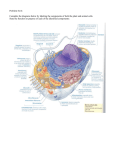
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Lecture 30 – Quiz on Wednesday: fatty acid synthesis (You will have 15 min to take the quiz; we will still have a lecture on WED.) – Membranes – Final is from 8AM-10AM on Dec. 12 Page 933 Fatty acid biosynthesis 1. Acetyl-CoA is converted by MAT to Acetyl ACP 2. Acetyl-ACP is attached to KS (condensation reaction). 3. Malonyl ACP is formed by MAT. 4. Acetyl-group is coupled to beta carbon of malonyl-ACP with release of CO2 to form acetoacetyl-ACP(2b) by KS. 5. Reduction of acetoacetyl-ACP with NADPH to form D--hydroxybutyrlACP by DH 6. Dehydration of D--hydroxybutyrlACP by ER to form a,b-transbutenoyl-ACP 7. Reduction of the double bond to form butyryl-ACP 8. Repeat until Palmitoyl-ACP (C16) is formed. 9. ACP is cleaved by TE releasing free fatty acid. Polar Lipids • Differ from triglycerides since they have one or more polar head groups. • 2 main types: glycerol or sphingosine based • Main class are phospholipids • involved in membrane structure • Amphipathic Fat molecules are hydrophobic, whereas phospholipids are amphipathic. (A) Triacylglycerol, a fat molecule, is entirely hydrophobic. (B) Phospholipids such as phosphatidyl-ethanolamine are amphipathic, containing both hydrophobic and hydrophilic portions. The hydrophobic parts are shaded red, and the hydrophilic parts are shaded blue and green. (The third hydrophobic tail of the triacylglycerol molecule is drawn here facing upward for comparison with the phospholipid, although normally it is depicted facing down.) Examples of polar lipids - Glycerophospholipids and Sphingolipids • The membrane lipids are composed of the glycerophospholipids and sphingolipids, which have polar and nonpolar regions. Figure 9.7 Glycerophospholipids Phosphatidic acid 4 major glycerophospholipids are polar/charged: Other examples of phosphoglycerol derivatives • Can be in sugars (glycolipids) • Diphosphatidyl glycerol O O RCO-CH2 O CH2-OCR O RCO-CH CH-OCR O OH O CH2-O-P-O-CH2-CH-CH2-O-P-O-CH2 O- glycerol O- Figure 9.8 Sphingolipid Structure sphingosine Fatty acid Polar head 4 major sphingolipids are: Found in nerve cell membranes brain, nervous system. Tay-Sachs disease: accumulation of ganglioside in brain and spleen causes death by age 4. Sphingolipids • Sphingomyelin (also considered a phospholipid) • Important in myelin sheath of nerve cell membrane. • Cerebrosides are glycolipids - found in brain and other tissues. • Sugar attached at P. • Ganglosides are sphingolipids with several sugars as head groups. Terminal sugar is sialic acid. Lipid/membrane consitutents that cannot be saponified. • Triacylglycerides, phospholipids, and sphingolipids can be saponified (hydrolyzed with OH-) • Some cannot be saponified: steroids (cholesterol-based) and terpenes. The molecular structure common to all steroids Steroid structures have four fused rings, A, B, C, and D. Figure 9-10b Cholesterol molecular structure - a steroid • Cholesterol has a polar head group (OH) and a nonpolar tail. • Cholesterol and ester derivatives are abundant in (blood) plasma proteins called lipoproteins. • Lipoproteins transport cholesterol to tissues for use in cell membranes and hormone precursors. Figure 9-10 The molecular structure common to all steroids • C30 Cholesterol molecule is derived from 5-carbon isoprene subunit. Cholesterol is traditionally only associated with animal cells, but derivatives are also identified in plants. Figure 9-10 (c) A cholesterol fatty acid ester • Cholesterol ester formed between the cholesterol hydroxyl group and a fatty acid with a long aliphatic side chain. • This is a common modification of cholesterol under physiological conditions. Figure 7-6 Structure of the Cholesterol, a Component of Mammalian Cell Membranes (c) The most common membrane sterols are cholesterol in animals and several related phytosterols in plants. Cholesterol and Eggs in the diet From Jennifer Moll,Your Guide to Cholesterol. About fifteen years ago, egg consumption was discouraged by many health care practitioners because of their high cholesterol content. The average intact egg contains about 210 mg of cholesterol, whereas the recommended intake of cholesterol is 300 mg. However, a