Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Introduced species wikipedia , lookup
Habitat conservation wikipedia , lookup
Latitudinal gradients in species diversity wikipedia , lookup
Biological Dynamics of Forest Fragments Project wikipedia , lookup
Mascarene Islands wikipedia , lookup
Cocos Island wikipedia , lookup
Biodiversity of New Caledonia wikipedia , lookup
Theoretical ecology wikipedia , lookup
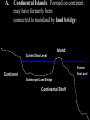
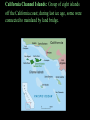
Lecture 5 Insular Gigantism and Dwarfism-island biogeography • Island giants are aplenty: Komodo has its dragons. Madagascar has its giant hissing cockroach. Until about 1,000 years ago, New Zealand had its colossal bird, the moa. • Of dwarves, the world has witnessed everything from foxes, rabbits, and snakes that are smaller than their mainland counterparts, to that ultimate oxymoron(矛盾), the pygmy mammoth, which once existed in various forms from California's Channel Islands to Wrangel Island in the Siberian Arctic • Why does this happen? • What factors encourage a species to alter its dimensions on islands? • What, in short, determines whether a creature will get Brobdingnagian or Lilliputian? insular gigantism Giant Weta Elephant Bird Japanese Spider Crab Giant Isopod Giant Isopod in in cold, deep waters of the Atlantic a good example of deep-sea gigantism 3-foot, 22-lb rabbit East Timor Giant Rat Galapagos Giant Tortoise • Komodo Dragon Komodo Dragon Giant moe Giant Squid Insular Dwarfism Little People of Flores Could a tiny sub-species of in the genus Homo have co- existed in Indonesia with humans as recent as 12,000 years ago? First dubbed a “hobbit-like human ancestor”, it was soon discovered that Homo floresiensis was in fact its own species, standing just three feet tall, about the height of a modern human toddler. Nine skeletons were found in Flores, Indonesia in 2003 The team that discovered H. floresiensis believe the species is an example of insular dwarfism, with their growth restricted by a limited choice of food on the island Dwarf Elephants prehistoric dwarf elephants evolved to be much smaller than modern elephants due to their insularity on islands around the world including Crete, Cyprus, Timor and the same island of Flores, Indonesia where pygmy human relatives were found dwarf elephants really were small: the Cyprus dwarf elephant likely weighed around 440 pounds THE ISLAND RULE The table reveals some interesting trends :Rodents tend toward gigantism, while carnivores, lagomorphs (兔形类,rabbits and hares), and artiodactyls (偶蹄 类,deer, hippos, and other even-toed ungulates) are more likely to become dwarfed. Overall, amongst mammal species that colonize islands, big ones have a tendency to shrink while small ones are apt to enlarge. Biologists have come to call Foster's generalization the "island rule." Evolution of Mammals on Islands." Nature 202: 234-235 (18 April 1964). Giants and dwarfs Three hypothesis for gigantism of island species (Schwaner & Sarre 1988) ①Predation hypothesis: either there is selective release if no predation occurs or there is selective advantage to escape a window of vulnerability ②Social-sexual hypothesis: due to high densities that occur among island populations, intraspecific competition among males and females selects for larger body size ③Food availability hypothesis: increase in the mean and variance in food supply/demand ratio selects for giants Features of isolated island endemics 1: Size changes • Birds and insects may become giant and/or flightless. (Giant Earwig of St Helena, Dodo of Mauritius, elephant bird of Madagascar, Flightless rails all that used to be all over the Pacific - now confined to Henderson island). • Mammals if present may become dwarf: Cypress had pygmy hippos. Mallorca had an endemic dormouse and an elephant, both about the same size! Komodo dragons evolved to predate pygmy elephants. • Tortoises where present become giant Galapagos and Aldabra. Features of isolated island endemics 2: Lifestyle changes • The Laysan finch looks like a sparrow, but lives like a vampire bat, sucking blood from albatrosses. • Galapagos finches have evolved to use cactus spines as a tool. • Hawaii has a caterpillar which catches flying insects. • In Hawaii Lobelias are giant trees. • A Seychelles tree Pisonia grandis has large sticky flowers which catch nestling terns. The tree benefits from their nutrients as they decay carnivorous flowers. Features of isolated island endemics 3: Vulnerability • Almost all island endemics are automatically a conservation worry due to small geographical range. • In addition: – They have no fear of predation. – They tend to be K selected - few large offspring. – They have no tolerance of disease. Predation • It is quite normal for wild birds in remote systems to see humans as useful landing posts! This lack of fear reflects evolutionary heritage, but is a disaster in terms of survival. • A consistent pattern is that remote islands used to hold giant flightless birds, until humans arrived. – – – – Geese in Hawaii Moas in New Zealand Kakapos in New Zealand Giant owls in the bahamas. • It seems clear that in many cases we simply ate the species to extinction. Biowarfare • Far worse damage was done by the species we introduced. Rats, cats, pigs and goats are the worst, but deer, ferrets and possums are also causing damage in New Zealand. • One lighthouse keeper’s cat brought home one entire population of the Chatham island robin Petroica traversi home, dead, one by one! (a 2nd popn survived - now 100 birds descended from 1 female) • Rats swim ashore from shipwrecks, and are destructive predators