICES Advice 2013, Book 1 7 MSY Ecosystem impact boundary PA exploitation boundary Figure 1.2.1 8 Limits to the exploitation of living marine resources that arise from a precautionary approach regarding single-stock exploitation, an ecosystem approach and an MSY approach, as an integrated framework of nested boundaries ICES Advice 2013, Book 1 1.2.2 The Technical basis of the ICES advice Fisheries management under the above philosophy may be carried out using a variety of approaches involving the specification and control of both input (numbers of vessels, days at sea) and outputs (catches, landings). Within the currently agreed European allocation scheme and regulatory framework the primary measure is the output or removals from the stock. Thus, ICES is typically requested to provide catch advice on a stock-by-stock basis, as fisheries on most of the stocks for which ICES provides advice are managed using stock-specific total allowable catches (TACs). In practice most fisheries in the ICES area are currently managed through constraints on landings. Those landings figures could be deduced from the catch advice based on the assumption that discards will remain a stable incremental factor (as observed in the recent past) additional to the landings. In many cases, other fishery management measures are used as well, such as technical regulations (e.g. closed areas, mesh sizes, days-at-sea limitations and minimum landing sizes). To support the European stock by stock management system, the ICES framework for fisheries advice needs to be applicable to individual stocks. This does not obviate the need to modify stock-specific advice to take account of technical interactions (e.g. bycatch in mixed-species fisheries) or of biological interactions (e.g. predator–prey relationship), but the structure for ICES fisheries advice remains the individual fish stock. 1.2.2.1 Maximum Yield and the Precautionary Approach Fisheries affect fish stocks through catches which can be expressed in a proportional way as the fishing mortality rate (F) applied to these stocks. Production of a fish stock is the sum of the population weight (biomass) augmented by the young each year (recruitment or incoming year class) growth in weight of existing year classes minus the loss of numbers of individuals from natural mortality. Production can be highly variable but, on average, it is related to stock size (often expressed as spawning-stock biomass or SSB), which in turn depends on F. That is, for each F, there is a long-term average production and an average stock size. The relationship between F, production, and stock size is called the production function. Surplus production at a given F is the catch that can be harvested without changing the average stock size in the long term. The peak of the production function is MSY, and the fishing mortality generating this peak is FMSY. Figure 1.2.2 gives a hypothetical production function versus F and Figure1.2.3 shows surplus production versus spawning-stock biomass. MSY Reference Points Yield SSB 200 400 B MSY 100 300 MSY 50 200 SSB Yield 150 100 F MSY 0 0 0 0.5 1 1.5 Fishing mortality Figure 1.2.2 Example of a yield (production) versus fishing mortality (F) for a hypothetical fishery. SSB: spawning-stock biomass. ICES Advice 2013, Book 1 9 Surplus Production Surplus Production 100 80 60 40 BMSY 20 0 0 200 400 600 800 SSB Figure 1.2.3 Example of surplus production versus spawning-stock biomass (SSB) for a hypothetical stock. The theory is that harvesting around 80 units when the stock is at BMSY would keep the stock in full reproductive state at an SSB capable of delivering MSY. On the stock at the carrying capacity (800 units of SSB) there is no surplus production, all the production is needed to support the high stock size. If a fishery begins on a stock at carrying capacity (i.e. the virgin state), the SSB is reduced and surplus production is generated. Fishing mortality (F) is the only variable in the production function that can be directly controlled by fisheries management. Fisheries management cannot control SSB, it can only influence it through F. SSB is also subject to natural variability that on a year-to-year basis can overwhelm the influence of F. MSY is a long-term average, and the maximum constant yield that could be taken sustainably would be lower than MSY. Fisheries that harvest variable yields in response to the natural variability in stock size will be on average nearer to the long term MSY. Due to the natural variability in SSB, beyond the influence of F, there may be situations where the spawning stock is so low that a significant risk exists that reproduction is impaired. A precautionary approach implies that fisheries management in such situations should be more cautious. For stocks where quantitative information is available, a reference point Blim may be identified as the stock size below which there may be reduced reproduction resulting in reduced recruitment. A precautionary safety margin incorporating the uncertainty in ICES stock estimates leads to a precautionary reference point Bpa, which is a biomass reference point designed to avoid reaching Blim. Therefore, when SSB is above Bpa the probability of impaired recruitment is expected to be low. For short-lived species, the biomass can fluctuate over the full range between years. A precautionary approach in this situation implies that a minimum stock size, Bescapement, should remain every year in the sea after fishing (Figure 1.2.4). 10 ICES Advice 2013, Book 1 Biomass Reference Points 900 800 MSY Btrigger or Bpa or MSY Bescapement Recruitment 700 600 500 400 300 200 100 0 0 100 Blim 200 300 BMSY 400 500 SSB Figure 1.2.4 Illustration of biomass-based biological reference points. Blim and Bpa are precautionary reference points related to the risk of impaired reproductive capacity, while MSY Bescapement often equal to Bpa is used in the advice framework for short-lived species. MSY Btrigger is the parameter in the ICES MSY framework which triggers advice on a reduced fishing mortality relative to FMSY. BMSY is the average biomass expected if the stock is exploited at FMSY. Diamonds show the variable recruitment verses SSB that have been observed over the years. Recruitment can be seen to be generally lower below Blim. The ICES framework for fisheries advice recognizes that the characteristics of fish stocks are different, and the information available on individual stocks also varies. The ICES approach uses both fishing mortality rates and biomass reference points. In general, FMSY should be lower than Fpa, (a precautionary buffer to avoid that true fishing mortality is at Flim the rate associated with long term stock decline and ultimately crash) and MSY Btrigger should be equal to or higher than Bpa. This is appropriate since a precautionary approach is a necessary boundary to ensure sustainability, but not sufficient, condition for achieving the maximum sustainable yield implied by the MSY framework. Although most of the stock for which ICES provides advice are considered in terms of exploitation using fishing mortality rates, for a very small number of stocks, such as Icelandic cod and saithe and some Nephrops stocks, ICES advises based on harvest rates (HR) a slightly different measure of exploitation. The HR is defined so that the recommended catch is a fraction of a reference biomass. The reference biomass can be based on total stock biomass, SSB, biomass above a minimum size or minimum age. The choice is tailored to the most suitable biomass for the stock concerned. In these cases the fishery may legitimately catch ages or sizes outside the reference range but the HR is still defined in terms of the selected reference. The HR can also be defined directly on the size or ages selected in the fishery, in this case the reference biomass is described as the fishable biomass. In stable fisheries with relatively stable recruitment F and HR can be related directly to one another. If recruitment is very variable or age or size selection changes in the fishery the relationship can change. Advising using a HR is most suitable for fisheries that exploit intermittent year-classes such as haddock or where age information is not available such as Nephrops. 1.2.2.2 ICES stock categories ICES recognise six main categories of stocks, and application of the ICES framework to each of these categories is discussed below. ICES has used the following biological and information based categorizations: ICES Advice 2013, Book 1 11 Category 1 – Stocks with quantitative assessments This type of stock can be considered in two sub categories a) stocks with several year classes contributing to the fishery that includes stocks with full analytical assessments and forecasts as well as stocks with quantitative assessments based on production models.; and b) short lived species stocks with quantitative assessments. These are the stocks that have short life cycles with catches dominated by single year classes. They are not considered data-limited and this category includes stocks with full analytical assessments and forecasts as well as stocks with quantitative assessments based on production models. Category 2 – stocks with analytical assessments and forecasts that are only treated qualitatively This category includes stocks with quantitative assessments and forecasts which for a variety of reasons are considered indicative of trends in fishing mortality, recruitment, and biomass. Category 3 – stocks for which survey-based assessments indicate trends This category includes stocks for which survey indices (or other indicators of stock size such as reliable fishery-dependant indices; e.g. lpue, cpue, and mean length in the catch) are available that provide reliable indications of trends in stock metrics such as total mortality, recruitment, and biomass. Category 4 – stocks for which only reliable catch data are available This category includes stocks for which a time-series of catch can be used to approximate MSY. Category 5 – Landings only stocks This category includes stocks for which only landings data are available. Category 6 – negligible landings stocks and stocks caught in minor amounts as bycatch This category includes stocks where landings are negligible in comparison to discards. It also includes stocks that are part of stock complexes and are primarily caught as bycatch species in other targeted fisheries. The development of indicators may be most appropriate for such stocks. 1.2.2.2.1a Long-lived stocks with population size estimates For stocks with population size estimates, ICES can calculate the catch that will achieve a desired fishing mortality rate. For most stocks with population size estimates, ICES can also forecast future stock size as a function of catch (i.e. for a range of catch options). In stocks naturally having several age groups, future stock size is not overly dependent on recruitment because many older animals exist in the population (unless the stock age composition has been truncated due to high fishing mortality). When population projections are too dependent on recruitment, projections are less reliable because recruitment can be variable and difficult to measure or predict. For long-lived stocks with population size estimates, ICES bases its approach on attaining a fishing mortality rate at, or below, FMSY. Annex 2 of the UN Fish Stocks Agreement (UNFSA; UN, 1995 see Section 1.2.1) states that “The fishing mortality rate which generates maximum sustainable yield should be regarded as a minimum standard for limit reference points. For stocks which are not overfished, fishery management strategies shall ensure that fishing mortality does not exceed that which corresponds to maximum sustainable yield, and that the biomass does not fall below a predefined threshold.” The World Summit for Sustainable Development (WSSD, Johannesburg; UN, 2002) states that “To achieve sustainable fisheries, the following actions are required at all levels: (a) Maintain or restore stocks to levels that can produce the maximum sustainable yield with the aim of achieving these goals for depleted stocks on an urgent basis and where possible not later than 2015.” The first statement refers to FMSY as an upper limit to fishing mortality. From a starting point of excessive exploitation (until recently this was the case for many stocks in the ICES area), the latter statement can be considered as an intermediate step towards fulfilling the UNFSA requirements as it establishes an intermediate target for fishing mortality at FMSY, so that stocks are restored by 2015. Competent authorities advised by ICES have based their implementation on the WSSD and the interpretation that fishing mortality should be reduced to FMSY by 2015 where possible (e.g. EC, 2006). The ICES MSY approach is thus based on this interpretation. In this approach, both fishing mortality and biomass reference points are used; these reference points are FMSY and MSY Btrigger. The approach does not currently use a BMSY estimate. BMSY is a notional value around which stock size fluctuates when F = FMSY. Recent stock size trends may not be informative about BMSY (e.g., when F has exceeded FMSY for many 12 ICES Advice 2013, Book 1 years or when current ecosystem conditions and spatial stock structure are, or could be, substantially different from those in the past). BMSY strongly depends on the interactions between the fish stock and the environment it lives in, including biological interactions between different species. MSY Btrigger is considered the lower bound of SSB fluctuation around BMSY. It is a biomass reference point that triggers a cautious response. The cautious response is to reduce fishing mortality to allow a stock to rebuild and fluctuate around a notional value of BMSY (even though the notional value is not specified in the framework). The concept of MSY Btrigger evolves from the PA reference point Bpa that ICES has used as a basis for fisheries advice since the late 1990s (see Figure 1.2.4). Bpa is derived from Blim based on the precision of the assessment, often taken as a standard value such that is in most cases Bpa = Blim*1.4. The evolution in the determination of MSY Btrigger requires contemporary data with fishing at FMSY to identify the normal range of fluctuations in biomass when stocks are fished at this fishing mortality rate. The ICES approach as specified in a hypothetical Harvest Control Rule (HCR)),) which shows how the target F should change with SSB is depicted in Figure 1.2.5. FMSY-HCR FMSY SSB (in the year for which advice is given) MSY Btrigger Figure 1.2.5 Approach shown in the ICES harvest control rule. Vertical axis is fishing mortality (F). Horizontal axis is spawning-stock biomass (SSB). Dotted section indicates stock below Blim Conceptually, SSB in the HCR is the estimated SSB at the beginning (or at spawning time) of the year to which the advice applies (advice year). For example, for an assessment performed in 2012 using data through 2011, the reference SSB will be the projected SSB at the beginning of 2013. FMSY-HCR is the fishing mortality rate used to calculate a catch option for the advice year. However, it may not be possible to project SSB to the beginning of the advice year, or the projections themselves may introduce so much additional uncertainty that it would be better to use a current SSB estimate. In such cases, the SSB used in the HCR will be the most recent reliable estimate. Since MSY Btrigger is intended to safeguard against an undesirable or unexpected low SSB when fishing at FMSY, the trigger reference point should be based on the natural variation in SSB and the assessment uncertainty, once FMSY has been reached. Ideally, FMSY should take account of selectivity, recruitment, growth, and natural mortality under current or recent ecosystem conditions, and be derived through stochastic simulations of target F in the context of a harvest control rule. However, recruitment functions are typically very noisy and poorly defined. It is therefore common to use proxies for FMSY, such as Fmax, F0.1, M, and F20-40%SPR 2 (Figure 1.2.6). Thus FMSY is used as a generic term for a robust estimate of a fishing mortality rate associated with high long-term yield. These proxies do not take into account the full range of stock dynamic directly but attempt to give good approximations to Fmsy where insufficient data is available to carry out a fuller evaluation. ICES will generally indicate when the advice is based on proxies. Conceptually these proxies have the following properties 2 F20-40%SPR are fishing mortalities that reduce the life time reproductive output of a year class to 20–40% of the reproductive output without fishing. ICES Advice 2013, Book 1 13 Fmax: The maximum yield point without accounting for the dependence of recruitment on SSB or its annual variability. Some stocks have a well defined Fmax at low F that is a good approximation for Fmsy. For other stocks the peak is either poorly defined at high F or not defined at all and the value is unsuitable. Fmax is sensitive to changes in the selection pattern / selectivity. The point where the increase in yield with increasing F is 10% of the rate at very low (around zero) F. This point F0.1: is often stable and well defined potentially giving a small reduction in yield relative to Fmsy, but may be quite close to Fmsy once dependence of recruitment on SSB and is annual variability is included. It is not suitable for stocks with higher natural mortality. M: Fmsy taken equal to natural mortality (M). Most suited to stocks with high natural mortality F20-40%SPR The fishing mortality that reduces the life time reproductive output of a year class to 20–40% of the reproductive output without fishing. It is based on a study of a wide range of stock biology. It has characteristics similar to F0.1 but is sensitive to assumptions of natural mortality as it depends on the unexploited biomass. Changes in selectivity, growth, natural mortality implies a re-estimation of those reference points. 0.8 120% Proxy ref. Points 0.7 100% 80% 0.5 Yield/R SSB/R 0.4 0.3 60% SSB/R Yield/R 0.6 40% 0.2 30% SSB/R 0.1 F 0 0 20% F 30%SPR F max 0.2 0.4 0% 0.6 0.8 1 Fishing mortality Figure 1.2.6 Illustrations of various proxies for FMSY. For SSB/R 100% is at F = 0. Other numerical values are illustrative only and will vary from stock to stock. As an initial option, MSY Btrigger is set at Bpa when this reference point is available, unless there is a sound basis for using a different value. In the future when there are sufficient observations of SSB fluctuations associated with fishing around FMSY, the MSY Btrigger should be re-estimated to correspond to the lower bound of the range of stock sizes associated with MSY. In general, re-estimated values of MSY Btrigger will be higher than Bpa because Bpa forms a lower boundary under the precautionary approach. The ICES harvest control rule (Figure 1.2.5) is designed to promote recovery of the stock to the normal range of stock sizes associated with FMSY when the stock is below this range (i.e. when it is below the MSY Btrigger). For most fisheries, recovery should theoretically occur at a fishing mortality of FMSY 3. The likelihood and speed of recovery is increased by reducing F whenever the stock is below the stock size range associated with fishing at FMSY. However, at very low stock sizes, the normal tendency for stock recovery at F less than or equal to FMSY may not hold. In these cases, the fishing 3 The theory is that fish populations compensate for fishing by increasing their production per unit stock size as stock size decreases. This type of response is known as compensatory. Production functions typically exhibit compensation. However, it is possible that at low stock sizes, production per unit stock size decreases as stock size decreases. This type of response is known as depensatory. It is difficult to observe (in part because there are few observations of stocks at very low stock sizes), but there are mechanisms that potentially result in depensation. Depensation has the potential to lead to extinction of a population. 14 ICES Advice 2013, Book 1 mortality rate derived from the HCR is likely to be so low that fishing may cease anyway. Nevertheless, when the stock size is so low that recruitment failure is a concern (e.g. well below Blim as estimated for a precautionary approach), additional conservation measures may be recommended to prevent a further decline. The special consideration given at low stock sizes is depicted by a broken line in Figure 1.2.5. Competent authorities receiving ICES advice have adopted several management plans in the spirit of the harvest control rule described above. When these plans are considered consistent with the precautionary approach and if no competent authority with a legitimate interest rejects a plan as the basis for the advice, the advice on the first page of the ICES advisory document will be based on the management plan. Other options will be also included in the body of the advisory report. The transition to advice based on reaching FMSY in 2015 Based on policy documents of management authorities and discussions with managers, there is general agreement that fisheries on the stocks for which ICES is requested to provide advice should be managed according to an MSY approach by 2015, but the transition should be gradual. Significant progress has been made in recent years developing and implementing precautionary management plans. Most of these plans are already developed based on delivering high long term yield and are conceptually similar to exploitation under MSY. These plans should not be jeopardized but they can, if necessary, be revised to be consistent with an MSY approach (as well as being precautionary). Over the next few years, ICES will advise on options that take account of this evolving situation. Although the World Summit on Sustainable Development (UN, 2002) called for stocks to be restored to levels that can produce MSY by 2015 where possible (which requires that overfishing relative to MSY be ended well in advance of 2015, but for many stocks it is already too late), this is not the policy of the European Commission (see EC, 2006). The EC and other management bodies that request advice from ICES have indicated they favour a gradual transition to implementing an MSY approach. Currently MSFD requires meeting Good Environmental Status (GES) which included F <Fmsy by 2020. Direct application of the ICES MSY approach In 2010 ICES introduced an MSY framework for fisheries advice. ICES provides catch options by direct application of the ICES HCR to give catch for Fmsy and where necessary catch options consistent with the MSY transition scheme. During the transition period (for advice in 2011–2015) where F is above FMSY and/or current biomass is below MSY Btrigger, ICES applies a stepwise transition to reach FMSY by 2015. The transition is in equal steps beginning with the year in which the transition was initiated. ICES Advice 2013, Book 1 15 Transition scheme If an estimate or a proxy of FMSY was available, the transition began in 2011 and F was to be reduced in five equal steps. Consequently, the catch option for 2014 will be: FMSY-HCR-transition (2014) = Min{0.2 • F (2010) + 0.8 • FMSY-HCR (2014); Fpa} whereas for 2015: FMSY-HCR-transition (2015) = Min{0.0 • F (2010) + 1.0 • FMSY-HCR (2015); Fpa} where F (2010) is the current year estimate of the fishing mortality in 2010 and FMSY-HCR (2014) is according to the ICES HCR in Figure 1.2.5, being equal to FMSY if SSB in 2014 is at or above MSY Btrigger and reduced linearly if SSB is below. The FMSY-HCR-transition values are capped at Fpa to maintain consistency with a precautionary approach. The plan for transition to MSY recognizes that managers requested a gradual transition, although they have not formally agreed to such a plan. However, there may be situations where a gradual transition is not appropriate because stock size is low (e.g. below Blim) and unless fishing mortality is reduced more rapidly the outlook is for a further decline (e.g. as a result of low recruitment). In such cases, ICES advises on a more rapid transition or application of the FMSY-HCR as soon as possible. 1.2.2.2.1b Short-lived stocks with population size estimates The future size of a short-lived fish stock is very sensitive to recruitment because there are only a few age groups in the natural population. Incoming recruitment is often the main or only component of the fishable stock. In addition, care must be given to ensure a sufficient spawning-stock size as the future of the stock is highly dependent on annual recruitment. For short-lived species, estimates or predictions of incoming recruitment are typically imprecise, as are the accompanying catch forecasts. For most short-lived stocks, the ICES MSY approach is aimed at achieving a target escapement (MSY Bescapement, the amount of biomass left to spawn, see Figure 1.2.4), which is robust against low SSB and recruitment failure and includes a biomass buffer to account for recruitment uncertainty. The yearly catches corresponds to the stock biomass in excess of the target escapement. No catch should be allowed unless this escapement can be achieved every year. For some short-lived species, assessments are so sensitive to incoming recruitment that the amount of biomass in excess of the target escapement cannot be reliably estimated until data obtained just prior to the fishery (or during the fishing year) have been analyzed. Therefore, an adaptive framework may be applied as follows: 1. 2. 3. Set a preliminary TAC that ensures a high likelihood of the target escapement being achieved or exceeded. This preliminary TAC is likely to be considerably below the final TAC (step 3). Assess the stock just before or during the fishing year, typically based on a survey or an experimental fishery. Adjust the TAC based on the assessment in step 2, ensuring that escapement is at, or above, the target. The MSY Bescapement should be set so there is sufficient biomass to provide the ecosystem services of a forage fish species and a low risk of future recruitment being impaired, similar to the basis for estimating Bpa in a precautionary approach. For short-lived species, where most of the annual surplus production is from recruitment (not growth), MSY Btrigger and Bpa might be expected to be similar. Therefore Bpa is a reasonable initial estimate of MSY Bescapement. 1.2.2.2.2 Stocks without population size estimates Of the more than 200 stocks for which ICES provides advice, ICES (2012a) determined that approximately half do not have population estimates from which catch options can be derived using the existing MSY framework. Up to and including 2011, ICES provided qualitative advice regarding the future exploitation of such stocks for which there is either limited knowledge about their biology or lack of data about their exploitation. Advice recipients have, however, expressed a strong interest in ICES developing quantitative advice based on the information available. In 2012, ICES has therefore developed a framework for quantitative advice regarding such stocks. This framework will, as other advice approaches, be refined in the future. The principles underlying this framework is that the available information should be used, that the advice to the extent possible should be based on the same principles as applied for stocks with analytical assessments and catch forecasts, and that a precautionary approach should be followed. The latter implies that as information becomes increasingly limited more conservative reference points should be used and a further margin of precaution should be adopted when the stock 16 ICES Advice 2013, Book 1 status is poorly known. The margin of risk tolerance is a management prerogative, but in the absence of any proposal by managers ICES has applied values which are given below. Unlike the classic fishery management problem of estimating maximum sustainable yield (MSY), fishery analysis on stocks without quantitative assessments must estimate a yield that is likely to be sustainable. The overall approach adopted by ICES has been developed under WKLIFE (ICES 2012a) and WKLIFE2 (ICES 2012c) and is explained in ICES report on the implementation of RGLIFE advice on Data Limited Stocks (ICES 2012c). The majority of the data-limited stocks have more information available than merely either catch or landings. The starting point for this analysis is therefore a categorization of the stocks according to the data and analyses that are available. The categorization of stocks is intended to reflect the decreasing availability of data, and thus the conclusions on the fishing pressure and state of the stock are likely to be less certain as one goes down the categories. As a consequence, a precautionary approach implies that exploitation rates advised for stocks below the data rich stocks (Category 1) will be more conservative than FMSY. Here we provide only a basic overview of the 5 additional categories that have been defined. Category 2 – stocks with analytical assessments and forecasts that are only treated qualitatively A quantitative assessment is available, but for a variety of reasons the information from the assessment has been used only as a description of trends such as fishing mortality, recruitment, biomass and future catches, rather than as an analytical assessment. Previously such analytic assessments were presented without Y-axis values. This approach uses a modification of the HCR from De Oliveira, J.A.A. et al., (2010). The general approach is to apply F0.1 as a robust and generally precautionary proxy for FMSY to account for the additional uncertainty associated with an assessment that cannot reliably estimate the size of the stock. Category 3 – stocks for which survey-based assessments indicate trends Surveys or other relative abundance or biomass indices are available for these stocks and they provide reliable indications of trends in total mortality, recruitment and abundance or biomass, but no quantitative, analytic assessment is available for the stock. The general concept of survey-based catch advice is based on Russell’s (1931) non-equilibrium definition of overfishing, in which catch exceeds biological production and causes a reduction in the stock. Therefore, decreasing surveys suggest catch should be incrementally decreased and vice versa. Category 4 – stocks for which only reliable catch data are available Only catch or landings data are available, and the data may not be continuous or consistent over time for a variety of reasons. The approach is to use estimates of Depletion-Corrected Average Catch (DCAC) to give approximations of stock depletion over the catch time series, it requires the ratio of FMSY/M and M. It assumes that the average catch has been sustainable in the past if abundance has not changed. Depletion Corrected Average Catch is an approximation of MSY. However, a catch advice based on MSY is only appropriate for stocks near BMSY (See Figure 1.2.3). For situations in which DCAC is much greater than recent catch, stock size may be less than BMSY and if F is decreased catch advice should increase slowly toward DCAC. Decreases or increases in catch are incremental and slow. Category 5 – Landing only stocks In the rare situation that only landings data are available, and no relevant life-history or fishery information can be gleaned from similar stocks or species in the eco-region or beyond the first approach should be to compile as much additional information as possible on fishery and survey data to transition to another category. However, in the meantime a Productivity and Susceptibility Analysis (PSA) risk assessment is used to evaluate the risks. Category 6 – negligible landings stocks and stocks caught in minor amounts as bycatch This category includes stocks where landings are negligible in comparison to discards. It also includes stocks that are part of stock complexes and are primarily caught as bycatch species in other targeted fisheries. The development of indicators may be most appropriate for such stocks. For each of these categories, methods have been employed to provide quantitative advice in 2012. Category one accounts for a precautionary approach through the use of PA reference and limit points. In order to apply a precautionary approach for categories 2-6 the framework for these stocks includes the following considerations regarding uncertainty and precaution have been applied in sequence: ICES Advice 2013, Book 1 17 - - As the methodologies used to estimate stock status, trends, and forecasts, due to the limited data available, are expected to be more susceptible to noise than methods used to produce forecasts for data-rich stocks, a change limit of ±20% (uncertainty cap) has been applied in the advice. This change limit is relative to the reference on which it is based and may be, e.g. recent average catches or a projection of a trend. A principle of an increasing precautionary margin with decreasing knowledge about the stock status has been applied: o The reference points for exploitation used have, when proxies could be identified, been selected on the lower margins of FMSY – either at the lower range of an interval as F0.1 or similar. o A precautionary margin of −20% (precautionary buffer) has been applied for those cases when the stock status relative to candidate reference points for stock size or exploitation is unknown. Exceptions to this rule have been made in cases where expert judgement determines that the stock is not reproductively impaired, and where there is evidence that the stock size is increasing or that exploitation has reduced significantly – for instance, on basis of survey indices or a reduction in fishing effort in the main fishery if the stock is taken as a bycatch species. This approach is intended to move in the direction of sustainable exploitation, having due regard for the species’ biological characteristics and uncertainty in the information. This implies that advice is applicable to a time-frame which is compatible with a measurable response in the metrics used as the basis for the advice; i.e. in the simplest case, and where the least information is available, this would imply a multi-annual constant catch advice. Where least information is available, including cases where the 20% precautionary margin has been applied, ICES therefore considers that the advice is not expected to be changed for a fixed and determined period such as, for example, three years, unless important new knowledge emerges regarding a stock which may justify a revision of the advice. 1.2.2.3 Frequency of advice and updates in 2013 In 2012 most stocks were allocated to categories and advice was provided. Many of these stocks have little new information on an annual basis and the advice can be used for two or more years. The 2012 advice will generally not be updated in 2013 if: o Biennial advice was provided in 2012 (e.g. elasmobranches, deep sea species and nephrops). o Only landings data are available and changes are negligible (categories 5 and 6 stocks). o The advice in 2012 was for lowest possible landings or zero catch advice (e.g. some elasmobranches, Irish sea cod) and there are no changes in the perception of the stock. o The PA buffer has been applied in 2012, except Pandalus in IIIa and IVa East for which an interbenchmark process is underway and for anchovy in IXa which is a short-lived species. o The DCAC method (category 4) was applied. o The advice has been the same for the last 3 years or more and the stock index has changed by less than 10%. The 2012 assessment and advice may be updated in 2013 if: o The PA buffer has not been applied and the stock index has declined by more than 10% compared to that used as the basis of advice in 2012. o There are doubts about the method applied in 2012 and a more appropriate method can be put forward. o Benchmarks have been held 1.2.2.3 Ecosystem considerations in fisheries advice The move toward an ecosystem approach to management (UN, 1992b, 2002; FAO, 1995) implies that human activities should be managed such that the overall health of the marine ecosystem is not placed at risk. This means that management of fisheries must consider not only the direct effects on fishery targets, but also the impacts on biodiversity, marine ecosystem structure, functioning, marine habitats and interactions between the fish populations. The advice must also be based on the best available knowledge about the interactions between the fish populations and their environment, be it other fish populations, other organisms, or the physico-chemical environment. A first step is to incorporate knowledge about the interactions between the various fish populations, for which advice is given. Two types of interactions should be considered in fisheries management, as described below. One type of interaction (referred to as “technical interactions”) results from the non-selective nature of many fishing operations. That is, the fishery captures a mixture of species and it is not entirely possible to control which species and how much of each is caught. For a mixed-species fishery, it may not be possible to achieve the single-stock MSYs (translated into TACs) of all the stocks simultaneously. Either the recommended maximum catches for some stocks will be exceeded in trying to catch the TACs of other stocks, or the TACs for some stocks will not be caught in order to prevent recommended maximum catches of other stocks from being exceeded. ICES has developed a mixed-species fisheries 18 ICES Advice 2013, Book 1 model (ICES, 2009a, 2010). The full value of this model (and future models of this type) will be realized with input from managers and stakeholders on trade-offs between species in the catch. The choice among different trade-offs and whether to fish only below or also above Fmsy for the different species in the fishery is not a scientific but a societal issue. However, ICES does consider that the precautionary approach and Fpa forms an upper bound for exploitation under these conditions. Estimates of MSY reference points depend on the size and age selectivity of the fishery. In many cases, both a higher yield and a larger stock size can be obtained by changing fishing practices (e.g. mesh size, fishing area and season) to achieve more favourable size and age selectivity. However, changing fishing practices to favour one species may put other species at a disadvantage in a mixed-species fishery. In the future, mixed-species fisheries advice should provide information to inform trade-offs between species in terms of changes in fishing practices that influence selectivity. As in 2012 ICES provides catch options in 2013 that incorporate technical interactions regarding demersal fisheries in the North Sea. The options are given as scenarios and not as the basis for the advice as single-stock management plans are currently in force, and mixed-fisheries advice would require an agreed policy for mixed-fisheries management by the relevant authorities. Another type of interaction results from ’biological interactions‘: some fish eat other fish, which means growth for the predator and mortality for the prey: fish populations also compete for food or habitat. Such interactions mean that as populations of one species increase by higher numbers and increased growth, populations of other species are likely to decrease because their mortality increases due to predation. It also means that as a population of fish increases one cannot expect that growth and mortality for that species remains constant as there will be increasing competition for food and habitat within that population. This is what is referred to in ecology as ‘density-dependence’ and is the reason that it is not realistic to assume simple projections of the growth of biomasses from low population sizes as the fishing pressure is reduced, for instance towards FMSY. This is also the basic reason for ICES to refrain from defining rebuilding targets based on a BMSY concept and the reason BMSY is not a part of the ICES approach to MSY. The implication is that all of the predicted increases in stock size based on applying an MSY approach on an individual stock basis are unlikely to occur simultaneously. Some stocks will increase substantially, but biological interactions may prevent other stocks from increasing as much as anticipated, and there may even be stocks that decrease in abundance as they are predated on by larger predator populations or are exposed to increased competition for food or habitat. ICES has for a number of years incorporated such multispecies considerations in the single-species framework by applying natural mortality or growth rates that are derived from models of species interactions using size, age, and stomach data for several species in Baltic, Barents Sea and North Sea. ICES has routinely incorporated short term changes in growth and maturation in short term projections in order to account for competition and food supply. ICES also expects to update MSY reference points regularly (typically as part of the benchmark process) again to reflect current dynamics. These aspects are a first step toward incorporating species interactions in fish stock assessments and advice and helps in making short-term predictions, where the surrounding ecosystem can be considered constant for a few years relative to the stock in question, more accurate. The utility of this approach is, however, very limited when it comes to medium-term forecasts or the exploration of long term reference points in an ecosystem context because the populations of all the various fish species are expected to change with changing fishing regimes and the interactions can therefore not be considered to be constant. This means that a full-fledged MSY approach cannot be implemented on a stock-by-stock basis. Basic MSY reference points such as FMSY, BMSY, and MSY Btrigger are conditional on a variable surrounding ecosystem and the other predator or prey fish populations living in it because growth and natural mortality, both of which are influenced by other fish populations, are determinants of these reference points. This means that the references to MSY reference points in UNFSA (UN, 1995) and other international agreements ultimately must be interpreted as features of the fish community or even the ecosystem rather than as constant parameters of a fish stock. Although biological interactions thus are important in terms of the response of stocks to a change in fishing pressure within a MSY approach, there are relatively few situations where the response of a multispecies community of fish to changes in fishing mortality can be reliably predicted. In the few cases where such predictions are possible, multispecies fishing mortality strategies can be developed to achieve MSY on a multispecies basis and to evaluate trade-offs between species based on preferences from managers and stakeholders. In situations where predictive models accounting for biological interactions are not reliable, it will be necessary to adopt a stock-by-stock MSY approach based on the observed response of these stocks once they have been fished at FMSY. As very few stocks have a history of exploitation at MSY target reference points, biomasses reference points can be expected to evolve as more and information become available. ICES Advice 2013, Book 1 19 As in 2012 ICES provides considerations in 2013 on options to incorporate biological interactions between herring, sprat, and cod in the Baltic in advice and fisheries management. The options are not presented as the basis for the advice as there are single-stock management plans in force, and multispecies fisheries advice would require an agreed policy for relevant authorities to consider biological interactions in fisheries management. Where relevant ICES indicates which options are considered precautionary. Achieving single- or multi-species MSY is not necessarily sufficient to assure all aspects of a healthy ecosystem and may need to be supplemented with measures to mitigate undesirable impacts on ecosystems. This need for supplementary measures is also considered in ICES advice. Reducing fishing mortality or changing selectivity should also reduce: (a) bycatch of non-target and sensitive species; (b) impacts on habitat and biodiversity; (c) the risk of truncated age structure; and (d) alterations that could possibly affect ecosystem functionality. In some cases, advice has included considerations of the impacts of fisheries on other components of the ecosystems. An example is the advice regarding sandeel, which is based on an escapement strategy to ensure that there is sufficient sandeel biomass to support populations of other biota that feed on sandeel. Where specific marine environmental management policies exist that require the regulation of fisheries to achieve their objectives, the fisheries advice will be restricted within the limitations required to achieve these objectives. In the EU context, this may be the case regarding fishing impacts on habitats relative to the Habitats Directive (Council Directive 92/43/EEC on the Conservation of natural habitats and of wild fauna and flora), and fisheries impacts on biodiversity, sea floor integrity, and foodwebs relative to the Marine Strategy Framework Directive (Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 establishing a framework for community action in the field of marine environmental policy). In the NEAFC context, advice has already been provided on fishing practices and fishing limitations to protect habitats of cold-water corals. 1.2.2.4 Management plan evaluations Recovery, or long-term, management plans have already been agreed for a number of fish stocks or fisheries within the ICES area, and new plans are being proposed. ICES has evaluated such management plans according to their compliance with a precautionary approach regarding risks to maintenance of reproductive capacity, and now also evaluates them according to the likelihood that high yields will be produced in the longer term. Stakeholders and authorities may raise other issues that may also be addressed in a specific management plan evaluation, such as stability of yield and risks under specific recruitment regimes. ICES has adopted one precautionary criterion for all medium/long lived stocks and a second similar criterion for short lived species. – 𝑴𝑴𝑴𝑴𝑴𝑴𝑴𝑴𝑴𝑴𝑴𝑴 𝒐𝒐𝒐𝒐 𝑳𝑳𝑳𝑳𝑳𝑳𝑳𝑳 𝒍𝒍𝒍𝒍𝒍𝒍𝒍𝒍𝒍𝒍: Management plan is precautionary if the maximum probability that 𝑆𝑆𝑆𝑆𝑆𝑆 is below 𝐵𝐵𝑙𝑙𝑙𝑙𝑙𝑙 is ≤ 5%, where the maximum (of the annual probabilities) is taken over all years in the plan (i.e. short and long terms), accounting for modification for recovery plans or initial recovery phases within long-term management plans. – 𝑺𝑺𝑺𝑺𝑺𝑺𝑺𝑺𝑺𝑺 𝒍𝒍𝒍𝒍𝒍𝒍𝒍𝒍𝒍𝒍 (a) If under natural conditions of no fishing the long-term annual probability of SSB being below 𝐵𝐵𝑙𝑙𝑙𝑙𝑙𝑙 is ≤ 5%, then the same criteria as for medium or long lived species is used. (b) If under natural conditions of no fishing, the long-term annual probability of SSB being below 𝐵𝐵𝑙𝑙𝑙𝑙𝑙𝑙 is > 5%, then the management plan is precautionary if the maximum probability that SSB is below 𝐵𝐵𝑙𝑙𝑙𝑙𝑙𝑙 is ≤ 5% (after the fishery) in any year when a fishery takes place. In all other years the fishery should be closed. Accepted plans with the above or more stringent criteria should not imply an increase of the long-term annual probability of SSB being below 𝐵𝐵𝑙𝑙𝑙𝑙𝑙𝑙 by more than a factor of 2 compared to natural conditions of no fishing. The management plans in place by 2012 were generally agreed prior to the introduction of MSY in the ICES advice, and on the basis of plan compliance with a precautionary approach. Some plans have since been evaluated with regard to generating high long-term yields, and these plans are considered also to be in accordance with an MSY approach. While the probability of avoiding a limit point should be less than 5%, ICES considers that a target point is reached if the associated probability of being above or below is 50%. 20 ICES Advice 2013, Book 1 It is generally anticipated that in the future competent authorities will aim at management plans (or replacement of management plans) being consistent with MSY. Management plan evaluations will be conducted to determine how plans perform in terms of long-term average catch, average stock size, average fishing mortality rate, and the statistical distributions of these variables. The ICES HCR will be used as a reference in comparing plan performance, although ICES does not expect that this HCR is superior to other HCRs, as it is selected to provide a simple approach with only the magnitude of yield as a consideration. Unless managers agree on specific performance criteria, the management plan evaluation can only be comparative; that is, ICES would not recommend one plan over another and would have no basis for rejecting a management plan if it is consistent with an MSY approach and it does not violate the precautionary approach. 1.2.3 ICES processes to provide stock status and single-stock advice ICES uses specific terminology and symbols or pictograms to describe the status of stocks. The wording aims at using a nomenclature which is less prone to misinterpretation, but at the same time allows for a match to the legal description, which still uses “safe biological limits“. ICES discontinued the use of this wording in 2008 as “safe biological limits” has in some cases misled the recipients of ICES advice and other stakeholders to consider stocks described as being “outside safe biological limits” to be biologically threatened (i.e. close to extinction). The terminology now uses different wording for the description of the stock status for biomass and fishing mortality and for the comparison to reference points based on an MSY approach, a precautionary approach, and existing and implemented management plans. The structure and the associated symbols and text are given below in Table 1.2.1 and 12.2.2 for MSY and precautionary approaches: Table 1.2.1 Symbols and text for MSY status MSY reference points Fishing mortality (FMSY) Biomass (MSY Btrigger) Explanation F < FMSY and Sign Text Appropriate F <<< FMSY (~ 0) Below target F > FMSY Above target No reference point Undefined Stock status unknown Unknown SSB = MSY Btrigger or SSB > MSY Btrigger SSB < MSY Btrigger At trigger or Above trigger No reference point Undefined Stock status unknown Unknown Below trigger Table 1.2.2 Symbols and text for precautionary status Precautionary reference points Fishing mortality (Fpa,Flim) Biomass (Bpa,Blim) Explanation Sign Text F =< Fpa Harvested sustainably Flim > F > Fpa Increased risk F > Flim Harvested unsustainably No reference point Undefined Stock status unknown Unknown SSB ≥ Bpa Full reproductive capacity Blim < B < Bpa Increased risk SSB < Blim Reduced reproductive capacity No reference point Undefined Stock status unknown Unknown In the case of management plans, the terminology changes depending on the characteristics of a specific reference point; namely, if the reference point is considered a target or a limit. If considered a target, this reference point would usually come with a target range, which means that a green symbol can be used when the stock is within the estimated or defined range, although for most of the stocks a range has not been defined. It is necessary to identify whether the reference points are defined as targets or as limits for each individual plan. Table 1.2.3 Symbols and text for status of stocks fished under management plans. ICES Advice 2013, Book 1 21 Management plan 4 reference points Fishing mortality (FMP) Biomass (SSBMP) Explanation Sign Text F< F mgt target / limit Below target /Below limit F within defined range At target or Within target range F>F mgt target / limit Above target / limit SSB > target, limit or trigger biomass Above target/limit/trigger SSB within defined range At target or Within target range SSB < target, limit or trigger biomass Below target/limit/trigger In situations where very limited information is available and the stock status table is filled with grey question mark symbols, ICES provides additional, qualitative information where available. For example, this information could be based on survey information and give an indication of stock status or trend. Table 1.2.3 Symbols and text for stocks with limited information. Qualitative evaluation Fishing mortality or exploitation rate Biomass If only trends are known Explanation Sign Text If there is an idea of the exploitation of this stock in relation to any possible reference points: If F is very high i.e. F > possible Short description reference points If F is very low i.e. F < possible Short description reference points If there is an idea of the state of this stock in relation to any possible reference points: If SSB is very low, i.e. SSB < possible Short description reference points If SSB is very high i.e. SSB > possible Short description reference points If parameter increases Increasing If parameter decreases Decreasing If trend is stable Stable The production of ICES advice can be separated into four distinct temporal phases (Figure 1.2.7): • • • • The first phase is the assembly of data up to December 31st of the year preceding the assessment year. The second phase is the assessment of the state of the stock at 1 January of the assessment year, This phase is looking at the past only and dealing with the stock status. The third phase between the assessment and the forecast is the assessment (interim) year. As incomplete data are available for this year (the year is not over yet), ICES has to make a number of assumptions on the fishery and biology. Some of these so-called interim year assumptions can significantly influence the catch forecast for the next year, but these assumptions are uncertain. If these assumptions prove to be markedly different from reality in subsequent assessments, stock status may be different than that forecasted. The fourth phase is the prognosis (forecast) on catch options to be taken next year (the year for which advice is given) and the state of the stock resulting from the different options.). Figure 1.2.7 Timeline of the production of ICES advice In some cases where stocks are short-lived (see above) this three phase approach is compressed into two years with catch options provided for the current year for in year management. The framework for the statement regarding future fisheries (third phase) has been developed in consultation with the relevant competent authorities. From among the range of catch options presented ICES provides a single main advice option which is based on the following principles: 4 Only included when plan is considered consistent with PA by ICES and agreed on by all relevant clients 22 ICES Advice 2013, Book 1 1. 2. 3. 1.2.4 If competent authorities with an interest in the stock have agreed that a management plan can be the basis for advice and this management plan has been found to be precautionary, this management plan will be the basis for the ICES advice. If this does not apply, the advice will be based on the ICES MSY framework. If there is no basis for giving MSY-based advice, advice will be based on precautionary considerations (see introductory section to the 2009 advice report). Advice to inform an ecosystem approach to marine management At the 13th Dialogue Meeting between ICES and the Clients (ICES 2004), the ICES plans for the introduction of an ecosystem approach into the advice were discussed. In 2008 ICES provided ecosystem overviews for the different sea regions (ICES, 2008). In 2012, these reviews will be updated starting with the Baltic Sea region. A new approach will be included in which an ecosystem description is combined with long-term trends in specific species or groups of species, and with long-term trends in drivers of ecosystem change such as climate and fishing pressure. Depending on the availability of long-term data, these overviews will be made available for the different ecoregions during 2013 and 2014. The organisation of the advisory report in ecoregions facilitates an ecosystem approach to marine management which is currently narrowly focussed on fisheries management. In future, non-fisheries parts of the trophic chains will be considered and integrated into advice going beyond fisheries management; e.g. aspects of eutrophication in the Baltic Sea linked with the abundance of the top predator cod. Our understanding of the functioning of the ecosystems is confined to certain ecosystem components. Work is underway to expand the number of ecosystem components beyond fisheries that are included in the ICES advice. However, this understanding is not uniform among ecosystems; some ecosystems have more data and the critical processes are understood better than in other ecosystems. Sources Anon. 1990. Icelandic Fisheries Management Act (No. 38, 15 May 1990). Anon. 1994. Faroe Islands Fisheries Management Act (Løgtingslóg nr. 28 um vinnuligan fiskiskap frá 10. mars 1994). Anon. 2004. Russian Federal Law onfFisheries and conservation of biological resources in the waters. N 166-P3 20/12/2004. Beddington, J. R., and May, R. M. 1977. Harvesting Natural Populations in a Randomly Fluctuating Environment. Science, 197 (4302): 463–465 (DOI: 10.1126/science.197.4302.463). De Oliveira, J.A.A., Darby, C.D., Earl, T.J. and C.M. O'Brien. 2010. Technical Background Evaluation of Annex IV Rules. ICES CM 2010/ACOM:58: 28 pp. EC. 2002. Council Regulation (EC) No. 2371/2002 of 20 December 2002 on the conservation and sustainable exploitation of fisheries resources under the Common Fisheries Policy. EC. 2006. Implementing sustainability in EU fisheries through maximum sustainable yield. Communication from the Commission to the Council and the European Parliament. COM (2006) 360 (final). EC. 2008. Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 establishing a framework for community action in the field of marine environmental policy (Marine Strategy Framework Directive). FAO. 1995. Code of Conduct for Responsible Fisheries. FAO Fisheries Technical Paper 350. http://www.fao.org/WAICENT/FAOINFO/FISHERY/agreem/codecond/codecon.asp. FAO. 2001. Reykjavik Conference on Responsible Fisheries in the Marine Ecosystem. Iceland, 1–4 October 2001. http://www.refisheries2001.org/. ICES. 2004. Report of the Thirteenth ICES Dialogue Meeting: Advancing scientific advice for an ecosystem approach to management: collaborating amongst managers, scientists, and other stakeholders. Dublin, Ireland, 26–27 April 2004. 2004. ICES Cooperative Research Report, 267. ICES. 2008. Report of the ICES Advisory Committee 2008. ICES Advice, 2008. ICES. 2009a. Report of the Ad hoc Group on Mixed Fisheries in the North Sea (AGMIXNS), 3–4 November 2009. ICES CM 2009/ACOM:52. ICES. 2009b. Report of the ICES Advisory Committee 2009. ICES Advice, 2009. Book 1: Introduction, Overviews and Special Requests. ICES. 2010. Report of the Working Group on Mixed Fisheries Advice for the North Sea (WGMIXFISH), 31 August–3 September 2010. ICES CM 2010\ACOM:35. ICES. 2012a. Report of the Workshop on the Development of Assessments based on LIFE history traits and Exploitation Characteristics (WKLIFE), 13-17 February 2012, Lisbon, Portugal. ICES CM 2012/ACOM:36. 121 pp. (in prep.) ICES 2012b. ICES implementation of advice for data limited stocks in 2012. Report in support of ICES advice. ICES CM 2012/ACOM:68. ICES Advice 2013, Book 1 23 ICES 2012c Report of The Workshop to Finalize the ICES Data-limited Stock (DLS) Methodologies Documentation in an Operational Form for the 2013 Advice Season and to make Recommendations on Target Categories for Data limited Stocks (WKLIFE2) ICES CM 2012/ACOM:79 Lovdata. 2008. Lov om forvaltning av viltlevande marine ressursar LOV-2008-06-06-37. http://www.lovdata.no/all/hl20080606-037.htmlhttp://www.lovdata.no/all/hl-20080606-037.html. Russel F. S. (1931). Some theoretical considerations on the “overfishing” problem. J Cons. Cons. Int. Explor. Mer. 6(1) 3-27 Sissenwine, M. P. 1978. Is MSY and adequate foundation for optimum yield? Fisheries, 3: 22–42. (doi: 10.1577/15488446(1978)003<0022:IMAAFF>2.0.CO;2). UN. 1982. United Nations Convention on the Law of the Sea (UNCLOS). http://www.un.org/Depts/los/convention_agreements/convention_overview_convention.htm. UN. 1992a United Nations Conference on Environment and Development (UNCED), Rio de Janeiro, Brazil. http://www.un.org/esa/dsd/agenda21/res_agenda21_00.shtml. UN. 1992b. Convention on Biological Diversity. http://www.cbd.int/convention/text/. UN. 1995. United Nations Conference on Straddling Fish Stocks and Highly Migratory Fish Stocks. http://www.un.org/Depts/los/convention_agreements/convention_overview_fish_stocks.htm. UN. 2002. World Summit on Sustainable Development (WSSD), Johannesburg, South Africa. http://www.un.org/jsummit/html/documents/summit_docs.html. WCED. 1987. Our Common Future. Report of the UN World Commission on Environment and Development 1987 http://www.un-documents.net/wced-ocf.htm. 24 ICES Advice 2013, Book 1 1.3 Technical basis for the advice The conceptual basis for the ICES advice on fisheries management is described in section 1.2 of this report. This section describes the technical basis for the advice including data and assessments. 1.3.1 Data used and data quality Catch and effort data The quality of the fish stock assessments is closely linked to the quality of the fisheries data, and ICES has expressed the greatest concern over the quality of catch and effort data for some of the important fisheries in the ICES area. The stock assessments presented in this report are carried out using the best possible estimates of the total catch. These estimates are not necessarily identical with the official landings statistics because they may include estimates of unreported landings and corrections for misallocation of catches by area and species. In the past there have been problems associated with discrepancies between the official landing figures reported to ICES by member countries and the corresponding catch data used by ICES. ICES recognises the need for a clear identification of the categories of the catch data. ICES attempts to identify factors contributing to the total removals from the various stocks through: • recorded landings, • discards at sea, • slipping of unwanted catches, • losses due to burst nets, etc., • unreported landings, • catch reported as other species, • catch reported as taken in other areas, • catch taken as bycatch in other (e.g. industrial) fisheries. The discards, slipped fish, unreported landings and industrial bycatches may vary considerably between different stocks and fisheries. It may not always be possible to reveal the sources of the estimated removals because of restrictions on how the data has been made available to ICES (e.g. confidentiality clauses). As a minimum, ICES describes the origin of the data (sampling programmes, field observations, interviews, etc.) so that interested parties can evaluate the quality of the information. Estimates of by-catches from the industrial fisheries are included in the assessments wherever the data is available. In recent years more information on discards has been collected through observer programmes and this information is increasingly made available to ICES for assessment purposes.. The catch data used by ICES are collated on a stock basis and not on an area basis so that direct comparisons between these figures and the official statistics are not always appropriate. ICES attempts to correct the shortcomings in the catch data. For non-reported landings such corrections, by their very nature, are difficult to document and are obviously open to debate. The stock assessments that are based on these data are of poor quality but they are still expected to be the best possible assessment of the state of the stocks. The fishing industry has on various occasions strongly disagreed with ICES’ estimates and has blamed ICES for not performing well. ICES does not accept the responsibility for quantifying non-reporting fisheries or ensuring access to proper discard data. The responsibility for discards and non-reporting and the uncertainty regarding the extent of these phenomena rests with the national authorities and the industry. When catch data could not be estimated, the trends in the stocks have sometimes been evaluated using research vessel data. This will only allow relative trends to be estimated and cannot be translated into a numerical advice on removals or effort. . Research vessel data Research vessel surveys are an essential fishery-independent source of information for scientists and a vital cross-check to the figures gathered from the international landings and from sampling onboard fishing boats. On research vessel surveys, scientists sample demersal fish such as cod, haddock, hake and plaice or pelagic fish such as mackerel and herring. ICES Advice 2013, Book 1 25 To sample fish on or near the seabed scientists use bottom trawls in the same way that fishers do. But whereas fishers target hotspot areas and continually try to upgrade their fishing gear to maximise their catch, fisheries scientists don’t want to maximise their catch but instead collect a representative sample. They also have to compare their results with previous years to follow trends, so it is vital that they use the same standard fishing gear each year rather than continually improving it. Research vessel surveys are carried out by national research institutes. ICES has an important role in internationally coordinating and analysing the surveys. Information from the fishing industry There is an increasing interaction between scientists and fishers during the collection of data in harbours and through observer programmes onboard fishing vessels. There have been a number of joint research projects between the fishing industry and scientists that have aimed to collect additional information on e.g. catch rates or catch compositions. In recent years, fishers in the North Sea have also been filling in questionnaires recording their perception of the state of key fish stock. This information is considered during the process of deriving ICES advice. Commercial Catch per Unit Effort (CPUE) series have been used in several stocks assessment as an indicator of stock abundance. In most cases the catch is then disaggregated by age through a market sampling process. A major difficulty in the use of CPUE series in stock assessment is the standardisation of fishing effort. The increasing efficiency of fishing vessels (e.g. through technical developments, GPS devices, new gear materials etc.) needs to taken into account in an estimate of effective fishing effort. This is not always possible due to lack of the relevant data for standardisation. The collaborations between the fishing industry and scientists has provided information which has been included as part of the assessment process. Such information has contributed to the understanding of the fisheries, and is increasingly provided in a form which enables direct inclusion in quantitative assessments. 1.3.2 Assessing the status of fish stocks Stock sizes and fishing mortalities are estimated in a stock assessment model. Most stock assessment models use catch at age information from the commercial fisheries and use additional information to “calibrate” the assessment. The additional information is mostly research survey indicators or catch rates in the commercial fishery (CPUE information). The estimated catches can be subject to serious bias if there are significant amounts of unreported landings or when information on discards at sea is not available. Catch information tends to become most unreliable when management measures are most restrictive (if they were implemented). In recent years several stocks have been at a low level and catch information has deteriorated for many fisheries. The consequence is that the ability to provide reliable, quantitative catch forecasts has decreased. Most management strategies in the ICES area rely on some forecast of the outcome of fisheries management in the management year. Under these conditions the Management Option table is an important part of the ICES advice. The catch options rely on estimates of recent stock size and fishing mortality and require an assumption about the total catch in the current or “assessment” year, because the fishery is rarely over when the assessment is carried out. 1.3.3 Evaluations of management plans When fisheries management plans have been agreed or proposed, ICES will evaluate the consistency of the management plan with international agreements and commitments. The main comparison will be in relation to the consistency with the precautionary approach. The methods for evaluating management plans differ by area, species and type of plan, but the general characteristics are that both fish populations and the management measures are simulated in a computer simulation process. The results of the simulations are scored in relation to the probability with which the stocks would be expected to be below Blim in near to medium term future. If the evaluation of a management plans indicates that a stock has a low probability (e.g. less that 5%) of being below Blim in the medium term, ICES considers the plan in accordance with the precautionary approach even when the stock is below the precautionary biomass level (Bpa) or above the precautionary fishing mortality (Fpa). 1.3.5 Quality of the advice ICES is dedicated to being transparent on the quality of the advice. Since 2004 competent authorities and a number of stakeholder organization are invited as observers to reviews and the advisory committee meetings. The quality of the 26 ICES Advice 2013, Book 1 advice can further be assessed by information in the advice on the basis for the advice relating to the subsequent years and, for stocks where analytical assessments could be carried out, a comparison between the most recent assessment and the previous assessments. ICES Advice 2013, Book 1 27 1.4 Structure of the Report The ICES advisory report is based on a regional orientation in so-called “Ecoregions” that allows the further development of an ecosystem approach in European waters. A review of existing biogeographical and management regions against a series of evaluation criteria has demonstrated that no existing regions could be adopted as ecoregions (ICES, 2004 5). The proposed ecoregions (Figure 1) are based on biogeographic and oceanographic features and existing political, social, economic and management divisions: • Iceland and East Greenland Seas • Barents Sea • Faroe Plateau Ecosystem • Norwegian Sea • Celtic Sea and west of Scotland • North Sea • South European Atlantic Shelf – Bay of Biscay and Iberian Seas • Baltic Sea • Oceanic northeast Atlantic The ecoregions Barents Sea and Norwegian Sea are presented in one single volume (3). The map also shows regions that are not covered by the ICES Advisory Report • Mediterranean Ecoregions: o Western Mediterranean Sea o Adriatic-Ionian Seas o Aegean-Levantine Seas • Black Sea The widely distributed and migratory species and the deepwater species for which stock identity have not been established, are addressed in Volume 9. The North Atlantic salmon stocks that are of interest to the North Atlantic Salmon Commission (NASCO) are treated in Volume 10. 5 ICES. 2004. Report of the ICES Advisory Committee on Fishery Management and Advisory Committee on Ecosystems, 2004. ICES Advice. Volume 1: 115–131. 28 ICES Advice 2013, Book 1 Figure 1.4.1 Proposed ecoregions for the implementation of the ecosystem approach in European waters, combined with ICES areas. ICES Advice 2013, Book 1 29 1.5 Answers to non-Ecoregion specific Special Requests 1.5.1 EU DG Mare 1.5.1.1 Special request, Advice April 2013 ECOREGION SUBJECT General advice Request from EU concerning monitoring of bycatch of cetaceans and other protected species Advice summary 1. Monitoring schemes. Sampling under the Data Collection Framework (DCF) can contribute to the assessment of bycatch of cetaceans and other species, but is not sufficient on its own as currently implemented by Member States. Not all fisheries are adequately covered and many issues, including design and sampling protocols would need to be modified/extended if DCF monitoring was to be the sole source of information. Monitoring under Regulation 812/2004 is much more specific for cetaceans, and has included the use of dedicated observers and remote electronic video recording. Development of remote electronic video recording seems likely to be a costeffective way of assessing bycatch in the future. 2. Acoustic Deterrent Devices (ADDs). ICES advises that regulation should not inhibit the development of more effective devices to deter harbour porpoises and other marine mammals from fishing gear. The characteristics of existing ADDs, which can deter harbour porpoises from fishing gear, are known. These characteristics cannot though be used to define all effective devices. Further studies would be needed to define standards for harbour porpoises and for ADDs that would be effective for other marine mammal species. To allow further development of ADDs, ICES recommends that a performance standard should be set. For an ADD to become acceptable, it should have a proven ability to reduce bycatch of the relevant species in the setting of a commercial fishery. 3. Reference points. Robust methods for setting reference points for bycatch of protected species already exist. ICES recommends that a process involving both managers and scientists be established to set species- and, where relevant, population-specific reference points. ICES proposes that a Bycatch Risk Approach be used to classify fisheries in terms of risk to protected species. Request ICES has been requested by EU to respond to the following request: “1. Assess the extent to which current fishery monitoring schemes, including inter alia those conducted under the DCF and Regulation 812/2004, provide an acceptable means of assessing the nature and scale of cetaceans and other protected species bycatch. Consider alternative means and other sources of data that could be used to improve our understanding of the conservation threat posed to cetaceans and protected species by bycatch in European fisheries. 2. Advise on how Annex II of Regulation 812/2004 defining technical specifications and conditions of use Acoustic Deterrent Devices could be best revised in light of technical and scientific progress in this field. 3 Based on the methodology used and the estimates of bycatch limits (take limits) generated by region at WKREV812 and other relevant analyses, propose effective ways to define limits or threshold reference points to bycatch that could be incorporated into management targets under the reformed CFP. Limits or threshold reference points should take account of uncertainty in existing bycatch estimates, should allow current conservation goals to be met, and should enable managers to identify fisheries that require further monitoring, and those where mitigation measures are most urgently required.” In the context of this request, ICES interprets “protected species” as those marine species listed in the relevant annexes of the Habitats Directive. The advice may also be relevant to bycatches of other marine species. ICES will be providing further advice on the bycatch of seabirds later in 2013. Elaboration of the advice 1. 30 Monitoring schemes. Sampling under the DCF tends to focus on the metiers that discard the most fish; these are not necessarily the same metiers that have the largest catch of protected species. Thus, bottom trawling is ICES Advice 2013, Book 1 generally oversampled with respect to monitoring of protected species bycatch, while in some specific fishing areas setnets, longlines, and purse-seines are undersampled. Some Member States have also undertaken additional observation schemes to meet the requirements of Regulation 812/2004 and those of the Habitats Directive (92/43/EEC). It would be possible to better define requirements on Member States under the DCF, but much will depend on how other data collection requirements will change under the revised Common Fisheries Policy. Alternatives to monitoring by human observers on-board vessels could include remote electronic video recording (e.g. Remote Electronic Monitoring), monitoring from vessels visiting a fishing fleet, interviews, and fishers’ logbooks. Indications of a bycatch problem may be obtained through schemes that examine stranding of protected species on shorelines. Of these alternatives remote electronic video recording seems to have the greatest potential to be developed to meet many of the needs of the existing DCF and also to improve monitoring of bycatch of protected species. 2. Acoustic Deterrent Devices. The standards currently listed in Annex II of Regulation 812/2004 are not fully appropriate. ICES advises that several commercially available ADDs have been proved effective in deterring harbour porpoises from fishing gear under specified conditions of use in commercial fisheries (Table 1.5.1.1). Other ADDs with the same specifications (and usage conditions) are therefore likely to be effective. It should be noted that many of these specifications are linked – thus for example, a higher source level ADD would not need to be spaced as close to another ADD as one with a lower source level. Some ADDs have been used to deter other species, and in some cases their efficacy for these species has been demonstrated. It is assumed that ADDs with similar key properties are as effective as those that have been tested. ICES recommends that ADDs targeted at reducing bycatch for these other species and under further development for harbour porpoises should only be authorised under Regulation 812/2004 if they have been proven by rigorous experiment to reduce bycatch rates by at least 80% with a 95% confidence. Experiments should be conducted in such a way that parties with a vested interest in the results cannot influence outcomes; they should include at least one control group and one treatment group. Experiments should be covered 100% by independent on-board observations and bycatch rates should be based on statistically independent bycatch events. Table 1.5.1.1 Overview of commercially available ADDs that have proven effective in deterring harbour porpoises from fishing gear. ADDs listed in italics do not have a published study of their effectiveness, but have the same specification as those with such a study. ADD type Source levels dB re 1 μPa rms at 1 m Signal frequency (kHz) Pulse duration (nominal) Interpulse interval (s) Dukane Netmark 1000 Fumunda 10 kHz Aquamark 300 Aquamark 100 132 10 300 ms 4 Maximum distance between any netting and the nearest ADD 200 m 132 132 145 10 10 20–160 300 ms 300 ms 200–300 ms 4 4 5–30 100 m 100 m 455 m DDD-03 L 174 5–500 0.5–9 s Random 4 km DDD-03 N 174 5–500 0.5–9 s Random 4 km 3. Reference Gönener and Bilgin (2009) Larsen and Krog (2007) Northridge et al. (2011) Reference points. Several methods have been used in defining limits or threshold reference points to bycatch of cetaceans (Table 1.5.1.2). The robustness of the various models to uncertain information varies. All rely on a public authority to define the overall conservation objective in terms that can be used in mathematical models, so their derivation requires not just the input of scientists, but also of relevant authorities. ICES cannot therefore provide advice on limits or threshold reference points, but instead recommends that the European Commission establishes a process involving both scientists and managers to derive these limits, using the most appropriate of these approaches for populations of species believed to be at most risk from bycatch. ICES advises that harbour porpoise, and common, striped, and bottlenose dolphin populations appear to be the species most at risk from bycatch in European waters at present. Table 1.5.1.2 Existing procedures to set limits and reference points for bycatch of marine mammals. Algorithm Catch Limit Algorithm (CLA)/Revised Management Procedure (RMP) ICES Advice 2013, Book 1 Management framework International Whaling Commission Conservation objectives 72% carrying capacity on average (50% of the time) Simulation timeframe 100 years 31 Potential Biological Removal (PBR) 1.7% of best available population estimate USA under Marine Mammal Protection Act ASCOBANS (for harbour porpoises in non-depleted areas) 50% of carrying capacity (95% of the time) 80% of carrying capacity (95% of the time) 100 years 100 years ICES advises that a Bycatch Risk Approach be used to identify areas and fisheries posing the greatest likely conservation threat to cetacean species due to bycatch (Figure 1.5.1.1). This approach can also be used for protected species other than cetaceans. The approach splits the population numbers of each protected species into different Management Areas (MA) and calculates take limits of species by area for any bycatch threshold level used. By using an expected bycatch rate (numbers per day or per unit of catch) multiplied by the total fishing effort, an approximate total number of bycaught animals can be estimated for each fishery and compared with any proposed take limit (see above). Figure 1.5.1.1 The Bycatch Risk Approach. Basis of advice 1. Monitoring schemes. The Data Collection Framework (DCF) is currently undergoing review and revision and will become the Data Collection Multi-Annual Programme (DCMAP). DCMAP will guide future fishery monitoring and data collection within the EU, covering a broad range of objectives. The contents of DCMAP are not yet determined, so ICES bases this advice on the current DCF as it seems likely that the principles will apply also to DCMAP. One of the current uncertainties in the DCMAP is the forthcoming EU ‘discards ban’ and how this will be implemented in detail. This ban may have profound consequences for fisheries monitoring in future. It could, for example, result in much greater emphasis on port-based sampling schemes, rather than seagoing observer schemes. It seems likely that bycatch of protected species will continue to be discarded and returned to the sea as they are “non-commercial” and unlicensed possession of such species is illegal in most EU Member States. In formulating the advice, ICES compared the coverage of current sampling under the DCF and the EU Regulation 812/2004 programmes with known abundances of cetaceans (and approximate indications of abundances of other protected species), with an index of bycatch vulnerability and with minimum estimates of fishing effort by metier (ICES, 2013b). Metiers/areas were identified where the risk to populations of certain species (groups) being adversely affected was greatest, and where coverage of the present monitoring schemes was relatively poor. 2. 32 Acoustic Deterrent Devices. Information was extracted from a recent review of the effectiveness of Acoustic Deterrent Devices (Dawson et al., 2013). ICES also approached a small number of underwater acousticians and manufacturers of ADDs and asked them for their views on how Annex II of Regulation 812/2004 could be best revised. A number of existing ADDs are effective in reducing harbour porpoise bycatch, but the full bounds of the technical specifications between an effective and an ineffective ADD are not known. ICES considers important that in advising on effective ADD specifications, there should be no constraints on the development of more effective devices for harbour porpoises or devices effective in deterring other species. ICES thus recommends that rigorous experiments are needed to demonstrate the effectiveness of new ADDs, while prescriptive constraints are unhelpful. ICES bases its advice for an >80% reduction in bycatch on the results of all the successful pinger experiments listed by Dawson et al. (2013), with a minimum confidence level of 95% as suggested/used for scientific studies. ICES Advice 2013, Book 1 3. Reference points. ICES has reviewed the existing procedures to establish limits and reference points (CLA, PBR, and 1.7%) several times in the past decade (SGFEN, 2002a, 2002b; ICES, 2012b). In all cases it was found that the choice of the most appropriate procedure depended on choices by managers in defining precisely the conservation objectives. These objectives essentially describe a societally chosen balance between exploitation of resources and conservation of protected species. The most appropriate way of working is therefore jointly between managers and scientists to explore and define conservation objectives. Furthermore, the choice of the most appropriate procedure to be adopted to achieve the conservation or management goal should be driven by the availability of suitable data. Sources Dawson, S. M., Northridge, S., Waples, D, and Read, A. J. 2013. To ping or not to ping: the use of active acoustic devices in mitigating interactions between small cetaceans and gillnet fisheries. Endangered Species Research, 19: 201−221. Gönener, S., and Bilgin, S. 2009. The effect of pingers on harbour porpoise, Phocoena phocoena, bycatch and fishing effort in the turbot gill net fishery in the Turkish Black Sea coast. Turkish Journal of Fisheries and Aquatic Sciences, 157: 151−157. ICES. 2010. Report of the Workshop to evaluate aspects of EC Regulation 812/2004 (WKREV812). ICES CM 2010/ACOM:66. 65 pp. ICES. 2012a. Report of the Study Group on Practical Implementation of Discard Sampling Plans (SGPIDS). ICES CM 2012/ACOM:51. 81 pp. ICES. 2012b. Report of the Working Group on Marine Mammal Ecology (WGMME). ICES CM 2012/ACOM:27. 140 pp. ICES. 2013a. Report of the Working Group on Bycatch of Protected Species (WGBYC). ICES CM 2013/ACOM:27. 83 pp. ICES. 2013b. Report of the Workshop on Bycatch of Cetaceans and other Protected Species, 2013 (WKBYC). ICES CM 2013/ACOM:36. 41 pp. Larsen, F., and Krog, C. 2007. Fishery trials with increased pinger spacing. Paper presented to the Scientific Committee of the International Whaling Commission. IWC SC/ 59/ - SM2, International Whaling Commission, Cambridge. Northridge, S., Kingston, A., Mackay, A., and Lonergan, M. 2011. Bycatch of vulnerable species: understanding the process and mitigating the impacts. Final report to Defra, Project MF1003. 99 pp. Available at: http://randd.defra.gov.uk/Document.aspx?Document=MF1003-FINALRevisedAugust2011.pdf SGFEN. 2002a. Incidental catches of small cetaceans. Report of the meeting of the subgroup on fishery and the environment (SGFEN) of the Scientific, Technical and Economic Committee for Fisheries (STECF), Brussels, 10–14 December 2001. SEC (2002) 376. 83 pp. SGFEN. 2002b. Incidental catches of small cetaceans. Report of the second meeting of the subgroup on fishery and the environment (SGFEN) of the Scientific, Technical and Economic Committee for Fisheries (STECF), Brussels, 11–14 June 2002. SEC (2002) 1134. 63 pp. ICES Advice 2013, Book 1 33 1.5.1.2 ECOREGION SUBJECT Special request, Advice June 2013 General advice New information regarding the impact of fisheries on other components of the ecosystem Advice summary ICES advises that the following seven areas contain habitats sensitive to bottom fishing activities: Hebrides Terrace Seamount; Rosemary Bank Seamount; Porcupine Sea Bight; Faroe–Shetland Channel and Tampen area; Irish Margin/Bay of Biscay; Gulf of Cadiz; Northwest Rockall Bank. Request Provide any new information regarding the impact of fisheries on other components of the ecosystem incl. small cetaceans and other marine mammals, seabirds and habitats. This should include any new information on the location of habitats sensitive to particular fishing activities. [Memorandum of understanding between the European Union and the International Council for the Exploration of the Sea, 2013]. ICES advice The advice in this section covers new information on the location of habitats sensitive to particular fishing activities (i.e. vulnerable marine ecosystems, VMEs). Advice on the impact of fisheries on seabirds is being collated under the heading of another special request from the European Commission that will be answered in the second half of 2013. The focus of ICES work in 2013 on bycatch of marine mammals and other components of the marine ecosystem has been more towards reviewing and advising on an improved monitoring and assessment system. This has been advised upon already to the European Commission (on 25 April 2013; ICES, 2013a). The only new data available to ICES in 2013 on the issue of bycatch is that contained in Member States’ reports to the Commission under Regulation 812/2004. ICES considers that no new issues have arisen as a consequence of the information in those reports. New information on the location of habitats sensitive to particular fishing activities New data that indicate the presence of VMEs were submitted to ICES in 2013 (ICES, 2013b). Some of these data were within the EEZs of Member States of the EU. ICES has no information (i.e. VMS data) available on actual fishing activity in these areas at the spatial resolution required to evaluate the pressure on the habitats. 1) Hebrides Terrace Seamount The Hebrides Terrace Seamount lies to the west of the UK, being partially joined to the continental slope. The summit is around 1000 m and the steep-sided flanks descend to below 2000 m. In 2012 a research survey completed two ROV (Remotely Operated Vehicle) transects of the steep flanks of the seamount and one transect across the summit. On the seamount summit, three ROV still images contained VME indicator species and none were at densities that would indicate actual VMEs. On the steep flanks, however, between the depths of 1200 m and 1700 m, coldwater corals were consistently observed at high densities, indicating VMEs (Figure 1.5.1.2.1). 34 ICES Advice 2013, Book 1 Figure 1.5.1.2.1 Observations of VME indicators on the Hebrides Terrace Seamount. 2) Rosemary Bank Seamount The Rosemary Bank Seamount lies at the north end of the Rockall Trough. In 2012 a trawl sample was obtained from the lower muddy slope on the eastern side of the bank at a depth of around 1300 m (59.245°N; −9.525°W). A large bycatch (>1000 kg) of Geodia sponges (a species vulnerable to bottom fisheries) was taken. 3) Porcupine Sea Bight Positions of deep-sea sponge aggregations reported by Rice et al. (1990) have become available. These were collected during scientific trawl surveys at depths between approximately 1000 and 1500 m (Figure 1.5.1.2.2). ICES Advice 2013, Book 1 35 Figure 1.5.1.2.2 Positions of deep-sea sponges in the Porcupine Seabight area. 4) Faroe–Shetland Channel and Tampen area A recent (2011) trawl bycatch record of a deep-sea sponge aggregation (estimated weight >1000 kg) confirms the northern part of the Faroe–Shetland Channel and Tampen area as important for deep-sea sponge habitat (Figure 1.5.1.2.3). Figure 1.5.1.2.3 36 Positions of deep-sea sponge observations in the Tampen area. The red circle indicates the new observation. ICES Advice 2013, Book 1 5) Whittard Canyon (Irish Margin) ROV transects found VMEs throughout the Whittard Canyon along with dead coral rubble fields (Figure 1.5.1.2.4). VME indicator species observed included Schizopathidae, Carophyliidae, Gorgonacea, Alcyoniidae, Paragorgiidae, Chrysogorgiidae, Isididae, Stylasteridae, Primnoidae, and Pennatulacea. Figure 1.5.1.2.4 Positions of VME indicator species in the Whittard Canyon. 6) Gulf of Cadiz (Spain) There are cold-seeps and mud-volcanoes on the Guadalquivir Diapiric ridge in the Gulf of Cadiz (36.5°N; −7.25°W). These VMEs are at depths of approximately 550 m and therefore potentially at risk from bottom fishing activity. 7) Northwest Rockall Bank Rockall Bank is a large plateau that lies partly in EU waters and partly in international waters regulated by NEAFC. Six further video transects revealed extensive patches of coral reefs in the centre and toward the southern end of the current closure (Figure 1.5.1.2.5). Another video transect revealed new observations of coral reefs outside the eastern part of the closure. No VMEs were observed along a transect to the north of the closure. ICES Advice 2013, Book 1 37 Figure 1.5.1.2.5 Map of Northwest Rockall showing locations of video transects and new findings of VMEs. References ICES. 2013a. Report of the ICES Advisory Committee 2013. ICES Advice, 2013. Book 1, Section 1.5.1. ICES. 2014b. Report of the ICES\NAFO Joint Working Group on Deep-water Ecology (WGDEC), ICES CM 2013/ACOM:28. 370 pp. Rice, A. L., Thurston, M. H., and New, A. L. 1990. Dense aggregations of a hexactinellid sponge, Pheronema carpenteri, in the Porcupine Sea Bight (northeast Atlantic Ocean), and possible causes. Progress in Oceanography, 24: 179– 196. 38 ICES Advice 2013, Book 1 1.5.1.3 Special request, Advice December 2013 ECOREGION General advice SUBJECT EU request on monitoring of bycatch of seabirds Advice summary ICES advises on a series of fisheries where bycatch is likely to pose a risk to seabird populations. Fisheries within each of the four European DCF regions as well as some parts of the larger DCF areas were reviewed for level IV métiers. Information on bycatch remains relatively sparse and is becoming increasingly dated. Priority should be given to monitoring in trammelnets and set gillnets in the Baltic, North Sea, and North Atlantic, and in set long-line fisheries in the Atlantic and Mediterranean/Black Sea. A framework, including metrics and criteria, to define a seabird bycatch problem is described and explored. Further work would be needed to assess the risk of bycatch adversely affecting seabird populations if this framework was adopted. A standard data format is under development and the database developed by ICES for marine mammal bycatch is designed also to store seabird bycatch data. Request The European Commission (DG MARE) requests ICES to consider the following: • • • To review and update current seabird bycatch data and identify fisheries where appropriate follow up monitoring to establish bycatch levels would be desirable. To explore the criteria and/or metrics that could be used to define a seabird bycatch problem. (This request is partially addressed in earlier request by the Commission to ICES on cetacean bycatch) but ICES should tailor this advice to specifically cover seabird bycatch. Establish a standard data reporting format for recording seabird bycatch and develop a database of seabird bycatch data in EU fisheries, similar to the database developed by WGBYC for marine mammal bycatch. ICES advice 1. Fisheries where appropriate follow-up monitoring to establish bycatch levels would be desirable Table 1.5.1.3.1 lists those fisheries for which ICES advises that bycatch of seabirds should be monitored. Fisheries are specified at métier level 4 (gear type) for each of the DCF regions Baltic Sea, North Sea, North Atlantic, and Mediterranean Sea/Black Sea. Only those fisheries are listed where bycatch of seabirds is known or at least can be suspected. The decision on whether to recommend monitoring in individual fisheries was based on one of these situations: • • Information/data already exist that indicate seabird bycatch. This information might originate either from dedicated seabird bycatch monitoring schemes, from data collected during the DCF observer programmes, or from specific projects on seabird bycatch. Expert judgement determines that bycatch could be suspected/expected/is likely in this particular fishery. This judgement could be based either on anecdotal information about seabird bycatch, or on transfer of knowledge about the same fishing method from other marine areas. It could also be based on biological information, for instance that life habits (such as foraging strategies) of similar bird species make interactions with the specific fishing method likely. In Table 1.5.1.3.1, countries were named only to indicate where information about seabird bycatch comes from. For most regions, it can be assumed that any bycatch occurring in one fishing fleet is likely to occur in similar fleets of other countries using the same gear. It is likely that there is variance in risk at a sub-regional scale due to variation in bird occurrence and in location of fisheries. Any bycatch recording scheme that is being established should, if possible, take account of any knowledge of such variation. If monitoring for seabird bycatch is indicated in the table as desirable, this does not necessarily mean that a full and regular monitoring scheme should be implemented. In many cases, especially where seabird bycatch is only suspected, the first step could be to run specific monitoring pilot projects to gain better knowledge about the amount and extent of seabird bycatch. It should also be noted that the status of seabird populations has not been taken into account; monitoring of bycatch in an increasing population may be less urgent than in a decreasing population. Analysis of existing information ICES Advice 2013, Book 1 39 on causes of mortality (e.g. from returns of seabirds that have been marked and found dead) may also be useful in prioritizing monitoring. 40 ICES Advice 2013, Book 1 Table 1.5.1.3.1 Gear types where bird bycatch is known or suspected and ICES advice on the desirability of monitoring. Colours in the gear type cells symbolize level of activity: orange – common fishing gear/activity in this ecoregion; green – negligible or non-existent; white – not known. In the ”Bird bycatch” column, “Yes” means data exist and ”Suspected” means expert judgement has been applied, with the justification for this judgement being summarized in the final column. An orange colour in the ”Monitoring desirable” column indicates a monitoring priority based on likely scale of bycatch. Baltic Gear type Bird bycatch Monitoring desirable Remarks and rationale for monitoring Drifting longlines [LLD] Set longlines [LLS] Suspected Yes Yes Yes Pots and traps [FPO] Suspected Yes No data are available on bycatch of birds in the Baltic, but susceptible species that occur in the Baltic are caught elsewhere in this gear. Bycatch has been recorded in fisheries targeting flatfish, cod, flounder, and eel in German waters; the species caught include great cormorant, auks, and seaduck. Evidence from elsewhere indicates that bycatch of species such as great cormorant can occur. Fykenets [FYK] Yes Yes Great cormorants have been recorded as drowning in fykenets in Danish waters. Stationary uncovered poundnets [FPN] Yes Yes This gear is used on all Baltic coasts. A low bycatch of great cormorant has been reported in Denmark, Germany, and Sweden. Trammelnet [GTR] Set gillnet [GNS] Yes Yes Bycatch in gillnets and trammelnets (most studies of seabird bycatch do not distinguish between the two) has been found to be high and possibly unsustainable in a number of Baltic countries. Diving birds that are affected include divers, grebes, great cormorant, seaduck, and auks. ICES Advice 2013, Book 1 41 North Sea Gear type Bird bycatch Monitoring desirable Remarks and rationale for monitoring Bottom otter trawl [OTB] Yes No Small numbers of common guillemots have been caught in sandeel trawls in the North Sea during the seabird breeding season; this fishery is short in duration. Midwater otter trawl [OTM] Set longlines [LLS] Yes Yes Northern gannets have been caught in the herring trawl fisheries off the northern and northeastern coasts of the UK. Suspected Yes No data are available on bycatch of birds in set longlines, but species that occur in the North Sea are caught in this gear elsewhere. Fykenets [FYK] Yes Yes Great cormorants have been recorded as drowning in fykenets in Dutch waters. Stationary uncovered poundnets [FPN] Yes Yes Three species of auk have been recorded bycaught in UK J- and T-nets targeting salmon and sea trout; that fishery is closed if bycatch gets too high. High levels of seabird bycatch have also been noted further north in UK. Trammelnet [GTR] Set gillnet [GNS] Yes Yes From the 1980s, bycatch of many thousands of seaduck and great crested grebe birds has been recorded off the Netherlands; despite mitigation measures, bycatch still occurs. There are few recent data but bycatch of seaduck and diving seabirds still occurs in UK waters and possibly also off Sweden, Germany, and Denmark where similar fisheries operate. Driftnet [GND] Yes No Bycatch of great cormorants, northern gannets, gulls, and common guillemot has been recorded in the UK bycatch monitoring scheme, notably in salmon and sea trout fisheries off Northumberland and Yorkshire. This fishery is being phased out. 42 ICES Advice 2013, Book 1 North Atlantic Gear type Bird bycatch Monitoring desirable Remarks and rationale for monitoring Bottom otter trawl [OTB] Midwater otter trawl [OTM] Yes Yes Yes Yes Midwater pair trawl [PTM] Trolling lines [LTL] Yes Yes Suspected Yes Drifting longlines [LLD] Set longlines [LLS] Yes Yes Yes Yes Pots and traps [FPO] Yes Yes Bycatch, including common guillemot, razorbill, and great cormorant, occurs regularly in the seabass fishery in the western Channel off the UK and France. Some bycatch of gulls occurs in the French anchovy fishery. There are no data on bycatch from the EU albacore fishery that operates mainly in the Bay of Biscay; seabird bycatch occurs in trolling operations elsewhere. Low bycatch, including northern gannet, shearwaters, and yellow-legged gull occurs in fisheries targeting tuna and other large fish in the waters of Spain, Portugal, and France. Throughout the region, fisheries targeting a wide range of fish species bycatch many species of seabird, in some cases in substantial numbers – for example tens of thousands of northern fulmar and great shearwater. Both surface-feeding birds such as gulls and diving seabirds such as cormorants and auks are affected. There is some bycatch of European shags in Spain. Fykenets [FYK] Suspected Yes Great cormorants have been recorded as drowning in fykenets in other waters. Trammelnet [GTR] Set gillnet [GNS] Yes Yes There is a regular and widespread bycatch that includes auks, shearwaters (notably Balearic shearwater), northern gannet, cormorants, seaduck, and divers. Driftnet [GND] Suspected Yes Purse seine [PS] Yes Yes Beach and boat seine [SB] [SV] Yes Yes Small driftnet fisheries in the UK and France target a range of fish species. The available information indicates no bycatch in these, but bycatch does occur in driftnets in other regions and the species affected also occur in the North Atlantic. Bycatch, especially of Balearic shearwaters occurs in the Portuguese fishery during the non-breeding period. Seabird mortality has also been reported in the Spanish fishery. Bycatch of common scoter (and black-headed gull) has been recorded in the Portuguese fishery. The common scoter has an important wintering area off the Portuguese coast. ICES Advice 2013, Book 1 Bycatch has been recorded in Spanish, Portuguese, and French waters in multispecies fisheries. The species caught include northern gannet, gulls, shearwaters, and cormorants. Bycatch has been recorded in the Portuguese seabass fishery. Bycatch of northern gannets has been recorded in the argentine fishery off northwestern UK, and also in fisheries to the west of the UK targeting herring, mackerel, and horse mackerel. 43 Mediterranean Sea and Black Sea Gear type Bird bycatch Monitoring desirable Remarks and rationale for monitoring Bottom otter trawl [OTB] Yes Yes Some bycatch of Balearic shearwater, northern gannet, and gulls occurs in the Spanish multispecies fisheries. Trolling lines [LTL] Yes Yes Scopoli’s and Yelkouan shearwaters are bycaught in the Ionian and Aegean Seas respectively, and there is a bycatch of Mediterranean shag in the Ionian Sea. Drifting longlines [LLD] Yes Yes Bycatch has been recorded mainly in the western Mediterranean. The swordfish fishery has bycatches of principally Scopoli’s shearwater, but also Balearic shearwater and yellow-legged gull are bycaught. In the eastern Mediterranean Scopoli’s shearwater has been bycaught. No bycatch has been recorded in the Aegean and no information is available for certain parts of the Mediterranean (e.g. the Adriatic). Set longlines [LLS] Yes Yes Bycatch of seabirds including shearwaters (Balearic, Yelkouan, Scopoli’s), Audouin’s gull, Mediterranean gull, yellow-legged gull, Mediterranean shag, black-legged kittiwake, and skuas occurs in various fisheries targeting mainly hake, Sparidae, and other demersal fish. Pots and traps [FPO] Yes Yes Mediterranean shag is bycaught in Spanish lobster fishing; no information for other parts of the Mediterranean. Trammelnet [GTR] Set gillnet [GNS] Driftnet [GND] Yes Yes Bycatch has been recorded of a wide range of seabird species, including Mediterranean shag and great cormorant, seaduck, razorbill, Yelkouan (and potentially Balearic) shearwaters, and Scopoli’s shearwater. Suspected Yes Bycatch of species that occur in the Mediterranean has been reported in other areas for this gear. Purse-seine [PS] Yes Yes Bycatch of Balearic (and potentially Yelkouan) shearwaters has been reported in Spanish fisheries for sardine, anchovy, and other small pelagics. Beach and boat seine [SB] [SV] Recreational fisheries Suspected Yes Bycatch of species that occur in the Mediterranean has been reported in other areas for this gear. Yes Yes Bycatch of Mediterranean shag, shearwaters, and Audouin’s gull has been reported in Spain and Greece (particularly hand and pole, trolling lines). 44 ICES Advice 2013, Book 1 2. Criteria and/or metrics that could be used to define a seabird bycatch problem What is a bycatch problem for seabirds? The definition of what constitutes a problem is of course societal rather than scientific. Societal choices can be derived from legislation and internationally agreed guidelines. In Europe, Directive 2009/147/EC on the conservation of wild birds (the Birds Directive) is the most important statute. This sets an overall objective for bird populations: “Member States shall take the requisite measures to maintain the population of the [bird species] at a level which corresponds in particular to ecological, scientific and cultural requirements, while taking account of economic and recreational requirements, or to adapt the population of these species to that level. (Art. 2).” In only a very few cases is the population of a seabird species believed to be too great, so it may be assumed that bycatch that causes a population decline is a problem, and for a number of cases where a species is believed to be depleted, bycatch that overly affects the ability of a population size to recover may also be a problem. In addition to the “legislative” approach above, public perception of waste and unnecessary death is relevant; thus in the case of oil spills, most members of the public regard this as a waste (both of oil and of seabirds) that is unnecessary as oil spills can largely be prevented or avoided. Seabird bycatch does not have the same public profile as oil spills (partly because it is characteristically chronic and continuous, as compared with the episodic and catastrophic nature of high profile oil spills). It is also the case that it is difficult to prevent or avoid some seabird bycatch if some particular fisheries are to continue. It is likely that a high (but not biologically important) bycatch in a fishery where mitigation is possible would be regarded as wasteful and as a problem. This would be consistent with the General Principles section of the FAO Code of Conduct for Responsible Fishing, whereby “States and users of aquatic ecosystems should minimize waste, catch of non-target species, both fish and non-fish species, and impacts on associated or dependent species.” Framework, criteria, and metrics for defining whether bycatch poses a problem to a seabird species As indicated in the IPOA (International Plan of Action)–Seabirds and reiterated by FAO (2008), there are key considerations in identifying whether or not a seabird bycatch problem exists: “When defining a seabird incidental catch problem, States and RFMO/As [Regional Fisheries Management Organisations/Arrangements should consider the following: (i) (ii) (iii) (iv) Defining the rationale for determining if a problem does, or does not, exist. The rationale should be based on: (a) the magnitude of seabird bycatch (rate or number); (b) species that are incidentally caught, and their conservation status; and (c) spatial and temporal overlap of fishing effort with seabirds.” Reviewing available data relevant to the incidental mortality of seabirds. Validating sources of information and where appropriate follow up with more detailed investigations. Adopting a precautionary approach where information is lacking or uncertain.” ICES advises that these considerations, when taken broadly and in combination with one another, provide sufficient criteria for defining a seabird bycatch problem when applied within a step-wise assessment framework. ICES advises that the application of Potential Biological Removal (PBR) may be utilized to serve as the primary metric for determining whether a seabird bycatch problem exists. PBR is derived from a formula that combines information on the maximum annual recruitment rate of populations, a conservative estimate of population size, and a recovery factor. PBR = ½ RMAX × Nmin × f RMAX is the maximum annual recruitment rate, Nmin is a conservative estimate of population size (e.g. the 20th percentile of the population estimate), and f is a recovery factor usually between 0.1 and 1. RMAX can be estimated from the annual adult survival rate in the absence of human effects and the average age at first breeding. The recovery factor f is usually set in categories relating to the amount by which a population is believed to be depleted or is declining and can be designated by broad taxonomic groups, as has been done for marine mammals (in the USA) to account for differences in their life histories. ICES advises the following assessment framework (Figure 1.5.1.3.1) be used in the identification of a seabird bycatch problem, based upon whether or not seabird bycatch is a substantial portion of the anthropogenic impact that causes losses greater than PBR of a particular species or population. ICES suggests that as a criterion “a substantial portion” could be interpreted as when bycatch exceeds 30% of anthropogenic mortality to the species or population. ICES is aware of some projects to evaluate the relative contribution of bycatch to overall anthropogenic mortality, but these projects are rare. Further information may be available from analyses of the causes of death of ringed birds. ICES Advice 2013, Book 1 45 Does Total Mortality Exceed PBR by Species or Population? Initial/Risk Assessment No Yes Is a Substantial Portion of the Mortaliy from Fisheries? No Yes Mitigation Measures Figure 1.5.1.3.1 Maybe Collect Information/Increase Monitoring A proposed framework for exploring the criteria and metrics that could be used to define a seabird bycatch problem. Stage 1: The PBR (metric) for each seabird species or population is calculated, using suitable proxies where there is insufficient information for each species or population. Stage 2: Once the PBR has been calculated, an initial rapid assessment (similar to a risk assessment) should be conducted to determine whether, based on existing information (including inferred information) there is a risk that the total estimate of anthropogenic mortality exceeds the PBR, and whether bycatch is a substantial portion of that mortality. The information to be used for this assessment may include (but is not limited to): • • • • • • observations of bycatch, the results of interviews and/or questionnaires, information regarding the nature of the fishery interactions, the geographic overlap between a given species and a fishery/gear type, information regarding non-fisheries-related mortality, and inferred information regarding the likelihood of certain species interacting with one or more fisheries. If the initial assessment results in a determination that it is not likely that PBR has been exceeded for all anthropogenic mortality, then a seabird bycatch problem does not exist for this species or population. No further immediate action needs to be taken. ICES recommends that this initial assessment be conducted per species/population at least every five years. If the initial assessment results in a determination that it is likely that PBR has been exceeded and fisheries are contributing a substantial portion of the removals, then a seabird bycatch problem may exist for this species or population. In this case, a third step is recommended. Stage 3: A more detailed evaluation of existing information, possibly including further studies to verify the existing data and to further examine the nature and magnitude of the interactions, should be conducted. If this more detailed analysis confirms that the total estimate of anthropogenic mortality exceeds the PBR, and that bycatch is a substantial portion of that mortality, then a determination would be made that a seabird bycatch problem exists for this species. ICES recommends that bycatch mitigation measures for all involved fisheries should be considered and put into effect as rapidly 46 ICES Advice 2013, Book 1 as possible. If the results of the more detailed study are ambiguous, it can be reaffirmed that a bycatch problem may exist. In these circumstances, ICES recommends that additional information, through scientific study or through monitoring, be collected. The results of these efforts will provide additional information from which to conduct future assessments of whether bycatch is exceeding PBR. For seabird species for which a bycatch problem may be occurring, a review of any new information should be conducted until either a seabird bycatch problem has been identified to exist or not to exist. Where information remains uncertain regarding the existence of a bycatch problem, ICES recommends a precautionary approach, including prioritizing monitoring efforts among other things, until sufficient information is collected to determine whether or not a bycatch problem exists. In the event of relevant new information becoming available, including for fisheries that are not believed to pose a risk at present, the framework may be used to review and (re)categorize the risk. ICES advises that sufficient information exists to identify fisheries where seabird bycatch is likely to be a problem (Table 1.5.1.3.1). Information to parameterize PBR is available for many seabird species/populations. ICES recommends that in cases where information is not available, suitable precautionary values are used, rather than investing heavily in improving e.g. knowledge on population abundance. It would be valuable to develop guidance for the use of various recovery factors (f), possibly by broad taxonomic groups or by status of population. A further issue is that insufficient effort data are reported and/or available in many fisheries either known or expected to have high bycatch levels. This applies particularly to smaller vessels; these can set relatively large quantities of static gear that can pose risk to birds. An improvement in the spatial and temporal resolution of effort reporting would be needed to avoid the risk of excessively precautionary advice. Outside this recommended framework, ICES is aware that there may be other factors contributing to the identification of a seabird bycatch problem. In particular, a nation may wish to undertake monitoring and/or mitigation measures based on a less biologically-based approach, such as one of “societal choice” regarding indiscriminate removal of birds and/or general wastefulness even where there may not be an immediate conservation risk. Any waste is unnecessary and waste reduction should be encouraged. 3. Data reporting format and database for seabird bycatch ICES advises that seabird bycatch information be stored in the database on bycatch of marine mammals, birds, turtles, and rare and/or endangered fish that is in a late stage of development within ICES. This ICES database was set up to collate and review data from the National reports required under Council Regulation (EC) No. 812/2004. In parallel, a standard reporting format was developed. While the standard reporting format was designed specifically for the tasks under Council Regulation (EC) No. 812/2004, it was designed in such a way that it can contain bycatch data on other groups than cetaceans. ICES advises the use of the standard reporting format defined for recording of marine mammal bycatch under Council Regulation (EC) No. 812/2004. The following adaptations are needed for reporting information on bycatch of seabirds: 1. 2. 