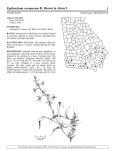
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Botanical Journal of the Linnean Society, 2009, 161, 1–19. With 5 figures INVITED REVIEW Orchid pollination: from Darwin to the present day CLAIRE MICHENEAU1*, STEVEN D. JOHNSON2 and MICHAEL F. FAY boj_995 1..19 FLS3 1 UMR 53 Peuplements Végétaux et Bio-Agresseurs en Milieu Tropical, Université de La Réunion, 15 avenue René Cassin BP 7151, 97715 Saint-Denis Messag Cedex 9, La Réunion, France 2 School of Biological and Conservation Sciences, University of KwaZulu-Natal, P Bag X01, Scottsville, Pietermaritzburg 3209, South Africa 3 Jodrell Laboratory, Royal Botanic Gardens, Kew, Surrey TW9 3AB, Richmond, UK Received 22 April 2009; accepted for publication 3 August 2009 In this year celebrating the bicentenary of the birth of Darwin and the sesquicentennial of the publication of Darwin’s On the Origin of Species, the present paper aims to assess the impact of Darwin’s legacy on the history of orchid pollination biology. To illustrate the major contribution of Darwin to this fascinating biological field, we focus on the large angraecoid orchid group and propose an overview of the complex relationships that these orchids have developed with specific pollinators. We further discuss how Darwin’s seminal work on the angraecoid orchid Angraecum sesquipedale triggered the beginning of a long debate about the evolution of long floral spurs and why his idea of reciprocal evolution or ‘coevolution’ was one of the great contributions to evolutionary biology. © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19. ADDITIONAL KEYWORDS: angraecoid orchids – bird pollination – hawkmoth pollination – long spur – Madagascar – Mascarene Islands – Orchidaceae. INTRODUCTION For the celebration of the bicentenary of Charles Darwin’s birth and the sesquicentennial of the publication of his On the Origin of Species (Darwin, 1859), the world is re-exploring the impact of Darwin and his theory of evolution on science and society. However, it is not possible to commemorate Darwin’s legacy without consideration of orchids, which took a central place in his main achievements, and to which he devoted many years of study: ‘I never was more interested in my life in any subject than this of orchids’ (Darwin, 1861 in a letter addressed to Hooker). The subject of the present paper is to revisit the history of orchid pollination biology from the time of Darwin to the present day, with an emphasis on the *Corresponding author. E-mail: [email protected] pollination mechanisms involved in the angraecoid orchids, including the famous long-spurred Madagascan species, which, through Darwin’s meticulous observations and innovative reasoning, served to bring the orchid family into the heart of evolutionary research on plant–pollinator interactions. ORCHID POLLINATION: A BRIEF HISTORY FROM DARWIN’S TIME TO THE 1930S Although, by the end of the 18th century, earlier botanists including Bernard de Jussieu and Michel Adanson had contributed to the better understanding of the general structure of orchid flowers, pollination mechanisms in orchids were first fully detailed by Darwin in his famous book, On the various contrivances by which British and foreign orchids are fertilised by insects and the good effects of intercrossing (Darwin, 1862). In addition to being ‘an inspiration to floral ecologists of his and later time’ (van der Pijl & Dodson, 1966), Darwin’s book remains a classic today. © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 1 2 C. MICHENEAU ET AL. In this book, he described his own observations on the pollination of many British orchids, his personal interpretations of the pollination mechanisms involved in some tropical species, mainly based on floral morphology, and his views on homologies in orchid flowers. The second edition, published in 1877, included an extended part dedicated to tropical orchids, based mainly on the observations made by his contemporary Fritz Müller (Darwin, 1877). These two books, in which Darwin clearly demonstrated that orchids provide strong evidence for natural selection and for adaptations that promote crosspollination, were written partly in support of his theory of evolution, which invoked much criticism after the publication of On the Origin of Species in 1859 (Darwin, 1859). However, Darwin’s idea that orchids provide evidence for adaptations to insect pollination found strong support among naturalists and influenced as never before the future directions of the fascinating field of orchid pollination biology. From that time until the 1920s, the Western world raced to gather information on insect–flower interactions, involving ‘most of our knowledge on the pollination mechanisms and insect visitors of European and North American flowers’ (Proctor, Yeo & Lack, 1996). The next major contribution to orchid pollination had to wait for the end of this descriptive period and the discovery of pseudocopulation in the genus Ophrys L. in Algeria (Correvon & Pouyanne, 1916; Pouyanne, 1917) and, soon after, in the genus Cryptostylis R.Br. in south-eastern Australia (Coleman, 1927). Sexual deception in flowers which mimic the signals of female insects, in some cases so accurately that they elicit copulatory behaviour in male insects (pseudocopulation), is almost exclusive to orchids and constitutes one of the most fascinating pollination mechanisms found in the family, if not in all flowering plants. Male insects are sexually attracted to the flower by a floral scent that imitates the olfactory cue or sex pheromone(s) of the female, often enhanced with a resemblance to the female in shape, colour and texture, and usually pick up and deposit pollen while they attempt copulation with flowers. Since its discovery in the 1920s, sexual deception has been found in a number of other orchids and remains an active topic of interest in orchid pollination and evolution (e.g. Schiestl et al., 1999, 2003; Ayasse et al., 2002; Singer, 2002; Singer et al., 2004). FROM THE 1930S TO THE 1960S During the next three decades, interest in orchid floral biology and pollinator observations declined, as the attention of biologists shifted to new disciplines such as cytology and genetics. It was only in 1966 that the second classic book of orchid pollination, Orchid flowers, their pollination and evolution, was published to commemorate the centenary of the publication of Darwin’s book (van der Pijl & Dodson, 1966). In addition to being a review of one century of orchid–pollinator observations, this book constituted a major update, taking advantage of recent observations by the authors themselves and contributions provided by other famous orchidologists including R. L. Dressler and B. Kullenberg. Van der Pijl and Dodson presented a modern appreciation of orchid pollination syndromes with an emphasis of the major role played by pollinators in the evolutionary history of the orchid family. In this, their views were deeply rooted in Darwinian thought. SINCE THE 1960S From the end of the 1960s, there was a resurgence of interest in pollination studies. This can be attributed to the development of ecology and new evolutionary concepts and models, many of which were integrated into the field known at that time as ‘biosystematics’ (Proctor et al. 1996). In addition, the formalization of new principles and methods (Jones & Little, 1983; Willson, 1983; Dafni, 1992; Kearns & Inouye, 1993) contributed to a redefinition of the scope of pollination studies as an integrated part of broader studies of reproductive biology and population dynamics. Especially in orchids, in which pollinator activity could be easily quantified by pollinarium removal and deposition, an incredible volume of quantitative data on pollination and reproductive success, both in time and space, has been gathered throughout the world: in Europe and North America (e.g. Nilsson, 1981, 1983a; Gigord, Macnair & Smithson, 2001; Smithson & Gigord, 2001; Smithson, 2002), the Neotropics and Australia (e.g. Romero & Nelson, 1986; Peakall, Beattie & James, 1987; Peakall, 1989; Zimmerman & Aide, 1989; Ackerman & Montalvo, 1990; RodríguezRobles, Meléndez & Ackerman, 1992; Ackerman, Rodríguez-Robles & Meléndez, 1994; Ackerman, Melendez-Ackerman & Salguero-Faria, 1997; Tremblay et al., 1998; Borba & Semir, 1999; Borba, Shepherd & Semir, 1999; Salguero-Faría & Ackerman, 1999; Singer & Cocucci, 1999; Parra-Tabla et al., 2000; Borba & Semir, 2001; Singer, 2001; Borba et al., 2002; Singer, 2002; Schiestl et al., 2003; Singer et al., 2004) and in South Africa (Steiner, Withehead & Johnson, 1994; Johnson & Steiner, 1997; Johnson, 1997a, b; Johnson & Kurzweil, 1998; Johnson, Linder & Steiner, 1998; Ellis & Johnson, 1999). Although noteworthy books and reviews contributed to enhancing our knowledge of the identity and specificity of orchid pollinators and their role in orchid evolution (e.g. Ackerman, 1986; Nilsson, 1992; Tremblay, 1992; © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 ORCHID POLLINATION van der Cingel, 1995; Johnson & Steiner, 2000; van der Cingel, 2001; see also Dressler, 1993), three publications, in particular, have synthesized the main conclusions that arose from this productive period. Firstly, Neiland and Wilcock (1998) undertook a comparative study of the reproductive success among 117 nectariferous and nectarless orchids, from both temperate and tropical regions. They confirmed the hypothesis that nectariferous orchids are more successful in setting fruit than nectarless species, but they also expressed concern about the lack of data on reproductive success in the tropical regions. Secondly, Tremblay et al. (2005) presented an extensive overview of orchid floral biology and reproductive success and their evolutionary consequences for orchid diversification. They reaffirmed that the seed production in representatives of the family is frequently strongly pollinator-limited and suggested that genetic drift, related to the small number of individuals that participate in reproduction when there is pollen