Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
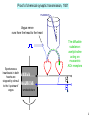
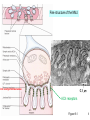
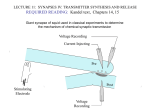
Henry Lester’s “office” hours Mon, 1:15-2 PM, Fri 1:15-2 outside the Red Door Bi/CNS 150 Lecture 5 Wednesday, October 9, 2013 Revised after lecture 10/9/13 Presynaptic transmitter release Chapters 9, 12 (co-written by T. Sudhof, one of this week’s Nobel Prize awardees) 1 Proof of chemical synaptic transmission, 1921 Vagus nerve runs from the head to the heart Spontaneous heartbeats in both hearts are stopped by stimuli to the “upstream” vagus The diffusible substance: acetylcholine acting on muscarinic ACh receptors smoked drum 2 electrical transmission in axons: electric field open closed Past lectures: V-gated Na+ channels V-gated K+ channels Today: V-gated Ca2+ channels chemical transmission at synapses: [neurotransmitter] closed open Friday: ACh-gated excitatory cation (Na+ / K+ / Ca2+) channels & GABA- and glycine-gated inhibitory anion (Cl- channels Next week: Glutamate-gated excitatory (Na+ / K+ / Ca2+) channels 3 Many basic principles of chemical transmission and developmental neuroscience were discovered at the neuromuscular junction (nerve-muscle synapse); acetylcholine is the transmitter. Figure 9-1 4 Fine structure of the NMJ 0.3 µm Incl. acetylcholinesterase ACh receptors Figure 9-1 5 Life cycle of a synaptic vesicle Figure 12-10 6 Caught by flash-freezing, invented at Caltech ~ 50 yr ago A. Van Harreveld Presynaptic terminal postsynaptic cell Like Figure 12-7 7 Proteins associated with synaptic vesicles, slide 1 Vesicles can be isolated from brain tissue by cell biological methods A. Homogenize brain in isotonic sucrose. B. Isolate synaptosomes (cut-off nerve terminals) by differential and sucrose gradient centrifugation C. Lyse synaptosomes in hypotonic solution to release vesicles. D. Isolate vesicles by glass bead column chromatography. 8 Proteins associated with synaptic vesicles, slide 2 Synaptophysin Synaptotagmin (the Ca2+ sensor) Snares (residents of either the vesicle [v-snare] or the target membrane [t-snare]) VAMP (also called synaptobrevin), a v-snare Syntaxin, a t-snare that also associates with Ca2+ channels SNAP-25, a t-snare (~25 kD) ATP-driven proton pump creates concentration gradient that drives neurotransmitter uptake against concentration gradient (one of three transporters that function in transmitter release) Lecture 1 asked, “Could cells utilize plasma membrane H+ fluxes?” “Probably not. There are not enough protons to make a bulk flow, required for robustly maintaining the ion concentration gradients. (but some very small organelles (~ 0.1 mm) and bacteria do indeed store energy as H+ gradients).” Mary Kennedy’s work 9 vesicle interior How synaptic vesicles fill from the cytosol H+ Transporter #1: ATP-driven proton pump cytosol Transporter #2: Proton-coupled neurotransmitter transporter Neurotransmitter and ATP cytosol (3,000 to 10,000 molecules of each) vesicle interior ~ isotonic! See Figure 13-1A 10 From Lecture #1 Transporter # 3. Na+-coupled cell membrane neurotransmitter transporters: Antidepressants (“SSRIs” = serotonin-selective reuptake inhibitors): Prozac, Zoloft, Paxil, Celexa, Luvox Drugs of abuse: MDMA Na+-coupled cell membrane serotonin transporter Attention-deficit disorder medications: Ritalin, Dexedrine, Adderall Trademarks: Drugs of abuse: cocaine amphetamine Presynaptic terminals Na+-coupled cell membrane dopamine transporter cytosol NH 3+ HO outside HO N H See Figure 13-1B, C HO H2 C C H2 NH3+ 11 From a previous recent lecture Atomic-scale structure of (bacterial) Na+ channels (2011, 2012) As of fall 2013, there are no crystal structures of voltagegated Ca2+ channels. From the similarities in sequence, we expect the secondary and tertiary structures to resemble those of K+ and Na+ channels. A voltage-gated Na+ channel can be changed to a voltagegated Ca2+ channel by mutating ... just 2 out of 1800 amino acids See Table 12-1 12 Electricity, then chemistry triggers synaptic vesicle fusion docked vesicle nerve impulse Na+ and K+ channels neurotransmitter voltage-gated Ca2+ channel We’ll show a more complete animation in a few minutes See 1st part of Chapter 12 13 Electricity, then chemistry triggers synaptic vesicle fusion docked vesicle nerve impulse Na+ and K+ channels Ca2+ neurotransmitter voltage-gated Ca2+ channel We’ll show a more complete animation in a few minutes See 1st part of Chapter 12 14 Electricity, then chemistry triggers synaptic vesicle fusion 1. The Na+ channels have produced the voltage change (depolarization); the K+ channels have rendered it brief (~ 1 ms) fused vesicle Ca2+ 2. The Ca2+ channels produce some depolarization, but their main function: to introduce the intracellular messenger Ca2+ See 1st part of Chapter 12 neurotransmitter We’ll show a more complete animation in a few minutes 15 Synaptotagmin is the calcium sensor Synaptotagmin has as many as 40 Ca2+-binding sites. Perhaps binding of more Ca2+ increases the rate of fusion and/or pushes the vesicle toward the “slow track” and full fusion. Like Figure 12-13 Animation of “full collapse fusion”: http://stke.sciencemag.org/content/vol2004/issue264/images/data/re19/DC2/slowtrack2.swf 16 Other proteins that act on synaptic vesicles II. Peripheral membrane proteins A. Synapsins anchor vesicles to cytoskeleton. B. Rab 3A is a GTPase perhaps involved in vesicle trafficking III. Soluble proteins that participate in vesicle fusion and release A. SM proteins Munc-18-1 binds to the N-terminus of syntaxin and participates in vesicle docking and priming. Munc-13 - essential for all forms of synaptic vesicle fusion, participates in vesicle priming. B. Complexins interact with SNARE complex and stabilize SNARE complex. C. NSF and its associated proteins are needed for SNARE recovery. 