study published in the Journal of the American Medical Association, in addition to several other studies, refute this. This study looked at the effects of egg consumption in 100,000 men and women, and concluded that eggs alone do not contribute to high cholesterol. In fact, when cholesterol was omitted from the diet of these subjects, their total cholesterol levels decreased only by 1%. What researchers did discover was that individuals who consumed eggs also consumed bacon, ham, butter, and other food products that could contribute to high cholesterol levels. Not only do these foods have high cholesterol, they also contain high amounts of saturated fats and trans-fats--both of which contribute to high cholesterol levels and atherosclerosis. Given these studies and the fact that eggs are an excellent source of nutrition, the American Heart Association now recommends that you can eat one egg a day, as opposed to three or four per week it previously allowed. Eggs are a rich source of protein, containing the essential amino acids required by your body. In addition to protein, eggs also contain many vitamins, minerals, and a fatty molecule called lecithin, which aids in transporting and metabolizing fats in the body .It is cautioned that if you do consume one egg a day, you might need to watch your total cholesterol levels since too much cholesterol could raise your LDL levels. Figure 7-8 The Structure of Hopanoids Abundant in petroleum (crude oil) deposits, suggesting a prokaryotic (bacterial) role in formation of oil deposits (a) A hopanoid, one of a class of sterol-like molecules that appear to function in the plasma membranes of at least some prokaryotes as sterols do in the membranes of eukaryotic cells. (b) The structure of cholesterol, for comparison. A weakly hydrophilic side chain (CH2OH or -OH) protrudes from each molecule. Figure 9-11(b-d) Structures of bioactive products produced from cholesterol. b) Estradiol - a female sex hormone. c) Testosterone - a male sex hormone. d) Cortisol - a regulator of glucose metabolism. Figure 9-11 (e,f) Structures of bile salts produced from cholesterol. e) Cholate - a bile salt derived from cholic acid. f) Glycholate - a bile salt derived from glycholic acid. • Bile salts are stored in the gallbladder, secreted into intestines to solubilize, adsorb dietary fats. • Bile salts have carboxylic acid groups that ionize at physiological pH (>pH 5.0) Terpenes • Terpene class of lipids includes all molecules biosynthesized from isoprenes (including cholesterol). • Important terpenes include beta-carotene, lycopene, squalene. • Terpenes often have strong odors. Eicosanoids • • Three classes of the Eicosanoid class of lipids: 1. Prostaglandins - isolated from prostrate gland, found in nearly all tissues. 2. Thromboxanes - 6-membered rings with oxygen - may help in blood clotting. 3. Leukotrienes - isolated from leukocytes, cause contraction of smooth muscle. All are derived from polyunsaturated 20-carbon fatty acid, arachidonate (20:4 D5,8,11,14). Three classes of the Eicosanoid lipids are all derived from polyunsaturated 20-carbon fatty acid, arachidonate (20:4 D5,8,11,14). •PGE2 induces wakefulness •PGD2 promotes sleep Table 9.4 Common fat-soluble vitamins (terpenes) Figure 9.14 Two insect pheromones Housefly attractant Honeybee queen attractant Key Functions of Cell Membranes • Cell membranes have at least five distinct functional roles: 1. Define the boundaries of the cell and its organelles. 2. Serve as locations for specific functions. 3. Provide for and regulate transport processes. 4. Contain the receptors needed to detect external signals. 