limitation, could have played a role in the extraordinary diversification of the family. Thirdly, Jersáková, Johnson & Kindlmann (2006) reviewed the mechanisms of deceptive pollination in the orchid family and the various hypotheses that have been put forward to account for their evolution. They concluded that promotion of cross-pollination was the most compelling argument for the evolutionary maintenance of floral deception in about 40% of species in the family. In addition, the combined development of molecular biology (DNA sequencing) and phylogenetic methods during the last two decades opened a vast new field of investigation, allowing the scientific community to revisit hypotheses about relationships among organisms. The mapping of pollination syndromes (or pollinator-linked floral traits) onto phylogenetic trees has provided new insights into the ecological determination of evolutionary transitions, their polarity, the extent to which changes were evolutionarily labile or constrained and the extent to which radiations can be explained by key trait innovations (e.g. Larsen et al., 2008). A comprehensive phylogeny of the orchid family based on molecular data was lacking until the contribution of Chase et al. (2003) and such phylogenies have been crucial in determining evolutionary transitions, especially for orchids in which extensive floral convergence as a result of pollinator-mediated selection has occurred. Recently, Ramírez et al. (2007) estimated ages for the origin of the orchid family and each subfamily based on pollinia found fossilized in amber. We can now explore how orchid–pollinator interactions developed over time, which may allow us to distinguish between coevolution (plants and pollinators evolve together reciprocally) vs. sequential evolution (when the evolution of one group follows another). 3 Pollination remains an active field of research and papers are still being published in large numbers as new data are accumulated. Some groups of orchids have been and still are the focus of study, including various members of the largest subfamily, Epidendroideae (e.g. Stpiczyńska, Davies & Gregg, 2007; Brys, Jacquemyn & Hermy, 2008; Chase et al., 2009; Pansarin, de Moraes Castro & Sazima, 2009), the bee orchids and their allies (Ophrys L., Orchidoideae, e.g. Cortis et al., 2009; Devey et al., 2009; Schlüter et al., 2009) and the lady’s slipper orchids (species of Cypripedium L., Cypripedioideae, e.g. Bänziger, Sun & Luo, 2008; Case & Bradford, 2009; Li et al., 2008), for which an elegant new method for studying pollination was recently published by Case & Bradford (2009). In a recent special issue of Annals of Botany focusing on orchid biology (Fay & Chase, 2009, and associated papers), pollination of orchids was discussed in several papers. In addition to two reporting pollination studies (Cheng et al., 2009; Peter & Johnson, 2009), these discussed pollination in relation to taxonomy and phylogenetic relationships (Chase et al., 2009; Hopper, 2009; Salazar et al., 2009; van den Berg et al., 2009), species delimitation (Cortis et al., 2009; Devey et al., 2009; Hedrén & Nordström, 2009), inbreeding vs. outbreeding (Duffy et al., 2009); invasiveness (Cohen & Ackerman, 2009), conservation (Swarts & Dixon, 2009) and evolutionary development (Mondragón-Palomino & Theißen, 2009). CASE STUDY: ANGRAECOID ORCHID POLLINATION BACKGROUND The angraecoid orchids (Epidendroideae Lindl., Vandeae Lindl., Angraecinae Summerh. sensu lato; see below) are a large group of monopodial epiphytic orchids, including more than 760 species. Two genera have diversified in Central and South America and the Caribbean (Campylocentrum Benth. and Dendrophylax Rchb.f.), but most of these orchids are the product of an extensive radiation in the western Indian Ocean, including Africa, Madagascar and nearby islands. Although angraecoid orchids were historically referred to two subtribes, Angraecinae (c. 17 genera, 410 species, mostly from Madagascar) and Aerangidinae Summerh. (c. 32 genera, 350 species, mostly from Africa) (e.g. Schlechter, 1926; Dressler, 1981, 1993; Chase et al., 2003), recent insights from molecular data provided evidence to support recognition of a single subtribe, Angraecinae, which includes all angraecoid orchids (i.e. Angraecinae sensu lato; Carlsward et al., 2006). The Old World angraecoid orchids are known for their white, spectacularly long-spurred flowers highly © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 4 C. MICHENEAU ET AL. specialized for hawkmoth (sphingid) pollination. The study of the pollination biology of these orchids started with the famous prediction by Darwin, which, in spite of being widely known, deserves to be repeated here. Darwin received some specimens of Angraecum sesquipedale Thou. and was particularly intrigued by the peculiar flowers of this strange orchid which, for the plants he examined, had a nectar tube approximately ‘11 and a half inches long’ (almost 30 cm). After meticulous floral observations and different experiments of manual pollination, he concluded that only a giant hawkmoth with a ‘wonderfully long proboscis’ would be able to pollinate this spectacular orchid. The predicted pollinator, Xanthopan morganii var. praedicta, was found 41 years later in the primary forests of Madagascar (Rothschild & Jordan, 1903), but effective pollination of the orchid by this animal was only demonstrated 135 years after Darwin’s prediction (Wasserthal, 1997)! In addition to predicting the existence of a giant hawkmoth on Madagascar, Darwin suggested that the extraordinary length of the spur of A. sesquipedale was driven by natural selection because of an advantage for reproduction success when the spur is longer than the moth proboscis and the head of the moth presses firmly against the orchid column, enhancing pollination. Conversely, if the orchid spur was longer than the proboscis, selection would favour a longer proboscis because this would enable the moth to gain access to more nectar. Darwin thus posited that there would be a kind of arms race between the spur length of the orchid and the proboscis of the moth (see Nilsson, 1988; Anderson & Johnson, 2008; Pauw, Stofberg & Waterman, 2009). This idea of reciprocal evolution or ‘coevolution’ was one of Darwin’s great contributions to evolutionary biology. Since this famous prediction, the large genus Angraecum Bory has received much attention from evolutionary biologists (e.g. Wallace, 1867, 1871; Nilsson, 1992, 1998b; Wasserthal, 1997). However, floral morphology is diverse within the genus, and in the subtribe, suggesting variation in pollination systems. Notably on the Mascarene islands (Mauritius, Reunion and Rodriguez, Indian Ocean), three species (Angraecum striatum Thou., A. bracteosum Balf.f. & S.Moore and A. cadetii Bosser) are particularly interesting because their white flowers have short, wide spurs. These features were sufficiently uncommon in the genus to prompt Bosser (1987) to revise section Hadrangis Schltr. to include only these three species, making the section endemic to the Mascarenes. Pollinator observations have shown that two of these species are pollinated by small songbirds from the family Zosteropidae (white-eyes) on Reunion (Micheneau, Fournel & Pailler, 2006; Micheneau et al., 2008c), whereas the third species, A. cadetii (Reunion and Mauritius) has a highly surprising pollinator, a raspy cricket (Gryllacrididae from the order Orthoptera) (Micheneau et al., in press). At the opposite end of the scale, all endemic long-spurred species are self-pollinated on Reunion (Micheneau, 2005; Micheneau et al., 2008b; see also Jacquemyn et al., 2005). These evolutionary shifts (from outcrossing to selfing and between different pollinators) are related to recent dispersal to the Mascarene Islands, where specific long-tongue pollinators of ancestral orchid colonists were probably absent. From combined phylogenetic, biogeographic and ecological data, it now appears that angraecoid orchid diversification was strongly associated with interactions with the pollinator fauna on Reunion (Micheneau et al., 2008a). In contrast, self-pollinating species appear less likely to be involved in adaptive radiations. In the following, we survey the current state of knowledge of the pollination of angraecoid orchids. METHODS INVOLVED IN POLLINATOR OBSERVATION Various methods have been used to observe pollinators foraging on angraecoid orchid flowers and these are representative of methods that may be used to observe pollinator activity on plants more generally (see also Kearns & Inouye, 1993). Pollinators may be identified directly in the field by observing their foraging on flowers. Direct observations are commonly undertaken in a time period that corresponds to the main activity of the expected pollinators (i.e. the evening in the case of hawkmoth pollinators, e.g. Martins & Johnson, 2007). However direct observations may be time-consuming, particularly if there are low visitation rates or if pollinator behaviour is altered by human presence (e.g. birds). Depending on the behaviour of pollinators, direct observations may also involve difficulties in distinguishing simple visitors from efficient pollinators. Videotape observations in the field offer a good alternative for observing pollinator visits without influencing their behaviour and without spending days in front of flowers while nothing is happening. The development of hard-disk camcorders with night option, combined with the use of long-duration rechargeable batteries and a waterproof casing, now allow the observation of pollinators for more than 12 h, whatever the time of day or night and whatever the weather (Micheneau et al., 2006, 2008c, in press). It is also possible to identify pollinators using indirect techniques: identifiable pollinaria may be found on pollinators caught on flowers of non-orchid species or at a light trap (e.g. Dressler, 1981; Nilsson et al., 1985, 1987; Luyt & Johnson, 2001) and identifiable insect scales and hairs may be found on orchid stigmas (e.g. Nilsson & Rabakonandrianina, 1988). In these cases, pollination © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 ORCHID POLLINATION mechanisms can be deduced on the basis of floral morphology and the attachment of pollinaria on the body of the pollinators, even if observations have been largely unsuccessful in the field (e.g. Nilsson & Rabakonandrianina, 1988; Luyt & Johnson, 2001). BREEDING SYSTEMS All angraecoid orchids that have been studied are self-compatible (Nilsson & Rabakonandrianina, 1988; Luyt & Johnson, 2001; Micheneau, 2005; Micheneau et al., 2006, 2008b,c; Martins & Johnson, 2007; see also Dressler, 1993). In almost all cases, fruit set is not significantly different between self- and crosspollinated flowers. However, in Mystacidium venosum Harv. ex Rolfe, self-pollination significantly decreased fruit weight, fruit length and the percentage of seeds with embryos, which could reflect either partial selfincompatibility or, more likely, strong inbreeding depression (Luyt & Johnson, 2001). These patterns are like those observed at the family level: selfcompatibility, as a result of the capacity to set fruits from self-pollen is largely dominant within Orchidaceae (self-incompatible is estimated to occur in at least 10% of all species; Dressler, 1993), resulting in no difference in the number of fruits between selfed and crossed treatments (Tremblay et al., 2005). However, self-pollination, via a significant decrease in embryo formation in mature capsules, tends to affect seed production (see Darwin, 1875; Tremblay et al., 2005). True self-incompatibility is known from just a few orchids. Darwin stated in the second edition of The variation of animals and plants under domestication (1875): ‘As these orchids had been grown under unnatural conditions in hot-houses, I concluded that their self-sterility was due to this cause. But Fritz Müller informs me that at Desterro, in Brazil, he fertilised above one hundred flowers of the abovementioned Oncidium flexuosum Lodd., which is there endemic, with its own pollen and with that taken from distinct plants: all the former were sterile, whilst those fertilised by pollen from any other plant of the same species were fertile’. [For a discussion of this and other examples of the extensive correspondence between Darwin and Müller, see West (2003)]. According to Tremblay et al. (2005), ‘Given the apparent costs of self-pollination in most orchids, mechanisms that promote outcrossing are to be expected. What is unusual is that in orchids these are often structural (movement of rostellum, movement of column, stigmatic maturity) and not the result of genetic selfincompatibility, dichogamy, or unisexuality (although examples of all of these mechanisms exist in orchids)’, which led them to address an open question for future research: ‘is it the very existence of pollinaria and column that has promoted the reliance of orchids on structural mechanisms that promote outcrossing?’ 5 Although all angraecoid orchids studied so far are self-compatible, most, but not all, require pollinator services to achieve reproduction. A notable exception to this general rule is found on the Mascarenes, where the six long-spurred species endemic to the archipelago are all able to set fruit without pollen vectors (Jacquemyn et al., 2005; Micheneau, 2005; Micheneau et al., 2008b). Self-pollination was not expected within the angraecoid orchids, which, as mentioned before, have long been considered as a highly moth-pollinator-dependent group. Within the tribe, Dressler (1993) estimated the number of selfpollinating vandoid species as < 1%, whereas estimates of self-pollination in Orchidaceae as a whole range between 5 and 20% of species (Catling, 1990). However, high rates of self-pollination are not surprising in island floras; shifts to self-pollination are thought to provide reproductive assurance when the pollinator fauna is lacking, sparse or unspecialized (Ackerman, 1985; Barrett, 1985; Catling, 1990; Barrett, 1996; Eckert, Samis & Dart, 2006). On the isolated volcanic island of Reunion, the proportion of self-pollinating orchids is however, among the highest in the world, estimated at 46% of species (i.e. 56 out of 121 species of orchids; Jacquemyn et al., 2005). To date, only the boreal flora of the northern hemisphere has been reported to include a greater proportion of self-pollinated species (c. 50% of c. 40 species; Catling, 1990). On Reunion, the distribution of self-pollinating orchids is significantly correlated with the high elevational gradient found on the island (c. 2500 m; Jacquemyn et al., 2005). However, angraecoid orchids do not follow this pattern; self-pollination in these species is strongly positively correlated with spur length (Fig. 1), suggesting that species specialized for pollination by long-tongued hawkmoths underwent selection for self-pollination because these moths were absent or scarce at the time of colonization. POLLINATOR IDENTITY AND SPECIFICITY WITHIN THE ANGRAECOID ORCHIDS Pollinator identity A list of published pollinators of angraecoid orchid species is shown in Table 1. These records include long-tongued sphingids from Madagascar and Africa, white-eye species from the Mascarenes and insular orthopterans (see also Fig. 2). As expected, the two famous orchid species with the longest spurs (often > 30 cm in length), Angraecum sesquipedale and A. sororium Schltr., are pollinated by the two sphingid species that possess the longest tongues in Madagascar (often > 20 cm), Xanthopan morganii var. praedicta and Coelonia solani, respectively (Wasserthal, 1997; Table 1). However, although the hawkmoth fauna of Madagascar (c. 60 species) is highly endemic and © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 6 C. MICHENEAU ET AL. Figure 1. Distribution of self-pollinated angraecoid orchids on Reunion according to (A) altitude and (B) spur length (data from Jacquemyn et al., 2005; Micheneau, 2005). Table 1. Checklist of published pollinators of angraecoid orchids with the place of the attachment of pollinaria on the body of the pollinator (S, sphingids; B, birds; O, orthopterans) Orchid species Attachment of pollinarium Spur length > 20 cm (Madagascar) Angraecum sesquipedale† Xanthopan morganii (S) Angraecum sororium† Coelonia solani (S) Base of the proboscis Base of the proboscis Spur length < 20 and > 10 cm (Madagascar, Kenya) Angraecum arachnites*§ Panogena lingens (S) Angraecum compactum†§ Xanthopan morganii, Panogena lingens (S) Aerangis ellisii‡ Agrius convolvuli, Panogena lingens (S) Neobathiea grandidierana§ Panogena lingens (S) Jumellea teretifolia§ Panogena lingens (S) Aerangis fuscata§ Panogena lingens (S) Rangaeris amaniensis¶ Agrius convolvuli (S) Aerangis brachycarpa¶ Agrius convolvuli, Coelonia fulvinotata (S) Aerangis thomsonii¶ Coelonia fulvinotata (S) Base of the proboscis ventrally Base of the proboscis dorsally N/A¶¶ Base of the proboscis dorsallly Base of the proboscis dorsallly Frons or palps of the head Head or base of the proboscis¶¶ Base at the proboscis dorsally Head or base of the proboscis*** Spur length c. 5 cm (Kenya, South Africa) Mystacidium venosum** Nephele accentifera aceentifera (S) Aerangis confusa¶ Hippotion celerio, Daphnis nerii (S) Base of the proboscis dorsally Base of the proboscis dorsally Spur length < 2 cm (Reunion) Angraecum striatum†† Angraecum bracteosum‡‡ Angraecum cadetii§§ Base of the beak Tip of the beak Top of the head Zosterops borbonicus (B) Zosterops olivaceus, Z. borbonicus (rare) (B) Glomeremus sp. (O) *Nilsson et al. (1985). †Wasserthal (1997). ‡Nilsson & Rabakonandrianina (1988). §Nilsson et al. (1987). ¶Martins & Johnson (2007). **Luyt & Johnson (2001). ††Micheneau et al. (2006). ‡‡Micheneau et al. (2008c). §§Micheneau et al. (in press). ¶¶Pollinators were identified from hairs found on orchid stigmas. ***Not clearly precised by the authors. N/A: Not applicable. © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 Figure 2. Illustrations of different pollinator groups. A, Hippotion celerio visiting flowers of Mystacidium capense (L.f.) Schltr. (South Africa, picture from S. D. Johnson). B, C, Angraecum striatum and its bird-pollinator Zosterops borbonicus (grey white-eyes) with pollinaria attached on its beak (pictures taken from videocaptures, Reunion, C. Micheneau and J. Fournel). D, Angraecum cadetii and its raspy cricket-pollinator Glomeremus sp. with pollinaria attached on its head (pictures taken from videocaptures with night-shot option, Reunion, C. Micheneau and J. Fournel). E, Illustration of a whole plant of A. cadetii with a visit of the grey white-eyes (note that the bird is a simple flower visitor and does not pollinate this orchid species; pictures taken from videocaptures, Reunion, C. Micheneau and J. Fournel). ORCHID POLLINATION © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 7 8 C. MICHENEAU ET AL. comparatively rich in long-tongued taxa, one species, Panogena lingens, is by far the most common pollinator of the Madagascan angraecoid species and the sole pollen vector of at least four orchid species (Table 1). This hawkmoth is widely distributed in Madagascar and shows a bimodal distribution in tongue length and width. Two distinct morphs have been recognized, the long- and slender-tongued morph with a mean proboscis length of c. 11–12 cm and the shorter-tongued morph with a mean proboscis length of c. 7–8 cm. The long-tongued morphs were often the only or most effective pollinators (Nilsson et al., 1987; Nilsson & Rabakonandrianina, 1988; see however, Wasserthal, 1997). In Africa, Agrius convolvuli and Coelonia fulvinotata were the two most common pollinators of Aerangis Rchb.f. and Rangaeris (Schltr.) Summerh. species (Fig. 2A, Martins & Johnson, 2007). On the island of Reunion, our studies conducted on the shortspurred angraecoid orchids, Angraecum striatum and A. bracteosum, revealed that the two white-eye species that occur on the island, the grey white-eye, Zosterops borbonicus, and the olive white-eye, Z. olivaceus, were both implicated in the pollination of these orchids: grey