of ground-nesting birds. Removing rats from the Isle of May involved 2 tons of warfarin. Saving the dark herald petrel in Pitcairn involved spreading 2 tons of brodifacoum on a 65ha island. • The biggest single killer of native Hawaiian birds came from one barrel of water thrown overboard in the 1880s. This introduced mosquitos, which vectored avian malaria. Now the surviving endemic birds are in high, cold mosquito-free forests. Almost all bird life on Guam has been wiped out by the introduction of a Solomon-island bird eating snake Boiga irregularis, which stowed away with the US military. Woods are now full of spiders webs, as there are no birds to eat spiders or snap their webs. Island Biogeography Definition: A subdivision of biogeography that relates the manner in which species distributions are influenced and restricted by “islands.” The “island” is any area of habitat surrounded by an inhospitable matrix to the species occurring on that island. Image credit: http://www.okstate.edu/artsci/botany/bisc3034/lnotes/islands.htm Types of Islands A. Continental Islands: Formed on continent; may have formerly been connected to mainland by land bridge: Island Current Sea Level Former Continent Sea Level Submerged Land Bridge Continental Shelf Examples of Continental Islands 1. British Isles 2. California Channel Islands 3. Block Island, Nantucket, Martha’s Vineyard British Isles: Land mass is part of European continent. During the last ice age, Britain was connected to Europe by a plateau called Doggerland. Doggerland Source: New Scientist, 8 Nov. 2008 As Ice Age ended, rising sea level flooded Doggerland and formed English Channel. North Sea Dogger Bank, an upland area of Doggerland, outlined in red. England France California Channel Islands: Group of eight islands off the California coast; during last ice age, some were connected to mainland by land bridge. B. Oceanic Islands: Never connected to continent; usually formed by volcanic activity and isolated from continent by deep ocean. Oceanic Island Current Sea Level Continental Shelf Former Sea Level Undersea Volcano Sea Floor Examples of Oceanic Islands • Iceland • Japan • Aleutians • Bermuda • Caribbean Islands • Hawaiian Islands • South Pacific Atolls • Et al. Many Caribbean islands were formed by volcanic activity at subduction zone. Virtual Islands Isolated communities separated via some sort of barrier Ex. Sky Islands (southeastern Arizona/ southwestern New Mexico) Ex. Discontiguous habitats created via fragmentation Ex. Caves! Islands are important natural laboratories for the study of biogeography, ecology, population genetics, evolutionary biology, etc. Early naturalists (e.g., 16th-18th centuries) exploring isolated islands noted new types of plants and animals, which were often distinctive for each island or island group For several centuries, scientific focus was on cataloging the diversity of island organisms Darwin observed dozens of animal species unique to the Galapagos …including 13 species of Galapagos Finches 1859 - Publication of “On the Origin of Species” Darwin speculated on possible means by which organisms colonized islands and evolved into new species (e.g., Galapagos finches) Species-Area Relationship and Size Isolation and Species Area Species Isolation Species Isolation Species Turnover Species Turnover Island Size Richness = island size and distance from mainland Small islands Less habitat Smaller populations Higher rates of extinction (intra,inter-specific competition) Island Distance Richness = island size and distance from mainland Distant islands Lower rates of colonization However, this does depend on dispersal mechanism of the species! Dispersal vs. Vicariance Hypotheses 1. Dispersal Hypothesis: Species originated in one area and dispersed to other areas. 2. Vicariance Hypothesis: Areas were formerly contiguous, and were occupied by a common ancestor. Speciation occurred once barriers arose. “Theories, like islands, are often reached by stepping stones…” MacArthur and Wilson (1967) Theory of Island Biogeography MacArthur and Wilson (1963): The number of species of a given taxon that become established on an island represents a dynamic equilibrium controlled by the rate of immigration of new species and the rate of extinction of previously established species. Image Credit: http://www.sscnet.ucla.edu/anthro/faculty/arnold/california-lab.htm Formation of a New Island • Island of Krakatau – Massive volcanic eruption in 1883. – Destroyed two-thirds of island. Also, eradicated life on neighboring islands of Rakata, Sertung, and Panjang. Formation of a New Island • 1930, a new island was formed from volcanic activity (Anak Krakatau). • Recolonization studies: – Nine months after 1883 eruption: first colonist of Rakata was a spider. – 1896, 11 species of ferns and 15 species of flowering plants. 