3. References to Council Regulation (EC) No. 812/2004 are redundant and should be removed. In the section on acoustic deterrent devices, the two tables on acoustic deterrent devices are redundant. Any mitigation should be described and referred to here. In the section on observer schemes, the number of tables should be expanded to cover each of the gear types monitored (i.e. “Longlines” should be added). The column with the unit of effort would need to be altered depending on the gear monitored. The column “Days at Sea” should be retained in any additional table as this is the most flexible and widely used unit of effort. All EU Member States should report data in a standard electronic format to ensure easy import to the database. The ICES database is in the process of being made web-accessible. Sources FAO. 2008. Report of the Expert Consultation on Best Practice Technical Guidelines for IPOA/NPOA–Seabirds, Bergen, Norway, 2–5 September 2008. FAO Fisheries and Aquaculture Report, No. 880. FAO, Rome. 37 pp. ICES. 2013. Report of the Workshop to Review and Advise on Seabird Bycatch (WKBYCS). ICES CM 2013/ACOM:61. 79 pp. ICES Advice 2013, Book 1 47 1.5.2 EU DGENV 1.5.2.1 Special request, Advice June 2013 ECOREGION General advice SUBJECT Request from EU for Scientific advice on data collection issues Advice summary Review of existing indicators A summary of the qualities and the future utility of the existing Data Collection Framework (DCF) Annex XIII indicators is provided. ICES advises that data collection and assessment to support indicators 5, 6, and 7 should continue. For indicators 2, 3 and 4, ICES does not advise further implementation as there are challenges to target setting, and response times are slow and variable. Indicators 1 and 8 have little further utility as they either do not address the priority issues in the most effective way (indicator 1) or will shortly be superseded by legal or regulatory changes (indicator 8). New indicators In addition to development of existing DCF Annex XIII indicators 5, 6, and 7, ICES advises that new indicators are required to track and to guide the management of the effects of fisheries on the ecosystem and so aid in the implementation of the Marine Strategy Framework Directive (MSFD) through its descriptors. These indicators can be classified in the following manner: Removal 6 of protected and sensitive species (including bycatch of non-target species) (MSFD Descriptor 1) An indicator of fishing effects on Endangered, Threatened and Protected (hereafter “protected”) and sensitive species will be useful for the EU and its Member States to meet a number of policy and legislative targets. Information is lacking on the abundance of, and bycatch of, many protected and sensitive species. Coverage of fisheries under the DCF is biased away from those fisheries carrying the greatest risk of catching many protected and sensitive species. Development of remote electronic video recording seems likely to be a cost-effective way of assessing bycatch in the future as it can be applied to all parts of the fishing fleet (metiers and/or fleet segments) as defined in DC-MAP (Data Collection – Multi-Annual Plan). Foodweb effects (MSFD Descriptor 4) Stock assessments of all forage fish species that account for >5% of the total fish biomass, or that are important in the diet of dependent species (especially when these are protected species), are required. These assessments should take account of the distribution and availability of the forage species to dependent predators. This indicator will indicate whether sufficient prey are available for important predators in the foodweb. Among the indicators that can describe changes in foodwebs, forage fish abundance and distribution is one of the few that can respond in a defined way to a fishing activity and is relevant to Descriptor 4 of MSFD. Impacts on seafloor habitats and associated communities (Damage to the seafloor and its biological communities) (MSFD Descriptor 6) ICES recommends some changes to the existing pressure indicators addressing this issue (DCF Annex XIII indicators 5, 6, 7) to enhance their ability to assess impacts on seabed habitats. ICES advises that species indicative of seabed habitat type caught in surveys, and by commercial vessels with on-board observers, be recorded. As observers are already on some of these vessels, benthic data collection will be relatively costeffective. This will further provide links to MSFD criteria 6.1 and 6.2 (seafloor integrity) and 1.6 (biological diversity). ICES recommends that fishing positions of all vessels, including those less than 12 m, be recorded and reported at 30-minute intervals. 6 The word “removal” refers to extra mortality caused by fishing, including direct catch, bycatch and lethal interactions caused by collision with fishing gear. 48 ICES Advice 2013, Book 1 Request ICES is requested to assist in the identification of new data to be collected in support of the implementation of the Common Fisheries Policy (CFP) and the Marine Strategy Framework Directive (MSFD). ICES should also assist in the review of the existing environmental indicators to measure the effects of fisheries on the marine ecosystem (2010/93/EU, Appendix XIII) [question 1] and in the selection and development of new indicators to measure the impacts of fisheries on the marine ecosystem, including by-catch of non-target species, the food web and damage to the seafloor and its biological communities, for each MSFD marine region or sub-region and finally make proposals in time for the new DC-MAP 2014-2020 review [question 2]. ICES interpretation of the request is based on the understanding that it is from DG Environment and it is to explore the overlap area between CFP and MSFD data needs – and not to describe all data needs for CFP and MSFD. Therefore, some types of data that are not currently collected under the DCF but whose inclusion in the DC-MAP would be relevant are not included in this response as they are considered to be solely related to the CFP. Advice 7 Review of existing indicators (question 1) ICES assessed the capacity of the DCF (2010/93/EU) Appendix XIII indicators 1 to 8 to detect and measure the effects of fisheries on the marine ecosystem. Table 1.5.2.1.1 summarizes the future utility of each indicator. ICES notes that the existing DCF Annex XIII indicators were intended to track fishing effects on the ecosystem, and that targets cannot be set for all of these indicators. For this reason, ICES advises that if indicators are to be progressed to support MSFD it is a condition that targets can be set to determine (a) when measures to achieve Good Environmental Status (GES) have been established and (b) whether GES has subsequently been achieved. New indicators (question 2) ICES advises that two groups of indicators would be needed to measure the impacts of fisheries on the marine ecosystem: (1) Pressure indicators that are suitable in describing the impacts of fisheries (i.e. metier and/or fleet segment) on the marine ecosystem. It is important that data are collected and stored at the highest resolved metier and/or fleet segment in DC-MAP. (2) State and pressure indicators for which targets are set at the regional or sub-regional scale. The state indicators would be used to describe the state of the ecosystem in relation to targets (e.g. targets consistent with achieving GES in the MSFD). Corresponding pressure indicators would be used to define the levels of fishing pressure (e.g. as mortality rates, spatial distribution of fishing activity) that would need to be achieved to meet the targets for state. Indicators in this group would describe state in relation to targets for MSFD descriptors 1) biodiversity, 3) commercial fishes, 4) foodwebs, and 6) seafloor integrity, as these are directly affected by fishing. 7 In this advice, the use of ‘regions’ and ‘sub-regions’ is consistent with EC (2008). ICES Advice 2013, Book 1 49 Table 1.5.2.1.1 50 Advice on existing environmental indicators to measure the effects of fisheries on the marine ecosystem, in relation to utility for implementing MSFD. Codes in italics indicate potential relevance to MSFD descriptors and criteria. Low – recommend discontinuation within DC-MAP. Medium – keep for research but indicator not recommended for management purposes. High – develop indicator further; DC-MAP should be developed to ensure that these indicators are calculated and reported. Indicator Definition Future utility 1 Conservation status of fish species Indicator of biodiversity to be used for synthesizing, assessing, and reporting trends in the biodiversity of vulnerable fish species. Low; this indicator does not address pressure and state on the most sensitive species. 2 Proportion of large fish (D1.3, D4.2) Indicator for the proportion of large fish by weight in the assemblage, reflecting the size structure and life history composition of the fish community. Medium; this indicator has a long response time to the effects of fishing and the responses are variable. Although targets have been proposed they are not linked to a clear consequence or benefit and may be perceived as having low policy relevance. Even if a target is set, it cannot be used to guide management of specific metiers and/or fleet segments. 3 Mean maximum length of fishes (D1.3) Indicator for the life history composition of the fish community. Medium; this indicator has a long response time to the effects of fishing and the responses are variable. Although targets have been proposed they are not linked to a clear consequence or benefit and may be perceived as having low policy relevance. 4 Size at maturation of exploited fish species (D1.3) Indicator of the potential ‘genetic effects’ on a population. Low; targets cannot be set, trends not linked to a clear consequence or benefit. Management response to achieve targets not defined. 5 Distribution of fishing activities (D6.1) Indicator of the spatial extent of fishing activity. Reported in conjunction with ‘Aggregation of fishing activities’. 6 Aggregation of fishing activities (D6.1) Indicator of the extent to which fishing activity is aggregated. It would be reported in conjunction with the indicator for ‘Distribution of fishing activities’. 7 Areas not impacted by mobile bottom gears Indicator of the area of seabed that has not been impacted by mobile bottom fishing gears in the last year, computed for a series of bathymetric strata and potential substrate type. It responds to changes in the distribution of bottom fishing activity resulting from catch controls, effort controls, or technical measures (including MPAs established in support of conservation legislation) and to the development of any other human activities that displace fishing activity (e.g. wind farms). High. Methods exist for analysis. Spatial footprints can be mapped by metier. 8 Discarding rates of commercially exploited species (discarding can also include unwanted bycatch that is landed) Indicator of the rate of discarding of commercially exploited species in relation to landings. Low; ICES notes that future policy will be to avoid all discards of commercial species; there is thus no value in pursuing this indicator. ICES Advice 2013, Book 1 Removal of protected and sensitive species (including bycatch of non-target species) Knowledge of removal rate per unit effort, catch weight, or catch value will be required. To generate an indicator of the consequences of removal of protected and sensitive species, it is necessary to know or assess the number of animals killed in fisheries and the abundance of each species. ICES advises that reliable schemes to establish population abundance of animals likely to be affected by fisheries removal and the number of animals being caught should be established to measure population effects on protected and sensitive species and to set targets for acceptable rates of removal. ICES notes that it is a simple matter to generate a list of species protected under EU legislation. ICES suggests that a risk assessment should be undertaken to focus DC-MAP data collection on those species most likely to be adversely affected by fishing removal. Foodweb effects ICES advises that developing indicators of foodwebs is complex. While many indicators can describe changes, few respond in a defined way to a manageable pressure. There is an exception to this advice; large ‘forage’ fish stocks which provide important prey for other fishes, marine mammals, and seabirds. For these stocks, biomass and fishing mortality should be assessed in relation to reference points. ICES advises that when stock assessments of forage fish include estimates of natural mortality that incorporate top predators, the biomass limit reference points are then robust indicators of the impact of fisheries on the provision of forage fish for the foodweb. ICES notes that although most large populations of forage fish are currently assessed, some are not (for example sprats and sandeels to the west of the United Kingdom) as they are presently not exploited. As a guideline, where there are indications that these populations constitute more than ca.5% of the fish biomass in a region, data should be collected to allow these populations to be brought into the assessment process. A process will be required to identify these stocks. ICES advises that data be provided on the spatial structure of forage fish stocks and incorporated into stock assessments, to allow biomass limit reference points to be set that prevent local depletions of forage fish by fisheries that would impact on predators. Impacts on seafloor habitats and associated communities (Damage to the seafloor and its biological communities) ICES advises that the existing DCF Annex XIII indicators 5, 6, and 7 be extended to all metiers/fleet segments (this includes smaller vessels that are currently not included in Vessel Monitoring System (VMS) regulations) and that more frequent (30-minute) position updates be transmitted and recorded. ICES notes that data on the distribution of main indicator species of benthic habitats and substrate (including. biogenic habitats) could be identified to use in conjunction with the pressure indicators 5, 6, and 7. ICES advises that species indicative of seabed habitat type caught in surveys, and on-board commercial vessels with on-board observers, be recorded. This can be used to identify and prioritize for management gear/habitat interactions, and provide stronger links to MSFD criteria 6.1 and 6.2 (seafloor integrity) and 1.6 (habitat condition). Priorities for the collection of data on the ecosystem effects of fishing in DC-MAP If collection of data to allow calculation of all the recommended indicators cannot be resourced, then the relative priorities for the collection of data are: 1. Removal rates of protected species. 2. VMS data for all fleet segments and/or metiers. 3. Assessments of state of forage stocks. 4. Removal rates of sensitive species (defined following risk assessment). 5. Data on interactions between bottom fisheries and habitat. Basis of the advice Review of existing indicators The review of existing indicators to measure the impacts of fisheries on the marine ecosystem was based on: • An analysis of existing DCF Annex XIII indicators made (see Table 1.5.2.1.2) considering whether: ICES Advice 2013, Book 1 51 a. b. c. d. e. • the indicator has been tested or assessed, a target can be set for the indicator, the indicator is suitable for tracking fishing effects on the marine ecosystem, there are challenges obtaining data used to calculate the indicator, the indicator can be applied to all regions and sub-regions; A review of the impacts of fisheries on the marine ecosystem that were not addressed by existing indicators but would (a) indicate differences in the environmental impacts of fisheries and (b) indicate the extent to which fisheries management measures influenced progress towards potential or stated targets. Future assessment of DCF indicators For the existing DCF Annex XIII indicators ICES will be calculating and reporting time-series for the existing indicators this autumn (2013), recognising that four full years of data are now available for reporting. In conjunction with existing ICES analyses and reports (ICES, 2012a, 2012b) this process will provide further insight into the performance of these indicators and any constraints affecting their calculation and use. ICES will be making a data call in June 2013 to collate the data required for this analysis. 52 ICES Advice 2013, Book 1 Table 1.5.2.1.2 Review of existing DCF Annex XIII indicators. Codes in italics indicate potential relevance to MSFD descriptors and criteria. Has indicator been tested/assessed? Can target be set? Suitability for tracking fisheries impact? Challenges to data provision Generic or region specific Conservation status of fish species The indicator has been assessed by ICES (2012a, 2012b), in EC project reports (e.g. Borges et al., 2011) and in scientific literature (Dulvy et al., 2006). Targets have been proposed for purposes of investigating performance of the indicator but they are not explicitly linked to existing legislation. Captures trends in status of some fish species but does not address well the most sensitive species that are potentially at highest risk from fishing mortality (ICES 2012a, 2012b). Can be calculated from trawl survey data as collected to meet the requirements of the current DCF. Can be applied in any region where demersal trawl surveys are conducted. Proportion of large fish (D1.3, D4.2) The indicator has been comprehensively assessed by ICES (2012a, 2012b), EC projects Piet et al. (2011), Bloomfield et al. (2011), Borges et al. (2011) and in the scientific literature (e.g. Shephard et al., 2011; Greenstreet et al., 2011) Targets have been proposed but they are not explicitly linked to existing legislation or linked to a clear consequence or benefit. Captures trends in response of fish community to fishing. Principal concerns are slow and variable responses to changes in management, and technical issues with methods (ICES 2012a, 2012b). Can be calculated from trawl survey data as collected to meet the requirements of the current DCF. Can be applied in any region where demersal trawl surveys are conducted. The indicator has been assessed by ICES (2012a, 2012b) and in EC projects Piet et al. (2011), Bloomfield et al. (2011), and Borges et al. (2011). Targets have been proposed but they are not explicitly linked to existing legislation or linked to a clear consequence or benefit. Captures trends in response of fish community to fishing. Principal concerns are slow and variable responses to changes in management and technical issues with methods (ICES 2012a, 2012b). Can be calculated from trawl survey data as collected to meet the requirements of the current DCF. Can be applied in any region where demersal trawl surveys are conducted. Size at maturation of exploited fish species (D1.3) The indicator has been assessed by ICES (2012b). There is scientific uncertainty over whether genetic change is induced by fishing (e.g., Hutchings and Fraser (2008), Kuparinen and Merilä (2007)). Targets have not been proposed, indicator recommended to track trends in size at maturation. Unknown: performance not comprehensively assessed. Data to calculate this indicator are collected on the current DCF but for relatively few species (ICES, 2012b). Can be applied in any region where size at maturity is estimated. Distribution of fishing activities (D6.1) Further developed into two specific and operational indicators: (a) Total area fished and (b) Proportion of surface area fished. The indicator has been assessed by ICES (2012b). At present no target value exists. Only suitable for tracking fishing impact in relation to D6 if based on fishing metiers that actually disturb the seafloor. In that case (b) is the preferred indicator as this shows the proportion of the seafloor disturbed annually. Confidentiality issues often prevent access to VMS data in a format to calculate the indicators at the appropriate spatial and temporal scale. Not all fisheries have VMS. This indicator can be calculated for any area. They can even be calculated for a specific habitat (potential relevance for D1) if data exist. Indicator 1 2 3 4 Mean maximum length of fishes (D1.3) 5 ICES Advice 2013, Book 1 53 Indicator Aggregation of fishing activities (D6.1) 6 Areas not impacted by mobile bottom gears 7 8 54 Discarding rates of commercially exploited species (discarding can also include unwanted bycatch that is landed) Has indicator been tested/assessed? Further developed into two specific and operational indicators, i.e. (a) Proportion of surface area fished by specific proportion of effort, and (b) Proportion of surface area fished at specific intensity. Indicator has been assessed by ICES (2012b). Further developed into two specific and operational indicators, i.e. (a) Cumulative proportion of surface area not impacted over a specific time period, and (b) Proportion of surface area not impacted at specific level of confidence. Indicator has been assessed by ICES (2012b). Not tested systematically though discard rates are used in some parts of ICES advice. Can target be set? At present no target value exists. Suitability for tracking fisheries impact? Only suitable for tracking fishing impact in relation to D6 if based on fishing metiers that actually disturb the seafloor. In that case (b) is the preferred indicator, showing the proportion of the seafloor fished more than once a year. Generic or region specific Challenges to data provision Confidentiality issues often prevent access to VMS data in a format to calculate the indicators at the appropriate spatial and temporal scale. Not all fisheries have VMS. This indicator can be calculated for any area. It can even be calculated for a specific habitat (potential relevance for D1) if data exist. At present no target value exists. This indicator was intended to show the impact on the seafloor and is therefore the most suitable. In this case (a) is the preferred indicator showing, the proportion of the seafloor not impacted over a period long enough to assume it is recovered even if disturbed prior to this period. As such this proportion is by definition in Good Environmental Status (GES) if not impacted by other human activities. Confidentiality issues often prevent access to VMS data in a format to calculate the indicators at the appropriate spatial and temporal scale. Not all fisheries have VMS. The absence of VMS coverage for vessels under 12 m is of particular importance in inshore and coastal areas. For potential relevance to D1 benthic bycatch and substrate should be reported. This indicator can be calculated for any specific area. It can even be calculated for a specific habitat (potential relevance for D1) if data exist. At present no target value exists This tracks additional mortality on commercial fish stocks, and is necessary to understand total pressure on a stock No major challenges to data provision but relies on unbiased sampling of fisheries. This indicator can be calculated for any specific area. ICES Advice 2013, Book 1 New indicators ICES advises that three new classes of indicators should be supported by DC-MAP. These are: ‘removal of protected and sensitive species’, ‘foodweb effects’, and ‘impact on seafloor habitats and associated communities’. Table 1.5.2.1.3 summarizes the new proposals for indicators. These new indicators are general rather than technical specifications of the indicator. ICES recommends that when/if indicators are carried forward, detailed specifications must be prepared and analytical methods supported by common analytical tools. Table 1.5.2.1.3 Issue Removal rates of protected and sensitive species Foodweb effects Impacts on seafloor habitats and associated communities Proposed indicators to address the three issues identified in the advice, the relevance of these indicators to the MSFD and their use (comparing state and pressure in relation to reference points). MSFD relevance D1(D4) D4.3 D6.1 Indicator(s) Frequency Usage Removal rates of protected species in relation to population size by region and sub-region. Removal rates of sensitive species in relation to population size by region and sub-region. Fishing mortality rate on forage fish stocks expressed in relation to reference points. Spawning-stock biomass of forage fish stocks expressed in relation to reference points. Spatial distribution of forage fish stocks expressed in relation to reference points. Distribution of fishing activities (based on DCF Annex XIII indicator 5) by fleet segment/metiers, expressed in relation to fisheries effort, catch rate, and catch value. Aggregation of fishing activities (based on DCF Annex XIII indicator 6) by fleet segment/metiers, expressed in relation to fisheries effort, catch rate, and catch value. Areas not impacted by mobile bottom gears (based on DCF Annex XIII indicator 7) by fleet segment/metiers at small scales and for combined fleets at regional and sub-regional gears. Calculated annually. Calculated annually. Comparison of removal rates of protected species with reference points. Comparison of removal rates of sensitive species with reference points. Comparison of mortality on forage fish stocks with reference points. Calculated annually. Comparison of status of forage fish stocks with reference points. Calculated annually. Comparison of distribution of forage fish stocks with reference points. Calculated annually. Comparison of relative effects of different fleet segments/metiers on seabed habitats. No reference point. Calculated annually. Comparison of relative effects of different fleet segments/metiers on seabed habitats. No reference point. Calculated annually. Assessment of extent of areas unimpacted by towed bottom gears. No reference point. Calculated annually. Use of DC-MAP data and indicators to support MSFD The MSFD provides a clear context for indicator development because the CFP is required to be used as the primary instrument to manage the impacts of fisheries on the marine ecosystem to the extent necessary to achieve GES. For the DC-MAP, ICES considers that the priority in relation to this request is to provide the data needed to report indicators for impacts of fisheries on the marine ecosystem that pose the greatest risk and are most likely to be unsustainable. ICES assumes that DC-MAP will be implemented through the EU Member States at a sub-regional, fishery, or fleet level, whereas the MSFD targets are the responsibility of the EU Member States alone. Whilst the EU Member States are encouraged to work together to define MSFD sub-regional targets, the setting of targets could be done in isolation from other regional EU Member States. This contrasts with the expected approach in DC-MAP. ICES notes that indicators to measure the impacts of fisheries on the marine ecosystem that are based on data collected in DC-MAP are most valuable if there is sufficient consistency in the implementation of MSFD among EU Member States, ensuring that all EU Member States in a given region or sub-region can use DC-MAP data and associated indicators. Alternatively, DC-MAP indicators would need to be developed to take account of the intentions of EU Member States in a given region or sub-region. ICES notes that surveys supported by DC-MAP also provide EU Member States with the opportunity to use the surveys as platforms of opportunity, collecting data that describe the impacts of fishing on marine ecosystems that may be relevant nationally. ICES Advice 2012, Book 1 55 Removal of protected and sensitive species (including bycatch of non-target species) Many species are caught that are not targeted. The indicators for fish will include some non-target species, but the main species of fish that may be adversely affected by bycatch are large species with low reproductive rates such as sharks and rays. Species that are included in the formal ICES fish stock assessment process need not be included in this group of indicators, but all others should be. Other non-target species that are affected include mammals, seabirds, and turtles. The bycatch of mammals and turtles should be monitored under the EU Habitats Directive (92/43/EEC); bycatch of cetaceans is also covered by Council (EC) Regulation No. 812/2004. In 2012 the Commission adopted an Action Plan to reduce incidental catches of seabirds in fishing gears; this plan includes requirements for data collection. An equivalent Action Plan for sharks was adopted in 2009. At present, the bycatch of some species is reported by observers, working under DCF, on-board vessels. The forthcoming ‘discards ban’ may affect future monitoring and data collection. It could, for example, result in much greater emphasis on port-based sampling schemes, rather than sea-going observer schemes. It seems likely that bycatch of protected species will continue to be returned to the sea as they are “non-commercial” and unlicensed possession of some of these species (especially cetaceans) is illegal in most EU Member States. The proposed indicator is the removal rate of all species protected by legislation and of all sensitive species. Removal rates would be reported as numbers and sizes of individuals caught and expressed in relation to fishing effort and/or catch weight and/or catch value in the metier and/or fleet segment. To identify the species that will be monitored in all regions and sub-regions a list of these species should be compiled for each region and sub-region. ICES recommends the application of a risk assessment process (e.g. as reviewed by ICES, 2012b) that considers the sensitivity of the species (to removal) in the first instance, followed by an assessment, if possible, of its exposure to fishing. The process should consider all fishing operations in all regions, stratified by the most highly resolved level of fleet classification used to collect data for fisheries management in DC-MAP. In conjunction with information on the population size of protected and sensitive species, the removal rates could be used to assess impacts on the populations in relation to reference points. Limit removal rates for some species have been agreed in some political settings (e.g. for harbour porpoise by the regional conservation agreement for small cetaceans, ASCOBANS), and could potentially be set, for other populations, and the values of indicators in relation to targets can be used to report on progress towards meeting existing commitments and adopt appropriate measures to achieve GES for MSFD Descriptor Criterion 1.3. Collection of reliable information on removal rates of sensitive and protected species is most commonly undertaken by on-board human observers. Sampling under the current Data Collection Framework (DCF) tends to focus on the metiers that discard the most fish; these are not necessarily the same metiers that have the largest catch of species of interest. Thus, bottom trawling is generally well sampled, while in some specific fishing areas set nets, longlines, and purse-seines are undersampled. Some EU Member States have undertaken additional observation schemes to meet the requirements of Council Regulation No. 812/2004 and those of the Habitats Directive (92/43/EEC). It would be possible to better define requirements on EU Member States under the DCF, but much will depend on how other data collection requirements will change under the revised Common Fisheries Policy (CFP). Alternatives to monitoring by human observers on-board vessels could include remote electronic video recording and monitoring from vessels visiting a fishing fleet. Of these alternatives remote electronic video recording seems to have the greatest potential to meet many of the needs of the existing DCF and also to improve monitoring of bycatch of noncommercial species, and has the advantage of being useable on metiers and/or fleet segments where the carriage of a human observer poses logistical problems. Foodweb effects ICES considers that when stock assessments of forage fish include estimates of natural mortality that incorporate top predators, the biomass limit reference points from the MSY approach are then robust indicators of the impact of fisheries on the provision of forage fish for the foodweb. When a stock assessment does not incorporate realistic estimates of predator-induced mortality, then MSY limit reference points may not be appropriate, and some alternative mechanism for ensuring forage fish biomass for predators may need to be found. Some large populations of forage fish are not assessed, primarily because they are not exploited to any great extent. In this situation, assessments of the populations should be carried out if the populations are thought to be of a significant size (>5% of the total fish biomass) based on catches, bird food needs, acoustic surveys, ichythoplankton surveys, and other analytical methods. The choice of 5% is arbitrary and offered as provisional guidance. 56 ICES Advice 2013, Book 1 Impacts on seafloor habitats and associated communities (Damage to the seafloor and its biological communities) VMS data are currently collected for assessing compliance with the CFP as detailed in the ‘Control Regulation’ (EC, 2009). In this advice, modifications to VMS collection procedures and greater VMS data exchange are proposed to improve the existing DCF Annex XIII indicators and to support interpretation of new state indicators. ICES recognises that changes in the collection of VMS data may not be affected through DC-MAP, but considers that indicators based on VMS data will be necessary to assess the effects of fishing on marine ecosystems in support of the MSFD and other legislation. Calculation of the VMS indicators will require that VMS data for all metiers and/or fleet segments included in DC-MAP are available from EU Member States. To facilitate exchange and allow calculation of the indicators for all metiers, ICES recommends that data are shared by EU Member States, using gridded data that record the number of VMS fishing records by month and metier and/or fleet segment. ICES recognises that grids can introduce some small biases when indicators are calculated with VMS data allocated to fixed grids (e.g. Piet and Hintzen, 2010; Gerritsen et al., 2013), but the exchange of data will rely on providing individual vessels with sufficient anonymity to support full international exchange of all VMS data. ICES recommends the sharing of gridded point data at 0.05 resolution, as already trialled in EC projects (Bloomfield et al., 2011). Metiers and/or fleet segments to which position records are allocated should be determined by the fisheries management requirements of the DC-MAP. ICES recommends that the fishing positions of all vessels are recorded and reported at 30-minute intervals. The indicators are calculated for all metiers and/or fleet segments fishing with towed bottom gears, but VMS coverage will be required for all other vessels to track their interactions with the environment. ICES recommends that this should include all fishing vessels in all waters in regions and sub-regions, including smaller and inshore vessels not currently monitored with VMS. Recording benthic bycatch and substrate (including biogenic habitats) will enhance the interpretation of these indicators. They can then be used to identify and prioritize for management gear/habitat interactions and extend the value of these pressure indicators to provide direct links to MSFD criteria 6.1 and 6.2 (seafloor integrity) and 1.6 (habitat condition) (ICES, 2012a). Sources Bloomfield, H., Nolan, C., Le Quesne, W. J. F., Raakjær, J., Christensen, A. S., Aanesen, M., Armstrong, C., Piet, G. J., and Frid, C. 2011. Fisheries Ecosystem Plan: North Western Waters. Making the European Fisheries Ecosystem Plan Operation (MEFEPO): Work Package 7 Report. Borges, M. F., Mendes, H., Bloomfield, H. J., Raakaer, J., Pinho, M. R., Duchêne, J., Porteiro, C., Velasco, F., Hilly, C., Aanesen, A., Armstrong, C., Piet, G. J., and Frid, C. L. J 2011. Fisheries Ecosystem Plan: South Western Waters. Making the European Fisheries Ecosystem Plan Operational (MEFEPO): Work Package 7 Report. Dulvy, N. K., Jennings, S., Rogers, S. I., and Maxwell, D. L. 2006. Threat and decline in fishes: an indicator of marine biodiversity. Canadian Journal of Fisheries and Aquatic Science, 63:1267–1275. EC. 2008. Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 establishing a framework for community action in the field of marine environmental policy (Marine Strategy Framework Directive). Official Journal of the European Union, L164: 19–40. EC. 2009. Council Regulation (EC) No. 1224/ 2009 of 20 November 2009 establishing a Community control system for ensuring compliance with the rules of the Common Fisheries Policy, amending Regulations (EC) No. 847/96, (EC) No 2371/2002, (EC) No. 811/2004, (EC) No. 768/2005, (EC) No. 2115/2005, (EC) No. 2166/2005, (EC) No. 388/2006, (EC) No. 509/2007, (EC) No. 676/2007, (EC) No. 1098/2007, (EC) No. 1300/2008, (EC) No. 1342/2008 and repealing Regulations (EEC) No. 2847/93, (EC) No. 1627/94 and (EC) No.1966/2006. Official Journal of the European Union, L343: 1–50. EC. 2010. Commission decision of 1 September 2010 on criteria and methodological standards on good environmental status of marine waters. Official Journal of the European Union, L232: 14–24. Gerritsen, H. D., Minto, C., and Lordan, C. 2013. How much of the seabed is impacted by mobile fishing gear? Absolute estimates from Vessel Monitoring System (VMS) point data. ICES Journal of Marine Science, 70: 523–531. Greenstreet, S. P. R., Rogers, S. I., Rice, J. C., Piet, G. J., Guirey, E. J., Fraser, H. M., and Fryer, R. J. 2011. Development of the EcoQO for the North Sea fish community. ICES Journal of Marine Science, 68:1–11. Hutchings, J. A., and Fraser, D. J. 2008. The nature of fisheries- and farming-induced evolution. Molecular Ecology, 17: 294–313. ICES. 2012a. Marine Strategy Framework Directive – Descriptor 3+. ICES CM 2012/ACOM:62. ICES. 2012b. Report of the Working Group on the Ecosystem Effects of Fishing Activities (WGECO). ICES CM 2012/ACOM:26. ICES Advice 2012, Book 1 57 Kuparinen, A., and Merilä, J. 2007. Detecting and managing fisheries-induced evolution. Trends in Ecology and Evolution, 22: 652–659. Piet, G. J., Bloomfield, H. J., Rockmann, C., Miller, D., van Hal, R., Raakjær, J., Christensen, A. S., Aanesen, M., Armstrong, C., and Frid, C. L. J. 2011. Fisheries Ecosystem Plan: North Sea. Making the European Fisheries Ecosystem Plan Operation (MEFEPO): Work Package 7 Report. Piet, G. J., and Hintzen, N. T. 2012. Indicators of fishing pressure and seafloor integrity. ICES Journal of Marine Science, 69: 1850–1858. Shephard, S, Reid, D. G., and Greenstreet, S. P. R. 2011. Interpreting the large fish indicator for the Celtic Sea. ICES Journal of Marine Science, 68: 1963–1972. 58 ICES Advice 2013, Book 1 1.5.2.2 Special request, Advice November 2013 ECOREGION SUBJECT General advice Request from EU for scientific advice on data collection issues – part 2 Advice summary ICES provides an inventory (in the form of annexed tables) of the main bycatch issues for fisheries in each of Europe’s Data Collection Framework (DCF) regions (Baltic, Atlantic, Mediterranean, Black Sea). An assessment is made as to how well the current DCF meets the need to understand these fishery interactions in the context of the EU’s Marine Strategy Framework Directive (MSFD). The detailed design of catch and bycatch observation schemes should integrate the needs for data both for fish stock assessment and for ecosystem assessment purposes. ICES advises that detailed planning, dependent on decisions at policy level, will be required to integrate all these data collection needs into existing survey protocols while ensuring that existing monitoring commitments are not compromised. The design of on-board observer schemes, whether carried out by humans or by remote technology (e.g. closed-circuit television CCTV) will also need care, and will need to take account of likely future changes in human observer coverage as a consequence of the ‘landings obligation’ being introduced. Detailed data on fishing effort will be required for any assessment of the effect of fishing. Data requirements are of higher temporal and spatial resolution and differ from those currently used for fish stock assessments. Further research and additional survey effort (including additional surveys) will be needed to interpret several of the DCF and MSFD indicators, especially if they are to be used in management. Request According to the MoU between ICES and the European Commission, ICES shall provide further scientific advice on data collection issues. ICES is therefore requested to assist in the identification of new data to be collected in support of the implementation of the Common Fisheries Policy (CFP) and the Marine Strategy Framework Directive. Indicators for impacts on the ecosystem from fisheries can contribute to assessments for MSFD Descriptors 1, 4 and 6. As such they would need to be integrated with assessments of non-fishery impacts in order to provide an overall assessment for these descriptors in each (sub-) region. This will require discussion with Member States (via the Regional Sea Conventions) on how to incorporate such indicators. ICES should provide recommendations on how this can be included in the MSFD assessment and reporting process, as well as the implementation of the EU Biodiversity Strategy, including the periodic assessment of the data and access to the data and assessments. ICES interpretation of this request is based on the understanding that the DG Environment wants to consider how further data could be collected via the DCF/DC-MAP that would also be useful in the implementation of the MSFD and the EU Biodiversity Strategy. The European Commission clarified the request by suggesting a simplified scope to address the request in a logical sequence: (a) (b) (c) Produce an inventory of the main fisheries per region: Identify the main bycatch issues per fishery (initially whether it is birds, mammals, reptiles, noncommercial fish or benthos); Coarsely quantify which fisheries are the main threats to each of these groups – some sort of high-level prioritisation; i.e. bird bycatch is an important issue for fishery X and Y in region Z). Assess how well current DCF addresses the main threats – to indicate where current DCF is OK and hence priorities for revision. Consider what methods for data collection (e.g. observers, use of CCTV etc.), sampling regime and periodicity would lead to the collection of data suitable for an indicator (Note there may be several options and costs might vary considerably, depending on links to observer systems for commercial stocks etc.) – it may be too early to pin down precisely the best methods. ICES provided advice on other parts of the request from DG ENV (not quoted above) earlier in 2013: http://www.ices.dk/sites/pub/Publication%20Reports/Advice/2013/Special%20requests/EU_%20data_%20collection_is sues.pdf. ICES Advice 2012, Book 1 59 ICES advice Inventory of main fisheries and their bycatch Summary overviews of the main fisheries (metiér level 4) for each of the Data Collection Framework regions and their potential for bycatch have been made for the vertebrate groups birds, mammals, reptiles, elasmobranchs, and bony fish (Annex 1). The first three of these groups include all species that occur in the respective region, while the overviews for fish are based on the threatened and declining fish species listed by HELCOM, OSPAR, and the Barcelona Convention. Roughly half of the listed species are sampled for biological variables in commercial fisheries under the DCF. Most elasmobranch species are sampled. Non-target and non-commercial fish are taken as bycatch in many fisheries, notably by trawls, nets, and seines. This includes many sharks and rays among elasmobranchs, and also many species of bony fish. Species of marine mammals are taken as bycatch in some fisheries, including by pelagic trawls and nets, as well as in longline fisheries in the Mediterranean Sea. Seabirds are taken in some fisheries, notably in nets and longline fisheries. Reptiles are taken as bycatch in net and trawl fisheries in the Mediterranean Sea. Summary overviews of the potential to physically impact benthic habitats are provided for the same fisheries (Annex 2). Habitats chosen for this evaluation are relevant to each DCF region. The degree to which each habitat is affected by bottom gears depends on the scale of the fishing operations and the sensitivity of the habitat. Bottom trawls generally affect the largest areas and have the potential to cause most damage to sensitive habitats. Pelagic trawls on the other hand do not generally come in contact with the seafloor and have no impacts. Set nets, longlines, seines, and traps have more spatially limited contact with the seafloor and generally smaller and more local effects compared to bottom trawls. There is limited information on the potential impacts from some gears on benthic habitats, as reflected by question marks in the tables. It should be noted that these overview assessments represent a wide span in real or potential impacts or threats, from low levels that might be assessed as insignificant, up to substantial and serious threats. Some impacts may be local while others will be widespread. If these assessments are to be used in management or targeting of studies/research, it will be necessary to carry out more detailed assessments in order to scale the impacts or threats for the various combinations of fishing activities and species or habitats. Because of this, ICES is not able to directly quantify all impacts. Current DCF sampling in relation to main threats There are gaps in the bycatch data for marine mammals, seabirds, reptiles, and of species indicative of benthic habitats, because there is no obligation in the current DCF to record this information. The gaps identified for monitoring the status of threatened and declining fish species are for coastal and inshore (some anadromous) listed species, pelagic sharks, and demersal deep-water species south of ICES Subarea VI. There is currently a gap in information on the spatial distribution of fishing activities by vessels under 12 m (in some regions, for example the Mediterranean, VMS data from fishing vessels of less than 15 m in length are not available). Data collection methods and recommendations The DCF requires a sampling programme of commercial fisheries that includes landings on shore and total catches (including discards) at sea. Sampling effort is stratified according to the relative contribution of a particular metiér to overall landings, value of landings, and fishing effort. As a consequence, this sampling effort may not be adequate to provide sufficient data on fisheries that have a particularly high bycatch of certain protected, threatened, or declining species. The detailed design of catch and bycatch observation schemes should integrate the needs for data both for fish stock assessment and for ecosystem assessment purposes. Monitoring should be based on statistically sound sampling schemes and will need a number of inputs, including a detailed set of quality targets and some knowledge of the required precision for indicators. Guidance on the desired precision, development, and provision of quality metrics of collected data would make it possible to advise on sampling intensity and the stratification of sampling. Guidance on prioritizing differing needs for time and personnel resources will also be needed. These are societal, not scientific, judgements that require the input of policy-makers. 60 ICES Advice 2013, Book 1 There are several general principles to follow when establishing an observer scheme (regardless of method of observation). • • • • Vessels with gears that apparently cause most unwanted bycatch should have highest sampling, based on a riskbased approach. If possible, vessels should be selected randomly (ideally working with the industry, perhaps through fishing federations). Relevant parameters of the fishing operation should be collected, such as technical features of the fishing gear, location, frequency of the fishing activities, etc. Power analyses can be used to estimate and stratify studies; it may not be necessary to observe every ship every year to estimate the amount of bycatch per gear. Additional information on the state of threatened and/or declining species and habitats is needed. The use of existing monitoring programmes may not be sufficient to get adequate information on many species and habitats. Recording the (by-)catch of these species by observers or scientific surveys might fill an information gap, as long as it is a limited list of readily recognisable species (including epifauna as indicators for their associated threatened and declining habitats) that can provide valuable information on, e.g. distribution of these species and habitats. The use of observation schemes on commercial vessels may not be sufficient to determine the state or condition of benthic species and habitats. This is due to the variation in catchability between benthic species, and our lack of detailed knowledge of this variation. A better assessment may be made based on models linking data on the location of fishing activities and their effects on benthic habitats. However, if an observer is on board and benthic species are bycaught, photos can be made for determination. This information could also be used to improve the quality of modelling. Increasing the scope of observer programmes and of CCTV observations of bycatch and discards need to be considered under the perspective likely decisions of the EU to introduce landing obligations. Assuming that there is compliance with the landing obligation, it is likely that observations of catch (and commercial species bycatch) could switch from being predominantly at sea to being predominantly on land due to cost–benefit reasons. This would reduce the potential for atsea observations of other features such as bycatch by human observers. When using CCTV cameras on board vessels, the following issues will need to be addressed. • • • • A catch (and bycatch) sampling scheme will still be required to confirm some information (e.g. species identity, age). Compulsory CCTV schemes, and schemes that recompense or incentivize fishers are more likely to be successful than voluntary schemes. Protocols for the analysis of CCTV records will need to be developed. Additional requirements for recording in logbooks (e.g. for rare species, species listed under the Habitats and Birds Directives, or under the Regional Sea Conventions) could be required when CCTV is in use, in order to compensate for the loss of records from on-board observers. The association of position (from integration of electronic logbook to GPS) could provide spatial details on the area where species were caught. The technical feasibility of such an approach might be experimentally tested on portions of the fleets. Species known to be indicative of seabed habitat type, caught in surveys and on-board commercial vessels with on-board observers, should be recorded. This information can be used to identify and prioritize the management of gear/habitat interactions and to provide stronger links to implementation by EU Member States working through the Regional Sea Commissions of MSFD criteria 6.1 and 6.2 (seafloor integrity) and 1.6 (habitat condition). The inclusion of benthic taxa to be assessed by on-board observation may need the development of guidelines and the availability of proper taxonomic expertise. Assessing the surface and sub-surface disturbance by bottom contact gears for the proposed MSFD indicators (of HELCOM and OSPAR) requires data on the extent of physical damage and loss of seabed habitats resulting from human activities. Fishing activity information therefore needs to be provided on the spatial distribution of fishing, including: location (maximum 30-minute position updates, preferably shorter), gear type (Levels 4 and5), gear width, vessel speed, and whether fishing is occurring or not. Effort should be recorded in appropriate units (e.g. hours towed, km of net × soak time, number of hooks set). ICES Advice 2012, Book 1 61 To better interpret the results from monitoring under DCF, ICES offers the following further advice. a) Additional research is required to better understand the ‘fishing pressure – benthic state’ relationship and to develop predictive tools for this means. b) The spatial distribution of most habitat features is generally only known at a coarse scale. If impacts are to be assessed at a finer scale, further research and survey will be required (in some cases modelling may be adequate). c) Equally, knowledge of the population abundance and the spatial and temporal distribution of many species is inadequate. Dedicated surveys are likely to be required to remedy this situation. d) If indicators are to be used to inform management, then it will be important to distinguish between effects caused by human activities and those that may be described as natural (e.g. hydrographic change). This may require data collection beyond that needed to assess the state of the indicators. Annually collected DCF data and Regional Seas indicators on fisheries impacts could contribute to the overall assessment and reporting for D1, 4, and 6 of the MSFD. Integrated assessment Data collected under DCF in the period 2014–2020 can be used to (further) develop, test, and evaluate the Regional Seas indicators. Information from adopted indicators can be included in the reporting process of the Regional Sea Commissions and included in Regional Sea Commission Assessments, leading to a common report for each (sub)region of the MSFD assessment. This will support Member States in reporting for the assessment required in 2018 under the MSFD. Furthermore, the interim data of DCF may be used for future state of the environment of the Regional Sea Commissions (such as the OSPAR Quality Status Report in 2021 and future HELCOM reports on the Ecosystem Health of the Baltic Sea). Integrated assessment (IA, or integrated ecosystem assessment, IEA) is seen as a core element of the ecosystem approach to management, being the step where information from monitoring is compiled and evaluated as a basis for scientific advice on management measures. ICES advises that IA should be used as a mechanism for integrating information collected for the various purposes under the CFP and MSFD, including indicators for impact of fisheries on other components of the ecosystem than commercial fish. ICES has established working groups for integrated assessment for some of the MSFD regions, including the Baltic Sea and the North Sea. These working groups can contribute to the overall holistic assessments of the status and trends of the regional ecosystems supporting the reporting by EU member states according to MSFD (and other) requirements. Sources Anderson, O. R. J., Small, C. J., Croxall, J. P., Dunn, E. K., Sullivan, B. J., Yates, O., and Black, A. 2011. Global seabird bycatch in longline fisheries. Endangered Species Research, 14: 91–106. Bazairi, H., Ben Haj, S., Boero, F., Cebrian, D., De Juan, S., Limam, A., Lleonart, J., Torchia, G., and Rais, C. (Eds.) 2010. The Mediterranean Sea Biodiversity: state of the ecosystems, pressures, impacts and future priorities. UNEPMAP RAC/SPA, Tunis. 100 pp. Casale, P. 2011. Sea turtle by-catch in the Mediterranean. Fish and Fisheries, 12: 299–316. Cebrian Menchero, D. 2010. Mitigation measures needed for reducing by-catch of seabirds in the Mediterranean region. General Fisheries Commission for the Mediterranean. Meetings of the GFCM Subsidiary Committees. SCMEE. 21st November–3rd December 2010. Cosgrove, R., and Browne, D. 2007. Cetacean by-catch rates in Irish gillnet fisheries in the Celtic Sea. Bord Iascaigh Mhara. Marine Technical Report, June 2007. GFCM. 2008. Report of the meeting of the ByCBAMS projecT (jointly between ACCOBAMS and SCMEE). Rome, Italy, 17–18 September 2008. General Fishery Commission for the Mediterranean. Scientific Advisory Committee. ICES. 2013a. Report of the Working Group on Bycatch of Protected Species (WGBYC), 4–8 February 2013, Copenhagen, Denmark. ICES CM 2013/ACOM:27. 73 pp. ICES. 2013b. Report of the Workshop on Data Collection – Assessments of non-fishery impacts (WKDCF-NF), 8–10 October 2013, Copenhagen, Denmark. ICES CM 2013/ACOM:74. 65 pp. OSPAR. 2010. Quality Status Report 2010. OSPAR Commission, London. Tudela, S. 2004. Ecosystem effect of fishing in the Mediterranean. General Fisheries Commission for the Mediterranean. Studies and Reviews, N74. 44 pp. Zydelis, R., Small, C., and French, G. 2013. The incidental catch of seabirds in gillnet fisheries: A global review. Biological Conservation, 162: 76–88. 62 ICES Advice 2013, Book 1 Annex 1 Potential bycatch of vertebrate groups in fishing metiérs for the four DCF regions NA (not applicable) – the vertebrate group does not occur; N (no) – the fishing metiér does not impact the vertebrate group (or bycatch is minimal); Y (yes) – a real or potential impact on the vertebrate group; ? – the fishing metiér and vertebrate group are likely to overlap in space and time but with insufficient information to make a judgement. Colours of fishing gear cells symbolize level of activity: orange – common fishing gear/activity in this ecoregion; green – negligible or non-existent; white – not known. Table 1.5.2.2.1 Baltic region (based on Anderson et al., 2011; Cosgrove and Browne, 2007; ICES, 2013a; Zydelis et al., 2013). Threatened and declining Gear groups Bottom trawls Gear type Birds Mammals Reptiles Elasmobranch Bony fish Bottom otter trawl [OTB] Y Y NA Y Y Multi-rig otter trawl [OTT] ? ? NA Y Y Bottom pair trawl [PTB] ? ? NA Y Y Midwater otter trawl [OTM] ? Y NA N Y Midwater pair trawl [PTM] ? Y NA N Y Rods and Lines Hand and Pole lines [LHP] [LHM] N N NA Y N Longlines Drifting longlines [LLD] ? N NA ? N Set longlines [LLS] ? N NA ? N Pots and Traps [FPO] N Y NA N N Fykenets [FYK] N Y NA N N Stationary uncovered poundnets [FPN] N Y NA N N Trammelnet [GTR] Y Y NA Y Y Set gillnet [GNS] Y Y NA Y Y Surrounding nets Purse-seine [PS] ? N NA ? Y Seines Fly-shooting seine [SSC] N N NA ? Y Anchored seine [SDN] N N NA ? Y Pair-seine [SPR] N N NA ? Y Beach and boat seine [SB] [SV] N N NA ? ? Yes N NA ? Y Pelagic trawls Traps Nets Recreational fisheries ICES Advice 2012, Book 1 63 Table 2 North Sea (based on Anderson et al., 2011; Cosgrove and Browne, 2007; ICES, 2013a; Zydelis et al., 2013). Threatened and declining Gear groups Dredges Gear type Birds Mammals Reptiles Elasmobranch Bony fish Boat dredge [DRB] N N NA N Y Mechanized / Suction dredge [HMD] N N NA N N Bottom otter trawl [OTB] Y N NA Y Y Multi-rig otter trawl [OTT] ? N NA Y Y Bottom pair trawl [PTB] ? N NA Y Y Beam trawl [TBB] N N NA Y Y Midwater otter trawl [OTM] N Y NA N N Midwater pair trawl [PTM] N Y NA N N Rods and Lines Hand and Pole lines [LHP] [LHM] N N NA N N Longlines Set longlines [LLS] ? N NA N N Traps Pots and Traps [FPO] N Y NA N N Fykenets [FYK] N N NA ? ? Trammelnet [GTR] Y Y NA Y Y Set gillnet [GNS] Y Y NA Y Y Driftnet [GND] Y ? NA N N Surrounding nets Purse-seine [PS] N N NA Y Y Seines Fly-shooting seine [SSC] N N NA Y Y Anchored seine [SDN] N N NA Y Y Pair-seine [SPR] N N NA Y Y Beach and boat seine [SB] [SV] N N NA N Y N N NA Y N Bottom trawls Pelagic trawls Nets Recreational fisheries 64 ICES Advice 2013, Book 1 Table 1.5.2.2.3 Atlantic region (based on Anderson et al., 2011; Cosgrove and Browne, 2007; ICES, 2013a; Zydelis et al., 2013). Threatened and declining Gear groups Dredges Gear type Birds Mammals Reptiles Elasmobranch Bony fish Boat dredge [DRB] N N N N Y Mechanized / Suction dredge [HMD] N N N N N Bottom otter trawl [OTB] Y N ? Y Y Multi-rig otter trawl [OTT] ? N N Y Y Bottom pair trawl [PTB] ? N N Y Y Beam trawl [TBB] ? N N Y Y Midwater otter trawl [OTM] N Y N Y Y Midwater pair trawl [PTM] N Y N Y Y Hand and Pole lines [LHP] [LHM] N N N N N Trolling lines [LTL] N Y N N N Drifting longlines [LLD] Y N N N N Set longlines [LLS] Y N N N N Pots and Traps [FPO] N Y Y N N Fykenets [FYK] N N N N N Stationary uncovered poundnets [FPN] N N N N N Trammelnet [GTR] Y Y ? Y Y Set gillnet [GNS] Y Y Y Y Y Driftnet [GND] Y Y Y Y N Surrounding nets Purse-seine [PS] Y Y N N Y Seines Fly-shooting seine [SSC] N N N Y Y Anchored seine [SDN] N N N Y Y Pair-seine [SPR] N N N Y Y Beach and boat seine [SB] [SV] N Y N N N N N N N N Bottom trawls Pelagic trawls Rods and Lines Longlines Traps Nets Recreational fisheries ICES Advice 2012, Book 1 65 Table 1.5.2.2.4 Mediterranean and Black Sea region (based on Casale, 2011; Cebrian Menchero, 2010; GFCM, 2008; ICES, 2013a; Tudela, 2004). Threatened and declining Gear groups Gear type Birds Mammals Reptiles Elasmobranch Bony fish Dredges Boat dredge [DRB] N N N N ? Bottom trawls Bottom otter trawl [OTB] Y Y Y Y Y Multi-rig otter trawl [OTT] ? ? Y Y Y Bottom pair trawl [PTB] ? ? Y Y Y Beam trawl [TBB] N N N Y Y Midwater otter trawl [OTM] ? ? Y Y Y Pelagic pair trawl [PTM] ? Y Y Y Y Hand and Pole lines [LHP] [LHM] N ? ? ? ? Trolling lines [LTL] Y ? ? ? ? Drifting longlines [LLD] Y Y Y Y Y Set longlines [LLS] Y Y Y Y Y Pots and Traps [FPO] N Y N N ? Fykenets [FYK] ? ? ? ? Stationary uncovered poundnets [FPN] ? ? Y ? Trammelnet [GTR] Y Y Y Y Y Set gillnet [GNS] Y Y Y Y Y Driftnet [GND] Y Y Y Y Y Purse-seine [PS] ? Y Y ? Y Lampara nets [LA] ? ? ? ? Y Fly-shooting seine [SSC] N N ? N Y Anchored seine [SDN] N N ? N Y Pair-seine [SPR] N N ? N Y Beach and boat seine [SB] [SV] N N ? ? ? ? Y ? Y ? Pelagic trawls Rods and Lines Longlines Traps Nets Surrounding nets Seines Recreational fisheries Y Y . 66 ICES Advice 2013, Book 1 Annex 2 Tables of fishing metiér and their potential to impact relevant benthic habitats NA (not applicable) – the fishing metiér is not employed or would not contact the benthic habitat; N (no) – the fishing metiér does contact, but does not impact, the benthic habitat; Y (yes) – a real or potential impact on the benthic habitat; ? – the fishing metiér and benthic habitat are likely to overlap in space and time, but with insufficient information to make a judgement. For the Mediterranean and Black Sea, the NA and N categories are combined as NA. Colours of fishing gear cells symbolize level of activity: orange – common fishing gear/activity in this ecoregion; green – negligible or nonexistent; white – not known. Table 1.5.2.2.5 Baltic Sea region Gear group Gear type Seagrasses Mud Sea pens Sand Gravel Muddy gravel Mixed sediments Kelp forests Bivalve reefs (Modiolus, Musculus, Ostrea) Bottom trawls Bottom otter trawl [OTB] NA Y NA Y Y Y Y Y NA Multi-rig otter trawl [OTT] Bottom pair trawl [PTB] NA Y NA Y Y Y Y Y NA NA Y NA Y Y Y Y Y NA Midwater otter trawl [OTM] Midwater pair trawl [PTM] Hand and Pole lines [LHP] [LHM] Drifting longlines [LLD] Set longlines [LLS] NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA N N N N N N N N N NA NA NA NA NA NA NA NA NA Y ? ? N ? ? ? Y ? Pots and Traps [FPO] Y ? ? N ? ? ? Y ? Fykenets [FYK] Y ? ? N ? ? ? Y ? Stationary uncovered poundnets [FPN] Trammelnet [GTR] NA NA NA NA NA NA NA NA NA Y ? ? N ? ? ? Y ? Set gillnet [GNS] Y ? ? N ? ? ? Y ? Surrounding nets Purse-seine [PS] ? ? ? ? ? ? ? ? ? Seines Fly-shooting seine [SSC] Y ? ? N ? ? ? Y ? Anchored seine [SDN] Y ? ? N ? ? ? Y ? Pair-seine [SPR] Y ? ? N ? ? ? Y ? Beach and boat seine [SB] [SV] Recreational fisheries Y ? ? N ? ? ? Y ? ? N ? N N N N Y Y Pelagic trawls Rods and Lines Longlines Traps Nets ICES Advice 2013, Book 1 67 Table 1.5.2.2.6 North Sea Gear groups Gear type Hard biogeni c reefs (Lophe lia, carbon ate mound s, maerl, etc.) Sabe llari a reefs Seagrass es Mu d Sea pen s Sa nd Gra vel Mudd y grave l Mixe d sedi ment s Kel p fore sts Circa littora l reefs incl. coral garde ns, spong es Bivalv e reefs (Modio lus, Muscul us, Ostrea ) Deepwater spong es Dredges Boat dredge [DRB] Y ? ? Y Y Y Y Y Y Y NA Y Y Mechanized / Suction dredge [HMD] NA NA NA NA ? Y NA NA Y NA NA NA NA Bottom otter trawl [OTB]1 Y Y NA Y Y Y Y Y Y Y Y Y Y Multi-rig otter trawl [OTT] Y Y NA Y Y Y Y Y Y Y Y Y Y Bottom pair trawl [PTB] Y Y NA Y Y Y Y Y Y Y Y Y Y Beam trawl [TBB] Y Y NA Y Y Y Y Y Y Y Y Y Y Midwater otter trawl [OTM] NA NA NA NA NA N A NA NA NA NA NA NA NA Midwater pair trawl [PTM] NA NA NA NA NA N A NA NA NA NA NA NA NA Rods and Lines Hand and Pole lines [LHP] [LHM] N N N N N N N N N N N N N Longlines Set longlines [LLS] Y Y Y ? ? N ? ? ? ? Y ? Y Traps Pots and Traps [FPO] Y Y Y ? ? N ? ? ? Y Y ? Y Fykenets [FYK] NA NA N ? NA N N N ? NA NA ? NA Trammelnet [GTR] Y Y Y ? ? N ? ? ? Y Y ? Y Set gillnet [GNS] Y Y Y ? ? N ? ? ? Y Y ? Y Driftnet [GND] N N N N N N N N N N N N N Surrounding nets Purse-seine [PS] N N N ? ? ? ? ? ? N N N N Seines Fly-shooting seine [SSC] Y Y Y ? ? N ? ? ? Y Y ? Y Anchored seine [SDN] Pair-seine [SPR] Y Y Y ? ? N ? ? ? Y Y ? Y Y Y Y ? ? N ? ? ? Y Y ? Y Beach and boat seine [SB] [SV] Y Y Y ? ? N ? ? ? Y Y ? Y Y ? ? N ? N N N N ? Y Y NA Bottom trawls Pelagic trawls Nets Recreational fisheries 68 ICES Advice 2013, Book 1 Table 1.5.2.2.7 Atlantic region Gear group Gear type Dredges Boat dredge [DRB] Bivalv e reefs (Modi olus, Muscu lus, Ostrea ) Deep water spong es NA Circ alittor al reefs incl. coral gard ens NA Y ? Y NA NA NA NA Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA NA N N N N N N N N N N N N NA NA NA NA NA NA NA NA NA NA NA NA NA Drifting longlines [LLD] NA NA NA NA NA NA NA NA NA NA NA NA NA Set longlines [LLS] Y Y Y ? ? ? ? ? ? Y Y ? Y Pots and Traps [FPO] Y Y Y ? ? ? ? ? ? Y Y ? Y Fykenets [FYK] NA NA N ? NA N N N ? NA NA ? NA Stationary uncovered poundnets [FPN] NA NA NA NA NA NA NA NA NA NA NA NA NA Trammelnet [GTR] Y Y NA Y Y Y Y Y Y Y Y Y Y Set gillnet [GNS] Y Y ? Y Y Y Y Y Y Y Y Y Y Driftnet [GND] N N N N N N N N N N N N N Surrounding nets Purse-seine [PS] N N N ? ? ? ? ? ? N N N N Seines Fly-shooting seine [SSC] Y Y Y ? ? N ? ? ? Y Y ? NA Anchored seine [SDN] Y Y Y ? ? N ? ? ? Y Y ? NA Pair-seine [SPR] Y Y Y ? ? N ? ? ? Y Y ? NA Beach and boat seine [SB] [SV] Y Y Y ? ? N ? ? ? Y Y ? NA Y ? ? N ? N N N N Y Y Y NA Bottom trawls Pelagic trawls Rods and Lines Longlines Traps Nets Hard biogeni c reefs (Lophe lia, carbon ate mound s Y Sabel laria reefs Sea gras ses Mu d Sea pen s San d Gra vel Mudd y grave l Mixe d sedim ents Kel p fore sts Y Y Y Y Y Y Y Y Mechanized / Suction dredge [HMD] N N N N N Y ? Y Bottom otter trawl [OTB] Y Y NA Y Y Y Y Multi-rig otter trawl [OTT] Y Y NA Y Y Y Bottom pair trawl [PTB] Y Y NA Y Y Beam trawl [TBB] Y Y NA Y Midwater otter trawl [OTM] NA NA NA Midwater pair trawl [PTM] NA NA Hand and Pole lines [LHP] [LHM] N Trolling lines [LTL] Recreational fisheries ICES Advice 2013, Book 1 69 Table 1.5.2.2.8 Gear groups Mediterranean and Black Sea (based on Tudela, 2004; Bazairi et al., 2010). Gear type INFRALITTORAL CIRCALITTORAL BATHYAL ABYSSA L Fine muddy -sands Coarse sands in very shallo w waters Stones and pebbles (incl. biogenic reefs: maerl, Rhodolites , etc.) Posidoni a oceanica meadows Hard Mud beds s and rock s Sand s Hard beds and rocks (coralligenous ) Mud s Sand s hard rocks (incl. deepsea corals ) Muds Dredges Boat dredge [DRB] Y NA ? NA NA Y Y Y NA NA NA NA Bottom trawls Bottom otter trawl [OTB] Y NA Y Y NA Y Y Y Y Y Y NA Multi-rig otter trawl [OTT] Y NA ? Y NA Y Y ? Y Y Y NA Bottom pair Y trawl [PTB] NA ? Y NA Y Y ? Y Y Y NA Beam trawl [TBB] Y NA ? NA NA Y Y Y NA NA NA NA Midwater otter trawl [OTM] NA NA NA NA NA NA NA NA NA NA NA NA Pelagic pair NA trawl [PTM] NA NA NA NA NA NA NA NA NA NA NA Hand and Pole lines [LHP] [LHM] NA NA NA NA NA NA NA NA NA NA NA NA Trolling lines [LTL] NA NA NA NA NA NA NA NA NA NA NA NA Drifting longlines [LLD] NA NA NA NA NA NA NA NA NA NA NA NA Set longlines [LLS] ? ? ? ? ? ? ? ? NA NA NA NA Pots and Traps [FPO] Y ? Y ? Y Y Y Y NA NA NA NA Fykenets [FYK] Y ? Y ? ? Y Y Y NA NA NA NA Stationary uncovered poundnets [FPN] ? ? ? ? ? ? ? ? NA NA NA NA Trammelne t [GTR] Y Y Y Y Y Y Y Y NA NA NA NA Set gillnet [GNS] Y Y Y Y Y Y Y Y NA NA NA NA Driftnet [GND] NA NA NA NA NA NA NA NA NA NA NA NA Purse-seine [PS] NA NA NA NA NA NA NA NA NA NA NA NA Lampara nets [LA] NA NA NA NA NA NA NA NA NA NA NA NA ? Flyshooting seine [SSC] ? ? ? ? ? ? ? NA NA NA NA ? ? ? ? ? ? ? ? NA NA NA NA Pelagic trawls Rods and Lines Longlines Traps (d) Nets Surrounding nets Seines Anchored seine [SDN] 70 ICES Advice 2013, Book 1 Gear groups Gear type INFRALITTORAL CIRCALITTORAL BATHYAL ABYSSA L Fine muddy -sands Coarse sands in very shallo w waters Stones and pebbles (incl. biogenic reefs: maerl, Rhodolites , etc.) Posidoni a oceanica meadows Hard Mud beds s and rock s Sand s Hard beds and rocks (coralligenous ) Mud s Sand s hard rocks (incl. deepsea corals ) Muds Pair-seine [SPR] ? ? ? ? ? ? ? ? NA NA NA NA Beach and boat seine [SB] [SV] ? ? NA Y ? NA NA NA NA NA NA NA ? ? ? ? Y ? ? ? NA NA NA NA Recreational fisheries ICES Advice 2013, Book 1 71 1.5.3 HELCOM No advice has been requested for 2013 1.5.4 NASCO The advice provided in response to special requests from the North Atlantic Salmon Conservation Organisation (NASCO) can be found in Book 10 of the ICES Advice 2013 Report. 72 ICES Advice 2013, Book 1 1.5.5 NEAFC 1.5.5.1 ECOREGION SUBJECT Special request, Advice June 2013 General advice Vulnerable deep-water habitats in the NEAFC Regulatory Area Advice summary ICES advises that new data are available on the presence of VMEs in the NEAFC Regulatory Area. ICES advises NEAFC that new closures to bottom fisheries or revision of existing closure boundaries in the following five areas would enable the protection of these VMEs. • • • • • Northwest Rockall Bank; Southwest Rockall Bank; Hatton–Rockall Basin; Hatton Bank; Josephine Seamount. Request NEAFC requests ICES to continue to provide all available new information on distribution of vulnerable habitats in the NEAFC Convention Area and fisheries activities in and in the vicinity of such habitats. ICES advice New information on the location of vulnerable habitats in the NEAFC Regulatory Area New data that indicate the presence of vulnerable habitats (i.e. VMEs) were submitted to ICES in 2013. Some of these data are in the NEAFC Regulatory Area (NEAFC RA). Five areas are detailed below which contain information on new findings of VMEs, and potential closure boundaries are advised. ICES has no new information (i.e. VMS data) available on actual fishing activity in these areas at the spatial resolution required to evaluate the pressure on the habitats. 1) Northwest Rockall Bank Rockall Bank is a large plateau that lies partly in EU waters and partly in international waters regulated by NEAFC. An area in the northwestern part of Rockall Bank has been closed to bottom fishing since 2007 for the protection of VMEs. In 2012, ICES advised a boundary modification to the area based on new information. Six further video transects within or close to the Northwest Rockall Bank closure were completed in 2012 (Figure 1.5.5.1.1). The video footage revealed extensive patches of coral reefs in the centre and toward the southern end (that which lies within the NEAFC RA) of the current closure. In part of the proposed eastern extension to the closure to bottom fishing advised by ICES in 2012, a video transect revealed new observations of coral reefs, reaffirming the appropriateness of this extension. No VMEs were observed in a transect to the north of the closure. ICES reiterates its advice from 2012 to extend the closure. The boundary of this closure is detailed in Figure 1.5.5.1.1 and Table 1.5.5.1.1. ICES Advice 2013, Book 1 73 Figure 1.5.5.1.1 74 Map of NW Rockall Bank showing locations of video transects and new findings of VMEs. ICES Advice 2013, Book 1 Table 1.5.5.1.1 P OINT N UMBER 2) Coordinates of points for recommended closure to bottom fishing in Northwest Rockall Bank. L ATITUDE (N) (DEGREES MINUTES S ECONDS) L ONGITUDE (W) (D EGREES MINUTES S ECONDS) L ATITUDE ( DECIMAL ) L ONGITUDE ( DECIMAL ) 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 58 02 49.20 57 51 35.92 57 47 50.42 57 43 22.15 57 37 15.49 57 42 33.62 57 49 48.97 57 56 05.67 57 53 37.50 57 50 05.13 57 45 18.43 57 28 59.98 57 22 00.01 56 55 59.98 56 55 59.98 57 00 00.00 57 37 00.01 57 50 15.79 57 50 42.00 57 59 35.30 58 09 29.55 58 13 05.91 58 13 43.32 58 12 14.22 13 22 25.96 13 07 30.14 13 02 59.42 13 02 17.37 13 14 55.75 13 16 28.56 13 23 09.02 13 43 26.11 13 52 28.16 13 56 22.56 14 08 24.00 14 19 00.01 14 19 00.01 14 36 00.00 14 51 00.00 14 52 59.98 14 42 00.00 14 28 44.22 14 28 25.86 14 23 11.18 14 03 48.85 13 53 17.88 13 49 41.37 13 43 52.32 58.04700 57.85998 57.79734 57.72282 57.62097 57.70934 57.83027 57.93491 57.89375 57.83476 57.75512 57.48333 57.36667 56.93333 56.93333 57.00000 57.61667 57.83772 57.84500 57.99314 58.15821 58.21831 58.22870 58.20395 -13.37388 -13.12504 -13.04984 -13.03816 -13.24882 -13.27460 -13.38584 -13.72392 -13.87449 -13.93960 -14.14000 -14.31667 -14.31667 -14.60000 -14.85000 -14.88333 -14.70000 -14.47895 -14.47385 -14.38644 -14.06357 -13.88830 -13.82816 -13.73120 25 58 07 11.71 13 34 29.10 58.11992 -13.57475 Southwest Rockall Bank In the southwestern section of Rockall Bank a closure to bottom fisheries has been in effect since 2007 to protect VMEs. New data were available from three towed video transects in 2012 (Figure 1.5.5.1.2). Coral reefs were recorded, as were extensive stretches of seabed without coral reefs. Most of the coral reefs recorded were inside the currently closed area, but some were observed outside the currently closed area. If NEAFC wishes to protect these VMEs, ICES advises NEAFC: 1) to extend the boundary of the closure in the western corner to incorporate the new video records. 2) to extend the boundary in the southern corner to encompass new video records and the 2012 bycatch records. Boundaries recommended by ICES have been drawn to include a buffer zone of three times the water depth. In the northern part of the Southwest Rockall Bank closed area there is now some video evidence that coral reefs are not present in an area where they have previously been reported. Should further research (including analysis of fishing data) demonstrate the absence of VMEs, this area may be considered for reopening to bottom fishing in the future. ICES Advice 2013, Book 1 75 Figure 1.5.5.1.2 Southwest Rockall Bank bottom fishing closure boundary modifications. The geographic coordinates for the two closure extensions are provided in Tables 1.5.5.1.2 and 1.5.5.1.3. 76 ICES Advice 2013, Book 1 Table 1.5.5.1.2 Geographic coordinates for the Southwest Rockall Bank western closure extension. L ATITUDE ( DECIMAL DEGREES) L ONGITIDE ( DECIMAL DEGREES) L ATITUDE (DMS) L ONGITUDE (DMS) 55.9694 55.9706 -16.2196 -16.0427 55° 58' 10" N 55° 58' 14" N 16° 13' 11" W 16° 2' 34" W 55.9144 -16.0925 55° 54' 52" N 16° 5' 33" W Table 1.5.5.1.3 Geographic coordinates for the Southwest Rockall Bank southern corner closure extension. L ATITUDE ( DECIMAL DEGREES) L ONGITIDE ( DECIMAL DEGREES) L ATITUDE (DMS) L ONGITUDE (DMS) 55.9310 55.8500 55.7977 -15.6806 -15.6167 -15.8968 55° 55' 52" N 55° 51' 0" N 55° 47' 52" N 15° 40' 50" W 15° 37' 0" W 15° 53' 48" W 55.8215 -15.9399 55° 49' 17" N 15° 56' 24" W 3) Hatton–Rockall Basin The Hatton-Rockall basin is an expanse of deep-water sedimentary habitat of depths between 1100–1500 m. In a research survey in 2012, a benthic sampling net contained at least two undescribed species of a genus of chemosymbiotic clams. To date, species from this genus have only been found in association with active cold seeps/gas hydrate ecosystems. This is the first indication of an active cold seep ecosystem at the Hatton–Rockall Basin. Cold seeps are considered to be VMEs under the FAO guidelines and since the site is at approximately 1200 m depth, it is vulnerable to bottom contact fishing. Estimates from the Ship’s log and sensors attached to the net give an approximate position with a radial uncertainty of around 3 km (Figure 1.5.5.1.3). Should NEAFC wish to protect this VME, ICES advises a closure to bottom fisheries centred around the circle in Figure 1.5.5.1.3 with a buffer zone of 2400 m (twice water depth), corresponding to the coordinates in Table 1.5.5.1.4. Future research should be able to provide a more precise location and estimate of the extent of the ecosystem, at which point the appropriateness of the closure boundary can be assessed. Figure 1.5.5.1.3 Approximate location of chemosynthetic clams found within the Hatton–Rockall Basin. ICES Advice 2013, Book 1 77 Table 1.5.5.1.4 Geographic coordinates for the advised closure. POINT DECIMAL DECIMAL LATITUDE LONGITUDE 1 58.0025 -15.4538 2 58.0025 -15.6377 3 57.90317 -15.4538 4 57.90317 -15.6377 Howell et al. (2013) observed aggregations of sponges in visual surveys in the centre of the Hatton–Rockall Basin at approximately 1150 m water depth. Should NEAFC want to protect these VMEs, ICES advises a closure to bottom fishing for the area shown in Figure 1.5.5.1.4, with the geographic coordinates provided in Table 1.5.5.1.5. Figure 1.5.5.1.4 Location of sponge aggregations at the Hatton–Rockall Basin with the advised closure boundary. Table 1.5.5.1.5 Geographic coordinates for the advised Hatton–Rockall Basin closure. L ATITUDE ( DECIMAL DEGREES) L ONGITUDE ( DECIMAL DEGREES) L ATITUDE (DMS) L ONGITUDE (DMS) 58.1840 58.2166 58.1832 -16.5190 -16.4205 -16.3624 58° 11' 2" N 58° 12' 60" N 58° 10' 59" N 16° 31' 8" W 16° 25' 14" W 16° 21' 45" W 58.1348 -16.4511 58° 8' 5" N 16° 27' 4" W 4) Hatton Bank This year no new VME data were available for the Hatton Bank area, but new information in graphic format was provided on Spanish fishing activity for the period 2000–2011 (Figure 1.5.5.1.5). This clearly demonstrates where the fishing activity occurs and highlights the southwestern slopes of the Hatton Bank as areas where VMEs are unlikely to be present due to intensive trawling. Therefore ICES considers that Area 3 in Figure 1.5.5.1.5 requires modifications (a reduction in the VME closed area) to reflect this. ICES assessed the significance of bycatch levels of seapens against the NAFO threshold levels and determined that the records of seapens on the southwestern slopes of Hatton Bank are insufficient to describe the area as a VME. Based on the information on where Spanish vessels fish and existing information on VMEs in the area, ICES advises that two areas, one to the southeast (Figure 1.5.5.1.6 and Table 1.5.5.1.6) and one to the southwest (Figure 1.5.5.1.6 and Table 1.5.5.1.7) of Hatton Bank would be suitable for closure to bottom fishing, should NEAFC wish to protect these VMEs. The closure boundaries are based on: i. ii. 78 The presence of carbonate mounds which classify as VME elements (area to the southwest of Hatton Bank); The presence of large bycatch (exceeding 400 kg) of sponges in the east (area to the southeast of Hatton Bank); ICES Advice 2013, Book 1 iii. iv. v. vi. vii. The presence of small bycatch of gorgonians in the area (area to the southwest of Hatton Bank); A ‘knoll’ area (VME element) to the southwest of Hatton Bank, visible from the bathymetric data and records of gorgonians from the summit (< 1000 m); Two areas of outcropped rock (VME elements) on the western slope, evident from multibeam data; Evidence of fishing (trawling) activity in the sedimentary areas (Hatton Drift) of the western slope (see Figure 1.5.5.1.5); All VME indicator records outside the advised closed area are below the threshold used to determine the presence of VMEs by NAFO or the current thresholds adopted by NEAFC. Figure 1.5.5.1.5 Fishery footprint for the Spanish bottom-trawl fishery (period 2000–2011), based on VMS data provided by the Spanish Government (modified from Durán Muñoz et al., 2012). Grey patches and points are VMS data (speed = 2–4 knots); Black crosses are absence of VME indicator taxa records in the bycatch; Triangles are records of cold-water corals pooled; Circles are records of sponges pooled; Squares are carbonate mounds (VME records from ICES database). The NEAFC closed area is represented by the purple polygons. An area advised for closure by ICES in 2012 is indicated by the black polygon. A multibeam bathymetry map provided by the Instituto Español de Oceanografía (IEO) showing the western slope of the bank is also presented. ICES Advice 2013, Book 1 79 Figure 1.5.5.1.6 The two areas of Hatton Bank proposed to be closed to bottom fishing, one to the southeast (blue boundary) and one to the southwest (yellow boundary). Where the boundary was drawn around VME indicator species a buffer of twice the depth is included. Table 1.5.5.1.6 Geographic coordinates for the advised closure extension to the southeast of Hatton Bank. L ATITUDE ( DECIMAL DEGREES) L ONGITUDE ( DECIMAL DEGREES) L ATITUDE (DMS) L ONGITUDE (DMS) 57.8626 57.9167 58.0500 57.8850 -18.0978 -17.5000 -17.5000 -16.9388 57° 51' 45" N 57° 54' 60" N 58° 3' 0" N 57° 53' 6" N 18° 5' 52" W 17° 29' 60" W 17° 30' 0" W 16° 56' 20" W 57.5851 -18.0335 57° 35' 6" N 18° 2' 0" W 80 ICES Advice 2013, Book 1 Table 1.5.5.1.7 Geographic coordinates for the advised closure extension to the soutwest of Hatton Bank. L ATITUDE ( DECIMAL DEGREES) L ONGITUDE ( DECIMAL DEGREES) L ATITUDE (DMS) L ONGITUDE (DMS) 57.9993 57.7500 57.8345 57.5188 57.2348 57.0368 56.8853 56.8370 56.7780 57.0007 57.1718 57.5445 -19.0842 -19.2500 -18.3970 -18.3547 -19.4738 -19.4588 -19.4828 -19.5604 -19.8954 -20.0704 -19.9207 -19.8773 57° 59' 58" N 57° 44' 60" N 57° 50' 4" N 57° 31' 8" N 57° 14' 5" N 57° 2' 12" N 56° 53' 7" N 56° 50' 13" N 56° 46' 41" N 57° 0' 2" N 57° 10' 18" N 57° 32' 40" N 19° 5' 3" W 19° 15' 0" W 18° 23' 49" W 18° 21' 17" W 19° 28' 26" W 19° 27' 32" W 19° 28' 58" W 19° 33' 37" W 19° 53' 43" W 20° 4' 13" W 19° 55' 15" W 19° 52' 38" W 57.7780 -19.6310 57° 46' 45" N 19° 37' 46" W 5) Josephine Seamount Josephine Seamount is one of six seamounts included in a cluster just over 200 nm north of the Island of Madeira. Josephine Seamount is classed by NEAFC as ‘an existing bottom fishing area’. In 2011, OSPAR designated the Josephine Seamount (and the five other seamounts in the immediate vicinity) as an MPA on the basis of information that included VME indicator species such as hexactinellid sponges and gorgonians (OSPAR, 2011). In 2012, ICES presented further historical evidence showing the presence of gorgonians on and around the Josephine Seamount. Taken together, these data indicate that there is a high likelihood that Josephine and the surrounding seamounts have VMEs. ICES did not have access to recent data on fishing activity for this area and thus cannot directly assess the risk of impact to these VMEs. However, in view of the present status of the Josephine Seamount MPA as an ‘existing fishing area’, ICES considers that there is a risk of significant adverse impacts to VMEs in the area from bottom fishing gears. Should NEAFC wish to protect these VMEs, ICES advises a closure to bottom fisheries, the boundary of which should correspond to the Josephine Seamount High Seas MPA established by OSPAR (OSPAR Decision 2010/5) (Figure 1.5.5.1.7 and Table 1.5.5.1.8). Such a closure would encompass the seamount, the documented locations of recent VME indicator records, and the five other nearby seamounts that are within the NEAFC RA. As a consequence of enclosing the seamounts in one protective management measure, some surrounding deep areas of high topographic relief (and thus likely to contain VMEs) would also be protected from potential impacts. Figure 1.5.5.1.7 Table 1.5.5.1.8 Map of Josephine Seamount showing the distribution of gorgonian corals and the proposed bottom fishing closure that corresponds precisely with the OSPAR High Seas MPA. The red square in the overview map shows the approximate location of the closure. Geographic coordinates for the proposed Josephine Seamount NEAFC bottom fishing closure. ICES Advice 2013, Book 1 81 L ATITUDE ( DECIMAL DEGREES) L ONGITUDE ( DECIMAL DEGREES) L ATITUDE (DMS) L ONGITUDE (DMS) 37.460 37.630 36.860 36.180 36.450 -14.650 -13.750 -13.420 -14.450 -15.390 37° 27' 36" N 37° 37' 48" N 36° 51' 36" N 36° 10' 48" N 36° 27' 0" N 14° 39' 0" W 13° 45' 0" W 13° 25' 12" W 14° 26' 60" W 15° 23' 24" W 36.760 -15.720 36° 45' 36" N 15° 43' 12" W ICES recommendation NEAFC has requested ICES to provide all available new information on fisheries activities in, and in the vicinity of, VME habitats. ICES cannot address this request further without data on fishing activities at high spatial resolution. ICES therefore recommends that all recent VMS data in the NEAFC RA are made available. This data should include information on gear type and vessel speed in order to define the impact on the VMEs by bottom trawling and by longlines. Sources Durán Muñoz, P., Sacau, M., and Sayago-Gil, M., on behalf of the ECOVUL/ARPA Team. 2012. The EU’s experience in the protection of cold-water corals in the high seas: The Hatton Bank (NEAFC Regulatory Area) – a case study. NEAFC–PECMAS symposium on its bottom fisheries. North East Atlantic Fisheries Commission. London, 25 June 2012. Howell, K. L., Huvenne, V., Piechaud, N., Roberts, K., and Ross, R. 2013. JC060 Data Analysis: Comprising data from Darwin Mounds (NE Rockall Trough), Rockall–Hatton Basin Polygonal Faults, NW Rockall Bank, and E Rockall Bank Escarpment. Report to the Joint Nature Conservation Committee. Unpublished. ICES 2013. Report of the ICES/NAFOJoint Working Group on Deep-water Ecology (WGDEC). ICES CM 2013/ACOM: 28. OSPAR. 2011. Background document on the Josephine Seamount Marine Protected Area. 82 ICES Advice 2013, Book 1 1.5.5.2 Special request, Advice June 2013 ECOREGION SUBJECT General advice Evaluation of the appropriateness of buffer zones Advice summary Both the VME location accuracy and a buffer zone are considered when delineating the closure boundary around VMEs. ICES is confident that the buffer zone considerations used to define the boundaries around the area closures are appropriate and therefore adequate for the protection of VMEs. A schematic diagram of the approach to generate buffer zones is presented. The buffer zones will always be included in ICES advice and will be illustrated where appropriate to the scale of the closure. Request ICES is requested to evaluate whether buffer zones applied in the current bottom fishing closures are appropriate. Additionally, ICES is requested to include, specify and illustrate buffer zones in its future advice on closures in the Regulatory Area as appropriate. Advice Two different considerations are used to delineate buffer zones around VMEs; one is linked to the VME location accuracy, the other to setting a buffer zone around the VME location (Figure 1.5.5.2.1). Example 1 Example 2 Example 3 Estimated location of VME Trawl track VME area Buffer zone Figure 1.5.5.2.1 Three conceptual examples of the two considerations for delineating buffer zones around VMEs, applied to three theoretical examples of VME closures. Example 1: isolated VME detection with low geospatial certainty (e.g. trawl track); Example 2: isolated VME detection with high geospatial certainty (e.g. ROV observation); and Example 3: area identified as hosting a VME. Consideration 1. VME location accuracy The data used by ICES to assess the likelihood of VME presence consists of mainly point records of species (Figure 1.5.5.2.1). While recognising this is the best available data, there are varying levels of spatial uncertainty associated with the records, ranging from trawl bycatch with low spatial accuracy (Example 1) to dynamically positioned ROV observations with high spatial accuracy (Example 2) and areas identified as hosting VMEs (Example 3). This uncertainty in VME location is dealt with by outlining the minimum boundary that encompasses the records. In the case of records derived from trawling, the deviation perpendicular to the track is considered negligible relative to the length of the track and is not taken into account in the VME location. While spatial accuracy of the position of VMEs has improved over time, there are still a high number of records where the location accuracy is unknown. In such cases a simple buffer is applied (see Consideration 2). Consideration 2. Buffer zone around VME location ICES considers a buffer zone to be a spatial margin of assurance around the VME to avoid adverse impact (Figure 1.5.5.2.1). The spatial extension of the buffer zone may vary and is based on the following: • The potential for fishing gear to stray into the VME is related to the uncertainty of the location of the fishing gear relative to the known location of the vessel. This will be a function of water depth and the trawl warp length deployed. In deep-water trawling, the typical warp length deployed decreases with water depth, from around 3:1 ICES Advice 2013, Book 1 83 • • at 200 m to 2:1 at 500 m and more. For VMEs that occur on flat or undulating seabed a buffer zone of approximately two (>500 m depth) or three times (< 500 m depth) the local depth is advised. In the case of VMEs on very steep slopes, the risk of straying of bottom trawls is mitigated by the fishers’ own incentive to avoid the steep slopes and cliff edges, in which case the buffer zone may be reduced. In some cases the presence of geomorphological features are used to define boundaries for closures on the basis that they are considered to be VME elements, in which case the VME reflects the topographic relief of the VME element without a buffer zone. As both the VME location accuracy and a buffer zone are considered when advising on a closure boundary around VMEs, ICES is confident that the buffer zone considerations used to extend closures beyond the immediate estimated position of a VME are appropriate and therefore adequate for the protection of VMEs. The buffer zone approach described here does not take into account any issues specifically related to enforcement. Source ICES 2013. Report of the ICES/NAFO Joint Working Group on Deep-water Ecology (WGDEC). ICES CM 2013/ACOM: 28. 84 ICES Advice 2013, Book 1 1.5.5.3 ECOREGION SUBJECT Special request, Advice June 2013 General advice Assessment of the list of VME indicator species and elements Advice summary 1) Taxa should be considered by habitat type and/or at the taxonomic level of family rather than listing all the likely species that would be indicators of VMEs in the NEAFC Regulatory Area (RA). A table of relevant families and habitat types is provided. 2) Three maps of VME elements in the NEAFC RA at depths less than 2000 m, including the Mid-Atlantic Ridge, the Rockall–Hatton area, and isolated seamounts, are provided. Canyon-like features and steep flanks and slopes are known only from the Mid-Atlantic Ridge and the Rockall–Hatton area. There is not sufficient information to map all knolls. 3) There are only five known or inferred vent sites within the NEAFC RA. The vent sites, other than the one inferred site on the Reykjanes Ridge, are below 2000 m depth and are not likely to be impacted by fishing activities. It is advised that the site on the Reykjanes Ridge be closed to bottom-contacting gear. Request 1) ICES is requested to assess whether the list of VME indicator species is exhaustive and suggest possible addition to that list. The basis for the assessment should be the FAO Guidelines specifying taxa and habitats that may be relevant. ICES should focus on taxa (species or assemblages of species) that tend to form dense aggregations of assumed particular functional significance. NAFO SC has in 2012 conducted a similar assessment and revision and to the extent scientifically valid harmonization with NAFO lists would be beneficial. 2) ICES is furthermore asked to map VME elements (i.e. geomorphological features) in the NEAFC RA. This would include seamounts and knolls at fishable depths (with summits shallower than 2000m), canyons, and steep flanks. Also in this exercise, harmonization with NAFO SC evaluations would be beneficial. 3) ICES is specifically requested to advise NEAFC on the occurrence of hydrothermal vents and measures applicable to protect hydrothermal vents and associated communities in the RA. ICES advice 1) VME indicator species ICES advises that in the NEAFC RA, VME indicators should be considered by habitat type and/or at the taxonomic level of family rather than by an exhaustive list of all likely species that could be indicators of VMEs. This approach avoids the risk of excluding or misidentifying any potential species, while ensuring that VMEs are appropriately recognised. Table 1.5.5.3.1 lists seven VME habitat types for the Northeast Atlantic with the taxa that are most likely to be found in these habitats. For comparison and harmonization the equivalent set of species from the NAFO list is also presented. All the habitats listed in Table 1.5.5.3.1 should contain significant aggregations of the representative taxa, and those taxa will most commonly meet the criteria of long-lived, functional significance or fragility. For the most part the families in both NEAFC and NAFO areas are comparable, but there are differences at species level. ICES Advice 2013, Book 1 85 Table 1.5.5.3.1 List of deep-water VMEs and their characteristic taxa. NAFO species have been aligned with the proposed VME habitat type for NEAFC and their representative taxa. In some VME habitat types, no species were listed by the NAFO Scientific Council. P ROPOSED NEAFC VME HABITAT T YPE R EPRESENTATIVE NEAFC T AXA C ORRESPONDING NAFO SPECIES 1. Cold-water coral reef A. Lophelia pertusa reef Lophelia pertusa Lophelia pertusa* Solenosmilia variabilis Solenosmilia variabilis* ANTHOTHELIDAE ANTHOTHELIDAE Anthothela grandiflora* CHRYSOGORGIIDAE Chrysogorgia sp. Metallogorgia melanotrichos Iridogorgia sp. ISIDIDAE, KERATOISIDINAE Acanella arbuscula Acanella eburnea Keratoisis ornata* Keratoisis sp.* Lepidisis sp. PLEXAURIDAE Swiftia sp.* Paramuricea grandis Paramuricea placomus* Paramuricea sp. Placogorgia sp. Placogorgia terceira ACANTHOGORGIIDAE Acanthogorgia armata* CORALLIIDAE Corallium bathyrubrum Corallium bayeri PARAGORGIIDAE Paragorgia arborea* Paragorgia johnsoni PRIMNOIDAE Calyptrophora sp. Parastenella atlantica Primnoa resedaeformis* Thouarella grasshoffi Narella laxa B. Solenosmilia variabilis reef 2. Coral garden A. Hard-bottom coral garden i. Hard-bottom gorgonian and black coral gardens CHRYSOGORGIIDAE ISIDIDAE, KERATOISIDINAE PLEXAURIDAE ACANTHOGORGIIDAE CORALLIIDAE PARAGORGIIDAE PRIMNOIDAE SCHIZOPATHIDAE ii. Colonial scleractinians on rocky outcrops Lophelia pertusa Solenosmilia variabilis iii. Non-reefal scleractinian aggregations Enallopsammia rostrata Madrepora oculata Enallopsammia rostrata Madrepora oculata* CHRYSOGORGIIDAE CHRYSOGORGIIDAE Radicipes gracilis* CARYOPHYLLIIDAE FLABELLIDAE --------- B. Soft-bottom coral gardens i. Soft-bottom gorgonian and black coral gardens ii. Cup-coral fields 86 ICES Advice 2013, Book 1 P ROPOSED NEAFC VME HABITAT T YPE R EPRESENTATIVE NEAFC T AXA C ORRESPONDING NAFO SPECIES iii. Cauliflower coral fields NEPHTHEIDAE ----- 3. Deep-sea sponge aggregations A. Ostur sponge aggregations GEODIIDAE GEODIIDAE Geodia barretti* Geodia macandrewii* Geodia phlegraei* ANCORINIDAE Stelletta normani* Stelletta sp. Stryphnus ponderosus* PACHASTRELLIDAE Thenea muricata* ANCORINIDAE PACHASTRELLIDAE B. Hard-bottom sponge gardens ----AXINELLIDAE ----MYCALIDAE POLYMASTIIDAE TETILLIDAE C. Glass sponge communities 4. Seapen fields ROSSELLIDAE PHERONEMATIDAE ANTHOPTILIDAE PENNATULIDAE FUNICULINIDAE HALIPTERIDAE KOPHOBELEMNIDAE PROTOPTILIDAE UMBELLULIDAE VIRGULARIIDAE ICES Advice 2013, Book 1 ACARNIDAE Iophon piceum* AXINELLIDAE Axinella sp.* Phakellia sp.* ESPERIOPSIDAE Esperiopsis villosa* MYCALIDAE Mycale (Mycale) lingua* POLYMASTIDAE Polymastia sp.* Weberella bursa* Weberella sp. TETILLIDAE Craniella cranium* ROSELLIDAE Asconema foliatum* ----ANTHOPTILIDAE Anthoptilum grandiflorum PENNATULIDAE Pennatula aculeata* Pennatula grandis Pennatula sp. FUNICULINIDAE Funiculina quadrangularis* HALIPTERIDAE Halipteris cf. christii* Halipteris finmarchica* Halipteris sp.* KOPHOBELEMNIDAE Kophobelemnon stelliferum* PROTOPTILIDAE Distichoptilum gracile Protoptilum sp.* UMBELLULIDAE Umbellula lindahli VIRGULARIIDAE Virgularia cf. mirabilis* 87 P ROPOSED NEAFC VME HABITAT T YPE 5. Tube-dwelling anemone patches R EPRESENTATIVE NEAFC T AXA CERIANTHIDAE C ORRESPONDING NAFO SPECIES 6. Mud- and sand-emergent fauna BOURGETCRINIDAE BOURGETCRINIDAE Conocrinus lofotensis ANTEDONTIDAE Trichometra cubensis HYOCRINIDAE Gephyrocrinus grimaldii ----- ANTEDONTIDAE HYOCRINIDAE XENOPHYOPHORA SYRINGAMMINIDAE 7. Bryzoan patches ----- Pachycerianthus borealis* EUCRATEIDAE Eucratea loricata * Species common to the NEAFC and the NAFO areas. 2) Maps of VME elements ICES has mapped three areas with VME elements (geomorphological features likely to contain VMEs): the Mid-Atlantic Ridge, the Rockall–Hatton area, and a set of isolated seamounts. Mid-Atlantic Ridge ICES advises that the Mid-Atlantic Ridge be regarded as one continuous combination of VME elements, southwards from the boundary of the Icelandic EEZ to the boundary of the EEZ north of the Azores (Figure 1.5.5.3.1). It is a chain of pinnacles, knolls, seamounts, ridges, and troughs that together make up one contiguous VME element. ICES has not attempted to provide a boundary for the Mid-Atlantic Ridge. 88 ICES Advice 2013, Book 1 Figure 1.5.5.3.1 The extent of the Mid-Atlantic Ridge from the Azores in the south to Iceland in the north showing peaks shallower than 2000 m depth. The NEAFC RA area is shown as a red boundary. Rockall–Hatton area ICES advises that the Rockall–Hatton area contains VME elements. These include some of the areas already closed to fishing due to the certain presence of VMEs. ICES has mapped the following VME elements outside the currently protected areas (Figure 1.5.5.3.2). • • • • • South Rockall Escarpment and Lorian Bank is an area of steep flanks, rising from 2000 m to the top of Lorian Bank at a depth of 800 m. Fangorn Bank is a knoll of very high rugosity, rising from 2000 m to around 1500 m. Edora’s Bank Western Approach consists of two seamounts in this area known as the Eridor Seamounts, both with summits above 2000 m and a steep flanked ridge running toward Edora’s Bank that is above 2000 m. A third seamount known as Rohan Seamount lies south of Edora’s Bank, with the summit at or just below 2000 m. The South Hatton Knoll has records of VME indicator species. The southwest corner of Lousy Bank is the lower slope of a seamount. ICES Advice 2013, Book 1 89 Figure 1.5.5.3.2 The Rockall–Hatton area, showing areas containing geomorphological features (black circles and polygons) that can be considered VME elements at depths shallower than 2000 m (black contour) that are currently open to bottom fisheries. Also shown are areas currently closed by NEAFC to bottom fishing. Background multibeam bathymetry courtesy of Irish Geological Survey, Instituto Español de Oceanografía (IEO) (Spain), and DTI (UK), with depth contours based on Gebco. Isolated seamounts ICES advises that all isolated seamounts (outside the Mid-Atlantic Ridge) whose summits are shallower than 2000 m depth, be regarded as VME elements. Some of these seamounts might be grouped into broad areas (e.g. immediately SW of the Azores EEZ), but equally they could be regarded as individual features (Figure 1.5.5.3.3). 90 ICES Advice 2013, Book 1 Figure 1.5.5.3.3 3) Isolated seamounts outside the Mid-Atlantic Ridge with summits shallower than 2000 m depth. The NEAFC RA area is shown as a red boundary. Hydrothermal vents There are only five known or inferred vent sites within the NEAFC RA (Figure 1.5.5.3.4). The vent sites, other than the one inferred site on the Reykjanes Ridge, are deeper than 2000 m and are hence unlikely to be impacted by fishing activities. The exact location of the site on the Reykjanes Ridge is not known, but due to the likely fragility of habitats and organisms it is advised that all bottom-contacting fishing gear should be prohibited from the area. ICES Advice 2013, Book 1 91 Figure 1.5.5.3.4 Confirmed or inferred hydrothermal vents in the North Atlantic. Three vents are clustered SW of the Azores (circled). The NEAFC RA area is shown as a red boundary. Background: 1) VME indicator species in the NEAFC and NAFO Regulatory Areas FAO provides criteria for species indicative of VMEs. These are: 1) 2) 3) 4) 5) unique or rare; functionally significant; fragile; have unusual life-history traits, such as being long-lived, slow growing, late maturing, recruit unpredictably; contribute to the structural complexity of the ecosystem. NAFO’s list of taxa, used as indicators of VMEs in the Northwest Atlantic was drawn up by considering each of the candidate VME indicator species in relation to the criteria outlined by the FAO guidelines. It is important to appreciate that in the NEAFC RA, there are several biogeographic provinces, whereas the NAFO RA consists of just one. A large number of taxa and some species are common to the NE and NW Atlantic (see Table 92 ICES Advice 2013, Book 1 1.5.5.3.1). For the most part, the families in both areas are comparable. There are some species and families that are not on the NAFO list, but may represent VMEs in the NEAFC RA. These are cup corals, cauliflower coral gardens dominated by soft corals of the family Nephtheidae, and fragile Xenophyophores. In the NAFO area there are species listed which are not considered to be of relevance in the NEAFC RA, such as a species of sea lily (crinoid) in the family Hyocrinidae that is currently unrecorded in NEAFC and bryozoan patches of the family Eucrateidae (Eucratea loricata), which only occur in shallow waters. 2) VME elements ICES applied NAFO’s VME element classification framework to features in the NEAFC RA. Table 1.5.5.3.2 lists five VME elements (and the NAFO equivalent) identified by ICES that have a high likelihood of supporting VMEs. In some cases there is evidence of VME indicators associated with these elements, but for other areas this cannot be verified at the current time. Table 1.5.5.3.2 List of VME elements known to occur in the NAFO and NEAFC Regulatory Areas (table modified from NAFO SCS Doc 12/19, 2012). P HYSICAL VME I NDICATOR E LEMENTS Explanation ICES/NEAFC Examples from Nomenclature NEAFC RA Isolated seamounts Figure 1.5.5.3.2 Non-MAR seamounts NAFO Nomenclature Examples from NAFO RA Seamounts Fogo Seamounts (Division 3O, 4Vs) Newfoundland Seamounts (Division 3MN) Corner Rise Seamounts (Division 6GH) New England Seamounts (Division 6EF) Steep-slopes and peaks on mid-ocean ridges Mid-Atlantic Ridge (Figure 1.5.5.3.1) Steep ridges and peaks support coral gardens and other VME species in high density Not present Knolls Hatton Bank, Fangorn Bank (within Figure 1.5.5.3.3) A typographic feature that rises less than 1000 m from the seafloor Knolls Orphan Knoll (Division 3K) Beothuk Knoll (Division 3 LMN) Southeast Shoal Tail of the Grand Bank spawning grounds (Division 3N) Canyon-like features Loury Canyon, margin of Edora’s Bank (within Figure 1.5.5.3.3) A steep sided ‘catchment’ feature not necessarily associated with a shelf, island or bank margin Canyons Shelf-indenting canyon; Tail of the Grand Bank (Division 3N) Canyons with head >400 m depth; South of Flemish Cap and Tail of the Grand Bank (Division 3MN) Canyons with heads >200 m depth; Tail of the Grand Bank (Division 3O) Steep flanks >6.4º SE Rockall (within Figure 1.5.5.3.3) from NAFO SCR Doc 11/73 Steep flanks >6.4º South and southeast of Flemish Cap. (Division 3 LM) ICES Advice 2013, Book 1 93 Description of VME elements within the NEAFC Regulatory Area i. The Mid-Atlantic Ridge as one contiguous VME element The Mid-Atlantic Ridge (MAR) between Iceland and the Azores may be characterised as one contiguous VME element with a complex topography, comprising the axial valley and flanks with hills and valleys of various depths and configurations and including many steep and seamount-like structures. Due to the structural complexity, mapping of individual VME elements has not been attempted. It is likely that most features shallower than 2000 m on the MAR are potential VME elements, but it should be noted that much of the area has a covering of sediment. Some major fracture zones occur where the ridge axis is broken and include deep east–west steep-walled canyon-like troughs. The major double fracture is the Charlie-Gibbs Fracture zone at about 52˚N. ii. VME elements in the Rockall–Hatton area The Rockall–Hatton area is a topographically complex area that has numerous VME elements within it, including seamounts, banks, steep flanks, and knolls shallower than 2000 m. Several of these have been previously identified by ICES as sites of VMEs, e.g. the upper section of Hatton Bank, Edora’s Bank, and several areas in the northwest and southwest of Rockall Bank. NEAFC has closed bottom fisheries in these areas. However, there are additional areas that ICES has identified as containing VME elements (Figure 1.5.5.3.3). The Fangorn Bank has a single longline bycatch record of black coral; there are no other records of VME indicator species from these additional areas. iii. Isolated seamounts (with summit shallower than 2000 m) Seamounts by definition rise 1000 m or more from the surrounding seafloor. Isolated seamounts are distributed throughout the NEAFC RA (Table 1.5.5.3.3). 94 ICES Advice 2013, Book 1 Table 1.5.5.3.3 L ATITUDE ( DECIMAL ) 36.4246 36.0525 36.4796 35.989 36.4723 36.3057 36.4544 36.4507 36.4875 36.328 36.9826 36.6708 37.0739 37.0222 37.0316 37.0182 37.0541 43.4153 43.4194 43.59 44.7278 44.7219 44.4216 44.5803 44.9656 44.0997 45.1114 43.994 44.5156 44.5443 45.0443 44.6221 52.4838 56.0755 52.5106 59.9183 54.9042 54.5966 57.8537 54.8042 55.42 43.5762 43.574 43.5941 44.5381 43.4113 43.0192 44.1213 44.6749 43.0233 43.3909 41.3219 41.3236 43.971 43.5406 45.0787 43.7909 44.6842 44.0691 44.1033 Seamounts with summit shallower than 2000 m from the NEAFC area (from Morato et al., in press). L ONGITUDE ( DECIMAL ) -33.8668 -33.7383 -33.8358 -33.6681 -33.7754 -34.3175 -33.4795 -33.378 -34.0619 -33.9304 -34.8651 -38.0743 -35.5123 -35.1158 -35.1771 -35.0318 -35.3837 -32.2388 -37.6799 -38.672 -34.3646 -34.0554 -40.2322 -33.9406 -40.9232 -38.9873 -39.4312 -36.5227 -40.4651 -40.5186 -40.9905 -40.9294 -41.0124 -37.3454 -40.5756 -34.1654 -25.285 -25.4485 -26.5802 -22.2819 -30.3948 -22.4496 -22.3927 -22.4984 -25.2685 -26.7975 -24.7639 -22.1207 -24.3592 -25.0386 -14.1022 -20.1974 -20.173 -21.7314 -22.2267 -13.4134 -22.9436 -25.4386 -21.8688 -21.9925 ICES Advice 2013, Book 1 S UMMIT DEPTH (M) 434 802 936 968 985 1011 1098 1106 1131 1229 1717 1774 1869 1958 1968 1972 1986 895 943 1058 1240 1379 1491 1607 1729 1798 1855 1861 1885 1904 1940 1951 1558 1754 1823 1962 1533 1645 1681 1724 1842 958 961 993 1098 1124 1226 1231 1234 1297 1316 1407 1407 1529 1588 1682 1714 1747 1750 1798 H EIGHT (M) 2036 1355 1507 1111 1483 1263 1281 1253 921 1342 984 1637 1110 1063 1095 970 1022 2601 2926 3194 2228 1716 2736 1612 2187 2321 2077 2084 2157 2108 1912 2216 1980 961 1714 976 1478 1081 1108 1623 1127 1952 2033 1971 1780 1942 2214 2169 1742 2072 3715 1947 1993 1832 1620 2506 1562 1150 1462 1330 B ASAL AREA ( KM 2 ) 934 981 1111 1099 867 1119 783 841 449 488 1145 1088 914 792 878 560 688 841 1147 1413 1263 877 1584 999 1504 1161 1328 1074 1504 1408 1600 1252 1130 784 1372 698 1137 1264 883 1036 969 1568 1568 1600 1600 652 716 936 1274 979 988 1291 1303 1408 1472 1382 1317 1207 1363 965 95 41.4425 42.4846 43.6296 42.5894 41.9835 44.5478 43.5319 42.8153 44.2914 43.3793 41.5813 42.6668 36.7903 36.6345 36.8565 36.7007 36.669 37.0303 36.2319 36.304 36.6783 36.5771 37.5095 37.3402 -21.2099 -19.0334 -23.7155 -26.1673 -19.9793 -25.0516 -23.0083 -21.5212 -22.9451 -25.7231 -20.0606 -21.1705 -14.3063 -14.2357 -14.4412 -14.2756 -14.2468 -13.8794 -14.5563 -14.5609 -13.9654 -14.9466 -13.9323 -14.5041 1801 1801 1823 1859 1872 1888 1915 1981 1983 1985 1997 1999 120 153 162 188 192 846 925 1000 1197 1213 1335 1356 1713 2425 1256 1479 2052 941 1052 1371 1747 1048 1535 1532 1714 2251 1593 1915 2081 1830 2287 2026 2192 1263 2393 1567 1297 1520 1335 1029 1327 779 724 1365 1206 997 1309 1360 1328 1270 1600 1552 1221 1520 1199 1268 538 1424 1269 1477 37.4694 -14.137 1456 1721 1107 iv. Knolls Knolls are topographic features that rise less than 1000 m from the surrounding seafloor. The resolution of standard regional-scale bathymetric data does not allow a comprehensive analysis of this class of VME element. v. Canyon-like features Most canyons in the ICES area are found along the European continental margins and there appears to be very few in the NEAFC RA that are not part of the Mid-Atlantic Ridge or Rockall–Hatton area. vi. Steep flanks and slopes Using bathymetric data, NAFO has classified areas of 6.4° or steeper slopes as VME elements (Murillo et al., 2011). However, existing regional-scale bathymetric data are not resolved to a fine enough scale to make a comprehensive analysis of this class of VME element. 3) Hydrothermal vents In the NEAFC RA, hydrothermal vents occur on the Mid-Atlantic Ridge. While some have been visually surveyed and studied, some remain unconfirmed and are inferred based on chemical plume detections in the overlying water column. Three vent fields in the NEAFC RA are located southwest of the Azores, one lies north of the Azores, and one lies south of Iceland on the Reykjanes Ridge (Figure 1.5.5.3.4 and Table 1.5.5.3.4). Only the vent on the Reykjanes Ridge is at a depth shallower than 2000 m. Table 1.5.5.3.4 Positions of known or inferred hydrothermal vents in the NEAFC RA. A REA V ENT NAME Rainbow L ATITUDE ( DECIMAL 36.21667 SW Azores L ONGITUDE ( DECIMAL ) -33.9 SW Azores AMAR 36.3833 -33.65 SW Azores S AMAR 1 36.083 -34.083 North Azores Moytirra 45.4833 -27.85 Reyjkanes Ridge Reykjanes Ridge, Area A 62.45 -25.433 Measures to protect hydrothermal vents 96 ICES Advice 2013, Book 1 In 2010, a workshop sponsored by the International Seabed Authority (ISA) was held to formulate general guidelines for the conservation of vent and seep ecosystems at regional and global scales. A number of anthropogenic pressures arising from indirect commercial activities, such as shipping, cable laying, and waste disposal may impact upon seeps and vents. The most severe threats to natural ecosystem structure and function at vents and seeps were considered to be the extractive industries (minerals, hydrocarbons) and bottom trawl fisheries (International Seabed Authority, 2011). Sources ICES 2013. Report of the ICES/NAFO Joint Working Group on Deep-water Ecology (WGDEC). ICES CM 2013/ACOM: 28. International Seabed Authority. 2011. Environmental Management of Deep-Sea Chemosynthetic Ecosystems: Justification of and Considerations for a Spatially Based Approach. Available online at: http://www.isa.org.jm/files/documents/EN/Pubs/TS9/index.html (accessed May 2013). Morato, T., Kvile, K. Ø., Taranto, G. H., Tempera, F., Narayanaswamy, B. E.,. Hebbeln, D., Menezes, G., Wienberg, C., Santos, R. S., and. Pitcher, T. J. In press. Seamount physiography and biology in North-East Atlantic and Mediterranean Sea. Biogeosciences. Murillo, F. J., Kenchington, E., Sacau, M., Piper, D. J. W., Wareham, V., and Muñoz, A. 2011. New VME indicator species (excluding corals and sponges) and some potential VME elements of the NAFO Regulatory Area. NAFO SCR Doc. 11/73. ICES Advice 2013, Book 1 97 1.5.5.4 ECOREGION SUBJECT Special request, Advice May 2013 General advice Advice on threshold levels for longline fishing Advice summary ICES advises the use of a threshold of 10 VME indicators caught per 1000 hook segment or per 1200 m section of long line, whichever is the shorter, to indicate the presence of a VME. Request ICES is requested to advice on the appropriateness of applying the threshold levels for VME indicator species for long line fishing as adopted in the SEAFO, and CCMLAR, in the NEAFC RA. ICES advice ICES advises that NEAFC set specific threshold levels for VME indicator bycatch by longlines. This is because of the substantial difference in the bycatch of VME indicators on longlines compared to trawls. ICES considers that the application of threshold levels for VME indicators for longline fishing adopted by SEAFO and CCMLAR would be an improvement on the current criteria to identify VMEs because it offers a way to standardize effort across fleets. However, the application of the SEAFO and CCMLAR thresholds are complicated, rely upon 100% observer coverage and are therefore unlikely to be operational in the NEAFC RA. ICES therefore advises to adapt the SEAFO and CCMLAR thresholds and to use a threshold of 10 hooks with VME indicators per caught per 1000 hook segment or per 1200 m section of long line, whichever is the shorter, to indicate the presence of a VME. Background Currently NEAFC operates a simple VME encounter measure for all fishing gear types including longlines, stating that if over 30 kg of live corals or 400 kg of live sponges is taken as bycatch per set a VME is considered to have been encountered and must be reported. This rule is considered inappropriate for long lines mainly because of the different selectivity of long lines compared to trawls and because of the heterogeneity of the long line fleet in the NEAFC RA. The long line fisheries in the NEAFC RA vary considerably according to target species, characteristics of the fishing areas and traditions, ranging from technically advanced and highly mechanized fisheries on large vessels to small-scale traditional artisanal fisheries. The differentiation also includes variability in the number of hooks. SEAFO and CCMLAR consider a ‘VME indicator unit’ either one litre of the type of VME indicator organisms that can be placed in a 10-litre container, or one kilogramme of the type of VME indicator organisms that do not fit into a 10-litre container. The threshold is exceeded when 10 or more VME indicator units are collected per 1000 hook segment or a 1200 m section of line, whichever is the shorter. Sources CCAMLR. 2010. http://www.ccamlr.org/en/measure-22-07-2010. ICES. 2013. Report of the ICES/NAFO Joint Working Group on Deep-water Ecology (WGDEC). ICES CM 2013/ACOM: 28. SEAFO. 2011. http://www.seafo.org/ConservationMeasures/Conservation%20Measure%202211%20on%20Bottom%20Fishing% 20Activities%20in%20the%20SEAFO%20Convention%20Area.pdf. 98 ICES Advice 2013, Book 1 1.5.5.5 Special request, Advice June 2013 ECOREGION SUBJECT General advice OSPAR/NEAFC special request on review of the results of the Joint OSPAR/NEAFC/CBD Workshop on Ecologically and Biologically Significant Areas (EBSAs) Advice summary ICES reviewed the ecological evidence supporting the ten proposed ecologically and biologically significant areas (EBSAs) from the OSPAR/NEAFC/CBD Workshop of September 2011, as presented in the annexes to that report. The review applied standard ICES practices and used primarily the references cited in the relevant annexes, but augmented those references with other publications and data sources. In nine of the ten proposed EBSAs, ICES came to different conclusions than were contained in the OSPAR/NEAFC/CBD Workshop report, with regard to the rankings of the Convention on Biological Diversity (CBD) EBSA criteria. Of the ten proposed EBSAs, ICES supports the conclusion of the OSPAR/NEAFC/CBD workshop that the Arctic Ice area (Area 10) meets one or more EBSA criteria and this area could go forward at this time, possibly with minor suggested changes to the rationale. In four proposed EBSAs (Reykjanes Ridge south of Iceland EEZ (Area 1); Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge (Area 2); Mid-Atlantic Ridge north of the Azores (Area 3); the Hatton and Rockall banks and Hatton–Rockall Basin (Area 4)), ICES considers that much of the area within the proposed EBSAs do not meet any of the EBSA criteria and for this reason the boundaries of these proposals need to be revised. More restricted parts of the proposed EBSAs meet several of the EBSA criteria and could go forward after boundary revision. ICES notes great similarities in the pro forma describing Areas 1 and 3 and part of Area 2 (OSPAR/NEAFC/CBD, 2011). A boundary revision to encompass the relevant parts of these areas as a single extended Mid-Atlantic Ridge proposed EBSA could be considered a step forwards. ICES recommends changes also to the pro forma rankings for all of these proposed EBSAs. Only a small part of the proposed EBSA for the Arctic Front – Greenland/Norwegian seas (Area 9) possibly meets some of the EBSA criteria. However, another part of the general area might meet some of the EBSA criteria. ICES recommends that further data analyses followed by an evaluation of the new results against the EBSA criteria be undertaken before any further decision is taken. The rationales for four proposed EBSAs (around the Pedro Nunes and Hugo de Lacerda seamounts (Area 5); Northeast Azores–Biscay Rise (Area 6); Evlanov Seamount region (Area 7); and Northwest of Azores EEZ (Area 8)) are not well supported by the information presented in the relevant annexes. There is a need for further data and analyses in these areas, particularly in relation to seabirds, and another evaluation of the areas against the EBSA criteria. ICES found no clear evidence of additional EBSAs in areas beyond national jurisdiction (ABNJ) of the Northeast Atlantic meeting the CBD scientific criteria. Request a) Review the description of areas meeting one or more of the CBD EBSA scientific criteria developed as an outcome of the Joint OSPAR/NEAFC/CBD Scientific Workshop, in particular: 1. Review each of the ten area delineations and descriptions in line with the CBD EBSA Scientific criteria and the most up-to-date scientific data and information, specifying any additional scientific data and information that is available; 2. Provide, if appropriate, revised EBSA proposals in the format of proformas adopted by the CBD b) If there is clear evidence for additional areas in ABNJ of the North-East Atlantic meeting the CBD EBSA scientific criteria, present a description with supporting scientific data and information for such areas, including CBD EBSA proformas for each. ICES advice ICES made the following conclusions and proposals. ICES Advice 2013, Book 1 99 Area 1. Reykjanes Ridge south of Iceland EEZ: Much of the area in the proposed EBSA does not meet any of the EBSA criteria. A more restricted area down the spine of the Mid-Atlantic Ridge and defined by depth ranges of deep-water coral and sponge concentrations does meet several EBSA criteria and the boundary delineation, ranking, and full rationale could be developed based on this new boundary. Area 2. Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge: Some areas in the proposed EBSA do not meet any of the EBSA criteria. A complex combination of the area down the spine of the Mid-Atlantic Ridge, the benthic area aligned with and close to the main fractures, and the water column in which the Subpolar Front is found throughout the year, does meet several EBSA criteria and the boundary delineation, ranking and full rationale could be developed at based on this new boundary. Area 3. Mid-Atlantic Ridge north of the Azores: Much of the area in the proposed EBSA does not meet any of the EBSA criteria. A more restricted area down the spine of the Mid-Atlantic Ridge and defined by depth ranges of deep-water coral and sponge concentrations does meet several EBSA criteria and the boundary delineation, ranking, and full rationale could be developed based on this new boundary. Area 4. The Hatton and Rockall banks and Hatton–Rockall Basin: Much of the area in the proposed EBSA does not meet any of the EBSA criteria. A more restricted area down to approximately 1500–1800 m depth, but excluding the abyssal plain does meet several EBSA criteria and the boundary delineation, ranking, and full rationale could be developed based on this new boundary. Area 5. Around the Pedro Nunes and Hugo de Lacerda seamounts: The proposed EBSA is not supported well by the information presented in the pro forma. There is a need for further analyses of those data already considered, as well as any additional relevant data on seabird foraging and other information. When these analyses are done, including for the additional data, another evaluation of the area against the CBD EBSA criteria would make it possible to advise which areas, if any, meet EBSA criteria. Area 6. Northeast Azores–Biscay Rise: The proposed EBSA is not supported well by the information presented in the pro forma. There is a need for further analyses of those data already considered as well as any additional relevant data on seabird foraging and other information. When these analyses are done, including for the additional data, another evaluation of the area against the CBD EBSA criteria would make it possible to advise which areas, if any, meet EBSA criteria. Area 7. Evlanov Seamount region: The proposed EBSA is not supported well by the information presented in the pro forma. There is a need for further analyses of those data already considered as well as any additional relevant data on seabird foraging and other information. When these analyses are done, including for the additional data, another evaluation of the area against the CBD EBSA criteria would make it possible to advise which areas, if any, meet EBSA criteria. Area 8. Northwest of Azores EEZ: The proposed EBSA is not supported well by the information presented in the pro forma. There is a need for further analyses of those data already considered as well as any additional relevant data on seabird foraging and other information. When these analyses are done, including for the additional data, another evaluation of the area against the CBD EBSA criteria would make it possible to advise which areas, if any, meet EBSA criteria. Area 9. The Arctic Front – Greenland/Norwegian seas: Only a small part of the area proposed by the OSPAR/NEAFC/CBD Workshop as an EBSA was considered to possibly meet some of the criteria. However, another part of the general area might meet some EBSA criteria. There is a need for more analyses of productivity and diversity data for the more southerly part of the main area, and then a re-evaluation of the new results against the EBSA criteria, before any areas might be considered as possibly meeting EBSA criteria. Area 10. The Arctic Ice: The rationale for concluding that this area meets one or more EBSA criteria can be improved, but the review by ICES generally supports the conclusions of the OSPAR/NEAFC/CBD workshop. A suggested revised proforma is attached as Annex 1.5.6.5.1 to this advice. With regard to new proposed areas that meet EBSA criteria, ICES has no additional information. However, ICES suggests a potential alternative configuration to the areas in proposed EBSAs 1, 2, and 3 that would comprise two areas meeting EBSA criteria, one covering the specified depths of the entire Mid-Atlantic Ridge, and one for the Charlie-Gibbs Fracture Zone and pelagic area of the Subpolar Front. Each of the two areas would have coherent, but different ecological rationales. 100 ICES Advice 2013, Book 1 ICES has provided its rankings for the revised proposed EBSAs and the rationale for those rankings. ICES has not revised the narrative or the references in the existing pro forma of proposed EBSAs other than for Arctic Ice habitat (Annex 1.5.6.5.1). Once OSPAR and NEAFC have made a decision on the configuration of the Mid-Atlantic Ridge proposed EBSAs, ICES could revise the pro forma for these EBSAs by the end of September 2013. Background Method ICES conducted its review informed by the content of the CBD Decisions IX/20 and X/29 on Marine and Coastal Biodiversity (CBD, 2008, 2010), the reports from the ‘Azores Workshop’ (CBD, 2007) and the ‘Ottawa Workshop’ (CBD, 2009), and the UNGA Resolution 58/240 (United Nations, 2004). ICES considers that the application of the criteria was intended to be a comparative or relative process, such that areas should be evaluated against other generally comparable areas (e.g. of comparable depth and latitude). In addition, even though the application of EBSA criteria is not guided directly by management considerations, potential benefits of spatial management measures are a relevant consideration in the evaluation. However, the appropriate baseline is not the absence of all management, but the presence of measures sufficient to ensure human uses are sustainable in areas typical of the zone of evaluation. ICES is responding to a request about EBSAs, and ICES stresses that this advice does not imply that any areas reviewed should or should not be considered as VMEs (but ICES notes that there is an overlap with advice provided recently on vulnerable marine ecosystems (VMEs; ICES, 2013a)). ICES notes that all areas found to meet criteria for VMEs would be expected to meet one or more criteria for EBSAs as well. However, the reverse is not necessarily true and EBSAs do not necessarily contain VMEs. There is neither a policy nor an ecological rationale for automatically excluding bottom fishing (or any other activity) from areas proposed as EBSAs. The expected initial response of regulatory authorities is to conduct risk or threat assessments of the activities they regulate relative to the properties considered ecologically or biologically significant, and to subsequently undertake management appropriate to the outcome of these assessments. ICES advice is based on applying several standards during its review, including: • • • • • Assigned rankings on a criterion should apply to at least most of the area included in a proposed EBSA, and not just to a small subset of the total area. Higher assigned rankings required the proposed area to differ from adjacent areas and other areas of similar depth and latitude on the property represented by the criterion. Some evidence must be available to justify awarding a higher ranking, noting that comprehensive data for the high seas cannot be expected. Rankings should not be based on the presence or history of threats to the features represented by the criteria, but on the biological, ecological, and geomorphological features of the area. Although EBSAs are not defined by or linked to any particular management actions by any authorities, it is appropriate to consider whether or not spatial management tools might benefit the conservation or sustainable use of the relevant features. For each of the ten areas in the OSPAR/NEAFC/CBD report proposed to meet EBSA criteria, ICES assigned one of three categories: • • Proceed with boundaries proposed by the OSPAR/NEAFC/CBD Workshop, with a rationale revised by ICES. Proceed with developing a proposed EBSA with different boundaries than those proposed by the OSPAR/NEAFC/CBD Workshop, and with a rationale provided by ICES. • Do not proceed with proposing an EBSA at this time, but rather undertake further collation and analysis of information and reconsider when the additional work is completed. Although ICES review included an evaluation and commentary on all pro forma contents of the OSPAR/NEAFC/CBD 2011 Workshop report, this advice presents only ICES conclusions as to whether the areas meet the EBSA criteria. This advice takes the form of biological properties that ICES concludes would define the boundaries, rankings, and rationales of those areas against the EBSA criteria. Proposed EBSAs that need minor revisions to the rationale Area 10. The Arctic Ice A suggested revised pro forma is attached to this advice as Annex 1.5.6.5.1. Proposed EBSAs that should have redefined boundaries before proceeding ICES Advice 2013, Book 1 101 Area 1. Reykjanes Ridge south of Iceland EEZ ICES advises that an area along the Reykjanes Ridge can be justified as meeting one or more EBSA criteria. This would be a much smaller area than the one proposed in the OSPAR/NEAFC/CBD Workshop report. Appropriate boundaries for such a proposed area that meets EBSA criteria would be a depth contour that runs in the deeper of two properties: 1. 2. Including a large portion (arbitrarily, perhaps 90%) of all hard volcanic substrates; habitats are reported to host the larger known coral formations and their associated communities Including a large portion (arbitrarily, 90%) of the records of large sponge communities in the overall northern Mid-Atlantic Ridge. In addition, the proposed area will include the only known hydrothermal vent in the area (Olafsson et al., 1991; German et al., 1994; German and Parsons, 1998; Mironov and Gebruk, 2007), whatever depth contour is used. Information on water masses should also be consulted, allowing proper identification of benthic and fish fauna to be included in a revised narrative to a pro forma for this area. In the time available ICES did not have the geological data to delineate the depth contour that would meet the first criterion, but such information should be readily available in marine geology databases. Nor did ICES have access to all of the records of where the large sponge deposits were taken. However, references to sources for those data are in the OSPAR/NEAFC/CBD Workshop pro forma and should be tracked back to find the appropriate depth contour for the second criterion in the tables below. Evaluation of the revised area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking Uniqueness rarity or Low Some High X (X) The bracketed “High” ranking is for a very restricted area covering the only known hydrothermal vent on the Reykjanes Ridge. Though this is a unique feature it occupies only a small part of the area proposed here as meeting this EBSA criterion (Olafsson et al., 1991; German et al., 1994; German and Parsons, 1998; Mironov and Gebruk, 2006). The MarEco sampling of corals and sponges reported a few species new to science and these may be restricted to the proposed area, although a firm conclusion on this cannot be drawn until more extensive sampling is undertaken. Areas that are required for a Special X importance for population to survive and thrive. life-history stages of species Explanation for ranking Although many populations undoubtedly complete their life cycles within the large area proposed in the OSPAR/NEAFC/CBD Workshop Report as an EBSA, this would be true of any marine area of comparable size. There is no evidence that the life history of any species is strongly dependent on any specific features of the area proposed as an EBSA. There is evidence from other areas of the Northeast Atlantic that areas of high coral density may be important as eggcase and nursery areas of deep-water sharks and rays. The area proposed here is targeted on the depths and substrates associated with higher coral and sponge densities, and if sharks and rays also concentrate spawning and early development in these habitats, then the score would be “Some” or “High” on this criterion as well. 102 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Areas containing habitats for the survival and recovery of endangered, threatened, or declining species, or areas with significant assemblages of such species. Explanation for ranking Importance for threatened, endangered, or declining species and/or habitats Low Some High X Large formations of corals and sponges are found in the area proposed by ICES as meeting this criterion. Habitats containing these species are listed by OSPAR and also feature as VME indicator species; the majority of these would be included in the proposed EBSA. Additional explanation regarding corals and sponges is included in the rationale for the criterion on Vulnerability. The possible role of the area proposed by ICES in the life histories of sharks and rays is discussed in the criterion on Vulnerability and Sensitivity. Areas that contain a relatively high Vulnerability, X proportion of sensitive habitats, fragility, sensitivity, or biotopes, or species that are functionally fragile (highly slow recovery susceptible to degradation or depletion by human activity or by natural events) or with slow recovery. ICES Advice 2013, Book 1 103 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High Explanation for ranking With regard to corals and sponges, Mortensen et al. (2008) found cold-water corals “at every sample station … observed at depths between 800 and 2400 m, however were commonly found shallower than 1400 m …, with species richness being very high. … no major reef structures were recorded, with the maximum colony size approximately 0.5 m in diameter. The number of coral taxa was strongly correlated with the percentage cover of hard bottom substrate ….” The area proposed here is targeted at the seamount peaks and slopes where hard substrates dominate. For sponges, no actual large expanses of sponge reef were reported in the OSPAR/NEAFC/CBD Workshop report. However, the pro forma in that report (OSPAR/NEAFC/CBD, 2011) notes that overall sampling of the area is patchy and cites three studies that found local patches with high densities of sponges, although in no cases were the sizes of the patches documented. These observations of widespread occurrences of corals and sponges are supported by the records in the GRID–Arendal data, which show both taxa to be presented in nearly every appropriate sample taken along the cruise tracks in the database. Biological productivity Areas containing species, populations, or communities with comparatively higher natural biological productivity. X Explanation for ranking Although benthic productivity of the proposed smaller EBSA may be higher than benthic productivity on the abyssal plain, productivity integrated over the entire water column and seafloor seems typical of systems of comparable depth and latitude globally. Areas containing a Biological diversity X comparatively higher diversity of ecosystems, habitats, communities, or species, or with higher genetic diversity. 104 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High Explanation for ranking The possible, but incompletely documented presence of large coral and sponge stands, and the documented high diversity of benthic and associated fish when corals or sponges are present in moderate or high density imply that some areas may have high diversity. Aside from these benthic communities of somewhat restricted distribution, the biodiversity otherwise appears typical of biotic communities at similar depths and latitude. Area 2. Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge ICES advises that an area with the following boundaries, capturing three distinct features, would meet one or more EBSA criteria. The area would include the following features: i. ii. iii. Subpolar Frontal Zone (coinciding with the Charlie-Gibbs Fracture Zone): The northern and southern boundaries for this feature should be set according to the known northernmost and southernmost meandering of the frontal system at 53°N and 48°N, respectively (Søiland et al., 2008). The eastern and western boundaries for this feature should be set according to the eastern and westernmost extension of the Charlie-Gibbs Fracture Zone (topography; approx. at 27°W and 42°W, respectively). Charlie-Gibbs Fracture Zone: The eastern and western boundaries for this feature should be set according to the east–west extension of the Fracture Zone (approx. at 27°W and 42°W, respectively). The northern and southern boundaries for this feature should be set with a view to encompass the characteristic bathymetry, topography, and substrates of the Fracture Zone. Sections of the Mid-Atlantic Ridge: The northernmost and southernmost boundaries would coincide respectively with the southern boundary of proposed Area 1 (Reykjanes Ridge south of Iceland EEZ) and the northern boundary of proposed Area 3 (Mid-Atlantic Ridge north of the Azores). Boundaries to the east and to the west would be a depth contour running in the deeper of two properties: • • including a large portion (arbitrarily, perhaps 90%) of all hard volcanic substrates; habitats reported to host the larger known coral deposits and their associated communities; including a large portion (arbitrarily, 90%) of the larger known deep-sea sponge aggregations. Where the Charlie-Gibbs Fracture Zone crosses the Mid-Atlantic Ridge, the benthic boundaries of the Fracture Zone in feature ii) may extend to the east and west beyond the area defined by feature iii) for the Mid-Atlantic Ridge. In those cases the benthic area of the Fracture Zone is proposed for inclusion in the EBSA proposed here. However, any part of the seafloor and associated benthos that lies below the pelagic feature i) (the total area occupied by the Subpolar Front during its annual movement) but does not meet either feature ii) or feature iii) is not included in the area proposed as meeting EBSA criteria. Only those parts of the water column where the Subpolar Front is prominent at some time during the year are proposed as meeting one or more of the EBSA criteria. Moreover, in the entire pelagic area described by i), at any given time only those parts of the total area where the Subpolar Front is located would be expected to meet some of the criteria. Although maps will show the full pelagic area is proposed as part of this complex EBSA, any conservation measures for ecological properties of the water column would need to take into account the position of the Subpolar Front to be fully effective. For each of the above-mentioned features a set of geographic coordinates delineating their respective boundaries needs to be determined. ICES Advice 2013, Book 1 105 Evaluation of the revised area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking Uniqueness rarity or Low Some X High (X) The portion of the proposed area encompassing both the Charlie-Gibbs Fracture Zone and Subpolar Front Zone are unique. Both represent unique or unusual geomorphological or oceanographic features in the Northeast Atlantic. Other portions of the proposed EBSA are part of the Mid-Atlantic Ridge and, as discussed for Area 1 (Reykjanes Ridge south of Iceland EEZ), may host some unique benthic species based on Mar-Eco sampling. Areas that are required for a Special X importance for population to survive and thrive. life-history stages of species Explanation for ranking There is no evidence available suggesting a significant importance of the area for life-history stages of widespread species in comparison to other marine areas of similar size and depth range. Importance for Areas containing habitats for the X survival and recovery of threatened, endangered, or endangered, threatened, or declining species, or areas with significant declining species and/or assemblages of such species. habitats Explanation for ranking There is good evidence that the area contains a significant assemblage of species and habitats that are assessed to be threatened, endangered, or declining, including the following: orange roughy (Hoplostethus atlanticus), leafscale gulper shark (Centrophorus squamosus), gulper shark (Centrophorus granulosus), Portuguese dogfish (Centroscymnus coelepis), Sei whale (Balaenoptera borealis), sperm whale (Physeter macrocephalus), leatherback turtle (Dermochelys coriacea), Lophelia pertusa reefs, and deep-sea sponge aggregations. Depending on the species, the special features of the Fracture Zone, the Subpolar Frontal Zone, or in a few cases the Mid-Atlantic Ridge, all provide important biological functions to the species which aggregate along each one. Areas that contain a relatively high Vulnerability, X proportion of sensitive habitats, fragility, sensitivity, or biotopes, or species that are functionally fragile (highly slow recovery susceptible to degradation or depletion by human activity or by natural events) or with slow recovery. Explanation for ranking The Charlie-Gibbs Fracture Zone and sections of the Mid-Atlantic Ridge through its associated substrate, current, and feeding conditions, provide habitats to a number of sensitive/vulnerable species and communities both on soft and hard substrate and in the water column. In particular biogenic habitats such as those formed by cold-water corals and sponges are considered vulnerable, often fragile, and slow (if at all) to recover from damage. Some fish species associated with the Fracture Zone and Mid-Atlantic Ridge also show slow growth, late maturity, irregular reproduction, and long generation time, as well as community characteristics of high diversity at low biomass. However, there is no evidence available suggesting that the area contains a significantly higher proportion of species that are functionally fragile or with slow recovery than other areas of comparable structure and depth range along the Mid-Atlantic Ridge. 106 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Areas containing species, populations, or communities with comparatively higher natural biological productivity. Explanation for ranking Biological productivity Low X Some High (X) There is good evidence that, due to the Subpolar Front, the pelagic area where the front is located at any particular time is characterized by an elevated abundance and diversity of many taxa, including an elevated standing stock of phytoplankton. This justifies a ranking of “High” for the pelagic area around the Subpolar Front, as it moves seasonally. However, there is no evidence of relatively elevated productivity in the benthic communities of the Fracture Zone and Mid-Atlantic Ridge. Areas containing a comparatively Biological X (X) higher diversity of ecosystems, diversity habitats, communities, or species, or with higher genetic diversity. Explanation for ranking The area of the Fracture Zone is characterized by a very high structural complexity, offering a diverse range of habitats. The area of the Subpolar Front is a feature where species are documented to assemble seasonally, and the sections of the Mid-Atlantic Ridge north and south of the Fracture Zone represent different biogeographic settings and their respective characteristic communities. Consequently, each of the three features characterizing this area contribute to a relatively higher diversity of ecosystems, habitats, communities, and species in comparison to other areas of similar size along the Mid-Atlantic Ridge. Area 3. Mid-Atlantic Ridge north of the Azores ICES considers that one or more EBSA criteria would be met by an area with boundaries including all the following properties: • • • Including a large portion (arbitrarily, perhaps 90%) of all hard volcanic substrates on the Mid-Atlantic Ridge south of the summer location of the Subpolar Front. Including the Moytirra Hydrothermal Vent Field. Incluings the area in the Mid-Atlantic Ridge included in the 50% density Kernel for foraging Cory’s shearwater. ICES Advice 2013, Book 1 107 Evaluation of the proposed area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking Uniqueness rarity or Low X Some High (X) There is support for qualification under this criterion only from the hydrothermal vent field in the area which is considered to be rare, and its associated communities which may be unique. This habitat is known from a single discrete location (thus the brackets) and so it cannot be considered to offer justification for the entire extent of the proposed area. ICES does not consider that any of the other evidence presented here supports qualification under this criterion. Areas that are required for a Special X importance for population to survive and thrive. life-history stages of species Explanation for ranking There is some support for qualification under this criterion from the occurrence of an important long-range foraging area for Cory’s shearwaters during their breeding season. However, the core area encompassing 50% of locations at sea is relatively small and does not justify the entire extent of the proposed area. If research finds that deep-sea sharks and rays use the denser coral deposits as important spawning and nursery grounds, as has been reported in the Hatton–Rockall Bank area proposed by ICES, then there would be additional justification for a score of “Some” on this criterion. Importance for Areas containing habitats for the survival and X recovery of endangered, threatened, or declining threatened, endangered, or species, or areas with significant assemblages of such species. declining species and/or habitats Explanation for ranking The only recognised threatened and/or declining species identified in the report as occurring in the area are the deepwater sharks Centrophorus squamosus and Centroscymnus coelolepis, both of which are included on the OSPAR list of threatened and/or declining species and habitats. Since this area is not considered to have special importance for their survival (compared with other areas of similar depth and latitude elsewhere) they would not qualify under this criterion. However, in addition to the threatened and declining species mentioned in the proposal, the OSPAR listed habitats ‘seamount communities’ and ‘coral gardens’ are likely to exist in this area. If these were taken into account together with the sharks, this might be regarded as a significant assemblage of threatened and declining species and habitats and the ranking would be “Some”. Areas that contain a relatively high proportion of Vulnerability, X sensitive habitats, biotopes, or species that are fragility, sensitivity, or functionally fragile (highly susceptible to degradation or depletion by human activity or by slow recovery natural events) or with slow recovery. Explanation for ranking The proposed area focuses on areas with high abundance of seamounts, stony corals, and coral gardens. Recently a new hydrothermal vent was discovered on the ridge at 45ºN. Seamounts, ocean ridges with hydrothermal vents, coral 108 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know reefs, and coral gardens are all considered priority habitats in need of protection by the OSPAR convention for the protection and conservation of the Northeast Atlantic. There is good support for qualification under this criterion from the occurrence of vulnerable marine ecosystems, including seamounts, stony corals, coral gardens, and hydrothermal vents. Areas containing species, populations, or Biological X communities with comparatively higher natural productivity biological productivity. Explanation for ranking There is no evidence that the productivity in the revised area is any different from the expected productivity of marine systems of similar depth and latitude. Areas containing a comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or diversity with higher genetic diversity. Explanation for ranking The presence of a hydrothermal vent field does not in itself indicate high diversity, but it does provide some evidence of habitat heterogeneity from which species diversity may be inferred. In addition, there is a mingling of benthic faunas characteristic of both warmer southern and cooler northern ocean environments, giving the area as a whole somewhat higher net biological diversity, although the diversity in any individual site is not markedly enhanced. Area 4. The Hatton and Rockall banks and the Hatton–Rockall Basin The Hatton and Rockall banks, and associated slopes, represent unique offshore bathyal habitats (200 to 3000 m) and constitute a prominent feature of the Northeast Atlantic continental margin south of the Greenland to Scotland ridges. The banks and slopes have high habitat heterogeneity and support a wide range of benthic and pelagic faunas. They are also subject to significant fishing impact, including bottom trawling, longlining, and midwater fisheries. The banks encompass a large depth range and consequently the seabed communities encounter strong environmental gradients (e.g. temperature, pressure, and food availability). These factors cause large-scale changes in species composition with depth and give rise to a high diversity of species and habitats. The area is influenced by a number of different water masses and there is considerable interaction between the topography and physical oceanographic processes, in some areas focusing internal wave and tidal energy which results in strong currents and greater mixing and resuspension. ICES recommend that additional work is needed primarily to refine the boundaries set out for this proposed EBSA. In particular the evidence base to use the 3000 m contour as the southern and western limits of this proposed EBSA is questionable since no evidence was provided that ecosystems meeting EBSA criteria are present at these depths. The features contributing to the uniqueness and rarity, threatened and declining species, vulnerability/fragility/sensitivity, and importance for life-history criteria stages all occur exclusively at relatively shallow depths (< 1500 m) with most being < 1200 m, and the only additional benefit gained by extending the boundary to 3000 m is an increase in the overall depth range covered and hence additional biological diversity. It is unclear whether the additional diversity conferred by the inclusion of the 1500 to 3000 m depth zone is any different from that present in any other area at comparable depth and latitude. ICES therefore recommends that proposed EBSA should go forward with a revised boundary approximating to the 1500 m depth contour. If further work can establish significant additional value in the inclusion of greater depths, the boundary could be adjusted accordingly in the future. Evaluation of the proposed area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Uniqueness rarity or The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, ICES Advice 2013, Book 1 Low Some High X (X) 109 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking The area has considerable environmental heterogeneity, and therefore biological diversity, as a result of its large depth range and strong environmental gradients. Habitat-forming sessile benthic communities, such as those of giant protozoans and sponges, are common. Although distinctive these features are not rare per se. Large areas of cold-water corals and sponges have been reported in the area. Some of these have been impacted by bottom trawl and longline fishing and past periods of bottom gillnet fishing, but some areas of large coral frameworks still exist, including areas such as the Logachev coral carbonate mound province which spans both national EEZ (Ireland) and international waters. Many of these coral frameworks are now protected as VMEs. An area of polygonal faults may be a unique seabed feature. It is currently poorly investigated but may host important biological communities (e.g. cold-seeps). The polygonal faults do not themselves appear to support unique biological communities or species but may be indicative of possible presence of active hydrocarbon seeps. One such active seep has recently been discovered in this area, supporting a rare chemosynthetic community that hosts species that have not been recorded elsewhere (hence the bracketed “High” score). There is support for qualification under this criterion from the occurrence of polygonal faults and an active cold hydrocarbon seep. These features exist within a very restricted area of the site and, as described, the uniqueness and rarity criterion would only apply where these habitats occur. If further information is provided on the occurrence of large areas of cold-water coral reef, this may provide further support for this criterion over a wider geographical area. Areas that are required for a population to survive Special X importance for and thrive. life-history stages of species Explanation for ranking Cold-water corals and areas of natural coral rubble provide highly diverse habitats. Recent observations show that Lophelia pertusa reefs provide nursery grounds for deep-water sharks, and egg cases of deep-water rays were recorded with small patches of Solenosmilia variabilis framework on the Hebrides Terrace Seamount during the RRS James Cook 073 Changing Oceans Expedition in June 2012. New evidence from RRS James Cook cruises 073 and 060 shows that small patch reefs of L. pertusa on Rockall Bank are used as refuge by gravid Sebastes viviparous. Parts of the Hatton–Rockall area are important as spawning areas for blue whiting, and the area is used as a corridor for a range of migrating species, including turtles. Blue whiting has a widespread spawning area from the Faroe–Shetland Channel in the north to the Porcupine Bank in the south. Three areas of blue ling spawning aggregations are known on the shallow parts of Hatton and Lousy banks. These are significant since they represent three of the six known or suspected spawning locations for the southern stock of blue ling. Importance for Areas containing habitats for the survival and X recovery of endangered, threatened, or declining threatened, endangered, or species, or areas with significant assemblages of such species. declining species and/or habitats Explanation for ranking The OSPAR threatened and declining habitats and species “carbonate mounds” and Lophelia pertusa are confirmed to be present in the area, and indicator species for “deep-sea sponge aggregations” and “coral gardens” have been recorded. The presence of these habitats has been confirmed in some areas that are now protected as VMEs. 110 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know The cold-water corals and natural rubble contain very large numbers of invertebrate species, including giant protozoans (xenophyophores), vase-shaped white sponges, actiniarians, antipatharian corals, hydroids, bryozoans, asteroids, ophiuroids, echinoids, holothurians, and crustaceans. The distribution of cold-water coral has been severely reduced in the area over the last 30 years. The deep-water sharks C. coelolepis and C. squamosus are also listed in the OSPAR list. Both occur in the area, but there is no information to indicate that this area is important for either species in the sense of having a significant proportion of the population or higher density than other areas of similar depth in the region. Both Zino’s petrel (endangered) and Fea’s petrel (near threatened) are listed on the IUCN Red List. A further five species of seabirds listed in Annex I of the European Union Bird’s Directive are found within the area. However, tracking for the two petrel species (data in Figure 2 of the proposal; OSPAR/NEAFC/CBD, 2011) appears to show that the area is of relatively low importance (5 to 10% of tracked birds) during a very short period (one month). Knowledge of cetaceans in the area is poor, but the critically endangered northern right whale (Eubalaena glacialis) has been observed in this area. However, this single observation is insufficient to demonstrate importance for this species. Areas that contain a relatively high proportion of Vulnerability, X sensitive habitats, biotopes, or species that are fragility, sensitivity, or functionally fragile (highly susceptible to degradation or depletion by human activity or by slow recovery natural events) or with slow recovery. Explanation for ranking It is uncertain how “a relatively high proportion” is defined in this context, but there is good evidence for vulnerable habitats and benthic species in the area (records of cold-water coral reefs and carbonate mounds, and indicator species for coral gardens, deep-water sponge aggregations). Distribution is not uniform across the area and many of the areas where they occur are now protected as VMEs. There is a high diversity of corals, including bamboo coral (Isididae), black coral (Antipatharia), as well as the reefforming stony corals (Scleractinia), though some of these may now be reduced in distribution and occurring in patches. Cold-water coral habitats are easily impacted and recover very slowly. Some species of cold-water corals can live for more than 4000 years. Many of the demersal fish have life histories of deep-water fish fauna with very slow recovery times as a result of their slow reproductive rate compared to pelagic fish. These fish may be more exposed to fishing pressure because trawlable habitat is more common in this area than is typical at these depths. Stocks have already been diminished in some areas. There is good support for qualification under this criterion from the occurrence of vulnerable marine ecosystems, including stony corals, carbonate mounds, possible coral gardens and deep-sea sponge aggregations, and an active cold seep. Although comparative studies have not been done, it is probable that occurrence of corals in the Rockall–Hatton area is higher than in other areas of comparable depth and latitude. This would therefore constitute a “relatively high proportion”. The species or habitats discussed in this rationale are generally found in depths above 1500 m and thus this proposed EBSA should be limited to this depth contour. Areas containing species, populations, or Biological X communities with comparatively higher natural productivity biological productivity. Explanation for ranking It is likely that pelagic production may be enhanced relative to surrounding areas due to upwelling, but benthic secondary production in deep-water environments is generally considered to be low compared to other environments. Areas containing a comparatively higher diversity Biological X of ecosystems, habitats, communities, or species, diversity or with higher genetic diversity. ICES Advice 2013, Book 1 111 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High Explanation for ranking The area comprises a patchwork of habitats with species changing consistently with both habitat type and increasing depth. Some habitats are threatened by direct impacts (e.g. bottom fishing), others may suffer indirectly (e.g. through the creation of sediment plumes by impacts of fishing gear in sensitive areas). Seabed communities include cold-water corals, rocky reefs, carbonate mounds, polygonal fault systems, sponge aggregations, and steep and gentle sedimented slopes. Cold-water corals provide diverse habitats for other invertebrates and fish. This area spans more than one biogeography province; consequently, overall diversity is likely to be higher than in other areas with comparable depth and habitat range. Rare habitats such as cold seeps and highly diverse habitats such as cold-water coral reef and rubble further contribute to the overall diversity. Proposal for a different configuration of EBSAs than those presented in the OSPAR/NEAFC/CBD Workshop report Proposed EBSAs 1, 2, and 3 (Reykjanes Ridge south of Iceland EEZ, Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge, and Mid-Atlantic Ridge north of the Azores) all encompass the hard substrates running roughly north–south in the higher elevations of the Mid-Atlantic Ridge, and for proposed EBSAs 1 to 3 the southern boundary of each aligns with the northern boundary of the next. The boundaries between them were defined primarily by the extreme limits of the position of the east–west-oriented Subpolar Front, a major pelagic oceanographic feature in proposed EBSA 2 that moves seasonally northward spring and summer) and southward (autumn and winter). The Subpolar Front has affinities with the Charlie-Gibbs Fracture Zone, but not with the Mid-Atlantic Ridge. If the Charlie-Gibbs Fracture Zone, running roughly east to west, and associated Subpolar Frontal Zone were treated separately from the Mid-Atlantic Ridge, then all the areas delineated by the features of the Mid-Atlantic Ridge specified for the proposed EBSAs 1, 2, and 3 would share a consistent geomorphological feature (the emergent hard substrates primarily of volcanic origin) with associated benthic fauna present from the northern boundary of proposed EBSA 1 to the southern boundary of proposed EBSA 3, with the Charlie-Gibbs Fracture Zone itself simply serving as an interruption in this feature. The entire Mid-Atlantic Ridge feature would score “Some” or “High” on several criteria, and for generally the same biological rationales for the entire ridge. The species composition of the benthic biota does change from north to south, but aside from the structural interruption caused by the transverse fractures, there is no strong evidence that discontinuities in benthic community composition exist along the ridge. Thus, an alternative area to proposed EBSAs 1, 2, and 3 can be justified along the entire Mid-Atlantic Ridge in the OSPAR/NEAFC area, defined by the features specified in the description of proposed EBSA 1, and ranked as “Some” or “High” on several EBSA criteria with a common justification for that entire area. A second alternative EBSA could then be proposed, consisting of the Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone, taken together. This area would have its own set of ecological properties and associated rankings on the EBSA criteria. It would be ranked “Some” or “High” on several criteria for justifications specific to the Fracture Zone and Subpolar Front, but in several cases for justifications very different from that of the Mid-Atlantic Ridge. These two potential proposed EBSAs would replace proposed EBSAs 1, 2, and 3 from the OSPAR/NEAFC/CBD Workshop report. It would also require a separate consideration of seabird foraging in the southern third of the area, jointly with the additional analyses already recommended for the OSPAR/NEAFC/CBD Workshop proposed EBSAs 5– 8. Alternative proposed EBSA for the Mid-Atlantic Ridge The Mid-Atlantic Ridge runs from the southern boundary of the Icelandic EEZ to the northern boundary of the Portuguese EEZ in the Azores and includes all area above a depth contour that runs in the deeper of two properties: 1. 2. 112 Including a large portion (arbitrarily, perhaps 90%) of all hard volcanic substrates; habitats are reported to host the larger known coral deposits and their associated communities. Included a large portion (arbitrarily, 90%) of the records of large sponge communities in the overall Mid-Atlantic Ridge. ICES Advice 2013, Book 1 In addition the proposed area should include all known hydrothermal vents along the ridge, if any of these are deeper than the contours meeting the properties above. Evaluation of the proposed area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know The area contains either (i) unique (“the only one of X (X) its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking The qualified “High” ranking is for restricted areas of the few known hydrothermal vents along the ridge. These are globally rare features; only a small part of the area proposed here meets this EBSA criterion. Uniqueness rarity or The MarEco sampling of corals and sponges reported several species new to science as it sampled the Mid-Atlantic Ridge. However, it is not possible to draw a firm conclusion on the presence of unique species along the ridge until more extensive sampling is undertaken. Areas that are required for a population to survive Special X importance for and thrive. life-history stages of species Explanation for ranking Although many populations undoubtedly complete their life cycles within the harder-substrate areas of the MidAtlantic Ridge, this would be true of any marine area of comparable size. There is no evidence that the life history of any species is strongly dependent on any specific features of the area proposed as an alternative EBSA. There is evidence from other areas of the Northeast Atlantic that areas of high coral density may be important as eggcase and nursery areas of deep-water sharks and rays. The area proposed here is targeted on the depths and substrates associated with higher coral and sponge densities, and if sharks and rays also concentrate spawning and early development in these habitats, then the score would be “Some” or “High” on this criterion as well. However, it has not yet been documented that skates and rays do preferentially use the coral and sponge formations for these life history functions in the Mid-Atlantic Ridge. For the more southern portions of the Mid-Atlantic Ridge in particular, there are reports of areas being important for foraging by seabirds, including Cory’s shearwater. The evidence available is not considered strong, however, and this aspect of the ecological functionality of the central ridge area should be considered as part of the reanalysis of EBSAs 5–8 proposed in the OSPAR/NEAFC/CBD Workshop report. Importance for Areas containing habitats for the survival and X recovery of endangered, threatened, or declining threatened, endangered, or species, or areas with significant assemblages of such species. declining species and/or habitats Explanation for ranking There are large deposits of corals and sponges found in the alternative area proposed by ICES as meeting this criterion. Habitats containing these species are listed by OSPAR and are also VME indicator species, and the majority of these would be included in the alternative proposed EBSA. Additional explanation regarding corals and sponges is included in the rationale for the criterion on Vulnerability. The possible role of the alternative proposed Mid-Atlantic Ridge area in the life histories of sharks and rays is discussed in the criterion on Vulnerability and Sensitivity. If a dependency between the breeding or early life history of threatened or endangered skates or rays were documented, there would be additional justification for a “High” ranking of this criterion. ICES Advice 2013, Book 1 113 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know Areas that contain a relatively high proportion of sensitive habitats, biotopes, or species that are functionally fragile (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery. Explanation for ranking Vulnerability, fragility, sensitivity, or slow recovery X With regard to corals and sponges, Mortensen et al. (2008) found cold-water corals “at every sample station … observed at depths between 800 and 2400 m, however were commonly found shallower than 1400 m …, with species richness being very high. … no major reef structures were recorded, with the maximum colony size approximately 0.5 m in diameter. The number of coral taxa was strongly correlated with the percentage cover of hard bottom substrate ….” The area proposed here is targeted at the seamount peaks and slopes where hard substrates dominate. For sponges, no actual large expanses of sponge reef were reported in the OSPAR/NEAFC/CBD Workshop report. However, the pro forma in that report notes that overall sampling of the area is patchy and cites three studies that found local patches with high densities of sponges, although in no cases were the sizes of the patches documented. If research finds that deep-sea sharks and rays use the denser coral deposits as important spawning and nursery grounds, as has been reported in the Hatton–Rockall Bank area proposed by ICES, then there would be additional justification for a score of “Some” on this criterion. Areas containing species, populations, or Biological X communities with comparatively higher natural productivity biological productivity. Explanation for ranking Although benthic productivity of the alternative proposed Mid-Atlantic Ridge EBSA may be higher than benthic productivity on the abyssal plain, productivity integrated over the entire water column and seafloor seems typical of systems of comparable depth and latitude globally. Areas containing comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or diversity with higher genetic diversity. Explanation for ranking The presence of comparatively large coral and sponge formations, and the documented high diversity of benthic and associated fish when corals or sponges are present in moderate or high density imply that some areas along the ridge may have high diversity. From north to south there is a mingling of benthic and demersal fish faunas characteristic of both cooler northern and warmer southern ocean environments, giving the area as a whole a somewhat higher net biological diversity, even if the diversity in any individual site is not markedly enhanced. Aside from the benthic communities of somewhat restricted distribution associated with the biogenic habitats, the biodiversity otherwise appears typical of biotic communities at similar depths and latitude. Alternative proposed EBSA – The Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone The area would include: i. ii. Subpolar Frontal Zone (coinciding with the Charlie-Gibbs Fracture Zone): The northern and southern boundaries for this feature should be set according to the known northernmost and southernmost locations of the frontal system (at approximately 53°N and 48°N). The eastern and western boundaries for this feature should be set according to the eastern and westernmost extension of the Charlie-Gibbs Fracture Zone (at approximately 27°W and 42°W). Charlie-Gibbs Fracture Zone: The eastern and western boundaries for this feature should be set according to the east–west extension of the Fracture Zone (at approximately 27°W and 42°W). The northern and southern seabed boundaries for this feature should be set with a view to encompass the characteristic topography and substrates of the Fracture Zone. Any area of the seafloor and associated benthos that lies below the pelagic feature i) (the total area occupied by the Subpolar Front during its annual movement) but does not meet feature ii) is not included in the area proposed as meeting EBSA criteria. Only those parts of the water column where the Subpolar Front is prominent at some time during the year 114 ICES Advice 2013, Book 1 are proposed as meeting one or more of the EBSA criteria. Moreover, in the entire pelagic area described by i), at any given time only that part of the total area where the Subpolar Front is located would be expected to meet some of the criteria. Although maps will show that the full pelagic area is proposed as part of this complex EBSA, any conservation measures for ecological properties of the water column would need to take into account the position of the Subpolar Front to be fully effective. Evaluation of the proposed area against the EBSA criteria CBD EBSA Criterion Description The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking Uniqueness rarity or Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know X The Charlie-Gibbs Fracture Zone is a set of geomorphological features unique to the entire North Atlantic, and the Subpolar Front is an oceanographic feature also unique to the Northeast Atlantic. Together these features justify a “High” score for this criterion. Areas that are required for a population to survive Special X importance for and thrive. life-history stages of species Explanation for ranking No evidence is available suggesting a significant importance of the area for life-history stages of widespread species in comparison with other marine areas of similar size and depth range. It is possible that there are species with special affinities for the unique geophysical features of the deep faults, but clear documentation of species with such affinities was not found in the references provided by the OSPAR/NEAFC/CBD Workshop report, and was not otherwise known to ICES. Importance for Areas containing habitats for the survival and X recovery of endangered, threatened, or declining threatened, endangered or species, or areas with significant assemblages of such species. declining species and/or habitats Explanation for ranking There is good evidence that the area contains a significant assemblage of species and habitats that are assessed to be threatened, endangered, or declining, including leafscale gulper shark (Centrophorus squamosus), gulper shark (Centrophorus granulosus), Portuguese dogfish (Centroscymnus coelepis), Sei whale (Balaenoptera borealis), sperm whale (Physeter macrocephalus), leatherback turtle (Dermochelys coriacea), as well as cold-water coral reefs and deep-sea sponge aggregations. Depending on the species, the special features of the Fracture Zone and the Subpolar Front are inferred to provide important biological functions to the species which aggregate along each one. Areas that contain a relatively high proportion of Vulnerability, X sensitive habitats, biotopes, or species that are fragility, sensitivity, or functionally fragile (highly susceptible to degradation or depletion by human activity or by slow recovery natural events) or with slow recovery. ICES Advice 2013, Book 1 115 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know Explanation for ranking The Charlie-Gibbs Fracture Zone geophysical structure provides habitats for a number of sensitive/vulnerable species and communities both on soft and hard substrate and in the associated water column. In particular biogenic habitats such as those formed by cold-water corals and sponges are considered vulnerable, are often fragile, and slow (if at all) to recover from damage. Some fish species associated with the Fracture Zone and the Subpolar Front also show slow growth, late maturity, irregular reproduction, and long generation time, as well as community characteristics of high diversity at low biomass. However, the documentation that the species with vulnerable life histories are especially closely affiliated with the Fracture Zone and frontal habitats is weak, and it is clear that these vulnerable species and biogenic habitats are not consistently present throughout the entire Fracture Zone and Subpolar Front. Areas containing species, populations, or Biological X (X) communities with comparatively higher natural productivity biological productivity. Explanation for ranking There is good evidence that, because of the Subpolar Front, the pelagic area where the front is located at any particular time is characterized by an elevated abundance and diversity of many taxa, including an elevated standing stock of phytoplankton. This justifies a ranking of “High” for the pelagic area around the Subpolar Front, as it moves seasonally. However, there is no evidence of relatively elevated productivity in the benthic communities of the Fracture Zone. Areas containing a comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or diversity with higher genetic diversity. Explanation for ranking The area of the Fracture Zone is characterized by a very high structural complexity, offering a diverse range of habitats. The area of the Subpolar Front is a feature where species are documented to assemble seasonally. Consequently, both features characterizing this area contribute to a relatively higher diversity of ecosystems, habitats, communities, and species in comparison to other areas of the Northeast Atlantic. Proposed EBSAs for which there is insufficient scientific justification For five of the ten areas proposed as meeting one or more EBSA criteria in the OSPAR/NEAFC/CBD Workshop report, ICES concluded that there is insufficient scientific justification at this time to propose their delineated area, or any subset of it, as meeting EBSA criteria. In all cases ICES recommends that additional information needs to be collated and analysed, and a new evaluation be conducted when those results are available. ICES provides reasoning for its advice and recommendations for further work in each of these areas below. Area 5. Around the Pedro Nunes and Hugo de Lacerda seamounts and Area 6. Northeast Azores–Biscay Rise The OSPAR/NEAFC/CBD Workshop concluded that both areas ranked as “High” on criterion Special Importance to Life History of Species, and “Some” on criteria Uniqueness and Rarity; Importance to Threatened, Endangered, or Declining Species; and Vulnerability, Sensitivity, etc., the latter primarily for seabirds. ICES questions the basis for these conclusions, noting that the data used to assess all the criteria specified were incomplete and often incorrectly interpreted, with the proposed boundaries not matching the information in the cited sources. ICES recommends that all available data on foraging activity of the Zino’s petrel, Cory’s shearwater, and other relevant species be examined. This should include published and any other available data. Occurrence data may be used as well, provided the rationale details how occurrence and foraging data are used to derive EBSA boundaries. Area 7. Evlanov Seamount region The OSPAR/NEAFC/CBD Workshop concluded that the area they delineated ranked as “High” on criterion Importance to Threatened, Endangered, or Declining Species, and “Some” on criteria Uniqueness and Rarity; Special Importance to Life History of Species; and Vulnerability, Sensitivity, etc., the latter primarily for seabirds. ICES questions the basis for these conclusions, noting that the data used to assess all the criteria specified were incomplete and often incorrectly 116 ICES Advice 2013, Book 1 interpreted, with the proposed boundaries not matching the information in the cited sources. In particular the sample sizes for Fea’s petrel were very small, and the information for sooty shearwater did not seem to differentiate the proposed EBSA area from most of surrounding area. ICES advises not to proceed with proposing any portion of this area as an EBSA at this time, but rather undertake further collation and analysis of information and reconsider when the additional work is completed. Area 8. Northwest of Azores EEZ The OSPAR/NEAFC/CBD Workshop concluded that the area they delineated ranked as “High” on criteria Special Importance for Life History of Stages of Species and Importance for Threatened, Endangered or Declining Species, and “Some” on criteria Uniqueness and Rarity; Vulnerability, Sensitivity, etc.; and Biological Diversity, the latter primarily for seabirds. ICES questions the basis for these conclusions, noting that the data used to assess all the criteria specified were incomplete and often incorrectly interpreted, with very small sample sizes for some of the species’ (e.g. Zino’s petrel) foraging areas, and questionable interpretation of the foraging areas of Cory’s shearwater. ICES does not support this area going forward as presented in the OSPAR/NEAFC/CBD EBSA Workshop report. ICES advises that improvements are needed to the supporting evidence in the narrative for criteria related to Special Importance for Life History Stages of Species and Importance for Threatened, Endangered or Declining Species and/or Habitats. It is necessary to augment information on how the area is being used (feeding, conditioning, migration) for the survival and recovery of the species and to put some scale on its importance to the species (some of which have very restricted breeding sites in the larger area). It is also necessary to document the proportion of species that are highly susceptible to degradation or depletion by human activity, in this case bycatch in longline fisheries. Area 9. The Arctic Front – Greenland/Norwegian seas The OSPAR/NEAFC/CBD Workshop concluded that the area they delineated ranked as “High” on criteria Special Importance to Life History of Species; Importance to Threatened, Endangered, or Declining Species; Biological Productivity; and Biological Diversity. ICES questions the basis for these conclusions, noting that (1) for several features the area proposed as meeting criteria were not noticeably different from the surrounding areas; (2) some of the rankings regarding importance appeared to be inferred from a belief that the area is high in productivity, but the rankings were not demonstrated otherwise; and (3) publications with contrasting conclusions were found for some of the key references cited in the OSPAR/NEAFC/CBD Workshop report. ICES recommends not to proceed with the proposed Arctic Front EBSA. There is no evidence of enhanced productivity at the Arctic Front which is the main rationale used to justify the proposed EBSA. However, there seems to be circumstantial evidence for an enhanced production that may attract feeding animals in the areas around and south of Jan Mayen, including the Jan Mayen Front. If parts of this area are located in the high seas, further analyses should be undertaken to determine if this area meets the EBSA criteria. Sources CBD. 2007. Expert Workshop on Ecological Criteria and Biogeographic Classification Systems for Marine Areas in Need of Protection. 2–4 October 2007, Azores, Portugal. (http://www.cbd.int/doc/meetings/mar/ewsebm01/official/ewsebm-01-02-en.pdf). CBD. 2008. CBD Decision IX/20 on Marine and Coastal Biodiversity. (http://www.cbd.int/doc/decisions/cop-09/cop-09dec-20-en.pdf). CBD. 2009. Expert Workshop on Scientific and Technical Guidance on the Use of Biogeographic Classification Systems and Identification of Marine Areas beyond National Jurisdiction in Need of Protection. 29 September–2 October 2009, Ottawa, Canada. (https://www.cbd.int/doc/meetings/sbstta/sbstta-14/information/sbstta-14-inf-04-en.pdf). CBD. 2010. CBD Decision X/29 on Marine and Coastal Biodiversity. (https://www.cbd.int/decision/cop/default.shtml?id=12295). German, C. R., and Parsons, L. M. 1998. Distributions of hydrothermal activity along the Mid-Atlantic Ridge: interplay of magmatic and tectonic controls. Earth and Planetary Science Letters, 160: 327–341. German, C. R., Briem, J., Chin, C., Danielsen, M., Holland, S., James, R., Jónsdottir, A., Ludford, E., Moser, C., Ólafsson, J., Palmer, M. R., and Rudnicki, M. D. 1994. Hydrothermal activity on the Reykjanes Ridge: the Steinahóll ventfield at 63°06’N. Earth and Planetary Science Letters, 121: 647–654. ICES. 2013a. Vulnerable deep-water habitats in the NEAFC Regulatory Area. In Report of ICES Advisory Committee, 2013, Section 1.5.5.1. ICES Advice, 2013, Book 1. ICES. 2013b. Report of the Workshop to Review and Advise on EBSA Proposed Areas (WKEBSA), 27–31 May 2013, Copenhagen, Denmark. In draft. ICES Advice 2013, Book 1 117 Mironov, A., and Gebruk, A. 2007. Deep-sea benthos of the Reykjanes Ridge: biogeographic analysis of the fauna living below 1000 m. Report on preliminary phase of the project patterns and processes of the ecosystems of the northern Mid-Atlantic (Mar-Eco). (http://www.mareco.no/sci/component_projects/epibenthos/deep-sea_benthos_of_the_reykjanes_ridge). Mortensen, P. B., Buhl-Mortensen, L., Gebruk, A. V., and Krylova, E. M. 2008. Occurrence of deep-water corals on the Mid-Atlantic Ridge based on Mar-Eco data. Deep-Sea Research, 55: 142–152. Olafsson, J., et al. 1991. A sudden cruise off Iceland. RIDGE Events Newsletter, 2(2): 35–38. OSPAR/NEAFC/CBD. 2011. Report of the Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic. 8–9 September 2011, Hyères, France. Annex 11. Søiland, H., Budgell, W. P., and Knutsen, Ø. 2008. The physical oceanographic conditions long the Mid-Atlantic Ridge north of the Azores in June–July 2004. Deep-Sea Research II, 55: 29–44. United Nations. 2004. UNGA Resolution 58/240. (http://daccess-dds-ny.un.org/doc/UNDOC/GEN/N03/508/92/PDF/N0350892.pdf?OpenElement). 118 ICES Advice 2013, Book 1 Annex 1.5.6.5.1 Revised pro forma for the Arctic Ice habitat Annex 1 Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic Hyères (Port Cros), France: 8 – 9 September 2011 ________________________________________ EBSA identification proforma for the North-East Atlantic - 10 Title/Name of the area - The Arctic Ice habitat - multiyear ice, seasonal ice and marginal ice zone Presented by WWF and reviewed by Participants at the Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas in the North-East Atlantic Contact: Sabine Christiansen [email protected] Abstract The permanently ice covered waters of the high Arctic provide a range of globally unique habitats associated with the variety of ice conditions. Multi-year sea ice only exists in the Arctic and although the projections of changing ice conditions due to climate change project a considerable loss of sea ice, in particular multiyear ice, the Eurasian Central Arctic high seas are likely to at least keep the ice longer than many other regions in the Arctic basin. Ice is a crucial habitat and source of particular foodweb dynamics, the loss of which will affect also a number of mammalian and avian predatory species. The particularly pronounced physical changes of Arctic ice conditions as already observed and expected for the coming decades, will require careful ecological monitoring and eventually measures to maintain or restore the resilience of the Arctic populations to quickly changing environmental conditions. Introduction Up until today most of the Eurasian part of the Arctic Basin, and in particular the high seas area in the Arctic Ocean (the waters beyond the 200 nm zones of coastal states, i.e. Norway, Russia, USA, Canada and Greenland/Denmark) is permanently ice covered. However, in recent years, much of the original multiyear pack ice has been replaced by seasonal (1 year) ice which made it possible for research and other vessels to reach the pole. In addition, the former fast pack-ice is now increasingly broken up by leads. This structural change in the Arctic ice quality will result in a substantial increase in light penetrating the thin ice and water column, in conjunction with the overall warming of surface waters and increased temperature and salinity stratification due to the melting of ice. In the near future, up to the end of the century, the permanent ice cover is expected to disappear completely in some models (Anisimov et al., 2007). This will result in significant changes in the structure and dynamics of the high Arctic ecosystems (CAFF, 2010; Gradinger, 1995; Piepenburg, 2005; Renaud et al., 2008; Wassmann, 2008, 2011) which should be closely monitored (Bluhm et al., 2011) as already envisaged by the Arctic Council (Gill et al., 2011; Mauritzen et al., 2011). Therefore, the area proposed here as EBSA is of particular scientific interest and may in the longterm, become relevant for the commercial exploitation of resources. ICES Advice 2013, Book 1 119 Location The Ecologically or Biologically Significant Marine Area (EBSA) proposed focusses on the presently permanently ice-covered waters in the OSPAR/NEAFC maritime areas, including the high seas section in the Central Arctic Basin north of the 200 nm zones of coastal states (see Fig. 1 attached). Therefore, the boundaries proposed extend from the North Pole (northernmost point of OSPAR/NEAFC maritime areas) to the southern limit of the summer sea ice extent and marginal ice zone, including on the shelf of East Greenland. The proposal currently only relates to features of the water column. Two legal states have to be distinguished: the Central Arctic high seas waters north of the 200 nm zones of adjacent coastal states, generally north of 84° N, and the waters within the Exclusive Economic Zones of Greenland, Russia and the fisheries protection zone of Norway around Svalbard. Figure 1 distinguishes between the high seas beyond national jurisdiction for which the „Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic“ has a mandate8, and national/nationally administered waters within the 200 nm zone, within which the OSPAR Contracting Parties have the responsiblity to report candidate EBSAs to the Convention on Biodiversity EBSA repository (OSPAR Commission, 2011). The seafloor of the respective region will likely fall on the extended continental shelves of several coastal states. It belongs to the „Arctic Basin“ region of (Gill et al., 2011). The coordinates of the overall area, as well as the high seas section are provided in Annex 1 (in decimals, shape files provided): c.f. Figure 1: Location of the Arctic Ice „Ecologically or Biologically Significant Area“ (EBSA) proposed by WWF in September 2011. The position of the Arctic and polar fronts was redrawn after (Rey, 2004, Fig. 5.7). Feature description The Ecologically or Biologically Significant Marine Area (EBSA) proposed focusses on the presently permanently ice-covered waters in the OSPAR/NEAFC maritime areas, including the high seas section in the Central Arctic Basin north of the 200 nm zones of coastal states, and the marginal ice zone (where the ice breaks up, also called seasonal ice zone) along its southern margins (see Fig. 1 attached). Due to the inflow of Atlantic water along the shelf of Svalbard, and the concurrent outflow of polar water and ice on the Greenland side of Fram Strait, the southern limit of the summer sea ice extent is much further south in the western compared to the eastern Framstrait, and in former times extended all along the Greenland coast. The high seas section of the OSPAR maritime area in the Central Arctic ocean is generally north of 84° N and is until today fully ice-covered also in summer, although the quantity of multiyear ice has already substantially decreased and the 1-year ice leaves increasingly large leads and open water spaces. The ice overlays a very deep water body of up to 5000 m depth far away from the surrounding continental shelves and slopes of Greenland and the Svalbard archipelago. The Nansen-Gakkel Ridge, a prolongation of the Mid-Atlantic Ridge north of the Fram Strait is structuring the deep Arctic basin in this section, separating the Central Nansen Basin to the south from the Amundsen Basin to the north. Abundant hydrothermal vent sites have been discovered on this ridge at about 85° 38 N (Edmonds et al., 2003). North of Spitsbergen, the Atlantic water of the West Spitsbergen Current enters the Arctic basin as a surface current. At around 83° N, a deep-reaching frontal zone separates the incoming Atlantic and shelf waters from those of the Central Nansen Basin (Anderson et al., 1989), as reflected in ice properties, nutrient concentrations, zooplankton communities, and benthic assemblages (Hirche and Mumm, 1992, and literature quoted). This water subsequently submerges under the less dense (less salinity, lower temperature) polar water and circulates, in opposite direction to the surface waters and ice, counterclockwise along the continental rises until turning south along the Lomonossov Ridge and through Fram Strait as East Greenland Current south to Danmark Strait (Aagaard, 1989; Aagaard et al., 1985). Connecting the more fertile shelves with the 8 Participant Briefing for a Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic. Invitation Annex 2, 2011 120 ICES Advice 2013, Book 1 deep central basin, these modified Atlantic waters supply the waters north of the Nansen-Gakkel Ridge, in the Amundsen basin, with advected organic material and nutrients which supplement the autochtonous production (Mumm et al., 1998). Due to the import of organic biomass from the Greenland Sea and the Arctic continental shelves, part of which may not be kept in the food web due to the polar conditions, the Arctic Ocean may also represent an enormous carbon sink (Hirche and Mumm, 1992). In the Fram Strait, the region between Svalbard to the east and Greenland to the west, the East Greenland Current is the main outflow of polar water and ice from the Arctic Basin (Maykut, 1985) (Aagaard and Coachman, 1968). The polar front (0° C isotherm and 34.5 isohaline at 50 m depth) extends approximately along the continental shelf of Greenland, separating the polar surface water from the Arctic (Intermediate) water and the marginal ice zone to the east (e.g. Aagaard and Coachman, 1968; Paquette et al., 1985). The ice cover is densest in polar water, its extent to the east depends on the wind conditions (compare also Angelen et al., 2011; Wadhams, 1981). The seasonal latitudinal progression of increasing and diminishing light levels, respectively, is the determining factor for the timing of the phytoplankton-related pelagic production. Therefore, the springbloom and ice break up progress from south to north in spring, reaching the Arctic area by about June/July. Because the currents in Fram Strait move in opposite direction, the polar East Greenland Current to the south, and the Atlantic West Spitsbergen Current to the north, there is a delay of about a month between biological spring and summer between the polar and the Atlantic side (Hirche et al., 1991). Therefore sea ice and the effect of melting ice are important determinants of the ecosystem processes all along the East Greenland polar front from the Greenland Sea through Fram Strait to the Arctic Basin (Legendre et al., 1992; Wassmann, 2011). Ice situation The Arctic Ocean develops towards a one-year instead of a multi-year sea-ice system with consequences for the entire ecosystem, including ecosystem shifts, biodiversity loss, for water mass modifications and for its role in the global overturning circulation. At its maximum, sea-ice covers 4.47 million km² in the Arctic Basin (Gill et al., 2011): According to data from ice satellite observations in 1973-76 (NASA, 1987, in (Gill et al., 2011)), permanent ice occupied 70-80% of the Arctic Basin area, and the interannual variability of this area did not exceed 2%. Seasonal ice occupied 6-17% (before the melting period of the mid-1970s). Only in the first decade of the 21st century, the permanent-ice area decreased to 6% in February 2008, concurrent with a rapid increase in seasonal- ice. Whereas multiyear ice used to cover 50-60% of the Arctic, it covered less than 30% in 2008, after a minimum of 10% in 2007. The average age of the remaining multiyear ice is also decreasing from over 20 % being at least six years in the mid- to late 1980s, to just 6% of ice six years old or older in 2008. c.f. Figure 2: Modelled ice age distribution in 1985-2000 (left) compared to February 2008 (right) (CAFF, 2010). This trend is likely to amplify in the coming years, as the net ocean-atmosphere heat output due to the current anomalously low sea ice coverage has approximately trippled compared to previous years, suggesting that the present sea ice losses have already initiated a positive feedback loop with increasing surface air temperatures in the Arctic (Kurtz et al., 2011). About 10% of the sea ice in the Arctic basin is exported each year through Fram Strait into the Greenland Sea (Maykut, 1985) which is therefore major sink for Arctic sea ice (Kwok, 2009). From 2001 to 2005, the summer ice cover was so low on the East Greenland shelf, that it was more of a marginal ice zone (Smith Jr and Barber, 2007), however the subsequent record lows in overall Arctic ice cover brought about an increase in ice cover off Greenland, which minimised the extent of the North East Water Polynia on the East Greenland shelf9, a previously seasonally ice-free stretch of water (Wadhams, 1981). Ice related biota 9 http://www.issibern.ch/teams/Polynya/ ICES Advice 2013, Book 1 121 Allover the Arctic, an inventory of ice-associated biota presently counts over 1000 protists, and more than 50 metazoan species (Bluhm et al., 2011). The regionally very variable ice fauna (depends i.e. on ice age, thickness, origin) consists of sympagic biota living within the caverns and brine channels of the ice, and associated pelagic fauna. The most abundant and diverse sympagic groups of the ice mesofauna in the Arctic seas are amphipods and copepods. Polar cod (Boreogadus saida) and partly Arctic cod (Arctogadus glacialis) are dependent on the sympagic macro- and mesofauna for food, themselves being important food sources for Arctic seals (such as ringed seal Phoca hispida) and birds, for example black guillemots Cephus grylle (Bradstreet and Cross, 1982; Gradinger and Bluhm, 2004 and literature reviewed; Horner et al., 1992; Süfke et al., 1998). The higher the light level in the ice, the higher is the biomass of benthic algae as well as meiofauna and microorganisms within the ice (Gradinger et al., 1991). Decreasing snow cover induces a feedback loop with enhanced algal biomass increasing the heat absorption of the ice which leads to changes in the ice structure, and ultimately the release of algae from the bottom layer (Apollonio, 1961 in Gradinger et al., 1991). Because of the distance to land and shelves, and the thickness and internal structure of the multiyear pack ice over deeper water, this type of ice has a fauna of its own (Carey, 1985; Gradinger et al., 1991). Arctic multiyear ice floes can have very high algal biomasses in the brine channels and in the bottom centimeters which serves as food for a variety of proto- and metazoans, usually smaller than 1 mm, over deep water (Gradinger et al., 1999). In the central Arctic, ice algal productivity can contribute up to 50 % of the total primary productivity, with lower contributions in the sea ice covered margins (Bluhm et al., 2011). In the boundary layer between ice floes and the water column, another specific community exists which forms the link between the ice based primary production and the pelagic fauna (Gradinger, 1995): large visible bands of diatoms hang down from the ice, exploited by amphipods such as Gammarus wilkitzki, and occacionally by water column copepods such as Calanus glacialis, which are important prey of for example polar cod Boreogadus saida. The caverns, wedges and irregularities of the ice provide important shelter from predators for larger ice associated species and provide an essential habitat for these species (Gradinger and Bluhm, 2004). During melt, the entire sympagic ice biota are released into the water column where they may initiate the spring algal plankton bloom (Smith and Sakshaug, 1990) or they may sink to the sea floor and serve as an episodic and first food pulse for benthic organisms before pelagic production begins (Arndt and Pavlova, 2005). In particular the shallow shelves and the shelf slope benthos has been shown to profit of this biomass input, reflected in very rich benthic communities (Klitgaard and Tendal, 2004; Piepenburg, 2005). The role of the polar front and marginal ice zone for the production system Primary production in the Arctic Ocean is primarily determined by light availability, which is a function of ice thickness, ice cover, snow cover, light attenuation), the abundance of both ice algae and phytoplankton, nutrient availability and surface water stratification. Generally, the spring bloom occurs later further north and in regions with a thick ice and snow cover. The current production period in the Arctic Ocean may extend to 120 days per year, with a total annual primary production in the central Arctic Ocean of probably up to 10 g C m-2 (Wheeler et al., 1996). Ice algae start primary production already at relatively low light levels when melting reduces the thickness of the ice and snow cover. Only after the ice breaks up, when melting releases the ice biota into the water column and meltwater leads to surface stratification, a major phytoplankton bloom of a few weeks develops, fuelling the higher trophic foodweb of the Arctic (Gradinger et al., 1999, and literature quoted). The marginal ice zones, i.e. where the ice gets broken up in warmer Atlantic or Arctic water, therefore play an important role in the overall production patterns of the Arctic Ocean. Due to the strong water column stratification and increased light levels involved with the melting of the ice, the location and recession of the ice edge in spring and summer determines the timing and magnitude of the spring phytoplankton bloom, which is generally earlier than in the open water (Gradinger and Baumann, 1991; Smith Jr et al., 1987). Wind- or eddy-induced upwelling in the marginal ice zone, as well as biological regeneration processes replenish the 122 ICES Advice 2013, Book 1 surface nutrient pool and therefore prolong the algal growth period (Gradinger and Baumann, 1991; Smith, 1987). The hydrographic variability explains the patchy patterns of primary and secondary production observed, as well as consequently the patchy occurrence of predators. The polar front separates to some degree the pelagic faunas of the polar and Arctic waters in the Greenland Sea and Fram Strait, each characterised by a few dominant copepod species with different life history strategies (Hirche et al., 1991; see also review in Melle et al., 2005): In polar waters, Calanus glacialis uses under ice plankton production and lipid reserves for initiating its spring reproduction phase, however depending on the phytoplankton bloom for raising its offspring (e.g. Leu et al., 2011). Somewhat later, on the warm side of the polar front in Arctic water, the Atlantic species Calanus finmarchicus uses the ice edge-related phytoplankton bloom for secondary production. Calanus hyperboreus, the third and largest of the charismatic copepod species has its core area of distribution in the Arctic waters of the Greenland Sea (Hirche, 1997; Hirche et al., 2006). Zooplankton of the Arctic Basin Overall zooplankton biomass decreases towards the central Arctic basin, reaching a minimum in the most northerly waters, i.e. the region with permanent ice cover (Mumm et al., 1998). However, investigations in recent years demonstrated increased biomasses compared to studies several decades earlier - possibly a consequence of the decrease in ice thickness and cover which only enabled the investigations to take place from ship board. There is a south-north decrease in zooplankton biomass, with a sharp decline north of 83°N (Hirche and Mumm, 1992), coinciding with differences in the species composition of the biomass-forming zooplankton species. Whereas the southern Nansen basin plankton is dominated by the Atlantic species Calanus finmarchicus, entering the Arctic Basin with the West Spitsbergen Current, the northernmost branch of the North Atlantic current, the Arctic and polar species Calanus hyperboreus and C. glacialis dominate the biomass in the high-Arctic Amundsen and Makarov Basins (Auel and Hagen, 2002; Mumm et al., 1998). The zooplankton species communities generally can be differentiated according to their occurrence in Polar Surface Water (0-50 m, temperature below –1.7°C, salinity less than 33.0), Atlantic Layer (200–900 m; temperature 0.5–1.5°C); salinity 34.5–34.8) and Arctic Deep Water (deeper than 1000 m, temperature -0,5-1° C, salinity > 34.9) (Auel and Hagen, 2002; Grainger, 1989; Kosobokova, 1982). The polar surface community in the upper 50 m of the water column consists of original polar species as well as species emerging from deeper Atlantic waters, alltogether leading to a high abundance and biomass peak in summer. Diversity and biomass are minimal in the impoverished Arctic basin deepwater community (Kosobokova 1982). Apart from a limited exchange with the Atlantic Ocean via the Fram Strait, the central Arctic deep-sea basins are isolated from the rest of the world ocean deepsea fauna. Therefore, the bathypelagic fauna consists of a few endemic Arctic species and some species of Atlantic origin. Due to the separation of the Eurasian and Canadian Basins by the Lomonosov Ridge, significant differences in hydrographic parameters (Anderson et al. 1994) and in the zooplankton composition occur between both basins (Auel and Hagen, 2002). Fish Polar cod, Boreogadus saida, is a keystone species in the ice-related foodwebs of the Arctic. Due to schooling behavior and high energy content polar cod efficiently transfer the energy from lower to higher trophic levels, such as seabirds, seals and some whales (Crawford and Jorgenson, 1993). Seabirds Ice cover is a physical feature of major importance to marine birds in high latitude oceans, providing access to resources, refuge from aquatic predators (Hunt, 1990). As seabirds are dependant on leads between ice floes or otherwise open water to access food, they search for the most productive waters in polynias (places within the ice which are permanently ice free) and marginal ice zones (Hunt, 1990). Here they forage both on the pelagic and sympagic ice-related fauna, especially the early stages of polar cod and the copepods Calanus hyperboreus and C. glacialis. Likely, they benefit of the structural complexity and good visibility of their prey near the ice (Hunt, 1990). ICES Advice 2013, Book 1 123 In the Greenland Sea and Fram Strait, major breeding colonies exist on Svalbard, Greenland and on Jan Mayen, all of these within reach of the seasonally moving marginal ice zone or a polynia (North East Water Polynia on the East Greenland shelf). Breeding seabirds like Little auks (Alle alle), from colonies in the northern Svalbard archipelago feed their offspring with prey caught in the vicinity of the nests, however intermittently travel at least 100 km to the marginal ice zone at 80° N to replenish their body reserves (Jakubas et al., Online 03 June 2011). Therefore, the distance of the marginal ice zone to the colony site is a critical factor determining the breeding success (e.g. Joiris and Falck, 2011). Opportunistically, the birds also use other zooplankton aggregations such as a in a cold core eddy in the Greenland Sea, closer to the nesting site (Joiris and Falck, 2011). A synopsis of seabird data for the period 1974–1993 (Joiris, 2000) showed that the little auk is one of the most abundant species, together with the fulmar Fulmarus glacialis, kittiwake Rissa tridactyla and Brünnich’s guillemot Uria lomvia in the European Arctic seas (mainly the Norwegian and Greenland Seas). In the Greenland Sea and the Fram Strait, little auks represented the main species in polar waters, at the ice edge and in closed pack ice, reaching more than 50% of all bird species (Joiris and Falck, 2011). In spring and autumn, millions of seabirds pass through the area when migrating between their breeding sites on Svalbard or the Russian Arctic and their wintering areas in Canada (Gill et al., 2011). There are several seabird species in the European Arctic which are only met in ice-covered areas, for example the Ivory gull Pagophila eburnea and the Thick-billed guillemot Uria lomvia (see e.g. CAFF, 2010): Both species spend the entire year in the Arctic, and breed in close vicinity to sea ice although Thick-billed guillemots were observed to fly up to 100 km from their colonies over open water to forage at the ice edge (Bradstreet 1979). The relatively rare Ivory gulls are closely associated with pack-ice, favouring areas with 70 – 90% ice cover near the ice edge, where they feed on small fish, including juvenile Arctic cod, squid, invertebrates, macro-zooplankton, carrion, offal and animal faeces (e.g. OSPAR Commission, 2009b). Ivory gulls have a low reproductive rate and breeding only takes place if there is sufficient food, which makes the population highly vulnerable to the effects of climate warming (e.g. OSPAR Commission, 2009b). Thick-billed guillemots are relatively long lived and slow to reproduce and has a low resistance to threats including oil pollution, by-catch in and competition with commercial fisheries operations, population declines due to hunting – particularly in Greenland (OSPAR Commission, 2009c). Ivory gull and Thick-billed guillemots are both listed by OSPAR as being under threat and/or decline, (OSPAR Commission, 2008) and in 2011 recommendations for conservation action were agreed (OSPAR Commission, 2011) which will be implemented in conjunction with the circumpolar conservation actions of CAFF (CAFF, 1996; Gilchrist et al., 2008). Marine mammals Several marine mammal species permanently associate with sea ice in the European Arctic. These include polar bear, walrus, and several seal species: bearded, Erignathus barbatus; ringed, Pusa hispida; hooded, Cystophora cristata; and harp seal Pagophilus groenlandicus. Three whale species also occupy Arctic waters year- round – narwhal, Monodon monoceros; beluga whale, Delphinapterus leucas; and bowhead whale, Balaena mysticetus. Polar bears Ursus maritimus are highly specialized for and dependent on the sea ice habitat and are therefore particularly vulnerable to changes in sea ice extent, duration and thickness. They have a circumpolar distribution limited by the southern extent of sea ice. Three subpopulations of polar bears occur in the European high Arctic: the East Greenland, Barents Sea and Arctic Basin sub-populations, all with an unknown population status (CAFF, 2010). Following the young-of-the-year ringed seal distribution, polar bears are most common close to land and over the shelves, however some also occur in the permanent multi-year pack ice of the central Arctic basin (Durner et al., 2009). Due to low reproductive rates and long lifetime, it is expected that the polar bears will not be able to adapt to the current fast warming of the Arctic and become extirpated from most of their range within the next 100 years (Schliebe et al., 2008). 124 ICES Advice 2013, Book 1 Walrusses, Odobenus rosmarus, inhabit the Arctic ice year-round. They are conservative benthic feeders, diving to 80-100 m depth for scaping off the rich mollusc fauna of the continental shelves, and need ice floes as resting and nursing platform close to their foraging grounds. Walrusses have been subject to severe hunting pressure from the end of the 18th century to the mid 20th century, and are still hunted today in Greenland (NAMMCO). By 1934, the estimated 70000-80000 individuals of the Atlantic population were reduced to 12001300, with none left on Svalbard (Weslawski et al., 2000). Todays relatively small sub-populations on the East Greenland and Svalbard-Franz Josef Land coasts have recently shown a slightly increasing trend, in the latter case reflecting the full protection of the species since the 1950´s (CAFF, 2010; NAMMCO). Apart from their sensitivity to direct human disturbance and pollution, it is expected that walrusses will suffer from the changing ice conditions (location, thickness for being used as haul-out site) as well as changes in ice-related productivity. The Atlantic subspecies of the bearded seal, Erignathus barbatus occurs south of 85° N from the central Canadian Arctic east to the central Eurasian Arctic, but no population estimates exist (Kovacs, 2008b). Because of their primarily benthic feeding habits they live in ice covered waters overlying the continental shelf. They are typically found in regions of broken free-floating pack ice; in these areas bearded seals prefer to use small and medium sized floes, where they haul out no more than a body length from water and they use leads within shore-fast ice only if suitable pack ice is not available (Kovacs, 2008b, and literature quoted). The Arctic ringed seal Pusa (Phoca) hispida hispida has a very large population size and broad distribution, however, there are concerns that future changes of Arctic sea ice will have a negative impact on the population, some of which have already been documented in some parts of the subspecies range (Kovacs et al., 2008). As the other seals, the ringed seal uses sea ice exclusively as their breeding, moulting and resting (haulout) habitat, and feed on small schooling fish and invertebrates. In a co-evolution with one of their main predators, the polar bear, they developed the ability to create and maintain breathing holes in relatively thick ice, which makes them well adapted to living in fully ice covered waters allover the year. The West Ice (or Is Odden) to the west of Jan Mayen, at approx. 72-73° N, in early spring a stretch of more of less fast drift ice, is of crucial importance as a whelping and moulting area for harp seals and hooded seals (summarised e.g. by ICES, 2008). Discovered in the early 18th century, up to 350000 seals (1920s) were killed per year, decimating the populations from an estimated one million individuals in the 1950s (Ronald et al., 1982) to today´s 70000 and 243000 of hooded and harp seals, respectively (Kovacs, 2008a, c). Hooded seal, Cystophora cristata, is a pack ice species, which is dependent on ice as a substrate for pupping, moulting, and resting and as such is vulnerable to reduction in extent or timing of pack ice formation and retreat, as well as ice edge related changes in productivity (Kovacs, 2008a, and literature quoted). Hooded Seals feed on a wide variety of fish and invertebrates, including species that occur throughout the water column. After breeding an moulting on the West Ice they follow the retreating pack ice to the north, but also spend significant periods of time pelagically, without hauling out (Folkow and Blix 1999) in (Kovacs, 2008a). The northeast Atlantic breeding stock has declined by 85-90 % over the last 40-60 years. The cause of the decline is unknown, but very recent data suggests that it is on-going (30% within 8 years), despite the protective measures that have been taken in the last few years. The species is therefore considered to be vulnerable (Kovacs, 2008a). Harp seals Pagophilus (Phoca) groenlandicus are the most numerous seal species in the Arctic seas. Their reproduction takes place in huge colonies, for example on the pack ice of the ‘‘West Ice’’ north of Jan Mayen, and after the breeding season they follow the retreating pack ice edge northwards up to 85° N, feeding mainly on polar cod under the ice (Kovacs, 2008c) . Narwhals Monodon monoceros primarily inhabit the ice-covered waters of the European Arctic, including the ice sheet off East Greenland (Jefferson et al., 2008b). For two months in summer, they visit the shallow fjords of East Greenland, spending all the rest of the year offshore, in deep ice-covered waters along the continental slope in the Greenland Sea and Arctic Basin (Heide-Jørgensen and Dietz, 1995). Narwhals are deep diving benthic feeders and forage on fish, squid, and shrimp, especially Arctic fish species, such as Greenland halibut, Arctic cod, and polar cod at up to 1500 m depth and mostly in winter. A recent assessment of the sensitivity of ICES Advice 2013, Book 1 125 all Arctic marine mammals to climate change ranked the narwhal as one of the three most sensitive species, primarily due to its narrow geographic distribution, specialized feeding and habitat choice, and high site fidelity (Laidre et al. 2008 in (Jefferson et al., 2008b)). Bowhead whales Balaena mysticetus are found only in Arctic and subarctic regions and a Svalbard-Barents population occurs from the coast of Greenland across the Greenland Sea to the Russian Arctic. They spend all of their lives in and near openings in the pack ice feeding on small to medium-sized zooplankton. They migrate to the high Arctic in summer, and retreat southward in winter with the advancing ice edge (Moore and Reeves 1993 in (Reilly et al., 2008)). Whaling has decimated the original bowhead whale populations to be rare nowadays, listed by OSPAR as being under threat and/or decline (OSPAR Commission, 2008). The species is considered to be very sensitive to changes in the ice-related ecosystem as well as sound disturbance, possible consequences of a progressive reduction of ice cover (OSPAR Commission, 2009a). Belugas Delphinapterus leucas prefer coastal and continental shelf waters with a broken-up ice cover. They have never been surveyed around Svalbard. Pods numbering into the thousands are sighted irregularly around the archipelago, and pods ranging from a few to a few hundred individuals are seen regularly (Gjertz and Wiig 1994; Kovacs and Lydersen 2006 in (Jefferson et al., 2008a)). Little is known about the populations of the larger fauna in the Central Arctic Basin over the deepsea basins and ridges. But it is not likely that it is currently an area of great abundance - too far from the coastal nesting sites of marine birds, and over too deep water to allow feeding on benthos, as most of the larger mammals would need, and currently of too low plankton production to feed the large whales. All of these groups have their distribution center along the continental shelves presently - however, following the receeding ice edge out to the central Arctic basin may be one of the options for the future. Feature condition, and future outlook This high Arctic region is particularly vulnerable to the the loss of ice cover and other effects of the anticipated global warming, including elevated UV radiation levels (Agustí, 2008). (Wassmann et al., 2010) summarise what changes may be expected within the subarctic/Arctic region: • northward displacement (range shifts) of subarctic and temperate species, and cross-Arctic transport of organisms; • increased abundance and reproductive output of subarctic species, decline and reduced reproductive success of some Arctic species associated with the ice and species now preyed upon by predators whose preferred prey have declined; • increased growth of some subarctic species and primary producers, and reduced growth and condition of animals that are bound to, associated with, or born on the ice; • anomalous behaviour of ice-bound, ice-associated, or ice-born animals with earlier spring events and delayed fall events; • changes in community structure due to range shifts of predators resulting in changes in the predator– prey linkages in the trophic network. (Wassmann, 2008) expects radical changes in the productivity, functional relationships and biodiversity of the Arctic Ocean. He suggests that a warmer climate with less ice cover will result in greater primary production, a reduction of the stratified water masses to the south, changes in the relationship between biological processes in the water column and the sediments, a reduction in niches for higher trophic levels and a displacement of Arctic by boreal species. On the shelves, increased sediment discharges are expected to lower the primary production due to higher turbidity, and enhance the organic input to the deep ocean. A more extensive review of expected or suspected consequences of climate change for the marine system of the Arctic is given in (Loeng et al., 2005). Figure 3, extracted from (Gill et al., 2011), presents the conceptual ideas about possible Arctic ecosystem changes mediated by human impact: 126 ICES Advice 2013, Book 1 The normal situation shown in the upper left panel consists of ice-dependent species and species that tolerate a broader range of temperatures and are found in waters with little or no sea ice. Primary production occurs in phytoplankton (small dots in the figure) in ice-free waters and in ice-attached algae and phytoplankton in icecovered waters. Phytoplankton (small t-shaped symbols in the figure) and ice algae are the main food sources for zooplankton and benthic animals. The fish community consists of both pelagic and demersal species. Several mammals are ice-associated, including polar bears and several species of seals. A number of sea bird species are also primarily associated with ice-covered waters. At moderate temperature increases (upper right) populations of ice-dependent species are expected to decline as sea ice declines, and sub-Arctic species are expected to move northwards. Arctic benthic species are expected to decline, especially if their distributions are pushed close to or beyond the continental slope. The expected effects from fisheries relate to the continental shelves. Two major effects are reductions in populations of benthic organisms due to disturbance from bottom trawling and removal of large individuals in targeted fish stocks. In addition, the size of targeted stocks, both demersal and pelagic, may be reduced. In addition, the effects of ocean acidification are considered (lower right). Ocean acidification will result in depletion of carbonate phases such as aragonite and calcite. This will alter the structure and function of calcareous organisms, particularly at lower trophic levels. Changes in pH can also alter metabolic processes in a range of organisms. It is not known how these changes will propagate to higher trophic levels, but the effects could be substantial. c.f. Figure 3: Conceptual models showing potential impacts on Arctic marine ecosystems under different scenarios (Gill et al., 2011). (Gill et al., 2011) conclude that the central part of the Arctic Basin is not a region for fisheries or oil and gas exploration. However, this region has played and will continue to play a very important role in the redistribution of pollutants, due to ice drift and/or currents between coastal and shelf areas and the Arctic Basin peripheries, far from sources of pollution. ICES Advice 2013, Book 1 127 Assessment against CBD EBSA Criteria Table 1. relation of each of the CBD criteria to the proposed area relating to the best available science. Note that a candidate EBSA may qualify on the basis of one or more of the criteria, the boundaries of the EBSA need not be defined with exact precision. CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Uniqueness or The area contains either (i) unique (“the only one rarity of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Low Some High x Explanation for ranking Arctic sea ice, in particular the multiyear ice of the Central Arctic is globally unique and hosts endemic species such as the Gammarid amphipod Gammarus wilkitzki and sea ice meiofauna which will disappear with the melting of the ice. Polar bears, walrusses, bowhead whales, narwhales, belugas, several seal species and many bird species are endemic to the high Arctic ice. While sea ice species such as G. wilkitzki are not endemic to the proposed EBSA they are endemic to the Arctic and unique within the OSPAR area Special Areas that are required for a population to importance for survive and thrive life-history stages of species x Explanation for ranking Sea ice is essential for its sympagic fauna, and to some extent also for the pelagic associated fauna which also depends on the right timing of biomass production (match/mismatch with bloom periods). The marginal ice zone and other openings in the ice are essential feeding grounds for a large number of ice-associated species which exploit the seasonallly high production there. At present the area covered by the proposed EBSA is ice-covered throughout the summer but although there is no marginal ice zone there will be an ice zone community present, thus the sea ice is essential to maintain the sympagic biological community and associated ecosystem functions. 128 ICES Advice 2013, Book 1 Importance for threatened, endangered or declining species and/or habitats Area containing habitat for the survival and recovery of endangered, threatened, declining species or area with significant assemblages of such species x Explanation for ranking The high arctic ice hosts endemic species such as the Gammarid amphipod Gammarus wilkitzki and sea ice meiofauna which will disappear with the melting of the ice. Many of the obligatory ice-related species are listed as vulnerable by IUCN, and/or listed as under threat and/or decline by OSPAR, examples include the Ivory gull, thick-billed guillemot, bowhead whale, hooded seal and polar bear. With the overall trend of retreating sea ice extent, the proposed EBSA may become increasingly important for all ice-dependent species in the future. Vulnerability, fragility, sensitivity, or slow recovery Areas that contain a relatively high proportion of sensitive habitats, biotopes or species that are functionally fragile (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery x Explanation for ranking The ice-related foodweb and ecosystem is highly sensitive to the ecological consequences of a warming climate. Beyond this the Arctic is at the forefront of the impacts of ocean acidification (Wicks & Roberts 2012). The largest changes in ocean pH will occur in the Arctic Ocean, with complete undersaturation of the Arctic Ocean water column predicted before the end of this century (Steinacher et al. 2009). Many of the seabird and mammal populations are particularly sensitive to changes due to their already low population numbers, and low fertility. If the retreat of the ice to the north will lead to increased shipping and oil and gas exploitation in Arctic waters, the increased risk of spills would also pose a potential hazard for example for guillemots, which are extremely susceptible to mortality from oil pollution (CAFF, 2010). In addition, some species like Ivory gull are sensitive to an increased heavy metal load in their prey. Biological productivity Area containing species, populations or communities with comparatively higher natural biological productivity Explanation for ranking This criterion was not evaluated in the OSPAR/NEAFC/CBD Workshop. ICES did not have enough information to evaluate this criterion. Biological diversity Area contains comparatively higher diversity of ecosystems, habitats, communities, or species, or has higher genetic diversity ICES Advice 2013, Book 1 129 Explanation for ranking This criterion was not evaluated in the OSPAR/NEAFC/CBD Workshop. ICES did not have enough information to evaluate this criterion. References Aagaard, K., 1989. A synthesis of Arctic Ocean circulation. Rapport Proces et Verbeaux Réunion du Conseil international pour l'Exploration de la Mer 188, 11-22. Aagaard, K., Coachman, L.K., 1968. The East Greenland Current north of Denmark Strait: Part II. Arctic 21, 267-290. Aagaard, K., Swift, J.H., Carmack, E.C., 1985. Thermohaline circulation in the Arctic mediterranean seas. Journal of Geophysical Research 90, 4833-4846. Agustí, S., 2008. Impacts of increasing ultraviolet radiation on the polar oceans. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.) Fundación BBVA pp. 25-46. Anderson, L.G., Jones, E.P., Koltermann, K.P., Schlosser, P., Swift, J.H., Wallace, D.W.R., 1989. The first oceanographic section across the Nansen Basin in the Arctic Ocean. Deep Sea Research 36, 475482. Angelen, J.H.v., Broeke, M.R.v.d., Kwok, R., 2011. The Greenland Sea Jet: A mechanism for wind‐driven sea ice export through Fram Strait. Geophysical Research Letters 38 (L12805). Anisimov, O.A., Vaughan, D.G., Callaghan, T.V., Furgal, C., Marchant, H., Prowse, T.D., Vilhjálmsson, H., Walsh, J.E., 2007. Polar regions (Arctic and Antarctic). In: Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Parry, M.L., Canziani, O.F., Palutikof, J.P., Linden, P.J.v.d., Hanson, C.E. (Eds.)Cambridge University Press, Cambridge pp. 653-685. Arndt, C., E. , Pavlova, O., 2005. Origin and fate of ice fauna in the Fram Strait and Svalbard area. Marine Ecology Progress Series 301, 55-66. Auel, H., Hagen, W., 2002. Mesozooplankton community structure, abundance and biomass in the central Arctic Ocean. Marine Biology 140, 1013-1021. Bluhm, B.A., Gebruk, A.V., Gradinger, R., Hopcroft, R.R., Huettmann, F., Kosobokova, K.N., Sirenko, B.I., Weslawski, J.M., 2011. Arctic marine biodiversity: An update of species richness and examples of biodiversity change. Oceanography 24 (3), 232-248. Bradstreet, M.S.M., Cross, W.E., 1982. Trophic relationships at high Arctic ice edges. Arctic 35 (1), 1-12. CAFF, 1996. International Murre conservation strategy and action plan. CAFF International Secretariat, CAFF Circumpolar Seabird Working Group, Akureyri, Iceland, pp. 1-16. CAFF, 2010. Arctic Biodiversity Trends 2010. Selected indicators of change. CAFF International Secretariat, , Akureyri, Iceland. Carey, A.G.I., 1985. Marine Ice Fauna. In: Arctic Sea Ice Biota. A., H.R. (Ed.)CRC Press, Boca Raton. Florida pp. 17-190 Crawford, R.E., Jorgenson, J.K., 1993. Schooling behaviour of arctic cod, Boreogadus saida in relation to drifting pack ice. Environmental Biology of Fishes 36 (4), 345-357. Durner, G.M., Douglas, D.C., Nielson, R.M., Amstrup, S.C., McDonald, T.L., Stirling, I., Mauritzen, M., Born, E.W., Wiig, Ø., Deweaver, E., Serreze, M.C., Belikov, S.E., Holland, M.M., Maslanik, J., Aars, J., 130 ICES Advice 2013, Book 1 Bailey, D.A., Derocher, A.E., 2009. Predicting 21st-century polar bear habitat distribution from global climate models. Ecological Monographs 79 (1), 25-58. Edmonds, H.N., Michael, P.J., Baker, E.T., Connelly, D.P., Snow, J.E., Langmuir, C.H., Dick, H.J.B., Mühe, R., German, C.R., Graham, D.W., 2003. Discovery of abundant hydrothermal venting on the ultraslowspreading Gakkel ridge in the Arctic Ocean. Nature 421, 252-256. Gilchrist, G., Strøm, H., Gavrilo, M.V., Mosbech, A., 2008. International Ivory Gull conservation strategy and action plan. CAFF International Secretariat, Circumpolar Seabird Group (CBird). CAFF Technical Report No. 18. Gill, M.J., Crane, K., Hindrum, R., Arneberg, P., Bysveen, I., Denisenko, N.V., Gofman, V., Grant-Friedman, A., Gudmundsson, G., Hopcroft, R.R., Iken, K., Labansen, A., Liubina, O.S., Melnikov, I.A., Moore, S.E., Reist, J.D., Sirenko, B.I., Stow, J., Ugarte, F., Vongraven, D., Watkins, J., 2011. Arctic Marine Biodiversity Monitoring Plan (CBMP-MARINE PLAN), CAFF Monitoring Series Report No.3, April 2011. CAFF International Secretariat,, Akureyri, Iceland. Gradinger, R., 1995. Climate change and biological oceanography of the Arctic Ocean. Phil. Trans. R. Soc. A 352, 277-286. Gradinger, R., Bluhm, B.A., 2004. In situ observations on the distribution and behavior of amphipods and Arctic cod (Boreogadus saida) under the sea ice of the high Arctic Canadian Basin. Polar Biology 27, 595-603. Gradinger, R., Friedrich, C., Spindler, M., 1999. Abundance, biomass and composition of the sea ice biota of the Greenland Sea pack ice. Deep Sea Research 46, 1457-1472. Gradinger, R., Spindler, M., Henschel, D., 1991. Development o Arctic sea-ice organisms under graded snow cover. In: Proceedings of the Pro Mare Symposium on Polar Marine Ecology. Sakshaug, E., E., H.C.C., Øritsland, N.A. (Eds.), Polar Research 10 (1), Trondheim pp. 295-307. Gradinger, R.R., Baumann, M.E.M., 1991. Distribution of phytoplankton communities in relation to the largescale hydrographical regime in the Fram Strait. Mar. Biol. 111, 311-321. Grainger, E.H., 1989. Vertical distribution of zooplankton in the central Arctic Ocean. In: Proc 6th Conf Comite´Arctique Int 1985. Rey, L., Alexander, V. (Eds.) Brill Leiden pp. 48–60. Heide-Jørgensen, M.P., Dietz, R., 1995. Some characteristics of narwhal, Monodon monoceros, diving behaviour in Baffin Bay. Canadian Journal of Zoology 73, 2106-2119. Hirche, H.J., 1997. Life cycle of the copepod Calanus hyperboreus in the Greenland Sea. Marine Biology 128 (4), 607-618. Hirche, H.J., Baumann, M.E.M., Kattner, G., Gradinger, R., 1991. Plankton distribution and the impact of copepod grazing on primary production in Fram Strait, Greenland Sea. Journal of Marine Systems 2 (3-4), 477-494. Hirche, H.J., Mumm, N., 1992. Distribution of dominant copepods in the Nansen Basin, Arctic Ocean, in summer. Deep Sea Research Part A. Oceanographic Research Papers 39 (2, Part 1), S485-S505. Hirche, H.J., Muyakshin, S., Klages, M., Auel, H., 2006. Aggregation of the Arctic copepod Calanus hyperboreus over the ocean floor of the Greenland Sea. Deep Sea Research Part I: Oceanographic Research Papers 53 (2), 310-320. Horner, R., Ackley, S.F., Dieckmann, G.S., Gulliksen, B., Hoshiai, T., Legendre, L., Melnikov, I.A., Reeburgh, W.S., Spindler, M., Sullivan, C.W., 1992. Ecology of sea ice biota. Habitat, terminology, and methodology. Polar Biology 12 (3), 417-427. Hunt, G.L.J., 1990. The pelagic distribution of marine birds in a heterogeneous environment. Polar Research 8, 43-54. ICES, 2008. Report of the ICES Advisory Committee In: ICES Advice, Book 3, The Barents and the Norwegian SEa. ICES Advice 2013, Book 1 131 Jakubas, D., Iliszko, L., Wojczulanis-Jakubas, K., Stempniewicz, L., Online 03 June 2011. Foraging by little auks in the distant marginal sea ice zone during the chick-rearing period. Polar Biology, 1-9. Jefferson, T.A., Karczmarski, L., Laidre, K., O’Corry-Crowe, G., Reeves, R.R., Rojas-Bracho, L., Secchi, E.R., Slooten, E., Smith, B.D., Wang, J.Y., Zhou, K., 2008a. Delphinapterus leucas In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Jefferson, T.A., Karczmarski, L., Laidre, K., O’Corry-Crowe, G., Reeves, R.R., Rojas-Bracho, L., Secchi, E.R., Slooten, E., Smith, B.D., Wang, J.Y., Zhou, K., 2008b. Monodon monoceros. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Joiris, C., Falck, E., 2011. Summer at-sea distribution of little auks Alle alle and harp seals Pagophilus (Phoca) groenlandica in the Fram Strait and the Greenland Sea: impact of small-scale hydrological events. Polar Biology 34 (4), 541-548. Joiris, C.R., 2000. Summer at-sea distribution of seabirds and marine mammals in polar ecosystems: a comparison between the European Arctic seas and the Weddell Sea, Antarctica. Journal of Marine Systems 27, 267-276. Klitgaard, A.B., Tendal, O.S., 2004. Distribution and species composition of mass occurrences of large-sized sponges in the northeast Atlantic. Progress in Oceanography 61, 57-98. Kosobokova, K.N., 1982. Composition and distribution of the biomass of zooplankton in the central Arctic Basin. Oceanology 22, 744-750. Kovacs, K., 2008a. Cystophora cristata. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Kovacs, K., 2008b. Erignathus barbatus. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Kovacs, K., 2008c. Pagophilus groenlandicus. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Kovacs, K., Lowry, L., Härkönen, T., 2008. Pusa hispida. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Kwok, R., 2009. Outflow of Arctic Ocean sea ice into the Greenland and Barents seas: 1979-2007. Journal of Climate 22, 2438-2456. Legendre, L., Ackley, S.F., Dieckmann, G.S., Gulliksen, B., Horner, R., Hoshiai, T., Melnikov, I.A., Reeburgh, W.S., Spindler, M., Sullivan, C.W., 1992. Ecology of sea ice biota. Polar Biology 12 (3), 429-444. Leu, E., Søreide, J.E., Hessen, D.O., Falk-Petersen, S., Berge, J., 2011. Consequences of changing sea ice cover for primary and secondary producers in the European Arctic shelf seas: timing, quantity, and quality. Progress in Oceanography 90, 18-32. Loeng, H., Brander, K., Carmack, E.C., Denisenko, S., Drinkwater, K., Hansen, B., Kovacs, K., Livingston, P., McLaughlin, F., Sakshaug, E., 2005. Marine systems. In: Arctic Climate Impact Assessment, ACIA. Symon, C., Arrisand, L., Heal, B. (Eds.),Cambridge University Press, Cambridge pp. 453-538. Mauritzen, C., Hansen, E., Andersson, M., Berx, B., Beszczynska-Möller, A., Burud, I., Christensen, K.H., Debernard, J., de Steur, L., Dodd, P., Gerland, S., Godøy, Ø., Hansen, B., Hudson, S., Høydalsvik, F., Ingvaldsen, R., Isachsen, P.E., Kasajima, Y., Koszalka, I., Kovacs, K.M., Køltzow, M., LaCasce, J., Lee, C.M., Lavergne, T., Lydersen, C., Nicolaus, M., Nilsen, F., Nøst, O.A., Orvik, K.A., Reigstad, M., Schyberg, H., Seuthe, L., Skagseth, Ø., Skar∂hamar, J., Skogseth, R., Sperrevik, A., Svensen, C., Søiland, H., Teigen, S.H., Tverberg, V., Wexels Riser, C., 2011. Closing the loop - Approaches to monitoring the state of the Arctic Mediterranean during the International Polar Year 2007-2008. Progress in Oceanography 90 (1-4), 62-89. 