white-eyes are the regular pollinators of A. striatum and occasionally pollinate A. bracteosum and olive white-eyes are the regular pollinators of A. bracteosum, but have never been observed foraging on A. striatum flowers (Fig. 2; Micheneau et al., 2006, 2008c). Although around 50 bird families have been reported as flower visitors (Proctor et al. 1996), three major avian radiations have independently evolved a specialized nectar diet: hummingbirds (Trochilidae) in the New World, sunbirds (Nectariniidae) in the Old World (primarily Africa) and honeyeaters (Meliphagidae) in Australasia (e.g. van der Pijl & Dodson, 1966; Wolf, Hainsworth & Gill., 1975; Stiles, 1981; Proctor et al., 1996; Anderson, 2003). Although these three groups of birds have been widely reported as orchid pollinators (e.g. van der Pijl & Dodson, 1966; Rodríguez-Robles et al., 1992; Johnson, 1996; Singer & Sazima, 2000; Johnson & Brown, 2004), pollination of orchids by birds outside these three main nectarivorous groups (white-eyes in this case) was not previously known. The family Zosteropidae includes around 100 species of small arboreal songbirds (Gill, 1971; Slikas et al., 2000). These white-eyes are known for their high dispersal capabilities and have colonized more oceanic islands than any other passerine family (Moreau, 1964, cited in Gill, 1971). Although some species of white-eyes play an important role in plant pollination (Gill, 1971; Proctor et al. 1996; Kunitake et al., 2004), white-eyes typically have a generalist diet which includes fruit, insects and nectar (Gill, 1971). More surprisingly, the third short-spurred species of the Mascarene endemic A. section Hadrangis is regularly pollinated at night by a medium-size ortho- pteran species from the subtribe Gryllacridinae. This recent discovery further constitutes the first recorded case of regular pollination by orthopterans in extant angiosperms (Micheneau et al., in press) and Orthoptera are mostly known as a herbivorous insect order (see Micheneau et al., in press, for further references on flower–orthopteran interactions). Pollinator specificity The observation of only a few pollinating moths per orchid species, despite the high number of potential pollinators observed in the vicinity of the orchids, revealed a high degree of specialization between longspurred angraecoid orchids and their specific hawkmoth pollinators (e.g. Nilsson et al., 1987). Similarly, it seems that each of the two bird-pollinated orchid species is specialized for efficient pollination by only one bird species (Micheneau et al., 2006). This was unexpected, as bird-pollination in orchids generally involves more than one bird pollinator. For example, three hummingbird species have been reported pollinating Sacoila lanceolata (Aubl.) Garay (Spiranthinae Lindl. ex Meisn., Orchidoideae Eaton) in southern Brazil (Phaethornis eurynome, Leucochloris albicollis and female Thalurania glaucopis) (Singer & Sazima, 2000) and, although only one hummingbird species (Chlorostilbon maugaeus) was proposed as the pollinator of Comparettia falcata Poepp. & Endl. (Oncidiinae Benth., Epidendroideae) in Puerto Rico (the only hummingbird species found in the study area), another hummingbird (Amazilia tzacatl) was recorded as the pollinator of this orchid in Peru (Rodríguez-Robles et al., 1992). Satyrium carneum (Dryand.) Sims and S. princeps Bolus (Satyriinae, Orchidoideae) are often pollinated by two or more sunbird species in South Africa (Nectarina spp.) (Johnson, 1996) and, even in the unusual cases in Disa chrysostachya Sw. and D. satyriopsis Kraenzl. (Disinae Benth., Orchidoideae), in which sunbirds transfer pollen with their feet from flower to flower, at least two species of sunbirds (Nectarina famosa and Cinnyris talatala) are involved (Johnson & Brown, 2004). The lack of records of specialization in birdpollination systems in orchids may result from the facts that (1) many birds are generalist foragers and (2) pollination by birds is difficult to observe because of their furtive behaviour (van der Pijl & Dodson, 1966). Difficulties involved in making such observations mean that the incidence of orchid–bird interactions is likely to have been underestimated. Despite the fact that the flowers of Angraecum cadetii were frequently visited during both night and day by a range of flower visitors (mostly birds and nocturnal arthropods; see Micheneau et al., in press, for further details), only raspy crickets were observed carrying pollinaria and pollinating orchid flowers © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 ORCHID POLLINATION (Micheneau et al., in press). This suggested that this unexpected interaction involved a highly specialized pollination system between the orchid and the orthopteran. Specialization towards a pollinator group or a single specific pollinator is thought to maximize the efficiency of pollination, by reducing the risk that a generalist visitor wastes pollen in visiting foreign flowers. Especially in orchids, in which most species have their pollen packed in a unique structure, the pollinarium, that is exported in its entirety at one time by a single visitor, the loss of pollen constitutes a massive loss of the male sex function (Tremblay, 1992; Johnson, Neal & Harder, 2005; Harder & Johnson, 2008). However, if the reduction of the number of pollinators enhances the efficiency of pollination, it also results in reproduction being strongly limited by pollinators (Tremblay et al., 2005), making orchids more dependent on their pollinators than the reverse (Nilsson, 1992). Such asymmetrical pollination systems are common among angiosperms (Bascompte, Jordano & Olesen, 2006) and are particularly well illustrated in the angraecoid orchids. Although long-spurred angraecoid species are often specialized to their specific hawkmoth pollinators (P. lingens in particular), sphingids, in turn, are generalist foragers, opportunistically exploiting a wide variety of orchids and other plants. Asymmetrical pollination systems are also evident in bird–orchid interactions. Both white-eye species are extreme generalists on Reunion, foraging on various flowers and adding fruits, seeds and insects to their diet. In addition, the flowering time of Angraecum spp., during which birds may survive on orchid nectar, is limited (i.e. approximately 1.5 months during the wet season). POLLINATOR BEHAVIOUR In all study cases, the crepuscular activity of the hawkmoths observed foraging on long-spurred flowers was extremely limited, only taking place in the evening between 17:15 and 19:30 h, whether the observations were made in Africa (Luyt & Johnson, 2001; Martins & Johnson, 2007) or in Madagascar (Nilsson et al., 1985; Nilsson & Rabakonandrianina, 1988). At the same time, most observations were undertaken during these crepuscular hours (rarely after 20.00 h). Local weather conditions may affect hawkmoth foraging, which is not observed in heavy rain or strong winds (Martins & Johnson, 2007). The average duration of flower visits ranged from 1 to 7 s, lasting less than 2 s in most observations. Typically, hawkmoths, ‘smelling them in the breeze’, reached the orchids in a ‘zigzag flight upwind’, indicating a long-distance attraction by the strong scent emitted by the flowers (Nilsson et al., 1987), the intensity of 9 which became stronger at dusk. At a short distance, hawkmoths hovered in front of the flowers, easily distinguishable by their white colour, and inserted their unrolled proboscis into the spur, pressing their heads towards the orchid column while collecting the nectar. In the majority of species, pollinaria became attached to the dorsal surface of the proboscis at a distance from the base which varied both among and within species, depending on the relationship between the length of the proboscis and orchid spur, except for Angraecum arachnites Schltr. in which pollinaria were attached to the ventral base of the proboscis of Panogena lingens because of the orientation of the orchid lip, and Aerangis fuscata (Rchb.f.) Schltr. in which all pollinaria were attached to the head of the moth (Table 1, Fig. 3). Martins & Johnson (2007) noted that pollinaria were often attached to the areas of the head that were worn and thus lacked scales. A moth may carry pollinaria of different orchid species and the number of pollinaria found on the proboscis of a moth was also variable (Nilsson et al., 1987; Nilsson & Rabakonandrianina, 1988). As a maximum, Luyt & Johnson (2001) found 20 pollinaria of Mystacidium venosum within 0.6 cm of each other on a single proboscis of Nephele accentifera. Pollinaria attach firmly to the pollinators by their sticky viscidia, which adhere most effectively to the smooth surfaces of the proboscis instead of hairy zones of their heads (in contrast to the powdery pollen of most angiosperms). This firm fixation allows the pollinaria to remain attached to the bodies of the pollinators for long periods, which in turn allows the pollinators to disperse pollen over long distances and to ensure effective outcrossing (e.g. Nilsson et al., 1985, 1987; Nilsson & Rabakonandrianina, 1988; Nilsson, Rabakonandrianina & Pettersson, 1992). For instance, Wasserthal (1997) reported that a pair of undeposited pollinia of A. sesquipedale remained undamaged on the base of the tongue of a female Xanthopan morganii var. praedicta for almost 25 days (see also Luyt & Johnson, 2001, for experiments on pollen longevity). In addition to the firm adhesion, bending of pollinaria has also been reported to occur in the majority of long-spurred species studied. Typically, a few minutes after the pollinarium is removed from a flower, the stipes undergo a bending movement of 90° from their initial position, changing the orientation of the pollinia, which become parallel to the axis of the proboscis, and correctly orient to allow pollen to be deposited on conspecific stigmas. These bending movements of pollinaria are assumed to preclude geitonogamy (selfing between flowers of the same plants) (e.g. Darwin, 1862; Wasserthal, 1997; Johnson & Edwards, 