16 species were dispersed by wind and another 8 by sea. Formation of a New Island – Recolonization of Rakata was greatly affected by how well plants were able to disperse. – Early plant communities were dominated by grasses. – 25 years later, plant communities were dominated by Cyrtandra bushes. – In the 1920s, the plant communities were dominated by Neonauclea trees. Formation of a New Island • Initially, wind and sea dispersed plants were more easily dispersed than those that required animals. – After 40 years, animal dispersed species became as common as wind and sea dispersed. Formation of a New Island • Recolonization of islands was based on the size of the island and the distance of the island to source of colonists, and the ability of an organism to disperse Theory of Island Biogeography • Equilibrium theory of insular zoogeography - first comprehensive theory of island biogeography: Robert MacArthur and E.O. Wilson (1963, 1967). – The number of species on an island tends toward an equilibrium number. Theory of Island Biogeography • This equilibrium number is the result of a balance between the rate of immigration and the rate of extinction. – Rate of immigration is highest when there are no species present on the island. – Rate of extinction is low at the time of first colonization. – Eventually, rate of extinction will equal rate of immigration. Theory of Island Biogeography • Both immigration and extinction lines should be curved. – Species arrive at an island at different rates. – Extinctions rise at accelerating rates. • As more species arrive, competition increases. • r-selected species arrive first (poor competitors followed by K-selected species (better competitors). Theory of Island Biogeography • The equilibrium number of species is determined only by the island’s area and position, which influences the rate of immigration and extinction. Theory of Island Biogeography • Equilibrium is dynamic; hence following colonization of an island: – Number of species remains constant. – Extinction = immigration. – Results in a turnover of species. Theory of Island Biogeography • Major modifications to MacArthur and Wilson’s theory of island biogeography. • Target effect (Whitehead and Jones 1969) – The rate of immigration depends on an island’s size. Theory of Island Biogeography • The rescue effect (Brown and Kodric-Brown 1977). – The distance from an island to a source pool of potential colonists affects both rate of extinction and rate of immigration. Theory of Island Biogeography • Target and rescue effects complete MacArthur model. Theory of Island Biogeography • Concept of an island – Patches of particular habitat on continents are viewed as islands in a sea of other unsuitable habitat. Theory of Island Biogeography • Strength of MacArthur-Wilson model: generated falsifiable predictions. • Prediction 1: the number of species should increase with increasing island size. Theory of Island Biogeography • Prediction 2: the number of species should decrease with increasing distance of the island from the source pool. Theory of Island Biogeography • Prediction 3: the turnover of species should be considerable – the number of species on the island might remain the same, but the identities of those species should change. Species-Area Effects • Oceanic islands - Studies of biogeography: Lesser Antilles – Islands enjoy a similar climate, surrounded by deep waters, and no historical connections to the mainland. Species-Area Effects • Ricklefs and Lovette (1999) summarized species richness for birds, bats, reptiles & amphibians, and butterflies over 19 islands that varied in area (13 – 1,510 km2). – Significant relationship between area and richness. Species-Area Effects • Habitat islands – James Brown (1978) - mountain ranges of the Great Basin. • Mountain ranges are essentially isolated from one another. • Significant relationship between species and area for mammals and birds. Species-Area Effects • As larger areas are sampled, fewer new species are added on continents than on islands. • Continents have more transient species. Species-Area Effects • In this study, the results for mammals were consistent with island results, while the results for birds showed less of an effect. Species-Area Effects • Species as islands – Species of host plants act as islands in a sea of other vegetation for the herbivores that eat from the plants. Species-Area Effects • Elaborated by Donald Strong (1974) – Found a species–area relationship between geographical area of distribution of British tree species and the number of insect herbivore species. Species-Area Effects • Entire island of Great Britain was divided up into 10-km2 grids. • Area the tree occupied in Britain was determined. • Number of insect herbivores per species of tree was determined. Species-Area Effects • Reasons for a species-area relationship (Hart and Horwitz 1991). – Extinction rates are greater on small islands. – The passive effect of increased sampling effort in bigger areas increases the number of rare species found. – Speciation may be more likely in bigger areas, an explanation also given for greater species richness in the tropics. Species-Area Effects • Larger areas contain more “core” areas, which are less affected by disturbances. – Perimeter areas contain more species that are sensitive to these disturbances. • The species-area relationship may more likely be the result of an increased diversity of habitats on large islands than just an increase in area relationships. Species-Area Effects • Larger areas often contain greater diversity of habitats. – Barry Fox (1983): investigated the relationship between species, area, and habitat diversity in Australian mammals. • Classified habitats into seven broad types. • Larger areas include more types of habitats. • Number of mammalian species is well predicted by area. • However, species richness was better predicted from the number of habitats than from area. Species-Area Effects • Dan Simberloff (1976a,b); investigated the effect of area alone on the richness of species. – Chose habitats that do not change as you sample bigger islands. – Studied islands of pure mangroves of varying size in the Florida Keys. – Collected every species that fed on the islands. Species-Area Effects • Reduction in area caused a reduction in the richness of invertebrate species. • Area of islands was reduced experimentally. – Seven months later, the insects became reestablished