17 An alternative form of Ca2+-dependent vesicle fusion, termed fast tracking, or “kiss and run” predominates at low frequency stimulation Animation: http://stke.sciencemag.org/content/vol2004/issue264/images/data/re19/DC2/newFasttrack2.swf 18 Transmitter release depends strongly on extracellular Ca concentration Experiments at the squid giant synapse, which excites the giant axon (See Figs. 12-1, 12-2, 12-3) Cooperative processes cause nonlinear relation between [Ca2+] and transmitter release HAL’s first paper, Nature 1970 19 Timing of synaptic events “Synaptic delay”, between the peak of the action potential and the start of transmitter release, is ~ 0.5 ms. Delay between the peak of the Ca2+ current and the beginning of the EPSP is ~ 0.2 ms (more at lower temperature). Most of the “synaptic delay” is caused in opening of Ca2+ channels during the action potential. mV The size and timing of the EPSP’s can be modulated by prolonging the action potential. Figure 12-1 20 stimulus to presynaptic motor axon, producing action potential measured postsynaptic response V large “synaptic potential” leads to muscle action potential +60 mV -60 1 Electrophysiological analysis of quantal synaptic transmission (slide 1) ms 5 subthreshold synaptic events (revealed in low Ca2+) (Figure 12-6, Box 12-1) 21 stimulus to presynaptic motor axon, producing action potential measured postsynaptic response V repeated identical stimuli to the presynaptic neuron . . . 5 mV Electrophysiological analysis of quantal synaptic transmission (slide 2) (Figure 12-6, Box 12-1) 5 ms . . . yield variable postsynaptic responses! 22 Electrophysiological analysis of quantal synaptic transmission (slide 3) repeated stimuli to presynaptic neuron 5 mV Fraction of Observations Analysis of Quantal Synaptic Transmission 1 0.9 0.8 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0 Stimulated Spontaneous 01 12 23 34 45 56 Amplitude of Postsynaptic Response (mV) no stimulus; spontaneous “miniature” postsynaptic potentials 50 - 1000 channels (differs among types of synapse). This is induced by the transmitter in a single vesicle. (Figure 12-6, Box 12-1) 23 Electrophysiological analysis of quantal synaptic transmission (slide 4): Binomial statistics of vesicle release N vesicles per terminal (3 in this example) p probability of release per vesicle what is the probability P of releasing n vesicles? (n = 2 for this action potential) N n P(n) p 1 p N n n As N and p 0, binomial distribution becomes Poisson distribution (Figure 12-6, Box 12-1) N and p sometimes change during memory, learning, and drug addiction 24 Electrophysiological analysis of quantal synaptic transmission (slide 5): Summary of the classical evidence: 1. Stimulated postsynaptic potentials (psp’s) have variable amplitudes 2. Spontaneous “miniature” postsynaptic potentials occur with only modest amplitude variability. 3. The amplitudes of the stimulated psp’s are integral multiples of the spontaneous “miniature” psp’s (Figure 12-6, Box 12-1) 25 A more direct electrical measurement of quantal release: Measuring the presynaptic capacitance increase due to vesicle fusion fused vesicle adds capacitance inside outside Na+ inside K+ Cl- G DC C E outside See Figure 12-8 26 Measuring the presynaptic capacitance increase due to vesicle fusion DC ~ 1 femtofarad Na+ = 1 fF = 10-15 F K+ Cl- G DC C E To measure the conductances, we set IC = CdV/dt = 0, but DG/dt 0. To measure capacitance, we set IC = CdV/dt 0, but DG/dt = 0. Phys1 reminders, as usual See Figure 12-8 27 On a time scale of seconds, Signaling at synapses occurs via 2 classes of mechanisms Discussed today 1. Chemical signaling is the dominant form in mammalian nervous systems. A. A chemical transmitter is secreted by the presynaptic terminal and diffuses within the gap or “cleft”, binding with specialized receptors in the membrane of the postsynaptic cell. B. The bound transmitter receptor can electrically excite or inhibit the postsynaptic cell. It sometimes also “modulates” the action of other transmitters. Not discussed today: 2. Electrical signaling results when current generated in one cell spreads to an adjacent cell through low resistance channels called “gap junctions” (see pages 178 – 185) 28 Reminder: Henry Lester’s “office” hours Mon, 1:15-2 PM, Fri, 1:15-2 PM outside the Red Door End of Lecture 5 29