2000; Luyt & Johnson, 2001; Peter & Johnson, 2006). White-eyes were active on flowers of Mascarene Angraecum spp. for the whole day from 06:02 h © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 10 C. MICHENEAU ET AL. Figure 3. Attachment distributions of pollinaria of angraecoid orchids on the proboscis of Panogena lingens. Aa, Angraecum arachnites; Ac, Angraecum compactum; Af, Aerangis fuscata; Jt, Jumellea teretifolia; Ng, Neobathiea grandidierana (modified from Nilsson et al., 1987). (earliest visit recorded) to 18.05 h (latest visit recorded), whether the weather was sunny or gloomy and rainy (Micheneau et al., 2006, 2008c; C. Micheneau & J. Fournel, unpubl. data). The two bird species, Z. olivaceus and Z. borbonicus, adopted the same behaviour while feeding on orchid flowers. Typically, they landed on a leaf or inflorescence and probed all ‘fresh-looking’ flowers within reach, moving around the inflorescence rachis if necessary. The time spent on orchids varied from 9 to 27 s (Micheneau et al., 2006). To reach the nectar in the spurs, the birds had to brush against the column and to pick up pollinaria as they retreated from the flowers. Pollinarium attachment on the beaks contrasted strongly between the two orchid species in relation to their floral morphology (i.e. flower opening and spur length). Foraging on A. striatum involved pollinarium attachment on both the upper and lower base of the beak (Fig. 2B), whereas pollinaria were always fixed at the extreme tip of the beak for A. bracteosum. Although birds may carry a large number of pollinaria, it was not rare to observe them cleaning their beaks on tree fern stipes or nearby hardwood species to remove their conspicuous packages. Raspy crickets were observed foraging on flowers of A. cadetii only during the darkest hours of night, between 19:50 and 04:40 h. In contrast to the furtive behaviour of both sphingids and birds, visits by these insular orthopterans could take a long time, up to 41 s for a single flower! ‘Typically, raspy crickets reached orchid flowers by climbing up the leaves or jumping across from neighbouring plants. In all successful pollination events, the raspy cricket positioned itself on the fleshy lip of the flower with its dorsal side orientated towards the orchid column and probed deep within the spur. Pollinaria of the orchid became stuck to the head of the crickets as they retreated from flowers’ (Micheneau et al., in press). POLLINATION AND REPRODUCTIVE SUCCESS Compilation of data from the literature indicated that pollination and fruiting success did not differ markedly between angraecoid orchids adapted to different pollinator groups, although pollination efficiency tends to be slightly higher in sphingid-pollinated flowers than in cases of bird-pollination (Fig. 4). Fruit set averaged approximately 17% for Madagascan– African moth-pollinated species, 13% for Mascarene bird-pollinated ones and 22% for Mascarene A. cadetii pollinated by orthopterans. Whatever the pollinator group, fruit set is however, close to the general mean of 17% reported for tropical orchids (Tremblay et al., 2005). Considering each pollinator group, fruit set resulting from hawkmoth pollination ranged from 3% for Aerangis confusa J.Stewart (Kenya, Martins and Johnson, 2007) to 41% for Angraecum arachnites (Madagascar, Nilsson et al. 1985) and averaged 17%. This mean value is much lower than the average of c. 45% calculated from data found in the literature (Tremblay et al., 2005) for tropical reward mothpollinated orchids (for comparison, fruit set reaches approximately 51% for temperate reward mothpollinated species; Tremblay et al., 2005). Reproductive success of bird-pollinated Angraecum spp. (c. 13%, Table 2) is also lower than the general median reported in the literature for bird-pollinated orchids (54%; Tremblay et al., 2005). However, this level of fruit set is close to the hummingbirdpollinated Comparettia falcata (c. 18%), which shares © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 ORCHID POLLINATION 11 Figure 4. Comparison of pollination and reproductive success between hawkmoth- and bird-pollinated angraecoid orchids. Mean (± SD) of pollinaria removal, pollinaria deposition and fruit set is presented. Data were compiled from Nilsson et al., 1985; Nilsson & Rabakonandrianina, 1988; Wasserthal, 1997; Luyt & Johnson, 2001; Martins & Johnson, 2007 for moth-pollinated species and from Micheneau et al., 2006, 2008c for bird-pollinated species. Table 2. Summary of floral features and reproductive success according to reproductive strategy Pollinator groups Hawkmoths* Birds† Orthopterans‡ Self-pollination§ Location Flower colour Madagascar White or greenish–white <5 4.6–33.3‡ Filiform, or spirally twisted Narrow 38.9 ± 57.1 (9) 15.2 ± 5.1 (12) Strong the evening Reunion White Reunion/Mauritius expected White/cream–white Reunion Cream–white >5 0.8–1.2 Conical <5 0.6 (mean) Conical 1 13.8 (mean) Filiform Large¶ 6.3 ± 2.0 (2) 9.7 ± 0.1 (2) Not detectable Very large 14.5 ± 13.7 mL (25) 12.3 ± 3.6 (21) Discrete (at night only) Narrow 6.1 ± 6.1 (9) 10.7 ± 1.3 (6) Not detectable Dominated by aromatics** 17.0 No volatile compound Dominated by (E)-b-ocimene at night 21.7 Indole at night Flowers/inflorescence Spur length (cm) Spur shape Spur opening Nectar volume (mL) % sugar (g/100 g) Floral scent (human nose) Floral scent composition Fruit set (%) 12.6 62.8 Mean ± SD (N) are presented for nectar properties. N corresponds to the number of species compiled from the literature, except for Orthopterans and self-pollination, for which N corresponds to the number of individuals sampled in the species Angraecum cadetii and Jumellea stenophylla, respectively. If variation were observed in nectar sugar concentration for a species, the highest value was chosen. *Compiled data from Nilsson et al. (1985, 1987); Nilsson & Rabakonandrianina (1988); Wasserthal (1997); Luyt & Johnson (2001); Martins & Johnson (2007) (study species are Angraecum arachnites, A. compactum Schltr., A. sororium, A. sesquipedale, Aerangis articulata (Rchb.f.) Schltr., A. brachycarpa (A.Rich.) Durand & Schinz, A. confusa, A. ellisii, A. fuscata, A. kotchyana, A. thomsonii, Jumellea teretifolia Schltr., Mystacidium venosum, Neobathiea grandidierana (Rchb.f.) Garay and Rangaeris amaniensis (Kraenzl.) Summerh). †Compiled data from Micheneau et al. (2006; 2008c) (study species are Angraecum bracteosum and A. striatum). ‡Data from Micheneau et al. (in press) (study species is Angraecum cadetii). §Data from Micheneau et al. (2008b) (study species is Jumellea stenophylla). ¶Angraecum bracteosum display a callus on the lip, which restricted flower’s entrance. **Kaiser (1993). © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 12 C. MICHENEAU ET AL. both an epiphytic habit and a tropical distribution (Rodríguez-Robles et al., 1992) with these Angraecum spp. Surprisingly, flower visits by orthopterans involved a high rate of fruiting success, averaging 22% at the study site of the Plaine des Palmistes in Reunion (average for 3 years). This rate is much higher than the 10% fruit set recorded for the bird pollinated sister species at the same study site and in the same years (Micheneau, 2005; Micheneau et al., 2008c). Reproductive success seems to be generally pollinator-limited in the angraecoid orchids (e.g. Wasserthal, 1997; Luyt & Johnson, 2001; Micheneau, 2005; Micheneau et al., 2006, 2008c). However, the spectacular rate of flower abortion found in Angraecum striatum (Micheneau et al., 2006) and the low rate of fruit set in Aerangis ellisii (Rchb.f.) Schltr. in spite of a high frequency of both pollen removal and deposition (Nilsson & Rabakonandrianina, 1988) suggest that in some species resource allocation may also affect reproductive success. FLOWER MORPHOLOGIES AND REPRODUCTIVE STRATEGIES Hawkmoth-pollinated flowers Long-spurred angraecoid orchids show a clear suite of floral adaptations to hawkmoth pollination, including floral traits involved in pollinator attraction (i.e. white flower and strong scent), pollinator fidelity (large sugar-rich nectar reward, situated in deep spurs easily accessible for long-tongued moths) and structural modifications to the column, the lip and/or the spur which all contribute to favourable contact between the head or proboscis of the moth and the orchid column and thus result in efficient pollinarium removal or deposition. The fact that in most species the lip lacks a landing platform is thought to facilitate pollination by hawkmoths, as they are able to probe nectar while hovering above the flowers without requiring any support from the flower (Luyt & Johnson, 2001). However, a landing platform may exist in some species, such as the very long-spurred Angraecum sesquipedale and A. sororium. Especially for these two species, the landing platform is thought to facilitate pollination by hawkmoths that swing– hover such as Xanthopan morganii praedicta and Coelonia solani (Wasserthal, 1997). In addition, particular shapes of lips may further function as a mechanical guide to orient the base of the proboscis into the spur (Nilsson et al., 1985; Luyt & Johnson, 2001). Specific adaptations of the rostellum may also extend over the mouth of the spur, flanking the entrance of the spur, presumably forcing the head of the pollinator into close contact with the orchid column (e.g. Aerangis elisii, Nilsson & Rabakonandri- anina, 1988; see also Wasserthal, 1997). Nectar production in the spur is another factor that has been viewed as being of crucial importance for successful pollination by hawkmoths (e.g. Darwin, 1862; Nilsson et al., 1985). In most flowers, the nectar is located only at the base of the spur, forcing the pollinator to probe deep into the spur to reach the nectar or ‘to drain the last drop’ (Darwin, 1862), so that its head makes the best contact with the orchid column to ensure successful pollen export and import (e.g. Darwin, 1862; Nilsson et al., 1985, 1987; Nilsson & Rabakonandrianina, 1988; Wasserthal, 1997; Martins & Johnson, 2007). In addition, spirally twisted