5. Provide mechanisms for cell-to-cell contact, communication, and adhesion. Figure 11–5 A typical membrane lipid molecule has a hydrophilic head and hydrophobic tails. Figure 9.9 Assembly of polar lipids into lipid bilayer structures (sheets) Hydrophobic interactions provide stabilizing energy to hold the bilayer together. Lipid aggregates • In aqueous solutions, amphiphilic molecules form micelles. • Have a hydrophobic interior to eliminate contacts between water and the hydrophobic tails. • Micelles form only after a critical micelle concentration is reached (dependent on the amphiphile). For short tails (dodecyl sulfate) need higher conc. (1 mM) whereas for longer hydrophobic tails in biological lipids need a lower concentration (<10-6) Lipid bilayers • • • Glycerolipids and sphingolipids form bilayers Structural basis for biological membranes Impermeable to most polar substances • A suspension of phospholipids can be disrupted using sonciation to form liposomes-closed self-sealing lipid vesicles bounded by a single-bilayer. Biological membranes are lipid bilayers with which proteins are associated. • Figure 11–11 Amphipathic phospholipids form a bilayer in water. (A) Schematic drawing of a phospholipid bilayer in water. (B) Computer simulation showing the phospholipid molecules (red heads and orange tails) and the surrounding water molecules (blue) in a cross section of a lipid bilayer. Figure 11–12 Phospholipid bilayers spontaneously close in on themselves to form sealed compartments. The closed structure is stable because it avoids the exposure of the hydrophobic hydrocarbon tails to water, which would be energetically unfavorable. Important Features of Lipid Distribution in Cell Membranes • Lipid bilayer membranes are fluid structures. • Rate of lipid diffusion within a lipid monolayer (lateral diffusion) is very rapid. • Rate of lipid exchange between layers (transverse diffusion) is very slow. • Specific proteins (e.g., in smooth ER) will move specific lipids from one membrane to the other, termed “flippases” or phospholipid translocators. Consequence: • Most membranes have unequal distribution of lipids in each monolayer of the bilayer structure: “membrane asymmetry” Figure 7-11 Demonstration of Lipid Mobility Within Membranes by Fluorescence Recovery After Photobleaching Lipid Fluidity in Cell Membranes • Lipid bilayer membranes are fluid structures. • Bilayer fluidity will vary with temperature. • Each lipid bilayer has a (Phase) Transition temperature Tm (melting temperature). • Tm increases with increasing chain length (butter vs. cooking oil). • Tm decreases with increasing degree of unsaturation (poor packing of bent tails). • Tm modulated by cholesterol: broadens effective Tm: Lowers Tm at high temp., increases Tm at low temp. Figure 11–6 Phosphatidylcholine is the most common phospholipid in cell membranes This particular phospholipid is built from five parts: the hydrophilic head, choline, is linked via a phosphate to glycerol, which in turn is linked to two hydrocarbon chains, forming the hydrophobic tail. The two hydrocarbon chains originate as fatty acids— that is, hydrocarbon chains with a –COOH group at one end—which become attached to glycerol via their –COOH groups. A kink in one of the hydrocarbon chains occurs where there is a double bond between two carbon atoms; it is exaggerated in these drawings for emphasis. The “phosphatidyl-” part of the name of phospholipids refers to the phosphate–glycerol–fatty acid portion of the molecule. Figure 7-14 The Effect of Unsaturated Fatty Acids on the Packing of Membrane Lipids (a) Membrane phospholipids with no unsaturated fatty acids fit together tightly because the fatty acid chains are parallel to each other. (b) Membrane lipids with one or more unsaturated fatty acids do not fit together as tightly because the cis double bonds cause bends in the chains, which interfere with packing. Each of the structures shown is a phosphatidylcholine molecule, with either two 18-carbon saturated fatty acids (stearate) (part a) or two 18-carbon fatty acids, one saturated (stearate) and the other with one double bond (oleate) (part b). Figure 7-13 The Effect of Chain Length and the Number of Double Bonds on the Melting Point of Fatty Acids • The melting point of fatty acids: 1. increases with chain length for saturated fatty acids. 