132 ICES Advice 2013, Book 1 Maykut, G.A., 1985. The ice environment. In: Sea-ice biota. Horner, R. (Ed.)CRC Press, Boca Raton pp. 2182. Melle, W., Ellertsen, B., Skjoldal, H.R., 2005. Zooplankton: The link to higher trophic levels. In: The Nordic Seas: An integrated perspective oceanography, climatology, biogeochemistry, and modelling. . Drange, H., Dokken, T., Furevik, T., Gerdes, R., Berger, W. (Eds.)Geophysical Monograph Series 158 pp. 137-202. Mumm, N., Auel, H., Hanssen, H., Hagen, W., Richter, C., Hirche, H.J., 1998. Breaking the ice: large-scale distribution of mesozooplankton after a decade of Arctic and transpolar cruises. Polar Biology 20 (3), 189-197. NAMMCO, The Atlantic Walrus. North Atlantic Marine Mammal Commission. Status of Marine Mammals in the North Atlantic, Tromsø, pp. 1-7. OSPAR Commission, 2008. OSPAR List of Threatened and/or Declining Species andHabitats. Reference number 2008-6. http://www.ospar.org/documents/dbase/decrecs/agreements/0806e_ospar%20list%20species%20and%20habitats.doc. OSPAR Commission, 2009a. Background Document for Bowhead whale Balaena mysticetus. OSPAR Commission, Biodiversity Series 494/2010, pp. 1-20. OSPAR Commission, 2009b. Background Document for Ivory gull Pagophila eburnea. OSPAR Commission, Biodiversity Series 410/2009, pp. 1-16. OSPAR Commission, 2009c. Background Document for Thick-billed murre Uria lomvia. OSPAR Commission, Biodiversity Series 416/2009, pp. 1-20. OSPAR Commission, 2011. Meeting of the OSPAR Commission (OSPAR) London: 20-24 June 2011. Summary Record OSPAR 11/20/1-E. OSPAR Commission, London. Paquette, R., Bourke, R., Newton, J., Perdue, W., 1985. The East Greenland Polar Front in autumn. Journal of Geophysical Research 90 (C3), 4866-4882. Piepenburg, D., 2005. Recent research on Arctic benthos: common notions need to be revised. Polar Biology 28 (10), 733-755. Reilly, S.B., Bannister, J.L., Best, P.B., Brown, M., , Brownell Jr., R.L., Butterworth, D.S., Clapham, P.J., Cooke, J., Donovan, G.P., Urbán, J., Zerbini, A.N., 2008. Balaena mysticetus. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Renaud, P.E., Caroll, M.L., Ambrose, W.G.J., 2008. Effects of global warming on Arctic seafloor communities and its consequences for higher trophic levels. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.)Fundación BBVA pp. 141-177. Rey, F., 2004. Phytoplankton: the grass of the sea. In: The Norwegian Sea Ecosystem. Skjoldal, H.R. (Ed.)Tapir Academic Press, Trondheim, Norway pp. 97-136. Ronald, K., Healey, P.J., Fisher, H.D., 1982. The harp seal, Pagophilus groenlandicus. In: Small cetaceans, seals, sirenians and otters. FAO Fisheries Series No. 5, Vol. IV, Food and Agriculture Organisation of the United Nations. Workding Party on Mammals. Schliebe, S., Wiig, Ø., Derocher, A.E., Lunn, N., 2008. Ursus maritimus. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Smith Jr, W.O., Barber, D., 2007. Polynyas and climate change: a view to the future. In: Polynays, windows to the world. Halpern, D. (Ed.), Elsevier Oceanography Serie 74, Elsevier, Amsterdam pp. 411-420. Smith Jr, W.O., Baumann, M.E.M., Wilson, D.L., Aletsee, L., 1987. Phytoplankton biomass and productivity in the Marginal Ice Zone of the Fram Strait during summer 1984. Journal of Geophysical Research 92 (C7), 6777-6786. ICES Advice 2013, Book 1 133 Smith, W.O.J., 1987. Phytoplankton dynamics in the marginal ice zones. Oceanography and Marine Biology Annual Review 25, 11-38. Smith, W.O.J., Sakshaug, E., 1990. Polar phytoplankton. In: Polar oceanography. Part B. Chemistry, biology and geology. Smith Jr, W.O. (Ed.) Academic Press, San Diego pp. 477-525. Steinacher, M., Joos, F., Frölicher, T., Plattner, G. & Doney, S.c. 2009. Imminent ocean acidification in the Arctic projected with the NcAR global coupled carbon cycle-climate model. Biogeosciences 6, 515– 533. Süfke, L., Piepenburg, D., Dorrien, C.C.v., 1998. Body size, sex ratio and diet composition of Arctogadus glacialis (Peters, 1874) (Pisces: Gadidae) in the Northeast Water Polynya (Greenland). Polar Biology 20, 357-363. Wadhams, P., 1981. The ice cover in the Greenland and Norwegian Seas. Rev. Geophys. Space Physics 19, 345-393. Wassmann, P., 2008. Impacts of global warming on Arctic pelagic ecosystems and processes. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.)Fundación BBVA pp. 113-148. Wassmann, P., 2011. Arctic marine ecosystems in an era of rapid climate change. Progress in Oceanography 90, 1-17. Wassmann, P., Duarte, C.M., Agustí, S., Seijr, M., 2010. Footprints of climate change in the Arctic Marine Ecosystem. Biological Global Change. Weslawski, J.M., Hacquebord, L., Stempniewicz, L., Malinga, M., 2000. Greenland whales and walruses in the Svalbard food web before and after exploitation. Oceanologia 42 (1), 37-56. Wheeler, P.A., Gosselin, M., Sherr, E., Thibault, D., Kirchman, D.L., Benner, R., Whitledge, T.E., 1996. Active cycling of organic carbon in the central Arctic Ocean. Nature 380, 697-699. Wicks, L., Roberts, J.M. (2012) Benthic invertebrates in a high CO2 world. Oceanography & Marine Biology: An Annual Review 50: 127 -188 Maps and Figures 134 ICES Advice 2013, Book 1 Figure 1: Location of the Ecologically or biologically significant areas (EBSA) proposed by WWF in September 2011. The position of the Arctic and polar fronts was redrawn after (Rey, 2004, Fig. 5.7). ICES Advice 2013, Book 1 135 Figure 2: Modelled ice age distribution in 1985-2000 (left) compared to February 2008 (right) (CAFF, 2010). Figure 3: Conceptual models showing potential impacts on Arctic marine ecosystems under different scenarios (Gill et al., 2011). 136 ICES Advice 2013, Book 1 1.5.5.6 Special request, Advice June 2013 ECOREGION SUBJECT General advice OSPAR/NEAFC special request on existing and potential new management measures for ecologically and biologically significant areas (EBSAs) Advice summary ICES provides a summary of management measures already implemented within ecologically and biologically significant areas (EBSAs). ICES notes that there are numerous measures already in place by both NEAFC and by OSPAR (as requests to Contracting Parties) and by other competent management authorities, and that both generic and targeted management measures may offer protection to the features that would qualify as meeting EBSA criteria. The performance of these measures has not been evaluated and it was not possible to assess whether these measures are sufficient to protect all of the features that would qualify under EBSA criteria from all potential threats, or whether further measures are required. ICES notes that some specific areas may need further protective measures. Request c) For the use of OSPAR and NEAFC Contracting Parties, as appropriate, provide a separate document with additional relevant information and up-to-date accounts of the relevant management measures already implemented within the individual EBSAs. d) In a separate document, describe potential new management measures within individual EBSAs if appropriate. ICES advice ICES summarized the existing management measures or recommendations of bodies with regulatory or coordinating authority for the major sectoral activities in the OSPAR and NEAFC areas, and, where they exist, those of the authorities for conservation and protection of special components of biodiversity. Some of the measures apply directly, while others require implementing legislation by national authorities (or in some cases by the EU). The power to require vessels to operate in specified ways lies with the Flag State of each vessel. ICES does not have information on the performance of many of the existing measures, and consequently has not been able to evaluate whether these management measures would be sufficient to protect all of the features that would qualify under EBSA criteria from all potential threats. Nonetheless a range of generic and spatially targeted protective management measures already exist within the EBSA areas. There are, however, some exceptions that may need improved protective measures: • • • Josephine Seamount The Hatton–Rockall basin The existing fishing areas that lie north of the Azores on the Mid-Atlantic Ridge. One of these is a potential threat to hydrothermal vents in the area. ICES advises that a systematic review of the performance of the existing measures and how they interact to reduce threats to EBSAs is needed before any new management measures could be sensibly considered through a gap analysis. ICES makes the following general suggestions that are likely to improve the protection of EBSAs in the Northeast Atlantic: • • • • Protective measures for vulnerable marine ecosystems (VMEs) should be made permanent. In many cases these measures are only temporary. This is not appropriate for the long-lived, sessile organisms within VMEs. Accessibility of existing ecological and fisheries data should be improved through a concerted effort between relevant authorities. Coordination of competent authorities should be improved in the selection and adoption of measures in order to help ensure efficient implementation. Observer coverage of fishing vessels operating within EBSAs should be increased. Management measures North-East Atlantic Fisheries Commission (NEAFC) Table 1.5.6.6.1 summarizes NEAFC regulations for fishing activities within nine of the ten proposed EBSAs. The Arctic ice habitat lies outside the regulatory area of NEAFC. The ten proposed EBSA areas are shown in Figure 1.5.6.6.1, together with existing bottom fisheries closures enforced by NEAFC and high seas MPAs established by OSPAR. Since ICES Advice 2013, Book 1 137 2005, NEAFC has closed 14 areas to bottom fishing to protect VMEs. In addition (for fisheries management rather than VME protection), NEAFC has closed an area for haddock on Rockall Bank since 2001 and enforced a seasonal closed area on the Reykjanes Ridge to protect a blue ling spawning ground. In total the area closed to protect VMEs is in excess of 600 000 square kilometres and covers all areas where the presence of VMEs have been validated up to the end of 2012. ICES has recently provided further advice on the boundaries of some of these VMEs (ICES, 2013a). NEAFC has different bottom-fishing regulations for existing fisheries areas and new fishing areas. Five of the proposed EBSAs are covered by the ‘New Fishing Area’ regulations only. Exploratory fishing in new areas is only authorized under strict conditions that include full observer coverage. There have been no applications for exploratory fishing since the regulations were adopted. Several points are notable: • • All the NEAFC closures lie within the proposed EBSAs. The majority of the overall area covered by the ten proposed EBSAs is classed by NEAFC as ‘new fishing areas’ and thus subject to strict regulation. Exceptions that are not ‘new fishing areas’ include an existing fishing area within the proposed EBSA 1 (Reykjanes Ridge), four within the proposed EBSA 3 (Mid-Atlantic Ridge north of Azores), and a further four within the proposed EBSA 4 (Hatton–Rockall area). The four major fisheries in the NEAFC Regulatory Area (herring, mackerel, blue whiting, and redfish) are pelagic trawl fisheries and therefore unlikely to pose threats to seabirds, taxa on the OSPAR list of threatened and declining species, or VMEs. In addition NEAFC regulations include: • • 138 A ban on the use of gillnet at depths greater than 200 m. A ban on directed fisheries for 17 different species of deep-sea sharks. ICES Advice 2013, Book 1 Figure 1.5.6.6.1 Map of the southern NEAFC Regulatory Area showing proposed EBSAs (Grey polygons; 1 = Reykjanes Ridge, 2 = Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone, 3 = Mid-Atlantic Ridge north of the Azores, 4 = Hatton–Rockall Plateau, 5 = Pedro Nunes and Hugo de Lacerda Seamounts, 6 = Northeast Azores–Biscay Rise, 7 = Evlanov Seamount, 8 = West of Azores). Existing NEAFC fishing areas are shown in white, NEAFC bottom fishery closures in pink, and OSPAR High Seas MPAs in green. ICES Advice 2013, Book 1 139 Table 1.5.6.6.1 Management measures adopted by NEAFC in each proposed EBSA. NEAFC closed areas or Rationale for NEAFC fishing regulations in the management measure proposed area Area 1. Reykjanes Ridge south of Iceland EEZ 1) Northern Mid-Atlantic 1,3,5) Protection of VME Ridge (MAR)1 (cold-water corals). 2) Seasonal blue ling 2) Protection for closure 2 spawning/aggregation 3)Existing fishing areas1 areas of blue ling. 4)New fishing areas1 NEAFC management regulations 1) Area closed to bottom trawling and fishing with static gear, including bottom-set gillnets and longlines 2009– 2015. 2) This area is closed for fishing blue ling from 15 February to 15 April until 2016. 3) Deep-sea demersal fisheries regulations: Certain gears are banned (gillnets) and actions against ghost fishing and lost gear are in place. 4) Authorization to go to new fishing areas follows a strict exploratory fishing protocol. Area 2. Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge Middle MAR1 Protection of VME (coldArea closed to bottom trawling and fishing with static water corals). gear, including bottom set gillnets and long-lines 20092015, including Charlie-Gibbs Fracture Zone and the Subpolar frontal zone. Area 3. Mid-Atlantic Ridge north of the Azores 1)Southern MAR; Altair Protection of VME (cold1) Area closed to bottom trawling and fishing with static Seamount, Antialtair water corals). gear, including bottom-set gillnets and longlines 2009– Seamount1 2015. 2)Existing fishing areas1 2) Deep-sea demersal fisheries regulations: Certain gears 3)New fishing areas1 are banned (gillnets) and actions against ghost fishing and lost gear are in place. 3) Authorization to go to new fishing areas follows a strict exploratory fishing protocol. Area 4. The Hatton and Rockall banks and Hatton–Rockall Basin 1) Rockall Haddock Box3 1) Haddock Box closed to 1) NEAFC has banned bottom trawling in this area. 2) Hatton4 protect juvenile haddock. 2–7) Area closed to bottom trawling and fishing with 2–7) Protection of VME 3) West Rockall Mounds static gear, including bottom-set gillnets and longlines (corals, coral reefs, and closure4 until 2015. 4) NW Rockall closure4 sponge grounds). 8) Deep-sea demersal fisheries regulations: Certain gears 5) SW Rockall closure4 7) Geomorphology. are banned (gillnets) and actions against ghost fishing 6) Logachev Mounds and lost gear are in place. closure 4 9) Authorization to go to new fishing areas follows a 7) Edora Bank closure5 strict exploratory fishing protocol. 8) Existing fishing areas1 9) New fishing areas1 140 ICES Advice 2013, Book 1 NEAFC closed areas or Rationale for NEAFC NEAFC management regulations fishing regulations in the management measure proposed area Area 5. Around Pedro Nunes and Hugo de Lacerda Seamounts – IBA MAO3 Area 6. Northeast Azores–Biscay Rise – IBA MAO3 Area 7. Evlanov Seamount Region Area 8. Northwest of Azores EEZ Area 9. The Arctic Front – Greenland/Norwegian Seas New fishing areas1 In most instances there is not enough research or Observers shall collect data in data to identify VMEs in “new fishing areas”. To accordance with a Vulnerable reduce risks to VMEs to a minimum these areas Marine Ecosystem Data are closed to normal commercial bottom fisheries Collection Protocol. under normal authorizations to fish from the Contracting Parties of NEAFC. Exploratory Bottom Fisheries Protocol. Vessels authorized under this protocol must have an observer on board. Exploratory fishing provides an opportunity to gather additional data on benthos and fish communities using industry vessels, and under strict controls on fishing. Area 10. The Arctic Ice habitat – multiyear ice, seasonal ice, and marginal ice zone This proposed area is outside of the NEAFC Regulatory Area. The following measures can be found at http://neafc.org/measures: 1 2 3 4 5 Rec. na 2011 on regulating bottom fishing as amended by Rec. 12 2013. Rec. 05 2013 Blue Ling seasonal closure. Rec. 03 2013 Rockall Haddock. Rec. 09 2013 Rockall Hatton VME closures. Rec. 08 2013 Edora Bank VME closure. International Commission for the Conservation of Atlantic Tuna (ICCAT) ICCAT has recommendations to mitigate the bycatch of seabirds, turtles, and pelagic shark species, but there are no specific spatial management measures relevant to the proposed EBSAs. OSPAR A range of measures have been adopted by the OSPAR Commission within the proposed EBSAs. These include legally binding decisions, which have been used for the establishment of seven marine protected areas (MPAs) in areas beyond national jurisdiction. Five of these lie within the proposed EBSAs. Two others lie outside the proposed EBSAs: the Milne Seamount complex MPA in the southwest of the OSPAR Area and the Josephine Seamount High Seas MPA in the southeast (Figure 1.5.6.6.1). For each MPA, OSPAR has also agreed recommendations for their management calling on Contracting Parties to undertake certain management actions within the competence of OSPAR (Table 1.5.6.6.2). In addition to these are three types of measures that are not spatially explicit and that apply across the OSPAR maritime area in the Northeast Atlantic (Table 1.5.6.6.3). These include a series of recommendations for the protection and conservation of threatened and/or declining species; a recommendation calling on Contracting Parties to specifically take into account OSPAR’s Listed of species and habitats in the conduct of Environmental Impact Assessments; as well as a code of conduct for undertaking marine research. ICES Advice 2013, Book 1 141 Table 1.5.6.6.2 Measures adopted by the OSPAR Commission specific to proposed EBSAs. Proposed area meeting the EBSA criteria Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the MidAtlantic Ridge Mid-Atlantic Ridge north of the Azores Table 1.5.6.6.3 • • • • • • • • • • • • Spatial management measures in place for specific sub-areas within the proposed EBSA (1.a) OSPAR Decision 2010/2 on the establishment of the Charlie-Gibbs South MPA (1.b) OSPAR Recommendation 2010/13 on the management of the Charlie-Gibbs South MPA (2.a) OSPAR Decision 2012/1 on the establishment of the Charlie-Gibbs North High Seas MPA (2.b) OSPAR Recommendation 2012/1 on the management of the Charlie-Gibbs North High Seas MPA (1.a) OSPAR Decision 2010/6 on the establishment of the Mid-Atlantic Ridge north of the Azores High Seas MPA (1.b) OSPAR Recommendation 2010/17 on the management of the Mid-Atlantic Ridge north of the Azores High Seas MPA (2.a) OSPAR Decision 2010/3 on the establishment of the Altair Seamount High Seas MPA (2.b) OSPAR Recommendation 2010/14 on the management of the Altair Seamount High Seas MPA (3.a) OSPAR Decision 2010/4 on the establishment of the Antialtair Seamount High Seas MPA (3.b) OSPAR Recommendation 2010/15 on the management of the Antialtair Seamount High Seas MPA General management measures taken by the OSPAR Commission that are common to all proposed EBSAs. OSPAR Code of Conduct for Responsible Marine Research in the Deep Seas and High Seas of the OSPAR Maritime Area (Reference number: 2008-1) OSPAR Recommendation 2010/5 on assessments of environmental impact in relation to threatened and/or declining species and habitats OSPAR Recommendation 2010/6 on furthering the protection and restoration of the common skate species complex, the white skate, the angel shark, and the basking shark in the OSPAR Maritime Area OSPAR Recommendation 2010/7 on furthering the protection and restoration of the Orange roughy (Hoplostethus atlanticus) in the OSPAR Maritime Area OSPAR Recommendation 2010/8 on furthering the protection and restoration of Lophelia pertusa reefs in the OSPAR Maritime Area OSPAR Recommendation 2010/9 on furthering the protection and restoration of coral gardens in the OSPAR Maritime Area OSPAR Recommendation 2011/2 on furthering the protection and conservation of the Ivory gull (Pagophila eburnea) OSPAR Recommendation 2011/3 on furthering the protection and conservation of the Little shearwater (Puffinus assimilis baroli) OSPAR Recommendation 2011/4 on furthering the protection and conservation of the Balearic shearwater (Puffinus mauretanicus) OSPAR Recommendation 2011/5 on furthering the protection and conservation of the Black-legged kittiwake (Rissa tridactyla tridactyla) OSPAR Recommendation 2011/6 on furthering the protection and conservation of the Roseate tern (Sterna dougallii) OSPAR Recommendation 2011/7 on furthering the protection and conservation of the Thick-billed murre (Uria lomvia) International Maritime Organization (IMO) and the London Convention No spatially based measures have been introduced by IMO or the London Convention for Areas Beyond National Jurisdiction in the Northeast Atlantic. Many general IMO measures (e.g. ballast water exchange protocols, disposal of waste) will apply to these areas. 142 ICES Advice 2013, Book 1 International Seabed Authority (ISA) According to UNCLOS ISA is the competent authority that regulates mining and mineral extraction in “The Area” (http://www.isa.org.jm/en/scientific/exploration). A memorandum of understanding was signed between the OSPAR Convention and the International Seabed Authority in June 2011 to facilitate consultation and sharing of data and information of relevance, with a view to promoting and enhancing a better understanding and coordination of their respective activities. There are three types of deep-sea mineral resources that are of commercial interest: polymetallic nodules, polymetallic sulphides, and cobalt-rich ferromanganese crusts. So far, ISA has only adopted the regulatory framework for the exploration of deep-sea mineral resources, including very strict regulations to ensure full environmental impact assessments are undertaken prior to any activity in “The Area”. The framework for the exploitation of these resources has yet to be developed and adopted. There are several areas in the Northeast Atlantic identified as having deposits of mineral resources of possible commercial interest, but exploratory mining cannot commence without adoption of a management framework. Arctic Council In follow-up to a recommendation to identify areas of heightened ecological and cultural significance, a report is being prepared for the Arctic Council’s Protection of the Arctic Marine Environment Working Group (PAME). If the areas are agreed, it is recommended that Arctic states, where appropriate, should encourage implementation of measures to protect these areas from the impacts of Arctic marine shipping, in coordination with all stakeholders and consistent with international law. National and European Union At the national and EU levels, many relevant measures for the protection of areas have not been systematically reviewed by ICES. Several of these measures implement more general international obligations, for instance under the Convention on Trade in Endangered Species. Sources ICES. 2013a. Vulnerable deep-water habitats in the NEAFC Regulatory Area. In Report of the ICES Advisory Committee, 2013, Section 1.5.5.1. ICES Advice, 2013, Book 1. ICES. 2013b. Report of the Workshop to Review and Advise on EBSA Proposed Areas (WKEBSA), 27–31 May 2013, Copenhagen, Denmark. In draft. ICES Advice 2013, Book 1 143 1.5.5.7 Special Request, Advice September 2013 ECOREGION General advice SUBJECT OSPAR/NEAFC special request on review and reformulation of four EBSA Proformas Advice summary ICES provided advice to OSPAR and NEAFC in June 2013 (OSPAR/NEAFC special request on review of the results of the Joint OSPAR/NEAFC/CBD Workshop on Ecologically and Biologically Significant Areas (EBSAs) (ICES Advice 2013 section 1.5.5.5). Following discussion with OSPAR and NEAFC, ICES (using experts of the review group) agreed to reformulate and revise four of the EBSAs and provide new updated maps. The material consists of scientifically updated Proformas for the following EBSAs: • • • • Mid-Atlantic Ridge North of the Azores and South of Iceland Charlie-Gibbs Fracture Zone (and the Sub-Polar Front) The Hatton and Rockall Banks and the Hatton-Rockall Basin The Arctic Ice habitat – multiyear ice, seasonal ice – marginal ice zone During the update on the Charlie-Gibbs Fracture Zone (CGFZ) it appeared that the Sub-Polar Front, which was included in the previous version of the Proforma, could not be scientifically supported with the evidence to hand at the time and was therefore excluded from the CGFZ Proforma. A short note explains the reasons and consequences for the exclusion. • Note on the Sub Polar Front. The five documents are scientifically updated technical documents and appended to this advice. 144 ICES Advice 2013, Book 1 ANNEX 1 Draft Proforma: The Hatton and Rockall Banks and the Hatton-Rockall Basin Presented by: The Joint OSPAR/NEAFC/CBD Scientific Workshop. Reviewed by an ICES expert group and revised by Francis Neat and J. Murray Roberts. Based on an original proposal submitted by: Dr David Billett and Dr Brian Bett (Deepseas Group, Ocean Biogeochemistry and Ecosystems Department, NOC, UK), Prof. Philip Weaver (Seascape Consultants Ltd and National Oceanography Centre, UK), Prof Callum Roberts and Ms Rachel Brown (Environment Department, University of York), Dr Murray Roberts and Dr Lea-Anne Henry, (Centre for Marine Biodiversity and Biotechnology, School of Life Sciences, Heriot-Watt University), Dr Kerry Howell and Dr Jason Hall-Spencer (Marine Biology and Ecology Research Centre, Marine Institute, University of Plymouth); Dr Andrew Davies (School of Ocean Sciences, Bangor University); Dr Bhavani Narayanaswamy (Scottish Association for Marine Science, Oban), Prof. Monty Priede (OceanLab, University of Aberdeen); Dr David Bailey (Division of Environmental and Evolutionary Biology, University of Glasgow); Prof. Alex Rogers (University of Oxford) and Mr Ben Lascelles (Global Seabird Programme, Bird Life International) Abstract The Hatton and Rockall Banks, associated slopes and connecting basin, represent offshore bathyal habitats between 200 to 1500 m that constitute a unique and prominent feature of the NE Atlantic. The area has high habitat heterogeneity and supports a wide range of benthic and pelagic species and ecosystems. There is significant fishing activity in the area, including bottom trawling, long-lining, and midwater fisheries. Introduction The Hatton and Rockall Banks are large isolated geomorphological features in the NE Atlantic. Formed from continental crust, they span depths from c. 200 to 2000m. The banks are linked by the Hatton-Rockall Basin at a depth of approximately 1300 m which has particular geomorphological features and habitats. The gently sloping banks and the basin provide a contrasting geological setting to the tectonically active Mid-Atlantic Ridge to the west and the generally steeper slopes of the European continental margin to East. The banks encompass a large depth range with strong environmental gradients (e.g. temperature, pressure, and food availability) that give rise to a high diversity of species and habitats (Billett, 1991; Bett, 2001; Howell et al., 2002; Davies et al. 2006; Roberts et al. 2008; Howell et al., 2009; Howell et al. 2010). Environmental heterogeneity is positively correlated with biological diversity at a variety of scales (Menot et al. 2010) as indicated by significantly elevated levels of species change across space (in areas such as Hatton Bank (Roberts et al. 2008). Changes in pressure and temperature have significant effects on the biochemistry of species, influencing cell membrane structure and enzyme characteristics (Gage and Tyler, 1991). In general, each species is adapted to a particular range of environmental conditions. Each may occur over a depth range of about 500 m, but the depths where any particular species is abundant, and therefore able to form viable populations, is generally limited to a much more restricted depth range of 100 to 200 m (Billett, 1991; Howell et al., 2002). There is evidence that such depth-related effects promote speciation (Howell et al., 2004). In addition, the progressive decrease in organic matter availability with increasing depth (with some patchiness depending on geomorphology) leads to a reduction of carnivores and an increase in detritus feeders (Billett, 1991). Taken together such environmental changes lead to a continuous sequential change in species composition with depth, and biological community characteristics that are radically different to those known in shelf seas. The area is influenced by a number of different water masses and there is considerable interaction between the topography and physical oceanographic processes, in some areas focusing internal wave and tidal energy (Ellett et al. 1986) which results in strong currents and greater mixing. This may give rise to highly localized and specialised biological communities such as sponge aggregations and coral gardens. The mixing of Arctic and Atlantic water in the North of the area means that species from both ecosystems are represented causing enhanced species diversity. The Rockall Bank supports shallow demersal fisheries targeting haddock, gurnard and monkfish (Neat & Campbell 2010). The slopes and the Hatton Bank are target areas for deep-water bottom fisheries for Ling (Molva molva), Blue Ling (Molva dypterygia), Tusk (Brosme brosme), Roundnose Grenadier (Coryphaenoides rupestris) and Black Scabbardfish (Aphanopus carbo). In the past deepwater sharks were also caught in the area, but this is now prohibited. A wide variety of other fish species are also taken as by-catch (Gordon et al., 2003; Large et al., 2003; ICES 2010). Some of the deepwater target species have characteristic low productivity and extended generation times. Deep-water fisheries have ICES Advice 2013, Book 1 145 significant effects not only on target fish species, but also on the benthic fauna (Le Guilloux et al., 2009; Clark et al. 2010). Major wide-ranging Northeast Atlantic epipelagic fish stocks, e.g. mackerel and blue whiting, use the Hatton-Rockall area for parts of their life cycle and are targeted by international fisheries. The slopes of the banks and channels between the banks have a diverse bathy- and mesopelagic fish community sustained by the zooplankton production in the epipelagic zone. The pelagic communities are similar to, and probably extensions of, those in adjacent oceanic waters along the European continental margin. Some invertebrate species, such as cold-water corals and sponges, provide important structural habitat heterogeneity. These habitats are highly susceptible to physical damage and may take hundreds, if not thousands, of years to reform (Hall-Spencer et al. 2002; Roberts et al. 2009; Söffker et al., 2011). Current fisheries control measures on Hatton and Rockall Bank have focused mainly on the protection of corals (HallSpencer et al., 2009) and sponges (ICES 2013). There is no evidence currently that the seabed at depth greater than 1500 m in the area is significantly different from comparable depths in the rest of the NE Atlantic. The majority of the features considered here occur at depths shallower than 1500 m and this therefore forms an appropriate delimitation of the EBSA. Location The EBSA would comprise the seabed and pelagic zones shallower than 1500 m above the Rockall and Hatton Banks and the adjoining Hatton-Rockall Basin. This extends into adjacent EEZs, but the current proposal refers to the ABNJ only. The area beyond national jurisdiction lies wholly within regions under consideration by the Commission on the Limits of the Continental Shelf. Feature description Benthic and pelagic communities to depths of 1500 m in and around the Hatton and Rockall Banks and Basin. Seabed communities include cold-water coral formations and sponge aggregations. Geomorphologically complex seabed types include rocky reefs, carbonate mounds, polygonal fault systems and sedimentary slopes, slides and fans. Pelagic communities include those inhabiting bathy-, meso- and epi-pelagic zones, including zooplankton, fish, cetaceans, turtles and seabirds. 1. Benthic and benthopelagic communities Cold-water coral Observations in the early 1970s found cold-water coral communities on the Rockall Bank down to a depth of 1,000 m (Wilson, 1979a). Thickets of Lophelia pertusa occurred principally at depths between 150-400m 10. Large coral growth features have recently (2011) been discoveredto be still present on the northern Rockall Bank (Huvenne et al., 2011, Roberts et al. 2013). Bottom-contact fishing can result in significant adverse impacts to these habitats. Frederiksen et al. (1992) reported a high diversity of corals on the northern Hatton Bank, including Paragorgia, Paramuricea, Isididae and Antipatharia as well as the scleractinians L. pertusa and M. oculata. Since these observations further records of coral frameworks have been noted throughout the Rockall, Hatton area, including the Logachev Mounds and the Western Rockall Bank Mounds (Kenyon et al., 2003; Roberts et al., 2003; Narayanaswamy et al., 2006; Howell et al., 2007; Durán Muñoz et al. 2009). Recent surveys identified many areas that contained the cold-water coral L. pertusa throughout the Rockall and Hatton Banks (Narayanaswamy et al., 2006; Howell et al., 2007; Roberts et al. 2008; Durán Muñoz et al. 2009). Several areas on the Hatton Bank contained pinnacles and mounds with extensive biogenic structures including areas of coral rubble around the flanks of the coral mounds. Coral frameworks are known from the Hatton Bank (Durán Muñoz et al. 2009), and are predicted to occur over focused regions of the Hatton Bank (Howell et al., 2011). Geophysical evidence suggests that these have formed by successive coral growth and sedimentation episodes, as in other regions (Roberts et al., 2006), forming coral carbonate mounds (Roberts et al. 2008). Single and clustered coral carbonate mounds have also been discovered on the southeast of Rockall Bank. These structures are comprised mostly of L. pertusa and can reach heights of 380 m in water depths of between 600-1000 m (Kenyon et al., 2003; Mienis et al., 2006; Mienis et al., 2007). 10 146 http://www.lophelia.org/lophelia/case_4.htm ICES Advice 2013, Book 1 Cold-water coral frameworks have been reported to support over 1,300 species in the Northeast Atlantic, some of which have yet to be described (Roberts et al., 2006). New species and associations have been reported recently (e.g. Myers & Hall-Spencer 2007; Le Guilloux et al., 2010; Söffker et al. 2011). The corals may provide an important habitat for certain fish species (Fosså et al., 2002; Söffker et al. 2011; Henry et al 2013), including commercial species Sebastes sp., Molva molva, Brosme brosme, Anarhichas lupus and Pollachius virens (Mortensen et al., 1995; Freiwald, 2002; Hall-Spencer et al., 2002). Pregnant Sebastes viviparus may use the reef as a refuge or as a nursery ground to raise their offspring (Fosså et al., 2002) as recently observed on the northern Rockall Bank (Huvenne et al., 2011, Roberts et al. 2013). As well as living reefs, dead coral framework and coral rubble provide a structural habitat. Jensen and Frederiksen (1992) collected Lophelia and found 256 species; a further 42 species were identified amongst coral rubble. There has been only limited research into linkages between coral and other deep-water ecosystems. Compared to the south-eastern US and Gulf of Mexico, molecular research has shown that northeastern Atlantic populations of L. pertusa are moderately differentiated (Morrison et al. 2011) and form distinct subpopulations, but also that Rockall Bank corals show some genetic similarity to those occurring on the New England Seamounts indicating some degree of connectivity (Morrison et al. 2011). Lophelia pertusa exhibits high levels of inbreeding through asexual reproduction at several sites in the NE Atlantic, suggesting a high incidence of self-recruitment in local populations (Le Goff-Vitry and Rogers, 2005). Further molecular studies are required in local areas to gauge the importance of the Rockall and Hatton Banks in the life history of regional coral populations. In summary the cold-water corals fit the following EBSA criteria: Uniqueness or rarity • Large areas of cold-water corals and sponges have been reported in the area. Some of these have been destroyed by demersal trawling, but in certain areas, e.g. SW Rockall Bank, extensive patches of coral framework still exist. Special importance for life-history stages • Cold-water corals and areas of natural coral rubble provide highly diverse habitats Importance for threatened, endangered or declining species/habitats • • The cold-water corals and natural rubble contain very large numbers of invertebrate species including giant protozoans on nearby sedimentary habitats (xenophyophores), vase shaped white sponges, actiniarians, antipatharian corals, hydroids, bryozoans, asteroids, ophiuroids, echinoids, holothurians and crustaceans. The distribution of cold-water coral has been severely reduced in the area over the last 30 years Vulnerability, fragility, sensitivity, or slow recovery • • There is a high diversity of corals, including bamboo coral (Isididae), black coral (Antipatharia) as well as the reef forming stony corals (Scleractinia), though some of these may now be reduced in distribution occurring in patches. Cold-water coral habitats are easily impacted and recover very slowly, if at all. Biological diversity • Cold-water corals provide diverse habitats for other invertebrates and fish. Sediment communities The Hatton and Rockall Banks support many different habitats each with their own depth-related species assemblages (Narayanaswamy et al., 2006; Howell et al., 2007; Roberts et al. 2008; Howell et al., 2009). Local seabed morphology in this region is ultimately controlled by hydrography and oceanography (Due et al. 2006; Sayago-Gil et al. 2010), which creates heterogeneity in sediment types including mud, exposed bedrock, fine sediments, living coral framework and coral debris that – this habitat heterogeneity has a major influence on species diversity and turnover (Roberts et al. 2008). A great variety of large invertebrate fauna (megafauna) occur in this region including giant protozoans (xenophyophores), vase shaped white sponges, actiniarians, antipatharian corals, hydroids, bryozoans, asteroids, ophiuroids, echinoids, holothurians and crustaceans (Narayanaswamy et al., 2006; Howell et al., 2007; Roberts et al. 2008). Large mega-infauna such as echiuran worms are evident from observations of their feeding traces. Little is known, however, of the smaller fauna living within the sediment. The Hatton-Rockall Basin is known to host a particular geomorphology known as a polygonal fault system (Berndt et al 2012). The faults in the Hatton-Rockall Basin have surface expression, i.e. a network of interlinked channels across the level seafloor. These fault structures were confirmed again in 2011 (Huvenne et al., 2011). The flanks of the gullies appear to support extensive, dense aggregations of mixed species sponge communities. ICES Advice 2013, Book 1 147 A key interest / conservation concern in such a geological setting would be the occurrence of cold-seep communities. Large carbonate blocks were encountered that were likely formed as a result of seafloor fluid escape. In 2012 the first evidence of an active cold-seep ecosystem in the area was suggested by the collection of chemosynthetic bivalves and polychaete worms (ICES 2013) on the eastern margin of Hatton-Rockall Basin at a depth of 1200 m. The species are new to science and suggest there is a lot still to learn of the seafloor and ecology of the Hatton and Rockall Banks. The megafauna on the Hatton and Rockall Banks are largely species known from the wider NE Atlantic continental margin (Gage et al. 1983; Gage et al., 1985; Mauchline et al., 1986; Harvey et al., 1988; Rice et al., 1991). These studies focused on sedimented areas within the EEZs of the UK and Ireland and provide a lot of information on the life history characteristics of the species including information on growth and reproduction. Apart from some species that produce small eggs (indicative of planktotrophic development) in a seasonal cycle, most species conform to the life history characteristics typical of the deep sea of larger egg size, lower fecundity and greater generation times (Gage and Tyler, 1991). This is an adaptation to the low food input to the deep sea, which leads to the rapid decrease in biomass with increasing depth (Lampitt et al., 1986; Wei et al., 2010). Fauna adapt to lower food availability in the deep sea by a number of trade-offs, one of which is a reduction in reproductive effort and longer generation times. The majority of species, therefore, are highly susceptible to repeated physical disturbance. In summary the sediment communities fit the following EBSA criteria: Uniqueness or rarity • • The area has considerable environmental heterogeneity, and therefore biological diversity, as a result of its large depth range and strong environmental gradients. Habitat-forming sessile benthic communities, such as those of giant protozoans and sponges, are common. The area of polygonal faults may be a unique seabed feature and the presence of newly described chemosynthetic bivalves and polychaete worms suggests the area may have unique communities. Importance for threatened, endangered or declining species/habitats • The area comprises a patchwork of habitats with species changing consistently with both habitat type and increasing to depths of 1500 m. Some habitats are threatened by direct impacts (e.g. trawling). Vulnerability, fragility, sensitivity, or slow recovery • Many of the species have reproductive cycles with long generation times leading to very slow and episodic recoveries following human impact. Most deep-sea species are particularly susceptible to degradation and depletion by human activity. Biological productivity • There are localised areas of concentrated production depending on geomorphology and hydrography, but little evidence that the area has an enhanced productivity relative to other areas. Biological diversity • Benthic and pelagic communities occupy all depths in and around the Hatton and Rockall Banks and Basin. Seabed communities include cold-water corals and sponge aggregations. Seabed geomorphology is diverse with examples of rocky reefs, carbonate mounds, polygonal fault systems, and steep and gentle sedimentary slopes. This high habitat heterogeneity supports a high number of species and diverse communities. Demersal fish The deep-water fish of the NE Atlantic continental margin are generally well-known following comprehensive and extensive surveys of the region (e.g. Gordon & Duncan, 1985; Merrett et al., 1991; Mauchline et al. 1986 and Rice et al. 1991). Species of commercial importance are reviewed by Gordon et al. (2003) and Large et al. (2003) and for fish associated with cold-water corals by Söffker et al. (2011). Fish species diversity increases to depths of approx. 1500 m on the continental slopes and declines thereafter (Campbell et al 2011). The shallow water fish assemblage on Rockall can be described as an impoverished sub-set of that found in adjacent continental shelf areas, but one that has a significantly different community composition (Neat & Campbell 2010). Recent surveys have found that the western slope of the Rockall Bank has a slightly different fish assemblage than the adjacent European slope with several species of a more southern affinity present (F. Neat unpublished data). Blue ling is known to spawn in a few locations on Rockall bank and at Hatton bank (Large et al 2008). 148 ICES Advice 2013, Book 1 The detailed sampling in the Porcupine Seabight in the 1970s and 1980s took place before the start of deep-water commercial fishing. More recent sampling of the same area in the 1990s and 2000s can be used to compare fish communities before and after bottom trawling (Bailey et al. 2009). These data show that over 70 fish species have been impacted by the fishing activity, of which only 4-5 are target commercial species. The area impacted is up to 2.5 times larger than the area fished because the home range of many the fish extends into considerably deeper waters. In the past decade, however, there is evidence that this initial steep decline in abundance has been halted, at least in one of the major groups of fishes, the grenadiers (Neat & Burns 2010). At the northern limits of the area where Arctic water masses mix with Atlantic water cold-water species such as Greenland Halibut and Roughhead Grenadier are present adding to the diversity of species in the area. In summary the demersal fish fit the following EBSA criteria: Vulnerability, fragility, sensitivity, or slow recovery • Many of the deep demersal fish have very slow recovery times as a result of their slow reproductive rate compared to pelagic fish. 2. Pelagic communities (plankton, nekton, birds) Fish: Mackerel, blue whiting and other wide-ranging pelagic fish such as epipelagic sharks and tuna-like species use the area during parts of their life-cycle, for feeding or as migration corridors. For blue whiting the slope area is used as a spawning area. Mackerel eggs and larvae from spawning areas further south drift through the area. Cetaceans: Phocoena phocoena have been observed over the shallower parts of Rockall Bank, but It is unlikely that the area is of particular importance for the species. Limited numbers of the endangered Blue whale (Balaenoptera musculus) and the critically endangered northern right whale (Eubalaena glacialis) have also been observed in this area (Cronin and Mackey, 2002; Hammond et al., 2006). Seabirds: Analyses of satellite tracking data hosted at www.seabirdtracking.org (Table 1) found the area to be used by multiple species through the year. The site is used by Manx Shearwaters (Puffinus puffinus) during the breeding season (Apr-Sept) from Iceland and UK colonies. From September until November tracked individuals of Cory’s Shearwater (Calonectris diomedea) from 3 colonies, Sooty Shearwater (Puffinus griseus), Fea’s Petrel (Pterodroma feae) and Zino’s Petrel (Pterodroma madeira) used the area. Studies of tracked Atlantic Puffin (Fratercula arctica) from Skomer and Isle of May colonies found the site to be important during the overwintering phase (Aug-Apr) (Harris et al. 2010, Guilford et al. 2011). In addition to tracking data, at-sea survey data confirms many more species within the area (e.g. Cronin and Mackey, 2002). Feature condition, and future outlook Demersal fish have been targets of extensive fisheries for decades, expanding primarily in the latter half of the 1980s. Although satisfactory stock assessments were seldom achieved, the probable declines in abundance and vulnerability of many of the target species have been reflected in advice from ICES for many years (ICES 1996 onwards, Large et al., 2003). A range of management actions by NEAFC and relevant coastal states have been implemented to reduce fishing effort and facilitate recovery of target species and some associated by-catch species. A similar range of measures applies to species inhabiting the shallowest areas, e.g. haddock. Epipelagic species such as mackerel and blue whiting, and large pelagic sharks and tuna-like species occurring in the area straddle between ABNJ and several EEZs and the fisheries are managed by relevant coastal states, NEAFC and ICCAT. Cetaceans are managed by the IWC. The management is based on recurrent stock assessments by ICES and other advisory bodies. Records of the physical impact of deep-water trawling west of Scotland extend back to the late 1980s (Roberts et al., 2000; Gage et al., 2005) and studies using VMS data show that fishing activity potentially affects much of the HattonRockall area (Hall-Spencer et al. 2009; Benn et al. 2010). Damage may occur to structural species such as corals and sponges, which may take hundreds to thousands of years to recover (Hall-Spencer et al., 2002; Davies et al. 2007; Roberts et al., 2009; Hogg et al. 2010). A recent survey (2011) has documented extensive destruction of coral framework on the northern Rockall Bank (Huvenne et al. 2011) in waters adjacent to the area currently being described. This expedition also encountered evidence of trawling impact on the megafauna of open sedimented areas, with photographic surveys in the area of the 'Haddock Box' (Rockall Bank) showing frequent occurrence of physically damaged holothurians - thought to be net escapees or discarded byICES Advice 2013, Book 1 149 catch. Cold seep communities are vulnerable to trawling impacts; they are typically highly localised and are of a relatively small scale such that they could be eliminated by a single trawl. Cold seeps are OSPAR priority habitats for which there are considerable concerns regarding the effects of bottom trawling (van Dover et al. 2011a, b). Some of the benthic communities of the Hatton and Rockall Banks have already been significantly affected by deep-water fishing (ICES WGDEC, 2007). The effects on deep-water fish may extend to waters deeper than those utilised by trawl fisheries (Bailey et al., 2009). Broad-scale multibeam surveys have revealed a diverse range of geomophological features and sediment types on Hatton Bank (Jacobs and Howell, 2007; Stewart and Davies, 2007; MacLachlan et al., 2008; Sayago-Gil et al., 2010). These physical environment maps, coupled with targeted biological surveys have resulted in the production of biological habitat maps for the region (Howell et al., 2011) which highlight the range and diversity of noncoral seabed features present in the area. Assessment against CBD EBSA Criteria Table 1. Relation of each of the CBD criteria to the proposed area relating to the best available science. Note that a candidate EBSA may qualify on the basis of one or more of the criteria, the boundaries of the EBSA need not be defined with exact precision. CBD EBSA Criterion Description The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Explanation for ranking Uniqueness or rarity Ranking of criterion relevance (please mark one column with an X) Don’t Low Some Know High X • The area has considerable environmental heterogeneity, and therefore biological diversity, as a result of its large depth range and strong environmental gradients. Habitat-forming sessile benthic communities, such as those of giant protozoans and sponges, are common. • Large areas of cold-water corals and sponges have been reported in the area. Some of these have been destroyed by demersal trawling, but some areas of large coral frameworks still exist. • An area of polygonal faults may be a unique seabed feature and the recent discovery of cold-seep species that are new to science suggests the area is very likely to be unique. Areas that are required for a population to survive and Special X thrive importance for life-history stages of species Explanation for ranking • • Cold-water corals and areas of natural coral rubble provide highly diverse habitats Parts of the Hatton-Rockall area are important as spawning areas for blue whiting, and the area is used as a corridor for a range of migrating species including turtles. Area containing habitat for the survival and recovery of Importance for X endangered, threatened, declining species or area with threatened, significant assemblages of such species endangered or declining species and/or habitats Explanation for ranking • • • 150 The cold-water corals and natural rubble contain very large numbers of invertebrate species including giant protozoans (xenophyophores), vase shaped white sponges, actiniarians, antipatharian corals, hydroids, bryozoans, asteroids, ophiuroids, echinoids, holothurians and crustaceans. The distribution of cold-water coral has been severely reduced in the area over the last 30 years The area comprises a patchwork of habitats with species changing consistently with both habitat type and increasing depth. Some habitats are threatened by direct impacts (e.g. trawling), others may suffer indirectly e.g. through the creation of sediment plumes by impacts of fishing gear in sensitive areas. ICES Advice 2013, Book 1 Areas that contain a relatively high proportion of sensitive Vulnerability, habitats, biotopes or species that are functionally fragile fragility, (highly susceptible to degradation or depletion by human sensitivity, or activity or by natural events) or with slow recovery slow recovery Explanation for ranking • • • • X There is a high diversity of corals, including bamboo coral (Isididae), black coral (Antipatharia) as well as the reef forming stony corals (Scleractinia), though some of these may now be reduced in distribution occurring in patches. Cold-water coral habitats are easily impacted and recover very slowly, if at all Many of the species have reproductive cycles with long generation times leading to very slow and episodic recoveries following human impact. Most deep-sea species are particularly susceptible to degradation and depletion by human activity and natural events. Many of the demersal fish have very slow recovery times as a result of their slow reproductive rate compared to pelagic fish. Stocks have already been diminished in some areas. Area containing species, populations or communities with Biological comparatively higher natural biological productivity productivity Explanation for ranking X • While pelagic organisms may be more concentrated over the banks in the area, there is little evidence to suggest overall enhanced productivity of the area. Area contains comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or has diversity higher genetic diversity Explanation for ranking • • • Benthic and pelagic communities occupy all depths in and around the Hatton and Rockall Banks and Basin. Seabed communities include cold-water corals, rocky reefs, carbonate mounds, polygonal fault systems, sponge aggregations, steep and gentle sedimented slopes. The Hatton and Rockall Banks and the Hatton-Rockall Basin have a high habitat heterogeneity that supports diverse seabed communities. Cold-water corals provide diverse habitats for other invertebrates and fish. References Bailey, D.M., Collins, M.A., Gordon, J.D.M., Zuur, A.F. & Priede, I.G. (2009) Long-term changes in deep-water fish populations in the northeast Atlantic: a deeper reaching effect of fisheries? Proc. Roy. Soc. Lond. B. 276, 19651969 Benn, A.R., Weaver, P.P.E, Billett, D.S.M., van den Hove, S., Murdock, A.P., Doneghan, G.B. & Le Bas, T. (2010). Human Activities on the Deep Seafloor in the North East Atlantic: An Assessment of Spatial Extent. PLoS One 5(9): e12730. doi:10.1371/journal.pone.0012730. Berndt, C, Jacobs, C. L., Evans, A. J., Gay, A., Elliot, G., Long, D. and Hitchen, K. (2012) Kilometre-scale polygonal seabed depressions in the Hatton Basin, NE Atlantic Ocean: Constraints on the origin of polygonal faulting Marine Geology, 332/334 . pp. 126-133. DOI 10.1016/j.margeo.2012.09.013. Bett, B.J. (2001) UK Atlantic Margin Environmental Survey: introduction and overview of bathyal benthic ecology. Cont. Shelf Res. 21, 917-956. Billett, D.S.M. (1991) Deep-sea holothurians. Oceanogr. mar. Biol. Ann. Rev. 29, 259-317. Campbell, N. et al. 2010. Taxonomic indicators of deep water demersal fish community diversity on the Northeast Atlantic continental slope. ICES J. Mar. Sci. 68, 365-378. Clark M.R., Rowden A.A., Schlacher T., Williams A., Consalvey M., Stocks K.I., Rogers A.D., O'Hara T.D., White M., Shank T.M. & Hall-Spencer J.M. (2010) The ecology of seamounts: structure, function and human impacts. Annu. Rev. Mar. Sci. 2, 253-278. Costello, M.J., McCrea, M., Freiwald, A., Lundälv, T., Jonsson, L., Bett, B.J., Van Weering, T.C.E., De Haas, H., Roberts, J.M. & Allen, D. (2005) Role of cold-water Lophelia pertusa reefs as fish habitat in the NE Atlantic. In: Freiwald, A., Roberts, J.M. (Eds.), Cold-water Corals and Ecosystems. Springer-Verlag, Berlin Heidelberg, pp. 771-805. Cronin, M., Mackey, M. (2002) Cetaceans and Seabirds of the Hatton-Rockall Region, Cruise Report of the Geological Survey of Ireland May 2002. Davies, A.J., Narayanaswamy, B.E., Hughes, D.J. & Roberts, J.M. (2006) An introduction of the benthic ecology of the Rockall-Hatton Area (SEA 7). Scottish Association for Marine Science, Oban, p. 94. http://www.offshoresea.org.uk/ Davies, A.J., Wisshak, M., Orr, J.C. & Roberts, J.M. (2008) Predicting suitable habitat for the cold-water coral Lophelia pertusa (Scleractinia). Deep-Sea Res. I 55, 1048-1062. Davies. A., Roberts. J.M. & Hall-Spencer, J.M. (2007) Preserving deep-sea natural heritage: emerging issues in offshore conservation and management. Biol. Cons. 138, 299-312. ICES Advice 2013, Book 1 151 Due, L., van Aken. H.M., Boldreel. L.O. & Kuijpers, A. (2006) Seismic and oceanographic evidence of present-day bottom-water dynamics in the Lousy Bank–Hatton Bank area, NE Atlantic. Deep-Sea Research I53:1729-1741 Durán Muñoz, P., Sayago-Gil, M., Cristobo, J., Parra, S., Serrano, A., Díaz del Rio, V., Patrocinio, T., Sacau, M., Murillo, F.J., Palomino, D. & Fernández-Salas, L.M. (2009) Seabed mapping for selecting cold-water coral protection areas on Hatton Bank, Northeast Atlantic. ICES J. Mar. Sci. 66, 2013-2025 Ellett, D.J., Edwards, A. & Bowers, R. (1986) The hydrography of the Rockall Channel – an overview. Conference Proceedings Symposium on the Oceanography of the Rockall Channel, Edinburgh (UK), 27-29 Mar 1985. Fosså, J.H., Mortensen, P.B. & Furevik, D.M. (2002) The deep-water coral Lophelia pertusa in Norwegian waters: distribution and fishery impacts. Hydrobiologia 471, 1-12. Frederiksen, R., Jensen, A. & Westerberg, H. (1992) The distribution of the scleractinian coral Lophelia pertusa around the Faeroe Islands and the relation to internal tidal mixing. Sarsia 77, 157-171. Freiwald, A. (2002) Reef-forming cold-water corals. In: Wefer, G., Billett, D., Hebbeln, D., Jorgensen, B.B., Schluter, M., Van Weering, T. (Eds.), Ocean Margin Systems. Springer-Verlag Berlin Heidelberg, Berlin, pp. 365-385. Gage, J.D. & Tyler, P.A. (1991) Deep-Sea Biology. A Natural History of Organisms at the Deep-Sea Floor. Cambridge University Press. 504pp. Gage, J.D., Billett, D.S.M., Jensen, M. & Tyler, P.A. (1985) Echinoderms of the Rockall Trough and adjacent areas. 2. Echinoidea and Holothurioidea. Bull. Br. Mus. Nat. Hist. (Zool.), 48, 173-213. Gage, J.D., Pearson, M., Clark, A.M., Paterson, G.L.J. & Tyler, P.A. (1983) Echinoderms of the Rockall Trough and adjacent areas. 1. Crinoidea, Asteroidea and Ophiuroidea. Bull. Br. Mus. nat. Hist. (Zool.) 45, 263-308. Gage, J.D., Roberts, J.M., Hartley, J.P. & Humphery, J.D. (2005) Potential impacts of deepsea trawling on the benthic ecosystem along the Northern European Continental Margin: A review. In: Barnes, P.W., Thomas, J.P. (Eds.), Benthic Habitats and the Effects of Fishing. American Fisheries Society, Bethesda, Maryland, pp. 503-517. Gordon, J. D. M., O. A. Bergstad, I. Figueiredo, and G. Menezes. 2003. Deep-water Fisheries of the Northeast Atlantic: I. Description and Current Trends. J. Northw. Atl. Fish. Sci. 31: 137-150. Gordon, J.D.M. & Duncan, J.A.R. (1985) The ecology of the deep-sea benthic and benthopelagic fish on the slopes of the Rockall Trough, northeastern Atlantic. Prog. Oceanogr. 15, 37-69. Guilford, T., Freeman, R., Boyle, D., Dean, B., Kirk, H., Phillips, R.A., Perrins, C. (2011) A dispersive migration in the Atlantic Puffin and its Implications for Migratory Navigation, Plos One, 6(7), e21336. Hall-Spencer J.M., Allain V. & Fossa J.H. (2002) Trawling damage to Northeast Atlantic ancient coral reefs. Proc. Roy. Soc. Lond. B. 269, 507-511. Hall-Spencer, J.M., Tasker, M., Söffker, M., Christiansen, S., Rogers, S., Campbell, M. & Hoydal, K. (2009) The design of Marine Protected Areas on High Seas and territorial waters of Rockall. Mar. Ecol. Prog. Ser. 397, 305-308. Harris, M.P., Daunt, F., Newell, M., Phillips, R.A., Wanless, S. (2010) Wintering areas of Atlantic Puffins Fratercula arctica from a North Sea colony as revealed by geolocation technology, Mar Biol, 157, 827-836 Harvey, R., Gage, J.D., Billett, D.S.M., Clark, A.M. and Paterson, G.L.J. (1988) Echinoderms of the Rockall Trough and adjacent areas. 3. Additional records. Bull. Br. Mus. (Nat. Hist.) (Zool.), 54 (4), 153-198. Henry, L-A. & Roberts, J.M. (2007) Biodiversity and ecological composition of macrobenthos on cold-water coral mounds and adjacent off-mound habitat in the bathyal Porcupine Seabight, NE Atlantic. Deep-Sea Res I 54, 654-672 Hogg, M.M., Tendal, O.S., Conway, K.W., Pomponi, S.A., van Soest, R.W.M., Gutt, J., Krautter, M. & Roberts, J.M. (2010) Deep-sea Sponge Grounds: Reservoirs of Biodiversity. UNEP-WCMC Biodiversity Series No. 32. UNEP-WCMC, Cambridge, UK Howell, K., Billett, D.S.M. & Tyler, P.A. (2002). Depth-related distribution and abundance of seastars (Echinodermata:Asteroidea) in the Porcupine Seabight and Porcupine Abyssal Plain, N.E. Atlantic. Deep-Sea Res. I 49, 1901-1920. Howell, K.L. (2010) A benthic classification system to aid in the implementation of marine protected area networks in the deep / high seas of the NE Atlantic. Biol. Cons. 143, 1041–1056. Howell, K.L., Davies J.S., Jacobs, C., and Narayanaswamy B.E. (2007). Broadscale Survey of the Habitats of Rockall Bank, and mapping of Annex I ‘Reef’ Habitat. Joint Nature Conservation Committee Report. No. 422, 165p. Howell, K.L., Davies J.S. & Narayanaswamy, B.E.(2010). Identifying deep-sea megafaunal epibenthic assemblages for use in habitat mapping and marine protected area network design. J. Mar. Biol. Ass. UK 90, 33-68 . Howell, K.L., Davies, J.S., Hughes, D.J. & Narayanaswamy, B.E. (2007) Strategic Environmental Assessment / Special Area for Conservation Photographic Analysis Report. Department of Trade and Industry, Strategic Environmental Assessment Report, UK, p. 163. Unpublished report. Howell, K.L., Holt, R., Pulido Endrino, I. & Stewart, H. (2011) When the species is also a habitat: comparing the predictively modelled distributions of Lophelia pertusa and the reef habitat it forms. Biol. Cons. Howell, K.L., Mowles S. & Foggo, A. (2010) Mounting evidence: near-slope seamounts are faunally indistinct from an adjacent bank. Mar. Ecol. 31, 52-62. Howell, K.L., Rogers, A., Tyler, P.A. & Billett, D.S.M. (2004). Reproductive isolation among morphotypes of the cosmopolitan species Zoroaster fulgens (Asteroidea:Echinodermata). Mar. Biol. 144, 977-984. Husebø, A., Nottestad, L., Fosså, J.H., Furevik, D.M. & Jorgensen, S.B. (2002) Distribution and abundance of fish in deep-sea coral habitats. Hydrobiologia 471, 91-99. 152 ICES Advice 2013, Book 1 Huvenne, V.A.I. et al. (2011) RRS James Cook Cruise 60, 09 May-12 Jun 2011. Benthic habitats and the impact of human activities in Rockall Trough, on Rockall Bank and in Hatton Basin. (National Oceanography Centre Cruise Report, No. 04) Southampton, UK: National Oceanography Centre, Southampton, 133pp. ICES (2007) Report of the Working Group on Deep-Water Ecology (WGDEC), 26–28 February 2007, ICES Cm 2007/aCE:01 Ref lRC. International Council for the Exploration of the Sea, Copenhagen, Denmark, 57pp. ICES 2010. Report of the Working group on the Biology and Assessemnt of Deepwater Fisheries Resources. www.ices.dk Jacobs, C.L., Howell, K.L. (2007) Habitat investigations within the SEA4 and SEA7 area of the UK continental shelf. MV Franklin Cruise 0206, 03-23 Aug 2006. Research and Consultancy Report No. 24. National Oceanography Centre, Southampton. UK. 95pp. Jensen, A. & Frederiksen, R. (1992) The fauna associated with the bank-forming deep-water coral Lophelia pertusa (Scleractinaria) on the Faroe Shelf. Sarsia 77, 53-69. Kenyon, N.H., Akhmetzhanov, A.M., Wheeler, A.J., van Weering, T.C.E., de Haas, H. & Ivanov, M.K. (2003) Giant carbonate mud mounds in the southern Rockall Trough. Marine Geology 195, 5-30. Lampitt, R.S., Billett, D.S.M. & Rice, A.L. (1986) The biomass of the invertebrate megabenthos from 500 to 4100m in the North East Atlantic. Mar. Biol. 93, 69-81. Large, P. A., C. Hammer, O. A. Bergstad, J. D. M. Gordon, and P. Lorance. 2003. Deep-water Fisheries of the Northeast Atlantic: II. Assessment and Management Approaches. J. Northw. Atl. Fish. Sci. 31: 151-163. Le Goff-Vitry & M.C. & Rogers, A.D. (2005) Molecular ecology of Lophelia pertusa in the NE Atlantic. In: Freiwald, A., Roberts, J.M. (Eds.). Cold-water Corals and Ecosystems. Springer-Verlag, Berlin Heidelberg, pp. 653-662. Le Guilloux, E., Hall-Spencer, J.M., Söffker, M.K. & Olu-Le Roy, K. (2010) Association between the squat lobster Gastroptychus formosus (Filhol, 1884) and cold-water corals in the North Atlantic. J. Mar Biol. Ass. UK 90, 1363-1369. MacLachlan SE, Elliot GM, Parson LM (2008) Investigations of the bottom current sculpted margin of Hatton bank, NE Atlantic. Mar. Geol. 253:170–184 Mauchline, J., Ellett, D.J., Gage, J.D., Gordon, J.D.M. & Jones, E.J.W. (1986) A bibliography of the Rockall Trough. Conference Proceedings Symposium on the Oceanography of the Rockall Channel, Edinburgh (UK), 27-29 Mar 1985. Menot, L., Sibuet, M., Carney, R.S., Levin, L.A., Rowe, G.T., Billett, D.S.M., Poore, G., Kitazato, H., Vanreusel, A., Galéron, J., Lavrado, H.P., Sellanes, J., Ingole, B. & Krylova, E. (2010) New Perceptions of Continental Margin Biodiversity. In: McIntyre, A., (Ed). Chapter 5. Life in the World’s Oceans: Diversity, Distribution and Abundance. Wiley-Blackwell. 79-101. Merrett, N.R., Gordon, J.D.M., Stehmann, M. & Haedrich, R.L. (1991) Deep demersal fish assemblage structure in the Porcupine Seabight (eastern North Atlantic): slope sampling by three different trawls compared. J. Mar. Biol, Ass. UK 71, 329-358. Mienis, F., de Stigter, H.C., de Haas, H. & van Weering, T.C.E. (2009) Near-bed particle deposition and resuspension in a cold-water coral mound area at the Southwest Rockall Trough margin, NE Atlantic. Deep-Sea Res. I 56, 10261038. Morrison, C., Ross, S., Nizinski, M., Brooke, S., Järnegren, J., Waller, R., Johnson, R. & King, T. (2011) Genetic discontinuity among regional populations of Lophelia pertusa in the North Atlantic Ocean. Cons. Genetics 12, 713-729 Mortensen, P.B., Hovland, M., Brattegard, T. & Farestveit, R. (1995) Deep-water bioherms of the scleractinian coral Lophelia pertusa (L) at 64 degrees N on the Norwegian Shelf – Structure and associated megafauna. Sarsia 80, 145-158. Myers, A.A. & Hall-Spencer, J.M. (2004) A new species of amphipod crustacean, Pleusymtes comitari sp. nov., associated with Acanthogorgia sp. gorgonians on deep-water coral reefs off Ireland. J. Mar. Biol. Ass. UK 84, 1029-1032. Narayanaswamy, B.E., Howell, K.L., Hughes, D.J., Davies, J.S., Roberts, J.M. & Black, K.D. (2006) Strategic Environmental Assessment Area 7 Photographic Analysis Report. 13. Department of Trade and Industry, Strategic Environmental Assessment Report, UK, p. 179. Unpublished report. Neat, F.C. & Burns, F. 2010. Stable abundance, but changing size structure in grenadier fishes (Macrouridae) over a decade (1998-2008) in which deepwater fisheries became regulated. Deep Sea Res. I. 57, 434-440. Neat, F. & Campbell, N. 2010. Demersal fish diversity of the isolated Rockall plateau compared with the adjacent west coast shelf of Scotland. Biol. J. Linn. Soc. Lond. 104, 138-147. Penny, A.J., Parker, S.J. & Brown, J.H. (2009) Protection measures implemented by New Zealand for vulnerable marine ecosystems in the South Pacific Ocean. Mar Ecol. Prog. Ser. 397, 341-354. Pollock, C & Barton, C. 2006. Offshore seabirds in the SEA 7 area. A report to the UK Department of Trade and Industry. Rice, A.L., Billett, D.S.M., Thurston, M.H. and Lampitt, R.S. (1991). The Institute of Oceanographic Sciences Biology Programme in the Porcupine Seabight: background and general introduction. J. Mar. Biol. Ass. U.K., 71, 281310. Roberts, J.M., Harvey, S.M., Lamont, P.A., Gage, J.D. & Humphery, J.D. (2000) Seabed photography, environmental assessment and evidence for deep-water trawling on the continental margin west of the Hebrides. Hydrobiologia 441: 173-183 ICES Advice 2013, Book 1 153 Roberts, J.M. and shipboard party (2013) Changing Oceans Expedition 2012. RRS James Cook cruise 073 Cruise Report. Heriot-Watt University. 224 pp. Roberts, J.M., Henry, L-A., Long, D. & Hartley, J.P. (2008) Cold-water coral reef frameworks, megafaunal communities and evidence for coral carbonate mounds on the Hatton Bank, north east Atlantic. Facies 54: 297-316 Roberts, J.M., Long, D., Wilson, J.B., Mortensen, P.B. & Gage, J.D. (2003) The cold-water coral Lophelia pertusa (Scleractinia) and enigmatic seabed mounds along the north-east Atlantic margin: are they related? Mar. Poll. Bull. 46, 7-20. Roberts, J.M., Wheeler, A., Freiwald, A. & Cairns, S.D. (2009) Cold-water corals: The biology and geology of deep-sea coral habitats. Cambridge University Press. 334pp. Roberts, J.M., Wheeler, A.J. & Freiwald, A. (2006) Reefs of the deep: The biology and geology of cold-water coral ecosystems. Science 213, 543-547. Rogers, A.D. & Gianni, M. (2010) The implementation of the UNGA Resolutions 61/105 and 64/72 in the management of deep-sea fisheries on the High Seas. Report of the Deep-Sea Conservation Coalition. International Programme on the State of the Ocean, London, UK. 97pp. Sayago-Gil M, Long D, Hitchen K, Díaz-del-Río V, Fernández-Salas LM, Durán-Muñoz P (2010) Evidence for currentcontrolled morphology along the western slope of Hatton Bank (Rockall Plateau, NE Atlantic Ocean). Geo-Mar. Lett. 30, 99-111 Söffker, M., Sloman, K.A. & Hall-Spencer, J.M. (2011) In situ observations of fish associated with coral reefs off Ireland. Deep-Sea Res. I 58, 818-825 Stewart, H A, and Davies, J S. 2007. Habitat investigations within the SEA7 and SEA4 areas of the UK continental shelf (Hatton Bank, Rosemary Bank, Wyville Thomson Ridge and Faroe–Shetland Channel). British Geological Survey Commercial Report, CR/07/051. Tittensor, D.P., Baco-Taylor, A.R., Brewin, P., Clark, M.R., Consalvey, M., Hall-Spencer, J.M., Rowden, A.A., Schlacher, T., Stocks, K. & Rogers, A.D. (2009) Predicting global habitat suitability for stony corals on seamounts. J. Biogeog. 36, 1111-1128 Van Dover, C., Smith CR, Ardron J, Dunn D, Gjerde K, Levin L, Smith S and the Dinard Workshop Contributors (2011a). Uncharted waters: Placing deep-sea chemosynthetic ecosystems in reserve. Marine Policy 36, 378-381. Van Dover, C., Smith CR, Ardron J, Dunn D, Gjerde K, Levin L, Smith S and the Dinard Workshop Contributors (2011b). Environmental management of deep-sea chemosynthetic ecosystems: justification of and considerations for a spatially-based approach. International Seabed Authority Technical Study 9. 29pp. Vanreusel, A., Fonseca, G., Danovaro, R., et al. (2010) The contribution of deep-sea macrohabitat heterogeneity to global nematode diversity. Mar. Ecol. 31, 66-77. Wei, C-L. et al. (2010). Global Patterns and Predictions of Seafloor Biomass Using Random Forests. PLoS One 5 (12) http://dx.plos.org/10.1371/journal.pone.0015323. Wilson, J.B., (1979a) The distribution of the coral Lophelia pertusa (L.) [L. prolifera (Pallas)] in the North-East Atlantic. J. Mar. Biol. Ass. UK 59, 149-164. Wilson, J.B., (1979b) ‘Patch’ development of the deep-water coral Lophelia pertusa (L.) on Rockall Bank. J. Mar. Biol. Ass. UK 59, 165-177. 154 ICES Advice 2013, Book 1 Maps and Figures Figure 1 Map of the ABNJ area in the NE Atlantic with boundary of the Hatton-Rockall EBSA outlined in red. This boundary approximates the 1500 m contour. ICES Advice 2013, Book 1 155 Table 2 Contributors of data for the analysis of seabird satellite tracking are as follows; full details about each dataset are available via www.seabirdtracking.org Species Site Owner Corys Shearwater Azores J. González-Solís Corys Shearwater Balearics J. González-Solís Corys Shearwater Canaries J. González-Solís Corys Shearwater Chafarinas J. González-Solís Corys Shearwater Berlengas P. Catry, J.P. Granadeiro Corys Shearwater Selvagens M.A. Dias, P. Catry Corys Shearwater Selvagens M.A. Dias, J.P. Granadeiro Corys Shearwater Veneguera J. González-Solís Sooty Shearwater Bay Fundy R. Ronconi Sooty Shearwater Falklands A. Hedd Sooty Shearwater Gough A. Hedd Great Shearwater Bay Fundy R. Ronconi Great Shearwater Inaccessible Island R. Ronconi, P. Ryan, M. Caroline Martin Manx Shearwater Iceland I.A. Sigurðsson, Y. Kolbeinsson, J. González-Solís Manx Shearwater UK A. Ramsay, J. González-Solís Fea’s Petrel Bugio I. Ramirez, V. Paiva Black-legged Kittiwake Norway T. Boulinier, D. Gremillet, J. González-Solís Little Shearwater Azores V. Neves, J. González-Solís Zino’s Petrel Madeira F. Zino, R.A. Phillips, M. Biscoito Figure 2 156 Species occurrence by month within the Hatton-Rockall area, showing percentage of tracked population for each species (and where relevant subpopulation) found within the area each month. ICES Advice 2013, Book 1 Rights and permissions – Any requests to use the seabird tracking data shown for this site in any publication need to be agreed with the data owners. An initial request should be sent to BirdLife International to coordinate this process. See http://www.seabirdtracking.org/terms.php for full terms of reference ICES Advice 2013, Book 1 157 ANNEX 2 Draft Proforma: Mid-Atlantic Ridge North of the Azores and South of Iceland Presented by: Based on the Joint OSPAR/NEAFC/CBD Scientific Workshop. Reviewed and revised by an ICES expert group. Abstract The Mid-Atlantic Ridge (MAR) is the major topographic feature of the Atlantic Ocean. Within the OSPAR/NEAFC area the Northern MAR separates the Newfoundland and Labrador Basins from the West-European Basin and the Irminger from the Iceland Basin. The ridge crest is generally cut by a deep rift valley along its length, bordered by high rift mountains, which are bordered by high fractured plateaus. This region is largely composed of volcanic rock and is the foundation of the proposed EBSA. Hydrothermal venting occurs along the ridges and small-scale physiographic features, including many small volcanoes (seamounts) and canyons, form near the ridge axis. The Moytirra vent field, within this EBSA, is the only high temperature hydrothermal vent known between the Azores and Iceland. Endemic vent fauna are associated with thermally active areas. The 2,500 m depth contour is used to inform the EBSA boundary. Introduction Mid-ocean ridge systems occupy a third of the ocean floor and are the site of the formation of new Earth’s crust (Heezen 1969). The Mid-Atlantic Ridge (MAR), a tectonic continental plate boundary, is the major topographic feature of the Atlantic Ocean, extending over 12,000 km from Iceland to the Bouvet Triple Junction in the South Atlantic (Figure 1). It divides the ocean longitudinally into two halves, each cut by secondary transverse ridges and interrupted by strike-slip transform faults that offset the ridge in opposing directions on either side of the axis of seafloor spreading (e.g., the Charlie Gibbs Fracture at 53ºN). Compared with other mid-ocean ridges, the MAR is a slow-spreading ridge where new oceanic floor is formed with an average spreading rate of 2.5 cm per year (Malinverno 1990). Hydrothermal venting occurs along the ridges and small-scale physiographic features, including many small volcanoes (seamounts) and canyons, form near the ridge axis; the crest consists mostly of hard volcanic rock. The MAR is an area which captures the Earth’s geological history, with outstanding representation of the major stages of Earth’s history, including the record of life, significant ongoing geological processes in the development of landforms and significant geomorphic or physiographic features. The general physiography of the MAR was documented some time ago (Heezen el al. 1959). The ridge crest is generally notched by a deep rift valley along its length, bordered by high rift mountains, which in turn are bordered by high fractured plateaus (Heezen et al. 1959). These crest zones are generally well defined and present along the full length of the MAR (Malinverno 1990). At approximately 50 -75 km from the axis of the ridge, the crest merges with sediment covered flanks which extend down to the abyssal plain (van Andel and Bowin 1968). The flanks are composed of a succession of smooth shelves, each from 2 to 100 km from the central axis and subdivided into upper, middle, and lower steps (Heezen el al. 1959) extending in some areas to depths of 4,572 m (Tolstoy and Ewing 1949). The flanks are generally covered with soft sediments. However, van Andel and Bowin (1968) describe considerable variability in sediment depth in the southern MAR, where the foothill region west of the ridge and the ridge slope are only thinly covered, while sedimentation in some valleys can range from nothing to a thickness of several hundred meters. The depth of the ridge crest is highly variable along its length. Malinverno (1990) conducted 46 profiles across the ridge axis from 0° to 50°N, with most of those conducted south of the Azores between 10° and 35°N. The average depth of the axial crest in those profiles was approximately 2,300 m but Malinverno demonstrated that depth was correlated with distance from the Azores and fracture zone characteristics. The MAR is divided into the Northern and Southern ridges near the equator by the deep Romanche Trench. Within the OSPAR area the Northern MAR separates the Newfoundland and Labrador Basins from the West-European Basin and the Irminger from the Iceland Basin. It has a profound role in the circulation of the water masses in the North Atlantic (Rossby 1999, Bower et al. 2002, Søiland et al. 2008) with currents crossing the MAR over deep gaps in the ridge and influencing upper-ocean circulation patterns (Bower et al. 2002). Canyons cut into the flanks may influence upward fluxes of water and abyssal mixing (Speer and Thurnherr 2005). Studies of volcanic rocks from the submerged MAR suggest that it consists largely of tholeiitic basalt with low values of K, Ti, and P. In contrast, the volcanic islands which form the elevated caps on the Ridge are built of alkali basalt with high values of Ti, Fe3+, P, Na, and K (Engel and Engel 1964). Variations in mineral content result from chemical and isotopic heterogeneity in the mantle (White and Schilling 1978). 158 ICES Advice 2013, Book 1 Moytirra Vent Field The Moytirra vent field is the only fully described high temperature hydrothermal vent known between the Azores and Iceland, making it a unique geophysical structure in the high seas of the North Atlantic and within the MAR. It is situated at 45°N on the 300 m high fault scarp of the eastern axial wall of the MAR, 3.5 km from the axial volcanic ridge crest (Wheeler et al. 2013). It is basalt-hosted and its position suggests that it is heated by an off-axis magma chamber. This type of base rock causes precipitation of iron and sulphide-rich minerals during mixing of the hot hydothermal vent fluids (200-400°C) with cold, oxygenated sea water- hence the term “black smoker” (Figure 2). The Moytirra vent field consists of three active vent sites emitting “black smoke" and producing a complex of chimneys and beehive diffusers. The largest chimney is 18 m tall and very actively venting. There may also be further unconfirmed vent sites on the MAR at 43 N, at 45 N and on the Reykjanes Ridge. In these areas, plumes and/or anomalously high concentrations of Mn in the water column have been detected http://www.interridge.org/irvents/ Location The Mid-Atlantic Ridge (MAR) extends over 12,000 km from Iceland to the Bouvet Triple Junction in the South Atlantic (Figure 1) and falls within the national jurisdictions of Iceland and the Azores. The proposed EBSA Mid-Atlantic Ridge North of the Azores and South of Iceland is for a portion of the MAR within the high-seas areas of OSPAR and NEAFC (Figure 3). Although the crest has an average depth of approximately 2,300 m (Malinverno 1990) it is variable, and the 2,500 m depth contour was used to inform the boundaries of the proposed EBSA as this captures the majority of the ridge crest, and known distribution of deep-water corals (maximum 2,400 m) (Figure 3, Table 1). Within the proposed EBSA the smaller physiographic feature, the Moytirra Vent Field, is located at latitude 45.833 and longitude -27.85 (Table 1). Feature description The Mid-Atlantic Ridge North of the Azores and South of Iceland is a unique geomorphological feature to the North Atlantic Ocean and to the high-seas areas of NEAFC and OSPAR. Within this feature is a smaller unique feature, the Moytirra vent field. The Moytirra vent field is the only high temperature hydrothermal vent known between the Azores and Iceland, making it a unique geophysical structure in the high seas of the North Atlantic and within the MAR. The fauna of the Northern MAR have not been fully described and it is premature to speculate on whether any species are endemic, excepting vent-endemic organisms associated with the hydrothermal vents. Some new species have been described and these may prove to be endemic to the proposed EBSA with further sampling. The benthic fauna associated with the Northern MAR are known from detailed observations at a few locations. Priede et al. (2013) used a variety of sampling gears to survey habitat, biomass and biodiversity in a segment of the Northern MAR as part of a multinational and multidisciplinary project (MAR-ECO). They found that primary production and export flux over the MAR were not enhanced compared with a nearby reference station over the Porcupine Abyssal Plain and biomass of benthic macrofauna and megafauna were similar to global averages at the same depths. Also as part of MAR-ECO, Mortensen et al. (2008) used an ROV to conduct video surveys along the MAR at 8 sites between the Reykjanes Ridge and the Azores. Deep-water corals were observed at all locations at depths less than 1400 m (range 800-2400 m) and 40 coral taxa were observed, including observations of patches of Lophelia pertusa. Crinoids, sponges, the bivalve Acesta excavata, and squat lobsters were associated with the Lophelia. None of those corals were recognized as new species to science and all likely have broader distributions extending along the continental slopes and seamounts at similar latitudes in the North Atlantic. Inevitably, 11 new species have been described arising from the MAR-ECO work and more are likely to be discovered as the samples are fully processed. These include a new genus and species of foram (Incola arantius gen. et sp. nov.), two new species of glass sponges of the genus Sympagella (Rossellidae), mushroom corals (Anthomastus gyratus sp. nov. and Heteropolypus sol sp. nov.), a deep-sea scavenging amphipod (Hirondellea namarensis sp. nov.), two new starfish (species of Hymenaster) and three species of elasipodid holothurians (Gebruk et al. 2013). MAR-ECO midwater and bottom trawls collected 54 species of cephalopods in 29 families (Vecchione et al. 2010). The squid Gonatus steenstrupi was the most abundant cephalopod in the samples, followed by the squids Mastigoteuthis agassizii and Teuthowenia megalops. A multispecies aggregation of large cirrate octopods dominated the demersal cephalopods. The demersal fish fauna of the MAR form two distinct groups with a faunal divide between 48 and 52°N (Hareide and Garnes 2001) and species-specific differences with depth (King et al. 2006, Bergstad et al. 2008). Hareide and Garnes (2001), using one trawl and three longline surveys, identified 56 species from 27 families of fish from between 400 and 2000 m depth along the MAR. In the northern part of the Ridge (north of 52°N) relatively common sub-Arctic species ICES Advice 2013, Book 1 159 such as Sebastes spp., tusk (Brosme brosme) and Greenland halibut (Reinhardtius hippoglossoides) were dominant while sub-tropical species such as golden eye perch (Beryx splendens) and cardinal fish (Epigonus telescopus) were dominant species below 48°N. During the 2004 MAR-ECO expedition to the MAR, 8518 fish, representing 40 species and 17 families were caught with longlines (Fossen et al. 2008). The 59 longline sets were distributed across the ridge axis at depths ranging from 400 to 4300 m at two locations: just north of the Azores archipelago and in the Charlie–Gibbs Fracture Zone. Chondrichthyans (primarily Etmopterus princeps) dominated the catches and contributed nearly 60% to both total biomass and abundance. King et al. (2006) recorded the scavenging fishes of the MAR using a baited autonomous lander equipped with a time-lapse camera between 924 and 3420 m water depth along 3 east–west transects at 42, 51 and 53°N across the MAR. They photographed 22 taxa with Synaphobranchus kaupii, Antimora rostrata and Coryphaenoides (Nematonurus) armatus dominant. Abyssal species in the axial valley region were C. armatus, Histiobranchus bathybius and Spectrunculus sp. No endemic demersal species have been reported although the zoarcid eelpout Pachycara thermophilum, is a vent-endemic species associated with hydrothermal vents of the MAR (Geistdoerfer 1994). Sutton et al. (2008) examined the assemblage structure and vertical distribution of deep-pelagic fishes relative to MAR with acoustic and discrete-depth trawling surveys in association with MAR-ECO. A 36-station, zig-zag survey along the Mid-Atlantic Ridge from Iceland to the Azores covered the full depth range (0 to >3000 m), from the surface to near the bottom, using a combination of gear types to sample the pelagic fauna. Dominant families of pelagic fish included Gonostomatidae, Melamphaidae, Microstomatidae, Myctophidae, and Sternoptychidae and 99 species of pelagic fish were found concentrated particularly north of the Charlie-Gibbs Fracture Zone. Sutton et al. (2008) found that abundance per volume of deep-pelagic fishes was highest in the epipelagic zone and within the benthic boundary layer (BBL; 0-200 m above the seafloor) while minimum fish abundance occurred at depths below 2300 m but above the BBL. Biomass per volume of deep-pelagic fishes over the MAR reached a maximum within the BBL, revealing a topographic association of a bathypelagic fish assemblage with the mid-ocean ridge system. The dominant component of deep-pelagic fish biomass over the MAR was a wide-ranging bathypelagic assemblage that was remarkably consistent along the length of the ridge from Iceland to the Azores. The authors conclude that special hydrodynamic and biotic features of mid-ocean ridge systems cause changes in the ecological structure of deep-pelagic fish assemblages relative to those at the same depths over abyssal plains. Moytirra Vent Field Due to the unique nature of the Moytirra vent field, the specialized vent fauna associated with it are also unique to the North Atlantic high-seas area. Wheeler et al. (2013) have documented aggregations of gastropods (Peltospira sp.) and populations of alvinocaridid shrimp (Mirocaris sp. and Rimicaris sp.) on the surfaces of the vent chimneys in addition to bythograeid crabs (Segonzacia sp.) and zoarcid fish (Pachycara sp.), all considered hydrothermal vent fauna (van Dover 1995). Feature condition, and future outlook Given the geophysical nature, location and size of the MAR it is unlikely that it will be affected by human activities. Despite its remoteness, the fauna associated with the MAR are not pristine. Starting in the early 1970s with Soviet/Russian trawlers stocks of roundnose grenadier (Coryphaenoides rupestris), orange roughy (Hoplostethus atlanticus) and alfonsino (Beryx splendens) associated with the MAR were exploited (Clark et al. 2007, ICES 2007). It can be assumed that most hills along the ridge were at least explored (usually by midwater trawls operating close to the seafloor), and at least 30 seamounts were also exploited for C. rupestris. After 1982, the targeted fishery for redfish developed, dwarfing the catches of roundnose grenadier. After the transition from Soviet to Russian fisheries, the Russian fishing effort and absolute catch on the MAR was significantly reduced, although catch per fishing day settled at relatively low levels by the end of 1990s and the fishery was still conducted periodically (ICES 2007). The fishery on C. rupestris takes deepwater redfish (Sebastes spp), orange roughy (H. atlanticus), blackscabbard fish (Aphanopus carbo) and deepwater sharks as bycatch (Clark et al. 2007). Longline fishing and near-bottom pelagic trawls have the potential to damage fragile benthic species such as deep-water corals and sponges. The scale of the impact that fishing and other human activities may have had on the MAR fauna is at present unquantified. In 2009 NEAFC adopted measures that close more than 330,000 km2 to bottom fisheries on the MAR until 2015 (Figure 4). According to the International Seabed Authority, exploratory mining for sulfide deposits has already been undertaken on the MAR, while ferromanganese nodules and cobalt-rich ferromanganese crusts have potential for mining interests. The potential impacts on the marine environment are removal of organisms and their habitats along with the mineral deposits and the smothering of adjacent communities by any sediment plume that may be created (http://www.isa.org.jm/en/about/faqs#16 ). Representatives from Norway, Iceland, Azores,the United Kingdom, IUCN and UNESCO have met to review the geological and biological heritage of the MAR in the North Atlantic (http://whc.unesco.org/en/activities/504/ ). 160 ICES Advice 2013, Book 1 Discussions focused on the areas where the highest peaks of the mountain chain reach sea-level and form islands. Most of the underwater ridge was not considered because it lies outside any national territory and therefore is not covered by provisions of the World Heritage Convention. There was agreement to encourage cooperation with other conventions in order to better protect the biological, cultural and geological heritage of the ridge. Assessment against CBD EBSA Criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Explanation for ranking Uniqueness or rarity High X The Mid-Atlantic Ridge (MAR) qualifies as a unique geomorphological feature in the North Atlantic. The Moytirra vent field is the only known high temperature hydrothermal vent between the Azores and Iceland, making it a unique geophysical structure in the high seas of the North Atlantic and within the MAR. Areas that are required for a population to survive and Special importance for thrive life-history stages of species Explanation for ranking Data deficient Importance for Area containing habitat for the survival and recovery of endangered, threatened, declining species or area with threatened, endangered or significant assemblages of such species declining species and/or habitats Explanation for ranking X X Data deficient Areas that contain a relatively high proportion of sensitive Vulnerability, habitats, biotopes or species that are functionally fragile fragility, sensitivity, or (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery slow recovery Explanation for ranking X Deep-water corals were observed at all 8 sites from 3 locations along the proposed EBSA at depths less than 1400 m (range 8002400 m) and 40 coral taxa were observed, including observations of patches of Lophelia pertusa. These taxa are fragile with slow recovery and highly susceptible to degradation or depletion by human activities including contact with bottom fishing gear (longlines, pots, trawls). Area containing species, populations or communities with Biological X comparatively higher natural biological productivity productivity Explanation for ranking The research conducted through the MAR-ECO project found that primary production and export flux over the MAR were not enhanced compared with a nearby reference station over the Porcupine Abyssal Plain and biomass of benthic macrofauna and megafauna were similar to global averages at the same depths. There is some evidence for pelagic fish concentrating in the benthic boundary layer (to 200 m above the seafloor) over the MAR in association with topographic features. Area contains comparatively higher diversity of ecosystems, habitats, communities, or species, or has higher genetic diversity Explanation for ranking Biological diversity X Data deficient. Diversity of habitats is greater than that of surrounding abyssal plain but with the exception of the vent fauna, habitats and species are generally shared with continental margins and seamounts not associated with the MAR. ICES Advice 2013, Book 1 161 Sharing experiences and information applying other international criteria (Optional) CBD EBSA Criterion Description Dependency: An area where ecological processes are highly dependent on biotically structured systems (e.g., coral reefs, kelp forests, mangrove forests, seagrass beds). Such ecosystems often have high diversity, which is dependent on the structuring organisms. Dependency also embraces the migratory routes of fish, reptiles, birds, mammals, and invertebrates. Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know Explanation for ranking Representativeness: An area that is an outstanding and illustrative example of specific biodiversity, ecosystems, ecological or physiographic processes Explanation for ranking Biogeographic importance: An area that either contains rare biogeographic qualities or is representative of a biogeographic “type” or types, or contains unique or unusual biological, chemical, physical, or geological features X Explanation for ranking The Mid-Atlantic Ridge (MAR) qualifies as a unique geomorphological feature in the North Atlantic. The Moytirra vent field is the only known high temperature hydrothermal vent between the Azores and Iceland, making it a unique geophysical structure in the high seas of the North Atlantic and within the MAR. Fauna endemic to the vents have adapted to the chemical and thermal properties of the environment. Structural complexity: An area that is characterized by complex physical structures created by significant concentrations of biotic and abiotic features. Explanation for ranking Natural Beauty: An area that contains superlative natural phenomena or areas of exceptional natural beauty and aesthetic importance. Explanation for ranking Earth’s geological history: An area with outstanding examples representing major stages of Earth’s history, including the record of life, significant on-going geological processes in the development of landforms, or significant geomorphic or physiographic features. X Explanation for ranking The Mid-Atlantic Ridge is the site of significant on-going geological processes (plate tectonics, formation of new Earth’s crust) and of significant physiographic features (axial rift valley, hydrothermal vent fields). [Other relevant criterion] Explanation for ranking [Other relevant criterion] Explanation for ranking 162 ICES Advice 2013, Book 1 References Bergstad, O.A., Menezes,G. and Å.S. Høines. 2008. Demersal fish on a mid-ocean ridge: Distribution patterns and structuring factors. Deep Sea Research Part II: Topical Studies in Oceanography 55(1–2): 185-202. Bower, A.S., Le Cann, B., Rossby, T., Zenk, W., Gould, J., Speer, K., Richardson, P.L., Prater, M.D. and H.-M. Zhang. 2002. Directly measured mid-depth circulation in the northeastern North Atlantic Ocean. Nature 419: 603- 607. Clark, M.R., Vinnichencko, V.I., Gordon, J.D.M., Beck-Bulat, G.Z., et al. 2007. Large-scale distant-water trawl fisheries on seamounts. In: Pitcher, T.J., Morato, T., Hart, P.J.B., Clark, M.R., Haggan, N. and Santos, R.S. (eds) Seamounts: Ecology, Conservation and Management. Fish and Aquatic Resources Series, Blackwell, Oxford, UK. Chapter 17, pp. 361 – 399. Engel, A.E.J. and C.G. Engel. 1964. Composition of basalts from the Mid-Atlantic Ridge. Science 144 (3624): 13301333. Fossen, I., Cotton, C.F., Bergstad, O.A. and J.E. Dyb. 2008. Species composition and distribution patterns of fishes captured by longlines on the Mid-Atlantic Ridge. Deep Sea Research Part II: Topical Studies in Oceanography 55(1–2): 203-217. Gebruk , A.V., Priede, I.G., Fenchel T. and F. Uiblein. 2013. Benthos of the sub-polar front area on the Mid-Atlantic Ridge: Results of the ECOMAR project. Marine Biology Research 9(5-6): 443-446. Geistdoerfer, P. 1994. Pachycara thermophilum, une nouvelle espèce de poisson Zoarcidae des sites hydrothermaux de la dorsale médio-atlantique. Cybium 18: 109-115. Hareide, N.-R. and G. Garnes. 2001. The distribution and catch rates of deep water fish along the Mid-Atlantic Ridge from 43 to 61°N. Fisheries Research 51(2–3): 297-310. Heezen, B.C.,Tharp, M. and M. Ewing. 1959. The Floors of the Oceans: I. The North Atlantic. The Geological Society of America Special Paper 65, 122 pp. Heezen, BC. 1969. The world rift system: An introduction to the symposium. Tectonophysics 8:269-279. ICES. 2007. Report of the Working Group on Deep-water Ecology (WGDEC), 26 – 28th February. ICES CM 2007/ACE:01 Ref. LRC. 61pp. King, N.J., P.M. Bagley and I.G. Priede. 2006. Depth zonation and latitudinal distribution of deep-sea scavenging demersal fishes of the Mid-Atlantic Ridge, 42 to 53ºN. Marine Ecology Progress Series 319: 263-274. Malinverno, A. 1990. A quantitative study of axial topography of the Mid-Atlantic Ridge. Journal of Geophysical Research 95: 2645-2660. Mortensen, P.B., Buhl-Mortensen, L., Gebruk, A.V. and E. M. Krylova. 2008. Occurrence of deep-water corals on the Mid-Atlantic Ridge based on MAR-ECO data. Deep-Sea Research II 55:142-152. Priede, I.G., Bergstad, O.A., Miller, P.I., Vecchione, M., Gebruk, A., et al. 2013. Does Presence of a Mid-Ocean Ridge Enhance Biomass and Biodiversity? PLoSONE 8(5): e61550. doi:10.1371/journal.pone.0061550 Rossby, T. 1999. On gyre interactions. Deep-Sea Research II 46: 139-164. Søiland, H., Budgell, W.P. and Ø Knutsen. 2008. The physical oceanographic conditions along the Mid-Atlantic Ridge north of the Azores in June-July 2004. Deep-Sea Research II 55: 29- 44. Speer, K. G. and A. M. Thurnherr. 2005. Abyssal Canyons and Mixing by Low-Frequency Flow. In: P. Muller and D. Henderson (Editors), Near-Boundary Processes and Their Parameterization, topics in physical oceanography, 'Aha Huliko' a Winter Workshop, pp. 17-19. Sutton, T.T., Porteiro, F.M., Heino, M., et al. 2008. Vertical structure, biomass and topographic association of deeppelagic fishes in relation to a mid-ocean ridge system. Deep-Sea Research II 55 (1–2):161–184. Tolstoy, I. and M. Ewing. 1949. North Atlantic hydrography and the Mid-Atlantic Ridge. Bulletin of the Geological Society of America 60(10): 1527-1540. van Andel, Tj. H. and C. O. Bowin. 1968. Mid-Atlantic Ridge between 22º and 23º north latitude and the tectonics of mid-ocean rises. Journal of Geophysical Research 73: 1279-1298. Van Dover, C.L. 1995. Ecology of Mid-Atlantic Ridge hydrothermal vents. Geological Society, London, Special Publications 1995, v. 87, p257-294. Vecchione, M., Young, R.E. and U. Piatkowski. 2010. Cephalopods of the northern Mid-Atlantic Ridge. Marine Biology Research 6: 25–52. Wheeler, A.J., Murton, B., Copley, J., Lim, A., Carlsson, J. et al. 2013. Moytirra: discovery of the first known deep-sea hydrothermal vent field on the slow-spreading Mid-Atlantic Ridge north of the Azores. Geochemistry, Geophysics, Geosystems, doi: 10.1002/ggge.20243. White, W.M. and J.-G. Schilling. 1978. The nature and origin of geochemical variation in Mid-Atlantic Ridge basalts from the Central North Atlantic. Geochimica et Cosmochimica Acta 42 (10): 1501-1516. ICES Advice 2013, Book 1 163 Tables, Maps and Figures Table 1 Boundaries for the proposed EBSA Mid-Atlantic Ridge North of the Azores and South of Iceland and location of the Moytirra Vent Field (see Figure 3). Feature Point 1 Point 2 Point 3 Point 4 Point 5 Point 6 EEZ of the Azores Latitude 63.59 55.85 52.97 52.50 47.11 43.18 Point 7 Point 8 Point 9 Point 10 Point 11 Point 12 EEZ of Iceland 42.50 46.43 49.43 52.50 53.78 60.13 Moytirra Vent Field 164 (dd) Longitude (dd) -30.91 -37.51 -36.70 -33.00 -28.09 -30.59 Details of Location Reference easterly following the EEZ boundary for the Azores to Point 7 45.4833 -27.55 -25.91 -26.63 -29.80 -33.69 -22.84 -27.85 westerly following the EEZ boundary for Iceland to join Point 1 300 m high fault scarp of the eastern axial wall, 3.5 km from the axial volcanic ridge crest Wheeler et al. (2013) ICES Advice 2013, Book 1 Figure 1 Location of the Mid-Atlantic Ridge (dashed lines). Image downloaded from: commons.wikimedia.org File:Mid-atlantic ridge.jpg - Wikimedia Commons. ICES Advice 2013, Book 1 165 Figure 2 166 Volcanically heated fluid rises from a deep-sea "black-smoker" in the Moytirra vent field. (Photo http://news.nationalgeographic.com/news/2011/08/110808-hydrothermal-ventsdownloaded from volcanic-animals-ocean-deep-sea-science-alien/ ) ICES Advice 2013, Book 1 Figure 3 Location of the proposed EBSA Mid-Atlantic Ridge North of the Azores and South of Iceland. The perimeter of the proposed boundary follows Table 1 with points numbered 1 through 12. The blue shaded areas represent the Exclusive Economic Zones of countries in the region. The light green line outlines the outer boundary of the OSPAR Convention Area which coincides with the NEAFC Convention Area at its western and southern boundary. Seafloor bathymetry is indicated in grayscale with the 2000 m (red) and 2500 m (light blue) General Bathymetric Chart of the Oceans (GEBCO) depth contours indicated. The red circle with the central star marks the location of the Moytirra Vent Field. See Table 1 for positional information (latitude and longitude). ICES Advice 2013, Book 1 167 Figure 4 168 Location of areas closed to bottom fishing (2009-2015) in the NEAFC regulatory area which includes the MAR http://www.neafc.org/page/closures. There is a small seasonal closure for Blue Ling near the boundary with Iceland EEZ which is not visible at this scale. ICES Advice 2013, Book 1 ANNEX 3 Draft Proforma: Charlie-Gibbs Fracture Zone Presented by: Based on the Joint OSPAR/NEAFC/CBD Scientific Workshop. Reviewed by revised by an ICES expert group Abstract Fracture zones are common topographic features of the global oceans that arise through plate tectonics. The CharlieGibbs Fracture Zone is an unusual left lateral strike-slip double transform fault in the North Atlantic Ocean along which the rift valley of the Mid-Atlantic Ridge is offset by 350 km near 52º30′N. It opens the deepest connection between the northwest and northeast Atlantic (maximum depth of approximately 4500 m) and is approximately 2000 km in length extending from about 25°W to 45°W. It is the most prominent interruption of the MAR between the Azores and Iceland and the only fracture zone between Europe and North America that has an offset of this size. Two named seamounts are associated with the transform faults: Minia and Hecate. The CGFZ is considered a unique geomorphological feature in the North Atlantic under the EBSA criteria; further, it captures the Earth’s geological history, including significant ongoing geological processes. Introduction Fracture zones are common topographic features of the global oceans that arise through plate tectonics. They are characterized by two strongly contrasting types of topography. Seismically active transform faults form near mid-ocean ridges where the continental plates move in opposing directions at their junction. Seismically inactive fracture zones, where the plate segments move in the same direction, extend beyond the transform faults often for 100s of kilometers. Their atypical crust thickness that can be as little as 2 km (Mutter et al. 1984, Cormier et al. 1984, Calvert and Whitmarsh 1986) allowing direct seismic investigations of the internal structure and composition of oceanic crusts used to model processes of seafloor spreading. In the Atlantic Ocean most fracture zones originate from the Mid-Atlantic Ridge (MAR) and are nearly perfectly west - east oriented. There are about 300 fracture zones occurring on average every 55 km along the ridge, with the offsets created by transform faults ranging from 9 to 400 km in length (Müller and Roest 1992). The Charlie-Gibbs Fracture Zone (CGFZ) is an unusual left lateral strike-slip double transform fault in the North Atlantic Ocean along which the rift valley of the MAR is offset by 350 km near 52º30′N (Figure 1). It opens the deepest connection between the northwest and northeast Atlantic (maximum depth of approximately 4500 m; Fleming et al. 1970) and is approximately 2000 km in length extending from about 25°W to 45°W. It is the most prominent interruption of the MAR between the Azores and Iceland and the only fracture zone between Europe and North America that has an offset of this size 11. Knowledge of its geomorphology is considered essential to the understanding of the plate tectonic history of the Atlantic north of the Azores (Olivet et al. 1974). For these reasons it is consider a unique geomorphological feature in the North Atlantic under the EBSA criteria; further, it captures the Earth’s geological history, including significant on-going geological processes. The CGFZ is comprised of two narrow parallel fracture zones (Fleming et al. 1970) which form deep trenches located at 30ºW (Charlie-Gibbs South Transform Fault) and at 35º15′W (Charlie-Gibbs North Transform Fault) and separated by a short (40 km) north-south seismically active (Bergman and Solomon 1988) spreading center (median transverse ridge) at 31º45’W (Figure 2; Searle 1981; Fleming et al. 1970, Olivet et al. 1974). The southern fault displaces the MAR, coming from the Azores, to the west over a distance of 120 km. It is at most 30 km wide (Searle 1981). The northern fault displaces the spreading ridge over another 230 km to the west before it connects to the northern part of the MAR going to Iceland. Both transform faults continue eastward and westward as inactive fracture zones (Figure 2). The CGFZ is characterized by rough morphology and the walls of the fracture valleys and the ridge in between them are broken and irregular with slopes of up to 29° (Fleming et al. 1970). The height of the ridge between the faults is at least 1000 m below the surface and as shallow as 636 m in parts (Fleming et al. 1970). Rock samples show the walls of the fracture zone to be both basaltic and ultramafic while the median transverse ridge contains gabbro (Hekinian and Aumento 1973). Earthquake epicentres are associated with the transform faults (Kanamori and Stewart 1976, Bergman and Solomon 1988) and an almost continuous belt of epicentres follow the southern end of the Reykjanes Ridge, along the northern transform valley, the central median valley and the southern transform valley to the north end of the MAR (Lilwall and Kirk 1985). Two named seamounts are associated with the transform faults: Minia Seamount (53°01′N 34°58′W) located near the junction of the Reykjanes Ridge and the northern transform fault and Hecate Seamount 11 The Spitzbergen and Jan Mayen fracture zones, of comparable offset (145 and 211 km respectively), lie between Greenland and Europe. ICES Advice 2013, Book 1 169 (52°17′N 31°00′W) located on the northern wall of the southern transform fault east of the short median transverse ridge (Figure 2; Fleming et al. 1970). Ridge and troughs along the CGFZ are mostly covered with muddy sediments (Fleming et al. 1970) although outcrops of sedimentary rock and boulder fields are exposed by recent faulting and current scour (Shor et al. 1980, Searle 1981) and the southern transform near 30°30′W has no sediment cover (Searle 1981). Considerable thicknesses of sediment are deposited in the northern transform valley from the Iceland-Scotland Overflow Water (ISOW) which carries a significant load of suspended sediment (25 μg I-1) as it passes through (Shor et al. 1980). Transverse ridges prevent the sediment reaching the southern valley (Searle 1981) which has less sediment cover, although it is still considered a depositional environment (Shor et al. 1980). The topography of the CGFZ has a major influence on deep water oceanographic circulation (Harvey and Theodorou 1986). A large component of the North Atlantic Deep Water originates in the Norwegian Sea and flows south over the sills between Scotland and Iceland (ISOW). It meets the CGFZ near the intersection of the transform faults and the spreading centre (Shor et al. 1980). There is then a westward movement of deep water passing through the fracture zone from east to west through to the Irminger Sea occurring from the core depth of the ISOW at about 2500 m to the sea floor (Garner 1972, Shor et al. 1980, Saunders 1994). Most of this water is carried through the northern transform fault where the overflow water first encounters the fracture zone. The topography of the CGFZ also is thought to have some influence on the circulation of surface waters, although they are not locked to the bottom features to the same extent as the ISOW (Rossby 1999, Bower et al. 2002). The northern branch of the North Atlantic Current defines the location of the sub-polar front between colder Sub Arctic Intermediate Water to the north and warmer North Atlantic Intermediate Water to the south (Søiland et al. 2008). The sub-polar front meanders between 48-53°N and surface flow is predominantly eastward. The CGFZ is therefore not only a topographic discontinuity in the MAR but the area also constitutes an oceanographic transition zone between waters of different temperatures and flow regimes (Priede et al. 2013). Location The Charlie-Gibbs Fracture Zone occurs at 52º30′N and extends from about 25°W to 45°W with the transform faults occurring between 30°W and 35°W (Olivet et al. 1974). The proposed Charlie-Gibbs Fracture Zone EBSA within the high-seas area of NEAFC and OSPAR takes the co-ordinates provided in Table 1 and illustrated in Figure 3. These are based on Olivet et al. (1974) and the location of the Minia Seamount which influences the northern boundary of the EBSA. The eastern boundary of the CGFZ is detectable beyond 42°W, the outer boundary of the NEFAC/OSPAR jurisdictions. The southern ridge continues uninterrupted to 45°W (Olivet et al. 1974). Feature description The Charlie-Gibbs Fracture Zone (CGFZ) is a unique geomorphological feature to the North Atlantic Ocean and to the high-seas areas of NEAFC and OSPAR. Owing to its remoteness, the fauna associated with the CGFZ are poorly studied and it is premature to speculate on whether any species are endemic based on first descriptions. For example, Gebruk (2008) described two species of holothurians and believed them to be endemic to the Mid-Atlantic Ridge but they subsequently were found on the European continental margin in the Whittard Canyon (Masson 2009). As part of the MAR-ECO project (Priede et al. 2013) manned submersibles were deployed on the axis (52°47′N) and the northern slopes (52°58′N) of the Charlie–Gibbs North transform fault and surveyed macroplankton (Vinogradov 2005), demersal nekton (Felley et al. 2008) and invertebrate megafauna (Gebruk and Krylova 2013). Pelagic shrimps, chaetognaths and gelatinous animals were numerically dominant in the plankton, with peak densities corresponding to the main pycnocline. Mucous houses of appendicularians were abundant at 150 m above the seabed, although this is common throughout the central Atlantic and not associated with specific bottom topography (Vinogradov 2005). Nekton included large and small macrourids (Coryphaenoides spp.), shrimp (infraorder Penaeidea), Halosauropsis macrochir, Aldrovandia sp., Antimora rostrata, and alepocephalids (Felley et al. 2008). Glass sponges were common between 1700 and 2500 m while the deeper parts of the fracture wall and the sea floor were dominated by isidid corals, other anthozoans, squat lobsters and echinoderms, especially holothurians. The elpidiid holothurian, Kolga nana, occurred at high density in the abyssal depression (Gebruk and Krylova 2013). Rogacheva et al. (2013) recorded 32 holothurian species from the CGFZ area through the ECOMAR project (http://www.oceanlab.abdn.ac.uk/ecomar/), including three elasipodid holothurian species new to science. In general, none of the fauna documented from the CGFZ showed distributions atypical of similar habitats in the broader North Atlantic, although Gebruk and Krylova (2013) discuss the known distribution of the holothurian Peniagone longipaillata and remark on the differences in relative abundance observed between the occurrence of this species, where 170 ICES Advice 2013, Book 1 it is common in the lower bathyl of the CGFZ, and the continental slopes in the Porcupine Seabight and Abyssal Plain areas and Whitard Canyon where it appears less so. There is weak evidence that the CGFZ may be important for juvenile zoarcids based on a high percentage of those observed with baited cameras being <100 mm in length (Kemp et al. 2013). General knowledge of seafloor benthos suggests that where the geo-morphological processes of the fracture zone have created steep walls along the fractures, the greater three-dimensional topographic complexity, combined with the strong water flows through the fractures, creates habitat that is likely to be more productive and support greater concentrations of fragile taxa such as deep-water corals and sponges than adjacent habitats (Miller et al. 2012). The sampling done along the fracture zone supports these inferences but the differences from other habitats in similar depths and latitudes have not been quantified yet. Feature condition, and future outlook Given the geophysical nature, location and size of the Charlie-Gibbs Fracture Zone (CGFZ) it is unlikely that it will be affected by human activities, although there is potential for mining of the rare minerals associated with the transform faults. In 2010 the Environmental Ministers of the OSPAR countries officially designated a marine protected area of 145,420 km2 in the southern part of the Charlie-Gibbs Fracture Zone (Figure 4) and adopted “significant and innovative measures to establish and manage the southern part of the originally proposed Charlie-Gibbs Fracture Zone MPA – “Charlie-Gibbs South MPA”-, for which the seabed and super adjacent waters are situated in areas beyond national jurisdiction” (OSPAR Commission 2010). That same year (2010) the OSPAR Commission and the International Seabed Authority signed a memorandum of understanding in order to conciliate the development of mineral resources with comprehensive protection of the marine environment. In this MOU, the Charlie Gibbs Fracture Zone is highlighted as an area where consultation between the two parties had been initiated. In 2012 OSPAR countries designated “ Charlie-Gibbs North High Seas Marine Protected Area”, an area of high seas of approximately 177,700 km2 (OSPAR Commission 2012), complementing the Charlie-Gibbs South MPA established previously (Figure 4). The scale of the impact that fishing and other human activities have had on the fauna of the CGFZ is at present unquantified and likely to be minor, although fishing has been reported on the Hectate Seamount (ICES 2007). In 2009 NEAFC closed more than 330,000 km2 to bottom fisheries on the Mid-Atlantic Ridge, including a large section of the CGFZ which includes the transform faults and median transverse ridge (http://www.neafc.org/page/closures) (Figure 4). Assessment against CBD EBSA Criteria [Discuss the case study in relation to each of the CBD criteria and relate the best available science. Note that a candidate EBSA may qualify on the basis of one or more of the criteria, the boundaries of the EBSA need not be defined with exact precision. And modeling may be used to estimate the presence of EBSA attributes. Please note where there are significant information gaps.] CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Explanation for ranking Uniqueness rarity or High X The Charlie-Gibbs Fracture Zone (CGFZ) is a unique geomorpholical feature in the high-sea between the Azores and Iceland. It is the only fracture zone with an offset of its size (350 km) between Europe and North America and opens the deepest connection between the northwest and northeast Atlantic. The fact that it is a double transform fault is an unusual feature. Areas that are required for a population to survive and Special importance for thrive life-history stages of species Explanation for ranking X Data deficient ICES Advice 2013, Book 1 171 Importance for Area containing habitat for the survival and recovery of endangered, threatened, declining species or area with threatened, endangered or significant assemblages of such species declining species and/or habitats Explanation for ranking X Data deficient Areas that contain a relatively high proportion of sensitive Vulnerability, habitats, biotopes or species that are functionally fragile fragility, sensitivity, or (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery slow recovery Explanation for ranking X Glass sponges were observed on hard substrates on the fault wall at depths between 1700 and 2500 m. These taxa are fragile with slow recovery and highly susceptible to degradation or depletion by human activities including contact with bottom fishing gear (longlines, pots, trawls).Inferring from the frequently documented presence of such species and communities in structurally complex deep-sea habitats elsewhere, further sampling is likely to document additional presence of sensitive habitats, biotopes, or species in the CGFZ fractures, Area containing species, populations or communities with X Biological comparatively higher natural biological productivity productivity Explanation for ranking There is no evidence that the CGFZ contains comparatively higher natural productivity. The strong current flows through the fractures and complex three dimensional habitats create conditions that may enhance productivity, but at present there are insufficient data to rank on this criterion. Area contains comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or has diversity higher genetic diversity Explanation for ranking Diversity of habitats is greater than that of surrounding abyssal plain but biotic diversity is poorly quantified and there is little basis for a comparative assessment on this criterion at this time. Sharing experiences and information applying other international criteria (Optional) CBD EBSA Criterion Description Dependency: An area where ecological processes are highly dependent on biotically structured systems (e.g., coral reefs, kelp forests, mangrove forests, seagrass beds). Such ecosystems often have high diversity, which is dependent on the structuring organisms. Dependency also embraces the migratory routes of fish, reptiles, birds, mammals, and invertebrates. Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know Explanation for ranking Representativeness: An area that is an outstanding and illustrative example of specific biodiversity, ecosystems, ecological or physiographic processes Explanation for ranking Biogeographic importance: An area that either contains rare biogeographic qualities or is representative of a biogeographic “type” or types, or contains unique or unusual biological, chemical, physical, or geological features X Explanation for ranking The CGFZ qualifies as a unique geomorphological feature in the North Atlantic being the largest transform fault separating Europe from North America. An area that is characterized by complex physical Structural complexity: structures created by significant concentrations of biotic and abiotic features. Explanation for ranking 172 ICES Advice 2013, Book 1 Natural Beauty: An area that contains superlative natural phenomena or areas of exceptional natural beauty and aesthetic importance. Explanation for ranking Earth’s geological history: An area with outstanding examples representing major stages of Earth’s history, including the record of life, significant on-going geological processes in the development of landforms, or significant geomorphic or physiographic features. X Explanation for ranking Fracture zones are of great geological interest due to their anomalous crust thickness that can be as little as 2 km allowing direct seismic investigations of the internal structure and composition of oceanic crusts used to model processes of seafloor spreading. Knowledge of the geomorphology of the CGFZ is considered essential to the understanding of the plate tectonic history of the Atlantic north of the Azores. [Other relevant criterion] Explanation for ranking [Other relevant criterion] Explanation for ranking References Bergman, E.A. and S.C. Solomon. 1988. Transform fault earthquakes in the North Atlantic: Source mechanisms and depth of faulting. Journal of Geophysical Research 93:9027-9057. Bower, A.S., Le Cann, B., Rossby, T., Zenk, W., Gould, J., Speer, K., Richardson, P.L., Prater, M.D. and H.-M. Zhang. 2002. Directly measured mid-depth circulation in the northeastern North Atlantic Ocean. Nature 419: 603- 607 Calvert, A.J. and R.B. Whitmarsh. 1986. The structure of the Charlie-Gibbs Fracture Zone. Journal of the Geological Society 1433: 819-821. Cormier, M.-H., Detrick, R. S. and G. M. Purdy. 1984. Anomalously thin crust in oceanic fracture zones: New seismic constraints from the Kane Fracture Zone. Journal of Geophysical Research 89:249–266. Felley, J.D., Vecchione, M. And R.R. Wilson Jr. 2008. Small-scale distribution of deep-sea demersal nekton and other megafauna in the Charlie-Gibbs Fracture Zone of the Mid-Atlantic Ridge. Deep Sea Research II 55: 153-160. Fleming, H.S., Cherkis, N.Z. and J.R. Heirtzler. 1970. The Gibbs Fracture Zone: A double fracture zone at 52°30′N in the Atlantic Ocean. Marine Geophysical Researches 1:37-45. Garner, D.M. 1972. Flow through the Charlie-Gibbs Fracture Zone, Mid-Atlantic Ridge. Canadian Journal of Earth Sciences 9: 116-121. Gebruk A.V. 2008. Holothurians (Holothuroidea, Echinodermata) of the northern Mid-Atlantic Ridge collected by the G.O. Sars MAR-ECO expedition with descriptions of four new species. Marine Biology Research 4, 48–60. Gebruk, A.V. and E.M. Krylova. 2013. Megafauna of the Charlie–Gibbs Fracture Zone (northern Mid-Atlantic Ridge) based on video observations. Journal of the Marine Biological Association of the United Kingdom 93: 1143-1150. doi:10.1017/S0025315412001890. Harvey, J.G. and A. Theodorou. 1986. The circulation of Norwegian Sea overflow water in the eastern North Atlantic. Oceanologica Acta 9: 393-402. Hekinian, R. and F. Aumento. 1973. Rocks from the Gibbs Fracture Zone and the Minia Seamount near 53°N in the Atlantic Ocean. Marine Geology 14: 47-72. ICES. 2007. Report of the Working Group on Deep-water Ecology (WGDEC), 26 – 28th February. ICES CM 2007/ACE:01 Ref. LRC. 61pp. Kanamori, H. and G.S. Stewart. 1976. Mode of the strain release along the Gibbs Fracture Zone, Mid-Atlantic Ridge. Physcis of the Earth and Planetary Interiors 11: 312-332. Kemp, K.M., Jamieson, A.J., Bagley, P.M., Collins, M.A. and I.G. Priede. 2008. A new technique for periodic bait release at a deep-sea camera platform: First results from the Charlie-Gibbs Fracture Zone, Mid-Atlantic Ridge. Deep-Sea Research II 55: 218-228. Lilwall, R.C. and R.E. Kirk. 1985. Ocean-bottom seismograph observations on the Charlie-Gibbs fracture zone. Geophysical Journal of the Royal Astronomical Society 80: 195-208. Masson, D.G. 2009. The Geobiology of Whittard Submarine Canyon. RRS James Cook Cruise 36, 19 June–28 July 2009. National Oceanography Centre, Southampton, 53 pp. http://www.eprints.soton.ac.uk/69504/1/nocscr041.pdf. ICES Advice 2013, Book 1 173 Miller, R.J., Hocevar, J., Stone, R.P. and D.V. Fedorov. 2012. Structure-forming corals and sponges and their use as fish habitat in Bering Sea submarine canyons. PLoS ONE 7(3): e33885. doi:10.1371/journal.pone.0033885 Müller, R.D. and W.R. Roest. 1992. Fracture zones in the North Atlantic from combined Geosat and Seasta data. Journal of Geophysical Research 97: 3337-3350. Mutter, J.C., Detrick, R.S. and North Atlantic Transect Study Group. 1984. Multichannel seismic evidence for anomalously thin crust at Blake Spur fracture zone. Geology 12: 534-537. Olivet, J.-L., Le Pichon, Xl, Monti, S. and B. Sichler. 1974. Charlie-Gibbs Fracture Zone. Journal of Geophysical Research 79: 2059-2072. OSPAR Commission. 2010. OSPAR Convention for the Protection of the Marine Environment of the North-East Atlantic. Meeting of the OSPAR Commission Bergen: 20-24 September 2010. Annex 49 (Ref. M6.2). http://www.ospar.org/content/content.asp?menu=01441000000000_000000_000000 OSPAR Commission. 2012. OSPAR Convention for the Protection of the Marine Environment of the North-East Atlantic. Meeting of the OSPAR Commission Bonn: 25-29 June 2012. Annex 6 (Ref. §5.19a). OSPAR Decision 2012/01. Priede, I.G., Billett, D.S.M., Brierley, A.S., Hoelzel, A.R., Inall, M., Miller, P.I., Cousins, N.J., Shields, M.A. and T. Fujii. 2013. The ecosystem of the Mid-Atlantic Ridge at the sub-polar front and Charlie-Gibbs Fracture Zone; ECO-MAR project strategy and description of the sampling programme 2007-2010. Deep-Sea Research II http://dx.doi.org/10.1016/j.dsr2.2013.06.012i. Rogacheva A, Gebruk A. and C.Alt. 2013. Deep-sea holothurians of the Charlie Gibbs Fracture Zone area, northern MidAtlantic Ridge. Marine Biology Research 9:587_623. Rossby, T. 1999. On gyre interactions. Deep-Sea Research II 46: 139-164. Saunders, P.M. 1994.The flux of overflow water through the Charlie–Gibbs Fracture Zone. Journal of Geophysical Research 99:12343–12355. Searle, R. 1981. The active part of the Charlie-Gibbs Fracture Zone: A study using sonar and other geophysical techniques. Journal of Geophysical Research 86: 243-262. Shor, A., Lonsdale, P., Hollister, C.D. and D. Spencer. 1980. Charlie-Gibbs fracture zone: bottom-water transport and its geological effects. Deep-Sea Research 27A: 325-245. Søiland, H., Budgell, W.P. and Ø Knutsen. 2008. The physical oceanographic conditions along the Mid-Atlantic Ridge north of the Azores in June-July 2004. Deep-Sea Research II 55: 29- 44. Vinogradov, G.M. 2005. Vertical distribution of macroplankton at the Charlie-Gibbs Fracture Zone (North Atlantic), as observed from the manned submersible “Mir-1”. Marine Biology 146: 325-331. 174 ICES Advice 2013, Book 1 Tables, Maps and Figures Table 1. Boundaries for the proposed EBSA Charlie-Gibbs Fracture Zone and location of the Minia and Hecate Seamounts (see Figure 3). Feature CGFZ Proposed EBSA Point 1 CGFZ Proposed EBSA Point 2 CGFZ Proposed EBSA Point 3 CGFZ Proposed EBSA Point 4 CGFZ Proposed EBSA Point 5 Northern Fracture (eastern portion) Southern Fracture (eastern portion) Northern Fracture (western portion) Southern Fracture (western portion) Minia Seamount Hecate Seamount ICES Advice 2013, Book 1 Latitude (dd) Longitude (dd) 51°N 42°W 53°05′N 42°W 53°05′N 30°W 52°30′N 25°W 51°45′N 25°W 53°05′N 42°W 51°N 45°W 52°30′N 27°W 51°45′N 25°W 53°01′N 34°58′W 53º 00.60' N 34º 49.80' W 52°17′N 31°00′W 52º 15.60' N 31º 03.00' W Details Location of Reference Olivet 8) Olivet 8) Olivet 11) Olivet 11) located near the junction of the Reykjanes Ridge and the northern transform fault et al. 1974 (p. 2062, fig. et al. 1974 (p. 2062, fig. et al. 1974 (p. 2063, fig. et al. 1974 (p. 2064, fig. Fleming et al. 1970 Seamount Catalogue http://earthref.org/SC/SMNT530N-0348W/ located on the northern wall of the southern transform fault east of the short median transverse ridge Fleming et al. 1970 Seamount Catalogue http://earthref.org/SC/SMNT530N-0348W/ 175 Figure 1 176 Location of the Charlie-Gibbs Fracture Zone (black lines) in the North Atlantic. The Mid-Atlantic Ridge runs through the centre of the Atlantic Ocean and its left lateral displacement can be clearly seen. Image downloaded from: commons.wikimedia.org File:Charlie-gibbs-full-extent.png - Wikimedia Commons. ICES Advice 2013, Book 1 Figure 2 Schematic of the Charlie-Gibbs Fracture Zone and the Mid-Atlantic Ridge (MAR) indicating the left lateral displacement of the MAR, the North and South transform faults and the central spreading axis. The relative location of two seamounts, Hecate and Minia are illustrated. Image downloaded from: commons.wikimedia.org File:Charliegibbsschema-en.svg- Wikimedia Commons. Figure 3. Proposed Charlie-Gibbs Fracture Zone EBSA (yellow lines). Numbers refer to the points in Table 1. The green line is the NEAFC/OSPAR outer boundary. ICES Advice 2013, Book 1 177 Figure 4. 178 Location of the OSPAR MPAs in the North Atlantic including the large Charlie-Gibbs South and CharlieGibbs North MPAs in the central area. The areas closed to bottom fishing by NEAFC are indicated by the yellow boundaries. Downloaded 10 Sep 2013 from: http://charlie-gibbs.org/charlie/node/70 ICES Advice 2013, Book 1 ANNEX 4 Draft Proforma - The Arctic Ice habitat - multiyear ice, seasonal ice and- marginal ice zone Presented by WWF and reviewed by Participants at the Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas in the North-East Atlantic. Reviewed and revised by an ICES expert group. Abstract The permanently ice covered waters of the high Arctic provide a range of globally unique habitats associated with the variety of ice conditions. In the northern hemisphere multi-year sea ice only exists in the Arctic and although the projections of changing ice conditions due to climate change project a considerable loss of sea ice, in particular multiyear ice, the Eurasian Central Arctic high seas are likely to at least keep the ice longer than many other regions in the Arctic basin. Ice is a crucial habitat and source of particular food web dynamics, the loss of which will affect also a number of mammalian and avian predatory species. The particularly pronounced physical changes of Arctic ice conditions as already observed and expected for the coming decades, will require careful ecological monitoring. Eventually measures will be needed to maintain or restore, to the extent possible the resilience of the Arctic populations to changing environmental conditions. Introduction Over many past millennia, most of the Eurasian part of the Arctic Basin, and in particular the high seas area in the Arctic Ocean (the waters beyond the 200 nm zones of coastal states, i.e. Norway, Russia, USA, Canada and Greenland/Denmark) have been permanently ice covered. However, in recent years, much of the multiyear permanent pack ice has been replaced by seasonal (1 year) ice. In addition, the former fast pack-ice is now increasingly broken up by leads. This structural change in the Arctic ice quality will result in a substantial increase in light penetrating the thin ice and water column, in conjunction with the overall warming of surface waters and increased temperature and salinity stratification due to the melting of ice. Some models predict that before the end of the century the permanent ice cover may disappear completely (Anisimov et al., 2007). The reduction and possible loss of permanent ice cover will result in significant changes in the structure and dynamics of the high Arctic ecosystems (CAFF, 2010; Gradinger, 1995; Piepenburg, 2005; Renaud et al., 2008; Wassmann, 2008, 2011). Understanding and studying the area proposed as EBSA is of particular scientific relevance as already envisaged by the Arctic Council (Gill et al., 2011; Mauritzen et al., 2011). Such studies may in the long-term, become relevant for the commercial exploitation of resources. Location The Ecologically or Biologically Significant Marine Area (EBSA) proposed focuses on the presently permanently icecovered waters in the OSPAR/NEAFC maritime areas, including the high seas section in the Central Arctic Basin north of the 200 nm zones of coastal states, and the area of contiguous seasonal ice directly connected to the multi-year permanent ice (see Fig. 1 attached). Therefore, the boundaries proposed extend from the North Pole (northernmost point of OSPAR/NEAFC maritime areas) to the southern limit of the summer sea ice extent and marginal ice zone, including on the shelf of East Greenland. The proposal currently only relates to features of the water column and the ice surface itself. Two legal states have to be distinguished: the Central Arctic high seas waters north of the 200 nm zones of adjacent coastal states, generally north of 84° N, and the waters within the Exclusive Economic Zones of Greenland, Russia and the fisheries protection zone of Norway around Svalbard. Figure 1 distinguishes between the high seas beyond national jurisdiction for which the ”Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic“ had a mandate12 and national/nationally administered waters within the 200 nm zone (national EEZs), within which the OSPAR Contracting Parties have the competence to report candidate EBSAs to the Convention on Biodiversity EBSA repository (OSPAR Commission, 2011). Participant Briefing for a Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic. Invitation Annex 2, 2011 12 ICES Advice 2013, Book 1 179 The seafloor of the respective region will likely fall on the extended continental shelves of several coastal states. It belongs to the ”Arctic Basin“ region of (Gill et al., 2011). Seafloor features were not considered in this assessment. Figure 1 shows the location of the Arctic Ice „Ecologically or Biologically Significant Area“ (EBSA) proposed. Feature description The Ecologically or Biologically Significant Marine Area (EBSA) proposed focusses on the presently permanently icecovered waters in the OSPAR/NEAFC maritime areas, including the high seas section in the Central Arctic Basin north of the 200 nm zones of coastal states, and the marginal ice zone (where the ice breaks up, also called seasonal ice zone) along its southern margins (see Fig. 1 attached). Due to the inflow of Atlantic water along the shelf of Svalbard, and the concurrent outflow of polar water and ice on the Greenland side of Fram Strait, the southern limit of the summer sea ice extent is much further south in the western compared to the eastern Fram Strait, and in former times extended all along the Greenland coast. Several of the key ecosystem functions and species dependencies are associated with the ice front. Statements made about those functions and dependencies apply to the area where the front is located at any particular time of the year, and not necessarily to either areas fully ice covered (permanently or seasonally) or open waters distant from the ice front in summer. The high seas section of the OSPAR maritime area in the Central Arctic ocean is generally north of 84° N and much is fully ice-covered also in summer, although the quantity of multiyear ice has already substantially decreased and the 1year ice leaves increasingly large leads and open water spaces. The ice overlays a very deep water body of up to 5000 m depth distinct from the surrounding continental shelves and slopes of Greenland and the Svalbard archipelago. The Nansen-Gakkel Ridge, a prolongation of the Mid-Atlantic Ridge north of the Fram Strait is structuring the deep Arctic basin in this section, separating the Central Nansen Basin to the south from the Amundsen Basin to the north. Abundant hydrothermal vent sites have been discovered on this ridge at about 85° 38 N (Edmonds et al., 2003). North of Spitsbergen, the Atlantic water of the West Spitsbergen Current enters the Arctic basin as a surface current. At around 83° N, a deep-reaching frontal zone separates the incoming Atlantic and shelf waters from those of the Central Nansen Basin (Anderson et al., 1989), a transition reflected in ice properties, nutrient concentrations, zooplankton communities, and benthic assemblages (Hirche and Mumm, 1992, and literature quoted). This water subsequently submerges under the less dense (less salinity, lower temperature) polar water and circulates, in opposite direction to the surface waters and ice, counterclockwise along the continental rises until turning south along the Lomonossov Ridge and through Fram Strait as East Greenland Current south to Danmark Strait (Aagaard, 1989; Aagaard et al., 1985). Connecting the more fertile shelves with the deep central basin, these modified Atlantic waters supply the waters north of the NansenGakkel Ridge, in the Amundsen basin, with advected organic material and nutrients which supplement the autochtonous production (Mumm et al., 1998). Due to the import of organic biomass from the Greenland Sea and the Arctic continental shelves, part of which may not be kept in the food web due to the polar conditions, the Arctic Ocean may also represent an enormous carbon sink (Hirche and Mumm, 1992). In the Fram Strait, the region between Svalbard to the east and Greenland to the west, the East Greenland Current is the main outflow of polar water and ice from the Arctic Basin (Maykut, 1985) (Aagaard and Coachman, 1968). The polar front (0° C isotherm and 34.5 isohaline at 50 m depth) extends approximately along the continental shelf of Greenland, separating the polar surface water from the Arctic (Intermediate) water and the marginal ice zone to the east (e.g. Aagaard and Coachman, 1968; Paquette et al., 1985). The ice cover is densest in polar water, its extent to the east depends on the wind conditions (compare also Angelen et al., 2011; Wadhams, 1981). The seasonal latitudinal progression of increasing and diminishing light levels, respectively, is the determining factor for the timing of the phytoplankton-related pelagic production. Therefore, the spring bloom and ice break up progress from south to north in spring, reaching the Arctic area by about June/July. Because the currents in Fram Strait move in opposite direction, the polar East Greenland Current to the south, and the Atlantic West Spitsbergen Current to the north, there is a delay of about a month between biological spring and summer between the polar and the Atlantic side (Hirche et al., 1991). Therefore, sea ice and the effect of melting ice are important determinants of the ecosystem processes all along the East Greenland polar front from the Greenland Sea through Fram Strait to the Arctic Basin (Legendre et al., 1992; Wassmann, 2011). Ice situation The Arctic Ocean is changing towards a one-year instead of a multi-year sea-ice system with consequences for the entire ecosystem, including ecosystem shifts, biodiversity changes, water mass modifications, and role in the global overturning circulation. At its maximum, sea-ice covers 4.47 million km² in the Arctic Basin (Gill et al., 2011): According to data 180 ICES Advice 2013, Book 1 from ice satellite observations in 1973-76 (NASA, 1987, in (Gill et al., 2011)), permanent ice occupied 70-80% of the Arctic Basin area, and the inter-annual variability of this area did not exceed 2%. Seasonal ice occupied 6-17% (before the melting period of the mid-1970s). By the end of the first decade of the 21st century, the permanent-ice area had decreased greatly, concurrent with a rapid increase in seasonal- ice. Whereas multiyear ice used to cover 50-60% of the Arctic, it covered less than 30% in 2008, after a minimum of 10% in 2007. The average age of the remaining multiyear ice is also decreasing from over 20 % being at least six years in the mid- to late 1980s, to just 6% of ice six years old or older in 2008. Figure 2: Modelled ice age distribution in 1985-2000 (left) compared to February 2008 (right) (CAFF, 2010). This trend is likely to amplify in the coming years, as the net ocean-atmosphere heat output due to the current anomalously low sea ice coverage has approximately tripled compared to previous years, suggesting that the present sea ice losses have already initiated a positive feedback loop with increasing surface air temperatures in the Arctic (Kurtz et al., 2011). About 10% of the sea ice in the Arctic basin is exported each year through Fram Strait into the Greenland Sea (Maykut, 1985) which is therefore major sink for Arctic sea ice (Kwok, 2009). From 2001 to 2005, the summer ice cover was so low on the East Greenland shelf, that it was more of a marginal ice zone (Smith Jr and Barber, 2007). However the subsequent record lows in overall Arctic ice cover brought about an increase in ice cover off Greenland, which minimised the extent of the North East Water Polynia on the East Greenland shelf13, a previously seasonally ice-free stretch of water (Wadhams, 1981). Ice related biota An inventory of ice-associated biota covering the entire Arctic presently counts over 1000 protists, and more than 50 metazoan species (Bluhm et al., 2011). The regionally variable ice fauna (dependent on, inter alia, ice age, thickness, origin) consists of sympagic biota living within the caverns and brine channels of the ice, and associated pelagic fauna. The most abundant and diverse sympagic groups of the ice mesofauna in the Arctic seas are amphipods and copepods. Polar cod (Boreogadus saida) and to a lesser extent Arctic cod (Arctogadus glacialis) are dependent on the sympagic macro- and mesofauna for food. The fish themselves are important food sources for Arctic seals (such as ringed seal Phoca hispida) and birds, for example black guillemots Cephus grylle (Bradstreet and Cross, 1982; Gradinger and Bluhm, 2004 and literature reviewed; Horner et al., 1992; Süfke et al., 1998). The higher the light level in the ice, the higher is the biomass of benthic algae as well as meiofauna and microorganisms within the ice (Gradinger et al., 1991). Decreasing snow cover induces a feedback loop with enhanced algal biomass increasing the heat absorption of the ice which leads to changes in the ice structure, and ultimately the release of algae from the bottom layer (Apollonio, 1961 in Gradinger et al., 1991). Because of the distance to land and shelves, and the thickness and internal structure of the multiyear pack ice over deeper water, this type of ice has a fauna of its own (Carey, 1985; Gradinger et al., 1991). Arctic multiyear ice floes can have very high algal biomasses in the brine channels and in the bottom centimeters which serves as food for a variety of proto- and metazoans, usually smaller than 1 mm, over deep water (Gradinger et al., 1999). In the central Arctic, ice algal productivity can contribute up to 50 % of the total primary productivity, with lower contributions in the sea ice covered margins (Bluhm et al., 2011). In the boundary layer between ice floes and the water column, another specific community exists which forms the link between the ice based primary production and the pelagic fauna (Gradinger, 1995). Large visible bands of diatoms hang down from the ice, and are exploited by amphipods such as Gammarus wilkitzki, and occasionally by water column copepods such as Calanus glacialis, which are important prey of for example polar cod Boreogadus saida. The caverns, wedges and irregularities of the ice provide important shelter from predators for larger ice associated species and provide an essential habitat for these species (Gradinger and Bluhm, 2004). During melt, the entire sympagic ice biota are released into the water column where they may initiate the spring algal plankton bloom (Smith and Sakshaug, 1990) or they may sink to the sea floor and serve as an episodic and first food pulse for benthic organisms before pelagic production begins (Arndt and Pavlova, 2005). In particular the shallow shelves and the shelf slope benthos has been shown to profit of this biomass input, reflected in very rich benthic communities (Klitgaard and Tendal, 2004; Piepenburg, 2005). The role of the polar front and marginal ice zone for the production system Primary production in the Arctic Ocean is primarily determined by light availability, which is a function of ice thickness, ice cover, snow cover, light attenuation), the abundance of both ice algae and phytoplankton, nutrient availability and surface water stratification. Generally, the spring bloom occurs later further north and in regions with a thick ice and snow 13 http://www.issibern.ch/teams/Polynya/ ICES Advice 2013, Book 1 181 cover. The current production period in the Arctic Ocean may extend to 120 days per year, with a total annual primary production in the central Arctic Ocean of probably up to 10 g C m-2 (Wheeler et al., 1996). Ice algae start primary production when light levels are relatively low, as melting reduces the thickness of the ice and snow cover. The major phytoplankton bloom develops only after the ice breaks up, when melting releases the ice biota into the water column and meltwater leads to surface stratification. The bloom lasts a few weeks, fuelling the higher trophic food web of the Arctic (Gradinger et al., 1999, and literature quoted). The marginal ice zones, i.e. where the ice gets broken up in warmer Atlantic or Arctic water, play an important role in the overall production patterns of the Arctic Ocean. Due to the strong water column stratification and increased light levels involved with the melting of the ice, the location and recession of the ice edge in spring and summer determines the timing and magnitude of the spring phytoplankton bloom, which is generally earlier than in the open water (Gradinger and Baumann, 1991; Smith Jr. et al., 1987). Wind- or eddy-induced upwelling in the marginal ice zone, as well as biological regeneration processes replenish the surface nutrient pool and therefore prolong the algal growth period (Gradinger and Baumann, 1991; Smith, 1987). The hydrographic variability explains the patchy patterns of primary and secondary production observed, as well as consequently the patchy occurrence of predators. The polar front separates to some degree the pelagic faunas of the polar and Arctic waters in the Greenland Sea and Fram Strait, each characterised by a few dominant copepod species with different life history strategies (Hirche et al., 1991; see also review in Melle et al., 2005): In polar waters, Calanus glacialis uses under ice plankton production and lipid reserves for initiating its spring reproduction phase, but depends on the phytoplankton bloom for raising its offspring (e.g. Leu et al., 2011). Somewhat later, on the warm side of the polar front in Arctic water, the Atlantic species Calanus finmarchicus uses the ice edge-related phytoplankton bloom for secondary production. Calanus hyperboreus, the third and largest of the charismatic copepod species has its core area of distribution in the Arctic waters of the Greenland Sea (Hirche, 1997; Hirche et al., 2006). Zooplankton of the Arctic Basin Overall zooplankton biomass decreases towards the central Arctic basin, reaching a minimum in the most northerly waters, i.e. the region with permanent ice cover (Mumm et al., 1998). However, investigations in recent years demonstrated increased biomasses compared to studies several decades earlier - possibly a consequence of the decrease in ice thickness and cover which only enabled the investigations to take place from ship board. There is a south-north decrease in zooplankton biomass, with a sharp decline north of 83°N (Hirche and Mumm, 1992), coinciding with differences in the species composition of the biomass-forming zooplankton species. Whereas the southern Nansen basin plankton is dominated by the Atlantic species Calanus finmarchicus, entering the Arctic Basin with the West Spitsbergen Current, the northernmost branch of the North Atlantic current, the Arctic and polar species Calanus hyperboreus and C. glacialis dominate the biomass in the high-Arctic Amundsen and Makarov Basins (Auel and Hagen, 2002; Mumm et al., 1998). The zooplankton species communities generally can be differentiated according to their occurrence in Polar Surface Water (0-50 m, temperature below –1.7°C, salinity less than 33.0), Atlantic Layer (200–900 m; temperature 0.5–1.5°C); salinity 34.5–34.8) and Arctic Deep Water (deeper than 1000 m, temperature -0,5--1° C, salinity > 34.9) (Auel and Hagen, 2002; Grainger, 1989; Kosobokova, 1982). The polar surface community in the upper 50 m of the water column consists of original polar species as well as species emerging from deeper Atlantic waters, altogether leading to a high abundance and biomass peak in summer. Diversity and biomass are minimal in the impoverished Arctic basin deepwater community (Kosobokova 1982). Apart from a limited exchange with the Atlantic Ocean via the Fram Strait, the central Arctic deep-sea basins are isolated from the rest of the world ocean deepsea fauna. Therefore, the bathypelagic fauna consists of a few endemic Arctic species and some species of Atlantic origin. Due to the separation of the Eurasian and Canadian Basins by the Lomonosov Ridge, significant differences in hydrographic parameters (Anderson et al. 1994) and in the zooplankton composition occur between both basins (Auel and Hagen, 2002). Fish Polar cod, Boreogadus saida, is a keystone species in the ice-related foodwebs of the Arctic. Due to schooling behavior and high energy content polar cod efficiently transfer the energy from lower to higher trophic levels, such as seabirds, seals and some whales (Crawford and Jorgenson, 1993). Seabirds Ice cover is a physical feature of major importance to marine birds in high latitude oceans, providing access to resources, and refuge from aquatic predators (Hunt, 1990). As seabirds are dependent on leads between ice floes or otherwise open water to access food, they search for the most productive waters in polynias (places within the ice which are permanently 182 ICES Advice 2013, Book 1 ice free) and marginal ice zones (Hunt, 1990). Here they forage both on the pelagic and sympagic ice-related fauna, especially the early stages of polar cod and the copepods Calanus hyperboreus and C. glacialis. Likely, they benefit from the structural complexity and good visibility of their prey near the ice (Hunt, 1990). In the Greenland Sea and Fram Strait, major breeding colonies exist on Svalbard, Greenland and on Jan Mayen, all of these within reach of the seasonally moving marginal ice zone or a polynia (North East Water Polynia on the East Greenland shelf). Breeding seabirds like Little auks (Alle alle), from colonies in the northern Svalbard archipelago feed their offspring with prey caught in the vicinity of the nests, however intermittently travel at least 100 km to the marginal ice zone at 80° N to replenish their body reserves (Jakubas et al., Online 03 June 2011). Therefore, the distance of the marginal ice zone to the colony site is a critical factor determining the breeding success (e.g. Joiris and Falck, 2011). Opportunistically, the birds also use other zooplankton aggregations such as a in a cold core eddy in the Greenland Sea, closer to the nesting site (Joiris and Falck, 2011). A synopsis of seabird data for the period 1974–1993 (Joiris, 2000) showed that the little auk is one of the most abundant species, together with the fulmar Fulmarus glacialis, kittiwake Rissa tridactyla and Brünnich’s guillemot Uria lomvia in the European Arctic seas (mainly the Norwegian and Greenland Seas). In the Greenland Sea and the Fram Strait, little auks represented the main species in polar waters, at the ice edge and in closed pack ice, reaching more than 50% of all bird species (Joiris and Falck, 2011). In spring and autumn, millions of seabirds pass through the area when migrating between their breeding sites on Svalbard or the Russian Arctic and their wintering areas in Canada (Gill et al., 2011). There are several seabird species in the European Arctic which are only met in ice-covered areas, for example the Ivory gull Pagophila eburnea and the Thick-billed guillemot Uria lomvia (see e.g. CAFF, 2010): Both species spend the entire year in the Arctic, and breed in close proximity to sea ice although Thick-billed guillemots were observed to fly up to 100 km from their colonies over open water to forage at the ice edge (Bradstreet 1979). The relatively rare Ivory gulls are closely associated with pack-ice, favouring areas with 70 – 90% ice cover near the ice edge, where they feed on small fish, including juvenile Arctic cod, squid, invertebrates, macro-zooplankton, carrion, offal and animal faeces (e.g. OSPAR Commission, 2009b). Ivory gulls have a low reproductive rate and breeding only takes place if there is sufficient food, which makes the population highly vulnerable to the effects of climate warming (e.g. OSPAR Commission, 2009b). Thick-billed guillemots are relatively long lived and slow to reproduce and has a low resilience to threats including oil pollution, by-catch in and competition with commercial fisheries operations, population declines due to hunting – particularly in Greenland (OSPAR Commission, 2009c). Ivory gull and Thick-billed guillemots are both listed by OSPAR as being under threat and/or decline, (OSPAR Commission, 2008) and in 2011 recommendations for conservation action were agreed (OSPAR Commission, 2011) which will be implemented in conjunction with the circumpolar conservation actions of CAFF (CAFF, 1996; Gilchrist et al., 2008). Marine mammals Several marine mammal species permanently associate with sea ice in the European Arctic. These include polar bear, walrus, and several seal species: bearded, Erignathus barbatus; ringed, Pusa hispida; hooded, Cystophora cristata; and harp seal Pagophilus groenlandicus. Three whale species also occupy Arctic waters year- round – narwhal, Monodon monoceros; beluga whale, Delphinapterus leucas; and bowhead whale, Balaena mysticetus. Polar bears Ursus maritimus are highly specialized for and dependent on the sea ice habitat and are therefore particularly vulnerable to changes in sea ice extent, duration and thickness. They have a circumpolar distribution limited by the southern extent of sea ice. Three subpopulations of polar bears occur in the European high Arctic: the East Greenland, Barents Sea and Arctic Basin sub-populations, all with an unknown population status (CAFF, 2010). Following the youngof-the-year ringed seal distribution, polar bears are most common close to land and over the shelves, however some also occur in the permanent multi-year pack ice of the central Arctic basin (Durner et al., 2009). Due to low reproductive rates and long lifetime, it has been predicted that the polar bears will not be able to adapt to the current fast warming of the Arctic and become extirpated from most of their range within the next 100 years (Schliebe et al., 2008). Walruses, Odobenus rosmarus, inhabit the Arctic ice year-round. They are conservative benthic feeders, diving to 80100 m depth for scaping off the rich mollusc fauna of the continental shelves, and need ice floes as resting and nursing platform close to their foraging grounds. Walruses have been subject to severe hunting pressure from the end of the 18th century to the mid-20th century, and are still hunted today in Greenland (NAMMCO). By 1934, the estimated 7000080000 individuals of the Atlantic population were reduced to 1200-1300, with none left on Svalbard (Weslawski et al., 2000). Today’s relatively small sub-populations on the East Greenland and Svalbard-Franz Josef Land coasts have recently shown a slightly increasing trend, in the latter case reflecting the full protection of the species since the 1950´s (CAFF, 2010; NAMMCO). Apart from their sensitivity to direct human disturbance and pollution, it is expected that ICES Advice 2013, Book 1 183 walruses will suffer from the changing ice conditions (location, thickness for being used as haul-out site) as well as changes in ice-related productivity. The Atlantic subspecies of the bearded seal, Erignathus barbatus occurs south of 85° N from the central Canadian Arctic east to the central Eurasian Arctic, but no population estimates exist (Kovacs, 2008b). Because of their primarily benthic feeding habits they live in ice covered waters overlying the continental shelf. They are typically found in regions of broken free-floating pack ice; in these areas bearded seals prefer to use small and medium sized floes, where they haul out no more than a body length from water and they use leads within shore-fast ice only if suitable pack ice is not available (Kovacs, 2008b, and literature quoted). The Arctic ringed seal Pusa (Phoca) hispida hispida has a very large population size and broad distribution, however, there are concerns that future changes of Arctic sea ice will have a negative impact on the population, some of which have already been documented in some parts of the subspecies range (Kovacs et al., 2008). As the other seals, the ringed seal uses sea ice exclusively as their breeding, moulting and resting (haulout) habitat, and feed on small schooling fish and invertebrates. In a co-evolution with one of their main predators, the polar bear, they developed the ability to create and maintain breathing holes in relatively thick ice, which makes them well adapted to living in fully ice covered waters the year round. The West Ice (or Is Odden) to the west of Jan Mayen, at approx. 72-73° N, in early spring a stretch of more of less fast drift ice, is of crucial importance as a whelping and moulting area for harp seals and hooded seals (summarised e.g. by ICES, 2008). Discovered in the early 18th century, up to 350000 seals (1920s) were killed per year, decimating the populations from an estimated one million individuals in the 1950s (Ronald et al., 1982) to today´s 70000 and 243000 of hooded and harp seals, respectively (Kovacs, 2008a, c). Hooded seal, Cystophora cristata, is a pack ice species, which is dependent on ice as a substrate for pupping, moulting, and resting and as such is vulnerable to reduction in extent or timing of pack ice formation and retreat, as well as ice edge related changes in productivity (Kovacs, 2008a, and literature quoted). Hooded Seals feed on a wide variety of fish and invertebrates, including species that occur throughout the water column. After breeding and moulting on the West Ice they follow the retreating pack ice to the north, but also spend significant periods of time pelagically, without hauling out (Folkow and Blix 1999) in (Kovacs, 2008a). The northeast Atlantic breeding stock has declined by 85-90 % over the last 40-60 years. The cause of the decline is unknown, but very recent data suggests that it is on-going (30% within 8 years), despite the protective measures that have been taken in the last few years. The species is therefore considered to be vulnerable (Kovacs, 2008a). Harp seals Pagophilus (Phoca) groenlandicus are the most numerous seal species in the Arctic seas. Their reproduction takes place in huge colonies, for example on the pack ice of the ‘‘West Ice’’ north of Jan Mayen, and after the breeding season they follow the retreating pack ice edge northwards up to 85° N, feeding mainly on polar cod under the ice (Kovacs, 2008c) . Narwhals Monodon monoceros primarily inhabit the ice-covered waters of the European Arctic, including the ice sheet off East Greenland (Jefferson et al., 2008b). For two months in summer, they visit the shallow fjords of East Greenland, spending all the rest of the year offshore, in deep ice-covered waters along the continental slope in the Greenland Sea and Arctic Basin (Heide-Jørgensen and Dietz, 1995). Narwhals are deep diving benthic feeders and forage on fish, squid, and shrimp, especially Arctic fish species, such as Greenland halibut, Arctic cod, and polar cod at up to 1500 m depth and mostly in winter. A recent assessment of the sensitivity of all Arctic marine mammals to climate change ranked the narwhal as one of the three most sensitive species, primarily due to its narrow geographic distribution, specialized feeding and habitat choice, and high site fidelity (Laidre et al. 2008 in (Jefferson et al., 2008b)). Bowhead whales Balaena mysticetus are found only in Arctic and subarctic regions and a Svalbard-Barents population occurs from the coast of Greenland across the Greenland Sea to the Russian Arctic. They spend all of their lives in and near openings in the pack ice feeding on small to medium-sized zooplankton. They migrate to the high Arctic in summer, and retreat southward in winter with the advancing ice edge (Moore and Reeves 1993 in (Reilly et al., 2008)). Whaling has decimated the original bowhead whale populations to be rare nowadays, listed by OSPAR as being under threat and/or decline (OSPAR Commission, 2008). The species is considered to be very sensitive to changes in the ice-related ecosystem as well as sound disturbance, possible consequences of a progressive reduction of ice cover (OSPAR Commission, 2009a). Belugas Delphinapterus leucas prefer coastal and continental shelf waters with a broken-up ice cover. They have never been surveyed around Svalbard. Pods numbering into the thousands are sighted irregularly around the archipelago, and pods ranging from a few to a few hundred individuals are seen regularly (Gjertz and Wiig 1994; Kovacs and Lydersen 2006 in (Jefferson et al., 2008a)). 184 ICES Advice 2013, Book 1 Little is known about the populations of the larger fauna in the Central Arctic Basin over the deepsea basins and ridges. But it is not likely that it is currently an area of great abundance - too far from the coastal nesting sites of marine birds, and over too deep water to allow feeding on benthos, as most of the larger mammals would need, and currently of too low plankton production to feed the large whales. All of these groups have their distribution center along the continental shelves presently - however, following the receding ice edge out to the central Arctic basin may be one of the options for the future. Feature condition, and future outlook This high Arctic region is particularly vulnerable to the the loss of ice cover and other effects of the anticipated global warming, including elevated UV radiation levels (Agustí, 2008). (Wassmann et al., 2010) summarise what changes may be expected within the subarctic/Arctic region: • • • • • northward displacement (range shifts) of subarctic and temperate species, and cross-Arctic transport of organisms; increased abundance and reproductive output of subarctic species, decline and reduced reproductive success of some Arctic species associated with the ice and species now preyed upon by predators whose preferred prey have declined; increased growth of some subarctic species and primary producers, and reduced growth and condition of animals that are bound to, associated with, or born on the ice; anomalous behaviour of ice-bound, ice-associated, or ice-born animals with earlier spring events and delayed fall events; changes in community structure due to range shifts of predators resulting in changes in the predator–prey linkages in the trophic network. (Wassmann, 2008) expects radical changes in the productivity, functional relationships and biodiversity of the Arctic Ocean. He suggests that a warmer climate with less ice cover will result in greater primary production, a reduction of the stratified water masses to the south, changes in the relationship between biological processes in the water column and the sediments, a reduction in niches for higher trophic levels and a displacement of Arctic by boreal species. On the shelves, increased sediment discharges are expected to lower the primary production due to higher turbidity, and enhance the organic input to the deep ocean. A more extensive review of expected or suspected consequences of climate change for the marine system of the Arctic is given in (Loeng et al., 2005). Figure 3, extracted from (Gill et al., 2011), presents the conceptual ideas about possible Arctic ecosystem changes mediated by human impact: The normal situation shown in the upper left panel consists of ice-dependent species and species that tolerate a broader range of temperatures and are found in waters with little or no sea ice. Primary production occurs in phytoplankton (small dots in the figure) in ice-free waters and in ice-attached algae and phytoplankton in ice-covered waters. Phytoplankton (small t-shaped symbols in the figure) and ice algae are the main food sources for zooplankton and benthic animals. The fish community consists of both pelagic and demersal species. Several mammals are ice-associated, including polar bears and several species of seals. A number of sea bird species are also primarily associated with ice-covered waters. At moderate temperature increases (upper right) populations of ice-dependent species are expected to decline as sea ice declines, and sub-Arctic species are expected to move northwards. Arctic benthic species are expected to decline, especially if their distributions are pushed close to or beyond the continental slope. The expected effects from fisheries relate to the continental shelves. Two major effects are changes in populations of benthic organisms due to disturbance from bottom trawling and removal of large individuals in targeted fish stocks. In addition, the size of targeted stocks, both demersal and pelagic, may be reduced. In addition, the effects of ocean acidification are considered (lower right). Ocean acidification will result in depletion of carbonate phases such as aragonite and calcite. This will alter the structure and function of calcareous organisms, particularly at lower trophic levels. Changes in pH can also alter metabolic processes in a range of organisms. It is not known how these changes will propagate to higher trophic levels, but the effects could be substantial. Figure 3: Conceptual models showing potential impacts on Arctic marine ecosystems under different scenarios (Gill et al., 2011). Gill et al. (2011) conclude that the central part of the Arctic Basin is not a region for fisheries or oil and gas exploration. However, this region has played and will continue to play a very important role in the redistribution of pollutants, due to ice drift and/or currents between coastal and shelf areas and the Arctic Basin peripheries, far from sources of pollution. ICES Advice 2013, Book 1 185 Assessment against CBD EBSA Criteria Table 1 CBD Criterion Relation of each of the CBD criteria to the proposed area relating to the best available science. Note that a candidate EBSA may qualify on the basis of one or more of the criteria, the boundaries of the EBSA need not be defined with exact precision. EBSA Description Uniqueness or rarity The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High x Explanation for ranking Arctic sea ice, in particular the multiyear ice of the Central Arctic is globally unique and hosts endemic species such as the Gammarid amphipod Gammarus wilkitzki and sea ice meiofauna which will disappear with the melting of the ice. Polar bears, walrusses, bowhead whales, narwhales, belugas, several seal species and many bird species are endemic to the high Arctic ice. While sea ice species such as G. wilkitzki are not endemic to the proposed EBSA they are endemic to the Arctic and unique within the OSPAR area Special importance Areas that are required for a population to survive and thrive for life-history stages of species x Explanation for ranking Sea ice is essential for its sympagic fauna, and to some extent also for the pelagic associated fauna which also depends on the right timing of biomass production (match/mismatch with bloom periods). The marginal ice zone and other openings in the ice are essential feeding grounds for a large number of ice-associated species which exploit the seasonally high production there. At present the area covered by the proposed EBSA includes both the area of permanent ice and, the area covered by seasonal ice and the ice edge. The community associated with the ice edge requires it special structural features for a number of ecological processes, including increased primary and secondary productivity, and feeding and resting of seabirds and marine mammals. Area containing habitat for the survival and recovery of x Importance for endangered, threatened, declining species or area with threatened, significant assemblages of such species endangered or declining species and/or habitats Explanation for ranking The high arctic ice hosts endemic species such as the Gammarid amphipod Gammarus wilkitzki and sea ice meiofauna which will disappear with the melting of the ice. Many of the obligatory ice-related species are listed as vulnerable by IUCN, and/or listed as under threat and/or decline by OSPAR, examples include the Ivory gull, thick-billed guillemot, bowhead whale, hooded seal and polar bear. With the overall trend of retreating sea ice extent, the proposed EBSA may become increasingly important for all ice-dependent species in the future. Areas that contain a relatively high proportion of sensitive Vulnerability, habitats, biotopes or species that are functionally fragile fragility, sensitivity, or slow (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery recovery 186 x ICES Advice 2013, Book 1 Explanation for ranking The ice-related foodweb and ecosystem is highly sensitive to the ecological consequences of a warming climate. Beyond this the Arctic is at the forefront of the impacts of ocean acidification (Wicks & Roberts 2012). The largest changes in ocean pH will occur in the Arctic Ocean, with complete undersaturation of the Arctic Ocean water column predicted before the end of this century (Steinacher et al. 2009). Many of the seabird and mammal populations are particularly sensitive to changes due to their already low population numbers, and low fertility. If the retreat of the ice to the north will lead to increased shipping and oil and gas exploitation in Arctic waters, the increased risk of spills would also pose a potential hazard for example for guillemots, which are extremely susceptible to mortality from oil pollution (CAFF, 2010). In addition, some species like Ivory gull are sensitive to an increased heavy metal load in their prey. Biological productivity Area containing species, populations or communities with comparatively higher natural biological productivity Explanation for ranking This criterion was not evaluated in the OSPAR/NEAFC/CBD Workshop. ICES did not have enough information to evaluate this criterion. Biological diversity Area contains comparatively higher diversity of ecosystems, habitats, communities, or species, or has higher genetic diversity Explanation for ranking This criterion was not evaluated in the OSPAR/NEAFC/CBD Workshop. ICES did not have enough information to evaluate this criterion. References Aagaard, K., 1989. A synthesis of Arctic Ocean circulation. Rapport Proces et Verbeaux Réunion du Conseil international pour l'Exploration de la Mer 188, 11-22. Aagaard, K., Coachman, L.K., 1968. The East Greenland Current north of Denmark Strait: Part II. Arctic 21, 267-290. Aagaard, K., Swift, J.H., Carmack, E.C., 1985. Thermohaline circulation in the Arctic mediterranean seas. Journal of Geophysical Research 90, 4833-4846. Agustí, S., 2008. Impacts of increasing ultraviolet radiation on the polar oceans. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.) Fundación BBVA pp. 25-46. Anderson, L.G., Jones, E.P., Koltermann, K.P., Schlosser, P., Swift, J.H., Wallace, D.W.R., 1989. The first oceanographic section across the Nansen Basin in the Arctic Ocean. Deep Sea Research 36, 475-482. Angelen, J.H.v., Broeke, M.R.v.d., Kwok, R., 2011. The Greenland Sea Jet: A mechanism for wind‐driven sea ice export through Fram Strait. Geophysical Research Letters 38 (L12805). Anisimov, O.A., Vaughan, D.G., Callaghan, T.V., Furgal, C., Marchant, H., Prowse, T.D., Vilhjálmsson, H., Walsh, J.E., 2007. Polar regions (Arctic and Antarctic). In: Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Parry, M.L., Canziani, O.F., Palutikof, J.P., Linden, P.J.v.d., Hanson, C.E. (Eds.)Cambridge University Press, Cambridge pp. 653-685. Arndt, C., E. , Pavlova, O., 2005. Origin and fate of ice fauna in the Fram Strait and Svalbard area. Marine Ecology Progress Series 301, 55-66. Auel, H., Hagen, W., 2002. Mesozooplankton community structure, abundance and biomass in the central Arctic Ocean. Marine Biology 140, 1013-1021. Bluhm, B.A., Gebruk, A.V., Gradinger, R., Hopcroft, R.R., Huettmann, F., Kosobokova, K.N., Sirenko, B.I., Weslawski, J.M., 2011. Arctic marine biodiversity: An update of species richness and examples of biodiversity change. Oceanography 24 (3), 232-248. Bradstreet, M.S.M., Cross, W.E., 1982. Trophic relationships at high Arctic ice edges. Arctic 35 (1), 1-12. CAFF, 1996. International Murre conservation strategy and action plan. CAFF International Secretariat, CAFF Circumpolar Seabird Working Group, Akureyri, Iceland, pp. 1-16. CAFF, 2010. Arctic Biodiversity Trends 2010. Selected indicators of change. CAFF International Secretariat, , Akureyri, Iceland. Carey, A.G.I., 1985. Marine Ice Fauna. In: Arctic Sea Ice Biota. A., H.R. (Ed.)CRC Press, Boca Raton. Florida pp. 17190 Crawford, R.E., Jorgenson, J.K., 1993. Schooling behaviour of arctic cod, Boreogadus saida in relation to drifting pack ice. Environmental Biology of Fishes 36 (4), 345-357. Durner, G.M., Douglas, D.C., Nielson, R.M., Amstrup, S.C., McDonald, T.L., Stirling, I., Mauritzen, M., Born, E.W., Wiig, Ø., Deweaver, E., Serreze, M.C., Belikov, S.E., Holland, M.M., Maslanik, J., Aars, J., Bailey, D.A., Derocher, A.E., 2009. Predicting 21st-century polar bear habitat distribution from global climate models. Ecological Monographs 79 (1), 25-58. ICES Advice 2013, Book 1 187 Edmonds, H.N., Michael, P.J., Baker, E.T., Connelly, D.P., Snow, J.E., Langmuir, C.H., Dick, H.J.B., Mühe, R., German, C.R., Graham, D.W., 2003. Discovery of abundant hydrothermal venting on the ultraslow-spreading Gakkel ridge in the Arctic Ocean. Nature 421, 252-256. Gilchrist, G., Strøm, H., Gavrilo, M.V., Mosbech, A., 2008. International Ivory Gull conservation strategy and action plan. CAFF International Secretariat, Circumpolar Seabird Group (CBird). CAFF Technical Report No. 18. Gill, M.J., Crane, K., Hindrum, R., Arneberg, P., Bysveen, I., Denisenko, N.V., Gofman, V., Grant-Friedman, A., Gudmundsson, G., Hopcroft, R.R., Iken, K., Labansen, A., Liubina, O.S., Melnikov, I.A., Moore, S.E., Reist, J.D., Sirenko, B.I., Stow, J., Ugarte, F., Vongraven, D., Watkins, J., 2011. Arctic Marine Biodiversity Monitoring Plan (CBMP-MARINE PLAN), CAFF Monitoring Series Report No.3, April 2011. CAFF International Secretariat,, Akureyri, Iceland. Gradinger, R., 1995. Climate change and biological oceanography of the Arctic Ocean. Phil. Trans. R. Soc. A 352, 277286. Gradinger, R., Bluhm, B.A., 2004. In situ observations on the distribution and behavior of amphipods and Arctic cod (Boreogadus saida) under the sea ice of the high Arctic Canadian Basin. Polar Biology 27, 595-603. Gradinger, R., Friedrich, C., Spindler, M., 1999. Abundance, biomass and composition of the sea ice biota of the Greenland Sea pack ice. Deep Sea Research 46, 1457-1472. Gradinger, R., Spindler, M., Henschel, D., 1991. Development o Arctic sea-ice organisms under graded snow cover. In: Proceedings of the Pro Mare Symposium on Polar Marine Ecology. Sakshaug, E., E., H.C.C., Øritsland, N.A. (Eds.), Polar Research 10 (1), Trondheim pp. 295-307. Gradinger, R.R., Baumann, M.E.M., 1991. Distribution of phytoplankton communities in relation to the large-scale hydrographical regime in the Fram Strait. Mar. Biol. 111, 311-321. Grainger, E.H., 1989. Vertical distribution of zooplankton in the central Arctic Ocean. In: Proc 6th Conf Comite´Arctique Int 1985. Rey, L., Alexander, V. (Eds.) Brill Leiden pp. 48–60. Heide-Jørgensen, M.P., Dietz, R., 1995. Some characteristics of narwhal, Monodon monoceros, diving behaviour in Baffin Bay. Canadian Journal of Zoology 73, 2106-2119. Hirche, H.J., 1997. Life cycle of the copepod Calanus hyperboreus in the Greenland Sea. Marine Biology 128 (4), 607618. Hirche, H.J., Baumann, M.E.M., Kattner, G., Gradinger, R., 1991. Plankton distribution and the impact of copepod grazing on primary production in Fram Strait, Greenland Sea. Journal of Marine Systems 2 (3-4), 477-494. Hirche, H.J., Mumm, N., 1992. Distribution of dominant copepods in the Nansen Basin, Arctic Ocean, in summer. Deep Sea Research Part A. Oceanographic Research Papers 39 (2, Part 1), S485-S505. Hirche, H.J., Muyakshin, S., Klages, M., Auel, H., 2006. Aggregation of the Arctic copepod Calanus hyperboreus over the ocean floor of the Greenland Sea. Deep Sea Research Part I: Oceanographic Research Papers 53 (2), 310-320. Horner, R., Ackley, S.F., Dieckmann, G.S., Gulliksen, B., Hoshiai, T., Legendre, L., Melnikov, I.A., Reeburgh, W.S., Spindler, M., Sullivan, C.W., 1992. Ecology of sea ice biota. Habitat, terminology, and methodology. Polar Biology 12 (3), 417-427. Hunt, G.L.J., 1990. The pelagic distribution of marine birds in a heterogeneous environment. Polar Research 8, 43-54. ICES, 2008. Report of the ICES Advisory Committee In: ICES Advice, Book 3, The Barents and the Norwegian SEa. Jakubas, D., Iliszko, L., Wojczulanis-Jakubas, K., Stempniewicz, L., Online 03 June 2011. Foraging by little auks in the distant marginal sea ice zone during the chick-rearing period. Polar Biology, 1-9. Jefferson, T.A., Karczmarski, L., Laidre, K., O’Corry-Crowe, G., Reeves, R.R., Rojas-Bracho, L., Secchi, E.R., Slooten, E., Smith, B.D., Wang, J.Y., Zhou, K., 2008a. Delphinapterus leucas In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Jefferson, T.A., Karczmarski, L., Laidre, K., O’Corry-Crowe, G., Reeves, R.R., Rojas-Bracho, L., Secchi, E.R., Slooten, E., Smith, B.D., Wang, J.Y., Zhou, K., 2008b. Monodon monoceros. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Joiris, C., Falck, E., 2011. Summer at-sea distribution of little auks Alle alle and harp seals Pagophilus (Phoca) groenlandica in the Fram Strait and the Greenland Sea: impact of small-scale hydrological events. Polar Biology 34 (4), 541-548. Joiris, C.R., 2000. Summer at-sea distribution of seabirds and marine mammals in polar ecosystems: a comparison between the European Arctic seas and the Weddell Sea, Antarctica. Journal of Marine Systems 27, 267-276. Klitgaard, A.B., Tendal, O.S., 2004. Distribution and species composition of mass occurrences of large-sized sponges in the northeast Atlantic. Progress in Oceanography 61, 57-98. Kosobokova, K.N., 1982. Composition and distribution of the biomass of zooplankton in the central Arctic Basin. Oceanology 22, 744-750. Kovacs, K., 2008a. Cystophora cristata. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Kovacs, K., 2008b. Erignathus barbatus. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Kovacs, K., 2008c. Pagophilus groenlandicus. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. 188 ICES Advice 2013, Book 1 Kovacs, K., Lowry, L., Härkönen, T., 2008. Pusa hispida. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Kwok, R., 2009. Outflow of Arctic Ocean sea ice into the Greenland and Barents seas: 1979-2007. Journal of Climate 22, 2438-2456. Legendre, L., Ackley, S.F., Dieckmann, G.S., Gulliksen, B., Horner, R., Hoshiai, T., Melnikov, I.A., Reeburgh, W.S., Spindler, M., Sullivan, C.W., 1992. Ecology of sea ice biota. Polar Biology 12 (3), 429-444. Leu, E., Søreide, J.E., Hessen, D.O., Falk-Petersen, S., Berge, J., 2011. Consequences of changing sea ice cover for primary and secondary producers in the European Arctic shelf seas: timing, quantity, and quality. Progress in Oceanography 90, 18-32. Loeng, H., Brander, K., Carmack, E.C., Denisenko, S., Drinkwater, K., Hansen, B., Kovacs, K., Livingston, P., McLaughlin, F., Sakshaug, E., 2005. Marine systems. In: Arctic Climate Impact Assessment, ACIA. Symon, C., Arrisand, L., Heal, B. (Eds.),Cambridge University Press, Cambridge pp. 453-538. Mauritzen, C., Hansen, E., Andersson, M., Berx, B., Beszczynska-Möller, A., Burud, I., Christensen, K.H., Debernard, J., de Steur, L., Dodd, P., Gerland, S., Godøy, Ø., Hansen, B., Hudson, S., Høydalsvik, F., Ingvaldsen, R., Isachsen, P.E., Kasajima, Y., Koszalka, I., Kovacs, K.M., Køltzow, M., LaCasce, J., Lee, C.M., Lavergne, T., Lydersen, C., Nicolaus, M., Nilsen, F., Nøst, O.A., Orvik, K.A., Reigstad, M., Schyberg, H., Seuthe, L., Skagseth, Ø., Skar∂hamar, J., Skogseth, R., Sperrevik, A., Svensen, C., Søiland, H., Teigen, S.H., Tverberg, V., Wexels Riser, C., 2011. Closing the loop - Approaches to monitoring the state of the Arctic Mediterranean during the International Polar Year 2007-2008. Progress in Oceanography 90 (1-4), 62-89. Maykut, G.A., 1985. The ice environment. In: Sea-ice biota. Horner, R. (Ed.)CRC Press, Boca Raton pp. 21-82. Melle, W., Ellertsen, B., Skjoldal, H.R., 2005. Zooplankton: The link to higher trophic levels. In: The Nordic Seas: An integrated perspective oceanography, climatology, biogeochemistry, and modelling. . Drange, H., Dokken, T., Furevik, T., Gerdes, R., Berger, W. (Eds.)Geophysical Monograph Series 158 pp. 137-202. Mumm, N., Auel, H., Hanssen, H., Hagen, W., Richter, C., Hirche, H.J., 1998. Breaking the ice: large-scale distribution of mesozooplankton after a decade of Arctic and transpolar cruises. Polar Biology 20 (3), 189-197. NAMMCO, The Atlantic Walrus. North Atlantic Marine Mammal Commission. Status of Marine Mammals in the North Atlantic, Tromsø, pp. 1-7. OSPAR Commission, 2008. OSPAR List of Threatened and/or Declining Species andHabitats. Reference number 20086. http://www.ospar.org/documents/dbase/decrecs/agreements/0806e_ospar%20list%20species%20and%20habitats.doc. OSPAR Commission, 2009a. Background Document for Bowhead whale Balaena mysticetus. OSPAR Commission, Biodiversity Series 494/2010, pp. 1-20. OSPAR Commission, 2009b. Background Document for Ivory gull Pagophila eburnea. OSPAR Commission, Biodiversity Series 410/2009, pp. 1-16. OSPAR Commission, 2009c. Background Document for Thick-billed murre Uria lomvia. OSPAR Commission, Biodiversity Series 416/2009, pp. 1-20. OSPAR Commission, 2011. Meeting of the OSPAR Commission (OSPAR) London: 20-24 June 2011. Summary Record OSPAR 11/20/1-E. OSPAR Commission, London. Paquette, R., Bourke, R., Newton, J., Perdue, W., 1985. The East Greenland Polar Front in autumn. Journal of Geophysical Research 90 (C3), 4866-4882. Piepenburg, D., 2005. Recent research on Arctic benthos: common notions need to be revised. Polar Biology 28 (10), 733-755. Reilly, S.B., Bannister, J.L., Best, P.B., Brown, M., , Brownell Jr., R.L., Butterworth, D.S., Clapham, P.J., Cooke, J., Donovan, G.P., Urbán, J., Zerbini, A.N., 2008. Balaena mysticetus. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Renaud, P.E., Caroll, M.L., Ambrose, W.G.J., 2008. Effects of global warming on Arctic seafloor communities and its consequences for higher trophic levels. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.)Fundación BBVA pp. 141-177. Rey, F., 2004. Phytoplankton: the grass of the sea. In: The Norwegian Sea Ecosystem. Skjoldal, H.R. (Ed.)Tapir Academic Press, Trondheim, Norway pp. 97-136. Ronald, K., Healey, P.J., Fisher, H.D., 1982. The harp seal, Pagophilus groenlandicus. In: Small cetaceans, seals, sirenians and otters. FAO Fisheries Series No. 5, Vol. IV, Food and Agriculture Organisation of the United Nations. Workding Party on Mammals. Schliebe, S., Wiig, Ø., Derocher, A.E., Lunn, N., 2008. Ursus maritimus. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Smith Jr, W.O., Barber, D., 2007. Polynyas and climate change: a view to the future. In: Polynays, windows to the world. Halpern, D. (Ed.), Elsevier Oceanography Serie 74, Elsevier, Amsterdam pp. 411-420. Smith Jr, W.O., Baumann, M.E.M., Wilson, D.L., Aletsee, L., 1987. Phytoplankton biomass and productivity in the Marginal Ice Zone of the Fram Strait during summer 1984. Journal of Geophysical Research 92 (C7), 6777-6786. Smith, W.O.J., 1987. Phytoplankton dynamics in the marginal ice zones. Oceanography and Marine Biology Annual Review 25, 11-38. ICES Advice 2013, Book 1 189 Smith, W.O.J., Sakshaug, E., 1990. Polar phytoplankton. In: Polar oceanography. Part B. Chemistry, biology and geology. Smith Jr, W.O. (Ed.) Academic Press, San Diego pp. 477-525. Steinacher, M., Joos, F., Frölicher, T., Plattner, G. & Doney, S.c. 2009. Imminent ocean acidification in the Arctic projected with the NcAR global coupled carbon cycle-climate model. Biogeosciences 6, 515–533. Süfke, L., Piepenburg, D., Dorrien, C.C.v., 1998. Body size, sex ratio and diet composition of Arctogadus glacialis (Peters, 1874) (Pisces: Gadidae) in the Northeast Water Polynya (Greenland). Polar Biology 20, 357-363. Wadhams, P., 1981. The ice cover in the Greenland and Norwegian Seas. Rev. Geophys. Space Physics 19, 345-393. Wassmann, P., 2008. Impacts of global warming on Arctic pelagic ecosystems and processes. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.)Fundación BBVA pp. 113-148. Wassmann, P., 2011. Arctic marine ecosystems in an era of rapid climate change. Progress in Oceanography 90, 1-17. Wassmann, P., Duarte, C.M., Agustí, S., Seijr, M., 2010. Footprints of climate change in the Arctic Marine Ecosystem. Biological Global Change. Weslawski, J.M., Hacquebord, L., Stempniewicz, L., Malinga, M., 2000. Greenland whales and walruses in the Svalbard food web before and after exploitation. Oceanologia 42 (1), 37-56. Wheeler, P.A., Gosselin, M., Sherr, E., Thibault, D., Kirchman, D.L., Benner, R., Whitledge, T.E., 1996. Active cycling of organic carbon in the central Arctic Ocean. Nature 380, 697-699. Wicks, L., Roberts, J.M. (2012) Benthic invertebrates in a high CO2 world. Oceanography & Marine Biology: An Annual Review 50: 127 -188 190 ICES Advice 2013, Book 1 Maps and Figures Figure 1 Location of the ecologically or biologically significant areas (EBSA) proposed by WWF in September 2011. Figure 2 Modelled ice age distribution in 1985-2000 (left) compared to February 2008 (right) (CAFF, 2010). ICES Advice 2013, Book 1 191 Figure 3 192 Conceptual models showing potential impacts on Arctic marine ecosystems under different (Gill et al., 2011). scenarios ICES Advice 2013, Book 1 Annex 5 Note on the Sub-Polar Front The OSPAR-NEAFC-CBD workshop report proposed an EBSA that included both the Sub-Polar Front (SPF) and the Charlie-Gibbs Fracture Zone (CGFZ). Although that report did not describe the physical oceanographic processes causing the linkage, it presented the information about the area in a way that implied there was a structural linkage between the benthic-seafloor features of the CGFZ and the pelagic features of the SPF. The ICES Review Group accepted the arguments in the earlier report for linkage between the two features. It only proposed an anchoring of the bottom of the SPF to CGFZ, so the front moved seasonally north and south in the water column but did not sweep over the entire seafloor between its northernmost and southernmost boundaries. Following further study of the summary descriptions of the physical oceanography of the SPF since the ICES Review Group discussions, the secondary sources all indicate that there is little or no structural linkage between the pelagic feature of the SPF and the benthic feature of the CGFZ. Two illustrative quotes are given below, one from a NOAA – Univ of Florida website on north Atlantic Oceanography, and one from the Mar-Eco website. “It [North Atlantic Current that forms the Sub Polar front] is recognized as a shallow, widespread and variable winddriven surface movement of warm water that covers a large part of the eastern subpolar North Atlantic and slowly spills into the Nordic Seas. It is also sometimes included as the Subarctic or Subpolar Front as it is thought of as the boundary between the cold, subpolar region and the warm, subtropical gyre of the Northeastern Atlantic.”. (Rosensteil Institute/NOAA) “Near the CGFZ is also the near-surface frontal zone between cold water to the north and warm saline water to the south, known as the Sub-polar Front.” (MarEco) The two features have some co-incidence in two-dimensional maps of the ocean, but much less (if any) connection in three dimensions. Merging them as a single proposed EBSA weakens the science rationale for either feature as an EBSA. If we rank only the benthic CGFZ against the EBSA criteria, it scores highly considering only the geo-morphology, and the limited information available on the benthic community strengthens the case. The case for the CGFZ is considered strong enough to stand alone and can be advanced immediately. In relation to the pelagic SPF, there appear to be several weaknesses in the case at present. First, we do not have a clear description of the three-dimensional structure of the SPF. Neither the description in the 2011 report (a huge rectangular box from surface to seabed) nor the May 2013 report (a sort of trapezoid, rectangular on the surface and for some hundreds of meters down, but then converging from both the northern summer and southern winter boundaries onto the CG fracture zone all along the east-west centre line of the fracture) appear to match the actual structure of the SPF. If the feature has not been described correctly, it is not possible to submit a reliable proforma about it. In addition, a rechecking of the cited sources indicate that there is no direct evidence that, even for the first several hundred metres of the water column (wherever the front is at a given time) actually is more productive, more diverse, or more important to life histories of species than adjacent areas. This lack of evidence may be due to a lack of study (or a lack of time to find such studies). ICES has found few or no analyses of satellite data on primary productivity along the front as it moves seasonally, zooplankton productivity (possibly from analyses of CPR data), fish data from ICCAT, and bird foraging data to determine if seabirds concentrate on the front. ICES is aware of scientists who may be able to put together the analyses needed to better evaluate the case for the SPF, but this cannot be done within the timescale needed for this round of the CBD process. ICES is aware of the work done to advance the case for the North Pacific Front (which included expertise from the relevant tuna commission equivalent to ICCAT). ICES concede that such a proforma on the SPF will miss this cycle for the CBD process, but considers that there is little cost to that miss. The threats to the biological features of such a pelagic EBSA are fishing and shipping. Both threats have been present for decades to centuries, and there are agencies that manage them at present. There is no knowledge of any imminent changes to either activity that would markedly increase risks before the next CBD cycle of reporting. Separating the pelagic SPF from the benthic CGFZ would allow both proformas to be soundly drafted. Putting the CGFZ forward now ensures it is into the CBD process when a new and possible imminent threat of seabed mining is present, and the case is made on its merits. It is not weakened by present issues with the information for the SPF. ICES Advice 2013, Book 1 193 The case for the SPF can be evaluated better than it is now. It will go forward by the next cycle, and in the meantime, existing management authorities (in particular ICCAT) can be brought more fully into process. A stronger proforma for the SPF, with input directly from the regulatory authorities that will be most affected, mean that there is much less risk of opposition when it does go forward. 194 ICES Advice 2013, Book 1 1.5.6.1 Special request, Advice January 2013 ECOREGION SUBJECT General advice Ecological quality objective for seabird populations in OSPAR Region III (Celtic Seas) Advice summary ICES collated and analysed the data for the ecological quality objective (EcoQO) indicator on breeding seabird population trends in OSPAR Region III for the period 1986–2011. ICES considers that the proposed EcoQO indicator was not achieved in 1986, 1989–1992, 1996 and in consecutive years during 2003–2011. ICES advises that special attention is given to six bird species (Northern fulmar, Arctic skua, European shag, herring gull, black-legged kittiwake, and roseate tern) that are all below the lower target levels. ICES made two separate assessments of the EcoQO: one using both the upper and the lower target level, the other using only the lower target level. ICES recommends excluding the upper target level of 130% when calculating the EcoQO. ICES recommends to continue testing the application of alternative statistical methods in order to reduce the uncertainty linked to the rather wide confidence intervals when calculating the EcoQO. ICES advises that the target levels of the EcoQO may also be used in determining good environmental status (GES). The lower target levels of 70% or 80% (depending on the number of eggs) can be considered as corresponding to GES for the individual species. Birds that are above the 130% target level and are likely to have significant negative impacts on other species may be of concern, and ICES advises to consider these cases when assessing the GES. Request ICES is requested to: i) ii) update the value of the draft EcoQO indicator on Seabird Population Trends in OSPAR Region III (Celtic Seas) and make any relevant recommendations and, consider whether or not the target thresholds [both a) the target for a species-specific trend in abundance (e.g. 70% or more of the baseline); and b) the target for the proportion of species meeting species-specific targets (e.g. 75% or more)] used in the EcoQO would be indicative of a seabird community that is at GES. ICES advice Request item i) Update the value of the draft EcoQO indicator on Seabird Population Trends in OSPAR Region III (Celtic Seas) and make any relevant recommendations. Introduction The EcoQO on seabird population trends was adopted by OSPAR’s Biodiversity Committee (BDC) in 2012 (OSPAR Commission, 2012): Changes in breeding seabird abundance should be within target levels for 75% of species monitored in any of the OSPAR regions or their sub‐divisions. To date, assessments of the EcoQO have used target levels originally suggested by ICES (2008): intra-specific annual abundance should be less than or equal to 130% of the baseline and more than or equal to 80% of the baseline, for species that lay only one egg, or more than or equal to 70% for species that lay more than one egg. It has been debated whether or not an upper target level should be applied when establishing the EcoQO. To help resolve the issue, ICES made two separate assessments of the EcoQO: one uses the upper target of 130% for all species, and another which does not use the upper target for any species. ICES (2012) collated and analysed the most recent data for the EcoQO indicator on Seabird Population Trends in OSPAR Region III (Celtic Seas) (Figure 1.5.5.1.1). ICES Advice 2013, Book 1 195 Figure 1.5.5.1.1 OSPAR Region III. Data for OSPAR Region III are collected as part of the UK and Ireland’s Seabird Monitoring Programme (SMP). The ICES (2012) advice included data from 1986–2010 on twelve species (Northern fulmar, European shag, herring gull, great black-backed gull, black-legged kittiwake, Sandwich tern, common guillemot, razorbill, Arctic skua, great cormorant, little tern, and roseate tern). In this update data from 1986–2011 are included and one more species – common tern – is added: the indicator is now based on thirteen species. Most colonies in OSPAR Region III were not surveyed in each year of the time-series, so imputation techniques were used to estimate the missing counts. The imputation methods and reference values used in this update are identical to those described and used in ICES (2010, 2011) and Annex 1. Development of the EcoQO for OSPAR Region III Using the ‘old targets’ option that includes the upper target level, the EcoQO was not achieved in 1986, 1989–1992, 1996, and in consecutive years during 2003–2011 (see Figure 1.5.5.1.2a). Using the ‘new target option’ based only on the lower target levels, the EcoQO was not achieved in 1986, 1989–90, 1992, and consecutively from 2005–2011 (see Figure 1.5.5.1.2b). 196 ICES Advice 2013, Book 1 a) ‘Old targets’ option b) ‘New targets’ option Figure 1.5.5.1.2 The proportion of species in OSPAR Region III that were within target levels of abundance during 1986–2011. The EcoQO was not achieved in years when the proportion dropped below 75%. a) The “Old targets” option with an upper target level of 130% of the baseline and a lower target of 80/70% of the baseline. b) The “New target” option with no upper target level. For both options, the lower target levels were not achieved by six species in 2011, showing no change compared to the last updates in 2009 and 2010 (ICES, 2010, 2011). The six species are: Northern fulmar, Arctic skua, European shag, herring gull, black-legged kittiwake, and roseate tern. An overview of species with their Latin names is given in Table 1.5.5.1.1 below. ICES Advice 2013, Book 1 197 Table 1.5.5.1.1 Species-specific assessment of relative breeding abundance in the Celtic Seas in 2011. Green cells indicate that the species-specific targets have been met; orange cells indicate that the lower speciesspecific target has been met but that relative abundance has exceeded 130%; red cells indicate that the lower species-specific targets have not been met. Arrows indicate recent trends in relative abundance (2010–2011) 14. See also Figure 1.5.5.1.A1 in Annex 1. 1 Species Fulmarus glacialis English name Northern fulmar ↓ 2 Carbo aristotelis European shag ↓ 3 Carbo carbo great cormorant ↓ 4 Stercorarius parasiticus Arctic skua ↓ 5 Sterna sandvicencis Sandwich tern ↓ 6 Sternula albifrons little tern ↑ 7 Sterna dougalii roseate tern ↑ 8 Sterna hirundo common tern ↓ 9 Larus argentatus herring gull ↓ 10 Larus marinus great black-backed gull ↓ 11 Rissa tridactyla black-legged kittiwake ↔ 12 Uria aalge common guillemot ↓ 13 Alca torda razorbill ↑ Roseate tern abundance has been below the lower target throughout 1986–2011, but has steadily increased during this period from 18% to 48% of the reference level. European shag abundance was relatively lower than roseate tern in 2011 (i.e. 29% of reference level). Shag numbers have been at or below the lower target since 1993, but have been declining further since 2004. Herring gull numbers have been in decline since the early 1970s, but the reference level was set at the mid-1980s level because numbers were thought to have been previously elevated by anthropogenic activities (e.g. commercial fisheries). Numbers have been steadily decreasing since 2000 and fell below target levels from 2003 onwards. They are currently at 53% of the reference level. Arctic skua numbers have been below the lower target since 2005 and were at 40% of the reference level in 2011. The decline in Northern fulmar numbers started in the mid-1990s but was steeper during 2005–2008. Their numbers dropped below the target level in 2007, remained stable at 73% of the reference level in 2009 and 2010, but declined to 68% in 2011. Black-legged kittiwake numbers have been declining since around 2000 and dropped just below the target level in 2008, 2010, and again in 2011. Great black-backed gull numbers have remained within target levels throughout 1986–2011 and have shown a slight decrease the last years. Razorbill and common guillemot numbers increased steadily during the 1980s and 1990s. Guillemot numbers are more or less stable. Razorbill numbers peaked between 2002 and 2005 but subsequently dropped a little and have remained within target levels since 2006. Since 2000 the numbers of great cormorant increased but have declined since 2009, returning within target levels. Common tern, Sandwich tern, and little tern have been increasing since late 1990s. Numbers of common tern and Sandwich tern, though lower in 2011 compared to 2010, remain substantially above the target level. Little tern numbers dipped dramatically between 2006 and 2010, but in 2011 numbers were once again well above the target level. 14 198 There may be a need to define how to assess the trends. ICES Advice 2013, Book 1 ICES notes that it is useful to report on the trends in the different bird species as well. However, it is unclear how these trends should be established and reported, e.g. over how many years etc. ICES recommends that clear guidelines for reporting of seabird trends are developed. Uncertainty and confidence intervals The inherent uncertainties of the recorded (mean) trends are a cause for concern (see Figure 1.5.5.1.A1 in Annex 1). Disregarding uncertainty when reporting the EcoQO limits the usefulness of the results. The results in terms of meeting the target levels are interpreted mainly from the mean trends. Accordingly, the uncertainty of the trends is not assessed in relation to the targets for species-specific trends in abundance and the target for the proportion of species meeting speciesspecific targets (e.g. 75% or more) used in the EcoQO. As an example, the lower target levels were not achieved by six species; Northern fulmar, Arctic skua, European shag, herring gull, black-legged kittiwake, and roseate tern. By inspection of the upper confidence levels, however, only Arctic skua and shag did not meet the species-specific target levels in 2011. Yet, the confidence intervals are not explicitly used when making conclusions, neither on these two species nor on the four species of seabirds for which the declines are dubious. As a result, the failure to meet the overall target of the EcoQO in Region III is not questioned. On the other hand, when using the lower confidence intervals, also great cormorant, great black-backed gull, and common guillemot are below the lower target. The problem with using the confidence intervals is obvious and ICES advises not only the continued use of the mean value when assessing the EcoQO, but also to consider improving the statistical uncertainty. Although the Seabird Trend Wizard (see Annex I) has provided reliable confidence intervals computed by bootstrapping the count data, the confidence intervals are rather wide for most species. Obviously, sources of variation exist in the data which influence the uncertainty of the estimated trends. In addition, the Wizard does not smooth the count data which makes it sub-optimal for reproducing long time-series, with alternating periods of increases and declines. Both issues may be addressed in the further application of the EcoQO. ICES recommends to continue testing the application of alternative statistical methods like TrendSpotter, Generalised Additive Models, and Bayesian time-series models capable of smoothing the time-series and including co-variables, which may reduce the amount of residual ‘noise’ present in the data. Considerations The failure to achieve the EcoQO in OSPAR Region III in consecutive years between 2005 and 2011 (for both target setting options) does give rise to concern as 4–6 of the thirteen species sampled were below the lower target levels during this period, and five species have shown substantial declines. ICES advises that special attention is given to discover the possible causes of decline of these species and to take appropriate action. The declines in three of these species: roseate tern, Arctic skua, and herring gull have already been highlighted within the UK and have been listed on the UK Biodiversity Action Plan and on the Red list of Birds of Conservation Concern in the UK. Roseate tern numbers have been increasing as a direct result of intensive management of colonies in Ireland. Arctic skua are relatively scarce in OSPAR Region III but the trend in the region is following a steeper decline in the neighbouring Northern Isles (OSPAR Region II) where impacts of climate and fishing on food supply have been exacerbated by increased predation and competition from great skua. The cause of the decline in herring gulls throughout the UK and Ireland is less well understood and ICES advises further work on this species. The EcoQO highlights a substantial decline in shag numbers in OSPAR Region III. Declines have occurred in the rest of the UK but not to the same extent. ICES advises further investigation into the cause of the decline. The recent declines in kittiwake and fulmar numbers in OSPAR Region III are worth continued monitoring and further investigation is required to determine likely causes. Kittiwake colonies within OSPAR Region III have been more successful than colonies on the east coast of Britain (in OSPAR Region II), which have been in decline in some areas since the late 1980s. A shortage of sandeels off the east coast is probably responsible for poor breeding there, but kittiwakes at colonies in western Britain tend to feed on other species of fish. ICES advises more research into the variation in availability of these prey species and the link with the decline of seabird populations. The continued increase in guillemot numbers may be surprising when other predators of small shoaling fish (kittiwakes and shag) have been declining and razorbill numbers have levelled off. The large increase in common terns and Sandwich tern numbers is probably due to improved protection from predators at colonies. Despite the 2011 resurgence, the declines in little tern numbers over the previous four years may be of concern and reasons for the decline should be investigated. ICES debated whether or not the upper target level should be applied when establishing the EcoQO. ICES (2011) considered applying the upper target level only to predatory species that are likely to have significant negative impacts on other species but recommended that the EcoQO should remain unaltered, because it should be “a value-free, objective ICES Advice 2013, Book 1 199 metric that makes no assumptions about the underlying causes of individual seabird species population change.” However, in 2012 a group of UK experts considering UK MSFD targets recommended that the EcoQO be used without the upper target threshold for abundance of any species. The lack of any upper threshold was considered to be more objective than applying it only to certain species and would mean that the EcoQO would be much more straightforward. ICES has the following considerations. In OSPAR Region III the old target level (130%) was exceeded by populations that were previously in poor health (most tern species) or by species that were very scarce (cormorant), whereas the 130% level should rather be a warning for possible negative interactions with other species. The terns are not considered to have negative impacts on other species. This might be different for cormorants that compete with other species for space and with fisheries. However, the carbo subspecies is relatively scarce with just 52 000 pairs globally, and is culled both legally and illegally in the UK and Ireland. Also, the historical baseline will often reflect the onset of national monitoring schemes that in many cases are established after periods of large human impact. This makes the 130% target level questionable as a valid parameter for use in the context of EcoQO. However, birds that are above the 130% target and are likely to have significant negative impacts on other species could be of concern. Therefore, ICES recommends that species above the 130% target are also reported. Consequently, ICES recommends excluding the upper target threshold of 130% when calculating the EcoQO. Further development is recommended with respect to complementary quality objectives based on parameters such as breeding success, development of interpretation models in relation to foodwebs (information on relevant prey trends), arranging trend data into functional groups of seabirds, and inclusion of data regarding relevant sea duck species (ICES, 2012). Request item ii) Consider whether or not the target thresholds [both a) the target for a species-specific trend in abundance (e.g. 70% or more of the baseline); and b) the target for the proportion of species meeting species-specific targets (e.g. 75% or more)] used in the EcoQO would be indicative of a seabird community that is at GES. ICES notes that GES is rather a national concern, whereas EcoQO targets refer to larger regions more in line with biogeographical populations. National fluctuations in seabird abundance can be very large for some species and might be linked to fluctuations in neighbouring countries, especially when considering numbers outside the breeding season. Setting the right national reference levels (baseline) and selecting the right species thus seems very important when the EcoQO is to be used as an indicative value of a seabird community in the context of GES. When suggesting lower target levels of 70% or 80% depending on the number of eggs, ICES (2008) considered them as values of abundance that management should be trying to maintain with high probability. This is the same rationale that underlies target-setting to reflect the achievement of GES under MSFD. ICES advises that the lower target levels of 70% or 80% can be considered as corresponding to GES for the individual species. There is clearly more debate required about the inclusion of an upper target threshold for species-specific abundance for the GES. It is questionable whether a GES only based on the lower target levels is adequate enough to establish the environmental status. Birds that are above the 130% target and are likely to have significant negative impacts on other species may be of concern and ICES advises to consider these when assessing the GES. The EcoQO target threshold of 75% or more species meeting their abundance targets was recently put out to public consultation in the UK as part of its implementation of MSFD. Several NGOs suggested raising the threshold to 90%. Examination of Figure 1.5.5.1.2 (and applying a 90% threshold) shows that in OSPAR Region III, the EcoQO would not have been met at all during 1986–2011 under the “old targets” option and met in just two years under the “new target” option. Instead, the UK decided to keep the 75% threshold with the caveat that no species should be consistently missing their individual targets, where the cause of that decline can be directly linked to human activity. ICES supports the conclusions of the UK and recommends to keep the 75% target threshold. Sources ICES. 2008. Report of the Workshop on Seabird Ecological Quality Indicator (WKSEQUIN), 8–9 March 2008, Lisbon, Portugal. ICES CM 2008/LRC:06. 60 pp. ICES. 2010. Report of the Working Group on Seabird Ecology (WGSE), 15–19 March 2010, ICES Headquarters, Copenhagen, Denmark. ICES CM 2010/SSGEF:10. 77 pp. ICES 2011 Report of the Working Group on Seabird Ecology (WGSE), 1–4 November 2011, Madeira, Portugal. ICES CM 2011/SSGEF:07. 87 pp. ICES. 2012. Report of the Joint ICES/OSPAR Ad hoc Group on Seabird Ecology (AGSE), 28–29 November 2012, Copenhagen, Denmark. ICES CM 2012/ACOM:82. 30 pp. 200 ICES Advice 2013, Book 1 Lloyd, C., Tasker, M. L., and Partridge, K. 1991. The status of seabirds in Britain and Ireland. Poyser, London. Mitchell, P. I., Newton, S. F., Ratcliffe, N., and Dunn, T. E. 2004. Seabird Populations of Britain and Ireland. T. & A. D. Poyser, London. OSPAR Commission. 2012. Summary Record of the Meeting of the Biodiversity Committee (BDC) in Brest: 13–17 February 2012. OSPAR Convention for the Protection of the Marine Environment of the North-East Atlantic, BDC 12/8/1-E. Thomas, G. E. 1993. Estimating annual total heron population counts. Applied Statistics, 42: 473-486. Whilde, A. 1985. The All-Ireland Tern Survey 1984. Unpublished IWC/RSPB Report, Dublin. ICES Advice 2013, Book 1 201 Annex I Methods Since the first assessment of the EcoQO (ICES, 2008), the Joint Nature Conservation Committee (JNCC) in collaboration with Biomathematics and Statistics Scotland developed an analytical ‘wizard’ for estimating trends in breeding numbers of individual species at various geographical scales, including OSPAR regions. The seabird trend wizard uses a modified chain method, first developed by Thomas (1993), to impute values of missing counts based on information in other years and sites (details of the Thomas method are given in Annex 3 of ICES (2008)). The wizard is a small Delphi application that retrieves counts from an Access database and generates script files and a DOS batch file that instruct R to conduct the trend analysis using the Thomas (1993) method. A further advantage of the new wizard is that the analyses can incorporate both whole colony counts and plot counts, even when they exist for the same colony in the same year. The accuracy and precision of the modelled regional trend for Northern fulmar were increased by restricting data input from only those colonies that had been surveyed for five years or more during 1986–2011. Data from all other species contained colonies that were surveyed in two or more years during 1986–2011 (as in ICES 2008, 2010, 2011). This reduced the sample size for fulmar to just 7% of the total number of pairs known to breed in OSPAR Region III (1998– 02 Census; Mitchell et al., 2004), compared to over 50% in all other species. Baselines for each species were the same baselines used in ICES (2008, 2010, 2011). 202 ICES Advice 2013, Book 1 Ecological quality objectives on seabird population trends in OSPAR Region III ICES Advice 2013, Book 1 203 Figure 1.5.5.1.A1 204 Trends in abundance of individual species in OSPAR Region III, 1986–2011. Fine dotted lines indicate the upper and lower boot-strapped confidence limits. Bold dashed lines indicate the upper and lower target levels; 100 = reference level (baseline). For fulmars only colonies with minimum five years of data are used. ICES Advice 2013, Book 1 1.5.6.2 Special request, Advice January 2013 ECOREGION SUBJECT General advice Data collection and storage to implement the OSPAR seabird recommendations Advice summary ICES advises that the OSPAR contracting parties share the responsibility of collating the relevant data for the seven bird species mentioned in OSPAR’s Recommendations 2011/1–7. It is suggested that each of the individual contracting parties (lead countries) is given the responsibility of collecting and storing data for one (or several) species. The data collection should follow, whenever possible, international standard methods. Data for each species should be stored by the responsible lead country as part of their national monitoring schemes. While some data are easily stored in standard formats, others will require the development of more sophisticated formats for convenient data exchange and comparison among contracting parties. Standardization and harmonization of data collection methods and reporting is crucial to ensure the comparability that is necessary for comprehensive and broader assessments at the regional and subregional levels. Several initiatives are currently working towards international standards for collection, exchange, and storage of seabird data. ICES does not at this stage recommend the creation of a central database, but a central portal might be considered. ICES wishes to explore, together with OSPAR, the further development of a common format for seabird data collection and reporting. A dedicated workshop is suggested as a possibility for relevant country experts to meet and design the framework for the required data collection and reporting. ICES recommends that principles similar to those stated above should apply to all OSPAR seabird data. Request ICES is requested to advise on suitable arrangements (including format) for data collection and storage resulting from the implementation of OSPAR Recommendations 2011/1-7 on seabirds, taking into account existing data collection arrangements and compatibility with current developments under MSFD implementation. ICES advice In 2011 OSPAR adopted seven Recommendations (OSPAR 2011/1-7) for furthering the protection and conservation of seven bird species: • • • • • • • Lesser black-backed gull (Larus fuscus fuscus) Ivory gull (Pagophila eburnea) Little shearwater (Puffinus assimilis baroli) Balearic shearwater (Puffinus mauretanicus) Black-legged kittiwake (Rissa tridactyla tridactyla) Roseate tern (Sterna dougallii) Thick-billed murre (Uria lomvia) The purpose of the Recommendations is to strengthen the protection of all life stages of these species. Article 3.2 in the Recommendations states: “Acting collectively within the framework of the OSPAR Commission, Contracting Parties should: develop and implement a monitoring and assessment strategy and data collection tools to promote and coordinate the collection of information on distribution, status of, threats to and impacts on the species, that can contribute to the implementation of the Marine Strategy Framework Directive, where appropriate, including… “ …species-specific requirements. 1. Arrangements for data collection and storage ICES suggests that the contracting parties share the responsibility for collating the relevant data for the seven seabird species (according to the OSPAR Recommendations). This could be achieved by each of the individual contracting parties (lead countries) being given the responsibility for collecting and storing data for one (or several) species. ICES Advice 2013, Book 1 205 There is considerable variability in the available methods for collecting and reporting seabird data. This might lead to lack of comparability, preventing comprehensive and broader assessments at the regional and subregional levels. Consequently, standardization and harmonization of data collection and reporting (including formats) is of major importance. Data collection should follow international standard methods whenever possible. Walsh et al. (1995) present a compilation of methods for survey and monitoring of seabirds. Schmeller et al. (2012) provide an overview of bird monitoring in Europe. Both may form a useful basis for further development and agreement at the OSPAR level. Data for each species should be stored by the responsible lead country as part of their national monitoring schemes. While some data are easily stored in standard formats (e.g. population size, breeding success), others will require the development of more sophisticated formats for convenient data exchange among contracting parties (e.g. tracking data, diet data). Barrett et al. (2007) review different methods used to collect dietary data from marine birds. Several other initiatives are currently working towards international standards for collection, exchange, and storage of seabird data and therefore ICES does not recommend the creation of a central database at this stage. However, it is important that data are stored in a format that allows easy comparison and exchange of data among the contracting parties. The creation of a central portal may therefore be considered. ICES recommends that principles similar to those stated above should apply to all OSPAR seabird data. 2. Data collection arrangements and compatibility In many cases, data are already being collected as part of national monitoring schemes or will be collected as part of the implementation of the Marine Strategy Framework Directive (MSFD) or other directives, conventions, and international agreements. Table 1.5.5.2.1 identifies known data collections and suggests a responsible contracting party for each species; this requires further discussion by contracting parties. Table 1.5.5.2.1 Known data collections by species. This list may not be complete. Species Lesser black-backed gull Ivory gull Population trends Breeding success Survival rates Diets X X X X Contaminants Movements Suggested Lead country X X Norway X X Norway X Portugal X Spain X UK X X Little shearwater X X Balearic shearwater Black-legged kittiwake Roseate tern X X X X X X X X X X X X UK Thick-billed murre X X X X X Iceland X When deciding on seabird data collection and storage at the OSPAR level, some other issues must be taken into account. Firstly, it should be determined whether the database(s) should be open to public access. For seabirds much data is collected by interest organzations and NGOs (e.g. ESAS) in addition to data collected through state-financed projects and monitoring programmes. Also, should access be given to data within all parameter categories? Close coordination and comparability with other commitments must be ensured and overlapping actions avoided. ICES is aware of funding problems related to the international collation, assessment, and reporting of seabird data. So far ICES experts have collated and reported on data for OSPAR regions II and III, but funding for experts participating in ICES seabird meetings is becoming increasingly scarce, a problem that should be addressed by OSPAR. ICES recommendation ICES wishes to explore, together with OSPAR, the further development of a common format for seabird data collection and reporting. A dedicated workshop is suggested as a possibility for relevant country experts to meet and design the framework for the required data collection and reporting. 206 ICES Advice 2013, Book 1 Sources Barrett, R. T., Camphuysen, K., Anker-Nilssen, T., Chardine, J. W., Furness, R. W., Garthe, S., Hüppop, O., Leopold, M. F., Montevecchi, W. A., and Veit, R. R. 2007. Diet studies of seabirds: a review and recommendations. ICES Journal of Marine Science, 64: 1675–1691. ICES. 2011. Report of the Working Group on Seabird Ecology (WGSE), 1–4 November, Madeira, Portugal. ICES CM 2011/SSGEF:07.87 pp. ICES. 2012. Report of the Joint ICES/OSPAR Ad hoc Group on Seabird Ecology (AGSE), 28–29 November 2012, Copenhagen, Denmark. ICES CM 2012/ACOM:82. 30 pp. Schmeller, D. S., Henle, K., Loyau, A., Besnard, A., and Henry, P-Y. 2012. Bird-monitoring in Europe – a first overview of practices, motivations and aims. Nature Conservation, 2: 41–57. (www.pensoft.net/journals/natureconservation/article/3644) Walsh, P. M., Halley, D. J., Harris, M. P., del Nevo, A., Sim, I. M. W., and Tasker, M. L. 1995. Seabird monitoring handbook for Britain and Ireland. JNCC/RSPB/ITE/Seabird Group, Peterborough, UK. ICES Advice 2013, Book 1 207 1.5.6.3 Special request, Advice June 2013 ECOREGION General advice SUBJECT OSPAR special request on review of the technical specification and application of common indicators under D1, D2, D4, and D6 Advice summary ICES undertook a simulation benchmarking exercise on the 35 common indicators whose technical specifications had been supplied by OSPAR. ICES advises that 17 of the indicators in the Greater North Sea can be considered highperforming; 14 are close to being fully operational and 19 are adequately monitored throughout the North Sea. ICES did not assess the performance of the indicators in regions other than the Greater North Sea, but considers that analysis of this region will help in the understanding and further development of indicators in other regions. ICES cannot define the precise nature of good environmental status (GES) as this is an EU Member State issue, but advises that in order to better understand GES, it would be helpful to develop further indicators, so that a more holistic ecosystem view can be achieved and potential additional monitoring needs can be identified. A technical review of the two OSPAR common indicators on non-indigenous species is provided. Request ICES is requested to undertake an independent peer review of the technical specifications and proposed operational implementation of the indicators (COBAM draft indicators) presented. The review should consider, from the perspective of producing a set of common indicators for the OSPAR Region: 1. 2. 3. 4. whether the indicators put forwards are appropriate to implement at a regional scale; whether the set of indicators is sufficient as a set to understand GES; identify any gaps; identify where there are difficulties in the operationalization of the indicators, with proposals for how to overcome these. Based on the outcomes of Request regarding maximising efficiencies for monitoring of biodiversity: 5. identify where there are opportunities to cluster indicators that can benefit from shared monitoring/ data collection. (OSPAR request 3/2013) ICES has addressed consideration 5 in its response to the request on maximizing the use of available sources of data for monitoring of biodiversity (ICES Advice 2013, Section 1.5.5.2). ICES advice Overall review Based on 16 criteria, 35 indicators were reviewed based on the information available in the Technical Specifications provided by OSPAR ICG–COBAM (for four indicators no technical specifications were made available). The criteria approach used here can help to guide future selection for currently underdeveloped indicators, providing a method for selecting one indicator over the other. The results of the assessment for the Greater North Sea are summarized below for three ecosystem aspects of the indicators. Overall performance A simulation benchmarking exercise was conducted on the 35 common indicators whose technical specifications had been supplied by OSPAR. Based on the scoring of the criteria and the thresholds set for the analysis ICES found that 17 of the indicators in the Greater North Sea can be considered high-performing; 14 are close to being fully operational, and 19 are adequately monitored throughout the North Sea. The interpretation of the indicator scores depends largely on the benchmarking procedure used and the resulting thresholds. The outcome of the assessment is affected by both the knowledge of the performance of the indicator and by the level of detail provided in the technical specifications. 208 ICES Advice 2013, Book 1 Operational implementation To improve the 21 indicators that are identified as less operational, in particular those that are currently categorized as core indicators, ICES recommends focussing further work on (1) integrating surveys and improving the current cost effectiveness of data collection (see reply to OSPAR request 4/2013 in ICES Advice 2013, Section 1.5.5.2) and (2) extending the spatial scale of the existing monitoring. Benthic and pelagic habitat indicators will need improvement in terms of monitoring and ability to understand how the metric can vary, whereas for species indicators, particularly for birds and mammals, improvements should be directed towards the spatial extent of monitoring. The results of the evaluation presented here only apply to the Greater North Sea (OSPAR Region II). Regional coverage ICES was unsure as to the meaning of “appropriate to implement at a regional scale” as this could mean at an subregional scale, or it might imply ‘appropriate for all OSPAR regions’. An analytical evaluation exercise was undertaken to assess the adequacy of current monitoring (as described in the technical specifications) for the Greater North Sea. Seventeen indicators have inadequate geographical monitoring within the North Sea (i.e. monitoring is undertaken across a limited fraction of the sub-region). Considering the Greater North Sea as a relatively data-rich region, the results are likely to be lower in other OSPAR regions. For many indicators, however, information on relevant monitoring programmes was limited in the technical specifications and this may have biased the results presented here. Understanding GES and gap analysis Identification of the critical ecosystem components and the most effective indicators to monitor these is essential to understand GES. Three of these ecosystem aspects were reviewed by ICES: 1) whether there are gaps in the list of OSPAR indicators compared to the indicators defined by the European Commission (Decision 2010/477/EU); 2) whether there are gaps in the list of indicators for important parts of the marine ecosystem; and finally 3) whether any of the present indicators are redundant. Gaps in OSPAR’s list of common indicators related to MSFD The 35 OSPAR common indicators do not address fully the Marine Strategy Framework Directive (MSFD) indicators listed in the EU Decision (2010/477/EU). ICES notes that it is not essential for all ecosystem aspects to be covered by OSPAR common indicators, as further indicators may be implemented individually by EU Member States to help in defining GES. • • • • • • • Within the three species-level ecosystem components (birds, mammals, and fish), no OSPAR common indicator addresses the requirement for indicators of ‘population genetic structure’ (MSFD indicator 1.3.2). No seabird or mammal OSPAR common indicator addresses the requirement for indicators of ‘composition and relative proportions of ecosystem components’ (MSFD indicator 1.7.1). None of the OSPAR common indicators addressing foodweb (MSFD indicators 4.1.1 and 4.3.1) use benthic invertebrate metrics. MSFD indicator 1.1.3 applies specifically to benthic species and habitats, yet none of the OSPAR common indicators address the requirements for this indicator. No OSPAR common indicator for fish and cephalopods is linked to the distribution range and distribution pattern (MSFD indicators 1.1.1 and 1.1.2), or to the population demographics (MSFD indicator 1.3.1). ICES notes that two distribution indicators (FC-7 and FC-8) and a proportion of mature fish (FC-6) indicator were listed in part B of ICG–COBAM’s report as potential candidate indicators, but no details were provided in the technical specifications. None of the benthic habitat OSPAR common indicators addressed the habitat-level MSFD indicator requirements for distributional range (1.4.1), distributional pattern (1.4.2), or volume (1.5.2). None of the OSPAR common indicators address the impact of non-indigenous species (MSFD indicators 2.2.1 and 2.2.2). In addition ICES found four major issues relevant to poor linkages between MSFD Decision indicators and OSPAR common indicators as specified in the ICG–COBAM document (Table 1.5.5.1.1). ICES Advice 2013, Book 1 209 Table 1.5.5.1.1 Problems with linkages between MSFD Decision indicators and OSPAR common indicators as specified in the ICG–COBAM document. The issues (left hand column) are: (1) The OSPAR common indicator does not seem to relate to the MSFD indicator; (2) This multimetric indicator does not fit the definition of the MSFD indicator, but is implicitly part of it; (3) This OSPAR indicator is considered relevant by OSPAR, but is not a MSFD indicator specified in the Decision document; (4) This linkage is tenuous, with insufficient detail provided in the ICG–COBAM Technical Specifications document to be convincing. No. of issue MSFD Decision indicator OSPAR ICG–COBAM indicator 1 1.2.1 (Population abundance and/or biomass) 1 1.3.1 (Population demographic characteristics) 1 1.3.1 (Population demographic characteristics) 1 1.3.1 (Population demographic characteristics) 1 1.6.1 (Condition of the typical species and communities) 1.6.2 (Relative abundance and/or biomass) FC-4 (Bycatch rates of Chondrichthyes) M-6 (Proportion of bycaught individuals within a species population) B-4 (Non-native/invasive mammal presence in island seabird colonies) B-5 (Mortality of marine birds from fishing (bycatch) and aquaculture) FC-3 (Mean maximum length of demersal fish and elasmobranchs) FC-2 (OSPAR EcoQO for proportion of large fish (LFI)) BH-2 (Multi-metric indices) 1 2 2 2 3 4 4 1.6.1 (Condition of the typical species and communities) 1.6.2 (Relative abundance and/or biomass) 6.2.1 (Presence of particularly sensitive and/or tolerant species) 4.3.1 (Abundance trends of functionally important selected groups/species) 6.2.2 (Multi-metric indices assessing benthic community condition and functionality) 6.2.2 (Multi-metric indices assessing benthic community condition and functionality) BH-2 (Multi-metric indices) BH-2 (Multi-metric indices) FW-9 (Ecological network analysis indicator (e.g. trophic efficiency, flow diversity) PH-1 (Changes in plankton functional types (life form) index ratio) FW-5 (Change in plankton functional types (life form) index ratio between: gelatinous zooplankton and fish larvae; copepods and phytoplankton; holoplankton and meroplankton Ecological gaps The proposed OSPAR common indicators reflect the seven OSPAR ecosystem components (seabirds, marine mammals and reptiles, fish and cephalopods, benthic and pelagic habitats, foodwebs, and non-indigenous species). However, focus on such broad groups may mean that other ecosystem components are ignored, and some of these may indeed have a bearing on determining whether GES at the whole ecosystem level has actually been achieved in any given OSPAR region. ICES therefore listed potential ecological gaps (Table 1.5.5.1.2), which may aid the further development of new indicators and identify potential additional monitoring needs. 