spurs found in Aerangis thomsonii (Rolfe) Schltr., A. kotschyana (Rchb.f.) Schltr. and Angraecum arachnites may also serve identical purposes. Spirally twisted spurs involve tight frictions between orchid spur and hawkmoth tongue during nectar consumption, which may encourage moths to probe deeper into the spur and to stay longer in close contact with the reproductive organs of the orchid. Bird-pollinated flowers Although they are phylogenetically close to Malagasy species that perfectly match the sphingophilous pollination syndrome (Micheneau et al., 2008a), the Mascarene bird-pollinated Angraecum spp. show the usual convergent adaptations observed in other birdpollinated orchids. Except for the colour of the flower, which is pure white rather than vividly coloured as in typical bird-pollinated flowers, A. striatum and A. bracteosum display multi-flowered inflorescences of unscented and short-spurred flowers, which are rather fleshy in texture. In addition, A. bracteosum flowers have a strong fold (callus) on the lip, restricting the opening of the flower. This callus is a convergent feature often found in bird-pollinated orchids and may force the birds to push their beaks against the orchid column to reach the nectar in the spur (e.g. van der Pijl & Dodson, 1966; van der Cingel, 2001). Orthopteran-pollinated flowers Flower morphology of A. cadetii appears to be the result of a suite of floral adaptations for efficient pollination by raspy crickets (Micheneau et al., in press). Sepals and petals are quite small and fleshy; no damage was observed on these flower parts, suggesting that orthopterans do not usually predate the orchids (Micheneau et al., in press). The shape and the size of the pollinia (quite large, flat and triangular), are also atypical for the genus Angraecum, especially if compared with the small ball-like pollinia of the bird-pollinated species of A. section Hadrangis (Fig. 5). In addition, there is a close match in size between the head of the insect and the nectar-spur opening (Micheneau et al., in press; S. Hugel, unpubl. © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 ORCHID POLLINATION 13 Figure 5. Comparison of the shape and the size of pollinia between bird- and orthopteran-pollinated species within Angraecum section Hadrangis. A, Orthopteran-pollinated A. cadetii. B, Bird-pollinated A. bracteosum. data). Finally, the floral bouquet of A. cadetii (i.e. nocturnal emission of monoterpene-dominated scent; Micheneau et al., in press) probably plays a major role in attracting raspy crickets, which use their long antennae to explore their surroundings at night (C. Micheneau, pers. observ.; S. Hugel, pers. comm.). Self-pollinating flowers The study of different floral traits related to plant– pollinator interactions, namely flower longevity, spur length, nectar properties and scent production, have been investigated in the autogamous long-spurred Jumellea stenophylla (Frapp. ex Cordem.) Schltr., which is endemic to Reunion (Micheneau et al., 2008b). Results have shown that flower longevity of this self-pollinating species was much shorter (only 4–5 days) than that of hawkmoth-pollinated orchids (c. 25 days, Luyt & Johnson, 2001). This short flower lifetime was associated with autonomous self-pollinations that activated flower senescence. Although spur length of J. stenophylla (c. 13.8 cm) is within the reported spur size of long-spurred angraecoid orchids studied, the species only produces a relatively small amount of nectar (c. 6.1 mL) and flowers do not emit a strong and sweet scent at dusk (see Micheneau et al., 2008b for further details) This lack of floral scent, associated with the high degree of auto-pollinated flowers with a short lifespan, makes the flowers highly unattractive for pollinators and consequently precludes any chance of this orchid being cross-pollinated. EVOLUTION OF POLLINATOR-LINKED FLORAL TRAITS Nectar properties Nectar volume and sugar concentration in angraecoid orchids show clear patterns in relation to pollinator group. In hawkmoth-pollinated species, nectar sugar concentration averages 15.2%, whereas it is only 9.7% in bird-pollinated flowers (Table 2). The values for hawkmoth-pollinated angraecoid orchids are similar to those found in hawkmoth-pollinated plants more generally (Pyke & Waser 1981) and the dilute nectar found in angraecoid orchids conforms almost exactly to the pattern observed for other plants pollinated by short-billed opportunistic birds (Johnson & Nicolson, 2008). Plants pollinated by specialist flower feeding birds (hummingbirds and sunbirds) tend to produce nectar with a sugar concentration around 20%, whereas those pollinated by short-billed opportunistic birds tend to produce nectar with much lower concentrations around 10% (Johnson & Nicolson, 2008). Gradients in nectar sugar concentrations along the length of the nectar column were recently reported to occur within spurs in African Aerangis and Rangaeris spp. (Martins & Johnson, 2007). In all study species, nectar was significantly more diluted towards the mouth of the spur than at the tip (concentrations ranged from c. 1% at the proximal end of the spur to © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 14 C. MICHENEAU ET AL. c. 20% at the distal end). Nectar sugar gradients may have the same function as spirally twisted spurs and the presence of the nectar only in the lower part of the lip, i.e. they encourage the moth to go as deep as possible into the spur while probing nectar, enhancing at the same time the chance to ensure efficient pollination (Martins & Johnson, 2007). Post-pollination reabsorption of sugars from spurs has also been reported for two aerangoid orchid species (Koopowitz & Marchant, 1998; Luyt & Johnson, 2002) and this may mobilize resources for fruit development (Koopowitz & Marchant, 1998; Luyt & Johnson, 2002). The sugar composition of nectar of only one angraecoid species has been investigated so far (Mystacidium venosum, Luyt & Johnson, 2001). The nectar of this hawkmoth-pollinated orchid proved to be especially sucrose-rich (87% sucrose, 10% fructose and 3% glucose). It would be worthwhile in future research to develop further these comparative studies of nectar sugar composition, with the aim of investigating evolutionary patterns of this floral trait between birdand moth-pollinated species within the angraecoid orchid group (Johnson & Nicolson, 2008). In particular, one could then test the prediction that there should be an association between opportunistic bird pollinators and hexose-dominated nectar, based on the inability of these birds to digest simple sugars (e.g. Martínez del Rio, Baker & Baker, 1992). Long nectar spur evolution Darwin’s seminal work on the angraecoid orchid Angraecum sesquipedale triggered the beginning of a long debate about the evolution of long floral spurs. The two competing explanations for these traits are the Darwinian ‘coevolutionary race’ model (1862) and the ‘pollinator shift’ model developed by Stebbins (1970). The Darwinian ‘coevolutionary race’ model attributes adaptive elongation of the tongues of the moths and the spurs of the flowers to reciprocal coevolution, as a result of a gradual increase as a result of mutual benefit (gain in reproduction efficiency for the orchid and gain in nectar foraging efficiency for the hawkmoth). The alternative ‘pollinator shift’ model states that long tongues in hawkmoths were present before the long spurs of the orchids and evolved without any reciprocal interactions with them (Wasserthal, 1997; Whittall & Hodges, 2007). This model attributes the evolution of long nectar spurs to punctuated shifts to successive pollinators with longer tongues. Both models agree, however, with Darwin’s idea that longer spurs evolved under directionally selective pressure exerted by long-tongued pollinators. Darwin did not exclude the possibility that longer tongues in hawkmoth may have evolved from selection on body size ‘in relation to their conditions of life, either in the larval or mature state’ (Darwin, 1862) or in response to the spur length of other plants. Much more controversial has been Wasserthal’s (1997) assertion that the evolution of long tongues in hawkmoths was driven by selection imposed by flower predators, rather than availability of nectar in deep tubular flowers (e.g. Wasserthal, 1997; Nilsson, 1998a, b; Svensson, Rydell & Töve, 1998; Wasserthal, 1998). Because the interactions between angraecoid orchids and hawkmoths are mostly asymmetric (Nilsson & Rabakonandrianina, 1988), it seems likely that coevolution was diffuse at the guild level and not strictly pairwise. It also seems likely that many members of these guilds became adapted to longtongued hawkmoths via a pollinator shift rather than through coevolution. However, the relationships between long-spurred angraecoid orchids and their hawkmoth pollinators remain largely unknown if we consider the small number of species that has been studied (< 20 species) against the number of existing species (> 760 species in Angraecinae s.l., i.e. including Aerangidinae). We need to expand our knowledge to better assess the evolutionary outcomes between these orchids and their pollinators. In particular, finescale studies of geographical variation in mutualistic traits such as tongue length and nectar depth may reveal co-evolutionary processes at the population level (see Anderson & Johnson, 2008). However, much of the evolutionary potential of these orchids and their pollinators may already have been lost as a result of the massive destruction of primary forests in Madagascar and the large number of fragmented populations (i.e. reduction in population size and genetic diversity can have a severe effect on the evolutionary potential, e.g. Hubbell et al., 2008). CONCLUDING REMARKS Orchids should be studied more because they epitomize evolution in its most dynamic aspect, the rapid production of an incredibly diverse array of species (Chase et al., 2003). Orchids have been a model system for studying floral adaptation and diversification ever