2. decreases dramatically with the number of double bonds for fatty acids with a fixed chain length. • The 18-carbon fatty acids in part b are stearate, oleate, linoleate, and linolenate, with 0, 1, 2, and 3 double bonds, respectively. Figure 11–16 Cholesterol stiffens cell membranes 11_16_Cholesterol.jpg (A) The structure of cholesterol. (B) How cholesterol fits into the gaps between phospholipid molecules in a lipid bilayer. The chemical formula of cholesterol is shown in Figure 11–7. Orientation of Cholesterol Molecules in a Lipid Bilayer (a) Cholesterol molecules are present in both lipid layers in the plasma membranes of most animal cells, but a specific molecule is localized to one of the two layers. (b) Each molecule orients itself in the lipid layer so that its single hydroxyl group is close to the polar head group of a neighboring phospholipid molecule, where it forms a hydrogen bond with the oxygen of the ester bond between the glycerol backbone and a fatty acid. Regulation of Cell Membrane Fluidity • Most organisms can regulate membrane fluidity. “homoviscous adaptation” in coldblooded poikilothermic organisms occurs by regulating lipid composition: • • • • Change ratio of 16-carbon to 18-carbon lipids via hydrolase enzyme (smaller lipids have lower Tm). Change degree of unsaturation (via desaturase enzyme). Important for bacteria, plants, reptiles, amphibians, hibernating mammals. Figure 9.16 Distribution of lipids in two monolayers of the human erythrocyte Figure 7-7 Phospholipid Composition of Several Kinds of Membranes The relative abundance of different kinds of phospholipids in biological membranes varies greatly with the source of the membrane. Table 9.5 Figure 11–1 Cell membranes act as selective barriers. 11_01_Cell.membranes.jpg Membranes serve as barriers between two compartments— either between the inside and the outside of the cell (A) or between two intracellular compartments (B). In either case the membrane prevents molecules on one side from mixing with those on the other. Figure 7-3 Timeline for the Development of the Fluid Mosaic Model •The fluid mosaic model of membrane structure that Singer and Nicholson proposed in 1972 was the culmination of studies that date back to the 1890s. •Future developments and revisions? Figure 11–15 Phospholipids can move within the plane of the membrane (Fast) (Very Slow) 11_15_Phospho,move.jpg (Fast) (Fast) The drawing shows the types of movement possible for phospholipid molecules in a lipid bilayer. Figure 7-10 Movements of Phospholipid Molecules Within Membranes A phospholipid molecule is capable of three kinds of movement in a membrane: rotation about its long axis; lateral diffusion by exchanging places with neighboring molecules in the same monolayer; and transverse diffusion, or "flip-flop," from one monolayer to the other. In a pure phospholipid bilayer at 37 degrees C, a typical lipid molecule exchanges places with neighboring molecules about 10 million times per second and can move laterally at a rate of about several micrometers per second. Flip-flops are rare, occur < 1 week-1 - Hr-1 Figure 9.18 The fluid mosaic model for biological membranes Figure 10–33 Freeze-fracture electron microscopy. This drawing shows how the technique provides images of both the hydrophobic interior of the cytosolic half of the bilayer (the P face) and the hydrophobic interior of the external half of the bilayer (the E face). After the fracturing process, the exposed fracture faces are shadowed with platinum and carbon, the organic material is digested away, and the resulting platinum replica is examined in an electron microscope. Figure 10–34 Freeze-fracture electron micrograph of human red blood cells. Note that the density of intramembrane particles on the cytosolic (P) face is higher than on the external (E) face. Figure 9–33 The thylakoid membranes from the chloroplast of a plant cell. In this freeze-fracture electron micrograph, the thylakoid membranes, which perform photosynthesis, are stacked up in multiple layers. The plane of the fracture has moved from layer to layer, passing through the middle of each lipid bilayer and exposing transmembrane proteins that have sufficient bulk in the interior of the bilayer to cast a shadow and show up as intramembrane particles in this platinum replica. The largest particles seen in the membrane are the complete photosystem II—a complex of multiple proteins. Figure 11–21 Membrane proteins can associate with the lipid bilayer in several different ways 11_21_proteins.associ.jpg (A) Transmembrane proteins can extend across the bilayer as a single a helix, as multiple a helices, or as a rolled-up sheet (called a barrel). (B) Some membrane proteins are anchored to the cytosolic surface by