210 ICES Advice 2013, Book 1 Table 1.5.5.1.2 Gaps in the OSPAR common indicators that may need to be filled to fully define GES. OSPAR ecosystem component Rocky and mixed benthic habitats Deep-water habitats and species Microplankton and microbenthos Cephalopods Coastal and inshore fish communities Highly migratory fish and reptiles Non-indigenous species Identified gaps There is little sampling of rocky and mixed benthic habitats for the status assessment of benthic fauna and fish species, partly due to the risk of sustaining damage to the trawl. Rocky habitats often host sensitive species and may provide refuge from fishing. The majority of surveys operate in coastal shelf seas and not in deeper waters, including those within the North Sea, Skagerrak, and Kattegat. For PH-1 and FW-5 (Table 1.5.5.1.1), OSPAR identified micro-, pico-, nano-phytoplankton and bacteria and micro-zooplankton including ciliates as being undersampled. These groups are essential components of the microbial loop of marine food webs but ICES notes that substantial effort would be required to obtain reasonably precise estimates given the extremepatchiness of such organisms in space and time. Cephalopods are caught in fisheries research surveys (e.g. IBTS) and hence data for the assessment of status of at least some species under FC-1, FC-7, and FC-8 (Table 1.5.5.1.1) should be available. Coastal fish communities are not required to be monitored under the Water Framework Directive. Existing surveys do not sample in shallower coastal zone waters. Shallow coastal waters are important during the juvenile phase of the life-history of many fish species. Sharks, tunas, and other highly migratory fish are only partly addressed by the OSPAR common indicators and none consider the status of reptiles. The importance of different pathways and associated vectors for each country should be assessed, after which a final decision on common indicators should be made. In addition to invasion vectors and pathways, the monitoring strategy should also depend on the taxa to be sampled. Impacts caused by non-indigenous species should be assessed. Redundant indicators In some cases, correlations between the health of different ecosystem components and/or indicators could be used to reduce the number of indicators needed. Potential correlation between indicators should be recognised in order to avoid misleading impressions of actual progress towards GES. ICES suggests a criterion “Indicators making up a suite of indicators should reflect variation in different attributes of the ecosystem component and thus be complementary” to identify situations where indicator redundancy could be an issue. Since the indicators are at varying stages of development and/or the information necessary to assess each OSPAR common indicator against this criterion was not provided in the ICG–COBAM Technical Specifications document, ICES was not able to assess whether any of the currently listed indicators contain elements of redundancy. Background ICES evaluated each indicator (based on the technical specifications supplied by OSPAR) using a standardized framework (ICES, 2013a). It is important to note the distinction between the indicator and its technical specification, as a poor specification could devalue a good indicator. ICES could only work with the specifications that had been provided and could not make assumptions about what was not in the specification. Table 1.5.5.1.3 lists the criteria used to evaluate the performance of OSPAR’s proposed common indicators and provides the importance weightings assigned to each criterion, their associated scores, and the guidelines for assessing the compliance of each indicator against each criterion. These criteria were essentially devised to assess the performance of ‘state’ indicators. However, the OSPAR common indicators also include ‘pressure’ indicators. It is inappropriate to evaluate such indicators against criteria for assessing the state indicators to variation in pressure. A pressure indicator should be, by definition, extremely sensitive and responsive to variation in the ecological pressure it purports to measure. Each indicator was therefore first assessed against criterion 1, which distinguished state indicators from pressure indicators. If the indicator was considered to be a pressure indicator, then it was automatically given a compliance score of zero against criteria 6, 8, 12, and 13 (highlighted in Table 1.5.5.1.3), as these are relevant only to state indicators. The importance weightings assigned to each criterion were given scores of Essential = 3, Desirable = 2, and Informative = 1, and the compliance fits were assigned scores of Fully met = 1, Partially met = 0.5, and Not met = 0. Multiplying these two values together provided a score for the performance of each indicator against each criterion. Summing these scores across all criteria then generated an overall score for the general performance of the indicator against all the criteria. The decision to give pressure indicators a compliance score of 0 introduces a bias in the assessment process in favour of state indicators. ICES notes the need for both “state” and “pressure” indicators so that the “pressure–state” relationship ICES Advice 2013, Book 1 211 can be adequately defined, and pressure indicators are essential in providing the scientific basis for advice regarding the most appropriate management measures required to achieve GES. Pressure indicators are also required for Descriptor 6. Criterion 16 considers correlations between indicators and was not used in the evaluation of the performance of the OSPAR common indicators. This criterion is intended to select metrics that measure different attributes of an ecosystem component’s condition, and to discourage selection of metrics that essentially perform similar functions. Therefore, this criterion should be applied after the main assessment process, to aid further selection between high performing indicators. 212 ICES Advice 2013, Book 1 Table 1.5.5.1.3 Criterion No. Criteria used to evaluate the performance of common indicators proposed by OSPAR to support implementation of the MSFD at the sub-regional and regional scales. The 16 criteria are grouped into five main categories, and the principle characteristic of each indicator’s performance examined by each criterion is given. The importance weightings, and their associated scores, are shown, as are the guidelines for assessing the level of compliance of each indicator against each criterion. Pale blue cells indicate criteria that were not used in the evaluation. In the compliance guidelines column, criteria are given a zero compliance score if the indicator relates to a ‘pressure’ (Criterion 1). Category Characteristic Criterion Type of Indicator State or pressure Is a “pressure” indicator being used for want of an appropriate “state” indicator? Importance Weighting Importance Score A 1 Quality of underlying data Existing and ongoing data Indicators must be supported by current or planned monitoring programmes that provide the data necessary to derive the indicator. Ideal monitoring programmes should have a time series capable of supporting baselines and reference point setting. Data should be collected on multiple sequential occasions using consistent protocols, which account for spatial and temporal heterogeneity. Indicators should Indicators should ideally be easily and accurately be concrete determined using technically feasible and quality assured methods, and have a high signal-to-noise ratio. Essential 3 Essential 3 Quantitative Quantitative measurements are preferred over versus qualitative qualitative, categorical measurements, which in turn are preferred over expert opinions and professional judgments. Relevant spatial Data should be derived from a large proportion of coverage the MSFD sub-region, at appropriate spatial resolution and sampling design, to which the indicator will apply. Desirable 2 Essential 3 2 Quality of underlying data 3 4 Quality of underlying data Quality of underlying data 5 ICES Advice 2013, Book 1 Guidelines for Compliance Assessment Score B Fully met (1): indicator is a “state” indicator. Not met (0): indicator is actually a “pressure” indicator. Although scoring 0 in this criterion, and linked criterion further on in this table, ICES recognises that pressure indicators are essential in management decision making and in indicators for D6. Fully met (1): long-term and ongoing data from which historical reference levels can be derived and past and future trends determined. Partially met (0.5): no baseline information, ongoing monitoring or historical data available, but monitoring programme discontinued; however, potential to re-establish the programme exists. Not met (0): data sources are fragmented, no planned monitoring programme in the future. Fully met (1): data and methods are technically feasible, widely adopted, and quality assured in all aspects, signal-tonoise ration is high. Partially met (0.5): potential issues with quality assurance, or methods not widely adopted, poor signal-to-noise ratio. Not met (0): indicator is not concrete or doubtful; noise excessively high due either to poor data quality or the indicator is unduly sensitive to environmental drivers. Fully met (1): all data for the indicator are quantitative. Partially met (0.5): data for the indicator are semiquantitative or largely qualitative. Not met (0): the indicator is largely based on expert judgement. Fully met (1): spatially extensive monitoring is undertaken across the sub-region. Partially met (0.5): monitoring does not cover the full sub-region, but is considered adequate to assess status at the sub-regional scale. Not met (0): monitoring is undertaken across a limited fraction of the sub-region and considered inadequate to assess status at the sub-regional scale. 213 Criterion No. Category Characteristic Criterion Importance Weighting Importance Score A Guidelines for Compliance Assessment Score B Quality of underlying data Reflects changes in ecosystem component that are caused by variation in any specified manageable pressures The indicator reflects change in the state of an ecological component, caused by specific significant manageable pressures (e.g. fishing mortality, habitat destruction). The indicator should therefore respond sensitively to particular changes in pressure. The response should be unambiguous and in a predictable direction, based on theoretical or empirical knowledge, thus reflecting the effect of change in pressure on the ecosystem component in question. Ideally the pressure–state relationship should be defined under both the disturbance and recovery phases. Essential 3 Management Relevant to MSFD management targets Relevant to management measures Clear targets that meet appropriate target criteria (absolute values or trend directions) for the indicator can be specified that reflect management objectives, such as achieving GES. Indicator links directly to management response. The relationship between human activity and resulting pressure on the ecological component is clearly understood. Desirable 2 Desirable 2 Comprehensible Indicators should be interpretable in a way that is easily understandable by policy-makers and other non-scientists (e.g. stakeholders) alike, and the consequences of variation in the indicator should be easy to communicate. Desirable 2 IF CRITERION 1 IS SCORED 0 THEN THE COMPLIANCE SCORE B MUST BE 0. Otherwise: Fully met (1): the indicator is primarily responsive to a single or multiple pressures and all the pressure–state 15 relationships are fully understood and defined, both under the disturbance and recovery phases of the relationship. Partially met (0.5): the indicator’s response to one or more pressures are understood, but the indicator is also likely to be significantly influenced by other non-anthropogenic (e.g. environmental) drivers, and perhaps additional pressures, in a way that is not clearly defined. Response under recovery conditions may not be well understood. Not met (0): no clear pressure–state relationship is evident. Fully met (1): an absolute target value for the indicator is set. Partially met (0.5): no absolute target is set for the indicator, but a target trend direction for the indicator is established. Not met (0): targets or trends unknown. IF CRITERION 1 IS SCORED 0 THEN THE COMPLIANCE SCORE B MUST BE 0. Otherwise: Fully met (1): both response-activity and activity–pressure relationships 16 are well defined – advice can be provided on both the direction AND the extent of any change in human activity required, and the precise management measures required to achieve this. Partially met (0.5): response– activity and activity–pressure relationships are not well understood, or only one of the relationships is defined, but not the other, so that the precise changes in pressure resulting from particular management actions cannot be predicted with certainty. Not met (0): no clear understanding of either relationship, so that the link between management response and pressure is completely obscure. Fully met (1): the indicator is easy to understand and communicate. Partially met (0.5): a more complex and difficult to understand indicator, but one for which the meaning of change in the indicator value is easy to 6 7 Management 8 Management 9 15 The term “pressure–state relationship” is used here in the sense described by Piet et al. (2007): e.g. fishing pressure (fishing mortality rate [F]) – state of the stock (stock biomass [B]). 16 Here the terms response–activity relationship and activity–pressure relationship are used in the sense described by Piet et al. (2007); e.g. management response (total allowable catch) – fishing activity (daysat-sea), and fishing activity (days-at-sea) – fishing pressure (fishing mortality rate [F]). 214 ICES Advice 2013, Book 1 Criterion No. Category Characteristic Criterion Importance Weighting Importance Score A Guidelines for Compliance Assessment Score B communicate. Not met (0): the indicator is neither easy to understand nor communicable. Management Established indicator Management Costeffectiveness Management Early warning Conceptual Scientific credibility 10 11 12 13 Conceptual 14 15 Conceptual Indicator suites 16 Indicators used in established management frameworks (e.g. EcoQO indicators) are preferred over novel indicators that perform the same role. Internationally used indicators should have preference over indicators used only at a national level. Sampling, measuring, processing, analysing indicator data, and reporting assessment outcomes, should make effective use of limited financial resources. Indicators that signal potential future change in an ecosystem attribute before actual harm is indicated are advantageous. These could facilitate preventive management, which could be less costly than restorative management. Scientific, peer-reviewed findings should underpin the assertion that the indicator provides a true representation of variation in the ecosystem attribute in question. Metrics relevance For descriptors D1 and D6, metrics should fit the to MSFD indicator function stated in the 2010 MSFD indicator Decision document. This requirement can be relaxed for D4 indicators because the Decision document stipulates the need for indicator development in respect of this Descriptor (but any newly proposed D4 indicators must still fulfil the overall goals stated for D4). Cross-application Metrics that are applicable to more than one MSFD indicator are preferable. Indicator Different indicators making up a suite of indicators correlation should each reflect variation in different attributes of the ecosystem component and thus be complementary. Potential correlation between indicators should be avoided. ICES Advice 2013, Book 1 Desirable 2 Fully met (1): the indicator is established and used in international policy frameworks. Partially met (0.5): the indicator is established as a national indicator. Not met (0): the indicator has not previously been used in a management framework. Desirable 2 Informative 1 Desirable 2 Essential 3 Fully met (1): little additional costs (no additional sampling is needed). Partially met (0.5): new sampling on already existing programmes is required. Not met (0): new sampling on new monitoring programmes are necessary. IF CRITERION 1 IS SCORED 0 THEN THE COMPLIANCE SCORE B MUST BE 0. Otherwise: Fully met (1): indicator provides early warning because of its high sensitivity to a pressure or environmental driver with short response time; Not met (0): relatively insensitive indicator that is slow to respond. IF CRITERION 1 IS SCORED 0 THEN THE COMPLIANCE SCORE B MUST BE 0. Otherwise: Fully met (1): peer-reviewed literature. Partially met (0.5): documented, but not peer-reviewed. Not met (0): not documented, or peer-reviewed literature is contradictory. Fully met (1): the metric complies with indicator function. Not met (0): the metric does not comply with indicator function. Desirable 2 Desirable 2 Fully met (1): metric is applicable across several MSFD indicators. Not met (0): no cross-application. Fully met (1): the indicators are uncorrelated. Partially met (0.5): correlation between some indicators. Not met (0): all indicators are correlated. 215 The analytical evaluation exercise was undertaken only for the Greater North Sea due to there being insufficient data and expertise on the other MSFD-relevant OSPAR sub-regions present at the relevant ICES expert group meetings. ICES considers that the process could be readily applied to the remaining OSPAR sub-regions. Evaluation of each of the OSPAR common indicators against criteria 2 to 15 was undertaken independently by at least three separate experts and the mean overall assessment scores, along with the range of overall scores, was determined (Figure 1.5.5.1.1). 17 Figure 1.5.5.1.1 Mean and range of three independent evaluations (four in the case of the foodweb (FW) indicators) of the performance of 35 OSPAR common indicators against criteria 2 to 15 listed in Table 1.5.5.1.3. Pressure (P) indicators were automatically assigned a zero compliance score against four criteria deemed not applicable to pressure indicators (see text and Table 1.5.5.1.3). For abbreviation of indicators see Table 1.5.5.1.4. The OSPAR common indicators are intended to enhance cooperation between Member States sharing marine regions so that status assessment can be made across whole MSFD sub-regions and regions. The mean scores given to each OSPAR common indicator against criterion 5 (relevant spatial coverage) was therefore determined to identify the most useful OSPAR common indicators in this respect (Figure 1.5.5.1.2). ICES assessed the degree to which each of the proposed OSPAR common indicators could be readily used, based on each indicator’s performance against criteria 2, 3, 4, 5, 10, and 11. The mean score given to each OSPAR common indicator against these criteria was determined (Figure 1.5.5.1.3). 17 It should be noted that ICES assessed the performance of 35 OSPAR common indicators. ICES is aware that several more common indicators are under consideration, but the version of the document “Report by ICG–COBAM on the development of an OSPAR common set of biodiversity indicators: Part C: Technical Specifications” available to expert groups only provided details for 35 indicators. 216 ICES Advice 2013, Book 1 Table 1.5.5.1.4 Abbreviations and categories of the 35 proposed biodiversity OSPAR common indicators as described in the ICG–COBAM Part C: Technical Specifications document. Code Indicator Mammals M-1 Distributional range and pattern of grey and harbour seal haul-out sites and breeding colonies M-2 Distributional range and pattern of cetaceans species regularly present M-3 Abundance of grey and harbour seal at haul-out sites M-4 Abundance at the relevant temporal scale of cetacean species regularly present M-5 Harbour seal and Grey seal pup production M-6 Numbers of individuals within species being bycaught in relation to population Marine birds Species-specific trends in relative abundance of non-breeding and breeding marine bird B-1 species B-2 Annual breeding success of kittiwake B-3 Breeding success/failure of marine birds B-4 Non-native/invasive mammal presence on island seabird colonies B-5 Mortality of marine birds from fishing (bycatch) and aquaculture B-6 Distributional pattern of breeding and non-breeding marine birds Fish and cephalopods FC-1 Population abundance/ biomass of a suite of selected species FC-2 OSPAR EcoQO for proportion of large fish (LFI) FC-3 Mean maximum length of demersal fish and elasmobranchs FC-4 Bycatch rates of Chondrichthyes Benthic habitat BH-1 Typical species composition BH-2 Multi-metric indices BH-3 Physical damage of predominant and special habitats BH-4 Area of habitat loss BH-5 Size–frequency distribution of bivalve or other sensitive/indicator species Pelagic habitat PH-1 Changes of plankton functional types (life form) index ratio PH-2 Plankton biomass and/or abundance PH-3 Changes in biodiversity index (s) Food webs FW-1 Reproductive success of marine birds in relation to food availability FW-2 Production of phytoplankton FW-3 Size composition in fish communities (LFI) FW-4 Changes in average trophic level of marine predators FW-5 Change of plankton functional types (life form) index ratio FW-6 Biomass, species composition, and spatial distribution of zooplankton FW-7 Fish biomass and abundance of dietary functional groups FW-8 Changes in average faunal biomass per trophic level FW-9 Ecological network analysis indicator (e.g. trophic efficiency, flow diversity) Non-indigenous species NIS-1 Pathways management measures NIS-2 Rate of new introductions of NIS (per defined period) ICES Advice 2013, Book 1 Category Core Core Core Core Core Core Core Core Core Candidate Core Core Core Core Candidate Core Core Candidate Candidate Candidate Core Core Core Core Core Core Core Core Candidate Candidate Candidate Candidate Candidate Candidate 217 Figure 1.5.5.1.2 Spatial coverage. Evaluation of the performance of 35 OSPAR common indicators (abbreviations in Table 1.5.5.1.4) against criterion 5 (relevant spatial coverage) listed in Table 1.5.5.1.3. Indicators B-4 and B-5 received zero scores. P indicates a “pressure” indicator. Figure 1.5.5.1.3 Useability. Evaluation of the performance of 35 OSPAR common indicators (abbreviations in Table 1.5.5.1.4) against criteria 2, 3, 4, 5, 10, and 11 (related to the ability to use the indicators immediately) listed in Table 1.5.5.1.3. P indicates a “pressure” indicator. Comparing between categories of indicators The process undertaken by ICES means that it is not possible to directly compare pressure and state indicators, nor their readiness for immediate use. It would be possible for experts to chose thresholds, but these would be partly arbitrary and would reflect the opinions (and possible biases) of experts involved in any given indicator assessment exercise; different groups of experts might therefore set different benchmark threshold scores, thus generally compromising the repeatability and consistency of such objective indicator assessment exercises. A simulation exercise was therefore undertaken (ICES, 218 ICES Advice 2013, Book 1 2013b) in order to understand the significance of scores in each of these categories (Figure 1.5.5.1.4, Table 1.5.5.1.5). ICES has based the advice on the performance of indicators on the thresholds that derive from this process. In Figure 1.5.5.1.4 is shown the simulated distribution of scores that will likely be generated when a very large set of virtual “state” and “pressure” indicators are put through the scoring system. Figure 1.5.5.1.4 indicates the mean score and the upper 25-percentile, upper 10-percentile, and upper 5-percentile benchmark scores on each distribution. For overall assessment against all 15 criteria, “pressure” indicators could not score as highly as “state” indicators because scores against four criteria were all automatically set to zero; hence the different distributions and benchmark thresholds illustrated in Figure 1.5.5.1.4. When considering the operational implementation of each indicator, only data from six criteria were used, giving the third distribution and set of benchmark scores. In this latter case all six criteria applied to both “state” and “pressure” indicators, so both indicator types had the same distribution and benchmark thresholds. The precise benchmark score thresholds are shown in Table 1.5.5.1.3. ICES considers that assessment scores falling above the upper 5-percentile benchmark threshold identify statistically significant high-performing indicators. Figure 1.5.5.1.4 Score distributions for 100 000 virtual indicators (for details see ICES, 2013b), showing mean (light blue), upper 25-percentile (red), upper 10-percentile (blue), and upper 5-percentile (green) scores, computed using randomly sampled categories (for score B) for state indicators (top), pressure indicators (middle), and for the subset of categories that were considered to reflect the operational implementation of each indicator (bottom). Table 1.5.5.1.5 Summary of simulated indicators: mean and upper percentiles for the distribution of randomized indicator scores. Summary statistic Mean Upper 25 percentile Upper 10 percentile Upper 5 percentile ICES Advice 2013, Book 1 State 0.500 0.578 0.656 0.688 Pressure 0.375 0.453 0.516 0.547 Operation 0.501 0.633 0.733 0.767 219 Table 1.5.5.1.6 shows the performance of the 35 common indicators against the benchmark thresholds for overall performance, adequate coverage of the Greater North Sea MSFD sub-region, and for how close each is to becoming fully operational. The analysis reflects the greater availability of appropriate fish community data and the relatively long history of developing and using ecological indicators based on these data. Table 1.5.5.1.6 Evaluation of the performance of the OSPAR common indicators against criteria 2 to 15; against the six criteria related to the ability to use the indicators, and against the criterion related to spatial coverage of the indicators. Green cells show where indicators meet the benchmark thresholds and red cells show where indicators do not meet the benchmark thresholds. For abbreviation of indicators see Table 1.5.5.1.4. Common Indicator M-1 M-2 M-3 M-4 M-5 M-6 B-1 B-2 B-3 B-4 B-5 B-6 FC-1 FC-2 FC-3 FC-4 BH-1 BH-2 BH-3 BH-4 BH-5 PH-1 PH-2 PH-3 FW-1 FW-2 FW-3 FW-4 FW-5 FW-6 FW-7 FW-8 FW-9 NIS-1 NIS-2 TOTAL Overall Operational Spatial Coverage 17 14 19 MSFD indicators 1.4.1, 1.4.2, 1.5.1, 1.5.2, 1.6.1, 1.6.2, and 1.6.3 all relate to ‘habitat-level of biodiversity’, and so are pertinent to both the benthic habitats and pelagic habitats ecosystem components, but not to birds, mammals, or fish. Despite this, the ICG–COBAM Technical Specification document links two fish component indicators, the large fish indicator (FC-2) and the mean maximum length (FC-3), to MSFD indicators 1.6.2 and 1.6.1, respectively. ICES considers that it would be more appropriate for these OSPAR common indicators to be linked to the ecosystem/community level of biodiversity covered by MSFD indicator 1.7.1 (a linkage is also specified in the ICG–COBAM Technical Specification document). 220 ICES Advice 2013, Book 1 Comments on indicators for non-indigenous species • • • • • • • • The technical description of the proposed NIS-1 indicator is insufficiently detailed and is incomplete. ICES considers that both the indicator and the description need further development. The OSPAR common indicators for non-indigenous species (NIS) do not provide information to support decisions on bioinvasion management options other than prevention. These include control, confinement, and mitigation of invasive species (Lodge at al., 2006; Olenin et al., 2011). It may be that country-specific indicators are more relevant to this issue than common indicators. The importance of different pathways and vectors differ between countries and this should be considered when deciding on appropriate common indicators (and in designing sampling, analysis, and assessments). An analysis should be carried out, taking into account the importance of different pathways and associated vectors for each country (see Olenin et al., 2010, 2011). Sampling methodology should be designed in relation to the organism groups to be monitored. It is unclear from the technical description for which organism groups/taxa/species monitoring will be planned, and therefore it is at this stage impossible to review the proposed methodology. In general, a sampling frequency of once per year is insufficient to obtain representative information for several organism groups, and is particularly poor for planktonic organisms with a reproduction cycle merely ranging a few days. The proposed site selection of two sites per country is likely to be insufficient. The site selection should be country-specific and rather depend on the analysis of the presence and importance of different pathways/vectors. ICES further suggests that the choice of monitoring location be based on likely entry points into regional seas. To assess impacts caused by NIS, distribution, abundance, and biomass data are also needed. For this reason also, more than two sites will need to be monitored. A possible way to decide which invasion pathways and habitats should be monitored is preparing a pathway/habitat matrix and leave the selection to individual countries based on their priorities and capabilities. Species identification is likely to pose serious problems, particularly for smaller organisms such as the microzooplankton or meio-benthos. The target setting should be redefined; a three-year period is likely to be too short, a longer assessment period is more appropriate (e.g. six years, as stated in the MSFD Directive (EU, 2008)). Sources EU. 2008. Directive of the European Parliament and the Council Establishing a Framework for Community Action in the Field of Marine Environmental Policy (Marine Strategy Framework Directive). European Commission. Directive 2008/56/EC, OJ L 164. ICES. 2013a. Report of the Working Group on the Ecosystem Effects of Fishing Activities (WGECO), 1–8 May 2013, Copenhagen, Denmark. ICES CM 2013/ACOM:25. 86 pp. ICES. 2013b. Report of the Working Group on Biodiversity Science (WGBIODIV). ICES CM 2013/SSGEF:02. 62 pp. ICES. 2013c. Report of the ICES Working Group on the Introduction and Transfers of Marine Organisms (WGITMO). ICES CM 2013/ACOM:30. 150 pp. ICES. 2013d. Report of the Working Group on Marine Habitat Mapping (WGMHM). In draft. ICES. 2013e. Report of the Working Group on Marine Mammal Ecology (WGMME). ICES CM 2013/ACOM:26. Lodge, D. M., Williams, S., MacIsaac, H. J., Hayes, K. R., et al. 2006. Biological invasions: recommendations for US policy and management. Ecological Applications, 16: 2035–2054. Olenin, S., Alemany, F., Cardoso, A. C., Gollasch, S., Goulletquer, P., Lehtiniemi, M., et al. 2010. Marine Strategy Framework Directive – Task Group 2 Report. Non-indigenous species. European Communities. EUR 24342 EN. ISBN 978-92-79-15655-7. ISSN 1018-5593. DOI 10.2788/87092. Luxembourg: Office for Official Publications of the European Communities. 44 pp. Olenin, S., Elliott, M., Bysveen, I., Culverhouse, P., Daunys, D., Dubelaar, G. B. J., et al. 2011. Recommendations on methods for the detection and control of biological pollution in marine coastal waters. Marine Pollution Bulletin, 62 (12): 2598–2604. OSPAR. 2013a. Report by ICG–COBAM on the development of an OSPAR common set of biodiversity indicators: Parts A and B. BDC 13/4/2-E. 41 pp. OSPAR. 2013b. Report by ICG–COBAM on the development of an OSPAR common set of biodiversity indicators: Part C: Technical Specifications. BDC 13/4/2 Add.1-E. 156 pp. Piet, G. J., Quirijns, F., Robinson, L., and Greenstreet, S. P. R. 2007. Potential pressure indicators for fishing and their data requirements. ICES Journal of Marine Science, 64: 110–121. ICES Advice 2013, Book 1 221 1.5.6.4 Special request, Advice June 2013 ECOREGION General advice SUBJECT OSPAR special request on maximizing the use of available sources of data for monitoring of biodiversity Advice summary ICES advises that two of the major annual coordinated surveys, the International Bottom Trawl Survey and the Beam Trawl Survey, used to provide information to support fish stock assessments, could also be used to provide information for several Marine Strategy Framework Directive (MSFD) indicators. In these two surveys, a small amount of additional sampling effort could deliver a much broader set of data. ICES advises that an ecosystem monitoring programme should be developed. Such a programme should be designed not only to observe status, but also to understand the processes underlying these observations, and the links between ecosystem components and with the physical environment. A further six existing surveys have the potential to collect appropriate data for nine of the eleven descriptors. Request The purpose of this request is to seek ICES advice on the potential sources of data and information that may be available to support the monitoring and assessment of biodiversity in relation to commitments under MSFD so as to maximise efficiencies in the use of available resources, for example: - - where efficiencies could be made to identify where there are monitoring programmes or data sources that can deliver multiple indicators, which may relate to different Descriptors, (e.g. The Data Collection Framework could be used to implement D3 and D1 indicators), or where with a small additional effort existing monitoring could be amplified to deliver a broader set of data. Advice is sought as to 1) the quality of these potential data sources and how they could be used, including but not limited to the relevance of outcomes identified in chapter 8 of the ICES MSFD D3+ report to Descriptors 1, 4 and 6. (OSPAR request 4/2013) ICES advice Scientific surveys Two major ICES coordinated annual surveys could, with relatively minor additional effort, be used to better inform the assessment of the state of multiple OSPAR indicators, relevant to several MSFD descriptors. For the International Bottom Trawl Survey, the main areas where data are available already are the Fish and Cephalopod indicators FC-1 to FC-8 18, with the exception of indicator 4 (on bycatch). Additional data products would be available contingent on a number of procedural developments, including the development of swept area estimation procedures and appropriate maturity estimation keys. Data support could be provided for Foodweb indicators (FW-4 and FW-7). FW-8 could be supported in terms of stomach sampling. For Bird indicators, the surveys could provide seabird data for indicator B-6, but it is suggested that this is more appropriate for other survey types (e.g. acoustic and icthyoplankton). For the Beam Trawl Survey, the conclusions in terms of the FC indicators are broadly similar, although swept area estimates are much more straightforward with this gear, and could be improved with use of covariates. No data for FC-6 is obtainable due to survey timing, but improvements could be made with new maturity keys and at-sea histological sampling. Biomass and abundance estimates of taxa that can be caught by the survey could support FW-4, FW-7, and FW-8. Again, the surveys could provide seabird data for indicator B-6, but it is suggested that this is more appropriate for other survey types (e.g. acoustic and icthyoplankton). 18 222 FC, FW, M, B, BH, PH, and NIS refer to the ICG–COBAM indicator numbering. ICES Advice 2013, Book 1 The combined response table to the OSPAR request for the two surveys is presented in Table1.5.5.2.1 (data availability) and Table 1.5.5.2.2 (possible improvements). ICES Advice 2013, Book 1 223 Table 1.5.5.2.1 EU Indicator ID 1.2.1 Possible contributions of the ICES International Bottom Trawl Surveys (IBTS) and Beam Trawl Surveys (BTS) to reporting under the MSFD, with regard to biodiversity-related indicators. The selected indicators are based on nomenclature in the EU Decision (COM 477/2010) and matching OSPAR indicator ID (2nd column). IBTS applies only to the North Sea and Northeast Atlantic; BTS is conducted in all areas except the Northeast Atlantic. OSPAR Indicator ID FC-1 Core Name Population abundance/ biomass of a suite of selected species. Survey IBTS BTS 4.2.1 3.3.2 N.A. (related to 4.3.1) FC-2; FW-3 Core FC-3 Core FC-5 Candidate OSPAR EcoQO for proportion of large fish (LFI). IBTS BTS Mean maximum length IBTS of demersal fish and elasmobranchs. BTS Conservation status of elasmobranch and demersal bony-fish species (IUCN). IBTS BTS 1.3.1; 3.3.1 224 FC-6 Candidate Proportion of mature fish in the populations of all species sampled adequately in international and national fish surveys. IBTS North Sea Northeast Atlantic Abundance estimates (per hour and per km2) of various fish species. Accuracy is species-dependent. Abundance (per km2) estimates for various fish species can be supplied. Accuracy is speciesdependent. Abundance estimates (per hour and per km2) of various fish species. Accuracy is species-dependent. Main source of data for this indicator. Main source of data for this indicator. Cut-off point and reference limit needs to be defined by the survey. Main source of data for this indicator. Main source of data for this indicator Abundance estimates (per hour and per km2) of various fish species. Accuracy is species-dependent. Abundance (per km2) estimates for various fish species can be supplied. Accuracy is speciesdependent. Main source of data for this indicator. Main source of data for IBTS target species, but depending on species-specific maturation process and hence sampling time (quarter). Data availability Western shelf France/Biscay Inshore Abundance (per km2) estimates for various fish species can be supplied. Accuracy is speciesdependent. Abundance (per km2) estimates for various fish species can be supplied. Accuracy is speciesdependent. The area covered is spatially restricted but will give additional information not available from other survey sources. Abundance (per km2) estimates for various fish species can be supplied. Accuracy is speciesdependent. Main source of data for this indicator. Cut-off point and reference limit needs to be defined by the survey. Main source of data for this indicator. Cut-off point and reference limit needs to be defined by the survey. Main source of data for this indicator. Cut-off point and reference limit needs to be defined by the survey. Main source of data for this indicator Main source of data for this indicator Main source of data for this indicator Abundance (per km2) estimates for various fish species can be supplied. Accuracy is speciesdependent. Abundance (per km2) estimates for various fish species can be supplied. Accuracy is speciesdependent. The area covered is spatially restricted but will give additional information not available from other survey sources. Abundance (per km2) estimates for various fish species can be supplied. Accuracy is speciesdependent. Main source of data for this indicator. Abundance estimates (per hour and per km2) of various fish species. Accuracy is species-dependent. Main source of data for IBTS target species, but depending on species-specific maturation process and hence sampling time (quarter). ICES Advice 2013, Book 1 EU Indicator ID 1.1.1 1.1.2 possibly related to 1.7.1 or 4.3.1 1.7.1; 4.3.1 OSPAR Indicator ID FC-7 Candidate FC-8 Candidate FW-4 Core FW-7 Candidate Name Survey North Sea Distributional range of a suite of selected species. IBTS Distributional pattern within the range of a suite of selected species. Changes in average trophic level of marine predators. IBTS Fish biomass and abundance of dietary functional groups. IBTS BTS BTS IBTS BTS BTS could be related to 4.2.1; 4.3.1 FW-8 Candidate Changes in average faunal biomass per trophic level (biomass trophic spectrum). ICES Advice 2013, Book 1 IBTS BTS Main source of data for this indicator. Main source of data for this indicator. Main source of data for this indicator. Main source of data for this indicator. Calculation of relative abundance is possible. Calculation of relative abundance is possible. Biomass and abundance estimates (per hour or per km2) of various fish species. Biomass and abundance estimates per km2 of various fish species. Data on biomass per haul for all fish species. Data on biomass per haul for fish species and benthic organisms available for some surveys and some years. Northeast Atlantic Data availability Western shelf France/Biscay Inshore Main source of data for this indicator. Main source of data for this indicator. Main source of data for this indicator. Main source of data for this indicator. Main source of data for this indicator. Main source of data for this indicator. Main source of data for this indicator. Calculation of relative abundance is possible. Calculation of relative abundance is possible. Calculation of relative abundance is possible. Biomass and abundance estimates per km2 of various fish species. Biomass and abundance estimates per km2 of various fish species. Biomass and abundance estimates per km2 of various fish species. Main source of data for this indicator. Calculation of relative abundance is possible. Biomass and abundance estimates (per hour or per km2) of various fish species Data on biomass per haul for all fish species. Data on biomass per haul for fish species and benthic organisms available for some surveys and some years. Data on biomass per haul for fish species available. Epibenthic biomass available for some surveys. 225 Table 1.5.5.2.2 EU Indicator ID 1.2.1 Possible improvements to the ICES International Bottom Trawl Surveys (IBTS) and Beam Trawl Surveys (BTS) in reporting under the MSFD, with regard to biodiversity-related indicators. The selected indicators are based on nomenclature in the EU Decision (COM 477/2010) and matching OSPAR indicator ID (2nd column). IBTS applies only to the North Sea and Northeast Atlantic; BTS is conducted in all areas except the Northeast Atlantic. The possible improvement of data availability in each of the survey areas, if extra effort is allocated to these surveys, is indicated. OSPAR Indicator ID FC-1 Core Name Population abundance/ biomass of a suite of selected species. Survey IBTS BTS 1.3.1; 3.3.1 Poss. related to 1.7.1 or 4.3.1 FC-6 Candidate FW-4 Core 226 For some species that are presently not always reported to species level (e.g. squids, gobies), species could be collected for taxonomic ID on shore. Precision could be improved through further development of sampling and statistical techniques. For some species that are presently not always reported to species level (e.g. squids, gobies), species could be collected for taxonomic ID on shore. For additional species theoretically possible, but requires extra resources for acquisition of maturity data. Guidelines needed for maturity keys / spawning times. Precision could be improved by further development of analysis of maturity. Changes in average trophic level of marine predators. Samples for fish predators can be provided on a regular basis (using stomach analyses or tissue samples for stable isotope analysis); sample processing requires extra analytical effort. Samples for fish predators can be provided (for stomach analyses or tissue samples for stable isotope analysis); sample processing requires extra analytical effort. BTS 1.7.1; 4.3.1 FW-7 Candidate Northeast Atlantic Proportion of IBTS mature fish in the populations of all species sampled adequately in international and BTS national fish surveys. IBTS Fish biomass and IBTS abundance of dietary functional groups. BTS Possible improvement with extra effort Western shelf North Sea Individual fish weights of nontarget species could be provided with extra effort. Individual fish weights of nontarget species could be provided with extra effort. France/ Biscay Inshore Precision could be improved through further development of sampling and statistical techniques. Precision could be improved through further development of sampling and statistical techniques. Precision could be improved through further development of sampling and statistical techniques. Precision could be improved by further development of analysis of maturity. Precision could be improved by further development of analysis of maturity. Samples for fish predators can be provided (for stomach analyses or tissue samples for stable isotope analysis); sample processing requires extra analytical effort. Samples for fish predators can be provided (for stomach analyses or tissue samples for stable isotope analysis); sample processing requires extra analytical effort. Samples for fish predators can be provided (for stomach analyses or tissue samples for stable isotope analysis); sample processing requires extra analytical effort. Individual fish weights of non-target species could be provided with extra effort. Individual fish weights of non-target species could be provided with extra effort. Individual fish weights of non-target species could be provided with extra effort. ICES Advice 2013, Book 1 EU OSPAR Indicator ID Indicator ID could be FW-8 related to Candidate 4.2.1; 4.3.1 1.1.2 B-6 Core ICES Advice 2013, Book 1 Name Survey North Sea Changes in average faunal biomass per trophic level (biomass trophic spectrum). Distributional pattern of breeding and non-breeding marine birds. IBTS BTS IBTS BTS Full benthic sort and sampling possible with extra resources. Northeast Atlantic Possible improvement with extra effort Western shelf Full benthic sort and sampling possible with extra resources. France/ Biscay Inshore Full benthic sort and sampling possible with extra resources. Full benthic sort and sampling possible with extra resources. Some vessels may be able to take bird observers aboard. Some vessels may be able to take bird observers aboard. 227 A further four surveys can provide information suitable for use in evaluating ecosystem indicators; one of these is outside the area of EU waters: • Norwegian Barents Sea survey – trawl, acoustic, and ecosystem survey. • French Biscay Pelagic ecosystem survey – acoustic. • German mackerel egg survey – icthyoplankton. • Scottish Nephrops TV survey. ICES analysed the strengths and weaknesses of these surveys and two key themes emerged: • • setting and prioritizing objectives; survey design and the need to be able to elucidate process by explicitly linking dynamics in different ecosystem components. Some of the strengths were mutually exclusive, either operationally or conceptually, and therefore an ‘ideal ecosystem survey’ on a single vessel, is unlikely to exist. The prioritization of surveys might be based on three factors: • • the characteristics of the ecosystem, particularly with respect to the spatial and temporal scales of variability. the available resources in ship time, but also expertise and financial considerations. International pooling of resources will aid to increase efficiency and improve regional ecosystem assessments across national boundaries. • the management and legal requirements and prioritizations for reporting. This is not a scientific criterion, but an ability to address the former will almost certainly have an impact on the availability of resources. Acoustic surveys may represent the best option for collecting data to evaluate the Pelagic Habitat (PH) indicators, and also some Benthic Habitat (BH) indicators by acoustic means including multi-beam technology. TV surveys can support BH-1, BH-3, BH-4, and FW-9. Icthyoplankton surveys provide potential to help evaluate both pelagic habitat and foodweb indicators. These surveys are also better platforms for the collection of data on birds and mammals at sea compared with slower trawl surveys. Integrated monitoring There are considerable benefits to designing and developing a cost-effective integrated monitoring programme that can address the existing information needs and those of MSFD in comparison with current discipline-specific monitoring programmes. The former may provide advice based on an understanding of ecosystem processes, which can help in identifying how to act rather than merely identifying current status in separate parts of the ecosystem. As none of the individual surveys can provide all the information that would be required to service the full suite of OSPAR common indicators, it makes the concept of clustering indicators rather difficult to put into practice. Perhaps the most effective cruise, if it were possible to organise, would be a bottom trawl survey where, in addition to the standard fish sampling: • • • • • • • Stomach and biological samples of a wide range of taxa were collected; Seabed habitat acoustic survey data were collected between fishing stations; Seabirds and marine mammals at sea were surveyed between fishing stations; A continuous plankton recorder was deployed while the vessel was underway between stations; Hydrographic data were collected continuously by on-board autonomous samplers; CTD data and water and plankton samples were collected at each sampling station; The night-time period was utilized to sample benthic invertebrates. Such a survey would collect data that could potentially service 65% of the OSPAR common indicators (M-2, M-4, B-1, B-6, FC-1, FC-2, FC-3, FC4, BH-1, BH-2, BH-5, PH-1, PH-2, PH-3, FW-2, FW-3, FW-4, FW-5, FW-6, FW-7, FW-8, FW-9, and NIS-2). 228 ICES Advice 2013, Book 1 Background This advice has focussed on research vessel surveys as potential sources of data. Other potential data sources include: • • • • • • Commercial landings data and logbooks. Commercial discard data (that will cease when discard bans come into force). Shore-based sampling, particularly for benthos, contaminants, hydrographic parameters and biological oceanography. Remote sensing i.e. satellites for e.g. sea surface temperature, ocean colour, sea surface elevation, waves, etc. Sea-going sampling systems such as continuous plankton recorders on platforms of opportunity, underwater cameras, etc. Aerial surveys (marine mammals, sea turtles, seabirds, sharks). Several ICES expert groups have been developing analyses of the opportunities for wider usage of fisheries research surveys (ICES, 2010a, 2010b, 2012a, 2012b, 2013a, 2013b, 2013c, 2013d). The request also refers to the Data Collection Framework (DCF) indicators used to evaluate the ecosystem impacts of fishing. Four of these in particular are relevant to MSFD indicators: • Conservation status of fish species (FC-5). • Proportion of large fish (FC-2, FW-3). • Mean maximum length of fishes (FC-3). • Size-at-maturation of exploited fish species (FC-6). ICES has recently advised the European Commission on this topic (ICES, 2013e). Table 1.5.5.2.3 summarizes some suggestions on how surveys may be adapted to fulfil the needs of OSPAR. . ICES Advice 2013, Book 1 229 ICES COORDINATED SURVEYS Fi s h s urveys us i ng nets (BTS, IBTS, etc) Acous ti c fi s h s urveys Vi deo s urveys (Nephrops ) Ichthyopl a nkton s urveys (fi s h l a rva e) 230 MAMMALS A A A A BIRDS A A FISH & CEPH E E E E BENTHIC HAB. PEL. HAB. A A A A A A A A A A A A A A A A A A A A A E A E A A A A A NIS-2 (Rate of new introductions of NIS) NIS-1 (pathways management measures) FW-9 (Ecological Network Analysis) FW-8 (Biomass Trophic Spectrum) FOOD WEBS E A A A A A FW-7 (fish biomass and dietary functional groups) FW-6 (zooplankton) FW-5 (change funct groups plankton) FW-4 (change in average trophic level of predators) FW-3 (Large Fish Indicator) FW-2 (production of phytoplankton) FW-1 (repr success in relation to food avail) PH-3 (changes biodiv index) PH-2 (biomass/abundance) PH-1 (changes functional types) BH-5 (size-freq sensitive species) BH-4 (area habitat loss) BH-3 (damage) BH-2 (multimetric indices) BH-1 (typical species) FC-4 (bycatch Chondrichthes) FC-3 (mean max length) FC-2 (LFI) FC-1 (abundance/biomass) B-6 (distribution) B-5 (bycatch) B-4 (mammals on seabird colonies) B-3 (breeding success/failure marine birds) B-2 (breeding success kitiwake) B-1 (trends in species) M-6 (bycatch) M-5 (seal pup production) M-4 (abundance cetaceans) M-3 (abundance grey & harbour seals) M-2 (range pattern cetaceans) M-1 (range pattern grey & harbour seals) Table 1.5.5.2.3 Summary of the OSPAR “common indicators” that could potentially be serviced using data derived from the additional (A) ecological sampling that might be feasible during different types of fisheries surveys. E = Existing data collection. Candidate indicators FC-5 to FC-8 are not shown as no technical specifications for these have been supplied to ICES. It may be that some surveys could help in data collection against these indicators. ALIENS A A A ICES Advice 2013, Book 1 Sources ICES. 2010a. Report of the Working Group on Integrating Surveys for the Ecosystem Approach (WGISUR), 26–28 January 2011, Dublin, Ireland. ICES CM 2010/SSGESST:08. 17 pp. ICES. 2010b. Report of the Workshop on Cataloguing Data Requirements from Surveys for the EAFM (WKCATDAT), 26–28 January 2011, Dublin, Ireland. ICES CM 2010/SSGESST:09. 38 pp. ICES. 2012a. Report of the Working Group on Integrating Surveys for the Ecosystem Approach (WGISUR), 24–26 January 2012, Ijmuiden, the Netherlands. ICES CM 2012/SSGESST:20. 24 pp. ICES. 2012b. Report of the Workshop on Evaluation of Current Ecosystem Surveys (WKECES), 20–22 November 2012, Bergen, Norway. ICES CM 2012/SSGESST:23. 59 pp. ICES. 2013a. Report of the International Bottom Trawl Survey Working Group (IBTSWG), 8–12 April 2013, Lisbon, Portugal. ICES CM 2013/SSGESST:10. ICES. 2013b. Report of the Working Group on Beam Trawl Surveys (WGBEAM), 23–26 April 2013, Ancona, Italy. ICES CM 2013/SSGESST:12. ICES. 2013c. Report of the Working Group on the Ecosystem Effects of Fishing Activities (WGECO), 1–8 May 2013, Copenhagen, Denmark. ICES CM 2013/ACOM:25. 86 pp. ICES. 2013d. Report of the Working Group on Biodiversity Science (WGBIODIV). ICES CM 2013/SSGEF:02. 62 pp. ICES. 2013e. Report of the ICES Advisory Committee. ICES Advice, 2013. Book 1, Section 1.5.2.1. ICES. 2013f. Report of the ICES Working Group on the Introduction and Transfers of Marine Organisms (WGITMO). ICES CM 2013/ACOM:30. 150 pp. ICES. 2013g. Report of the Working Group on Marine Habitat Mapping (WGMHM). In draft. Supplementary background documentation is available at: https://groupnet.ices.dk/wgeco2013/default.aspx?RootFolder=%2fwgeco2013%2fReport%202013%2fToR%20C&Fold erCTID=&View=%7b63F03C99%2d9F39%2d457D%2d8135%2d25A0BEA67AAF%7d ICES Advice 2013, Book 1 231 1.5.6.5 Special request, Advice June 2013 ECOREGION SUBJECT General advice OSPAR/NEAFC special request on review of the results of the Joint OSPAR/NEAFC/CBD Workshop on Ecologically and Biologically Significant Areas (EBSAs) Advice summary ICES reviewed the ecological evidence supporting the ten proposed ecologically and biologically significant areas (EBSAs) from the OSPAR/NEAFC/CBD Workshop of September 2011, as presented in the annexes to that report. The review applied standard ICES practices and used primarily the references cited in the relevant annexes, but augmented those references with other publications and data sources. In nine of the ten proposed EBSAs, ICES came to different conclusions than were contained in the OSPAR/NEAFC/CBD Workshop report, with regard to the rankings of the Convention on Biological Diversity (CBD) EBSA criteria. Of the ten proposed EBSAs, ICES supports the conclusion of the OSPAR/NEAFC/CBD workshop that the Arctic Ice area (Area 10) meets one or more EBSA criteria and this area could go forward at this time, possibly with minor suggested changes to the rationale. In four proposed EBSAs (Reykjanes Ridge south of Iceland EEZ (Area 1); Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge (Area 2); Mid-Atlantic Ridge north of the Azores (Area 3); the Hatton and Rockall banks and Hatton–Rockall Basin (Area 4)), ICES considers that much of the area within the proposed EBSAs do not meet any of the EBSA criteria and for this reason the boundaries of these proposals need to be revised. More restricted parts of the proposed EBSAs meet several of the EBSA criteria and could go forward after boundary revision. ICES notes great similarities in the pro forma describing Areas 1 and 3 and part of Area 2 (OSPAR/NEAFC/CBD, 2011). A boundary revision to encompass the relevant parts of these areas as a single extended Mid-Atlantic Ridge proposed EBSA could be considered a step forwards. ICES recommends changes also to the pro forma rankings for all of these proposed EBSAs. Only a small part of the proposed EBSA for the Arctic Front – Greenland/Norwegian seas (Area 9) possibly meets some of the EBSA criteria. However, another part of the general area might meet some of the EBSA criteria. ICES recommends that further data analyses followed by an evaluation of the new results against the EBSA criteria be undertaken before any further decision is taken. The rationales for four proposed EBSAs (around the Pedro Nunes and Hugo de Lacerda seamounts (Area 5); Northeast Azores–Biscay Rise (Area 6); Evlanov Seamount region (Area 7); and Northwest of Azores EEZ (Area 8)) are not well supported by the information presented in the relevant annexes. There is a need for further data and analyses in these areas, particularly in relation to seabirds, and another evaluation of the areas against the EBSA criteria. ICES found no clear evidence of additional EBSAs in areas beyond national jurisdiction (ABNJ) of the Northeast Atlantic meeting the CBD scientific criteria. Request a) Review the description of areas meeting one or more of the CBD EBSA scientific criteria developed as an outcome of the Joint OSPAR/NEAFC/CBD Scientific Workshop, in particular: 1. Review each of the ten area delineations and descriptions in line with the CBD EBSA Scientific criteria and the most up-to-date scientific data and information, specifying any additional scientific data and information that is available; 2. Provide, if appropriate, revised EBSA proposals in the format of proformas adopted by the CBD b) If there is clear evidence for additional areas in ABNJ of the North-East Atlantic meeting the CBD EBSA scientific criteria, present a description with supporting scientific data and information for such areas, including CBD EBSA proformas for each. ICES advice ICES made the following conclusions and proposals. 232 ICES Advice 2013, Book 1 Area 1. Reykjanes Ridge south of Iceland EEZ: Much of the area in the proposed EBSA does not meet any of the EBSA criteria. A more restricted area down the spine of the Mid-Atlantic Ridge and defined by depth ranges of deep-water coral and sponge concentrations does meet several EBSA criteria and the boundary delineation, ranking, and full rationale could be developed based on this new boundary. Area 2. Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge: Some areas in the proposed EBSA do not meet any of the EBSA criteria. A complex combination of the area down the spine of the Mid-Atlantic Ridge, the benthic area aligned with and close to the main fractures, and the water column in which the Subpolar Front is found throughout the year, does meet several EBSA criteria and the boundary delineation, ranking and full rationale could be developed at based on this new boundary. Area 3. Mid-Atlantic Ridge north of the Azores: Much of the area in the proposed EBSA does not meet any of the EBSA criteria. A more restricted area down the spine of the Mid-Atlantic Ridge and defined by depth ranges of deep-water coral and sponge concentrations does meet several EBSA criteria and the boundary delineation, ranking, and full rationale could be developed based on this new boundary. Area 4. The Hatton and Rockall banks and Hatton–Rockall Basin: Much of the area in the proposed EBSA does not meet any of the EBSA criteria. A more restricted area down to approximately 1500–1800 m depth, but excluding the abyssal plain does meet several EBSA criteria and the boundary delineation, ranking, and full rationale could be developed based on this new boundary. Area 5. Around the Pedro Nunes and Hugo de Lacerda seamounts: The proposed EBSA is not supported well by the information presented in the pro forma. There is a need for further analyses of those data already considered, as well as any additional relevant data on seabird foraging and other information. When these analyses are done, including for the additional data, another evaluation of the area against the CBD EBSA criteria would make it possible to advise which areas, if any, meet EBSA criteria. Area 6. Northeast Azores–Biscay Rise: The proposed EBSA is not supported well by the information presented in the pro forma. There is a need for further analyses of those data already considered as well as any additional relevant data on seabird foraging and other information. When these analyses are done, including for the additional data, another evaluation of the area against the CBD EBSA criteria would make it possible to advise which areas, if any, meet EBSA criteria. Area 7. Evlanov Seamount region: The proposed EBSA is not supported well by the information presented in the pro forma. There is a need for further analyses of those data already considered as well as any additional relevant data on seabird foraging and other information. When these analyses are done, including for the additional data, another evaluation of the area against the CBD EBSA criteria would make it possible to advise which areas, if any, meet EBSA criteria. Area 8. Northwest of Azores EEZ: The proposed EBSA is not supported well by the information presented in the pro forma. There is a need for further analyses of those data already considered as well as any additional relevant data on seabird foraging and other information. When these analyses are done, including for the additional data, another evaluation of the area against the CBD EBSA criteria would make it possible to advise which areas, if any, meet EBSA criteria. Area 9. The Arctic Front – Greenland/Norwegian seas: Only a small part of the area proposed by the OSPAR/NEAFC/CBD Workshop as an EBSA was considered to possibly meet some of the criteria. However, another part of the general area might meet some EBSA criteria. There is a need for more analyses of productivity and diversity data for the more southerly part of the main area, and then a re-evaluation of the new results against the EBSA criteria, before any areas might be considered as possibly meeting EBSA criteria. Area 10. The Arctic Ice: The rationale for concluding that this area meets one or more EBSA criteria can be improved, but the review by ICES generally supports the conclusions of the OSPAR/NEAFC/CBD workshop. A suggested revised proforma is attached as Annex 1.5.6.5.1 to this advice. With regard to new proposed areas that meet EBSA criteria, ICES has no additional information. However, ICES suggests a potential alternative configuration to the areas in proposed EBSAs 1, 2, and 3 that would comprise two areas meeting EBSA criteria, one covering the specified depths of the entire Mid-Atlantic Ridge, and one for the Charlie-Gibbs Fracture Zone and pelagic area of the Subpolar Front. Each of the two areas would have coherent, but different ecological rationales. ICES has provided its rankings for the revised proposed EBSAs and the rationale for those rankings. ICES has not revised the narrative or the references in the existing pro forma of proposed EBSAs other than for Arctic Ice habitat (Annex ICES Advice 2013, Book 1 233 1.5.6.5.1). Once OSPAR and NEAFC have made a decision on the configuration of the Mid-Atlantic Ridge proposed EBSAs, ICES could revise the pro forma for these EBSAs by the end of September 2013. Background Method ICES conducted its review informed by the content of the CBD Decisions IX/20 and X/29 on Marine and Coastal Biodiversity (CBD, 2008, 2010), the reports from the ‘Azores Workshop’ (CBD, 2007) and the ‘Ottawa Workshop’ (CBD, 2009), and the UNGA Resolution 58/240 (United Nations, 2004). ICES considers that the application of the criteria was intended to be a comparative or relative process, such that areas should be evaluated against other generally comparable areas (e.g. of comparable depth and latitude). In addition, even though the application of EBSA criteria is not guided directly by management considerations, potential benefits of spatial management measures are a relevant consideration in the evaluation. However, the appropriate baseline is not the absence of all management, but the presence of measures sufficient to ensure human uses are sustainable in areas typical of the zone of evaluation. ICES is responding to a request about EBSAs, and ICES stresses that this advice does not imply that any areas reviewed should or should not be considered as VMEs (but ICES notes that there is an overlap with advice provided recently on vulnerable marine ecosystems (VMEs; ICES, 2013a)). ICES notes that all areas found to meet criteria for VMEs would be expected to meet one or more criteria for EBSAs as well. However, the reverse is not necessarily true and EBSAs do not necessarily contain VMEs. There is neither a policy nor an ecological rationale for automatically excluding bottom fishing (or any other activity) from areas proposed as EBSAs. The expected initial response of regulatory authorities is to conduct risk or threat assessments of the activities they regulate relative to the properties considered ecologically or biologically significant, and to subsequently undertake management appropriate to the outcome of these assessments. ICES advice is based on applying several standards during its review, including: • • • • • Assigned rankings on a criterion should apply to at least most of the area included in a proposed EBSA, and not just to a small subset of the total area. Higher assigned rankings required the proposed area to differ from adjacent areas and other areas of similar depth and latitude on the property represented by the criterion. Some evidence must be available to justify awarding a higher ranking, noting that comprehensive data for the high seas cannot be expected. Rankings should not be based on the presence or history of threats to the features represented by the criteria, but on the biological, ecological, and geomorphological features of the area. Although EBSAs are not defined by or linked to any particular management actions by any authorities, it is appropriate to consider whether or not spatial management tools might benefit the conservation or sustainable use of the relevant features. For each of the ten areas in the OSPAR/NEAFC/CBD report proposed to meet EBSA criteria, ICES assigned one of three categories: • • Proceed with boundaries proposed by the OSPAR/NEAFC/CBD Workshop, with a rationale revised by ICES. Proceed with developing a proposed EBSA with different boundaries than those proposed by the OSPAR/NEAFC/CBD Workshop, and with a rationale provided by ICES. • Do not proceed with proposing an EBSA at this time, but rather undertake further collation and analysis of information and reconsider when the additional work is completed. Although ICES review included an evaluation and commentary on all pro forma contents of the OSPAR/NEAFC/CBD 2011 Workshop report, this advice presents only ICES conclusions as to whether the areas meet the EBSA criteria. This advice takes the form of biological properties that ICES concludes would define the boundaries, rankings, and rationales of those areas against the EBSA criteria. Proposed EBSAs that need minor revisions to the rationale Area 10. The Arctic Ice A suggested revised pro forma is attached to this advice as Annex 1.5.6.5.1. Proposed EBSAs that should have redefined boundaries before proceeding Area 1. Reykjanes Ridge south of Iceland EEZ 234 ICES Advice 2013, Book 1 ICES advises that an area along the Reykjanes Ridge can be justified as meeting one or more EBSA criteria. This would be a much smaller area than the one proposed in the OSPAR/NEAFC/CBD Workshop report. Appropriate boundaries for such a proposed area that meets EBSA criteria would be a depth contour that runs in the deeper of two properties: 3. 4. Including a large portion (arbitrarily, perhaps 90%) of all hard volcanic substrates; habitats are reported to host the larger known coral formations and their associated communities Including a large portion (arbitrarily, 90%) of the records of large sponge communities in the overall northern Mid-Atlantic Ridge. In addition, the proposed area will include the only known hydrothermal vent in the area (Olafsson et al., 1991; German et al., 1994; German and Parsons, 1998; Mironov and Gebruk, 2007), whatever depth contour is used. Information on water masses should also be consulted, allowing proper identification of benthic and fish fauna to be included in a revised narrative to a pro forma for this area. In the time available ICES did not have the geological data to delineate the depth contour that would meet the first criterion, but such information should be readily available in marine geology databases. Nor did ICES have access to all of the records of where the large sponge deposits were taken. However, references to sources for those data are in the OSPAR/NEAFC/CBD Workshop pro forma and should be tracked back to find the appropriate depth contour for the second criterion in the tables below. Evaluation of the revised area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking Uniqueness rarity or Low Some High X (X) The bracketed “High” ranking is for a very restricted area covering the only known hydrothermal vent on the Reykjanes Ridge. Though this is a unique feature it occupies only a small part of the area proposed here as meeting this EBSA criterion (Olafsson et al., 1991; German et al., 1994; German and Parsons, 1998; Mironov and Gebruk, 2006). The MarEco sampling of corals and sponges reported a few species new to science and these may be restricted to the proposed area, although a firm conclusion on this cannot be drawn until more extensive sampling is undertaken. Areas that are required for a Special X importance for population to survive and thrive. life-history stages of species Explanation for ranking Although many populations undoubtedly complete their life cycles within the large area proposed in the OSPAR/NEAFC/CBD Workshop Report as an EBSA, this would be true of any marine area of comparable size. There is no evidence that the life history of any species is strongly dependent on any specific features of the area proposed as an EBSA. There is evidence from other areas of the Northeast Atlantic that areas of high coral density may be important as eggcase and nursery areas of deep-water sharks and rays. The area proposed here is targeted on the depths and substrates associated with higher coral and sponge densities, and if sharks and rays also concentrate spawning and early development in these habitats, then the score would be “Some” or “High” on this criterion as well. Importance for Areas containing habitats for the X survival and recovery of threatened, threatened, or endangered, or endangered, declining species, or areas with ICES Advice 2013, Book 1 235 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High declining species significant assemblages of such species. and/or habitats Explanation for ranking Large formations of corals and sponges are found in the area proposed by ICES as meeting this criterion. Habitats containing these species are listed by OSPAR and also feature as VME indicator species; the majority of these would be included in the proposed EBSA. Additional explanation regarding corals and sponges is included in the rationale for the criterion on Vulnerability. The possible role of the area proposed by ICES in the life histories of sharks and rays is discussed in the criterion on Vulnerability and Sensitivity. Areas that contain a relatively high Vulnerability, X proportion of sensitive habitats, fragility, sensitivity, or biotopes, or species that are functionally fragile (highly slow recovery susceptible to degradation or depletion by human activity or by natural events) or with slow recovery. 236 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High Explanation for ranking With regard to corals and sponges, Mortensen et al. (2008) found cold-water corals “at every sample station … observed at depths between 800 and 2400 m, however were commonly found shallower than 1400 m …, with species richness being very high. … no major reef structures were recorded, with the maximum colony size approximately 0.5 m in diameter. The number of coral taxa was strongly correlated with the percentage cover of hard bottom substrate ….” The area proposed here is targeted at the seamount peaks and slopes where hard substrates dominate. For sponges, no actual large expanses of sponge reef were reported in the OSPAR/NEAFC/CBD Workshop report. However, the pro forma in that report (OSPAR/NEAFC/CBD, 2011) notes that overall sampling of the area is patchy and cites three studies that found local patches with high densities of sponges, although in no cases were the sizes of the patches documented. These observations of widespread occurrences of corals and sponges are supported by the records in the GRID–Arendal data, which show both taxa to be presented in nearly every appropriate sample taken along the cruise tracks in the database. Biological productivity Areas containing species, populations, or communities with comparatively higher natural biological productivity. X Explanation for ranking Although benthic productivity of the proposed smaller EBSA may be higher than benthic productivity on the abyssal plain, productivity integrated over the entire water column and seafloor seems typical of systems of comparable depth and latitude globally. Areas containing a Biological diversity X comparatively higher diversity of ecosystems, habitats, communities, or species, or with higher genetic diversity. ICES Advice 2013, Book 1 237 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High Explanation for ranking The possible, but incompletely documented presence of large coral and sponge stands, and the documented high diversity of benthic and associated fish when corals or sponges are present in moderate or high density imply that some areas may have high diversity. Aside from these benthic communities of somewhat restricted distribution, the biodiversity otherwise appears typical of biotic communities at similar depths and latitude. Area 2. Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge ICES advises that an area with the following boundaries, capturing three distinct features, would meet one or more EBSA criteria. The area would include the following features: iii. iv. v. Subpolar Frontal Zone (coinciding with the Charlie-Gibbs Fracture Zone): The northern and southern boundaries for this feature should be set according to the known northernmost and southernmost meandering of the frontal system at 53°N and 48°N, respectively (Søiland et al., 2008). The eastern and western boundaries for this feature should be set according to the eastern and westernmost extension of the Charlie-Gibbs Fracture Zone (topography; approx. at 27°W and 42°W, respectively). Charlie-Gibbs Fracture Zone: The eastern and western boundaries for this feature should be set according to the east–west extension of the Fracture Zone (approx. at 27°W and 42°W, respectively). The northern and southern boundaries for this feature should be set with a view to encompass the characteristic bathymetry, topography, and substrates of the Fracture Zone. Sections of the Mid-Atlantic Ridge: The northernmost and southernmost boundaries would coincide respectively with the southern boundary of proposed Area 1 (Reykjanes Ridge south of Iceland EEZ) and the northern boundary of proposed Area 3 (Mid-Atlantic Ridge north of the Azores). Boundaries to the east and to the west would be a depth contour running in the deeper of two properties: • • including a large portion (arbitrarily, perhaps 90%) of all hard volcanic substrates; habitats reported to host the larger known coral deposits and their associated communities; including a large portion (arbitrarily, 90%) of the larger known deep-sea sponge aggregations. Where the Charlie-Gibbs Fracture Zone crosses the Mid-Atlantic Ridge, the benthic boundaries of the Fracture Zone in feature ii) may extend to the east and west beyond the area defined by feature iii) for the Mid-Atlantic Ridge. In those cases the benthic area of the Fracture Zone is proposed for inclusion in the EBSA proposed here. However, any part of the seafloor and associated benthos that lies below the pelagic feature i) (the total area occupied by the Subpolar Front during its annual movement) but does not meet either feature ii) or feature iii) is not included in the area proposed as meeting EBSA criteria. Only those parts of the water column where the Subpolar Front is prominent at some time during the year are proposed as meeting one or more of the EBSA criteria. Moreover, in the entire pelagic area described by i), at any given time only those parts of the total area where the Subpolar Front is located would be expected to meet some of the criteria. Although maps will show the full pelagic area is proposed as part of this complex EBSA, any conservation measures for ecological properties of the water column would need to take into account the position of the Subpolar Front to be fully effective. For each of the above-mentioned features a set of geographic coordinates delineating their respective boundaries needs to be determined. 238 ICES Advice 2013, Book 1 Evaluation of the revised area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking Uniqueness rarity or Low Some X High (X) The portion of the proposed area encompassing both the Charlie-Gibbs Fracture Zone and Subpolar Front Zone are unique. Both represent unique or unusual geomorphological or oceanographic features in the Northeast Atlantic. Other portions of the proposed EBSA are part of the Mid-Atlantic Ridge and, as discussed for Area 1 (Reykjanes Ridge south of Iceland EEZ), may host some unique benthic species based on Mar-Eco sampling. Areas that are required for a Special X importance for population to survive and thrive. life-history stages of species Explanation for ranking There is no evidence available suggesting a significant importance of the area for life-history stages of widespread species in comparison to other marine areas of similar size and depth range. Importance for Areas containing habitats for the X survival and recovery of threatened, endangered, or endangered, threatened, or declining species, or areas with significant declining species and/or assemblages of such species. habitats Explanation for ranking There is good evidence that the area contains a significant assemblage of species and habitats that are assessed to be threatened, endangered, or declining, including the following: orange roughy (Hoplostethus atlanticus), leafscale gulper shark (Centrophorus squamosus), gulper shark (Centrophorus granulosus), Portuguese dogfish (Centroscymnus coelepis), Sei whale (Balaenoptera borealis), sperm whale (Physeter macrocephalus), leatherback turtle (Dermochelys coriacea), Lophelia pertusa reefs, and deep-sea sponge aggregations. Depending on the species, the special features of the Fracture Zone, the Subpolar Frontal Zone, or in a few cases the Mid-Atlantic Ridge, all provide important biological functions to the species which aggregate along each one. Areas that contain a relatively high Vulnerability, X proportion of sensitive habitats, fragility, sensitivity, or biotopes, or species that are functionally fragile (highly slow recovery susceptible to degradation or depletion by human activity or by natural events) or with slow recovery. Explanation for ranking The Charlie-Gibbs Fracture Zone and sections of the Mid-Atlantic Ridge through its associated substrate, current, and feeding conditions, provide habitats to a number of sensitive/vulnerable species and communities both on soft and hard substrate and in the water column. In particular biogenic habitats such as those formed by cold-water corals and sponges are considered vulnerable, often fragile, and slow (if at all) to recover from damage. Some fish species associated with the Fracture Zone and Mid-Atlantic Ridge also show slow growth, late maturity, irregular reproduction, and long generation time, as well as community characteristics of high diversity at low biomass. However, there is no evidence available suggesting that the area contains a significantly higher proportion of species that are functionally fragile or with slow recovery than other areas of comparable structure and depth range along the Mid-Atlantic Ridge. ICES Advice 2013, Book 1 239 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Areas containing species, populations, or communities with comparatively higher natural biological productivity. Explanation for ranking Biological productivity Low X Some High (X) There is good evidence that, due to the Subpolar Front, the pelagic area where the front is located at any particular time is characterized by an elevated abundance and diversity of many taxa, including an elevated standing stock of phytoplankton. This justifies a ranking of “High” for the pelagic area around the Subpolar Front, as it moves seasonally. However, there is no evidence of relatively elevated productivity in the benthic communities of the Fracture Zone and Mid-Atlantic Ridge. Areas containing a comparatively Biological X (X) higher diversity of ecosystems, diversity habitats, communities, or species, or with higher genetic diversity. Explanation for ranking The area of the Fracture Zone is characterized by a very high structural complexity, offering a diverse range of habitats. The area of the Subpolar Front is a feature where species are documented to assemble seasonally, and the sections of the Mid-Atlantic Ridge north and south of the Fracture Zone represent different biogeographic settings and their respective characteristic communities. Consequently, each of the three features characterizing this area contribute to a relatively higher diversity of ecosystems, habitats, communities, and species in comparison to other areas of similar size along the Mid-Atlantic Ridge. Area 3. Mid-Atlantic Ridge north of the Azores ICES considers that one or more EBSA criteria would be met by an area with boundaries including all the following properties: • • • 240 Including a large portion (arbitrarily, perhaps 90%) of all hard volcanic substrates on the Mid-Atlantic Ridge south of the summer location of the Subpolar Front. Including the Moytirra Hydrothermal Vent Field. Incluings the area in the Mid-Atlantic Ridge included in the 50% density Kernel for foraging Cory’s shearwater. ICES Advice 2013, Book 1 Evaluation of the proposed area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking Uniqueness rarity or Low X Some High (X) There is support for qualification under this criterion only from the hydrothermal vent field in the area which is considered to be rare, and its associated communities which may be unique. This habitat is known from a single discrete location (thus the brackets) and so it cannot be considered to offer justification for the entire extent of the proposed area. ICES does not consider that any of the other evidence presented here supports qualification under this criterion. Areas that are required for a Special X importance for population to survive and thrive. life-history stages of species Explanation for ranking There is some support for qualification under this criterion from the occurrence of an important long-range foraging area for Cory’s shearwaters during their breeding season. However, the core area encompassing 50% of locations at sea is relatively small and does not justify the entire extent of the proposed area. If research finds that deep-sea sharks and rays use the denser coral deposits as important spawning and nursery grounds, as has been reported in the Hatton–Rockall Bank area proposed by ICES, then there would be additional justification for a score of “Some” on this criterion. Importance for Areas containing habitats for the survival and X recovery of endangered, threatened, or declining threatened, endangered, or species, or areas with significant assemblages of such species. declining species and/or habitats Explanation for ranking The only recognised threatened and/or declining species identified in the report as occurring in the area are the deepwater sharks Centrophorus squamosus and Centroscymnus coelolepis, both of which are included on the OSPAR list of threatened and/or declining species and habitats. Since this area is not considered to have special importance for their survival (compared with other areas of similar depth and latitude elsewhere) they would not qualify under this criterion. However, in addition to the threatened and declining species mentioned in the proposal, the OSPAR listed habitats ‘seamount communities’ and ‘coral gardens’ are likely to exist in this area. If these were taken into account together with the sharks, this might be regarded as a significant assemblage of threatened and declining species and habitats and the ranking would be “Some”. Areas that contain a relatively high proportion of Vulnerability, X sensitive habitats, biotopes, or species that are fragility, sensitivity, or functionally fragile (highly susceptible to degradation or depletion by human activity or by slow recovery natural events) or with slow recovery. Explanation for ranking The proposed area focuses on areas with high abundance of seamounts, stony corals, and coral gardens. Recently a new hydrothermal vent was discovered on the ridge at 45ºN. Seamounts, ocean ridges with hydrothermal vents, coral ICES Advice 2013, Book 1 241 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know reefs, and coral gardens are all considered priority habitats in need of protection by the OSPAR convention for the protection and conservation of the Northeast Atlantic. There is good support for qualification under this criterion from the occurrence of vulnerable marine ecosystems, including seamounts, stony corals, coral gardens, and hydrothermal vents. Areas containing species, populations, or Biological X communities with comparatively higher natural productivity biological productivity. Explanation for ranking There is no evidence that the productivity in the revised area is any different from the expected productivity of marine systems of similar depth and latitude. Areas containing a comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or diversity with higher genetic diversity. Explanation for ranking The presence of a hydrothermal vent field does not in itself indicate high diversity, but it does provide some evidence of habitat heterogeneity from which species diversity may be inferred. In addition, there is a mingling of benthic faunas characteristic of both warmer southern and cooler northern ocean environments, giving the area as a whole somewhat higher net biological diversity, although the diversity in any individual site is not markedly enhanced. Area 4. The Hatton and Rockall banks and the Hatton–Rockall Basin The Hatton and Rockall banks, and associated slopes, represent unique offshore bathyal habitats (200 to 3000 m) and constitute a prominent feature of the Northeast Atlantic continental margin south of the Greenland to Scotland ridges. The banks and slopes have high habitat heterogeneity and support a wide range of benthic and pelagic faunas. They are also subject to significant fishing impact, including bottom trawling, longlining, and midwater fisheries. The banks encompass a large depth range and consequently the seabed communities encounter strong environmental gradients (e.g. temperature, pressure, and food availability). These factors cause large-scale changes in species composition with depth and give rise to a high diversity of species and habitats. The area is influenced by a number of different water masses and there is considerable interaction between the topography and physical oceanographic processes, in some areas focusing internal wave and tidal energy which results in strong currents and greater mixing and resuspension. ICES recommend that additional work is needed primarily to refine the boundaries set out for this proposed EBSA. In particular the evidence base to use the 3000 m contour as the southern and western limits of this proposed EBSA is questionable since no evidence was provided that ecosystems meeting EBSA criteria are present at these depths. The features contributing to the uniqueness and rarity, threatened and declining species, vulnerability/fragility/sensitivity, and importance for life-history criteria stages all occur exclusively at relatively shallow depths (< 1500 m) with most being < 1200 m, and the only additional benefit gained by extending the boundary to 3000 m is an increase in the overall depth range covered and hence additional biological diversity. It is unclear whether the additional diversity conferred by the inclusion of the 1500 to 3000 m depth zone is any different from that present in any other area at comparable depth and latitude. ICES therefore recommends that proposed EBSA should go forward with a revised boundary approximating to the 1500 m depth contour. If further work can establish significant additional value in the inclusion of greater depths, the boundary could be adjusted accordingly in the future. Evaluation of the proposed area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Uniqueness rarity 242 or The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, Low Some High X (X) ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking The area has considerable environmental heterogeneity, and therefore biological diversity, as a result of its large depth range and strong environmental gradients. Habitat-forming sessile benthic communities, such as those of giant protozoans and sponges, are common. Although distinctive these features are not rare per se. Large areas of cold-water corals and sponges have been reported in the area. Some of these have been impacted by bottom trawl and longline fishing and past periods of bottom gillnet fishing, but some areas of large coral frameworks still exist, including areas such as the Logachev coral carbonate mound province which spans both national EEZ (Ireland) and international waters. Many of these coral frameworks are now protected as VMEs. An area of polygonal faults may be a unique seabed feature. It is currently poorly investigated but may host important biological communities (e.g. cold-seeps). The polygonal faults do not themselves appear to support unique biological communities or species but may be indicative of possible presence of active hydrocarbon seeps. One such active seep has recently been discovered in this area, supporting a rare chemosynthetic community that hosts species that have not been recorded elsewhere (hence the bracketed “High” score). There is support for qualification under this criterion from the occurrence of polygonal faults and an active cold hydrocarbon seep. These features exist within a very restricted area of the site and, as described, the uniqueness and rarity criterion would only apply where these habitats occur. If further information is provided on the occurrence of large areas of cold-water coral reef, this may provide further support for this criterion over a wider geographical area. Areas that are required for a population to survive Special X importance for and thrive. life-history stages of species Explanation for ranking Cold-water corals and areas of natural coral rubble provide highly diverse habitats. Recent observations show that Lophelia pertusa reefs provide nursery grounds for deep-water sharks, and egg cases of deep-water rays were recorded with small patches of Solenosmilia variabilis framework on the Hebrides Terrace Seamount during the RRS James Cook 073 Changing Oceans Expedition in June 2012. New evidence from RRS James Cook cruises 073 and 060 shows that small patch reefs of L. pertusa on Rockall Bank are used as refuge by gravid Sebastes viviparous. Parts of the Hatton–Rockall area are important as spawning areas for blue whiting, and the area is used as a corridor for a range of migrating species, including turtles. Blue whiting has a widespread spawning area from the Faroe–Shetland Channel in the north to the Porcupine Bank in the south. Three areas of blue ling spawning aggregations are known on the shallow parts of Hatton and Lousy banks. These are significant since they represent three of the six known or suspected spawning locations for the southern stock of blue ling. Importance for Areas containing habitats for the survival and X recovery of endangered, threatened, or declining threatened, endangered, or species, or areas with significant assemblages of such species. declining species and/or habitats Explanation for ranking The OSPAR threatened and declining habitats and species “carbonate mounds” and Lophelia pertusa are confirmed to be present in the area, and indicator species for “deep-sea sponge aggregations” and “coral gardens” have been recorded. The presence of these habitats has been confirmed in some areas that are now protected as VMEs. ICES Advice 2013, Book 1 243 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know The cold-water corals and natural rubble contain very large numbers of invertebrate species, including giant protozoans (xenophyophores), vase-shaped white sponges, actiniarians, antipatharian corals, hydroids, bryozoans, asteroids, ophiuroids, echinoids, holothurians, and crustaceans. The distribution of cold-water coral has been severely reduced in the area over the last 30 years. The deep-water sharks C. coelolepis and C. squamosus are also listed in the OSPAR list. Both occur in the area, but there is no information to indicate that this area is important for either species in the sense of having a significant proportion of the population or higher density than other areas of similar depth in the region. Both Zino’s petrel (endangered) and Fea’s petrel (near threatened) are listed on the IUCN Red List. A further five species of seabirds listed in Annex I of the European Union Bird’s Directive are found within the area. However, tracking for the two petrel species (data in Figure 2 of the proposal; OSPAR/NEAFC/CBD, 2011) appears to show that the area is of relatively low importance (5 to 10% of tracked birds) during a very short period (one month). Knowledge of cetaceans in the area is poor, but the critically endangered northern right whale (Eubalaena glacialis) has been observed in this area. However, this single observation is insufficient to demonstrate importance for this species. Areas that contain a relatively high proportion of Vulnerability, X sensitive habitats, biotopes, or species that are fragility, sensitivity, or functionally fragile (highly susceptible to degradation or depletion by human activity or by slow recovery natural events) or with slow recovery. Explanation for ranking It is uncertain how “a relatively high proportion” is defined in this context, but there is good evidence for vulnerable habitats and benthic species in the area (records of cold-water coral reefs and carbonate mounds, and indicator species for coral gardens, deep-water sponge aggregations). Distribution is not uniform across the area and many of the areas where they occur are now protected as VMEs. There is a high diversity of corals, including bamboo coral (Isididae), black coral (Antipatharia), as well as the reefforming stony corals (Scleractinia), though some of these may now be reduced in distribution and occurring in patches. Cold-water coral habitats are easily impacted and recover very slowly. Some species of cold-water corals can live for more than 4000 years. Many of the demersal fish have life histories of deep-water fish fauna with very slow recovery times as a result of their slow reproductive rate compared to pelagic fish. These fish may be more exposed to fishing pressure because trawlable habitat is more common in this area than is typical at these depths. Stocks have already been diminished in some areas. There is good support for qualification under this criterion from the occurrence of vulnerable marine ecosystems, including stony corals, carbonate mounds, possible coral gardens and deep-sea sponge aggregations, and an active cold seep. Although comparative studies have not been done, it is probable that occurrence of corals in the Rockall–Hatton area is higher than in other areas of comparable depth and latitude. This would therefore constitute a “relatively high proportion”. The species or habitats discussed in this rationale are generally found in depths above 1500 m and thus this proposed EBSA should be limited to this depth contour. Areas containing species, populations, or Biological X communities with comparatively higher natural productivity biological productivity. Explanation for ranking It is likely that pelagic production may be enhanced relative to surrounding areas due to upwelling, but benthic secondary production in deep-water environments is generally considered to be low compared to other environments. Areas containing a comparatively higher diversity Biological X of ecosystems, habitats, communities, or species, diversity or with higher genetic diversity. 244 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High Explanation for ranking The area comprises a patchwork of habitats with species changing consistently with both habitat type and increasing depth. Some habitats are threatened by direct impacts (e.g. bottom fishing), others may suffer indirectly (e.g. through the creation of sediment plumes by impacts of fishing gear in sensitive areas). Seabed communities include cold-water corals, rocky reefs, carbonate mounds, polygonal fault systems, sponge aggregations, and steep and gentle sedimented slopes. Cold-water corals provide diverse habitats for other invertebrates and fish. This area spans more than one biogeography province; consequently, overall diversity is likely to be higher than in other areas with comparable depth and habitat range. Rare habitats such as cold seeps and highly diverse habitats such as cold-water coral reef and rubble further contribute to the overall diversity. Proposal for a different configuration of EBSAs than those presented in the OSPAR/NEAFC/CBD Workshop report Proposed EBSAs 1, 2, and 3 (Reykjanes Ridge south of Iceland EEZ, Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge, and Mid-Atlantic Ridge north of the Azores) all encompass the hard substrates running roughly north–south in the higher elevations of the Mid-Atlantic Ridge, and for proposed EBSAs 1 to 3 the southern boundary of each aligns with the northern boundary of the next. The boundaries between them were defined primarily by the extreme limits of the position of the east–west-oriented Subpolar Front, a major pelagic oceanographic feature in proposed EBSA 2 that moves seasonally northward spring and summer) and southward (autumn and winter). The Subpolar Front has affinities with the Charlie-Gibbs Fracture Zone, but not with the Mid-Atlantic Ridge. If the Charlie-Gibbs Fracture Zone, running roughly east to west, and associated Subpolar Frontal Zone were treated separately from the Mid-Atlantic Ridge, then all the areas delineated by the features of the Mid-Atlantic Ridge specified for the proposed EBSAs 1, 2, and 3 would share a consistent geomorphological feature (the emergent hard substrates primarily of volcanic origin) with associated benthic fauna present from the northern boundary of proposed EBSA 1 to the southern boundary of proposed EBSA 3, with the Charlie-Gibbs Fracture Zone itself simply serving as an interruption in this feature. The entire Mid-Atlantic Ridge feature would score “Some” or “High” on several criteria, and for generally the same biological rationales for the entire ridge. The species composition of the benthic biota does change from north to south, but aside from the structural interruption caused by the transverse fractures, there is no strong evidence that discontinuities in benthic community composition exist along the ridge. Thus, an alternative area to proposed EBSAs 1, 2, and 3 can be justified along the entire Mid-Atlantic Ridge in the OSPAR/NEAFC area, defined by the features specified in the description of proposed EBSA 1, and ranked as “Some” or “High” on several EBSA criteria with a common justification for that entire area. A second alternative EBSA could then be proposed, consisting of the Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone, taken together. This area would have its own set of ecological properties and associated rankings on the EBSA criteria. It would be ranked “Some” or “High” on several criteria for justifications specific to the Fracture Zone and Subpolar Front, but in several cases for justifications very different from that of the Mid-Atlantic Ridge. These two potential proposed EBSAs would replace proposed EBSAs 1, 2, and 3 from the OSPAR/NEAFC/CBD Workshop report. It would also require a separate consideration of seabird foraging in the southern third of the area, jointly with the additional analyses already recommended for the OSPAR/NEAFC/CBD Workshop proposed EBSAs 5– 8. Alternative proposed EBSA for the Mid-Atlantic Ridge The Mid-Atlantic Ridge runs from the southern boundary of the Icelandic EEZ to the northern boundary of the Portuguese EEZ in the Azores and includes all area above a depth contour that runs in the deeper of two properties: 5. 6. Including a large portion (arbitrarily, perhaps 90%) of all hard volcanic substrates; habitats are reported to host the larger known coral deposits and their associated communities. Included a large portion (arbitrarily, 90%) of the records of large sponge communities in the overall Mid-Atlantic Ridge. ICES Advice 2013, Book 1 245 In addition the proposed area should include all known hydrothermal vents along the ridge, if any of these are deeper than the contours meeting the properties above. Evaluation of the proposed area against the EBSA criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know The area contains either (i) unique (“the only one of X (X) its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking The qualified “High” ranking is for restricted areas of the few known hydrothermal vents along the ridge. These are globally rare features; only a small part of the area proposed here meets this EBSA criterion. Uniqueness rarity or The MarEco sampling of corals and sponges reported several species new to science as it sampled the Mid-Atlantic Ridge. However, it is not possible to draw a firm conclusion on the presence of unique species along the ridge until more extensive sampling is undertaken. Areas that are required for a population to survive Special X importance for and thrive. life-history stages of species Explanation for ranking Although many populations undoubtedly complete their life cycles within the harder-substrate areas of the MidAtlantic Ridge, this would be true of any marine area of comparable size. There is no evidence that the life history of any species is strongly dependent on any specific features of the area proposed as an alternative EBSA. There is evidence from other areas of the Northeast Atlantic that areas of high coral density may be important as eggcase and nursery areas of deep-water sharks and rays. The area proposed here is targeted on the depths and substrates associated with higher coral and sponge densities, and if sharks and rays also concentrate spawning and early development in these habitats, then the score would be “Some” or “High” on this criterion as well. However, it has not yet been documented that skates and rays do preferentially use the coral and sponge formations for these life history functions in the Mid-Atlantic Ridge. For the more southern portions of the Mid-Atlantic Ridge in particular, there are reports of areas being important for foraging by seabirds, including Cory’s shearwater. The evidence available is not considered strong, however, and this aspect of the ecological functionality of the central ridge area should be considered as part of the reanalysis of EBSAs 5–8 proposed in the OSPAR/NEAFC/CBD Workshop report. Importance for Areas containing habitats for the survival and X recovery of endangered, threatened, or declining threatened, endangered, or species, or areas with significant assemblages of such species. declining species and/or habitats Explanation for ranking There are large deposits of corals and sponges found in the alternative area proposed by ICES as meeting this criterion. Habitats containing these species are listed by OSPAR and are also VME indicator species, and the majority of these would be included in the alternative proposed EBSA. Additional explanation regarding corals and sponges is included in the rationale for the criterion on Vulnerability. The possible role of the alternative proposed Mid-Atlantic Ridge area in the life histories of sharks and rays is discussed in the criterion on Vulnerability and Sensitivity. If a dependency between the breeding or early life history of threatened or endangered skates or rays were documented, there would be additional justification for a “High” ranking of this criterion. 246 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know Areas that contain a relatively high proportion of sensitive habitats, biotopes, or species that are functionally fragile (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery. Explanation for ranking Vulnerability, fragility, sensitivity, or slow recovery X With regard to corals and sponges, Mortensen et al. (2008) found cold-water corals “at every sample station … observed at depths between 800 and 2400 m, however were commonly found shallower than 1400 m …, with species richness being very high. … no major reef structures were recorded, with the maximum colony size approximately 0.5 m in diameter. The number of coral taxa was strongly correlated with the percentage cover of hard bottom substrate ….” The area proposed here is targeted at the seamount peaks and slopes where hard substrates dominate. For sponges, no actual large expanses of sponge reef were reported in the OSPAR/NEAFC/CBD Workshop report. However, the pro forma in that report notes that overall sampling of the area is patchy and cites three studies that found local patches with high densities of sponges, although in no cases were the sizes of the patches documented. If research finds that deep-sea sharks and rays use the denser coral deposits as important spawning and nursery grounds, as has been reported in the Hatton–Rockall Bank area proposed by ICES, then there would be additional justification for a score of “Some” on this criterion. Areas containing species, populations, or Biological X communities with comparatively higher natural productivity biological productivity. Explanation for ranking Although benthic productivity of the alternative proposed Mid-Atlantic Ridge EBSA may be higher than benthic productivity on the abyssal plain, productivity integrated over the entire water column and seafloor seems typical of systems of comparable depth and latitude globally. Areas containing comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or diversity with higher genetic diversity. Explanation for ranking The presence of comparatively large coral and sponge formations, and the documented high diversity of benthic and associated fish when corals or sponges are present in moderate or high density imply that some areas along the ridge may have high diversity. From north to south there is a mingling of benthic and demersal fish faunas characteristic of both cooler northern and warmer southern ocean environments, giving the area as a whole a somewhat higher net biological diversity, even if the diversity in any individual site is not markedly enhanced. Aside from the benthic communities of somewhat restricted distribution associated with the biogenic habitats, the biodiversity otherwise appears typical of biotic communities at similar depths and latitude. Alternative proposed EBSA – The Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone The area would include: vi. vii. Subpolar Frontal Zone (coinciding with the Charlie-Gibbs Fracture Zone): The northern and southern boundaries for this feature should be set according to the known northernmost and southernmost locations of the frontal system (at approximately 53°N and 48°N). The eastern and western boundaries for this feature should be set according to the eastern and westernmost extension of the Charlie-Gibbs Fracture Zone (at approximately 27°W and 42°W). Charlie-Gibbs Fracture Zone: The eastern and western boundaries for this feature should be set according to the east–west extension of the Fracture Zone (at approximately 27°W and 42°W). The northern and southern seabed boundaries for this feature should be set with a view to encompass the characteristic topography and substrates of the Fracture Zone. Any area of the seafloor and associated benthos that lies below the pelagic feature i) (the total area occupied by the Subpolar Front during its annual movement) but does not meet feature ii) is not included in the area proposed as meeting EBSA criteria. Only those parts of the water column where the Subpolar Front is prominent at some time during the year are proposed as meeting one or more of the EBSA criteria. Moreover, in the entire pelagic area described by i), at any ICES Advice 2013, Book 1 247 given time only that part of the total area where the Subpolar Front is located would be expected to meet some of the criteria. Although maps will show that the full pelagic area is proposed as part of this complex EBSA, any conservation measures for ecological properties of the water column would need to take into account the position of the Subpolar Front to be fully effective. Evaluation of the proposed area against the EBSA criteria CBD EBSA Criterion Description The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations), or endemic species, populations, or communities, and/or (ii) unique, rare, or distinct habitats or ecosystems, and/or (iii) unique or unusual geomorphological or oceanographic features. Explanation for ranking Uniqueness rarity or Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know X The Charlie-Gibbs Fracture Zone is a set of geomorphological features unique to the entire North Atlantic, and the Subpolar Front is an oceanographic feature also unique to the Northeast Atlantic. Together these features justify a “High” score for this criterion. Areas that are required for a population to survive Special X importance for and thrive. life-history stages of species Explanation for ranking No evidence is available suggesting a significant importance of the area for life-history stages of widespread species in comparison with other marine areas of similar size and depth range. It is possible that there are species with special affinities for the unique geophysical features of the deep faults, but clear documentation of species with such affinities was not found in the references provided by the OSPAR/NEAFC/CBD Workshop report, and was not otherwise known to ICES. Importance for Areas containing habitats for the survival and X recovery of endangered, threatened, or declining threatened, endangered or species, or areas with significant assemblages of such species. declining species and/or habitats Explanation for ranking There is good evidence that the area contains a significant assemblage of species and habitats that are assessed to be threatened, endangered, or declining, including leafscale gulper shark (Centrophorus squamosus), gulper shark (Centrophorus granulosus), Portuguese dogfish (Centroscymnus coelepis), Sei whale (Balaenoptera borealis), sperm whale (Physeter macrocephalus), leatherback turtle (Dermochelys coriacea), as well as cold-water coral reefs and deep-sea sponge aggregations. Depending on the species, the special features of the Fracture Zone and the Subpolar Front are inferred to provide important biological functions to the species which aggregate along each one. Areas that contain a relatively high proportion of Vulnerability, X sensitive habitats, biotopes, or species that are fragility, sensitivity, or functionally fragile (highly susceptible to degradation or depletion by human activity or by slow recovery natural events) or with slow recovery. 248 ICES Advice 2013, Book 1 CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know Explanation for ranking The Charlie-Gibbs Fracture Zone geophysical structure provides habitats for a number of sensitive/vulnerable species and communities both on soft and hard substrate and in the associated water column. In particular biogenic habitats such as those formed by cold-water corals and sponges are considered vulnerable, are often fragile, and slow (if at all) to recover from damage. Some fish species associated with the Fracture Zone and the Subpolar Front also show slow growth, late maturity, irregular reproduction, and long generation time, as well as community characteristics of high diversity at low biomass. However, the documentation that the species with vulnerable life histories are especially closely affiliated with the Fracture Zone and frontal habitats is weak, and it is clear that these vulnerable species and biogenic habitats are not consistently present throughout the entire Fracture Zone and Subpolar Front. Areas containing species, populations, or Biological X (X) communities with comparatively higher natural productivity biological productivity. Explanation for ranking There is good evidence that, because of the Subpolar Front, the pelagic area where the front is located at any particular time is characterized by an elevated abundance and diversity of many taxa, including an elevated standing stock of phytoplankton. This justifies a ranking of “High” for the pelagic area around the Subpolar Front, as it moves seasonally. However, there is no evidence of relatively elevated productivity in the benthic communities of the Fracture Zone. Areas containing a comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or diversity with higher genetic diversity. Explanation for ranking The area of the Fracture Zone is characterized by a very high structural complexity, offering a diverse range of habitats. The area of the Subpolar Front is a feature where species are documented to assemble seasonally. Consequently, both features characterizing this area contribute to a relatively higher diversity of ecosystems, habitats, communities, and species in comparison to other areas of the Northeast Atlantic. Proposed EBSAs for which there is insufficient scientific justification For five of the ten areas proposed as meeting one or more EBSA criteria in the OSPAR/NEAFC/CBD Workshop report, ICES concluded that there is insufficient scientific justification at this time to propose their delineated area, or any subset of it, as meeting EBSA criteria. In all cases ICES recommends that additional information needs to be collated and analysed, and a new evaluation be conducted when those results are available. ICES provides reasoning for its advice and recommendations for further work in each of these areas below. Area 5. Around the Pedro Nunes and Hugo de Lacerda seamounts and Area 6. Northeast Azores–Biscay Rise The OSPAR/NEAFC/CBD Workshop concluded that both areas ranked as “High” on criterion Special Importance to Life History of Species, and “Some” on criteria Uniqueness and Rarity; Importance to Threatened, Endangered, or Declining Species; and Vulnerability, Sensitivity, etc., the latter primarily for seabirds. ICES questions the basis for these conclusions, noting that the data used to assess all the criteria specified were incomplete and often incorrectly interpreted, with the proposed boundaries not matching the information in the cited sources. ICES recommends that all available data on foraging activity of the Zino’s petrel, Cory’s shearwater, and other relevant species be examined. This should include published and any other available data. Occurrence data may be used as well, provided the rationale details how occurrence and foraging data are used to derive EBSA boundaries. Area 7. Evlanov Seamount region The OSPAR/NEAFC/CBD Workshop concluded that the area they delineated ranked as “High” on criterion Importance to Threatened, Endangered, or Declining Species, and “Some” on criteria Uniqueness and Rarity; Special Importance to Life History of Species; and Vulnerability, Sensitivity, etc., the latter primarily for seabirds. ICES questions the basis for these conclusions, noting that the data used to assess all the criteria specified were incomplete and often incorrectly interpreted, with the proposed boundaries not matching the information in the cited sources. In particular the sample sizes ICES Advice 2013, Book 1 249 for Fea’s petrel were very small, and the information for sooty shearwater did not seem to differentiate the proposed EBSA area from most of surrounding area. ICES advises not to proceed with proposing any portion of this area as an EBSA at this time, but rather undertake further collation and analysis of information and reconsider when the additional work is completed. Area 8. Northwest of Azores EEZ The OSPAR/NEAFC/CBD Workshop concluded that the area they delineated ranked as “High” on criteria Special Importance for Life History of Stages of Species and Importance for Threatened, Endangered or Declining Species, and “Some” on criteria Uniqueness and Rarity; Vulnerability, Sensitivity, etc.; and Biological Diversity, the latter primarily for seabirds. ICES questions the basis for these conclusions, noting that the data used to assess all the criteria specified were incomplete and often incorrectly interpreted, with very small sample sizes for some of the species’ (e.g. Zino’s petrel) foraging areas, and questionable interpretation of the foraging areas of Cory’s shearwater. ICES does not support this area going forward as presented in the OSPAR/NEAFC/CBD EBSA Workshop report. ICES advises that improvements are needed to the supporting evidence in the narrative for criteria related to Special Importance for Life History Stages of Species and Importance for Threatened, Endangered or Declining Species and/or Habitats. It is necessary to augment information on how the area is being used (feeding, conditioning, migration) for the survival and recovery of the species and to put some scale on its importance to the species (some of which have very restricted breeding sites in the larger area). It is also necessary to document the proportion of species that are highly susceptible to degradation or depletion by human activity, in this case bycatch in longline fisheries. Area 9. The Arctic Front – Greenland/Norwegian seas The OSPAR/NEAFC/CBD Workshop concluded that the area they delineated ranked as “High” on criteria Special Importance to Life History of Species; Importance to Threatened, Endangered, or Declining Species; Biological Productivity; and Biological Diversity. ICES questions the basis for these conclusions, noting that (1) for several features the area proposed as meeting criteria were not noticeably different from the surrounding areas; (2) some of the rankings regarding importance appeared to be inferred from a belief that the area is high in productivity, but the rankings were not demonstrated otherwise; and (3) publications with contrasting conclusions were found for some of the key references cited in the OSPAR/NEAFC/CBD Workshop report. ICES recommends not to proceed with the proposed Arctic Front EBSA. There is no evidence of enhanced productivity at the Arctic Front which is the main rationale used to justify the proposed EBSA. However, there seems to be circumstantial evidence for an enhanced production that may attract feeding animals in the areas around and south of Jan Mayen, including the Jan Mayen Front. If parts of this area are located in the high seas, further analyses should be undertaken to determine if this area meets the EBSA criteria. Sources CBD. 2007. Expert Workshop on Ecological Criteria and Biogeographic Classification Systems for Marine Areas in Need of Protection. 