since Darwin’s influential book on the subject was published in 1862. They provide some of the most exceptional examples of highly specialized interactions between animals and plants. Not surprisingly, orchid pollination has been intensively studied relative to that in other extant angiosperms, with entire books dedicated to the subject (e.g. Darwin, 1862, 1877; van der Pijl & Dodson, 1966, van der Cingel, 1995, 2001), book chapters (e.g. Proctor et al. 1996, Roberts, 2003; see also Dressler, 1981, 1993) and numerous high-impact publications (e.g. Nilsson, 1983b, 1992; Romero & Nelson, © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 ORCHID POLLINATION 1986; Nilsson et al., 1992; Johnson & Nilsson, 1999; Schiestl et al., 1999, 2003; Cozzolino & Widmer, 2005; Liu et al., 2006; Ramírez et al., 2007). The unique, unexpected and diverse orchid pollination systems provide excellent opportunities for determining how pollinator-mediated selection has shaped the evolution of floral traits (e.g. flower scent, colour and spur length) and how shifts between pollinators may be responsible for patterns of macroevolution at the species level and above. However, in spite of the large size of this model family and the long history of orchid pollination biology, the identity and specificity of most orchid pollinators are unknown and remain inadequately studied, especially in the tropics where the family has undergone extensive diversification. Finally, highly specialized pollination interactions, such as those exemplified by orchids, are highly vulnerable to changes in populations and habitats. As Darwin pointed out while the pollinating sphingid of A. sesquipedale was still undiscovered: ‘If such great moths were to become extinct in Madagascar, assuredly the Angræcum would become extinct’ (Darwin, 1862). Once again, Darwin led the way in drawing this important connection between evolutionary studies and the ‘real-world’ problem of conservation. This is especially relevant today because of the present context of pollinator decline (Buchmann & Nabham, 1996; Kearns, Inouye & Waser, 1998; Biesmeijer et al., 2006; Ghazoul, 2006) as a result of habitat loss (Harris & Johnson, 2004; Aguilar et al., 2006) and climate change (Deutsch et al., 2008). ACKNOWLEDGEMENTS We warmly thank T. Pailler and J. Fournel (University of La Réunion) for their contributions to research involved in pollination biology of Mascarene angraecoid orchids. REFERENCES Ackerman JD. 1985. Pollination mechanisms of temperate and tropical orchids. In: Tan KW, ed. Proceedings of the eleventh world orchid conference. Miami: World Orchid Conference, 98–101. Ackerman JD. 1986. Mechanisms and evolution of fooddeceptive pollination systems in orchids. Lindleyana 1: 108– 113. Ackerman JD, Melendez-Ackerman E, Salguero-Faria J. 1997. Variation in pollinator abundance and selection on fragrance phenotypes in an epiphytic orchid. American Journal of Botany 84: 1383–1390. Ackerman JD, Montalvo AM. 1990. Short- and long-term limitations to fruit production in a tropical orchid. Ecology 71: 263–272. Ackerman JD, Rodríguez-Robles JA, Meléndez EJ. 1994. 15 A meager nectar offering by an epiphytic orchid is better than nothing. Biotropica 26: 44–49. Aguilar R, Ashworth L, Galetto L, Aizen MA. 2006. Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis. Ecology Letters 9: 968–980. Anderson B, Johnson SD. 2008. The geographical mosaic of coevolution in a plant–pollinator mutualism. Evolution 62: 220–225. Anderson SH. 2003. The relative importance of birds and insects as pollinators of the New Zealand flora. New Zealand Journal of Ecology 27: 83–94. Ayasse M, Schiestl FP, Paulus HF, Ibarra F, Francke W. 2002. Pollinator attraction in sexually deceptive orchid by means of unconventional chemicals. Proceedings of the Royal Society of London, Series B: Biological Sciences 270: 517–522. Barrett SCH. 1985. Floral trimorphism and monomorphism in continental and island populations of Eichhornia paniculata (Speng.) Solms. (Pontederiaceae). Biological Journal of the Linnean Society 25: 41–60. Barrett SCH. 1996. The reproductive biology and genetics of island plants. Philosophical Transactions of the Royal Society of London, Series B 351: 725–733. Bascompte J, Jordano P, Olesen JM. 2006. Asymmetric coevolutionary networks facilitate biodiversity maintenance. Science 312: 431–433. Biesmeijer JC, Roberts SPM, Reemer M, Ohlemüller R, Edwards M, Peeters T, Schaffers AP, Potts SG, Kleukers R, Thomas CD, Settele J, Kunin WE. 2006. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313: 351–354. Borba EL, Semir J. 1999. Temporal variation in pollinarium size after its removal in species of Bulbophyllum: a different mechanism preventing self-pollination in Orchidaceae. Plant Systematics and Evolution 217: 197–204. Borba EL, Semir J. 2001. Pollinator specificity and convergence in fly-pollinated Pleurothallis (Orchidaceae) species: a multiple population approach. Annals of Botany 88: 75– 88. Borba EL, Shepherd GJ, Semir J. 1999. Reproductive systems and crossing potential in three species of Bulbophyllum (Orchidaceae) occurring in Brazilian ‘campo rupestre’ vegetation. Plant Systematics and Evolution 217: 205–214. Borba EL, Shepherd GJ, van den Berg C, Semir J. 2002. Floral and vegetative morphometrics of five Pleurothallis (Orchidaceae) species: correlation with taxonomy, phylogeny, genetic variability and pollination systems. Annals of Botany 90: 219–230. Bosser J. 1987. Contribution à l’étude des Orchidaceae de Madagascar et des Mascareignes. XXII. Adansonia 3: 249– 254. Brys R, Jacquemyn H, Hermy M. 2008. Pollination efficiency and reproductive patterns in relation to local plant density, population size, and floral display in the rewarding Listera ovata (Orchidaceae). Botanical Journal of the Linnean Society 157: 713–721. © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 16 C. MICHENEAU ET AL. Buchmann SL, Nabham GP. 1996. The forgotten pollinators. Washington: Island Press. Bänziger H, Sun H, Luo Y-B. 2008. Pollination of the wild lady’s slipper orchids Cypripedium yunnanense and C. flavum (Orchidaceae) in south-west China: why are there no hybrids? Botanical Journal of the Linnean Society 156: 51–64. Carlsward BS, Whitten MW, Williams NH, Bytebier B. 2006. Molecular phylogenetics of Vandeae (Orchidaceae) and the evolution of leaflessness. American Journal of Botany 93: 770–786. Case MA, Bradford ZR. 2009. Enhancing the trap of lady’s slippers: a new technique for discovering pollinators yields new data from Cypripedium parviflorum. Botanical Journal of the Linnean Society 160: 1–10. Catling PM. 1990. Auto-pollination in the Orchidaceae. In: Arditti J, ed. Orchid biology, reviews and perspectives, V. Portland: Timber Press Inc, 121–158. Chase MW, Cameron K, Barrett RL, Freudenstein JV. 2003. DNA data and Orchidaceae systematics: a new phylogenetic classification. In: Dixon KW, Barrett RL, Cribb P, eds. Orchid conservation. Kota Kinabalu: Natural History Publication (Borneo), 69–89. Chase MW, Williams NH, Donisete de Faria A, Neubig KM, Amaral MCE, Whitten WM. 2009. Floral convergence in Oncidiinae (Cymbidieae; Orchidaceae): an expanded concept of Gomesa and a new genus Nohawilliamsia. Annals of Botany 104: 387–402. Cheng J, Shi J, Shangguan F-Z, Dafni A, Deng Z-H, Luo Y-B. 2009. The pollination of a self-incompatible, foodmimic orchid, Coelogyne fimbriata (Orchidaceae), by female Vespula wasps. Annals of Botany 104: 565–571. Cohen IM, Ackerman JD. 2009. Oeceoclades maculata, an alien tropical orchid in a Caribbean rainforest. Annals of Botany 104: 557–563. Coleman E. 1927. Pollination of the orchid Cryptostylis leptochila. Victorian Naturalist 44: 20–22. Correvon H, Pouyanne A. 1916. Un curieux cas de mimétisme chez les ophrydées. Journal de la Société Nationale d’Horticulture de France 17: 29–31, 41–42, 84. Cortis P, Vereecken NJ, Schiestl FP, Barone Lumaga MR, Scrugli A, Cozzolino S. 2009. Pollinator convergence and the nature of species’ boundaries in sympatric Sardinian Ophrys (Orchidaceae). Annals of Botany 104: 497–506. Cozzolino S, Widmer A. 2005. Orchid diversity: an evolutionary consequence of deception? Trends in Ecology and Evolution 20: 487–494. Dafni A. 1992. Pollination ecology: a practical approach. New York: Oxford University Press. Darwin C. 1859. The origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray Darwin C. 1862. On the various contrivances by which British and foreign orchids are fertilised by insects, and on the good effect of intercrossing. London: John Murray. Darwin C. 1875. The variation of animals and plants under domestication. London: John Murray. Darwin C. 1877. The various contrivances by which orchids are fertilised by insects, 2nd edn. London: John Murray. Deutsch CA, Tewksbury JJ, Huey RB, Sheldon KS, Ghalambor CK, Haak DC, Martin PR. 2008. From the cover: impacts of climate warming on terrestrial ectotherms across latitude. Proceedings of the National Academy of Sciences of the United States of America 105: 6668–6672. Devey DS, Bateman RM, Fay MF, Hawkins JA. 2009. Genetic structure and systematic relationships within the Ophrys fuciflora aggregate (Orchidaceae: Orchidinae): high diversity in Kent and a wind-induced discontinuity bisecting the Adriatic. Annals of Botany 104: 483–495. Dressler RL. 1981. The orchids, natural history and classification. Cambridge: Harvard University Press. Dressler RL. 1993. Phylogeny and classification of the orchid family. Portland: Dioscorides Press. Duffy KJ, Scopece G, Cozzolino S, Fay MF, Smith RJ, Stout JC. 2009. Ecology and genetic diversity of the denseflowered orchid, Neotinea maculata, at the centre and edge of its range. Annals of Botany 104: 507–516. Eckert CG, Samis KE, Dart S. 2006. Reproductive assurance and the evolution of uniparental reproduction in flowering plants. In: Harder LD, Barrett SCH, eds. The ecology and evolution of flowers. Oxford: Oxford University Press, 183–203. Ellis AG, Johnson SD. 1999. Do pollinators determine hybridization patterns in sympatric Satyrium (Orchidaceae) species? Plant