Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
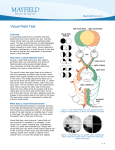
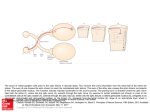
Preliminary Results of Optic Nerve Crush Damage in the 13 Lined Ground Squirrel Ryan 1 Bures , Ben 1 Sajdak , Grant 1 O’Connell , Cassandra 1 Piper , David 2,3 Henson and Dana 1 Merriman 1.UW Oshkosh Biology & Microbiology, University of Wisconsin Oshkosh, Oshkosh, WI 2. Hanover College 3. REU Student at Wisconsin Oshkosh The UW Oshkosh McNair Scholars Program is 100% funded through a TRIO grant from the United States Department of Education PR/Award Number P217A070188. For 2012/2013, the UW Oshkosh McNair Scholars Program will receive $213,000 each year in federal funds. Purpose Methods Whenever the central nervous system (CNS) of humans, monkeys, cats, rabbits, rats, or mice is damaged, neurons die and two other cell types, microglia and astrocytes, undergo activation to make emergency repairs to the tissue. Taken together, their activation is known as “reactive gliosis”. Upon insult to the CNS, microglia undergo a shift from a resting state to an activated state and proliferate. In this activated state, the microglia cells become phagocytes sequestering neuronal debris. They also release IL-2 that signals astrocytes to activate new gene expression programs involving motility and cytoskeletal reorganization. The reactive astrocytes form a glial scar that reinforces the blood-brain barrier (damaged by the initial insult) and fills the space vacated by dead neurons. Hallmarks of reactive gliosis include significant upregulation of the cytoplasmic intermediate filament proteins, vimentin and glial fibrillary acidic protein (GFAP), within astrocytes, as well as the extracellular deposition of chondroitin sulfate proteoglycans (CSPGs) in new basement membrane. These damage responses accomplish tissue repair, but inhibit the ability of surviving neurons or therapies such as stem cells to repair lost neuron circuitry. In very limited studies to date, the ground squirrel CNS shows a uniquely muted microglial and glial response to damage, with minimal reaction to neuron death. As such, the squirrel’s version of reactive gliosis could represent the best-case scenario for repairing neuron circuitry in human patients suffering CNS damage due to trauma or disease. Whereas cell signal pathways have been partially elucidated for full-version reactive gliosis in typical rodent models (rat and mouse), nothing is known of the attenuated pathways in ground squirrel. The long-term objective in our lab is to determine the differences in signal pathways that account for the different outcomes, with possible translational value to human medicine. The goal of this pilot study was to achieve proficiency in (a) a new-to-us damage model, known as “optic nerve crush” (ONC), and (b) initial immunolocalization of three key damage response markers: GFAP and vimentin expression and CSPG deposition. With these methods in hand, we will be positioned to compare and contrast signal pathways. Optic nerve crush damages TWO tissues: the nerve itself, which is devoid of neuronal cell nuclei (but is populated by nuclei of astrocytes and microglia); and retrograde apoptotic neuron death in the retina (Fig. 1). Therefore, we collected both optic nerve and retina tissue in our experiments. Long-Evans rats, which undergo the full reactive gliosis program after ONC, served as our positive control species. One eye from each animal received sham treatment as a negative control. A time-series of survival was arranged to attempt to “catch” the canonical chain of events after nerve crush, which is thought to be: • Microglial activation (and onset of neuron apoptosis) • Vimentin upregulation • GFAP upregulation • CSPG deposition In both rats and squirrels, we used (Fig. 2): • On-section immunolabeling using anti-vimentin, anti-GFAP, and antiCSPG to assess reactive gliosis. • Electron microscopy analysis of morphology changes. All animal procedures complied with the UW Oshkosh IACUC approval and all federal animal welfare guidelines. Thirteen-lined ground squirrels (Ictidomys tridecemlineatus) were obtained from the UW Oshkosh breeding colony. Rats (Rattus norvegicus) were obtained from Charles Rivers. McNair Scholars Program: Award Number P217A070188, National Science Foundation Research Experience for Undergraduates, 2012 Sources and Acknowledgements Tsung-I, Kuan-Ming F, Shun-Fen T. Role of Ciliary Neurotrophic Factors in Microglia Phagocytosis. Neurochemical Research. January 2009; 34(1):109-117 Rolls A, Shechter R, Schwartz M. The bright side of the glial scar in CNS repair. Nature Reviews Neuroscience. March 2009;10(3):235-241 Linberg K, Sakai T, Lewis G, Fisher S. Experimental retinal detachment in the cone-dominant ground squirrel retina; Morphology and basic immunocytochemistry. Visual Neuroscience. 2002; 19 603-619 Berlin Memorial Hospital for help embedding and sectioning tissue Fix 4% Paraformaldehyde Optic nerve crush site – local glial response Fix 2% Paraformaldehyde 2% Glutaraldehyde Embed Embed Optic nerve with axons, astrocytes, & microglia Section Section Immunolabel for GFAP, Vim, CSPG, and IBA 1 Figure 1. Representation of retina and optic nerve postcrush, not representative of scale or concentration Investigate Morphology by Electron Microscopy (data not shown) Arrow Key: Procedures completed Preliminary data obtained, troubleshooting ongoing Cross-section Immunolabeled Optic Nerve Vimentin 24 Hour Post-Crush GFAP Figure 3. Differential expression of proteins associated with glial scarring between the rat and squirrel postONC (400x). As expected, expression of vimentin and GFAP was higher in the rat than in the squirrel at 24 hours post-injury. These two markers clearly label hypertrophic astrocytic processes at this time period. The extracellular macromolecules CSPGs similarly show a higher deposition in the rat compared to the squirrel at 24 hours. While data concerning baseline expression of these proteins is not yet processed, these preliminary results suggest a more robust glial response in the damaged rat optic nerve. 24 Hour Post-Crush Ganglion cell nuclei 2 Weeks Post-Crush Retinas and Optic Nerves Figure 2. Flow Chart of methods 24 Hour Post-Crush Retina with neurons and astrocytes scattered neuron death and glial response 2 Weeks Post-Crush Left Eye: ONC injury Right Eye: Sham CSPGs Funding Support GFAP Immunolabeled Optic Nerve Figure 4. GFAP immunoreactivity at 24 hours and 2 weeks post crush. Damaged rat optic nerve shows more GFAP labeling at 24 hours than the squirrel optic nerve (A, B). Compared to 24 hr, GFAP expression at 2 weeks is higher in squirrel astrocytes (C), but they do not display the severe hypertrophic gliosis seen in the rat at 2 weeks (D). Conclusions & Future Work Vimentin, CSPG, and especially GFAP expression in the damaged optic nerve of the rat is consistently higher than that seen in damaged squirrel optic nerve. This was most evident at 2 weeks post-crush (Fig. 4C, D) when glial scars are well-developed in typical species such as the rat. These data corroborate our earlier demonstration that ground squirrel retina exhibits attenuated reactive gliosis following detachment (data available from presenter). We continue to troubleshoot procedures for electron microscopy and will soon begin to analyze our banked tissue (optic nerves and retina) for apoptotic cell death and microglial activation.