2–4 October 2007, Azores, Portugal. (http://www.cbd.int/doc/meetings/mar/ewsebm01/official/ewsebm-01-02-en.pdf). CBD. 2008. CBD Decision IX/20 on Marine and Coastal Biodiversity. (http://www.cbd.int/doc/decisions/cop-09/cop-09dec-20-en.pdf). CBD. 2009. Expert Workshop on Scientific and Technical Guidance on the Use of Biogeographic Classification Systems and Identification of Marine Areas beyond National Jurisdiction in Need of Protection. 29 September–2 October 2009, Ottawa, Canada. (https://www.cbd.int/doc/meetings/sbstta/sbstta-14/information/sbstta-14-inf-04-en.pdf). CBD. 2010. CBD Decision X/29 on Marine and Coastal Biodiversity. (https://www.cbd.int/decision/cop/default.shtml?id=12295). German, C. R., and Parsons, L. M. 1998. Distributions of hydrothermal activity along the Mid-Atlantic Ridge: interplay of magmatic and tectonic controls. Earth and Planetary Science Letters, 160: 327–341. German, C. R., Briem, J., Chin, C., Danielsen, M., Holland, S., James, R., Jónsdottir, A., Ludford, E., Moser, C., Ólafsson, J., Palmer, M. R., and Rudnicki, M. D. 1994. Hydrothermal activity on the Reykjanes Ridge: the Steinahóll ventfield at 63°06’N. Earth and Planetary Science Letters, 121: 647–654. ICES. 2013a. Vulnerable deep-water habitats in the NEAFC Regulatory Area. In Report of ICES Advisory Committee, 2013, Section 1.5.5.1. ICES Advice, 2013, Book 1. ICES. 2013b. Report of the Workshop to Review and Advise on EBSA Proposed Areas (WKEBSA), 27–31 May 2013, Copenhagen, Denmark. In draft. 250 ICES Advice 2013, Book 1 Mironov, A., and Gebruk, A. 2007. Deep-sea benthos of the Reykjanes Ridge: biogeographic analysis of the fauna living below 1000 m. Report on preliminary phase of the project patterns and processes of the ecosystems of the northern Mid-Atlantic (Mar-Eco). (http://www.mareco.no/sci/component_projects/epibenthos/deep-sea_benthos_of_the_reykjanes_ridge). Mortensen, P. B., Buhl-Mortensen, L., Gebruk, A. V., and Krylova, E. M. 2008. Occurrence of deep-water corals on the Mid-Atlantic Ridge based on Mar-Eco data. Deep-Sea Research, 55: 142–152. Olafsson, J., et al. 1991. A sudden cruise off Iceland. RIDGE Events Newsletter, 2(2): 35–38. OSPAR/NEAFC/CBD. 2011. Report of the Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic. 8–9 September 2011, Hyères, France. Annex 11. Søiland, H., Budgell, W. P., and Knutsen, Ø. 2008. The physical oceanographic conditions long the Mid-Atlantic Ridge north of the Azores in June–July 2004. Deep-Sea Research II, 55: 29–44. United Nations. 2004. UNGA Resolution 58/240. (http://daccess-dds-ny.un.org/doc/UNDOC/GEN/N03/508/92/PDF/N0350892.pdf?OpenElement). ICES Advice 2013, Book 1 251 Annex 1.5.6.5.1 Revised pro forma for the Arctic Ice habitat Annex 1 Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic Hyères (Port Cros), France: 8 – 9 September 2011 ________________________________________ EBSA identification proforma for the North-East Atlantic - 10 Title/Name of the area - The Arctic Ice habitat - multiyear ice, seasonal ice and marginal ice zone Presented by WWF and reviewed by Participants at the Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas in the North-East Atlantic Contact: Sabine Christiansen [email protected] Abstract The permanently ice covered waters of the high Arctic provide a range of globally unique habitats associated with the variety of ice conditions. Multi-year sea ice only exists in the Arctic and although the projections of changing ice conditions due to climate change project a considerable loss of sea ice, in particular multiyear ice, the Eurasian Central Arctic high seas are likely to at least keep the ice longer than many other regions in the Arctic basin. Ice is a crucial habitat and source of particular foodweb dynamics, the loss of which will affect also a number of mammalian and avian predatory species. The particularly pronounced physical changes of Arctic ice conditions as already observed and expected for the coming decades, will require careful ecological monitoring and eventually measures to maintain or restore the resilience of the Arctic populations to quickly changing environmental conditions. Introduction Up until today most of the Eurasian part of the Arctic Basin, and in particular the high seas area in the Arctic Ocean (the waters beyond the 200 nm zones of coastal states, i.e. Norway, Russia, USA, Canada and Greenland/Denmark) is permanently ice covered. However, in recent years, much of the original multiyear pack ice has been replaced by seasonal (1 year) ice which made it possible for research and other vessels to reach the pole. In addition, the former fast pack-ice is now increasingly broken up by leads. This structural change in the Arctic ice quality will result in a substantial increase in light penetrating the thin ice and water column, in conjunction with the overall warming of surface waters and increased temperature and salinity stratification due to the melting of ice. In the near future, up to the end of the century, the permanent ice cover is expected to disappear completely in some models (Anisimov et al., 2007). This will result in significant changes in the structure and dynamics of the high Arctic ecosystems (CAFF, 2010; Gradinger, 1995; Piepenburg, 2005; Renaud et al., 2008; Wassmann, 2008, 2011) which should be closely monitored (Bluhm et al., 2011) as already envisaged by the Arctic Council (Gill et al., 2011; Mauritzen et al., 2011). Therefore, the area proposed here as EBSA is of particular scientific interest and may in the longterm, become relevant for the commercial exploitation of resources. 252 ICES Advice 2013, Book 1 Location The Ecologically or Biologically Significant Marine Area (EBSA) proposed focusses on the presently permanently ice-covered waters in the OSPAR/NEAFC maritime areas, including the high seas section in the Central Arctic Basin north of the 200 nm zones of coastal states (see Fig. 1 attached). Therefore, the boundaries proposed extend from the North Pole (northernmost point of OSPAR/NEAFC maritime areas) to the southern limit of the summer sea ice extent and marginal ice zone, including on the shelf of East Greenland. The proposal currently only relates to features of the water column. Two legal states have to be distinguished: the Central Arctic high seas waters north of the 200 nm zones of adjacent coastal states, generally north of 84° N, and the waters within the Exclusive Economic Zones of Greenland, Russia and the fisheries protection zone of Norway around Svalbard. Figure 1 distinguishes between the high seas beyond national jurisdiction for which the „Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic“ has a mandate19, and national/nationally administered waters within the 200 nm zone, within which the OSPAR Contracting Parties have the responsiblity to report candidate EBSAs to the Convention on Biodiversity EBSA repository (OSPAR Commission, 2011). The seafloor of the respective region will likely fall on the extended continental shelves of several coastal states. It belongs to the „Arctic Basin“ region of (Gill et al., 2011). The coordinates of the overall area, as well as the high seas section are provided in Annex 1 (in decimals, shape files provided): c.f. Figure 1: Location of the Arctic Ice „Ecologically or Biologically Significant Area“ (EBSA) proposed by WWF in September 2011. The position of the Arctic and polar fronts was redrawn after (Rey, 2004, Fig. 5.7). Feature description The Ecologically or Biologically Significant Marine Area (EBSA) proposed focusses on the presently permanently ice-covered waters in the OSPAR/NEAFC maritime areas, including the high seas section in the Central Arctic Basin north of the 200 nm zones of coastal states, and the marginal ice zone (where the ice breaks up, also called seasonal ice zone) along its southern margins (see Fig. 1 attached). Due to the inflow of Atlantic water along the shelf of Svalbard, and the concurrent outflow of polar water and ice on the Greenland side of Fram Strait, the southern limit of the summer sea ice extent is much further south in the western compared to the eastern Framstrait, and in former times extended all along the Greenland coast. The high seas section of the OSPAR maritime area in the Central Arctic ocean is generally north of 84° N and is until today fully ice-covered also in summer, although the quantity of multiyear ice has already substantially decreased and the 1-year ice leaves increasingly large leads and open water spaces. The ice overlays a very deep water body of up to 5000 m depth far away from the surrounding continental shelves and slopes of Greenland and the Svalbard archipelago. The Nansen-Gakkel Ridge, a prolongation of the Mid-Atlantic Ridge north of the Fram Strait is structuring the deep Arctic basin in this section, separating the Central Nansen Basin to the south from the Amundsen Basin to the north. Abundant hydrothermal vent sites have been discovered on this ridge at about 85° 38 N (Edmonds et al., 2003). North of Spitsbergen, the Atlantic water of the West Spitsbergen Current enters the Arctic basin as a surface current. At around 83° N, a deep-reaching frontal zone separates the incoming Atlantic and shelf waters from those of the Central Nansen Basin (Anderson et al., 1989), as reflected in ice properties, nutrient concentrations, zooplankton communities, and benthic assemblages (Hirche and Mumm, 1992, and literature quoted). This water subsequently submerges under the less dense (less salinity, lower temperature) polar water and circulates, in opposite direction to the surface waters and ice, counterclockwise along the continental rises until turning south along the Lomonossov Ridge and through Fram Strait as East Greenland Current south to Danmark Strait (Aagaard, 1989; Aagaard et al., 1985). Connecting the more fertile shelves with the 19 Participant Briefing for a Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic. Invitation Annex 2, 2011 ICES Advice 2013, Book 1 253 deep central basin, these modified Atlantic waters supply the waters north of the Nansen-Gakkel Ridge, in the Amundsen basin, with advected organic material and nutrients which supplement the autochtonous production (Mumm et al., 1998). Due to the import of organic biomass from the Greenland Sea and the Arctic continental shelves, part of which may not be kept in the food web due to the polar conditions, the Arctic Ocean may also represent an enormous carbon sink (Hirche and Mumm, 1992). In the Fram Strait, the region between Svalbard to the east and Greenland to the west, the East Greenland Current is the main outflow of polar water and ice from the Arctic Basin (Maykut, 1985) (Aagaard and Coachman, 1968). The polar front (0° C isotherm and 34.5 isohaline at 50 m depth) extends approximately along the continental shelf of Greenland, separating the polar surface water from the Arctic (Intermediate) water and the marginal ice zone to the east (e.g. Aagaard and Coachman, 1968; Paquette et al., 1985). The ice cover is densest in polar water, its extent to the east depends on the wind conditions (compare also Angelen et al., 2011; Wadhams, 1981). The seasonal latitudinal progression of increasing and diminishing light levels, respectively, is the determining factor for the timing of the phytoplankton-related pelagic production. Therefore, the springbloom and ice break up progress from south to north in spring, reaching the Arctic area by about June/July. Because the currents in Fram Strait move in opposite direction, the polar East Greenland Current to the south, and the Atlantic West Spitsbergen Current to the north, there is a delay of about a month between biological spring and summer between the polar and the Atlantic side (Hirche et al., 1991). Therefore sea ice and the effect of melting ice are important determinants of the ecosystem processes all along the East Greenland polar front from the Greenland Sea through Fram Strait to the Arctic Basin (Legendre et al., 1992; Wassmann, 2011). Ice situation The Arctic Ocean develops towards a one-year instead of a multi-year sea-ice system with consequences for the entire ecosystem, including ecosystem shifts, biodiversity loss, for water mass modifications and for its role in the global overturning circulation. At its maximum, sea-ice covers 4.47 million km² in the Arctic Basin (Gill et al., 2011): According to data from ice satellite observations in 1973-76 (NASA, 1987, in (Gill et al., 2011)), permanent ice occupied 70-80% of the Arctic Basin area, and the interannual variability of this area did not exceed 2%. Seasonal ice occupied 6-17% (before the melting period of the mid-1970s). Only in the first decade of the 21st century, the permanent-ice area decreased to 6% in February 2008, concurrent with a rapid increase in seasonal- ice. Whereas multiyear ice used to cover 50-60% of the Arctic, it covered less than 30% in 2008, after a minimum of 10% in 2007. The average age of the remaining multiyear ice is also decreasing from over 20 % being at least six years in the mid- to late 1980s, to just 6% of ice six years old or older in 2008. c.f. Figure 2: Modelled ice age distribution in 1985-2000 (left) compared to February 2008 (right) (CAFF, 2010). This trend is likely to amplify in the coming years, as the net ocean-atmosphere heat output due to the current anomalously low sea ice coverage has approximately trippled compared to previous years, suggesting that the present sea ice losses have already initiated a positive feedback loop with increasing surface air temperatures in the Arctic (Kurtz et al., 2011). About 10% of the sea ice in the Arctic basin is exported each year through Fram Strait into the Greenland Sea (Maykut, 1985) which is therefore major sink for Arctic sea ice (Kwok, 2009). From 2001 to 2005, the summer ice cover was so low on the East Greenland shelf, that it was more of a marginal ice zone (Smith Jr and Barber, 2007), however the subsequent record lows in overall Arctic ice cover brought about an increase in ice cover off Greenland, which minimised the extent of the North East Water Polynia on the East Greenland shelf20, a previously seasonally ice-free stretch of water (Wadhams, 1981). Ice related biota 20 http://www.issibern.ch/teams/Polynya/ 254 ICES Advice 2013, Book 1 Allover the Arctic, an inventory of ice-associated biota presently counts over 1000 protists, and more than 50 metazoan species (Bluhm et al., 2011). The regionally very variable ice fauna (depends i.e. on ice age, thickness, origin) consists of sympagic biota living within the caverns and brine channels of the ice, and associated pelagic fauna. The most abundant and diverse sympagic groups of the ice mesofauna in the Arctic seas are amphipods and copepods. Polar cod (Boreogadus saida) and partly Arctic cod (Arctogadus glacialis) are dependent on the sympagic macro- and mesofauna for food, themselves being important food sources for Arctic seals (such as ringed seal Phoca hispida) and birds, for example black guillemots Cephus grylle (Bradstreet and Cross, 1982; Gradinger and Bluhm, 2004 and literature reviewed; Horner et al., 1992; Süfke et al., 1998). The higher the light level in the ice, the higher is the biomass of benthic algae as well as meiofauna and microorganisms within the ice (Gradinger et al., 1991). Decreasing snow cover induces a feedback loop with enhanced algal biomass increasing the heat absorption of the ice which leads to changes in the ice structure, and ultimately the release of algae from the bottom layer (Apollonio, 1961 in Gradinger et al., 1991). Because of the distance to land and shelves, and the thickness and internal structure of the multiyear pack ice over deeper water, this type of ice has a fauna of its own (Carey, 1985; Gradinger et al., 1991). Arctic multiyear ice floes can have very high algal biomasses in the brine channels and in the bottom centimeters which serves as food for a variety of proto- and metazoans, usually smaller than 1 mm, over deep water (Gradinger et al., 1999). In the central Arctic, ice algal productivity can contribute up to 50 % of the total primary productivity, with lower contributions in the sea ice covered margins (Bluhm et al., 2011). In the boundary layer between ice floes and the water column, another specific community exists which forms the link between the ice based primary production and the pelagic fauna (Gradinger, 1995): large visible bands of diatoms hang down from the ice, exploited by amphipods such as Gammarus wilkitzki, and occacionally by water column copepods such as Calanus glacialis, which are important prey of for example polar cod Boreogadus saida. The caverns, wedges and irregularities of the ice provide important shelter from predators for larger ice associated species and provide an essential habitat for these species (Gradinger and Bluhm, 2004). During melt, the entire sympagic ice biota are released into the water column where they may initiate the spring algal plankton bloom (Smith and Sakshaug, 1990) or they may sink to the sea floor and serve as an episodic and first food pulse for benthic organisms before pelagic production begins (Arndt and Pavlova, 2005). In particular the shallow shelves and the shelf slope benthos has been shown to profit of this biomass input, reflected in very rich benthic communities (Klitgaard and Tendal, 2004; Piepenburg, 2005). The role of the polar front and marginal ice zone for the production system Primary production in the Arctic Ocean is primarily determined by light availability, which is a function of ice thickness, ice cover, snow cover, light attenuation), the abundance of both ice algae and phytoplankton, nutrient availability and surface water stratification. Generally, the spring bloom occurs later further north and in regions with a thick ice and snow cover. The current production period in the Arctic Ocean may extend to 120 days per year, with a total annual primary production in the central Arctic Ocean of probably up to 10 g C m-2 (Wheeler et al., 1996). Ice algae start primary production already at relatively low light levels when melting reduces the thickness of the ice and snow cover. Only after the ice breaks up, when melting releases the ice biota into the water column and meltwater leads to surface stratification, a major phytoplankton bloom of a few weeks develops, fuelling the higher trophic foodweb of the Arctic (Gradinger et al., 1999, and literature quoted). The marginal ice zones, i.e. where the ice gets broken up in warmer Atlantic or Arctic water, therefore play an important role in the overall production patterns of the Arctic Ocean. Due to the strong water column stratification and increased light levels involved with the melting of the ice, the location and recession of the ice edge in spring and summer determines the timing and magnitude of the spring phytoplankton bloom, which is generally earlier than in the open water (Gradinger and Baumann, 1991; Smith Jr et al., 1987). Wind- or eddy-induced upwelling in the marginal ice zone, as well as biological regeneration processes replenish the surface nutrient pool and therefore prolong the algal growth period (Gradinger and Baumann, 1991; Smith, ICES Advice 2013, Book 1 255 1987). The hydrographic variability explains the patchy patterns of primary and secondary production observed, as well as consequently the patchy occurrence of predators. The polar front separates to some degree the pelagic faunas of the polar and Arctic waters in the Greenland Sea and Fram Strait, each characterised by a few dominant copepod species with different life history strategies (Hirche et al., 1991; see also review in Melle et al., 2005): In polar waters, Calanus glacialis uses under ice plankton production and lipid reserves for initiating its spring reproduction phase, however depending on the phytoplankton bloom for raising its offspring (e.g. Leu et al., 2011). Somewhat later, on the warm side of the polar front in Arctic water, the Atlantic species Calanus finmarchicus uses the ice edge-related phytoplankton bloom for secondary production. Calanus hyperboreus, the third and largest of the charismatic copepod species has its core area of distribution in the Arctic waters of the Greenland Sea (Hirche, 1997; Hirche et al., 2006). Zooplankton of the Arctic Basin Overall zooplankton biomass decreases towards the central Arctic basin, reaching a minimum in the most northerly waters, i.e. the region with permanent ice cover (Mumm et al., 1998). However, investigations in recent years demonstrated increased biomasses compared to studies several decades earlier - possibly a consequence of the decrease in ice thickness and cover which only enabled the investigations to take place from ship board. There is a south-north decrease in zooplankton biomass, with a sharp decline north of 83°N (Hirche and Mumm, 1992), coinciding with differences in the species composition of the biomass-forming zooplankton species. Whereas the southern Nansen basin plankton is dominated by the Atlantic species Calanus finmarchicus, entering the Arctic Basin with the West Spitsbergen Current, the northernmost branch of the North Atlantic current, the Arctic and polar species Calanus hyperboreus and C. glacialis dominate the biomass in the high-Arctic Amundsen and Makarov Basins (Auel and Hagen, 2002; Mumm et al., 1998). The zooplankton species communities generally can be differentiated according to their occurrence in Polar Surface Water (0-50 m, temperature below –1.7°C, salinity less than 33.0), Atlantic Layer (200–900 m; temperature 0.5–1.5°C); salinity 34.5–34.8) and Arctic Deep Water (deeper than 1000 m, temperature -0,5-1° C, salinity > 34.9) (Auel and Hagen, 2002; Grainger, 1989; Kosobokova, 1982). The polar surface community in the upper 50 m of the water column consists of original polar species as well as species emerging from deeper Atlantic waters, alltogether leading to a high abundance and biomass peak in summer. Diversity and biomass are minimal in the impoverished Arctic basin deepwater community (Kosobokova 1982). Apart from a limited exchange with the Atlantic Ocean via the Fram Strait, the central Arctic deep-sea basins are isolated from the rest of the world ocean deepsea fauna. Therefore, the bathypelagic fauna consists of a few endemic Arctic species and some species of Atlantic origin. Due to the separation of the Eurasian and Canadian Basins by the Lomonosov Ridge, significant differences in hydrographic parameters (Anderson et al. 1994) and in the zooplankton composition occur between both basins (Auel and Hagen, 2002). Fish Polar cod, Boreogadus saida, is a keystone species in the ice-related foodwebs of the Arctic. Due to schooling behavior and high energy content polar cod efficiently transfer the energy from lower to higher trophic levels, such as seabirds, seals and some whales (Crawford and Jorgenson, 1993). Seabirds Ice cover is a physical feature of major importance to marine birds in high latitude oceans, providing access to resources, refuge from aquatic predators (Hunt, 1990). As seabirds are dependant on leads between ice floes or otherwise open water to access food, they search for the most productive waters in polynias (places within the ice which are permanently ice free) and marginal ice zones (Hunt, 1990). Here they forage both on the pelagic and sympagic ice-related fauna, especially the early stages of polar cod and the copepods Calanus hyperboreus and C. glacialis. Likely, they benefit of the structural complexity and good visibility of their prey near the ice (Hunt, 1990). 256 ICES Advice 2013, Book 1 In the Greenland Sea and Fram Strait, major breeding colonies exist on Svalbard, Greenland and on Jan Mayen, all of these within reach of the seasonally moving marginal ice zone or a polynia (North East Water Polynia on the East Greenland shelf). Breeding seabirds like Little auks (Alle alle), from colonies in the northern Svalbard archipelago feed their offspring with prey caught in the vicinity of the nests, however intermittently travel at least 100 km to the marginal ice zone at 80° N to replenish their body reserves (Jakubas et al., Online 03 June 2011). Therefore, the distance of the marginal ice zone to the colony site is a critical factor determining the breeding success (e.g. Joiris and Falck, 2011). Opportunistically, the birds also use other zooplankton aggregations such as a in a cold core eddy in the Greenland Sea, closer to the nesting site (Joiris and Falck, 2011). A synopsis of seabird data for the period 1974–1993 (Joiris, 2000) showed that the little auk is one of the most abundant species, together with the fulmar Fulmarus glacialis, kittiwake Rissa tridactyla and Brünnich’s guillemot Uria lomvia in the European Arctic seas (mainly the Norwegian and Greenland Seas). In the Greenland Sea and the Fram Strait, little auks represented the main species in polar waters, at the ice edge and in closed pack ice, reaching more than 50% of all bird species (Joiris and Falck, 2011). In spring and autumn, millions of seabirds pass through the area when migrating between their breeding sites on Svalbard or the Russian Arctic and their wintering areas in Canada (Gill et al., 2011). There are several seabird species in the European Arctic which are only met in ice-covered areas, for example the Ivory gull Pagophila eburnea and the Thick-billed guillemot Uria lomvia (see e.g. CAFF, 2010): Both species spend the entire year in the Arctic, and breed in close vicinity to sea ice although Thick-billed guillemots were observed to fly up to 100 km from their colonies over open water to forage at the ice edge (Bradstreet 1979). The relatively rare Ivory gulls are closely associated with pack-ice, favouring areas with 70 – 90% ice cover near the ice edge, where they feed on small fish, including juvenile Arctic cod, squid, invertebrates, macro-zooplankton, carrion, offal and animal faeces (e.g. OSPAR Commission, 2009b). Ivory gulls have a low reproductive rate and breeding only takes place if there is sufficient food, which makes the population highly vulnerable to the effects of climate warming (e.g. OSPAR Commission, 2009b). Thick-billed guillemots are relatively long lived and slow to reproduce and has a low resistance to threats including oil pollution, by-catch in and competition with commercial fisheries operations, population declines due to hunting – particularly in Greenland (OSPAR Commission, 2009c). Ivory gull and Thick-billed guillemots are both listed by OSPAR as being under threat and/or decline, (OSPAR Commission, 2008) and in 2011 recommendations for conservation action were agreed (OSPAR Commission, 2011) which will be implemented in conjunction with the circumpolar conservation actions of CAFF (CAFF, 1996; Gilchrist et al., 2008). Marine mammals Several marine mammal species permanently associate with sea ice in the European Arctic. These include polar bear, walrus, and several seal species: bearded, Erignathus barbatus; ringed, Pusa hispida; hooded, Cystophora cristata; and harp seal Pagophilus groenlandicus. Three whale species also occupy Arctic waters year- round – narwhal, Monodon monoceros; beluga whale, Delphinapterus leucas; and bowhead whale, Balaena mysticetus. Polar bears Ursus maritimus are highly specialized for and dependent on the sea ice habitat and are therefore particularly vulnerable to changes in sea ice extent, duration and thickness. They have a circumpolar distribution limited by the southern extent of sea ice. Three subpopulations of polar bears occur in the European high Arctic: the East Greenland, Barents Sea and Arctic Basin sub-populations, all with an unknown population status (CAFF, 2010). Following the young-of-the-year ringed seal distribution, polar bears are most common close to land and over the shelves, however some also occur in the permanent multi-year pack ice of the central Arctic basin (Durner et al., 2009). Due to low reproductive rates and long lifetime, it is expected that the polar bears will not be able to adapt to the current fast warming of the Arctic and become extirpated from most of their range within the next 100 years (Schliebe et al., 2008). Walrusses, Odobenus rosmarus, inhabit the Arctic ice year-round. They are conservative benthic feeders, diving to 80-100 m depth for scaping off the rich mollusc fauna of the continental shelves, and need ice floes ICES Advice 2013, Book 1 257 as resting and nursing platform close to their foraging grounds. Walrusses have been subject to severe hunting pressure from the end of the 18th century to the mid 20th century, and are still hunted today in Greenland (NAMMCO). By 1934, the estimated 70000-80000 individuals of the Atlantic population were reduced to 12001300, with none left on Svalbard (Weslawski et al., 2000). Todays relatively small sub-populations on the East Greenland and Svalbard-Franz Josef Land coasts have recently shown a slightly increasing trend, in the latter case reflecting the full protection of the species since the 1950´s (CAFF, 2010; NAMMCO). Apart from their sensitivity to direct human disturbance and pollution, it is expected that walrusses will suffer from the changing ice conditions (location, thickness for being used as haul-out site) as well as changes in ice-related productivity. The Atlantic subspecies of the bearded seal, Erignathus barbatus occurs south of 85° N from the central Canadian Arctic east to the central Eurasian Arctic, but no population estimates exist (Kovacs, 2008b). Because of their primarily benthic feeding habits they live in ice covered waters overlying the continental shelf. They are typically found in regions of broken free-floating pack ice; in these areas bearded seals prefer to use small and medium sized floes, where they haul out no more than a body length from water and they use leads within shore-fast ice only if suitable pack ice is not available (Kovacs, 2008b, and literature quoted). The Arctic ringed seal Pusa (Phoca) hispida hispida has a very large population size and broad distribution, however, there are concerns that future changes of Arctic sea ice will have a negative impact on the population, some of which have already been documented in some parts of the subspecies range (Kovacs et al., 2008). As the other seals, the ringed seal uses sea ice exclusively as their breeding, moulting and resting (haulout) habitat, and feed on small schooling fish and invertebrates. In a co-evolution with one of their main predators, the polar bear, they developed the ability to create and maintain breathing holes in relatively thick ice, which makes them well adapted to living in fully ice covered waters allover the year. The West Ice (or Is Odden) to the west of Jan Mayen, at approx. 72-73° N, in early spring a stretch of more of less fast drift ice, is of crucial importance as a whelping and moulting area for harp seals and hooded seals (summarised e.g. by ICES, 2008). Discovered in the early 18th century, up to 350000 seals (1920s) were killed per year, decimating the populations from an estimated one million individuals in the 1950s (Ronald et al., 1982) to today´s 70000 and 243000 of hooded and harp seals, respectively (Kovacs, 2008a, c). Hooded seal, Cystophora cristata, is a pack ice species, which is dependent on ice as a substrate for pupping, moulting, and resting and as such is vulnerable to reduction in extent or timing of pack ice formation and retreat, as well as ice edge related changes in productivity (Kovacs, 2008a, and literature quoted). Hooded Seals feed on a wide variety of fish and invertebrates, including species that occur throughout the water column. After breeding an moulting on the West Ice they follow the retreating pack ice to the north, but also spend significant periods of time pelagically, without hauling out (Folkow and Blix 1999) in (Kovacs, 2008a). The northeast Atlantic breeding stock has declined by 85-90 % over the last 40-60 years. The cause of the decline is unknown, but very recent data suggests that it is on-going (30% within 8 years), despite the protective measures that have been taken in the last few years. The species is therefore considered to be vulnerable (Kovacs, 2008a). Harp seals Pagophilus (Phoca) groenlandicus are the most numerous seal species in the Arctic seas. Their reproduction takes place in huge colonies, for example on the pack ice of the ‘‘West Ice’’ north of Jan Mayen, and after the breeding season they follow the retreating pack ice edge northwards up to 85° N, feeding mainly on polar cod under the ice (Kovacs, 2008c) . Narwhals Monodon monoceros primarily inhabit the ice-covered waters of the European Arctic, including the ice sheet off East Greenland (Jefferson et al., 2008b). For two months in summer, they visit the shallow fjords of East Greenland, spending all the rest of the year offshore, in deep ice-covered waters along the continental slope in the Greenland Sea and Arctic Basin (Heide-Jørgensen and Dietz, 1995). Narwhals are deep diving benthic feeders and forage on fish, squid, and shrimp, especially Arctic fish species, such as Greenland halibut, Arctic cod, and polar cod at up to 1500 m depth and mostly in winter. A recent assessment of the sensitivity of all Arctic marine mammals to climate change ranked the narwhal as one of the three most sensitive species, 258 ICES Advice 2013, Book 1 primarily due to its narrow geographic distribution, specialized feeding and habitat choice, and high site fidelity (Laidre et al. 2008 in (Jefferson et al., 2008b)). Bowhead whales Balaena mysticetus are found only in Arctic and subarctic regions and a Svalbard-Barents population occurs from the coast of Greenland across the Greenland Sea to the Russian Arctic. They spend all of their lives in and near openings in the pack ice feeding on small to medium-sized zooplankton. They migrate to the high Arctic in summer, and retreat southward in winter with the advancing ice edge (Moore and Reeves 1993 in (Reilly et al., 2008)). Whaling has decimated the original bowhead whale populations to be rare nowadays, listed by OSPAR as being under threat and/or decline (OSPAR Commission, 2008). The species is considered to be very sensitive to changes in the ice-related ecosystem as well as sound disturbance, possible consequences of a progressive reduction of ice cover (OSPAR Commission, 2009a). Belugas Delphinapterus leucas prefer coastal and continental shelf waters with a broken-up ice cover. They have never been surveyed around Svalbard. Pods numbering into the thousands are sighted irregularly around the archipelago, and pods ranging from a few to a few hundred individuals are seen regularly (Gjertz and Wiig 1994; Kovacs and Lydersen 2006 in (Jefferson et al., 2008a)). Little is known about the populations of the larger fauna in the Central Arctic Basin over the deepsea basins and ridges. But it is not likely that it is currently an area of great abundance - too far from the coastal nesting sites of marine birds, and over too deep water to allow feeding on benthos, as most of the larger mammals would need, and currently of too low plankton production to feed the large whales. All of these groups have their distribution center along the continental shelves presently - however, following the receeding ice edge out to the central Arctic basin may be one of the options for the future. Feature condition, and future outlook This high Arctic region is particularly vulnerable to the the loss of ice cover and other effects of the anticipated global warming, including elevated UV radiation levels (Agustí, 2008). (Wassmann et al., 2010) summarise what changes may be expected within the subarctic/Arctic region: • northward displacement (range shifts) of subarctic and temperate species, and cross-Arctic transport of organisms; • increased abundance and reproductive output of subarctic species, decline and reduced reproductive success of some Arctic species associated with the ice and species now preyed upon by predators whose preferred prey have declined; • increased growth of some subarctic species and primary producers, and reduced growth and condition of animals that are bound to, associated with, or born on the ice; • anomalous behaviour of ice-bound, ice-associated, or ice-born animals with earlier spring events and delayed fall events; • changes in community structure due to range shifts of predators resulting in changes in the predator– prey linkages in the trophic network. (Wassmann, 2008) expects radical changes in the productivity, functional relationships and biodiversity of the Arctic Ocean. He suggests that a warmer climate with less ice cover will result in greater primary production, a reduction of the stratified water masses to the south, changes in the relationship between biological processes in the water column and the sediments, a reduction in niches for higher trophic levels and a displacement of Arctic by boreal species. On the shelves, increased sediment discharges are expected to lower the primary production due to higher turbidity, and enhance the organic input to the deep ocean. A more extensive review of expected or suspected consequences of climate change for the marine system of the Arctic is given in (Loeng et al., 2005). Figure 3, extracted from (Gill et al., 2011), presents the conceptual ideas about possible Arctic ecosystem changes mediated by human impact: ICES Advice 2013, Book 1 259 The normal situation shown in the upper left panel consists of ice-dependent species and species that tolerate a broader range of temperatures and are found in waters with little or no sea ice. Primary production occurs in phytoplankton (small dots in the figure) in ice-free waters and in ice-attached algae and phytoplankton in icecovered waters. Phytoplankton (small t-shaped symbols in the figure) and ice algae are the main food sources for zooplankton and benthic animals. The fish community consists of both pelagic and demersal species. Several mammals are ice-associated, including polar bears and several species of seals. A number of sea bird species are also primarily associated with ice-covered waters. At moderate temperature increases (upper right) populations of ice-dependent species are expected to decline as sea ice declines, and sub-Arctic species are expected to move northwards. Arctic benthic species are expected to decline, especially if their distributions are pushed close to or beyond the continental slope. The expected effects from fisheries relate to the continental shelves. Two major effects are reductions in populations of benthic organisms due to disturbance from bottom trawling and removal of large individuals in targeted fish stocks. In addition, the size of targeted stocks, both demersal and pelagic, may be reduced. In addition, the effects of ocean acidification are considered (lower right). Ocean acidification will result in depletion of carbonate phases such as aragonite and calcite. This will alter the structure and function of calcareous organisms, particularly at lower trophic levels. Changes in pH can also alter metabolic processes in a range of organisms. It is not known how these changes will propagate to higher trophic levels, but the effects could be substantial. c.f. Figure 3: Conceptual models showing potential impacts on Arctic marine ecosystems under different scenarios (Gill et al., 2011). (Gill et al., 2011) conclude that the central part of the Arctic Basin is not a region for fisheries or oil and gas exploration. However, this region has played and will continue to play a very important role in the redistribution of pollutants, due to ice drift and/or currents between coastal and shelf areas and the Arctic Basin peripheries, far from sources of pollution. 260 ICES Advice 2013, Book 1 Assessment against CBD EBSA Criteria Table 1. relation of each of the CBD criteria to the proposed area relating to the best available science. Note that a candidate EBSA may qualify on the basis of one or more of the criteria, the boundaries of the EBSA need not be defined with exact precision. CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Know Uniqueness or The area contains either (i) unique (“the only one rarity of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Low Some High x Explanation for ranking Arctic sea ice, in particular the multiyear ice of the Central Arctic is globally unique and hosts endemic species such as the Gammarid amphipod Gammarus wilkitzki and sea ice meiofauna which will disappear with the melting of the ice. Polar bears, walrusses, bowhead whales, narwhales, belugas, several seal species and many bird species are endemic to the high Arctic ice. While sea ice species such as G. wilkitzki are not endemic to the proposed EBSA they are endemic to the Arctic and unique within the OSPAR area Special Areas that are required for a population to importance for survive and thrive life-history stages of species x Explanation for ranking Sea ice is essential for its sympagic fauna, and to some extent also for the pelagic associated fauna which also depends on the right timing of biomass production (match/mismatch with bloom periods). The marginal ice zone and other openings in the ice are essential feeding grounds for a large number of ice-associated species which exploit the seasonallly high production there. At present the area covered by the proposed EBSA is ice-covered throughout the summer but although there is no marginal ice zone there will be an ice zone community present, thus the sea ice is essential to maintain the sympagic biological community and associated ecosystem functions. ICES Advice 2013, Book 1 261 Importance for threatened, endangered or declining species and/or habitats Area containing habitat for the survival and recovery of endangered, threatened, declining species or area with significant assemblages of such species x Explanation for ranking The high arctic ice hosts endemic species such as the Gammarid amphipod Gammarus wilkitzki and sea ice meiofauna which will disappear with the melting of the ice. Many of the obligatory ice-related species are listed as vulnerable by IUCN, and/or listed as under threat and/or decline by OSPAR, examples include the Ivory gull, thick-billed guillemot, bowhead whale, hooded seal and polar bear. With the overall trend of retreating sea ice extent, the proposed EBSA may become increasingly important for all ice-dependent species in the future. Vulnerability, fragility, sensitivity, or slow recovery Areas that contain a relatively high proportion of sensitive habitats, biotopes or species that are functionally fragile (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery x Explanation for ranking The ice-related foodweb and ecosystem is highly sensitive to the ecological consequences of a warming climate. Beyond this the Arctic is at the forefront of the impacts of ocean acidification (Wicks & Roberts 2012). The largest changes in ocean pH will occur in the Arctic Ocean, with complete undersaturation of the Arctic Ocean water column predicted before the end of this century (Steinacher et al. 2009). Many of the seabird and mammal populations are particularly sensitive to changes due to their already low population numbers, and low fertility. If the retreat of the ice to the north will lead to increased shipping and oil and gas exploitation in Arctic waters, the increased risk of spills would also pose a potential hazard for example for guillemots, which are extremely susceptible to mortality from oil pollution (CAFF, 2010). In addition, some species like Ivory gull are sensitive to an increased heavy metal load in their prey. Biological productivity Area containing species, populations or communities with comparatively higher natural biological productivity Explanation for ranking This criterion was not evaluated in the OSPAR/NEAFC/CBD Workshop. ICES did not have enough information to evaluate this criterion. Biological diversity 262 Area contains comparatively higher diversity of ecosystems, habitats, communities, or species, or has higher genetic diversity ICES Advice 2013, Book 1 Explanation for ranking This criterion was not evaluated in the OSPAR/NEAFC/CBD Workshop. ICES did not have enough information to evaluate this criterion. References Aagaard, K., 1989. A synthesis of Arctic Ocean circulation. Rapport Proces et Verbeaux Réunion du Conseil international pour l'Exploration de la Mer 188, 11-22. Aagaard, K., Coachman, L.K., 1968. The East Greenland Current north of Denmark Strait: Part II. Arctic 21, 267-290. Aagaard, K., Swift, J.H., Carmack, E.C., 1985. Thermohaline circulation in the Arctic mediterranean seas. Journal of Geophysical Research 90, 4833-4846. Agustí, S., 2008. Impacts of increasing ultraviolet radiation on the polar oceans. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.) Fundación BBVA pp. 25-46. Anderson, L.G., Jones, E.P., Koltermann, K.P., Schlosser, P., Swift, J.H., Wallace, D.W.R., 1989. The first oceanographic section across the Nansen Basin in the Arctic Ocean. Deep Sea Research 36, 475482. Angelen, J.H.v., Broeke, M.R.v.d., Kwok, R., 2011. The Greenland Sea Jet: A mechanism for wind‐driven sea ice export through Fram Strait. Geophysical Research Letters 38 (L12805). Anisimov, O.A., Vaughan, D.G., Callaghan, T.V., Furgal, C., Marchant, H., Prowse, T.D., Vilhjálmsson, H., Walsh, J.E., 2007. Polar regions (Arctic and Antarctic). In: Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Parry, M.L., Canziani, O.F., Palutikof, J.P., Linden, P.J.v.d., Hanson, C.E. (Eds.)Cambridge University Press, Cambridge pp. 653-685. Arndt, C., E. , Pavlova, O., 2005. Origin and fate of ice fauna in the Fram Strait and Svalbard area. Marine Ecology Progress Series 301, 55-66. Auel, H., Hagen, W., 2002. Mesozooplankton community structure, abundance and biomass in the central Arctic Ocean. Marine Biology 140, 1013-1021. Bluhm, B.A., Gebruk, A.V., Gradinger, R., Hopcroft, R.R., Huettmann, F., Kosobokova, K.N., Sirenko, B.I., Weslawski, J.M., 2011. Arctic marine biodiversity: An update of species richness and examples of biodiversity change. Oceanography 24 (3), 232-248. Bradstreet, M.S.M., Cross, W.E., 1982. Trophic relationships at high Arctic ice edges. Arctic 35 (1), 1-12. CAFF, 1996. International Murre conservation strategy and action plan. CAFF International Secretariat, CAFF Circumpolar Seabird Working Group, Akureyri, Iceland, pp. 1-16. CAFF, 2010. Arctic Biodiversity Trends 2010. Selected indicators of change. CAFF International Secretariat, , Akureyri, Iceland. Carey, A.G.I., 1985. Marine Ice Fauna. In: Arctic Sea Ice Biota. A., H.R. (Ed.)CRC Press, Boca Raton. Florida pp. 17-190 Crawford, R.E., Jorgenson, J.K., 1993. Schooling behaviour of arctic cod, Boreogadus saida in relation to drifting pack ice. Environmental Biology of Fishes 36 (4), 345-357. Durner, G.M., Douglas, D.C., Nielson, R.M., Amstrup, S.C., McDonald, T.L., Stirling, I., Mauritzen, M., Born, E.W., Wiig, Ø., Deweaver, E., Serreze, M.C., Belikov, S.E., Holland, M.M., Maslanik, J., Aars, J., Bailey, D.A., Derocher, A.E., 2009. Predicting 21st-century polar bear habitat distribution from global climate models. Ecological Monographs 79 (1), 25-58. ICES Advice 2013, Book 1 263 Edmonds, H.N., Michael, P.J., Baker, E.T., Connelly, D.P., Snow, J.E., Langmuir, C.H., Dick, H.J.B., Mühe, R., German, C.R., Graham, D.W., 2003. Discovery of abundant hydrothermal venting on the ultraslowspreading Gakkel ridge in the Arctic Ocean. Nature 421, 252-256. Gilchrist, G., Strøm, H., Gavrilo, M.V., Mosbech, A., 2008. International Ivory Gull conservation strategy and action plan. CAFF International Secretariat, Circumpolar Seabird Group (CBird). CAFF Technical Report No. 18. Gill, M.J., Crane, K., Hindrum, R., Arneberg, P., Bysveen, I., Denisenko, N.V., Gofman, V., Grant-Friedman, A., Gudmundsson, G., Hopcroft, R.R., Iken, K., Labansen, A., Liubina, O.S., Melnikov, I.A., Moore, S.E., Reist, J.D., Sirenko, B.I., Stow, J., Ugarte, F., Vongraven, D., Watkins, J., 2011. Arctic Marine Biodiversity Monitoring Plan (CBMP-MARINE PLAN), CAFF Monitoring Series Report No.3, April 2011. CAFF International Secretariat,, Akureyri, Iceland. Gradinger, R., 1995. Climate change and biological oceanography of the Arctic Ocean. Phil. Trans. R. Soc. A 352, 277-286. Gradinger, R., Bluhm, B.A., 2004. In situ observations on the distribution and behavior of amphipods and Arctic cod (Boreogadus saida) under the sea ice of the high Arctic Canadian Basin. Polar Biology 27, 595-603. Gradinger, R., Friedrich, C., Spindler, M., 1999. Abundance, biomass and composition of the sea ice biota of the Greenland Sea pack ice. Deep Sea Research 46, 1457-1472. Gradinger, R., Spindler, M., Henschel, D., 1991. Development o Arctic sea-ice organisms under graded snow cover. In: Proceedings of the Pro Mare Symposium on Polar Marine Ecology. Sakshaug, E., E., H.C.C., Øritsland, N.A. (Eds.), Polar Research 10 (1), Trondheim pp. 295-307. Gradinger, R.R., Baumann, M.E.M., 1991. Distribution of phytoplankton communities in relation to the largescale hydrographical regime in the Fram Strait. Mar. Biol. 111, 311-321. Grainger, E.H., 1989. Vertical distribution of zooplankton in the central Arctic Ocean. In: Proc 6th Conf Comite´Arctique Int 1985. Rey, L., Alexander, V. (Eds.) Brill Leiden pp. 48–60. Heide-Jørgensen, M.P., Dietz, R., 1995. Some characteristics of narwhal, Monodon monoceros, diving behaviour in Baffin Bay. Canadian Journal of Zoology 73, 2106-2119. Hirche, H.J., 1997. Life cycle of the copepod Calanus hyperboreus in the Greenland Sea. Marine Biology 128 (4), 607-618. Hirche, H.J., Baumann, M.E.M., Kattner, G., Gradinger, R., 1991. Plankton distribution and the impact of copepod grazing on primary production in Fram Strait, Greenland Sea. Journal of Marine Systems 2 (3-4), 477-494. Hirche, H.J., Mumm, N., 1992. Distribution of dominant copepods in the Nansen Basin, Arctic Ocean, in summer. Deep Sea Research Part A. Oceanographic Research Papers 39 (2, Part 1), S485-S505. Hirche, H.J., Muyakshin, S., Klages, M., Auel, H., 2006. Aggregation of the Arctic copepod Calanus hyperboreus over the ocean floor of the Greenland Sea. Deep Sea Research Part I: Oceanographic Research Papers 53 (2), 310-320. Horner, R., Ackley, S.F., Dieckmann, G.S., Gulliksen, B., Hoshiai, T., Legendre, L., Melnikov, I.A., Reeburgh, W.S., Spindler, M., Sullivan, C.W., 1992. Ecology of sea ice biota. Habitat, terminology, and methodology. Polar Biology 12 (3), 417-427. Hunt, G.L.J., 1990. The pelagic distribution of marine birds in a heterogeneous environment. Polar Research 8, 43-54. ICES, 2008. Report of the ICES Advisory Committee In: ICES Advice, Book 3, The Barents and the Norwegian SEa. Jakubas, D., Iliszko, L., Wojczulanis-Jakubas, K., Stempniewicz, L., Online 03 June 2011. Foraging by little auks in the distant marginal sea ice zone during the chick-rearing period. Polar Biology, 1-9. 264 ICES Advice 2013, Book 1 Jefferson, T.A., Karczmarski, L., Laidre, K., O’Corry-Crowe, G., Reeves, R.R., Rojas-Bracho, L., Secchi, E.R., Slooten, E., Smith, B.D., Wang, J.Y., Zhou, K., 2008a. Delphinapterus leucas In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Jefferson, T.A., Karczmarski, L., Laidre, K., O’Corry-Crowe, G., Reeves, R.R., Rojas-Bracho, L., Secchi, E.R., Slooten, E., Smith, B.D., Wang, J.Y., Zhou, K., 2008b. Monodon monoceros. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Joiris, C., Falck, E., 2011. Summer at-sea distribution of little auks Alle alle and harp seals Pagophilus (Phoca) groenlandica in the Fram Strait and the Greenland Sea: impact of small-scale hydrological events. Polar Biology 34 (4), 541-548. Joiris, C.R., 2000. Summer at-sea distribution of seabirds and marine mammals in polar ecosystems: a comparison between the European Arctic seas and the Weddell Sea, Antarctica. Journal of Marine Systems 27, 267-276. Klitgaard, A.B., Tendal, O.S., 2004. Distribution and species composition of mass occurrences of large-sized sponges in the northeast Atlantic. Progress in Oceanography 61, 57-98. Kosobokova, K.N., 1982. Composition and distribution of the biomass of zooplankton in the central Arctic Basin. Oceanology 22, 744-750. Kovacs, K., 2008a. Cystophora cristata. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Kovacs, K., 2008b. Erignathus barbatus. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Kovacs, K., 2008c. Pagophilus groenlandicus. IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Kovacs, K., Lowry, L., Härkönen, T., 2008. Pusa hispida. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Kwok, R., 2009. Outflow of Arctic Ocean sea ice into the Greenland and Barents seas: 1979-2007. Journal of Climate 22, 2438-2456. Legendre, L., Ackley, S.F., Dieckmann, G.S., Gulliksen, B., Horner, R., Hoshiai, T., Melnikov, I.A., Reeburgh, W.S., Spindler, M., Sullivan, C.W., 1992. Ecology of sea ice biota. Polar Biology 12 (3), 429-444. Leu, E., Søreide, J.E., Hessen, D.O., Falk-Petersen, S., Berge, J., 2011. Consequences of changing sea ice cover for primary and secondary producers in the European Arctic shelf seas: timing, quantity, and quality. Progress in Oceanography 90, 18-32. Loeng, H., Brander, K., Carmack, E.C., Denisenko, S., Drinkwater, K., Hansen, B., Kovacs, K., Livingston, P., McLaughlin, F., Sakshaug, E., 2005. Marine systems. In: Arctic Climate Impact Assessment, ACIA. Symon, C., Arrisand, L., Heal, B. (Eds.),Cambridge University Press, Cambridge pp. 453-538. Mauritzen, C., Hansen, E., Andersson, M., Berx, B., Beszczynska-Möller, A., Burud, I., Christensen, K.H., Debernard, J., de Steur, L., Dodd, P., Gerland, S., Godøy, Ø., Hansen, B., Hudson, S., Høydalsvik, F., Ingvaldsen, R., Isachsen, P.E., Kasajima, Y., Koszalka, I., Kovacs, K.M., Køltzow, M., LaCasce, J., Lee, C.M., Lavergne, T., Lydersen, C., Nicolaus, M., Nilsen, F., Nøst, O.A., Orvik, K.A., Reigstad, M., Schyberg, H., Seuthe, L., Skagseth, Ø., Skar∂hamar, J., Skogseth, R., Sperrevik, A., Svensen, C., Søiland, H., Teigen, S.H., Tverberg, V., Wexels Riser, C., 2011. Closing the loop - Approaches to monitoring the state of the Arctic Mediterranean during the International Polar Year 2007-2008. Progress in Oceanography 90 (1-4), 62-89. Maykut, G.A., 1985. The ice environment. In: Sea-ice biota. Horner, R. (Ed.)CRC Press, Boca Raton pp. 2182. ICES Advice 2013, Book 1 265 Melle, W., Ellertsen, B., Skjoldal, H.R., 2005. Zooplankton: The link to higher trophic levels. In: The Nordic Seas: An integrated perspective oceanography, climatology, biogeochemistry, and modelling. . Drange, H., Dokken, T., Furevik, T., Gerdes, R., Berger, W. (Eds.)Geophysical Monograph Series 158 pp. 137-202. Mumm, N., Auel, H., Hanssen, H., Hagen, W., Richter, C., Hirche, H.J., 1998. Breaking the ice: large-scale distribution of mesozooplankton after a decade of Arctic and transpolar cruises. Polar Biology 20 (3), 189-197. NAMMCO, The Atlantic Walrus. North Atlantic Marine Mammal Commission. Status of Marine Mammals in the North Atlantic, Tromsø, pp. 1-7. OSPAR Commission, 2008. OSPAR List of Threatened and/or Declining Species andHabitats. Reference number 2008-6. http://www.ospar.org/documents/dbase/decrecs/agreements/0806e_ospar%20list%20species%20and%20habitats.doc. OSPAR Commission, 2009a. Background Document for Bowhead whale Balaena mysticetus. OSPAR Commission, Biodiversity Series 494/2010, pp. 1-20. OSPAR Commission, 2009b. Background Document for Ivory gull Pagophila eburnea. OSPAR Commission, Biodiversity Series 410/2009, pp. 1-16. OSPAR Commission, 2009c. Background Document for Thick-billed murre Uria lomvia. OSPAR Commission, Biodiversity Series 416/2009, pp. 1-20. OSPAR Commission, 2011. Meeting of the OSPAR Commission (OSPAR) London: 20-24 June 2011. Summary Record OSPAR 11/20/1-E. OSPAR Commission, London. Paquette, R., Bourke, R., Newton, J., Perdue, W., 1985. The East Greenland Polar Front in autumn. Journal of Geophysical Research 90 (C3), 4866-4882. Piepenburg, D., 2005. Recent research on Arctic benthos: common notions need to be revised. Polar Biology 28 (10), 733-755. Reilly, S.B., Bannister, J.L., Best, P.B., Brown, M., , Brownell Jr., R.L., Butterworth, D.S., Clapham, P.J., Cooke, J., Donovan, G.P., Urbán, J., Zerbini, A.N., 2008. Balaena mysticetus. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . www.iucnredlist.org Downloaded on 31 August 2011. Renaud, P.E., Caroll, M.L., Ambrose, W.G.J., 2008. Effects of global warming on Arctic seafloor communities and its consequences for higher trophic levels. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.)Fundación BBVA pp. 141-177. Rey, F., 2004. Phytoplankton: the grass of the sea. In: The Norwegian Sea Ecosystem. Skjoldal, H.R. (Ed.)Tapir Academic Press, Trondheim, Norway pp. 97-136. Ronald, K., Healey, P.J., Fisher, H.D., 1982. The harp seal, Pagophilus groenlandicus. In: Small cetaceans, seals, sirenians and otters. FAO Fisheries Series No. 5, Vol. IV, Food and Agriculture Organisation of the United Nations. Workding Party on Mammals. Schliebe, S., Wiig, Ø., Derocher, A.E., Lunn, N., 2008. Ursus maritimus. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. www.iucnredlist.org Downloaded on 31 August 2011. Smith Jr, W.O., Barber, D., 2007. Polynyas and climate change: a view to the future. In: Polynays, windows to the world. Halpern, D. (Ed.), Elsevier Oceanography Serie 74, Elsevier, Amsterdam pp. 411-420. Smith Jr, W.O., Baumann, M.E.M., Wilson, D.L., Aletsee, L., 1987. Phytoplankton biomass and productivity in the Marginal Ice Zone of the Fram Strait during summer 1984. Journal of Geophysical Research 92 (C7), 6777-6786. Smith, W.O.J., 1987. Phytoplankton dynamics in the marginal ice zones. Oceanography and Marine Biology Annual Review 25, 11-38. 266 ICES Advice 2013, Book 1 Smith, W.O.J., Sakshaug, E., 1990. Polar phytoplankton. In: Polar oceanography. Part B. Chemistry, biology and geology. Smith Jr, W.O. (Ed.) Academic Press, San Diego pp. 477-525. Steinacher, M., Joos, F., Frölicher, T., Plattner, G. & Doney, S.c. 2009. Imminent ocean acidification in the Arctic projected with the NcAR global coupled carbon cycle-climate model. Biogeosciences 6, 515– 533. Süfke, L., Piepenburg, D., Dorrien, C.C.v., 1998. Body size, sex ratio and diet composition of Arctogadus glacialis (Peters, 1874) (Pisces: Gadidae) in the Northeast Water Polynya (Greenland). Polar Biology 20, 357-363. Wadhams, P., 1981. The ice cover in the Greenland and Norwegian Seas. Rev. Geophys. Space Physics 19, 345-393. Wassmann, P., 2008. Impacts of global warming on Arctic pelagic ecosystems and processes. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.)Fundación BBVA pp. 113-148. Wassmann, P., 2011. Arctic marine ecosystems in an era of rapid climate change. Progress in Oceanography 90, 1-17. Wassmann, P., Duarte, C.M., Agustí, S., Seijr, M., 2010. Footprints of climate change in the Arctic Marine Ecosystem. Biological Global Change. Weslawski, J.M., Hacquebord, L., Stempniewicz, L., Malinga, M., 2000. Greenland whales and walruses in the Svalbard food web before and after exploitation. Oceanologia 42 (1), 37-56. Wheeler, P.A., Gosselin, M., Sherr, E., Thibault, D., Kirchman, D.L., Benner, R., Whitledge, T.E., 1996. Active cycling of organic carbon in the central Arctic Ocean. Nature 380, 697-699. Wicks, L., Roberts, J.M. (2012) Benthic invertebrates in a high CO2 world. Oceanography & Marine Biology: An Annual Review 50: 127 -188 Maps and Figures ICES Advice 2013, Book 1 267 Figure 1: Location of the Ecologically or biologically significant areas (EBSA) proposed by WWF in September 2011. The position of the Arctic and polar fronts was redrawn after (Rey, 2004, Fig. 5.7). 268 ICES Advice 2013, Book 1 Figure 2: Modelled ice age distribution in 1985-2000 (left) compared to February 2008 (right) (CAFF, 2010). Figure 3: Conceptual models showing potential impacts on Arctic marine ecosystems under different scenarios (Gill et al., 2011). ICES Advice 2013, Book 1 269 1.5.6.6 Special request, Advice June 2013 ECOREGION SUBJECT General advice OSPAR/NEAFC special request on existing and potential new management measures for ecologically and biologically significant areas (EBSAs) Advice summary ICES provides a summary of management measures already implemented within ecologically and biologically significant areas (EBSAs). ICES notes that there are numerous measures already in place by both NEAFC and by OSPAR (as requests to Contracting Parties) and by other competent management authorities, and that both generic and targeted management measures may offer protection to the features that would qualify as meeting EBSA criteria. The performance of these measures has not been evaluated and it was not possible to assess whether these measures are sufficient to protect all of the features that would qualify under EBSA criteria from all potential threats, or whether further measures are required. ICES notes that some specific areas may need further protective measures. Request c) For the use of OSPAR and NEAFC Contracting Parties, as appropriate, provide a separate document with additional relevant information and up-to-date accounts of the relevant management measures already implemented within the individual EBSAs. d) In a separate document, describe potential new management measures within individual EBSAs if appropriate. ICES advice ICES summarized the existing management measures or recommendations of bodies with regulatory or coordinating authority for the major sectoral activities in the OSPAR and NEAFC areas, and, where they exist, those of the authorities for conservation and protection of special components of biodiversity. Some of the measures apply directly, while others require implementing legislation by national authorities (or in some cases by the EU). The power to require vessels to operate in specified ways lies with the Flag State of each vessel. ICES does not have information on the performance of many of the existing measures, and consequently has not been able to evaluate whether these management measures would be sufficient to protect all of the features that would qualify under EBSA criteria from all potential threats. Nonetheless a range of generic and spatially targeted protective management measures already exist within the EBSA areas. There are, however, some exceptions that may need improved protective measures: • • • Josephine Seamount The Hatton–Rockall basin The existing fishing areas that lie north of the Azores on the Mid-Atlantic Ridge. One of these is a potential threat to hydrothermal vents in the area. ICES advises that a systematic review of the performance of the existing measures and how they interact to reduce threats to EBSAs is needed before any new management measures could be sensibly considered through a gap analysis. ICES makes the following general suggestions that are likely to improve the protection of EBSAs in the Northeast Atlantic: • • • • Protective measures for vulnerable marine ecosystems (VMEs) should be made permanent. In many cases these measures are only temporary. This is not appropriate for the long-lived, sessile organisms within VMEs. Accessibility of existing ecological and fisheries data should be improved through a concerted effort between relevant authorities. Coordination of competent authorities should be improved in the selection and adoption of measures in order to help ensure efficient implementation. Observer coverage of fishing vessels operating within EBSAs should be increased. Management measures North-East Atlantic Fisheries Commission (NEAFC) Table 1.5.6.6.1 summarizes NEAFC regulations for fishing activities within nine of the ten proposed EBSAs. The Arctic ice habitat lies outside the regulatory area of NEAFC. The ten proposed EBSA areas are shown in Figure 1.5.6.6.1, together with existing bottom fisheries closures enforced by NEAFC and high seas MPAs established by OSPAR. Since 270 ICES Advice 2013, Book 1 2005, NEAFC has closed 14 areas to bottom fishing to protect VMEs. In addition (for fisheries management rather than VME protection), NEAFC has closed an area for haddock on Rockall Bank since 2001 and enforced a seasonal closed area on the Reykjanes Ridge to protect a blue ling spawning ground. In total the area closed to protect VMEs is in excess of 600 000 square kilometres and covers all areas where the presence of VMEs have been validated up to the end of 2012. ICES has recently provided further advice on the boundaries of some of these VMEs (ICES, 2013a). NEAFC has different bottom-fishing regulations for existing fisheries areas and new fishing areas. Five of the proposed EBSAs are covered by the ‘New Fishing Area’ regulations only. Exploratory fishing in new areas is only authorized under strict conditions that include full observer coverage. There have been no applications for exploratory fishing since the regulations were adopted. Several points are notable: • • All the NEAFC closures lie within the proposed EBSAs. The majority of the overall area covered by the ten proposed EBSAs is classed by NEAFC as ‘new fishing areas’ and thus subject to strict regulation. Exceptions that are not ‘new fishing areas’ include an existing fishing area within the proposed EBSA 1 (Reykjanes Ridge), four within the proposed EBSA 3 (Mid-Atlantic Ridge north of Azores), and a further four within the proposed EBSA 4 (Hatton–Rockall area). The four major fisheries in the NEAFC Regulatory Area (herring, mackerel, blue whiting, and redfish) are pelagic trawl fisheries and therefore unlikely to pose threats to seabirds, taxa on the OSPAR list of threatened and declining species, or VMEs. In addition NEAFC regulations include: • • A ban on the use of gillnet at depths greater than 200 m. A ban on directed fisheries for 17 different species of deep-sea sharks. ICES Advice 2013, Book 1 271 Figure 1.5.6.6.1 272 Map of the southern NEAFC Regulatory Area showing proposed EBSAs (Grey polygons; 1 = Reykjanes Ridge, 2 = Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone, 3 = Mid-Atlantic Ridge north of the Azores, 4 = Hatton–Rockall Plateau, 5 = Pedro Nunes and Hugo de Lacerda Seamounts, 6 = Northeast Azores–Biscay Rise, 7 = Evlanov Seamount, 8 = West of Azores). Existing NEAFC fishing areas are shown in white, NEAFC bottom fishery closures in pink, and OSPAR High Seas MPAs in green. ICES Advice 2013, Book 1 Table 1.5.6.6.1 Management measures adopted by NEAFC in each proposed EBSA. NEAFC closed areas or Rationale for NEAFC fishing regulations in the management measure proposed area Area 1. Reykjanes Ridge south of Iceland EEZ 1) Northern Mid-Atlantic 1,3,5) Protection of VME Ridge (MAR)1 (cold-water corals). 2) Seasonal blue ling 2) Protection for closure 2 spawning/aggregation 3)Existing fishing areas1 areas of blue ling. 4)New fishing areas1 NEAFC management regulations 1) Area closed to bottom trawling and fishing with static gear, including bottom-set gillnets and longlines 2009– 2015. 2) This area is closed for fishing blue ling from 15 February to 15 April until 2016. 3) Deep-sea demersal fisheries regulations: Certain gears are banned (gillnets) and actions against ghost fishing and lost gear are in place. 4) Authorization to go to new fishing areas follows a strict exploratory fishing protocol. Area 2. Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the Mid-Atlantic Ridge Middle MAR1 Protection of VME (coldArea closed to bottom trawling and fishing with static water corals). gear, including bottom set gillnets and long-lines 20092015, including Charlie-Gibbs Fracture Zone and the Subpolar frontal zone. Area 3. Mid-Atlantic Ridge north of the Azores 1)Southern MAR; Altair Protection of VME (cold1) Area closed to bottom trawling and fishing with static Seamount, Antialtair water corals). gear, including bottom-set gillnets and longlines 2009– Seamount1 2015. 2)Existing fishing areas1 2) Deep-sea demersal fisheries regulations: Certain gears 3)New fishing areas1 are banned (gillnets) and actions against ghost fishing and lost gear are in place. 3) Authorization to go to new fishing areas follows a strict exploratory fishing protocol. Area 4. The Hatton and Rockall banks and Hatton–Rockall Basin 1) Rockall Haddock Box3 1) Haddock Box closed to 1) NEAFC has banned bottom trawling in this area. 2) Hatton4 protect juvenile haddock. 2–7) Area closed to bottom trawling and fishing with 2–7) Protection of VME 3) West Rockall Mounds static gear, including bottom-set gillnets and longlines (corals, coral reefs, and closure4 until 2015. 4) NW Rockall closure4 sponge grounds). 8) Deep-sea demersal fisheries regulations: Certain gears 5) SW Rockall closure4 7) Geomorphology. are banned (gillnets) and actions against ghost fishing 6) Logachev Mounds and lost gear are in place. closure 4 9) Authorization to go to new fishing areas follows a 7) Edora Bank closure5 strict exploratory fishing protocol. 8) Existing fishing areas1 9) New fishing areas1 ICES Advice 2013, Book 1 273 NEAFC closed areas or Rationale for NEAFC NEAFC management regulations fishing regulations in the management measure proposed area Area 5. Around Pedro Nunes and Hugo de Lacerda Seamounts – IBA MAO3 Area 6. Northeast Azores–Biscay Rise – IBA MAO3 Area 7. Evlanov Seamount Region Area 8. Northwest of Azores EEZ Area 9. The Arctic Front – Greenland/Norwegian Seas New fishing areas1 In most instances there is not enough research or Observers shall collect data in data to identify VMEs in “new fishing areas”. To accordance with a Vulnerable reduce risks to VMEs to a minimum these areas Marine Ecosystem Data are closed to normal commercial bottom fisheries Collection Protocol. under normal authorizations to fish from the Contracting Parties of NEAFC. Exploratory Bottom Fisheries Protocol. Vessels authorized under this protocol must have an observer on board. Exploratory fishing provides an opportunity to gather additional data on benthos and fish communities using industry vessels, and under strict controls on fishing. Area 10. The Arctic Ice habitat – multiyear ice, seasonal ice, and marginal ice zone This proposed area is outside of the NEAFC Regulatory Area. The following measures can be found at http://neafc.org/measures: 6 7 8 9 10 Rec. na 2011 on regulating bottom fishing as amended by Rec. 12 2013. Rec. 05 2013 Blue Ling seasonal closure. Rec. 03 2013 Rockall Haddock. Rec. 09 2013 Rockall Hatton VME closures. Rec. 08 2013 Edora Bank VME closure. International Commission for the Conservation of Atlantic Tuna (ICCAT) ICCAT has recommendations to mitigate the bycatch of seabirds, turtles, and pelagic shark species, but there are no specific spatial management measures relevant to the proposed EBSAs. OSPAR A range of measures have been adopted by the OSPAR Commission within the proposed EBSAs. These include legally binding decisions, which have been used for the establishment of seven marine protected areas (MPAs) in areas beyond national jurisdiction. Five of these lie within the proposed EBSAs. Two others lie outside the proposed EBSAs: the Milne Seamount complex MPA in the southwest of the OSPAR Area and the Josephine Seamount High Seas MPA in the southeast (Figure 1.5.6.6.1). For each MPA, OSPAR has also agreed recommendations for their management calling on Contracting Parties to undertake certain management actions within the competence of OSPAR (Table 1.5.6.6.2). In addition to these are three types of measures that are not spatially explicit and that apply across the OSPAR maritime area in the Northeast Atlantic (Table 1.5.6.6.3). These include a series of recommendations for the protection and conservation of threatened and/or declining species; a recommendation calling on Contracting Parties to specifically take into account OSPAR’s Listed of species and habitats in the conduct of Environmental Impact Assessments; as well as a code of conduct for undertaking marine research. 274 ICES Advice 2013, Book 1 Table 1.5.6.6.2 Measures adopted by the OSPAR Commission specific to proposed EBSAs. Proposed area meeting the EBSA criteria Charlie-Gibbs Fracture Zone and Subpolar Frontal Zone of the MidAtlantic Ridge Mid-Atlantic Ridge north of the Azores Table 1.5.6.6.3 • • • • • • • • • • • • Spatial management measures in place for specific sub-areas within the proposed EBSA (1.a) OSPAR Decision 2010/2 on the establishment of the Charlie-Gibbs South MPA (1.b) OSPAR Recommendation 2010/13 on the management of the Charlie-Gibbs South MPA (2.a) OSPAR Decision 2012/1 on the establishment of the Charlie-Gibbs North High Seas MPA (2.b) OSPAR Recommendation 2012/1 on the management of the Charlie-Gibbs North High Seas MPA (1.a) OSPAR Decision 2010/6 on the establishment of the Mid-Atlantic Ridge north of the Azores High Seas MPA (1.b) OSPAR Recommendation 2010/17 on the management of the Mid-Atlantic Ridge north of the Azores High Seas MPA (2.a) OSPAR Decision 2010/3 on the establishment of the Altair Seamount High Seas MPA (2.b) OSPAR Recommendation 2010/14 on the management of the Altair Seamount High Seas MPA (3.a) OSPAR Decision 2010/4 on the establishment of the Antialtair Seamount High Seas MPA (3.b) OSPAR Recommendation 2010/15 on the management of the Antialtair Seamount High Seas MPA General management measures taken by the OSPAR Commission that are common to all proposed EBSAs. OSPAR Code of Conduct for Responsible Marine Research in the Deep Seas and High Seas of the OSPAR Maritime Area (Reference number: 2008-1) OSPAR Recommendation 2010/5 on assessments of environmental impact in relation to threatened and/or declining species and habitats OSPAR Recommendation 2010/6 on furthering the protection and restoration of the common skate species complex, the white skate, the angel shark, and the basking shark in the OSPAR Maritime Area OSPAR Recommendation 2010/7 on furthering the protection and restoration of the Orange roughy (Hoplostethus atlanticus) in the OSPAR Maritime Area OSPAR Recommendation 2010/8 on furthering the protection and restoration of Lophelia pertusa reefs in the OSPAR Maritime Area OSPAR Recommendation 2010/9 on furthering the protection and restoration of coral gardens in the OSPAR Maritime Area OSPAR Recommendation 2011/2 on furthering the protection and conservation of the Ivory gull (Pagophila eburnea) OSPAR Recommendation 2011/3 on furthering the protection and conservation of the Little shearwater (Puffinus assimilis baroli) OSPAR Recommendation 2011/4 on furthering the protection and conservation of the Balearic shearwater (Puffinus mauretanicus) OSPAR Recommendation 2011/5 on furthering the protection and conservation of the Black-legged kittiwake (Rissa tridactyla tridactyla) OSPAR Recommendation 2011/6 on furthering the protection and conservation of the Roseate tern (Sterna dougallii) OSPAR Recommendation 2011/7 on furthering the protection and conservation of the Thick-billed murre (Uria lomvia) International Maritime Organization (IMO) and the London Convention No spatially based measures have been introduced by IMO or the London Convention for Areas Beyond National Jurisdiction in the Northeast Atlantic. Many general IMO measures (e.g. ballast water exchange protocols, disposal of waste) will apply to these areas. ICES Advice 2013, Book 1 275 International Seabed Authority (ISA) According to UNCLOS ISA is the competent authority that regulates mining and mineral extraction in “The Area” (http://www.isa.org.jm/en/scientific/exploration). A memorandum of understanding was signed between the OSPAR Convention and the International Seabed Authority in June 2011 to facilitate consultation and sharing of data and information of relevance, with a view to promoting and enhancing a better understanding and coordination of their respective activities. There are three types of deep-sea mineral resources that are of commercial interest: polymetallic nodules, polymetallic sulphides, and cobalt-rich ferromanganese crusts. So far, ISA has only adopted the regulatory framework for the exploration of deep-sea mineral resources, including very strict regulations to ensure full environmental impact assessments are undertaken prior to any activity in “The Area”. The framework for the exploitation of these resources has yet to be developed and adopted. There are several areas in the Northeast Atlantic identified as having deposits of mineral resources of possible commercial interest, but exploratory mining cannot commence without adoption of a management framework. Arctic Council In follow-up to a recommendation to identify areas of heightened ecological and cultural significance, a report is being prepared for the Arctic Council’s Protection of the Arctic Marine Environment Working Group (PAME). If the areas are agreed, it is recommended that Arctic states, where appropriate, should encourage implementation of measures to protect these areas from the impacts of Arctic marine shipping, in coordination with all stakeholders and consistent with international law. National and European Union At the national and EU levels, many relevant measures for the protection of areas have not been systematically reviewed by ICES. Several of these measures implement more general international obligations, for instance under the Convention on Trade in Endangered Species. Sources ICES. 2013a. Vulnerable deep-water habitats in the NEAFC Regulatory Area. In Report of the ICES Advisory Committee, 2013, Section 1.5.5.1. ICES Advice, 2013, Book 1. ICES. 2013b. Report of the Workshop to Review and Advise on EBSA Proposed Areas (WKEBSA), 27–31 May 2013, Copenhagen, Denmark. In draft. 276 ICES Advice 2013, Book 1 1.5.6.7 Special Request, Advice September 2013 ECOREGION General advice SUBJECT OSPAR/NEAFC special request on review and reformulation of four EBSA Proformas Advice summary ICES provided advice to OSPAR and NEAFC in June 2013 (OSPAR/NEAFC special request on review of the results of the Joint OSPAR/NEAFC/CBD Workshop on Ecologically and Biologically Significant Areas (EBSAs) (ICES Advice 2013 section 1.5.5.5). Following discussion with OSPAR and NEAFC, ICES (using experts of the review group) agreed to reformulate and revise four of the EBSAs and provide new updated maps. The material consists of scientifically updated Proformas for the following EBSAs: • • • • Mid-Atlantic Ridge North of the Azores and South of Iceland Charlie-Gibbs Fracture Zone (and the Sub-Polar Front) The Hatton and Rockall Banks and the Hatton-Rockall Basin The Arctic Ice habitat – multiyear ice, seasonal ice – marginal ice zone During the update on the Charlie-Gibbs Fracture Zone (CGFZ) it appeared that the Sub-Polar Front, which was included in the previous version of the Proforma, could not be scientifically supported with the evidence to hand at the time and was therefore excluded from the CGFZ Proforma. A short note explains the reasons and consequences for the exclusion. • Note on the Sub Polar Front. The five documents are scientifically updated technical documents and appended to this advice. ICES Advice 2013, Book 1 277 ANNEX 1 Draft Proforma: The Hatton and Rockall Banks and the Hatton-Rockall Basin Presented by: The Joint OSPAR/NEAFC/CBD Scientific Workshop. Reviewed by an ICES expert group and revised by Francis Neat and J. Murray Roberts. Based on an original proposal submitted by: Dr David Billett and Dr Brian Bett (Deepseas Group, Ocean Biogeochemistry and Ecosystems Department, NOC, UK), Prof. Philip Weaver (Seascape Consultants Ltd and National Oceanography Centre, UK), Prof Callum Roberts and Ms Rachel Brown (Environment Department, University of York), Dr Murray Roberts and Dr Lea-Anne Henry, (Centre for Marine Biodiversity and Biotechnology, School of Life Sciences, Heriot-Watt University), Dr Kerry Howell and Dr Jason Hall-Spencer (Marine Biology and Ecology Research Centre, Marine Institute, University of Plymouth); Dr Andrew Davies (School of Ocean Sciences, Bangor University); Dr Bhavani Narayanaswamy (Scottish Association for Marine Science, Oban), Prof. Monty Priede (OceanLab, University of Aberdeen); Dr David Bailey (Division of Environmental and Evolutionary Biology, University of Glasgow); Prof. Alex Rogers (University of Oxford) and Mr Ben Lascelles (Global Seabird Programme, Bird Life International) Abstract The Hatton and Rockall Banks, associated slopes and connecting basin, represent offshore bathyal habitats between 200 to 1500 m that constitute a unique and prominent feature of the NE Atlantic. The area has high habitat heterogeneity and supports a wide range of benthic and pelagic species and ecosystems. There is significant fishing activity in the area, including bottom trawling, long-lining, and midwater fisheries. Introduction The Hatton and Rockall Banks are large isolated geomorphological features in the NE Atlantic. Formed from continental crust, they span depths from c. 200 to 2000m. The banks are linked by the Hatton-Rockall Basin at a depth of approximately 1300 m which has particular geomorphological features and habitats. The gently sloping banks and the basin provide a contrasting geological setting to the tectonically active Mid-Atlantic Ridge to the west and the generally steeper slopes of the European continental margin to East. The banks encompass a large depth range with strong environmental gradients (e.g. temperature, pressure, and food availability) that give rise to a high diversity of species and habitats (Billett, 1991; Bett, 2001; Howell et al., 2002; Davies et al. 2006; Roberts et al. 2008; Howell et al., 2009; Howell et al. 2010). Environmental heterogeneity is positively correlated with biological diversity at a variety of scales (Menot et al. 2010) as indicated by significantly elevated levels of species change across space (in areas such as Hatton Bank (Roberts et al. 2008). Changes in pressure and temperature have significant effects on the biochemistry of species, influencing cell membrane structure and enzyme characteristics (Gage and Tyler, 1991). In general, each species is adapted to a particular range of environmental conditions. Each may occur over a depth range of about 500 m, but the depths where any particular species is abundant, and therefore able to form viable populations, is generally limited to a much more restricted depth range of 100 to 200 m (Billett, 1991; Howell et al., 2002). There is evidence that such depth-related effects promote speciation (Howell et al., 2004). In addition, the progressive decrease in organic matter availability with increasing depth (with some patchiness depending on geomorphology) leads to a reduction of carnivores and an increase in detritus feeders (Billett, 1991). Taken together such environmental changes lead to a continuous sequential change in species composition with depth, and biological community characteristics that are radically different to those known in shelf seas. The area is influenced by a number of different water masses and there is considerable interaction between the topography and physical oceanographic processes, in some areas focusing internal wave and tidal energy (Ellett et al. 1986) which results in strong currents and greater mixing. This may give rise to highly localized and specialised biological communities such as sponge aggregations and coral gardens. The mixing of Arctic and Atlantic water in the North of the area means that species from both ecosystems are represented causing enhanced species diversity. The Rockall Bank supports shallow demersal fisheries targeting haddock, gurnard and monkfish (Neat & Campbell 2010). The slopes and the Hatton Bank are target areas for deep-water bottom fisheries for Ling (Molva molva), Blue Ling (Molva dypterygia), Tusk (Brosme brosme), Roundnose Grenadier (Coryphaenoides rupestris) and Black Scabbardfish (Aphanopus carbo). In the past deepwater sharks were also caught in the area, but this is now prohibited. A wide variety of other fish species are also taken as by-catch (Gordon et al., 2003; Large et al., 2003; ICES 2010). Some of the deepwater target species have characteristic low productivity and extended generation times. Deep-water fisheries have 278 ICES Advice 2013, Book 1 significant effects not only on target fish species, but also on the benthic fauna (Le Guilloux et al., 2009; Clark et al. 2010). Major wide-ranging Northeast Atlantic epipelagic fish stocks, e.g. mackerel and blue whiting, use the Hatton-Rockall area for parts of their life cycle and are targeted by international fisheries. The slopes of the banks and channels between the banks have a diverse bathy- and mesopelagic fish community sustained by the zooplankton production in the epipelagic zone. The pelagic communities are similar to, and probably extensions of, those in adjacent oceanic waters along the European continental margin. Some invertebrate species, such as cold-water corals and sponges, provide important structural habitat heterogeneity. These habitats are highly susceptible to physical damage and may take hundreds, if not thousands, of years to reform (Hall-Spencer et al. 2002; Roberts et al. 2009; Söffker et al., 2011). Current fisheries control measures on Hatton and Rockall Bank have focused mainly on the protection of corals (HallSpencer et al., 2009) and sponges (ICES 2013). There is no evidence currently that the seabed at depth greater than 1500 m in the area is significantly different from comparable depths in the rest of the NE Atlantic. The majority of the features considered here occur at depths shallower than 1500 m and this therefore forms an appropriate delimitation of the EBSA. Location The EBSA would comprise the seabed and pelagic zones shallower than 1500 m above the Rockall and Hatton Banks and the adjoining Hatton-Rockall Basin. This extends into adjacent EEZs, but the current proposal refers to the ABNJ only. The area beyond national jurisdiction lies wholly within regions under consideration by the Commission on the Limits of the Continental Shelf. Feature description Benthic and pelagic communities to depths of 1500 m in and around the Hatton and Rockall Banks and Basin. Seabed communities include cold-water coral formations and sponge aggregations. Geomorphologically complex seabed types include rocky reefs, carbonate mounds, polygonal fault systems and sedimentary slopes, slides and fans. Pelagic communities include those inhabiting bathy-, meso- and epi-pelagic zones, including zooplankton, fish, cetaceans, turtles and seabirds. 1. Benthic and benthopelagic communities Cold-water coral Observations in the early 1970s found cold-water coral communities on the Rockall Bank down to a depth of 1,000 m (Wilson, 1979a). Thickets of Lophelia pertusa occurred principally at depths between 150-400m 21. Large coral growth features have recently (2011) been discoveredto be still present on the northern Rockall Bank (Huvenne et al., 2011, Roberts et al. 2013). Bottom-contact fishing can result in significant adverse impacts to these habitats. Frederiksen et al. (1992) reported a high diversity of corals on the northern Hatton Bank, including Paragorgia, Paramuricea, Isididae and Antipatharia as well as the scleractinians L. pertusa and M. oculata. Since these observations further records of coral frameworks have been noted throughout the Rockall, Hatton area, including the Logachev Mounds and the Western Rockall Bank Mounds (Kenyon et al., 2003; Roberts et al., 2003; Narayanaswamy et al., 2006; Howell et al., 2007; Durán Muñoz et al. 2009). Recent surveys identified many areas that contained the cold-water coral L. pertusa throughout the Rockall and Hatton Banks (Narayanaswamy et al., 2006; Howell et al., 2007; Roberts et al. 2008; Durán Muñoz et al. 2009). Several areas on the Hatton Bank contained pinnacles and mounds with extensive biogenic structures including areas of coral rubble around the flanks of the coral mounds. Coral frameworks are known from the Hatton Bank (Durán Muñoz et al. 2009), and are predicted to occur over focused regions of the Hatton Bank (Howell et al., 2011). Geophysical evidence suggests that these have formed by successive coral growth and sedimentation episodes, as in other regions (Roberts et al., 2006), forming coral carbonate mounds (Roberts et al. 2008). Single and clustered coral carbonate mounds have also been discovered on the southeast of Rockall Bank. These structures are comprised mostly of L. pertusa and can reach heights of 380 m in water depths of between 600-1000 m (Kenyon et al., 2003; Mienis et al., 2006; Mienis et al., 2007). 21 http://www.lophelia.org/lophelia/case_4.htm ICES Advice 2013, Book 1 279 Cold-water coral frameworks have been reported to support over 1,300 species in the Northeast Atlantic, some of which have yet to be described (Roberts et al., 2006). New species and associations have been reported recently (e.g. Myers & Hall-Spencer 2007; Le Guilloux et al., 2010; Söffker et al. 2011). The corals may provide an important habitat for certain fish species (Fosså et al., 2002; Söffker et al. 2011; Henry et al 2013), including commercial species Sebastes sp., Molva molva, Brosme brosme, Anarhichas lupus and Pollachius virens (Mortensen et al., 1995; Freiwald, 2002; Hall-Spencer et al., 2002). Pregnant Sebastes viviparus may use the reef as a refuge or as a nursery ground to raise their offspring (Fosså et al., 2002) as recently observed on the northern Rockall Bank (Huvenne et al., 2011, Roberts et al. 2013). As well as living reefs, dead coral framework and coral rubble provide a structural habitat. Jensen and Frederiksen (1992) collected Lophelia and found 256 species; a further 42 species were identified amongst coral rubble. There has been only limited research into linkages between coral and other deep-water ecosystems. Compared to the south-eastern US and Gulf of Mexico, molecular research has shown that northeastern Atlantic populations of L. pertusa are moderately differentiated (Morrison et al. 2011) and form distinct subpopulations, but also that Rockall Bank corals show some genetic similarity to those occurring on the New England Seamounts indicating some degree of connectivity (Morrison et al. 2011). Lophelia pertusa exhibits high levels of inbreeding through asexual reproduction at several sites in the NE Atlantic, suggesting a high incidence of self-recruitment in local populations (Le Goff-Vitry and Rogers, 2005). Further molecular studies are required in local areas to gauge the importance of the Rockall and Hatton Banks in the life history of regional coral populations. In summary the cold-water corals fit the following EBSA criteria: Uniqueness or rarity • Large areas of cold-water corals and sponges have been reported in the area. Some of these have been destroyed by demersal trawling, but in certain areas, e.g. SW Rockall Bank, extensive patches of coral framework still exist. Special importance for life-history stages • Cold-water corals and areas of natural coral rubble provide highly diverse habitats Importance for threatened, endangered or declining species/habitats • • The cold-water corals and natural rubble contain very large numbers of invertebrate species including giant protozoans on nearby sedimentary habitats (xenophyophores), vase shaped white sponges, actiniarians, antipatharian corals, hydroids, bryozoans, asteroids, ophiuroids, echinoids, holothurians and crustaceans. The distribution of cold-water coral has been severely reduced in the area over the last 30 years Vulnerability, fragility, sensitivity, or slow recovery • • There is a high diversity of corals, including bamboo coral (Isididae), black coral (Antipatharia) as well as the reef forming stony corals (Scleractinia), though some of these may now be reduced in distribution occurring in patches. Cold-water coral habitats are easily impacted and recover very slowly, if at all. Biological diversity • Cold-water corals provide diverse habitats for other invertebrates and fish. Sediment communities The Hatton and Rockall Banks support many different habitats each with their own depth-related species assemblages (Narayanaswamy et al., 2006; Howell et al., 2007; Roberts et al. 2008; Howell et al., 2009). Local seabed morphology in this region is ultimately controlled by hydrography and oceanography (Due et al. 2006; Sayago-Gil et al. 2010), which creates heterogeneity in sediment types including mud, exposed bedrock, fine sediments, living coral framework and coral debris that – this habitat heterogeneity has a major influence on species diversity and turnover (Roberts et al. 2008). A great variety of large invertebrate fauna (megafauna) occur in this region including giant protozoans (xenophyophores), vase shaped white sponges, actiniarians, antipatharian corals, hydroids, bryozoans, asteroids, ophiuroids, echinoids, holothurians and crustaceans (Narayanaswamy et al., 2006; Howell et al., 2007; Roberts et al. 2008). Large mega-infauna such as echiuran worms are evident from observations of their feeding traces. Little is known, however, of the smaller fauna living within the sediment. The Hatton-Rockall Basin is known to host a particular geomorphology known as a polygonal fault system (Berndt et al 2012). The faults in the Hatton-Rockall Basin have surface expression, i.e. a network of interlinked channels across the level seafloor. These fault structures were confirmed again in 2011 (Huvenne et al., 2011). The flanks of the gullies appear to support extensive, dense aggregations of mixed species sponge communities. 280 ICES Advice 2013, Book 1 A key interest / conservation concern in such a geological setting would be the occurrence of cold-seep communities. Large carbonate blocks were encountered that were likely formed as a result of seafloor fluid escape. In 2012 the first evidence of an active cold-seep ecosystem in the area was suggested by the collection of chemosynthetic bivalves and polychaete worms (ICES 2013) on the eastern margin of Hatton-Rockall Basin at a depth of 1200 m. The species are new to science and suggest there is a lot still to learn of the seafloor and ecology of the Hatton and Rockall Banks. The megafauna on the Hatton and Rockall Banks are largely species known from the wider NE Atlantic continental margin (Gage et al. 1983; Gage et al., 1985; Mauchline et al., 1986; Harvey et al., 1988; Rice et al., 1991). These studies focused on sedimented areas within the EEZs of the UK and Ireland and provide a lot of information on the life history characteristics of the species including information on growth and reproduction. Apart from some species that produce small eggs (indicative of planktotrophic development) in a seasonal cycle, most species conform to the life history characteristics typical of the deep sea of larger egg size, lower fecundity and greater generation times (Gage and Tyler, 1991). This is an adaptation to the low food input to the deep sea, which leads to the rapid decrease in biomass with increasing depth (Lampitt et al., 1986; Wei et al., 2010). Fauna adapt to lower food availability in the deep sea by a number of trade-offs, one of which is a reduction in reproductive effort and longer generation times. The majority of species, therefore, are highly susceptible to repeated physical disturbance. In summary the sediment communities fit the following EBSA criteria: Uniqueness or rarity • • The area has considerable environmental heterogeneity, and therefore biological diversity, as a result of its large depth range and strong environmental gradients. Habitat-forming sessile benthic communities, such as those of giant protozoans and sponges, are common. The area of polygonal faults may be a unique seabed feature and the presence of newly described chemosynthetic bivalves and polychaete worms suggests the area may have unique communities. Importance for threatened, endangered or declining species/habitats • The area comprises a patchwork of habitats with species changing consistently with both habitat type and increasing to depths of 1500 m. Some habitats are threatened by direct impacts (e.g. trawling). Vulnerability, fragility, sensitivity, or slow recovery • Many of the species have reproductive cycles with long generation times leading to very slow and episodic recoveries following human impact. Most deep-sea species are particularly susceptible to degradation and depletion by human activity. Biological productivity • There are localised areas of concentrated production depending on geomorphology and hydrography, but little evidence that the area has an enhanced productivity relative to other areas. Biological diversity • Benthic and pelagic communities occupy all depths in and around the Hatton and Rockall Banks and Basin. Seabed communities include cold-water corals and sponge aggregations. Seabed geomorphology is diverse with examples of rocky reefs, carbonate mounds, polygonal fault systems, and steep and gentle sedimentary slopes. This high habitat heterogeneity supports a high number of species and diverse communities. Demersal fish The deep-water fish of the NE Atlantic continental margin are generally well-known following comprehensive and extensive surveys of the region (e.g. Gordon & Duncan, 1985; Merrett et al., 1991; Mauchline et al. 1986 and Rice et al. 1991). Species of commercial importance are reviewed by Gordon et al. (2003) and Large et al. (2003) and for fish associated with cold-water corals by Söffker et al. (2011). Fish species diversity increases to depths of approx. 1500 m on the continental slopes and declines thereafter (Campbell et al 2011). The shallow water fish assemblage on Rockall can be described as an impoverished sub-set of that found in adjacent continental shelf areas, but one that has a significantly different community composition (Neat & Campbell 2010). Recent surveys have found that the western slope of the Rockall Bank has a slightly different fish assemblage than the adjacent European slope with several species of a more southern affinity present (F. Neat unpublished data). Blue ling is known to spawn in a few locations on Rockall bank and at Hatton bank (Large et al 2008). ICES Advice 2013, Book 1 281 The detailed sampling in the Porcupine Seabight in the 1970s and 1980s took place before the start of deep-water commercial fishing. More recent sampling of the same area in the 1990s and 2000s can be used to compare fish communities before and after bottom trawling (Bailey et al. 2009). These data show that over 70 fish species have been impacted by the fishing activity, of which only 4-5 are target commercial species. The area impacted is up to 2.5 times larger than the area fished because the home range of many the fish extends into considerably deeper waters. In the past decade, however, there is evidence that this initial steep decline in abundance has been halted, at least in one of the major groups of fishes, the grenadiers (Neat & Burns 2010). At the northern limits of the area where Arctic water masses mix with Atlantic water cold-water species such as Greenland Halibut and Roughhead Grenadier are present adding to the diversity of species in the area. In summary the demersal fish fit the following EBSA criteria: Vulnerability, fragility, sensitivity, or slow recovery • Many of the deep demersal fish have very slow recovery times as a result of their slow reproductive rate compared to pelagic fish. 2. Pelagic communities (plankton, nekton, birds) Fish: Mackerel, blue whiting and other wide-ranging pelagic fish such as epipelagic sharks and tuna-like species use the area during parts of their life-cycle, for feeding or as migration corridors. For blue whiting the slope area is used as a spawning area. Mackerel eggs and larvae from spawning areas further south drift through the area. Cetaceans: Phocoena phocoena have been observed over the shallower parts of Rockall Bank, but It is unlikely that the area is of particular importance for the species. Limited numbers of the endangered Blue whale (Balaenoptera musculus) and the critically endangered northern right whale (Eubalaena glacialis) have also been observed in this area (Cronin and Mackey, 2002; Hammond et al., 2006). Seabirds: Analyses of satellite tracking data hosted at www.seabirdtracking.org (Table 1) found the area to be used by multiple species through the year. The site is used by Manx Shearwaters (Puffinus puffinus) during the breeding season (Apr-Sept) from Iceland and UK colonies. From September until November tracked individuals of Cory’s Shearwater (Calonectris diomedea) from 3 colonies, Sooty Shearwater (Puffinus griseus), Fea’s Petrel (Pterodroma feae) and Zino’s Petrel (Pterodroma madeira) used the area. Studies of tracked Atlantic Puffin (Fratercula arctica) from Skomer and Isle of May colonies found the site to be important during the overwintering phase (Aug-Apr) (Harris et al. 2010, Guilford et al. 2011). In addition to tracking data, at-sea survey data confirms many more species within the area (e.g. Cronin and Mackey, 2002). Feature condition, and future outlook Demersal fish have been targets of extensive fisheries for decades, expanding primarily in the latter half of the 1980s. Although satisfactory stock assessments were seldom achieved, the probable declines in abundance and vulnerability of many of the target species have been reflected in advice from ICES for many years (ICES 1996 onwards, Large et al., 2003). A range of management actions by NEAFC and relevant coastal states have been implemented to reduce fishing effort and facilitate recovery of target species and some associated by-catch species. A similar range of measures applies to species inhabiting the shallowest areas, e.g. haddock. Epipelagic species such as mackerel and blue whiting, and large pelagic sharks and tuna-like species occurring in the area straddle between ABNJ and several EEZs and the fisheries are managed by relevant coastal states, NEAFC and ICCAT. Cetaceans are managed by the IWC. The management is based on recurrent stock assessments by ICES and other advisory bodies. Records of the physical impact of deep-water trawling west of Scotland extend back to the late 1980s (Roberts et al., 2000; Gage et al., 2005) and studies using VMS data show that fishing activity potentially affects much of the HattonRockall area (Hall-Spencer et al. 2009; Benn et al. 2010). Damage may occur to structural species such as corals and sponges, which may take hundreds to thousands of years to recover (Hall-Spencer et al., 2002; Davies et al. 2007; Roberts et al., 2009; Hogg et al. 2010). A recent survey (2011) has documented extensive destruction of coral framework on the northern Rockall Bank (Huvenne et al. 2011) in waters adjacent to the area currently being described. This expedition also encountered evidence of trawling impact on the megafauna of open sedimented areas, with photographic surveys in the area of the 'Haddock Box' (Rockall Bank) showing frequent occurrence of physically damaged holothurians - thought to be net escapees or discarded by282 ICES Advice 2013, Book 1 catch. Cold seep communities are vulnerable to trawling impacts; they are typically highly localised and are of a relatively small scale such that they could be eliminated by a single trawl. Cold seeps are OSPAR priority habitats for which there are considerable concerns regarding the effects of bottom trawling (van Dover et al. 2011a, b). Some of the benthic communities of the Hatton and Rockall Banks have already been significantly affected by deep-water fishing (ICES WGDEC, 2007). The effects on deep-water fish may extend to waters deeper than those utilised by trawl fisheries (Bailey et al., 2009). Broad-scale multibeam surveys have revealed a diverse range of geomophological features and sediment types on Hatton Bank (Jacobs and Howell, 2007; Stewart and Davies, 2007; MacLachlan et al., 2008; Sayago-Gil et al., 2010). These physical environment maps, coupled with targeted biological surveys have resulted in the production of biological habitat maps for the region (Howell et al., 2011) which highlight the range and diversity of noncoral seabed features present in the area. Assessment against CBD EBSA Criteria Table 1. Relation of each of the CBD criteria to the proposed area relating to the best available science. Note that a candidate EBSA may qualify on the basis of one or more of the criteria, the boundaries of the EBSA need not be defined with exact precision. CBD EBSA Criterion Description The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Explanation for ranking Uniqueness or rarity Ranking of criterion relevance (please mark one column with an X) Don’t Low Some Know High X • The area has considerable environmental heterogeneity, and therefore biological diversity, as a result of its large depth range and strong environmental gradients. Habitat-forming sessile benthic communities, such as those of giant protozoans and sponges, are common. • Large areas of cold-water corals and sponges have been reported in the area. Some of these have been destroyed by demersal trawling, but some areas of large coral frameworks still exist. • An area of polygonal faults may be a unique seabed feature and the recent discovery of cold-seep species that are new to science suggests the area is very likely to be unique. Areas that are required for a population to survive and Special X thrive importance for life-history stages of species Explanation for ranking • • Cold-water corals and areas of natural coral rubble provide highly diverse habitats Parts of the Hatton-Rockall area are important as spawning areas for blue whiting, and the area is used as a corridor for a range of migrating species including turtles. Area containing habitat for the survival and recovery of Importance for X endangered, threatened, declining species or area with threatened, significant assemblages of such species endangered or declining species and/or habitats Explanation for ranking • • • The cold-water corals and natural rubble contain very large numbers of invertebrate species including giant protozoans (xenophyophores), vase shaped white sponges, actiniarians, antipatharian corals, hydroids, bryozoans, asteroids, ophiuroids, echinoids, holothurians and crustaceans. The distribution of cold-water coral has been severely reduced in the area over the last 30 years The area comprises a patchwork of habitats with species changing consistently with both habitat type and increasing depth. Some habitats are threatened by direct impacts (e.g. trawling), others may suffer indirectly e.g. through the creation of sediment plumes by impacts of fishing gear in sensitive areas. ICES Advice 2013, Book 1 283 Areas that contain a relatively high proportion of sensitive Vulnerability, habitats, biotopes or species that are functionally fragile fragility, (highly susceptible to degradation or depletion by human sensitivity, or activity or by natural events) or with slow recovery slow recovery Explanation for ranking • • • • X There is a high diversity of corals, including bamboo coral (Isididae), black coral (Antipatharia) as well as the reef forming stony corals (Scleractinia), though some of these may now be reduced in distribution occurring in patches. Cold-water coral habitats are easily impacted and recover very slowly, if at all Many of the species have reproductive cycles with long generation times leading to very slow and episodic recoveries following human impact. Most deep-sea species are particularly susceptible to degradation and depletion by human activity and natural events. Many of the demersal fish have very slow recovery times as a result of their slow reproductive rate compared to pelagic fish. Stocks have already been diminished in some areas. Area containing species, populations or communities with Biological comparatively higher natural biological productivity productivity Explanation for ranking X • While pelagic organisms may be more concentrated over the banks in the area, there is little evidence to suggest overall enhanced productivity of the area. Area contains comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or has diversity higher genetic diversity Explanation for ranking • • • Benthic and pelagic communities occupy all depths in and around the Hatton and Rockall Banks and Basin. Seabed communities include cold-water corals, rocky reefs, carbonate mounds, polygonal fault systems, sponge aggregations, steep and gentle sedimented slopes. The Hatton and Rockall Banks and the Hatton-Rockall Basin have a high habitat heterogeneity that supports diverse seabed communities. Cold-water corals provide diverse habitats for other invertebrates and fish. References Bailey, D.M., Collins, M.A., Gordon, J.D.M., Zuur, A.F. & Priede, I.G. (2009) Long-term changes in deep-water fish populations in the northeast Atlantic: a deeper reaching effect of fisheries? Proc. Roy. Soc. Lond. B. 276, 19651969 Benn, A.R., Weaver, P.P.E, Billett, D.S.M., van den Hove, S., Murdock, A.P., Doneghan, G.B. & Le Bas, T. (2010). Human Activities on the Deep Seafloor in the North East Atlantic: An Assessment of Spatial Extent. PLoS One 5(9): e12730. doi:10.1371/journal.pone.0012730. Berndt, C, Jacobs, C. L., Evans, A. J., Gay, A., Elliot, G., Long, D. and Hitchen, K. (2012) Kilometre-scale polygonal seabed depressions in the Hatton Basin, NE Atlantic Ocean: Constraints on the origin of polygonal faulting Marine Geology, 332/334 . pp. 126-133. DOI 10.1016/j.margeo.2012.09.013. Bett, B.J. (2001) UK Atlantic Margin Environmental Survey: introduction and overview of bathyal benthic ecology. Cont. Shelf Res. 21, 917-956. Billett, D.S.M. (1991) Deep-sea holothurians. Oceanogr. mar. Biol. Ann. Rev. 29, 259-317. Campbell, N. et al. 2010. Taxonomic indicators of deep water demersal fish community diversity on the Northeast Atlantic continental slope. ICES J. Mar. Sci. 68, 365-378. Clark M.R., Rowden A.A., Schlacher T., Williams A., Consalvey M., Stocks K.I., Rogers A.D., O'Hara T.D., White M., Shank T.M. & Hall-Spencer J.M. (2010) The ecology of seamounts: structure, function and human impacts. Annu. Rev. Mar. Sci. 2, 253-278. Costello, M.J., McCrea, M., Freiwald, A., Lundälv, T., Jonsson, L., Bett, B.J., Van Weering, T.C.E., De Haas, H., Roberts, J.M. & Allen, D. (2005) Role of cold-water Lophelia pertusa reefs as fish habitat in the NE Atlantic. In: Freiwald, A., Roberts, J.M. (Eds.), Cold-water Corals and Ecosystems. Springer-Verlag, Berlin Heidelberg, pp. 771-805. Cronin, M., Mackey, M. (2002) Cetaceans and Seabirds of the Hatton-Rockall Region, Cruise Report of the Geological Survey of Ireland May 2002. Davies, A.J., Narayanaswamy, B.E., Hughes, D.J. & Roberts, J.M. (2006) An introduction of the benthic ecology of the Rockall-Hatton Area (SEA 7). Scottish Association for Marine Science, Oban, p. 94. http://www.offshoresea.org.uk/ Davies, A.J., Wisshak, M., Orr, J.C. & Roberts, J.M. (2008) Predicting suitable habitat for the cold-water coral Lophelia pertusa (Scleractinia). Deep-Sea Res. I 55, 1048-1062. Davies. A., Roberts. J.M. & Hall-Spencer, J.M. (2007) Preserving deep-sea natural heritage: emerging issues in offshore conservation and management. Biol. Cons. 138, 299-312. 284 ICES Advice 2013, Book 1 Due, L., van Aken. H.M., Boldreel. L.O. & Kuijpers, A. (2006) Seismic and oceanographic evidence of present-day bottom-water dynamics in the Lousy Bank–Hatton Bank area, NE Atlantic. Deep-Sea Research I53:1729-1741 Durán Muñoz, P., Sayago-Gil, M., Cristobo, J., Parra, S., Serrano, A., Díaz del Rio, V., Patrocinio, T., Sacau, M., Murillo, F.J., Palomino, D. & Fernández-Salas, L.M. (2009) Seabed mapping for selecting cold-water coral protection areas on Hatton Bank, Northeast Atlantic. ICES J. Mar. Sci. 66, 2013-2025 Ellett, D.J., Edwards, A. & Bowers, R. (1986) The hydrography of the Rockall Channel – an overview. Conference Proceedings Symposium on the Oceanography of the Rockall Channel, Edinburgh (UK), 27-29 Mar 1985. Fosså, J.H., Mortensen, P.B. & Furevik, D.M. (2002) The deep-water coral Lophelia pertusa in Norwegian waters: distribution and fishery impacts. Hydrobiologia 471, 1-12. Frederiksen, R., Jensen, A. & Westerberg, H. (1992) The distribution of the scleractinian coral Lophelia pertusa around the Faeroe Islands and the relation to internal tidal mixing. Sarsia 77, 157-171. Freiwald, A. (2002) Reef-forming cold-water corals. In: Wefer, G., Billett, D., Hebbeln, D., Jorgensen, B.B., Schluter, M., Van Weering, T. (Eds.), Ocean Margin Systems. Springer-Verlag Berlin Heidelberg, Berlin, pp. 365-385. Gage, J.D. & Tyler, P.A. (1991) Deep-Sea Biology. A Natural History of Organisms at the Deep-Sea Floor. Cambridge University Press. 504pp. Gage, J.D., Billett, D.S.M., Jensen, M. & Tyler, P.A. (1985) Echinoderms of the Rockall Trough and adjacent areas. 2. Echinoidea and Holothurioidea. Bull. Br. Mus. Nat. Hist. (Zool.), 48, 173-213. Gage, J.D., Pearson, M., Clark, A.M., Paterson, G.L.J. & Tyler, P.A. (1983) Echinoderms of the Rockall Trough and adjacent areas. 1. Crinoidea, Asteroidea and Ophiuroidea. Bull. Br. Mus. nat. Hist. (Zool.) 45, 263-308. Gage, J.D., Roberts, J.M., Hartley, J.P. & Humphery, J.D. (2005) Potential impacts of deepsea trawling on the benthic ecosystem along the Northern European Continental Margin: A review. In: Barnes, P.W., Thomas, J.P. (Eds.), Benthic Habitats and the Effects of Fishing. American Fisheries Society, Bethesda, Maryland, pp. 503-517. Gordon, J. D. M., O. A. Bergstad, I. Figueiredo, and G. Menezes. 2003. Deep-water Fisheries of the Northeast Atlantic: I. Description and Current Trends. J. Northw. Atl. Fish. Sci. 31: 137-150. Gordon, J.D.M. & Duncan, J.A.R. (1985) The ecology of the deep-sea benthic and benthopelagic fish on the slopes of the Rockall Trough, northeastern Atlantic. Prog. Oceanogr. 15, 37-69. Guilford, T., Freeman, R., Boyle, D., Dean, B., Kirk, H., Phillips, R.A., Perrins, C. (2011) A dispersive migration in the Atlantic Puffin and its Implications for Migratory Navigation, Plos One, 6(7), e21336. Hall-Spencer J.M., Allain V. & Fossa J.H. (2002) Trawling damage to Northeast Atlantic ancient coral reefs. Proc. Roy. Soc. Lond. B. 269, 507-511. Hall-Spencer, J.M., Tasker, M., Söffker, M., Christiansen, S., Rogers, S., Campbell, M. & Hoydal, K. (2009) The design of Marine Protected Areas on High Seas and territorial waters of Rockall. Mar. Ecol. Prog. Ser. 397, 305-308. Harris, M.P., Daunt, F., Newell, M., Phillips, R.A., Wanless, S. (2010) Wintering areas of Atlantic Puffins Fratercula arctica from a North Sea colony as revealed by geolocation technology, Mar Biol, 157, 827-836 Harvey, R., Gage, J.D., Billett, D.S.M., Clark, A.M. and Paterson, G.L.J. (1988) Echinoderms of the Rockall Trough and adjacent areas. 