an amphipathic a helix. (C) Others are attached to either side of the bilayer solely by a covalent attachment to a lipid molecule (red zigzag lines). (D) Finally, many proteins are attached to the membrane only by relatively weak, noncovalent interactions with other membrane proteins. Figure 7-19 The Main Classes of Membrane Proteins Membrane proteins are classified according to their mode of attachment to the membrane. Integral membrane proteins contain one or more hydrophobic regions that are embedded within the lipid bilayer. Peripheral membrane proteins are too hydrophilic to penetrate into the membrane but are attached to the membrane by electrostatic and hydrogen bonds that link them to adjacent membrane proteins or to phospholipid head groups. Lipid-anchored proteins are hydrophilic and do not penetrate into the membrane; they are covalently bound to lipid molecules that are embedded in the lipid bilayer. (f) Proteins on the inner surface of the membrane are usually anchored by either a fatty acid or a prenyl group. (g) On the outer membrane surface, the most common lipid anchor is glycosylphosphatidylinositol (GPI). Page 403 Figure 12-29 Prenylated proteins. (a) A farnesylated protein and (b) a geranylgeranylated protein. Prenylated proteins • • Derived from isoprenoid groups Proteins associated with intracellular membrane but also facilitate protein-protein interactions. • For farnsylated and geranylgeranylated proteins, the prenyl group is attached to a specific C-terminal sequence. CaaX where C is Cys, a is an aliphatic amino acid and X is any amino acid. • • • • The aaX portion is cleaved after the prenylation of the protein. If X is Ala, Met, or Ser, the protein is farnyslated If X is Leu, the protein is geranylgeranylated Page 404 Figure 12-30 Core structure of the glycosylphosphatidyl inositol (GPI) anchors of proteins. Glycosylphosphatidylinositol (GPI) • • • Anchor proteins to the exterior of the eukaryotic membrane. Alternative to transmembrane polypeptides Proteins destined to be anchored to the surface of the membrane are synthesized with membrane spanning Cterminal sequences which are removed after GPI addition. Figure 11–20 Plasma membrane proteins have a variety of functions. Figure 9.19 Passive transport of solute molecules through a permeable membrane Figure 9.20 Three types of membrane transport Figure 9.23 Glucose permease of erythrocyte membrane Passive transport system - intracellular [glucose] =< plasma [glucose] concentration. Also transports epimers of glucose - mannose, galactose at slower rates (20%) Figure 9.23 Transport of sodium and potassium ions by Na+-K+ ATPase transporter (pump) Three Na+ ions are transported out of the cell for every two K+ that move inside Lipases • • How are lipids accessed for energy production? Know the differences between the triacylglycerol lipase and phospholipase A2 mechanisms. • Triacylglycerol lipase uses a catalytic triad similar to Ser proteases (Asp, His, Ser) Phospholipase A2 uses a catalytic triad but substitutes water for Ser. • Page 911 Figure 25-3a Substrate binding to phospholipase A2. (a) A hypothetical model of phospholipase A2 in complex with a micelle of lysophosphatidylethanolamine. Page 912 Figure 25-4a The X-ray structure of porcine phospholipase A2 (lavender) in complex with the tetrahedral intermediate mimic MJ33. Figure 25-4bThe catalytic mechanism of phospholipase A2. Page 912 What other mechanism does this look like? What are the differences? Fatty acid binding proteins • • • Fatty acids form complexes with intestinal fatty acidbinding protein (I-FABP) which makes them more soluble. Chylomicrons-transport exogenous (dietary) triacylglycerols and chloestorl packaged into lipoprotein molecules from the intestine to the tissues. Chylomicrons are released into the bloodstream via transport proteins named for their density. • VLDL (very low density lipoproteins), LDL (low density lipoproteins) - transport endogenous (internally produced) triacylglycerols and cholesterols from the liver to tissues “Bad” • HDL (high density lipoproteins), - transport endogenous cholesterol from the tissues to liver - “Good” Page 439 Table 12-6 Characteristics of the Major Classes of Lipoproteins in Human Plasma. Page 442