at equilibrium. – Insect densities dropped on all experimental islands. The Effect of Distance on Island Immigration • MacArthur & Wilson’s best evidence for the effect of distance on island immigration came from a study of the numbers of land and freshwater bird species on four groups of islands. The Effect of Distance on Island Immigration • The relationship between area and number of species is clear. • There is also a distinct effect of distance – nearer islands support more species. The Effect of Distance on Island Immigration • Jared Diamond (1972); relationship between distance and number of species. – Tabulated land birds on islands close to the source area (New Guinea), and assumed these islands had 100% of the available birds. –He documented drop-off in species with increasing distance from New Guinea. The Effect of Distance on Island Immigration • Degree of saturation: richness of bird species as a proportion of the number found on New Guinea. – Strong decline with increasing distance. • Supports MacArthur-Wilson’s predictions. Species Turnover • Francis Gilbert (1980); investigations of turnover. – Found 25 investigations to demonstrate turnover; determined that most of them suffered from fatal flaws. • Methodology, statistics, or quality of data. Species Turnover • Ex. Jared Diamond (1968) studied birds of California’s Channel Islands National Park. – Compared his list to that of A. B. Howell (1917). – Diamond reported that 5-10 species per island were no longer present, but just as many species not listed by Howell had apparently colonized the island – indicating turnover. Species Turnover • Results were challenged: Lynch and Johnson (1974) pointed out that Howell’s list was not exhaustive and just a summary of all known breeding records (some as old as 1860). • Comparing old list with new lists can be problematic. Species Turnover • Simberloff and Wilson (1969, 1970); only study of turnover with any merit. – Censused small (11 to 25 m in diameter) red mangrove islands in Florida Keys for all terrestrial arthropods. Species Turnover • Fumigated their experimental islands with methyl bromide to kill all arthropods. Species Turnover • Periodically after fumigation, they censused all islands for several years. • After 250 days, most islands had similar number of arthropod species that they began with. – Supporting MacArthurWilson theory. Species Turnover • Colonization and extinction rates were observed. – Colonization rates during the first 150 days were higher on nearer islands than far islands. • Supporting MacArthur-Wilson theory. – Calculated rates of turnover were very low (1.5 extinctions per year). • Data was weak support for MacArthur-Wilson theory of turnover . Species Turnover • Same species returned to island. – Indicates the existence of biological processes that shape the final community structure the same way every time the island is recolonized. • Contrary to the theory of biogeography. • Treats the dynamics of different colonizing species as equivalents. • Community properties unimportant. Species Turnover • Conclusion – Turnover involves only a subset of transient or unimportant species, with more important species becoming permanent after colonization. Species Turnover • Take home message: turnover rates are low, which gives little support to this part of MacArthur-Wilson theory. Theory of National Park Design • Shape, design and management of nature reserves. • Centered on island biogeography theory, which suggests that large parks hold more species than smaller ones. Theory of National Park Design • International Union for Conservation of Nature and Natural Reserves (IUCN) stated that refuge criteria and management practice should be based on the equilibrium theory of island biogeography. – Recommendations are on shaky ground. • Large areas cost a lot of money. Theory of National Park Design • Which is better, single large areas or several small ones? – Single large preserves may buffer populations against extinction. – Many studies show that multiple small sites contain more species (broader range of habitats). Theory of National Park Design • Fauna were shown to be richer in collections of small national parks than in large parks. • Smaller parks are better for maintaining diversity. • Implications for future land purchases. Summary • Island biogeography theory predicts that the equilibrium number for species on an island is determined by a balance between immigration of species onto that island and extinction of species already there. Summary • The theory suggests that the number of species is determined by an island's size and position relative to a source pool of colonists. • Extinction should increase on small islands, because of their smaller populations, and immigration should decrease on far islands, because colonists have a difficult time reaching distant places. Summary • Island biogeography theory also suggests that there is much turnover on islands as new species arrive and old ones become extinct. – There is little evidence, however, to support this prediction. – Most turnover that has been documented suggests that rates of turnover are low and center mainly on transient species. Summary • Island biogeography theory may be applied to "habitat islands" as well as real islands. • In the relationship between species richness and area, the slope of the line may be steeper for true islands than habitat islands and steeper for poor dispersers like mammals than for good dispersers like birds. A = Alaxa North QilianCorridor region Y = Yangtze block Q =Qiangtang block H = the main body of the North China block J = Jiangnan block