3. Additional records. Bull. Br. Mus. (Nat. Hist.) (Zool.), 54 (4), 153-198. Henry, L-A. & Roberts, J.M. (2007) Biodiversity and ecological composition of macrobenthos on cold-water coral mounds and adjacent off-mound habitat in the bathyal Porcupine Seabight, NE Atlantic. Deep-Sea Res I 54, 654-672 Hogg, M.M., Tendal, O.S., Conway, K.W., Pomponi, S.A., van Soest, R.W.M., Gutt, J., Krautter, M. & Roberts, J.M. (2010) Deep-sea Sponge Grounds: Reservoirs of Biodiversity. UNEP-WCMC Biodiversity Series No. 32. UNEP-WCMC, Cambridge, UK Howell, K., Billett, D.S.M. & Tyler, P.A. (2002). Depth-related distribution and abundance of seastars (Echinodermata:Asteroidea) in the Porcupine Seabight and Porcupine Abyssal Plain, N.E. Atlantic. Deep-Sea Res. I 49, 1901-1920. Howell, K.L. (2010) A benthic classification system to aid in the implementation of marine protected area networks in the deep / high seas of the NE Atlantic. Biol. Cons. 143, 1041–1056. Howell, K.L., Davies J.S., Jacobs, C., and Narayanaswamy B.E. (2007). Broadscale Survey of the Habitats of Rockall Bank, and mapping of Annex I ‘Reef’ Habitat. Joint Nature Conservation Committee Report. No. 422, 165p. Howell, K.L., Davies J.S. & Narayanaswamy, B.E.(2010). Identifying deep-sea megafaunal epibenthic assemblages for use in habitat mapping and marine protected area network design. J. Mar. Biol. Ass. UK 90, 33-68 . Howell, K.L., Davies, J.S., Hughes, D.J. & Narayanaswamy, B.E. (2007) Strategic Environmental Assessment / Special Area for Conservation Photographic Analysis Report. Department of Trade and Industry, Strategic Environmental Assessment Report, UK, p. 163. Unpublished report. Howell, K.L., Holt, R., Pulido Endrino, I. & Stewart, H. (2011) When the species is also a habitat: comparing the predictively modelled distributions of Lophelia pertusa and the reef habitat it forms. Biol. Cons. Howell, K.L., Mowles S. & Foggo, A. (2010) Mounting evidence: near-slope seamounts are faunally indistinct from an adjacent bank. Mar. Ecol. 31, 52-62. Howell, K.L., Rogers, A., Tyler, P.A. & Billett, D.S.M. (2004). Reproductive isolation among morphotypes of the cosmopolitan species Zoroaster fulgens (Asteroidea:Echinodermata). Mar. Biol. 144, 977-984. Husebø, A., Nottestad, L., Fosså, J.H., Furevik, D.M. & Jorgensen, S.B. (2002) Distribution and abundance of fish in deep-sea coral habitats. Hydrobiologia 471, 91-99. ICES Advice 2013, Book 1 285 Huvenne, V.A.I. et al. (2011) RRS James Cook Cruise 60, 09 May-12 Jun 2011. Benthic habitats and the impact of human activities in Rockall Trough, on Rockall Bank and in Hatton Basin. (National Oceanography Centre Cruise Report, No. 04) Southampton, UK: National Oceanography Centre, Southampton, 133pp. ICES (2007) Report of the Working Group on Deep-Water Ecology (WGDEC), 26–28 February 2007, ICES Cm 2007/aCE:01 Ref lRC. International Council for the Exploration of the Sea, Copenhagen, Denmark, 57pp. ICES 2010. Report of the Working group on the Biology and Assessemnt of Deepwater Fisheries Resources. www.ices.dk Jacobs, C.L., Howell, K.L. (2007) Habitat investigations within the SEA4 and SEA7 area of the UK continental shelf. MV Franklin Cruise 0206, 03-23 Aug 2006. Research and Consultancy Report No. 24. National Oceanography Centre, Southampton. UK. 95pp. Jensen, A. & Frederiksen, R. (1992) The fauna associated with the bank-forming deep-water coral Lophelia pertusa (Scleractinaria) on the Faroe Shelf. Sarsia 77, 53-69. Kenyon, N.H., Akhmetzhanov, A.M., Wheeler, A.J., van Weering, T.C.E., de Haas, H. & Ivanov, M.K. (2003) Giant carbonate mud mounds in the southern Rockall Trough. Marine Geology 195, 5-30. Lampitt, R.S., Billett, D.S.M. & Rice, A.L. (1986) The biomass of the invertebrate megabenthos from 500 to 4100m in the North East Atlantic. Mar. Biol. 93, 69-81. Large, P. A., C. Hammer, O. A. Bergstad, J. D. M. Gordon, and P. Lorance. 2003. Deep-water Fisheries of the Northeast Atlantic: II. Assessment and Management Approaches. J. Northw. Atl. Fish. Sci. 31: 151-163. Le Goff-Vitry & M.C. & Rogers, A.D. (2005) Molecular ecology of Lophelia pertusa in the NE Atlantic. In: Freiwald, A., Roberts, J.M. (Eds.). Cold-water Corals and Ecosystems. Springer-Verlag, Berlin Heidelberg, pp. 653-662. Le Guilloux, E., Hall-Spencer, J.M., Söffker, M.K. & Olu-Le Roy, K. (2010) Association between the squat lobster Gastroptychus formosus (Filhol, 1884) and cold-water corals in the North Atlantic. J. Mar Biol. Ass. UK 90, 1363-1369. MacLachlan SE, Elliot GM, Parson LM (2008) Investigations of the bottom current sculpted margin of Hatton bank, NE Atlantic. Mar. Geol. 253:170–184 Mauchline, J., Ellett, D.J., Gage, J.D., Gordon, J.D.M. & Jones, E.J.W. (1986) A bibliography of the Rockall Trough. Conference Proceedings Symposium on the Oceanography of the Rockall Channel, Edinburgh (UK), 27-29 Mar 1985. Menot, L., Sibuet, M., Carney, R.S., Levin, L.A., Rowe, G.T., Billett, D.S.M., Poore, G., Kitazato, H., Vanreusel, A., Galéron, J., Lavrado, H.P., Sellanes, J., Ingole, B. & Krylova, E. (2010) New Perceptions of Continental Margin Biodiversity. In: McIntyre, A., (Ed). Chapter 5. Life in the World’s Oceans: Diversity, Distribution and Abundance. Wiley-Blackwell. 79-101. Merrett, N.R., Gordon, J.D.M., Stehmann, M. & Haedrich, R.L. (1991) Deep demersal fish assemblage structure in the Porcupine Seabight (eastern North Atlantic): slope sampling by three different trawls compared. J. Mar. Biol, Ass. UK 71, 329-358. Mienis, F., de Stigter, H.C., de Haas, H. & van Weering, T.C.E. (2009) Near-bed particle deposition and resuspension in a cold-water coral mound area at the Southwest Rockall Trough margin, NE Atlantic. Deep-Sea Res. I 56, 10261038. Morrison, C., Ross, S., Nizinski, M., Brooke, S., Järnegren, J., Waller, R., Johnson, R. & King, T. (2011) Genetic discontinuity among regional populations of Lophelia pertusa in the North Atlantic Ocean. Cons. Genetics 12, 713-729 Mortensen, P.B., Hovland, M., Brattegard, T. & Farestveit, R. (1995) Deep-water bioherms of the scleractinian coral Lophelia pertusa (L) at 64 degrees N on the Norwegian Shelf – Structure and associated megafauna. Sarsia 80, 145-158. Myers, A.A. & Hall-Spencer, J.M. (2004) A new species of amphipod crustacean, Pleusymtes comitari sp. nov., associated with Acanthogorgia sp. gorgonians on deep-water coral reefs off Ireland. J. Mar. Biol. Ass. UK 84, 1029-1032. Narayanaswamy, B.E., Howell, K.L., Hughes, D.J., Davies, J.S., Roberts, J.M. & Black, K.D. (2006) Strategic Environmental Assessment Area 7 Photographic Analysis Report. 13. Department of Trade and Industry, Strategic Environmental Assessment Report, UK, p. 179. Unpublished report. Neat, F.C. & Burns, F. 2010. Stable abundance, but changing size structure in grenadier fishes (Macrouridae) over a decade (1998-2008) in which deepwater fisheries became regulated. Deep Sea Res. I. 57, 434-440. Neat, F. & Campbell, N. 2010. Demersal fish diversity of the isolated Rockall plateau compared with the adjacent west coast shelf of Scotland. Biol. J. Linn. Soc. Lond. 104, 138-147. Penny, A.J., Parker, S.J. & Brown, J.H. (2009) Protection measures implemented by New Zealand for vulnerable marine ecosystems in the South Pacific Ocean. Mar Ecol. Prog. Ser. 397, 341-354. Pollock, C & Barton, C. 2006. Offshore seabirds in the SEA 7 area. A report to the UK Department of Trade and Industry. Rice, A.L., Billett, D.S.M., Thurston, M.H. and Lampitt, R.S. (1991). The Institute of Oceanographic Sciences Biology Programme in the Porcupine Seabight: background and general introduction. J. Mar. Biol. Ass. U.K., 71, 281310. Roberts, J.M., Harvey, S.M., Lamont, P.A., Gage, J.D. & Humphery, J.D. (2000) Seabed photography, environmental assessment and evidence for deep-water trawling on the continental margin west of the Hebrides. Hydrobiologia 441: 173-183 286 ICES Advice 2013, Book 1 Roberts, J.M. and shipboard party (2013) Changing Oceans Expedition 2012. RRS James Cook cruise 073 Cruise Report. Heriot-Watt University. 224 pp. Roberts, J.M., Henry, L-A., Long, D. & Hartley, J.P. (2008) Cold-water coral reef frameworks, megafaunal communities and evidence for coral carbonate mounds on the Hatton Bank, north east Atlantic. Facies 54: 297-316 Roberts, J.M., Long, D., Wilson, J.B., Mortensen, P.B. & Gage, J.D. (2003) The cold-water coral Lophelia pertusa (Scleractinia) and enigmatic seabed mounds along the north-east Atlantic margin: are they related? Mar. Poll. Bull. 46, 7-20. Roberts, J.M., Wheeler, A., Freiwald, A. & Cairns, S.D. (2009) Cold-water corals: The biology and geology of deep-sea coral habitats. Cambridge University Press. 334pp. Roberts, J.M., Wheeler, A.J. & Freiwald, A. (2006) Reefs of the deep: The biology and geology of cold-water coral ecosystems. Science 213, 543-547. Rogers, A.D. & Gianni, M. (2010) The implementation of the UNGA Resolutions 61/105 and 64/72 in the management of deep-sea fisheries on the High Seas. Report of the Deep-Sea Conservation Coalition. International Programme on the State of the Ocean, London, UK. 97pp. Sayago-Gil M, Long D, Hitchen K, Díaz-del-Río V, Fernández-Salas LM, Durán-Muñoz P (2010) Evidence for currentcontrolled morphology along the western slope of Hatton Bank (Rockall Plateau, NE Atlantic Ocean). Geo-Mar. Lett. 30, 99-111 Söffker, M., Sloman, K.A. & Hall-Spencer, J.M. (2011) In situ observations of fish associated with coral reefs off Ireland. Deep-Sea Res. I 58, 818-825 Stewart, H A, and Davies, J S. 2007. Habitat investigations within the SEA7 and SEA4 areas of the UK continental shelf (Hatton Bank, Rosemary Bank, Wyville Thomson Ridge and Faroe–Shetland Channel). British Geological Survey Commercial Report, CR/07/051. Tittensor, D.P., Baco-Taylor, A.R., Brewin, P., Clark, M.R., Consalvey, M., Hall-Spencer, J.M., Rowden, A.A., Schlacher, T., Stocks, K. & Rogers, A.D. (2009) Predicting global habitat suitability for stony corals on seamounts. J. Biogeog. 36, 1111-1128 Van Dover, C., Smith CR, Ardron J, Dunn D, Gjerde K, Levin L, Smith S and the Dinard Workshop Contributors (2011a). Uncharted waters: Placing deep-sea chemosynthetic ecosystems in reserve. Marine Policy 36, 378-381. Van Dover, C., Smith CR, Ardron J, Dunn D, Gjerde K, Levin L, Smith S and the Dinard Workshop Contributors (2011b). Environmental management of deep-sea chemosynthetic ecosystems: justification of and considerations for a spatially-based approach. International Seabed Authority Technical Study 9. 29pp. Vanreusel, A., Fonseca, G., Danovaro, R., et al. (2010) The contribution of deep-sea macrohabitat heterogeneity to global nematode diversity. Mar. Ecol. 31, 66-77. Wei, C-L. et al. (2010). Global Patterns and Predictions of Seafloor Biomass Using Random Forests. PLoS One 5 (12) http://dx.plos.org/10.1371/journal.pone.0015323. Wilson, J.B., (1979a) The distribution of the coral Lophelia pertusa (L.) [L. prolifera (Pallas)] in the North-East Atlantic. J. Mar. Biol. Ass. UK 59, 149-164. Wilson, J.B., (1979b) ‘Patch’ development of the deep-water coral Lophelia pertusa (L.) on Rockall Bank. J. Mar. Biol. Ass. UK 59, 165-177. ICES Advice 2013, Book 1 287 Maps and Figures Figure 1 288 Map of the ABNJ area in the NE Atlantic with boundary of the Hatton-Rockall EBSA outlined in red. This boundary approximates the 1500 m contour. ICES Advice 2013, Book 1 Table 2 Contributors of data for the analysis of seabird satellite tracking are as follows; full details about each dataset are available via www.seabirdtracking.org Species Site Owner Corys Shearwater Azores J. González-Solís Corys Shearwater Balearics J. González-Solís Corys Shearwater Canaries J. González-Solís Corys Shearwater Chafarinas J. González-Solís Corys Shearwater Berlengas P. Catry, J.P. Granadeiro Corys Shearwater Selvagens M.A. Dias, P. Catry Corys Shearwater Selvagens M.A. Dias, J.P. Granadeiro Corys Shearwater Veneguera J. González-Solís Sooty Shearwater Bay Fundy R. Ronconi Sooty Shearwater Falklands A. Hedd Sooty Shearwater Gough A. Hedd Great Shearwater Bay Fundy R. Ronconi Great Shearwater Inaccessible Island R. Ronconi, P. Ryan, M. Caroline Martin Manx Shearwater Iceland I.A. Sigurðsson, Y. Kolbeinsson, J. González-Solís Manx Shearwater UK A. Ramsay, J. González-Solís Fea’s Petrel Bugio I. Ramirez, V. Paiva Black-legged Kittiwake Norway T. Boulinier, D. Gremillet, J. González-Solís Little Shearwater Azores V. Neves, J. González-Solís Zino’s Petrel Madeira F. Zino, R.A. Phillips, M. Biscoito Figure 2 Species occurrence by month within the Hatton-Rockall area, showing percentage of tracked population for each species (and where relevant subpopulation) found within the area each month. Rights and permissions – Any requests to use the seabird tracking data shown for this site in any publication need to be agreed with the data owners. An initial request should be sent to BirdLife International to coordinate this process. See http://www.seabirdtracking.org/terms.php for full terms of reference ICES Advice 2013, Book 1 289 ANNEX 2 Draft Proforma: Mid-Atlantic Ridge North of the Azores and South of Iceland Presented by: Based on the Joint OSPAR/NEAFC/CBD Scientific Workshop. Reviewed and revised by an ICES expert group. Abstract The Mid-Atlantic Ridge (MAR) is the major topographic feature of the Atlantic Ocean. Within the OSPAR/NEAFC area the Northern MAR separates the Newfoundland and Labrador Basins from the West-European Basin and the Irminger from the Iceland Basin. The ridge crest is generally cut by a deep rift valley along its length, bordered by high rift mountains, which are bordered by high fractured plateaus. This region is largely composed of volcanic rock and is the foundation of the proposed EBSA. Hydrothermal venting occurs along the ridges and small-scale physiographic features, including many small volcanoes (seamounts) and canyons, form near the ridge axis. The Moytirra vent field, within this EBSA, is the only high temperature hydrothermal vent known between the Azores and Iceland. Endemic vent fauna are associated with thermally active areas. The 2,500 m depth contour is used to inform the EBSA boundary. Introduction Mid-ocean ridge systems occupy a third of the ocean floor and are the site of the formation of new Earth’s crust (Heezen 1969). The Mid-Atlantic Ridge (MAR), a tectonic continental plate boundary, is the major topographic feature of the Atlantic Ocean, extending over 12,000 km from Iceland to the Bouvet Triple Junction in the South Atlantic (Figure 1). It divides the ocean longitudinally into two halves, each cut by secondary transverse ridges and interrupted by strike-slip transform faults that offset the ridge in opposing directions on either side of the axis of seafloor spreading (e.g., the Charlie Gibbs Fracture at 53ºN). Compared with other mid-ocean ridges, the MAR is a slow-spreading ridge where new oceanic floor is formed with an average spreading rate of 2.5 cm per year (Malinverno 1990). Hydrothermal venting occurs along the ridges and small-scale physiographic features, including many small volcanoes (seamounts) and canyons, form near the ridge axis; the crest consists mostly of hard volcanic rock. The MAR is an area which captures the Earth’s geological history, with outstanding representation of the major stages of Earth’s history, including the record of life, significant ongoing geological processes in the development of landforms and significant geomorphic or physiographic features. The general physiography of the MAR was documented some time ago (Heezen el al. 1959). The ridge crest is generally notched by a deep rift valley along its length, bordered by high rift mountains, which in turn are bordered by high fractured plateaus (Heezen et al. 1959). These crest zones are generally well defined and present along the full length of the MAR (Malinverno 1990). At approximately 50 -75 km from the axis of the ridge, the crest merges with sediment covered flanks which extend down to the abyssal plain (van Andel and Bowin 1968). The flanks are composed of a succession of smooth shelves, each from 2 to 100 km from the central axis and subdivided into upper, middle, and lower steps (Heezen el al. 1959) extending in some areas to depths of 4,572 m (Tolstoy and Ewing 1949). The flanks are generally covered with soft sediments. However, van Andel and Bowin (1968) describe considerable variability in sediment depth in the southern MAR, where the foothill region west of the ridge and the ridge slope are only thinly covered, while sedimentation in some valleys can range from nothing to a thickness of several hundred meters. The depth of the ridge crest is highly variable along its length. Malinverno (1990) conducted 46 profiles across the ridge axis from 0° to 50°N, with most of those conducted south of the Azores between 10° and 35°N. The average depth of the axial crest in those profiles was approximately 2,300 m but Malinverno demonstrated that depth was correlated with distance from the Azores and fracture zone characteristics. The MAR is divided into the Northern and Southern ridges near the equator by the deep Romanche Trench. Within the OSPAR area the Northern MAR separates the Newfoundland and Labrador Basins from the West-European Basin and the Irminger from the Iceland Basin. It has a profound role in the circulation of the water masses in the North Atlantic (Rossby 1999, Bower et al. 2002, Søiland et al. 2008) with currents crossing the MAR over deep gaps in the ridge and influencing upper-ocean circulation patterns (Bower et al. 2002). Canyons cut into the flanks may influence upward fluxes of water and abyssal mixing (Speer and Thurnherr 2005). Studies of volcanic rocks from the submerged MAR suggest that it consists largely of tholeiitic basalt with low values of K, Ti, and P. In contrast, the volcanic islands which form the elevated caps on the Ridge are built of alkali basalt with high values of Ti, Fe3+, P, Na, and K (Engel and Engel 1964). Variations in mineral content result from chemical and isotopic heterogeneity in the mantle (White and Schilling 1978). 290 ICES Advice 2013, Book 1 Moytirra Vent Field The Moytirra vent field is the only fully described high temperature hydrothermal vent known between the Azores and Iceland, making it a unique geophysical structure in the high seas of the North Atlantic and within the MAR. It is situated at 45°N on the 300 m high fault scarp of the eastern axial wall of the MAR, 3.5 km from the axial volcanic ridge crest (Wheeler et al. 2013). It is basalt-hosted and its position suggests that it is heated by an off-axis magma chamber. This type of base rock causes precipitation of iron and sulphide-rich minerals during mixing of the hot hydothermal vent fluids (200-400°C) with cold, oxygenated sea water- hence the term “black smoker” (Figure 2). The Moytirra vent field consists of three active vent sites emitting “black smoke" and producing a complex of chimneys and beehive diffusers. The largest chimney is 18 m tall and very actively venting. There may also be further unconfirmed vent sites on the MAR at 43 N, at 45 N and on the Reykjanes Ridge. In these areas, plumes and/or anomalously high concentrations of Mn in the water column have been detected http://www.interridge.org/irvents/ Location The Mid-Atlantic Ridge (MAR) extends over 12,000 km from Iceland to the Bouvet Triple Junction in the South Atlantic (Figure 1) and falls within the national jurisdictions of Iceland and the Azores. The proposed EBSA Mid-Atlantic Ridge North of the Azores and South of Iceland is for a portion of the MAR within the high-seas areas of OSPAR and NEAFC (Figure 3). Although the crest has an average depth of approximately 2,300 m (Malinverno 1990) it is variable, and the 2,500 m depth contour was used to inform the boundaries of the proposed EBSA as this captures the majority of the ridge crest, and known distribution of deep-water corals (maximum 2,400 m) (Figure 3, Table 1). Within the proposed EBSA the smaller physiographic feature, the Moytirra Vent Field, is located at latitude 45.833 and longitude -27.85 (Table 1). Feature description The Mid-Atlantic Ridge North of the Azores and South of Iceland is a unique geomorphological feature to the North Atlantic Ocean and to the high-seas areas of NEAFC and OSPAR. Within this feature is a smaller unique feature, the Moytirra vent field. The Moytirra vent field is the only high temperature hydrothermal vent known between the Azores and Iceland, making it a unique geophysical structure in the high seas of the North Atlantic and within the MAR. The fauna of the Northern MAR have not been fully described and it is premature to speculate on whether any species are endemic, excepting vent-endemic organisms associated with the hydrothermal vents. Some new species have been described and these may prove to be endemic to the proposed EBSA with further sampling. The benthic fauna associated with the Northern MAR are known from detailed observations at a few locations. Priede et al. (2013) used a variety of sampling gears to survey habitat, biomass and biodiversity in a segment of the Northern MAR as part of a multinational and multidisciplinary project (MAR-ECO). They found that primary production and export flux over the MAR were not enhanced compared with a nearby reference station over the Porcupine Abyssal Plain and biomass of benthic macrofauna and megafauna were similar to global averages at the same depths. Also as part of MAR-ECO, Mortensen et al. (2008) used an ROV to conduct video surveys along the MAR at 8 sites between the Reykjanes Ridge and the Azores. Deep-water corals were observed at all locations at depths less than 1400 m (range 800-2400 m) and 40 coral taxa were observed, including observations of patches of Lophelia pertusa. Crinoids, sponges, the bivalve Acesta excavata, and squat lobsters were associated with the Lophelia. None of those corals were recognized as new species to science and all likely have broader distributions extending along the continental slopes and seamounts at similar latitudes in the North Atlantic. Inevitably, 11 new species have been described arising from the MAR-ECO work and more are likely to be discovered as the samples are fully processed. These include a new genus and species of foram (Incola arantius gen. et sp. nov.), two new species of glass sponges of the genus Sympagella (Rossellidae), mushroom corals (Anthomastus gyratus sp. nov. and Heteropolypus sol sp. nov.), a deep-sea scavenging amphipod (Hirondellea namarensis sp. nov.), two new starfish (species of Hymenaster) and three species of elasipodid holothurians (Gebruk et al. 2013). MAR-ECO midwater and bottom trawls collected 54 species of cephalopods in 29 families (Vecchione et al. 2010). The squid Gonatus steenstrupi was the most abundant cephalopod in the samples, followed by the squids Mastigoteuthis agassizii and Teuthowenia megalops. A multispecies aggregation of large cirrate octopods dominated the demersal cephalopods. The demersal fish fauna of the MAR form two distinct groups with a faunal divide between 48 and 52°N (Hareide and Garnes 2001) and species-specific differences with depth (King et al. 2006, Bergstad et al. 2008). Hareide and Garnes (2001), using one trawl and three longline surveys, identified 56 species from 27 families of fish from between 400 and 2000 m depth along the MAR. In the northern part of the Ridge (north of 52°N) relatively common sub-Arctic species such as Sebastes spp., tusk (Brosme brosme) and Greenland halibut (Reinhardtius hippoglossoides) were dominant while ICES Advice 2013, Book 1 291 sub-tropical species such as golden eye perch (Beryx splendens) and cardinal fish (Epigonus telescopus) were dominant species below 48°N. During the 2004 MAR-ECO expedition to the MAR, 8518 fish, representing 40 species and 17 families were caught with longlines (Fossen et al. 2008). The 59 longline sets were distributed across the ridge axis at depths ranging from 400 to 4300 m at two locations: just north of the Azores archipelago and in the Charlie–Gibbs Fracture Zone. Chondrichthyans (primarily Etmopterus princeps) dominated the catches and contributed nearly 60% to both total biomass and abundance. King et al. (2006) recorded the scavenging fishes of the MAR using a baited autonomous lander equipped with a time-lapse camera between 924 and 3420 m water depth along 3 east–west transects at 42, 51 and 53°N across the MAR. They photographed 22 taxa with Synaphobranchus kaupii, Antimora rostrata and Coryphaenoides (Nematonurus) armatus dominant. Abyssal species in the axial valley region were C. armatus, Histiobranchus bathybius and Spectrunculus sp. No endemic demersal species have been reported although the zoarcid eelpout Pachycara thermophilum, is a vent-endemic species associated with hydrothermal vents of the MAR (Geistdoerfer 1994). Sutton et al. (2008) examined the assemblage structure and vertical distribution of deep-pelagic fishes relative to MAR with acoustic and discrete-depth trawling surveys in association with MAR-ECO. A 36-station, zig-zag survey along the Mid-Atlantic Ridge from Iceland to the Azores covered the full depth range (0 to >3000 m), from the surface to near the bottom, using a combination of gear types to sample the pelagic fauna. Dominant families of pelagic fish included Gonostomatidae, Melamphaidae, Microstomatidae, Myctophidae, and Sternoptychidae and 99 species of pelagic fish were found concentrated particularly north of the Charlie-Gibbs Fracture Zone. Sutton et al. (2008) found that abundance per volume of deep-pelagic fishes was highest in the epipelagic zone and within the benthic boundary layer (BBL; 0-200 m above the seafloor) while minimum fish abundance occurred at depths below 2300 m but above the BBL. Biomass per volume of deep-pelagic fishes over the MAR reached a maximum within the BBL, revealing a topographic association of a bathypelagic fish assemblage with the mid-ocean ridge system. The dominant component of deep-pelagic fish biomass over the MAR was a wide-ranging bathypelagic assemblage that was remarkably consistent along the length of the ridge from Iceland to the Azores. The authors conclude that special hydrodynamic and biotic features of mid-ocean ridge systems cause changes in the ecological structure of deep-pelagic fish assemblages relative to those at the same depths over abyssal plains. Moytirra Vent Field Due to the unique nature of the Moytirra vent field, the specialized vent fauna associated with it are also unique to the North Atlantic high-seas area. Wheeler et al. (2013) have documented aggregations of gastropods (Peltospira sp.) and populations of alvinocaridid shrimp (Mirocaris sp. and Rimicaris sp.) on the surfaces of the vent chimneys in addition to bythograeid crabs (Segonzacia sp.) and zoarcid fish (Pachycara sp.), all considered hydrothermal vent fauna (van Dover 1995). Feature condition, and future outlook Given the geophysical nature, location and size of the MAR it is unlikely that it will be affected by human activities. Despite its remoteness, the fauna associated with the MAR are not pristine. Starting in the early 1970s with Soviet/Russian trawlers stocks of roundnose grenadier (Coryphaenoides rupestris), orange roughy (Hoplostethus atlanticus) and alfonsino (Beryx splendens) associated with the MAR were exploited (Clark et al. 2007, ICES 2007). It can be assumed that most hills along the ridge were at least explored (usually by midwater trawls operating close to the seafloor), and at least 30 seamounts were also exploited for C. rupestris. After 1982, the targeted fishery for redfish developed, dwarfing the catches of roundnose grenadier. After the transition from Soviet to Russian fisheries, the Russian fishing effort and absolute catch on the MAR was significantly reduced, although catch per fishing day settled at relatively low levels by the end of 1990s and the fishery was still conducted periodically (ICES 2007). The fishery on C. rupestris takes deepwater redfish (Sebastes spp), orange roughy (H. atlanticus), blackscabbard fish (Aphanopus carbo) and deepwater sharks as bycatch (Clark et al. 2007). Longline fishing and near-bottom pelagic trawls have the potential to damage fragile benthic species such as deep-water corals and sponges. The scale of the impact that fishing and other human activities may have had on the MAR fauna is at present unquantified. In 2009 NEAFC adopted measures that close more than 330,000 km2 to bottom fisheries on the MAR until 2015 (Figure 4). According to the International Seabed Authority, exploratory mining for sulfide deposits has already been undertaken on the MAR, while ferromanganese nodules and cobalt-rich ferromanganese crusts have potential for mining interests. The potential impacts on the marine environment are removal of organisms and their habitats along with the mineral deposits and the smothering of adjacent communities by any sediment plume that may be created (http://www.isa.org.jm/en/about/faqs#16 ). Representatives from Norway, Iceland, Azores,the United Kingdom, IUCN and UNESCO have met to review the geological and biological heritage of the MAR in the North Atlantic (http://whc.unesco.org/en/activities/504/ ). Discussions focused on the areas where the highest peaks of the mountain chain reach sea-level and form islands. Most 292 ICES Advice 2013, Book 1 of the underwater ridge was not considered because it lies outside any national territory and therefore is not covered by provisions of the World Heritage Convention. There was agreement to encourage cooperation with other conventions in order to better protect the biological, cultural and geological heritage of the ridge. Assessment against CBD EBSA Criteria CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Explanation for ranking Uniqueness or rarity High X The Mid-Atlantic Ridge (MAR) qualifies as a unique geomorphological feature in the North Atlantic. The Moytirra vent field is the only known high temperature hydrothermal vent between the Azores and Iceland, making it a unique geophysical structure in the high seas of the North Atlantic and within the MAR. Areas that are required for a population to survive and Special importance for thrive life-history stages of species Explanation for ranking Data deficient Importance for Area containing habitat for the survival and recovery of endangered, threatened, declining species or area with threatened, endangered or significant assemblages of such species declining species and/or habitats Explanation for ranking X X Data deficient Areas that contain a relatively high proportion of sensitive Vulnerability, habitats, biotopes or species that are functionally fragile fragility, sensitivity, or (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery slow recovery Explanation for ranking X Deep-water corals were observed at all 8 sites from 3 locations along the proposed EBSA at depths less than 1400 m (range 8002400 m) and 40 coral taxa were observed, including observations of patches of Lophelia pertusa. These taxa are fragile with slow recovery and highly susceptible to degradation or depletion by human activities including contact with bottom fishing gear (longlines, pots, trawls). Area containing species, populations or communities with Biological X comparatively higher natural biological productivity productivity Explanation for ranking The research conducted through the MAR-ECO project found that primary production and export flux over the MAR were not enhanced compared with a nearby reference station over the Porcupine Abyssal Plain and biomass of benthic macrofauna and megafauna were similar to global averages at the same depths. There is some evidence for pelagic fish concentrating in the benthic boundary layer (to 200 m above the seafloor) over the MAR in association with topographic features. Area contains comparatively higher diversity of ecosystems, habitats, communities, or species, or has higher genetic diversity Explanation for ranking Biological diversity X Data deficient. Diversity of habitats is greater than that of surrounding abyssal plain but with the exception of the vent fauna, habitats and species are generally shared with continental margins and seamounts not associated with the MAR. ICES Advice 2013, Book 1 293 Sharing experiences and information applying other international criteria (Optional) CBD EBSA Criterion Description Dependency: An area where ecological processes are highly dependent on biotically structured systems (e.g., coral reefs, kelp forests, mangrove forests, seagrass beds). Such ecosystems often have high diversity, which is dependent on the structuring organisms. Dependency also embraces the migratory routes of fish, reptiles, birds, mammals, and invertebrates. Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know Explanation for ranking Representativeness: An area that is an outstanding and illustrative example of specific biodiversity, ecosystems, ecological or physiographic processes Explanation for ranking Biogeographic importance: An area that either contains rare biogeographic qualities or is representative of a biogeographic “type” or types, or contains unique or unusual biological, chemical, physical, or geological features X Explanation for ranking The Mid-Atlantic Ridge (MAR) qualifies as a unique geomorphological feature in the North Atlantic. The Moytirra vent field is the only known high temperature hydrothermal vent between the Azores and Iceland, making it a unique geophysical structure in the high seas of the North Atlantic and within the MAR. Fauna endemic to the vents have adapted to the chemical and thermal properties of the environment. Structural complexity: An area that is characterized by complex physical structures created by significant concentrations of biotic and abiotic features. Explanation for ranking Natural Beauty: An area that contains superlative natural phenomena or areas of exceptional natural beauty and aesthetic importance. Explanation for ranking Earth’s geological history: An area with outstanding examples representing major stages of Earth’s history, including the record of life, significant on-going geological processes in the development of landforms, or significant geomorphic or physiographic features. X Explanation for ranking The Mid-Atlantic Ridge is the site of significant on-going geological processes (plate tectonics, formation of new Earth’s crust) and of significant physiographic features (axial rift valley, hydrothermal vent fields). [Other relevant criterion] Explanation for ranking [Other relevant criterion] Explanation for ranking 294 ICES Advice 2013, Book 1 References Bergstad, O.A., Menezes,G. and Å.S. Høines. 2008. Demersal fish on a mid-ocean ridge: Distribution patterns and structuring factors. Deep Sea Research Part II: Topical Studies in Oceanography 55(1–2): 185-202. Bower, A.S., Le Cann, B., Rossby, T., Zenk, W., Gould, J., Speer, K., Richardson, P.L., Prater, M.D. and H.-M. Zhang. 2002. Directly measured mid-depth circulation in the northeastern North Atlantic Ocean. Nature 419: 603- 607. Clark, M.R., Vinnichencko, V.I., Gordon, J.D.M., Beck-Bulat, G.Z., et al. 2007. Large-scale distant-water trawl fisheries on seamounts. In: Pitcher, T.J., Morato, T., Hart, P.J.B., Clark, M.R., Haggan, N. and Santos, R.S. (eds) Seamounts: Ecology, Conservation and Management. Fish and Aquatic Resources Series, Blackwell, Oxford, UK. Chapter 17, pp. 361 – 399. Engel, A.E.J. and C.G. Engel. 1964. Composition of basalts from the Mid-Atlantic Ridge. Science 144 (3624): 13301333. Fossen, I., Cotton, C.F., Bergstad, O.A. and J.E. Dyb. 2008. Species composition and distribution patterns of fishes captured by longlines on the Mid-Atlantic Ridge. Deep Sea Research Part II: Topical Studies in Oceanography 55(1–2): 203-217. Gebruk , A.V., Priede, I.G., Fenchel T. and F. Uiblein. 2013. Benthos of the sub-polar front area on the Mid-Atlantic Ridge: Results of the ECOMAR project. Marine Biology Research 9(5-6): 443-446. Geistdoerfer, P. 1994. Pachycara thermophilum, une nouvelle espèce de poisson Zoarcidae des sites hydrothermaux de la dorsale médio-atlantique. Cybium 18: 109-115. Hareide, N.-R. and G. Garnes. 2001. The distribution and catch rates of deep water fish along the Mid-Atlantic Ridge from 43 to 61°N. Fisheries Research 51(2–3): 297-310. Heezen, B.C.,Tharp, M. and M. Ewing. 1959. The Floors of the Oceans: I. The North Atlantic. The Geological Society of America Special Paper 65, 122 pp. Heezen, BC. 1969. The world rift system: An introduction to the symposium. Tectonophysics 8:269-279. ICES. 2007. Report of the Working Group on Deep-water Ecology (WGDEC), 26 – 28th February. ICES CM 2007/ACE:01 Ref. LRC. 61pp. King, N.J., P.M. Bagley and I.G. Priede. 2006. Depth zonation and latitudinal distribution of deep-sea scavenging demersal fishes of the Mid-Atlantic Ridge, 42 to 53ºN. Marine Ecology Progress Series 319: 263-274. Malinverno, A. 1990. A quantitative study of axial topography of the Mid-Atlantic Ridge. Journal of Geophysical Research 95: 2645-2660. Mortensen, P.B., Buhl-Mortensen, L., Gebruk, A.V. and E. M. Krylova. 2008. Occurrence of deep-water corals on the Mid-Atlantic Ridge based on MAR-ECO data. Deep-Sea Research II 55:142-152. Priede, I.G., Bergstad, O.A., Miller, P.I., Vecchione, M., Gebruk, A., et al. 2013. Does Presence of a Mid-Ocean Ridge Enhance Biomass and Biodiversity? PLoSONE 8(5): e61550. doi:10.1371/journal.pone.0061550 Rossby, T. 1999. On gyre interactions. Deep-Sea Research II 46: 139-164. Søiland, H., Budgell, W.P. and Ø Knutsen. 2008. The physical oceanographic conditions along the Mid-Atlantic Ridge north of the Azores in June-July 2004. Deep-Sea Research II 55: 29- 44. Speer, K. G. and A. M. Thurnherr. 2005. Abyssal Canyons and Mixing by Low-Frequency Flow. In: P. Muller and D. Henderson (Editors), Near-Boundary Processes and Their Parameterization, topics in physical oceanography, 'Aha Huliko' a Winter Workshop, pp. 17-19. Sutton, T.T., Porteiro, F.M., Heino, M., et al. 2008. Vertical structure, biomass and topographic association of deeppelagic fishes in relation to a mid-ocean ridge system. Deep-Sea Research II 55 (1–2):161–184. Tolstoy, I. and M. Ewing. 1949. North Atlantic hydrography and the Mid-Atlantic Ridge. Bulletin of the Geological Society of America 60(10): 1527-1540. van Andel, Tj. H. and C. O. Bowin. 1968. Mid-Atlantic Ridge between 22º and 23º north latitude and the tectonics of mid-ocean rises. Journal of Geophysical Research 73: 1279-1298. Van Dover, C.L. 1995. Ecology of Mid-Atlantic Ridge hydrothermal vents. Geological Society, London, Special Publications 1995, v. 87, p257-294. Vecchione, M., Young, R.E. and U. Piatkowski. 2010. Cephalopods of the northern Mid-Atlantic Ridge. Marine Biology Research 6: 25–52. Wheeler, A.J., Murton, B., Copley, J., Lim, A., Carlsson, J. et al. 2013. Moytirra: discovery of the first known deep-sea hydrothermal vent field on the slow-spreading Mid-Atlantic Ridge north of the Azores. Geochemistry, Geophysics, Geosystems, doi: 10.1002/ggge.20243. White, W.M. and J.-G. Schilling. 1978. The nature and origin of geochemical variation in Mid-Atlantic Ridge basalts from the Central North Atlantic. Geochimica et Cosmochimica Acta 42 (10): 1501-1516. ICES Advice 2013, Book 1 295 Tables, Maps and Figures Table 1 Boundaries for the proposed EBSA Mid-Atlantic Ridge North of the Azores and South of Iceland and location of the Moytirra Vent Field (see Figure 3). Feature Point 1 Point 2 Point 3 Point 4 Point 5 Point 6 EEZ of the Azores Latitude 63.59 55.85 52.97 52.50 47.11 43.18 Point 7 Point 8 Point 9 Point 10 Point 11 Point 12 EEZ of Iceland 42.50 46.43 49.43 52.50 53.78 60.13 Moytirra Vent Field 296 (dd) Longitude (dd) -30.91 -37.51 -36.70 -33.00 -28.09 -30.59 Details of Location Reference easterly following the EEZ boundary for the Azores to Point 7 45.4833 -27.55 -25.91 -26.63 -29.80 -33.69 -22.84 -27.85 westerly following the EEZ boundary for Iceland to join Point 1 300 m high fault scarp of the eastern axial wall, 3.5 km from the axial volcanic ridge crest Wheeler et al. (2013) ICES Advice 2013, Book 1 Figure 1 Location of the Mid-Atlantic Ridge (dashed lines). Image downloaded from: commons.wikimedia.org File:Mid-atlantic ridge.jpg - Wikimedia Commons. ICES Advice 2013, Book 1 297 Figure 2 298 Volcanically heated fluid rises from a deep-sea "black-smoker" in the Moytirra vent field. (Photo http://news.nationalgeographic.com/news/2011/08/110808-hydrothermal-ventsdownloaded from volcanic-animals-ocean-deep-sea-science-alien/ ) ICES Advice 2013, Book 1 Figure 3 Location of the proposed EBSA Mid-Atlantic Ridge North of the Azores and South of Iceland. The perimeter of the proposed boundary follows Table 1 with points numbered 1 through 12. The blue shaded areas represent the Exclusive Economic Zones of countries in the region. The light green line outlines the outer boundary of the OSPAR Convention Area which coincides with the NEAFC Convention Area at its western and southern boundary. Seafloor bathymetry is indicated in grayscale with the 2000 m (red) and 2500 m (light blue) General Bathymetric Chart of the Oceans (GEBCO) depth contours indicated. The red circle with the central star marks the location of the Moytirra Vent Field. See Table 1 for positional information (latitude and longitude). ICES Advice 2013, Book 1 299 Figure 4 300 Location of areas closed to bottom fishing (2009-2015) in the NEAFC regulatory area which includes the MAR http://www.neafc.org/page/closures. There is a small seasonal closure for Blue Ling near the boundary with Iceland EEZ which is not visible at this scale. ICES Advice 2013, Book 1 ANNEX 3 Draft Proforma: Charlie-Gibbs Fracture Zone Presented by: Based on the Joint OSPAR/NEAFC/CBD Scientific Workshop. Reviewed by revised by an ICES expert group Abstract Fracture zones are common topographic features of the global oceans that arise through plate tectonics. The CharlieGibbs Fracture Zone is an unusual left lateral strike-slip double transform fault in the North Atlantic Ocean along which the rift valley of the Mid-Atlantic Ridge is offset by 350 km near 52º30′N. It opens the deepest connection between the northwest and northeast Atlantic (maximum depth of approximately 4500 m) and is approximately 2000 km in length extending from about 25°W to 45°W. It is the most prominent interruption of the MAR between the Azores and Iceland and the only fracture zone between Europe and North America that has an offset of this size. Two named seamounts are associated with the transform faults: Minia and Hecate. The CGFZ is considered a unique geomorphological feature in the North Atlantic under the EBSA criteria; further, it captures the Earth’s geological history, including significant ongoing geological processes. Introduction Fracture zones are common topographic features of the global oceans that arise through plate tectonics. They are characterized by two strongly contrasting types of topography. Seismically active transform faults form near mid-ocean ridges where the continental plates move in opposing directions at their junction. Seismically inactive fracture zones, where the plate segments move in the same direction, extend beyond the transform faults often for 100s of kilometers. Their atypical crust thickness that can be as little as 2 km (Mutter et al. 1984, Cormier et al. 1984, Calvert and Whitmarsh 1986) allowing direct seismic investigations of the internal structure and composition of oceanic crusts used to model processes of seafloor spreading. In the Atlantic Ocean most fracture zones originate from the Mid-Atlantic Ridge (MAR) and are nearly perfectly west - east oriented. There are about 300 fracture zones occurring on average every 55 km along the ridge, with the offsets created by transform faults ranging from 9 to 400 km in length (Müller and Roest 1992). The Charlie-Gibbs Fracture Zone (CGFZ) is an unusual left lateral strike-slip double transform fault in the North Atlantic Ocean along which the rift valley of the MAR is offset by 350 km near 52º30′N (Figure 1). It opens the deepest connection between the northwest and northeast Atlantic (maximum depth of approximately 4500 m; Fleming et al. 1970) and is approximately 2000 km in length extending from about 25°W to 45°W. It is the most prominent interruption of the MAR between the Azores and Iceland and the only fracture zone between Europe and North America that has an offset of this size 22. Knowledge of its geomorphology is considered essential to the understanding of the plate tectonic history of the Atlantic north of the Azores (Olivet et al. 1974). For these reasons it is consider a unique geomorphological feature in the North Atlantic under the EBSA criteria; further, it captures the Earth’s geological history, including significant on-going geological processes. The CGFZ is comprised of two narrow parallel fracture zones (Fleming et al. 1970) which form deep trenches located at 30ºW (Charlie-Gibbs South Transform Fault) and at 35º15′W (Charlie-Gibbs North Transform Fault) and separated by a short (40 km) north-south seismically active (Bergman and Solomon 1988) spreading center (median transverse ridge) at 31º45’W (Figure 2; Searle 1981; Fleming et al. 1970, Olivet et al. 1974). The southern fault displaces the MAR, coming from the Azores, to the west over a distance of 120 km. It is at most 30 km wide (Searle 1981). The northern fault displaces the spreading ridge over another 230 km to the west before it connects to the northern part of the MAR going to Iceland. Both transform faults continue eastward and westward as inactive fracture zones (Figure 2). The CGFZ is characterized by rough morphology and the walls of the fracture valleys and the ridge in between them are broken and irregular with slopes of up to 29° (Fleming et al. 1970). The height of the ridge between the faults is at least 1000 m below the surface and as shallow as 636 m in parts (Fleming et al. 1970). Rock samples show the walls of the fracture zone to be both basaltic and ultramafic while the median transverse ridge contains gabbro (Hekinian and Aumento 1973). Earthquake epicentres are associated with the transform faults (Kanamori and Stewart 1976, Bergman and Solomon 1988) and an almost continuous belt of epicentres follow the southern end of the Reykjanes Ridge, along the northern transform valley, the central median valley and the southern transform valley to the north end of the MAR (Lilwall and Kirk 1985). Two named seamounts are associated with the transform faults: Minia Seamount (53°01′N 34°58′W) located near the junction of the Reykjanes Ridge and the northern transform fault and Hecate Seamount 22 The Spitzbergen and Jan Mayen fracture zones, of comparable offset (145 and 211 km respectively), lie between Greenland and Europe. ICES Advice 2013, Book 1 301 (52°17′N 31°00′W) located on the northern wall of the southern transform fault east of the short median transverse ridge (Figure 2; Fleming et al. 1970). Ridge and troughs along the CGFZ are mostly covered with muddy sediments (Fleming et al. 1970) although outcrops of sedimentary rock and boulder fields are exposed by recent faulting and current scour (Shor et al. 1980, Searle 1981) and the southern transform near 30°30′W has no sediment cover (Searle 1981). Considerable thicknesses of sediment are deposited in the northern transform valley from the Iceland-Scotland Overflow Water (ISOW) which carries a significant load of suspended sediment (25 μg I-1) as it passes through (Shor et al. 1980). Transverse ridges prevent the sediment reaching the southern valley (Searle 1981) which has less sediment cover, although it is still considered a depositional environment (Shor et al. 1980). The topography of the CGFZ has a major influence on deep water oceanographic circulation (Harvey and Theodorou 1986). A large component of the North Atlantic Deep Water originates in the Norwegian Sea and flows south over the sills between Scotland and Iceland (ISOW). It meets the CGFZ near the intersection of the transform faults and the spreading centre (Shor et al. 1980). There is then a westward movement of deep water passing through the fracture zone from east to west through to the Irminger Sea occurring from the core depth of the ISOW at about 2500 m to the sea floor (Garner 1972, Shor et al. 1980, Saunders 1994). Most of this water is carried through the northern transform fault where the overflow water first encounters the fracture zone. The topography of the CGFZ also is thought to have some influence on the circulation of surface waters, although they are not locked to the bottom features to the same extent as the ISOW (Rossby 1999, Bower et al. 2002). The northern branch of the North Atlantic Current defines the location of the sub-polar front between colder Sub Arctic Intermediate Water to the north and warmer North Atlantic Intermediate Water to the south (Søiland et al. 2008). The sub-polar front meanders between 48-53°N and surface flow is predominantly eastward. The CGFZ is therefore not only a topographic discontinuity in the MAR but the area also constitutes an oceanographic transition zone between waters of different temperatures and flow regimes (Priede et al. 2013). Location The Charlie-Gibbs Fracture Zone occurs at 52º30′N and extends from about 25°W to 45°W with the transform faults occurring between 30°W and 35°W (Olivet et al. 1974). The proposed Charlie-Gibbs Fracture Zone EBSA within the high-seas area of NEAFC and OSPAR takes the co-ordinates provided in Table 1 and illustrated in Figure 3. These are based on Olivet et al. (1974) and the location of the Minia Seamount which influences the northern boundary of the EBSA. The eastern boundary of the CGFZ is detectable beyond 42°W, the outer boundary of the NEFAC/OSPAR jurisdictions. The southern ridge continues uninterrupted to 45°W (Olivet et al. 1974). Feature description The Charlie-Gibbs Fracture Zone (CGFZ) is a unique geomorphological feature to the North Atlantic Ocean and to the high-seas areas of NEAFC and OSPAR. Owing to its remoteness, the fauna associated with the CGFZ are poorly studied and it is premature to speculate on whether any species are endemic based on first descriptions. For example, Gebruk (2008) described two species of holothurians and believed them to be endemic to the Mid-Atlantic Ridge but they subsequently were found on the European continental margin in the Whittard Canyon (Masson 2009). As part of the MAR-ECO project (Priede et al. 2013) manned submersibles were deployed on the axis (52°47′N) and the northern slopes (52°58′N) of the Charlie–Gibbs North transform fault and surveyed macroplankton (Vinogradov 2005), demersal nekton (Felley et al. 2008) and invertebrate megafauna (Gebruk and Krylova 2013). Pelagic shrimps, chaetognaths and gelatinous animals were numerically dominant in the plankton, with peak densities corresponding to the main pycnocline. Mucous houses of appendicularians were abundant at 150 m above the seabed, although this is common throughout the central Atlantic and not associated with specific bottom topography (Vinogradov 2005). Nekton included large and small macrourids (Coryphaenoides spp.), shrimp (infraorder Penaeidea), Halosauropsis macrochir, Aldrovandia sp., Antimora rostrata, and alepocephalids (Felley et al. 2008). Glass sponges were common between 1700 and 2500 m while the deeper parts of the fracture wall and the sea floor were dominated by isidid corals, other anthozoans, squat lobsters and echinoderms, especially holothurians. The elpidiid holothurian, Kolga nana, occurred at high density in the abyssal depression (Gebruk and Krylova 2013). Rogacheva et al. (2013) recorded 32 holothurian species from the CGFZ area through the ECOMAR project (http://www.oceanlab.abdn.ac.uk/ecomar/), including three elasipodid holothurian species new to science. In general, none of the fauna documented from the CGFZ showed distributions atypical of similar habitats in the broader North Atlantic, although Gebruk and Krylova (2013) discuss the known distribution of the holothurian Peniagone longipaillata and remark on the differences in relative abundance observed between the occurrence of this species, where 302 ICES Advice 2013, Book 1 it is common in the lower bathyl of the CGFZ, and the continental slopes in the Porcupine Seabight and Abyssal Plain areas and Whitard Canyon where it appears less so. There is weak evidence that the CGFZ may be important for juvenile zoarcids based on a high percentage of those observed with baited cameras being <100 mm in length (Kemp et al. 2013). General knowledge of seafloor benthos suggests that where the geo-morphological processes of the fracture zone have created steep walls along the fractures, the greater three-dimensional topographic complexity, combined with the strong water flows through the fractures, creates habitat that is likely to be more productive and support greater concentrations of fragile taxa such as deep-water corals and sponges than adjacent habitats (Miller et al. 2012). The sampling done along the fracture zone supports these inferences but the differences from other habitats in similar depths and latitudes have not been quantified yet. Feature condition, and future outlook Given the geophysical nature, location and size of the Charlie-Gibbs Fracture Zone (CGFZ) it is unlikely that it will be affected by human activities, although there is potential for mining of the rare minerals associated with the transform faults. In 2010 the Environmental Ministers of the OSPAR countries officially designated a marine protected area of 145,420 km2 in the southern part of the Charlie-Gibbs Fracture Zone (Figure 4) and adopted “significant and innovative measures to establish and manage the southern part of the originally proposed Charlie-Gibbs Fracture Zone MPA – “Charlie-Gibbs South MPA”-, for which the seabed and super adjacent waters are situated in areas beyond national jurisdiction” (OSPAR Commission 2010). That same year (2010) the OSPAR Commission and the International Seabed Authority signed a memorandum of understanding in order to conciliate the development of mineral resources with comprehensive protection of the marine environment. In this MOU, the Charlie Gibbs Fracture Zone is highlighted as an area where consultation between the two parties had been initiated. In 2012 OSPAR countries designated “ Charlie-Gibbs North High Seas Marine Protected Area”, an area of high seas of approximately 177,700 km2 (OSPAR Commission 2012), complementing the Charlie-Gibbs South MPA established previously (Figure 4). The scale of the impact that fishing and other human activities have had on the fauna of the CGFZ is at present unquantified and likely to be minor, although fishing has been reported on the Hectate Seamount (ICES 2007). In 2009 NEAFC closed more than 330,000 km2 to bottom fisheries on the Mid-Atlantic Ridge, including a large section of the CGFZ which includes the transform faults and median transverse ridge (http://www.neafc.org/page/closures) (Figure 4). Assessment against CBD EBSA Criteria [Discuss the case study in relation to each of the CBD criteria and relate the best available science. Note that a candidate EBSA may qualify on the basis of one or more of the criteria, the boundaries of the EBSA need not be defined with exact precision. And modeling may be used to estimate the presence of EBSA attributes. Please note where there are significant information gaps.] CBD EBSA Criterion Description Ranking of criterion relevance (please mark one column with an X) Don’t Low Some Know The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Explanation for ranking Uniqueness rarity or High X The Charlie-Gibbs Fracture Zone (CGFZ) is a unique geomorpholical feature in the high-sea between the Azores and Iceland. It is the only fracture zone with an offset of its size (350 km) between Europe and North America and opens the deepest connection between the northwest and northeast Atlantic. The fact that it is a double transform fault is an unusual feature. Areas that are required for a population to survive and Special importance for thrive life-history stages of species Explanation for ranking X Data deficient ICES Advice 2013, Book 1 303 Importance for Area containing habitat for the survival and recovery of endangered, threatened, declining species or area with threatened, endangered or significant assemblages of such species declining species and/or habitats Explanation for ranking X Data deficient Areas that contain a relatively high proportion of sensitive Vulnerability, habitats, biotopes or species that are functionally fragile fragility, sensitivity, or (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery slow recovery Explanation for ranking X Glass sponges were observed on hard substrates on the fault wall at depths between 1700 and 2500 m. These taxa are fragile with slow recovery and highly susceptible to degradation or depletion by human activities including contact with bottom fishing gear (longlines, pots, trawls).Inferring from the frequently documented presence of such species and communities in structurally complex deep-sea habitats elsewhere, further sampling is likely to document additional presence of sensitive habitats, biotopes, or species in the CGFZ fractures, Area containing species, populations or communities with X Biological comparatively higher natural biological productivity productivity Explanation for ranking There is no evidence that the CGFZ contains comparatively higher natural productivity. The strong current flows through the fractures and complex three dimensional habitats create conditions that may enhance productivity, but at present there are insufficient data to rank on this criterion. Area contains comparatively higher diversity of Biological X ecosystems, habitats, communities, or species, or has diversity higher genetic diversity Explanation for ranking Diversity of habitats is greater than that of surrounding abyssal plain but biotic diversity is poorly quantified and there is little basis for a comparative assessment on this criterion at this time. Sharing experiences and information applying other international criteria (Optional) CBD EBSA Criterion Description Dependency: An area where ecological processes are highly dependent on biotically structured systems (e.g., coral reefs, kelp forests, mangrove forests, seagrass beds). Such ecosystems often have high diversity, which is dependent on the structuring organisms. Dependency also embraces the migratory routes of fish, reptiles, birds, mammals, and invertebrates. Ranking of criterion relevance (please mark one column with an X) Don’t Low Some High Know Explanation for ranking Representativeness: An area that is an outstanding and illustrative example of specific biodiversity, ecosystems, ecological or physiographic processes Explanation for ranking Biogeographic importance: An area that either contains rare biogeographic qualities or is representative of a biogeographic “type” or types, or contains unique or unusual biological, chemical, physical, or geological features X Explanation for ranking The CGFZ qualifies as a unique geomorphological feature in the North Atlantic being the largest transform fault separating Europe from North America. An area that is characterized by complex physical Structural complexity: structures created by significant concentrations of biotic and abiotic features. Explanation for ranking 304 ICES Advice 2013, Book 1 Natural Beauty: An area that contains superlative natural phenomena or areas of exceptional natural beauty and aesthetic importance. Explanation for ranking Earth’s geological history: An area with outstanding examples representing major stages of Earth’s history, including the record of life, significant on-going geological processes in the development of landforms, or significant geomorphic or physiographic features. X Explanation for ranking Fracture zones are of great geological interest due to their anomalous crust thickness that can be as little as 2 km allowing direct seismic investigations of the internal structure and composition of oceanic crusts used to model processes of seafloor spreading. Knowledge of the geomorphology of the CGFZ is considered essential to the understanding of the plate tectonic history of the Atlantic north of the Azores. [Other relevant criterion] Explanation for ranking [Other relevant criterion] Explanation for ranking References Bergman, E.A. and S.C. Solomon. 1988. Transform fault earthquakes in the North Atlantic: Source mechanisms and depth of faulting. Journal of Geophysical Research 93:9027-9057. Bower, A.S., Le Cann, B., Rossby, T., Zenk, W., Gould, J., Speer, K., Richardson, P.L., Prater, M.D. and H.-M. Zhang. 2002. Directly measured mid-depth circulation in the northeastern North Atlantic Ocean. Nature 419: 603- 607 Calvert, A.J. and R.B. Whitmarsh. 1986. The structure of the Charlie-Gibbs Fracture Zone. Journal of the Geological Society 1433: 819-821. Cormier, M.-H., Detrick, R. S. and G. M. Purdy. 1984. Anomalously thin crust in oceanic fracture zones: New seismic constraints from the Kane Fracture Zone. Journal of Geophysical Research 89:249–266. Felley, J.D., Vecchione, M. And R.R. Wilson Jr. 2008. Small-scale distribution of deep-sea demersal nekton and other megafauna in the Charlie-Gibbs Fracture Zone of the Mid-Atlantic Ridge. Deep Sea Research II 55: 153-160. Fleming, H.S., Cherkis, N.Z. and J.R. Heirtzler. 1970. The Gibbs Fracture Zone: A double fracture zone at 52°30′N in the Atlantic Ocean. Marine Geophysical Researches 1:37-45. Garner, D.M. 1972. Flow through the Charlie-Gibbs Fracture Zone, Mid-Atlantic Ridge. Canadian Journal of Earth Sciences 9: 116-121. Gebruk A.V. 2008. Holothurians (Holothuroidea, Echinodermata) of the northern Mid-Atlantic Ridge collected by the G.O. Sars MAR-ECO expedition with descriptions of four new species. Marine Biology Research 4, 48–60. Gebruk, A.V. and E.M. Krylova. 2013. Megafauna of the Charlie–Gibbs Fracture Zone (northern Mid-Atlantic Ridge) based on video observations. Journal of the Marine Biological Association of the United Kingdom 93: 1143-1150. doi:10.1017/S0025315412001890. Harvey, J.G. and A. Theodorou. 1986. The circulation of Norwegian Sea overflow water in the eastern North Atlantic. Oceanologica Acta 9: 393-402. Hekinian, R. and F. Aumento. 1973. Rocks from the Gibbs Fracture Zone and the Minia Seamount near 53°N in the Atlantic Ocean. Marine Geology 14: 47-72. ICES. 2007. Report of the Working Group on Deep-water Ecology (WGDEC), 26 – 28th February. ICES CM 2007/ACE:01 Ref. LRC. 61pp. Kanamori, H. and G.S. Stewart. 1976. Mode of the strain release along the Gibbs Fracture Zone, Mid-Atlantic Ridge. Physcis of the Earth and Planetary Interiors 11: 312-332. Kemp, K.M., Jamieson, A.J., Bagley, P.M., Collins, M.A. and I.G. Priede. 2008. A new technique for periodic bait release at a deep-sea camera platform: First results from the Charlie-Gibbs Fracture Zone, Mid-Atlantic Ridge. Deep-Sea Research II 55: 218-228. Lilwall, R.C. and R.E. Kirk. 1985. Ocean-bottom seismograph observations on the Charlie-Gibbs fracture zone. Geophysical Journal of the Royal Astronomical Society 80: 195-208. Masson, D.G. 2009. The Geobiology of Whittard Submarine Canyon. RRS James Cook Cruise 36, 19 June–28 July 2009. National Oceanography Centre, Southampton, 53 pp. http://www.eprints.soton.ac.uk/69504/1/nocscr041.pdf. Miller, R.J., Hocevar, J., Stone, R.P. and D.V. Fedorov. 2012. Structure-forming corals and sponges and their use as fish habitat in Bering Sea submarine canyons. PLoS ONE 7(3): e33885. doi:10.1371/journal.pone.0033885 ICES Advice 2013, Book 1 305 Müller, R.D. and W.R. Roest. 1992. Fracture zones in the North Atlantic from combined Geosat and Seasta data. Journal of Geophysical Research 97: 3337-3350. Mutter, J.C., Detrick, R.S. and North Atlantic Transect Study Group. 1984. Multichannel seismic evidence for anomalously thin crust at Blake Spur fracture zone. Geology 12: 534-537. Olivet, J.-L., Le Pichon, Xl, Monti, S. and B. Sichler. 1974. Charlie-Gibbs Fracture Zone. Journal of Geophysical Research 79: 2059-2072. OSPAR Commission. 2010. OSPAR Convention for the Protection of the Marine Environment of the North-East Atlantic. Meeting of the OSPAR Commission Bergen: 20-24 September 2010. Annex 49 (Ref. M6.2). http://www.ospar.org/content/content.asp?menu=01441000000000_000000_000000 OSPAR Commission. 2012. OSPAR Convention for the Protection of the Marine Environment of the North-East Atlantic. Meeting of the OSPAR Commission Bonn: 25-29 June 2012. Annex 6 (Ref. §5.19a). OSPAR Decision 2012/01. Priede, I.G., Billett, D.S.M., Brierley, A.S., Hoelzel, A.R., Inall, M., Miller, P.I., Cousins, N.J., Shields, M.A. and T. Fujii. 2013. The ecosystem of the Mid-Atlantic Ridge at the sub-polar front and Charlie-Gibbs Fracture Zone; ECO-MAR project strategy and description of the sampling programme 2007-2010. Deep-Sea Research II http://dx.doi.org/10.1016/j.dsr2.2013.06.012i. Rogacheva A, Gebruk A. and C.Alt. 2013. Deep-sea holothurians of the Charlie Gibbs Fracture Zone area, northern MidAtlantic Ridge. Marine Biology Research 9:587_623. Rossby, T. 1999. On gyre interactions. Deep-Sea Research II 46: 139-164. Saunders, P.M. 1994.The flux of overflow water through the Charlie–Gibbs Fracture Zone. Journal of Geophysical Research 99:12343–12355. Searle, R. 1981. The active part of the Charlie-Gibbs Fracture Zone: A study using sonar and other geophysical techniques. Journal of Geophysical Research 86: 243-262. Shor, A., Lonsdale, P., Hollister, C.D. and D. Spencer. 1980. Charlie-Gibbs fracture zone: bottom-water transport and its geological effects. Deep-Sea Research 27A: 325-245. Søiland, H., Budgell, W.P. and Ø Knutsen. 2008. The physical oceanographic conditions along the Mid-Atlantic Ridge north of the Azores in June-July 2004. Deep-Sea Research II 55: 29- 44. Vinogradov, G.M. 2005. Vertical distribution of macroplankton at the Charlie-Gibbs Fracture Zone (North Atlantic), as observed from the manned submersible “Mir-1”. Marine Biology 146: 325-331. 306 ICES Advice 2013, Book 1 Tables, Maps and Figures Table 1. Boundaries for the proposed EBSA Charlie-Gibbs Fracture Zone and location of the Minia and Hecate Seamounts (see Figure 3). Feature CGFZ Proposed EBSA Point 1 CGFZ Proposed EBSA Point 2 CGFZ Proposed EBSA Point 3 CGFZ Proposed EBSA Point 4 CGFZ Proposed EBSA Point 5 Northern Fracture (eastern portion) Southern Fracture (eastern portion) Northern Fracture (western portion) Southern Fracture (western portion) Minia Seamount Hecate Seamount ICES Advice 2013, Book 1 Latitude (dd) Longitude (dd) 51°N 42°W 53°05′N 42°W 53°05′N 30°W 52°30′N 25°W 51°45′N 25°W 53°05′N 42°W 51°N 45°W 52°30′N 27°W 51°45′N 25°W 53°01′N 34°58′W 53º 00.60' N 34º 49.80' W 52°17′N 31°00′W 52º 15.60' N 31º 03.00' W Details Location of Reference Olivet 8) Olivet 8) Olivet 11) Olivet 11) located near the junction of the Reykjanes Ridge and the northern transform fault et al. 1974 (p. 2062, fig. et al. 1974 (p. 2062, fig. et al. 1974 (p. 2063, fig. et al. 1974 (p. 2064, fig. Fleming et al. 1970 Seamount Catalogue http://earthref.org/SC/SMNT530N-0348W/ located on the northern wall of the southern transform fault east of the short median transverse ridge Fleming et al. 1970 Seamount Catalogue http://earthref.org/SC/SMNT530N-0348W/ 307 Figure 1 308 Location of the Charlie-Gibbs Fracture Zone (black lines) in the North Atlantic. The Mid-Atlantic Ridge runs through the centre of the Atlantic Ocean and its left lateral displacement can be clearly seen. Image downloaded from: commons.wikimedia.org File:Charlie-gibbs-full-extent.png - Wikimedia Commons. ICES Advice 2013, Book 1 Figure 2 Schematic of the Charlie-Gibbs Fracture Zone and the Mid-Atlantic Ridge (MAR) indicating the left lateral displacement of the MAR, the North and South transform faults and the central spreading axis. The relative location of two seamounts, Hecate and Minia are illustrated. Image downloaded from: commons.wikimedia.org File:Charliegibbsschema-en.svg- Wikimedia Commons. Figure 3. Proposed Charlie-Gibbs Fracture Zone EBSA (yellow lines). Numbers refer to the points in Table 1. The green line is the NEAFC/OSPAR outer boundary. ICES Advice 2013, Book 1 309 Figure 4. 310 Location of the OSPAR MPAs in the North Atlantic including the large Charlie-Gibbs South and CharlieGibbs North MPAs in the central area. The areas closed to bottom fishing by NEAFC are indicated by the yellow boundaries. Downloaded 10 Sep 2013 from: http://charlie-gibbs.org/charlie/node/70 ICES Advice 2013, Book 1 ANNEX 4 Draft Proforma - The Arctic Ice habitat - multiyear ice, seasonal ice and- marginal ice zone Presented by WWF and reviewed by Participants at the Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas in the North-East Atlantic. Reviewed and revised by an ICES expert group. Abstract The permanently ice covered waters of the high Arctic provide a range of globally unique habitats associated with the variety of ice conditions. In the northern hemisphere multi-year sea ice only exists in the Arctic and although the projections of changing ice conditions due to climate change project a considerable loss of sea ice, in particular multiyear ice, the Eurasian Central Arctic high seas are likely to at least keep the ice longer than many other regions in the Arctic basin. Ice is a crucial habitat and source of particular food web dynamics, the loss of which will affect also a number of mammalian and avian predatory species. The particularly pronounced physical changes of Arctic ice conditions as already observed and expected for the coming decades, will require careful ecological monitoring. Eventually measures will be needed to maintain or restore, to the extent possible the resilience of the Arctic populations to changing environmental conditions. Introduction Over many past millennia, most of the Eurasian part of the Arctic Basin, and in particular the high seas area in the Arctic Ocean (the waters beyond the 200 nm zones of coastal states, i.e. Norway, Russia, USA, Canada and Greenland/Denmark) have been permanently ice covered. However, in recent years, much of the multiyear permanent pack ice has been replaced by seasonal (1 year) ice. In addition, the former fast pack-ice is now increasingly broken up by leads. This structural change in the Arctic ice quality will result in a substantial increase in light penetrating the thin ice and water column, in conjunction with the overall warming of surface waters and increased temperature and salinity stratification due to the melting of ice. Some models predict that before the end of the century the permanent ice cover may disappear completely (Anisimov et al., 2007). The reduction and possible loss of permanent ice cover will result in significant changes in the structure and dynamics of the high Arctic ecosystems (CAFF, 2010; Gradinger, 1995; Piepenburg, 2005; Renaud et al., 2008; Wassmann, 2008, 2011). Understanding and studying the area proposed as EBSA is of particular scientific relevance as already envisaged by the Arctic Council (Gill et al., 2011; Mauritzen et al., 2011). Such studies may in the long-term, become relevant for the commercial exploitation of resources. Location The Ecologically or Biologically Significant Marine Area (EBSA) proposed focuses on the presently permanently icecovered waters in the OSPAR/NEAFC maritime areas, including the high seas section in the Central Arctic Basin north of the 200 nm zones of coastal states, and the area of contiguous seasonal ice directly connected to the multi-year permanent ice (see Fig. 1 attached). Therefore, the boundaries proposed extend from the North Pole (northernmost point of OSPAR/NEAFC maritime areas) to the southern limit of the summer sea ice extent and marginal ice zone, including on the shelf of East Greenland. The proposal currently only relates to features of the water column and the ice surface itself. Two legal states have to be distinguished: the Central Arctic high seas waters north of the 200 nm zones of adjacent coastal states, generally north of 84° N, and the waters within the Exclusive Economic Zones of Greenland, Russia and the fisheries protection zone of Norway around Svalbard. Figure 1 distinguishes between the high seas beyond national jurisdiction for which the ”Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic“ had a mandate23 and national/nationally administered waters within the 200 nm zone (national EEZs), within which the OSPAR Contracting Parties have the competence to report candidate EBSAs to the Convention on Biodiversity EBSA repository (OSPAR Commission, 2011). Participant Briefing for a Joint OSPAR/NEAFC/CBD Scientific Workshop on the Identification of Ecologically or Biologically Significant Marine Areas (EBSAs) in the North-East Atlantic. Invitation Annex 2, 2011 23 ICES Advice 2013, Book 1 311 The seafloor of the respective region will likely fall on the extended continental shelves of several coastal states. It belongs to the ”Arctic Basin“ region of (Gill et al., 2011). Seafloor features were not considered in this assessment. Figure 1 shows the location of the Arctic Ice „Ecologically or Biologically Significant Area“ (EBSA) proposed. Feature description The Ecologically or Biologically Significant Marine Area (EBSA) proposed focusses on the presently permanently icecovered waters in the OSPAR/NEAFC maritime areas, including the high seas section in the Central Arctic Basin north of the 200 nm zones of coastal states, and the marginal ice zone (where the ice breaks up, also called seasonal ice zone) along its southern margins (see Fig. 1 attached). Due to the inflow of Atlantic water along the shelf of Svalbard, and the concurrent outflow of polar water and ice on the Greenland side of Fram Strait, the southern limit of the summer sea ice extent is much further south in the western compared to the eastern Fram Strait, and in former times extended all along the Greenland coast. Several of the key ecosystem functions and species dependencies are associated with the ice front. Statements made about those functions and dependencies apply to the area where the front is located at any particular time of the year, and not necessarily to either areas fully ice covered (permanently or seasonally) or open waters distant from the ice front in summer. The high seas section of the OSPAR maritime area in the Central Arctic ocean is generally north of 84° N and much is fully ice-covered also in summer, although the quantity of multiyear ice has already substantially decreased and the 1year ice leaves increasingly large leads and open water spaces. The ice overlays a very deep water body of up to 5000 m depth distinct from the surrounding continental shelves and slopes of Greenland and the Svalbard archipelago. The Nansen-Gakkel Ridge, a prolongation of the Mid-Atlantic Ridge north of the Fram Strait is structuring the deep Arctic basin in this section, separating the Central Nansen Basin to the south from the Amundsen Basin to the north. Abundant hydrothermal vent sites have been discovered on this ridge at about 85° 38 N (Edmonds et al., 2003). North of Spitsbergen, the Atlantic water of the West Spitsbergen Current enters the Arctic basin as a surface current. At around 83° N, a deep-reaching frontal zone separates the incoming Atlantic and shelf waters from those of the Central Nansen Basin (Anderson et al., 1989), a transition reflected in ice properties, nutrient concentrations, zooplankton communities, and benthic assemblages (Hirche and Mumm, 1992, and literature quoted). This water subsequently submerges under the less dense (less salinity, lower temperature) polar water and circulates, in opposite direction to the surface waters and ice, counterclockwise along the continental rises until turning south along the Lomonossov Ridge and through Fram Strait as East Greenland Current south to Danmark Strait (Aagaard, 1989; Aagaard et al., 1985). Connecting the more fertile shelves with the deep central basin, these modified Atlantic waters supply the waters north of the NansenGakkel Ridge, in the Amundsen basin, with advected organic material and nutrients which supplement the autochtonous production (Mumm et al., 1998). Due to the import of organic biomass from the Greenland Sea and the Arctic continental shelves, part of which may not be kept in the food web due to the polar conditions, the Arctic Ocean may also represent an enormous carbon sink (Hirche and Mumm, 1992). In the Fram Strait, the region between Svalbard to the east and Greenland to the west, the East Greenland Current is the main outflow of polar water and ice from the Arctic Basin (Maykut, 1985) (Aagaard and Coachman, 1968). The polar front (0° C isotherm and 34.5 isohaline at 50 m depth) extends approximately along the continental shelf of Greenland, separating the polar surface water from the Arctic (Intermediate) water and the marginal ice zone to the east (e.g. Aagaard and Coachman, 1968; Paquette et al., 1985). The ice cover is densest in polar water, its extent to the east depends on the wind conditions (compare also Angelen et al., 2011; Wadhams, 1981). The seasonal latitudinal progression of increasing and diminishing light levels, respectively, is the determining factor for the timing of the phytoplankton-related pelagic production. Therefore, the spring bloom and ice break up progress from south to north in spring, reaching the Arctic area by about June/July. Because the currents in Fram Strait move in opposite direction, the polar East Greenland Current to the south, and the Atlantic West Spitsbergen Current to the north, there is a delay of about a month between biological spring and summer between the polar and the Atlantic side (Hirche et al., 1991). Therefore, sea ice and the effect of melting ice are important determinants of the ecosystem processes all along the East Greenland polar front from the Greenland Sea through Fram Strait to the Arctic Basin (Legendre et al., 1992; Wassmann, 2011). Ice situation The Arctic Ocean is changing towards a one-year instead of a multi-year sea-ice system with consequences for the entire ecosystem, including ecosystem shifts, biodiversity changes, water mass modifications, and role in the global overturning circulation. At its maximum, sea-ice covers 4.47 million km² in the Arctic Basin (Gill et al., 2011): According to data 312 ICES Advice 2013, Book 1 from ice satellite observations in 1973-76 (NASA, 1987, in (Gill et al., 2011)), permanent ice occupied 70-80% of the Arctic Basin area, and the inter-annual variability of this area did not exceed 2%. Seasonal ice occupied 6-17% (before the melting period of the mid-1970s). By the end of the first decade of the 21st century, the permanent-ice area had decreased greatly, concurrent with a rapid increase in seasonal- ice. Whereas multiyear ice used to cover 50-60% of the Arctic, it covered less than 30% in 2008, after a minimum of 10% in 2007. The average age of the remaining multiyear ice is also decreasing from over 20 % being at least six years in the mid- to late 1980s, to just 6% of ice six years old or older in 2008. Figure 2: Modelled ice age distribution in 1985-2000 (left) compared to February 2008 (right) (CAFF, 2010). This trend is likely to amplify in the coming years, as the net ocean-atmosphere heat output due to the current anomalously low sea ice coverage has approximately tripled compared to previous years, suggesting that the present sea ice losses have already initiated a positive feedback loop with increasing surface air temperatures in the Arctic (Kurtz et al., 2011). About 10% of the sea ice in the Arctic basin is exported each year through Fram Strait into the Greenland Sea (Maykut, 1985) which is therefore major sink for Arctic sea ice (Kwok, 2009). From 2001 to 2005, the summer ice cover was so low on the East Greenland shelf, that it was more of a marginal ice zone (Smith Jr and Barber, 2007). However the subsequent record lows in overall Arctic ice cover brought about an increase in ice cover off Greenland, which minimised the extent of the North East Water Polynia on the East Greenland shelf24, a previously seasonally ice-free stretch of water (Wadhams, 1981). Ice related biota An inventory of ice-associated biota covering the entire Arctic presently counts over 1000 protists, and more than 50 metazoan species (Bluhm et al., 2011). The regionally variable ice fauna (dependent on, inter alia, ice age, thickness, origin) consists of sympagic biota living within the caverns and brine channels of the ice, and associated pelagic fauna. The most abundant and diverse sympagic groups of the ice mesofauna in the Arctic seas are amphipods and copepods. Polar cod (Boreogadus saida) and to a lesser extent Arctic cod (Arctogadus glacialis) are dependent on the sympagic macro- and mesofauna for food. The fish themselves are important food sources for Arctic seals (such as ringed seal Phoca hispida) and birds, for example black guillemots Cephus grylle (Bradstreet and Cross, 1982; Gradinger and Bluhm, 2004 and literature reviewed; Horner et al., 1992; Süfke et al., 1998). The higher the light level in the ice, the higher is the biomass of benthic algae as well as meiofauna and microorganisms within the ice (Gradinger et al., 1991). Decreasing snow cover induces a feedback loop with enhanced algal biomass increasing the heat absorption of the ice which leads to changes in the ice structure, and ultimately the release of algae from the bottom layer (Apollonio, 1961 in Gradinger et al., 1991). Because of the distance to land and shelves, and the thickness and internal structure of the multiyear pack ice over deeper water, this type of ice has a fauna of its own (Carey, 1985; Gradinger et al., 1991). Arctic multiyear ice floes can have very high algal biomasses in the brine channels and in the bottom centimeters which serves as food for a variety of proto- and metazoans, usually smaller than 1 mm, over deep water (Gradinger et al., 1999). In the central Arctic, ice algal productivity can contribute up to 50 % of the total primary productivity, with lower contributions in the sea ice covered margins (Bluhm et al., 2011). In the boundary layer between ice floes and the water column, another specific community exists which forms the link between the ice based primary production and the pelagic fauna (Gradinger, 1995). Large visible bands of diatoms hang down from the ice, and are exploited by amphipods such as Gammarus wilkitzki, and occasionally by water column copepods such as Calanus glacialis, which are important prey of for example polar cod Boreogadus saida. The caverns, wedges and irregularities of the ice provide important shelter from predators for larger ice associated species and provide an essential habitat for these species (Gradinger and Bluhm, 2004). During melt, the entire sympagic ice biota are released into the water column where they may initiate the spring algal plankton bloom (Smith and Sakshaug, 1990) or they may sink to the sea floor and serve as an episodic and first food pulse for benthic organisms before pelagic production begins (Arndt and Pavlova, 2005). In particular the shallow shelves and the shelf slope benthos has been shown to profit of this biomass input, reflected in very rich benthic communities (Klitgaard and Tendal, 2004; Piepenburg, 2005). The role of the polar front and marginal ice zone for the production system Primary production in the Arctic Ocean is primarily determined by light availability, which is a function of ice thickness, ice cover, snow cover, light attenuation), the abundance of both ice algae and phytoplankton, nutrient availability and surface water stratification. Generally, the spring bloom occurs later further north and in regions with a thick ice and snow 24 http://www.issibern.ch/teams/Polynya/ ICES Advice 2013, Book 1 313 cover. The current production period in the Arctic Ocean may extend to 120 days per year, with a total annual primary production in the central Arctic Ocean of probably up to 10 g C m-2 (Wheeler et al., 1996). Ice algae start primary production when light levels are relatively low, as melting reduces the thickness of the ice and snow cover. The major phytoplankton bloom develops only after the ice breaks up, when melting releases the ice biota into the water column and meltwater leads to surface stratification. The bloom lasts a few weeks, fuelling the higher trophic food web of the Arctic (Gradinger et al., 1999, and literature quoted). The marginal ice zones, i.e. where the ice gets broken up in warmer Atlantic or Arctic water, play an important role in the overall production patterns of the Arctic Ocean. Due to the strong water column stratification and increased light levels involved with the melting of the ice, the location and recession of the ice edge in spring and summer determines the timing and magnitude of the spring phytoplankton bloom, which is generally earlier than in the open water (Gradinger and Baumann, 1991; Smith Jr. et al., 1987). Wind- or eddy-induced upwelling in the marginal ice zone, as well as biological regeneration processes replenish the surface nutrient pool and therefore prolong the algal growth period (Gradinger and Baumann, 1991; Smith, 1987). The hydrographic variability explains the patchy patterns of primary and secondary production observed, as well as consequently the patchy occurrence of predators. The polar front separates to some degree the pelagic faunas of the polar and Arctic waters in the Greenland Sea and Fram Strait, each characterised by a few dominant copepod species with different life history strategies (Hirche et al., 1991; see also review in Melle et al., 2005): In polar waters, Calanus glacialis uses under ice plankton production and lipid reserves for initiating its spring reproduction phase, but depends on the phytoplankton bloom for raising its offspring (e.g. Leu et al., 2011). Somewhat later, on the warm side of the polar front in Arctic water, the Atlantic species Calanus finmarchicus uses the ice edge-related phytoplankton bloom for secondary production. Calanus hyperboreus, the third and largest of the charismatic copepod species has its core area of distribution in the Arctic waters of the Greenland Sea (Hirche, 1997; Hirche et al., 2006). Zooplankton of the Arctic Basin Overall zooplankton biomass decreases towards the central Arctic basin, reaching a minimum in the most northerly waters, i.e. the region with permanent ice cover (Mumm et al., 1998). However, investigations in recent years demonstrated increased biomasses compared to studies several decades earlier - possibly a consequence of the decrease in ice thickness and cover which only enabled the investigations to take place from ship board. There is a south-north decrease in zooplankton biomass, with a sharp decline north of 83°N (Hirche and Mumm, 1992), coinciding with differences in the species composition of the biomass-forming zooplankton species. Whereas the southern Nansen basin plankton is dominated by the Atlantic species Calanus finmarchicus, entering the Arctic Basin with the West Spitsbergen Current, the northernmost branch of the North Atlantic current, the Arctic and polar species Calanus hyperboreus and C. glacialis dominate the biomass in the high-Arctic Amundsen and Makarov Basins (Auel and Hagen, 2002; Mumm et al., 1998). The zooplankton species communities generally can be differentiated according to their occurrence in Polar Surface Water (0-50 m, temperature below –1.7°C, salinity less than 33.0), Atlantic Layer (200–900 m; temperature 0.5–1.5°C); salinity 34.5–34.8) and Arctic Deep Water (deeper than 1000 m, temperature -0,5--1° C, salinity > 34.9) (Auel and Hagen, 2002; Grainger, 1989; Kosobokova, 1982). The polar surface community in the upper 50 m of the water column consists of original polar species as well as species emerging from deeper Atlantic waters, altogether leading to a high abundance and biomass peak in summer. Diversity and biomass are minimal in the impoverished Arctic basin deepwater community (Kosobokova 1982). Apart from a limited exchange with the Atlantic Ocean via the Fram Strait, the central Arctic deep-sea basins are isolated from the rest of the world ocean deepsea fauna. Therefore, the bathypelagic fauna consists of a few endemic Arctic species and some species of Atlantic origin. Due to the separation of the Eurasian and Canadian Basins by the Lomonosov Ridge, significant differences in hydrographic parameters (Anderson et al. 1994) and in the zooplankton composition occur between both basins (Auel and Hagen, 2002). Fish Polar cod, Boreogadus saida, is a keystone species in the ice-related foodwebs of the Arctic. Due to schooling behavior and high energy content polar cod efficiently transfer the energy from lower to higher trophic levels, such as seabirds, seals and some whales (Crawford and Jorgenson, 1993). Seabirds Ice cover is a physical feature of major importance to marine birds in high latitude oceans, providing access to resources, and refuge from aquatic predators (Hunt, 1990). As seabirds are dependent on leads between ice floes or otherwise open water to access food, they search for the most productive waters in polynias (places within the ice which are permanently 314 ICES Advice 2013, Book 1 ice free) and marginal ice zones (Hunt, 1990). Here they forage both on the pelagic and sympagic ice-related fauna, especially the early stages of polar cod and the copepods Calanus hyperboreus and C. glacialis. Likely, they benefit from the structural complexity and good visibility of their prey near the ice (Hunt, 1990). In the Greenland Sea and Fram Strait, major breeding colonies exist on Svalbard, Greenland and on Jan Mayen, all of these within reach of the seasonally moving marginal ice zone or a polynia (North East Water Polynia on the East Greenland shelf). Breeding seabirds like Little auks (Alle alle), from colonies in the northern Svalbard archipelago feed their offspring with prey caught in the vicinity of the nests, however intermittently travel at least 100 km to the marginal ice zone at 80° N to replenish their body reserves (Jakubas et al., Online 03 June 2011). Therefore, the distance of the marginal ice zone to the colony site is a critical factor determining the breeding success (e.g. Joiris and Falck, 2011). Opportunistically, the birds also use other zooplankton aggregations such as a in a cold core eddy in the Greenland Sea, closer to the nesting site (Joiris and Falck, 2011). A synopsis of seabird data for the period 1974–1993 (Joiris, 2000) showed that the little auk is one of the most abundant species, together with the fulmar Fulmarus glacialis, kittiwake Rissa tridactyla and Brünnich’s guillemot Uria lomvia in the European Arctic seas (mainly the Norwegian and Greenland Seas). In the Greenland Sea and the Fram Strait, little auks represented the main species in polar waters, at the ice edge and in closed pack ice, reaching more than 50% of all bird species (Joiris and Falck, 2011). In spring and autumn, millions of seabirds pass through the area when migrating between their breeding sites on Svalbard or the Russian Arctic and their wintering areas in Canada (Gill et al., 2011). There are several seabird species in the European Arctic which are only met in ice-covered areas, for example the Ivory gull Pagophila eburnea and the Thick-billed guillemot Uria lomvia (see e.g. CAFF, 2010): Both species spend the entire year in the Arctic, and breed in close proximity to sea ice although Thick-billed guillemots were observed to fly up to 100 km from their colonies over open water to forage at the ice edge (Bradstreet 1979). The relatively rare Ivory gulls are closely associated with pack-ice, favouring areas with 70 – 90% ice cover near the ice edge, where they feed on small fish, including juvenile Arctic cod, squid, invertebrates, macro-zooplankton, carrion, offal and animal faeces (e.g. OSPAR Commission, 2009b). Ivory gulls have a low reproductive rate and breeding only takes place if there is sufficient food, which makes the population highly vulnerable to the effects of climate warming (e.g. OSPAR Commission, 2009b). Thick-billed guillemots are relatively long lived and slow to reproduce and has a low resilience to threats including oil pollution, by-catch in and competition with commercial fisheries operations, population declines due to hunting – particularly in Greenland (OSPAR Commission, 2009c). Ivory gull and Thick-billed guillemots are both listed by OSPAR as being under threat and/or decline, (OSPAR Commission, 2008) and in 2011 recommendations for conservation action were agreed (OSPAR Commission, 2011) which will be implemented in conjunction with the circumpolar conservation actions of CAFF (CAFF, 1996; Gilchrist et al., 2008). Marine mammals Several marine mammal species permanently associate with sea ice in the European Arctic. These include polar bear, walrus, and several seal species: bearded, Erignathus barbatus; ringed, Pusa hispida; hooded, Cystophora cristata; and harp seal Pagophilus groenlandicus. Three whale species also occupy Arctic waters year- round – narwhal, Monodon monoceros; beluga whale, Delphinapterus leucas; and bowhead whale, Balaena mysticetus. Polar bears Ursus maritimus are highly specialized for and dependent on the sea ice habitat and are therefore particularly vulnerable to changes in sea ice extent, duration and thickness. They have a circumpolar distribution limited by the southern extent of sea ice. Three subpopulations of polar bears occur in the European high Arctic: the East Greenland, Barents Sea and Arctic Basin sub-populations, all with an unknown population status (CAFF, 2010). Following the youngof-the-year ringed seal distribution, polar bears are most common close to land and over the shelves, however some also occur in the permanent multi-year pack ice of the central Arctic basin (Durner et al., 2009). Due to low reproductive rates and long lifetime, it has been predicted that the polar bears will not be able to adapt to the current fast warming of the Arctic and become extirpated from most of their range within the next 100 years (Schliebe et al., 2008). Walruses, Odobenus rosmarus, inhabit the Arctic ice year-round. They are conservative benthic feeders, diving to 80100 m depth for scaping off the rich mollusc fauna of the continental shelves, and need ice floes as resting and nursing platform close to their foraging grounds. Walruses have been subject to severe hunting pressure from the end of the 18th century to the mid-20th century, and are still hunted today in Greenland (NAMMCO). By 1934, the estimated 7000080000 individuals of the Atlantic population were reduced to 1200-1300, with none left on Svalbard (Weslawski et al., 2000). Today’s relatively small sub-populations on the East Greenland and Svalbard-Franz Josef Land coasts have recently shown a slightly increasing trend, in the latter case reflecting the full protection of the species since the 1950´s (CAFF, 2010; NAMMCO). Apart from their sensitivity to direct human disturbance and pollution, it is expected that walruses will suffer from the changing ice conditions (location, thickness for being used as haul-out site) as well as changes in ice-related productivity. ICES Advice 2013, Book 1 315 The Atlantic subspecies of the bearded seal, Erignathus barbatus occurs south of 85° N from the central Canadian Arctic east to the central Eurasian Arctic, but no population estimates exist (Kovacs, 2008b). Because of their primarily benthic feeding habits they live in ice covered waters overlying the continental shelf. They are typically found in regions of broken free-floating pack ice; in these areas bearded seals prefer to use small and medium sized floes, where they haul out no more than a body length from water and they use leads within shore-fast ice only if suitable pack ice is not available (Kovacs, 2008b, and literature quoted). The Arctic ringed seal Pusa (Phoca) hispida hispida has a very large population size and broad distribution, however, there are concerns that future changes of Arctic sea ice will have a negative impact on the population, some of which have already been documented in some parts of the subspecies range (Kovacs et al., 2008). As the other seals, the ringed seal uses sea ice exclusively as their breeding, moulting and resting (haulout) habitat, and feed on small schooling fish and invertebrates. In a co-evolution with one of their main predators, the polar bear, they developed the ability to create and maintain breathing holes in relatively thick ice, which makes them well adapted to living in fully ice covered waters the year round. The West Ice (or Is Odden) to the west of Jan Mayen, at approx. 72-73° N, in early spring a stretch of more of less fast drift ice, is of crucial importance as a whelping and moulting area for harp seals and hooded seals (summarised e.g. by ICES, 2008). Discovered in the early 18th century, up to 350000 seals (1920s) were killed per year, decimating the populations from an estimated one million individuals in the 1950s (Ronald et al., 1982) to today´s 70000 and 243000 of hooded and harp seals, respectively (Kovacs, 2008a, c). Hooded seal, Cystophora cristata, is a pack ice species, which is dependent on ice as a substrate for pupping, moulting, and resting and as such is vulnerable to reduction in extent or timing of pack ice formation and retreat, as well as ice edge related changes in productivity (Kovacs, 2008a, and literature quoted). Hooded Seals feed on a wide variety of fish and invertebrates, including species that occur throughout the water column. After breeding and moulting on the West Ice they follow the retreating pack ice to the north, but also spend significant periods of time pelagically, without hauling out (Folkow and Blix 1999) in (Kovacs, 2008a). The northeast Atlantic breeding stock has declined by 85-90 % over the last 40-60 years. The cause of the decline is unknown, but very recent data suggests that it is on-going (30% within 8 years), despite the protective measures that have been taken in the last few years. The species is therefore considered to be vulnerable (Kovacs, 2008a). Harp seals Pagophilus (Phoca) groenlandicus are the most numerous seal species in the Arctic seas. Their reproduction takes place in huge colonies, for example on the pack ice of the ‘‘West Ice’’ north of Jan Mayen, and after the breeding season they follow the retreating pack ice edge northwards up to 85° N, feeding mainly on polar cod under the ice (Kovacs, 2008c) . Narwhals Monodon monoceros primarily inhabit the ice-covered waters of the European Arctic, including the ice sheet off East Greenland (Jefferson et al., 2008b). For two months in summer, they visit the shallow fjords of East Greenland, spending all the rest of the year offshore, in deep ice-covered waters along the continental slope in the Greenland Sea and Arctic Basin (Heide-Jørgensen and Dietz, 1995). Narwhals are deep diving benthic feeders and forage on fish, squid, and shrimp, especially Arctic fish species, such as Greenland halibut, Arctic cod, and polar cod at up to 1500 m depth and mostly in winter. A recent assessment of the sensitivity of all Arctic marine mammals to climate change ranked the narwhal as one of the three most sensitive species, primarily due to its narrow geographic distribution, specialized feeding and habitat choice, and high site fidelity (Laidre et al. 2008 in (Jefferson et al., 2008b)). Bowhead whales Balaena mysticetus are found only in Arctic and subarctic regions and a Svalbard-Barents population occurs from the coast of Greenland across the Greenland Sea to the Russian Arctic. They spend all of their lives in and near openings in the pack ice feeding on small to medium-sized zooplankton. They migrate to the high Arctic in summer, and retreat southward in winter with the advancing ice edge (Moore and Reeves 1993 in (Reilly et al., 2008)). Whaling has decimated the original bowhead whale populations to be rare nowadays, listed by OSPAR as being under threat and/or decline (OSPAR Commission, 2008). The species is considered to be very sensitive to changes in the ice-related ecosystem as well as sound disturbance, possible consequences of a progressive reduction of ice cover (OSPAR Commission, 2009a). Belugas Delphinapterus leucas prefer coastal and continental shelf waters with a broken-up ice cover. They have never been surveyed around Svalbard. Pods numbering into the thousands are sighted irregularly around the archipelago, and pods ranging from a few to a few hundred individuals are seen regularly (Gjertz and Wiig 1994; Kovacs and Lydersen 2006 in (Jefferson et al., 2008a)). Little is known about the populations of the larger fauna in the Central Arctic Basin over the deepsea basins and ridges. But it is not likely that it is currently an area of great abundance - too far from the coastal nesting sites of marine birds, 316 ICES Advice 2013, Book 1 and over too deep water to allow feeding on benthos, as most of the larger mammals would need, and currently of too low plankton production to feed the large whales. All of these groups have their distribution center along the continental shelves presently - however, following the receding ice edge out to the central Arctic basin may be one of the options for the future. Feature condition, and future outlook This high Arctic region is particularly vulnerable to the the loss of ice cover and other effects of the anticipated global warming, including elevated UV radiation levels (Agustí, 2008). (Wassmann et al., 2010) summarise what changes may be expected within the subarctic/Arctic region: • • • • • northward displacement (range shifts) of subarctic and temperate species, and cross-Arctic transport of organisms; increased abundance and reproductive output of subarctic species, decline and reduced reproductive success of some Arctic species associated with the ice and species now preyed upon by predators whose preferred prey have declined; increased growth of some subarctic species and primary producers, and reduced growth and condition of animals that are bound to, associated with, or born on the ice; anomalous behaviour of ice-bound, ice-associated, or ice-born animals with earlier spring events and delayed fall events; changes in community structure due to range shifts of predators resulting in changes in the predator–prey linkages in the trophic network. (Wassmann, 2008) expects radical changes in the productivity, functional relationships and biodiversity of the Arctic Ocean. He suggests that a warmer climate with less ice cover will result in greater primary production, a reduction of the stratified water masses to the south, changes in the relationship between biological processes in the water column and the sediments, a reduction in niches for higher trophic levels and a displacement of Arctic by boreal species. On the shelves, increased sediment discharges are expected to lower the primary production due to higher turbidity, and enhance the organic input to the deep ocean. A more extensive review of expected or suspected consequences of climate change for the marine system of the Arctic is given in (Loeng et al., 2005). Figure 3, extracted from (Gill et al., 2011), presents the conceptual ideas about possible Arctic ecosystem changes mediated by human impact: The normal situation shown in the upper left panel consists of ice-dependent species and species that tolerate a broader range of temperatures and are found in waters with little or no sea ice. Primary production occurs in phytoplankton (small dots in the figure) in ice-free waters and in ice-attached algae and phytoplankton in ice-covered waters. Phytoplankton (small t-shaped symbols in the figure) and ice algae are the main food sources for zooplankton and benthic animals. The fish community consists of both pelagic and demersal species. Several mammals are ice-associated, including polar bears and several species of seals. A number of sea bird species are also primarily associated with ice-covered waters. At moderate temperature increases (upper right) populations of ice-dependent species are expected to decline as sea ice declines, and sub-Arctic species are expected to move northwards. Arctic benthic species are expected to decline, especially if their distributions are pushed close to or beyond the continental slope. The expected effects from fisheries relate to the continental shelves. Two major effects are changes in populations of benthic organisms due to disturbance from bottom trawling and removal of large individuals in targeted fish stocks. In addition, the size of targeted stocks, both demersal and pelagic, may be reduced. In addition, the effects of ocean acidification are considered (lower right). Ocean acidification will result in depletion of carbonate phases such as aragonite and calcite. This will alter the structure and function of calcareous organisms, particularly at lower trophic levels. Changes in pH can also alter metabolic processes in a range of organisms. It is not known how these changes will propagate to higher trophic levels, but the effects could be substantial. Figure 3: Conceptual models showing potential impacts on Arctic marine ecosystems under different scenarios (Gill et al., 2011). Gill et al. (2011) conclude that the central part of the Arctic Basin is not a region for fisheries or oil and gas exploration. However, this region has played and will continue to play a very important role in the redistribution of pollutants, due to ice drift and/or currents between coastal and shelf areas and the Arctic Basin peripheries, far from sources of pollution. ICES Advice 2013, Book 1 317 Assessment against CBD EBSA Criteria Table 1 CBD Criterion Relation of each of the CBD criteria to the proposed area relating to the best available science. Note that a candidate EBSA may qualify on the basis of one or more of the criteria, the boundaries of the EBSA need not be defined with exact precision. EBSA Description Uniqueness or rarity The area contains either (i) unique (“the only one of its kind”), rare (occurs only in few locations) or endemic species, populations or communities, and/or (ii) unique, rare or distinct, habitats or ecosystems; and/or (iii) unique or unusual geomorphological or oceanographic features Ranking of criterion relevance (please mark one column with an X) Don’t Know Low Some High x Explanation for ranking Arctic sea ice, in particular the multiyear ice of the Central Arctic is globally unique and hosts endemic species such as the Gammarid amphipod Gammarus wilkitzki and sea ice meiofauna which will disappear with the melting of the ice. Polar bears, walrusses, bowhead whales, narwhales, belugas, several seal species and many bird species are endemic to the high Arctic ice. While sea ice species such as G. wilkitzki are not endemic to the proposed EBSA they are endemic to the Arctic and unique within the OSPAR area Special importance Areas that are required for a population to survive and thrive for life-history stages of species x Explanation for ranking Sea ice is essential for its sympagic fauna, and to some extent also for the pelagic associated fauna which also depends on the right timing of biomass production (match/mismatch with bloom periods). The marginal ice zone and other openings in the ice are essential feeding grounds for a large number of ice-associated species which exploit the seasonally high production there. At present the area covered by the proposed EBSA includes both the area of permanent ice and, the area covered by seasonal ice and the ice edge. The community associated with the ice edge requires it special structural features for a number of ecological processes, including increased primary and secondary productivity, and feeding and resting of seabirds and marine mammals. Area containing habitat for the survival and recovery of x Importance for endangered, threatened, declining species or area with threatened, significant assemblages of such species endangered or declining species and/or habitats Explanation for ranking The high arctic ice hosts endemic species such as the Gammarid amphipod Gammarus wilkitzki and sea ice meiofauna which will disappear with the melting of the ice. Many of the obligatory ice-related species are listed as vulnerable by IUCN, and/or listed as under threat and/or decline by OSPAR, examples include the Ivory gull, thick-billed guillemot, bowhead whale, hooded seal and polar bear. With the overall trend of retreating sea ice extent, the proposed EBSA may become increasingly important for all ice-dependent species in the future. Areas that contain a relatively high proportion of sensitive Vulnerability, habitats, biotopes or species that are functionally fragile fragility, sensitivity, or slow (highly susceptible to degradation or depletion by human activity or by natural events) or with slow recovery recovery 318 x ICES Advice 2013, Book 1 Explanation for ranking The ice-related foodweb and ecosystem is highly sensitive to the ecological consequences of a warming climate. Beyond this the Arctic is at the forefront of the impacts of ocean acidification (Wicks & Roberts 2012). The largest changes in ocean pH will occur in the Arctic Ocean, with complete undersaturation of the Arctic Ocean water column predicted before the end of this century (Steinacher et al. 2009). Many of the seabird and mammal populations are particularly sensitive to changes due to their already low population numbers, and low fertility. If the retreat of the ice to the north will lead to increased shipping and oil and gas exploitation in Arctic waters, the increased risk of spills would also pose a potential hazard for example for guillemots, which are extremely susceptible to mortality from oil pollution (CAFF, 2010). In addition, some species like Ivory gull are sensitive to an increased heavy metal load in their prey. Biological productivity Area containing species, populations or communities with comparatively higher natural biological productivity Explanation for ranking This criterion was not evaluated in the OSPAR/NEAFC/CBD Workshop. ICES did not have enough information to evaluate this criterion. Biological diversity Area contains comparatively higher diversity of ecosystems, habitats, communities, or species, or has higher genetic diversity Explanation for ranking This criterion was not evaluated in the OSPAR/NEAFC/CBD Workshop. ICES did not have enough information to evaluate this criterion. References Aagaard, K., 1989. A synthesis of Arctic Ocean circulation. Rapport Proces et Verbeaux Réunion du Conseil international pour l'Exploration de la Mer 188, 11-22. Aagaard, K., Coachman, L.K., 1968. The East Greenland Current north of Denmark Strait: Part II. Arctic 21, 267-290. Aagaard, K., Swift, J.H., Carmack, E.C., 1985. Thermohaline circulation in the Arctic mediterranean seas. Journal of Geophysical Research 90, 4833-4846. Agustí, S., 2008. Impacts of increasing ultraviolet radiation on the polar oceans. In: Impacts of global warming on polar ecosystems. Duarte, C.M. (Ed.) Fundación BBVA pp. 25-46. Anderson, L.G., Jones, E.P., Koltermann, K.P., Schlosser, P., Swift, J.H., Wallace, D.W.R., 1989. The first oceanographic section across the Nansen Basin in the Arctic Ocean. Deep Sea Research 36, 475-482. Angelen, J.H.v., Broeke, M.R.v.d., Kwok, R., 2011. The Greenland Sea Jet: A mechanism for wind‐driven sea ice export through Fram Strait. Geophysical Research Letters 38 (L12805). Anisimov, O.A., Vaughan, D.G., Callaghan, T.V., Furgal, C., Marchant, H., Prowse, T.D., Vilhjálmsson, H., Walsh, J.E., 2007. Polar regions (Arctic and Antarctic). In: Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Parry, M.L., Canziani, O.F., Palutikof, J.P., Linden, P.J.v.d., Hanson, C.E. (Eds.)Cambridge University Press, Cambridge pp. 653-685. Arndt, C., E. , Pavlova, O., 2005. Origin and fate of ice fauna in the Fram Strait and Svalbard area. Marine Ecology Progress Series 301, 55-66. Auel, H., Hagen, W., 2002. Mesozooplankton community structure, abundance and biomass in the central Arctic Ocean. Marine Biology 140, 1013-1021. Bluhm, B.A., Gebruk, A.V., Gradinger, R., Hopcroft, R.R., Huettmann, F., Kosobokova, K.N., Sirenko, B.I., Weslawski, J.M., 2011. Arctic marine biodiversity: An update of species richness and examples of biodiversity change. Oceanography 24 (3), 232-248. Bradstreet, M.S.M., Cross, W.E., 1982. Trophic relationships at high Arctic ice edges. Arctic 35 (1), 1-12. CAFF, 1996. International Murre conservation strategy and action plan. CAFF International Secretariat, CAFF Circumpolar Seabird Working Group, Akureyri, Iceland, pp. 1-16. CAFF, 2010. Arctic Biodiversity Trends 2010. Selected indicators of change. CAFF International Secretariat, , Akureyri, Iceland. Carey, A.G.I., 1985. Marine Ice Fauna. In: Arctic Sea Ice Biota. A., H.R. (Ed.)CRC Press, Boca Raton. Florida pp. 17190 Crawford, R.E., Jorgenson, J.K., 1993. Schooling behaviour of arctic cod, Boreogadus saida in relation to drifting pack ice. Environmental Biology of Fishes 36 (4), 345-357. Durner, G.M., Douglas, D.C., Nielson, R.M., Amstrup, S.C., McDonald, T.L., Stirling, I., Mauritzen, M., Born, E.W., Wiig, Ø., Deweaver, E., Serreze, M.C., Belikov, S.E., Holland, M.M., Maslanik, J., Aars, J., Bailey, D.A., Derocher, A.E., 2009. Predicting 21st-century polar bear habitat distribution from global climate models. Ecological Monographs 79 (1), 25-58. ICES Advice 2013, Book 1 319 Edmonds, H.N., Michael, P.J., Baker, E.T., Connelly, D.P., Snow, J.E., Langmuir, C.H., Dick, H.J.B., Mühe, R., German, C.R., Graham, D.W., 2003. Discovery of abundant hydrothermal venting on the ultraslow-spreading Gakkel ridge in the Arctic Ocean. Nature 421, 252-256. Gilchrist, G., Strøm, H., Gavrilo, M.V., Mosbech, A., 2008. International Ivory Gull conservation strategy and action plan. CAFF International Secretariat, Circumpolar Seabird Group (CBird). CAFF Technical Report No. 18. Gill, M.J., Crane, K., Hindrum, R., Arneberg, P., Bysveen, I., Denisenko, N.V., Gofman, V., Grant-Friedman, A., Gudmundsson, G., Hopcroft, R.R., Iken, K., Labansen, A., Liubina, O.S., Melnikov, I.A., Moore, S.E., Reist, J.D., Sirenko, B.I., Stow, J., Ugarte, F., Vongraven, D., Watkins, J., 2011. Arctic Marine Biodiversity Monitoring Plan (CBMP-MARINE PLAN), CAFF Monitoring Series Report No.3, April 2011. CAFF International Secretariat,, Akureyri, Iceland. Gradinger, R., 1995. Climate change and biological oceanography of the Arctic Ocean. Phil. Trans. R. Soc. A 352, 277286. Gradinger, R., Bluhm, B.A., 2004. In situ observations on the distribution and behavior of amphipods and Arctic cod (Boreogadus saida) under the sea ice of the high Arctic Canadian Basin. Polar Biology 27, 595-603. Gradinger, R., Friedrich, C., Spindler, M., 1999. Abundance, biomass and composition of the sea ice biota of the Greenland Sea pack ice. Deep Sea Research 46, 1457-1472. Gradinger, R., Spindler, M., Henschel, D., 1991. Development o Arctic sea-ice organisms under graded snow cover. In: Proceedings of the Pro Mare Symposium on Polar Marine Ecology. Sakshaug, E., E., H.C.C., Øritsland, N.A. (Eds.), Polar Research 10 (1), Trondheim pp. 295-307. Gradinger, R.R., Baumann, M.E.M., 1991. Distribution of phytoplankton communities in relation to the large-scale hydrographical regime in the Fram Strait. Mar. Biol. 111, 311-321. Grainger, E.H., 1989. Vertical distribution of zooplankton in the central Arctic Ocean. In: Proc 6th Conf Comite´Arctique Int 1985. Rey, L., Alexander, V. (Eds.) Brill Leiden pp. 48–60. Heide-Jørgensen, M.P., Dietz, R., 1995. Some characteristics of narwhal, Monodon monoceros, diving behaviour in Baffin Bay. Canadian Journal of Zoology 73, 2106-2119. Hirche, H.J., 1997. Life cycle of the copepod Calanus hyperboreus in the Greenland Sea. Marine Biology 128 (4), 607618. Hirche, H.J., Baumann, M.E.M., Kattner, G., Gradinger, R., 1991. Plankton distribution and the impact of copepod grazing on primary production in Fram Strait, Greenland Sea. Journal of Marine Systems 2 (3-4), 477-494. Hirche, H.J., Mumm, N., 1992. Distribution of dominant copepods in the Nansen Basin, Arctic Ocean, in summer. Deep Sea Research Part A. Oceanographic Research Papers 39 (2, Part 1), S485-S505. Hirche, H.J., Muyakshin, S., Klages, M., Auel, H., 2006. Aggregation of the Arctic copepod Calanus hyperboreus over the ocean floor of the Greenland Sea. Deep Sea Research Part I: Oceanographic Research Papers 53 (2), 310-320. Horner, R., Ackley, S.F., Dieckmann, G.S., Gulliksen, B., Hoshiai, T., Legendre, L., Melnikov, I.A., Reeburgh, W.S., Spindler, M., Sullivan, C.W., 1992. Ecology of sea ice biota. Habitat, terminology, and methodology. Polar Biology 12 (3), 417-427. Hunt, G.L.J., 1990. The pelagic distribution of marine birds in a heterogeneous environment. Polar Research 8, 43-54. ICES, 2008. Report of the ICES Advisory Committee In: ICES Advice, Book 3, The Barents and the Norwegian SEa. Jakubas, D., Iliszko, L