Systematics and Evolution 219: 137–150. Fay MF, Chase MW. 2009. Preface. Orchid biology: from Linnaeus via Darwin to the 21st century. Annals of Botany 104: 359–364. Ghazoul J. 2006. Buzziness as usual? Questioning the global pollination crisis. Trends in Ecology and Evolution 20: 367– 373. Gigord LDB, Macnair MR, Smithson A. 2001. Negative frequency-dependent selection maintains a dramatic flower color polymophism in the rewardless orchid Dactylorhiza sambucina (L.) Soó. Proceedings of the National Academy of Sciences of the United States of America 98: 6253– 6255. Gill FB. 1971. Ecology and evolution of the sympatric Mascarene white-eyes, Zosterops borbonica and Zosterops oliveacea. The Auk 88: 35–60. Harder LD, Johnson SD. 2008. Function and evolution of aggregated pollen in angiosperms. International Journal of Plant Sciences 169: 59–78. Harris LF, Johnson SD. 2004. The consequences of habitat fragmentation for plant–pollinator mutualisms. International Journal of Tropical Insect Science 24: 29–43. Hedrén M, Nordström S. 2009. Polymorphic populations of Dactylorhiza incarnata s.l. (Orchidaceae) on the Baltic island of Gotland: morphology, habitat preference and genetic differentiation. Annals of Botany 104: 527–542. Hopper SD. 2009. Taxonomic turmoil down-under: recent developments in Australian orchid systematics. Annals of Botany 104: 447–455. Hubbell SP, He F, Condit R, Borda-de-Água L, Kellner J, ter Steege H. 2008. How many tree species are there in the © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 ORCHID POLLINATION Amazon and how many of them will go extinct? Proceedings of the National Academy of Sciences 105: 11498–11504. Jacquemyn H, Micheneau C, Roberts DL, Pailler T. 2005. Elevation gradients of species diversity, breeding system and floral traits of orchid species on Réunion Island. Journal of Biogeography 32: 1751–1761. Jersáková J, Johnson SD, Kindlmann P. 2006. Mechanisms and evolution of deceptive pollination in orchids. Biological Reviews 81: 219–235. Johnson SD. 1996. Bird pollination in South African species of Satyrium (Orchidaceae). Plant Systematics and Evolution 203: 91–98. Johnson SD. 1997a. Pollination ecotypes of Satyrium hallackii (Orchidaceae) in South Africa. Botanical Journal of the Linnean Society 123: 225–235. Johnson SD. 1997b. Insect pollination and floral mechanisms in South African species of Satyrium (Orchidaceae). Plant Systematics and Evolution 204: 195–206. Johnson SD, Brown M. 2004. Transfer of pollinaria on bird’s feet: a new pollination system in orchids. Plant Systematics and Evolution 244: 181–188. Johnson SD, Edwards TJ. 2000. The structure and function of orchid pollinaria. Plant Systematics and Evolution 222: 243–269. Johnson SD, Kurzweil H. 1998. Systematics and phylogeny of the Satyrium erectum group (Orchidaceae), with descriptions of two new species from the Karoo region of South Africa. Botanical Journal of the Linnean Society 127: 179– 194. Johnson SD, Linder HP, Steiner KE. 1998. Phylogeny and radiation of pollination systems in Disa (Orchidaceae). American Journal of Botany 85: 402–411. Johnson SD, Neal PR, Harder LD. 2005. Pollen fates and the limits on male reproductive success in an orchid population. Biological Journal of the Linnean Society 86: 175– 190. Johnson SD, Nicolson SW. 2008. Evolutionary associations between nectar properties and specificity in bird-pollination systems. Biology Letters 4: 49–52. Johnson SD, Nilsson LA. 1999. Pollen carryover, geitonogamy, and the evolution of deceptive pollination systems in orchids. Ecology 80: 2607–2619. Johnson SD, Steiner KE. 1997. Long-tongued fly pollination and evolution of floral spur length in the Disa draconis complex (Orchidaceae). Evolution 51: 45–53. Johnson SD, Steiner KE. 2000. Generalization versus specialization in plant pollination systems. Trends in Ecology and Evolution 15: 140–143. Jones CE, Little RJ. 1983. Handbook of experimental pollination biology. New York: Van Nostrand Reinhold Co. Kaiser R. 1993. The scent of orchids: olfactory and chemical investigations. Amsterdam: Elsevier. Kearns CA, Inouye DW. 1993. Techniques for pollination biologists. Niwot: University Press of Colorado. Kearns CA, Inouye DW, Waser NM. 1998. Endangered mutualisms: the conservation of plant–pollinator interactions. Annual Review of Ecology and Systematics 29: 83–112. 17 Koopowitz H, Marchant TA. 1998. Postpollination nectar reabsorption in the African epiphyte Aerangis verdickii (Orchidaceae). American Journal of Botany 85: 508. Kunitake YK, Hasegawa M, Miyashita T, Higuchi H. 2004. Role of seasonally specialist bird Zosterops japonica on pollen transfer and reproductive success of Camellia japonica in a temperate area. Plant Species Biology 19: 197–201. Larsen MW, Peter C, Johnson SD, Olesen JM. 2008. Comparative biology of pollination systems in the African– Malagasy genus Brownleea (Brownleeinae: Orchidaceae). Botanical Journal of the Linnean Society 156: 65–78. Li P, Luo Y-B, Deng Y-X, Kou Y. 2008. Pollination of the lady’s slipper Cypripedium henryi Rolfe (Orchidaceae). Botanical Journal of the Linnean Society 156: 491–499. Liu K-W, Liu Z-J, Huang L, Li L-Q, Chen L-J, Tang G-D. 2006. Pollination: self-fertilization strategy in an orchid. Nature 441: 945–946. Luyt R, Johnson SD. 2001. Hawkmoth pollination of the African epiphytic orchid Mystacidium venosum, with special reference to flower and pollen longevity. Plant Systematics and Evolution 228: 49–62. Luyt R, Johnson SD. 2002. Postpollination nectar reabsorption and its implications for fruit quality in an epiphytic orchid. Biotropica 34: 442–446. Martins DJ, Johnson SD. 2007. Hawkmoth pollination of aerangoid orchids in Kenya, with special reference to nectar sugar concentration gradients in the floral spurs. American Journal of Botany 94: 650–659. Martínez del Rio C, Baker HG, Baker I. 1992. Ecological and evolutionary implications of digestive processes: bird preferences and the sugar constituents of floral nectar and fruit pulp. Experientia 48: 544–551. Micheneau C. 2005. Systématique moléculaire de la soustribu des Angraecinae: perspectives taxonomiques et implications de la relation plantes-pollinisateurs dans l’évolution des formes florales. Unpublished D. Phil. Thesis, Université de La Réunion. Micheneau C, Carlsward BS, Fay MF, Bytebier B, Pailler T, Chase MW. 2008a. Phylogenetics and biogeography of Mascarene angraecoid orchids (Vandeae, Orchidaceae). Molecular Phylogenetics and Evolution 46: 908–922. Micheneau C, Fournel J, Gauvin-Bialecki A, Pailler T. 2008b. Auto-pollination in a long-spurred endemic orchid (Jumellea stenophylla) on Reunion Island (Mascarene Archipelago, Indian Ocean). Plant Systematics and Evolution 272: 11–22. Micheneau C, Fournel J, Humeau L, Pailler T. 2008c. Orchid–bird interactions: a case study from Angraecum (Vandeae, Angraecinae) and Zosterops (white-eyes, Zosteropidae) on Reunion Island. Botany 86: 1143–1151. Micheneau C, Fournel J, Pailler T. 2006. Bird pollination in an angraecoid orchid on Reunion Island (Mascarene Archipelago, Indian Ocean). Annals of Botany 97: 965–974. Micheneau C, Fournel J, Warren BH, Hugel S, GauvinBialecki A, Pailler T, Strasberg D, Chase MW. In press. Orthoptera, a new order of pollinator. Annals of Botany. © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 18 C. MICHENEAU ET AL. Mondragón-Palomino M, Theißen G. 2009. Why are orchid flowers so diverse? Reduction of evolutionary constraints by paralogues of class B floral homeotic genes. Annals of Botany 104: 583–594. Neiland MRM, Wilcock CC. 1998. Fruit set, nectar reward, and rarity in the Orchidaceae. American Journal of Botany 85: 1657. Nilsson LA. 1981. The pollination ecology of Listera ovata (Orchidaceae). Nordic Journal of Botany 1: 461–480. Nilsson LA. 1983a. Processes of isolation and introgressive interplay between Platanthera bifolia (L.) Rich. and P. chlorantha (Custer) Reichb. (Orchidaceae). Botanical Journal of the Linnean Society 26: 1–19. Nilsson LA. 1983b. Mimesis of bellflower (Campanula) by the red helleborine orchid Cephelanthera rubra. Nature 305: 799–800. Nilsson LA. 1988. The evolution of flowers with deep corolla tubes. Nature 334: 147–149. Nilsson LA. 1992. Orchid pollination biology. Trends in Ecology and Evolution 7: 255–259. Nilsson LA. 1998a. Deep flowers for long tongues: reply from L. A. Nilsson. Trends in Ecology and Evolution 13: 509– 509. Nilsson LA. 1998b. Deep flowers for long tongues. Trends in Ecology and Evolution 13: 259–260. Nilsson LA, Jonsson L, Ralison L, Randrianjohany E. 1987. Angraecoid orchids and hawkmoths in central Madagascar: specialized pollination systems and generalist foragers. Biotropica 19: 310–318. Nilsson LA, Jonsson L, Rason L, Randrianjohany E. 1985. Monophily and pollination mechanisms in Angraecum arachnites Schltr. (Orchidaceae) in a guild of long-tongued hawk-moths (Sphingidae) in Madagascar. Biological Journal of the Linnean Society 26: 1–19. Nilsson LA, Rabakonandrianina E. 1988. Hawk-moth scale analysis and pollination specialization in the epilithic Malagasy endemic Aerangis ellisii (Reichenb. fil.) Schltr. (Orchidaceae). Botanical Journal of the Linnean Society 97: 49–61. Nilsson LA, Rabakonandrianina E, Pettersson B. 1992. Exact tracking of pollen transfer and mating in plants. Nature 360: 666–668. Pansarin LM, de Moraes Castro M, Sazima M. 2009. Osmophore and elaiophores of Grobya amherstiae (Catasetinae, Orchidaceae) and their relation to pollination. Botanical Journal of the Linnean Society 159: 408–415. Parra-Tabla V, Vargas CF, Magaña-Rueda S, Navarro J. 2000. Female and male pollination success of Oncidium ascendens Lindey (Orchidaceae) in two contrasting habitat patches: forest vs. agricultural field. Biological Conservation 94: 335–340. Pauw A, Stofberg J, Waterman RJ. 2009. Flies and flowers in Darwin’s race. Evolution 63: 268–279. Peakall R. 1989. The unique pollination of Leporella fimbriata (Orchidaceae): pollination by pseudocopulating male ants (Myrmecia urens, Formicidae). Plant Systematics and Evolution 167: 137–148. Peakall R, Beattie AJ, James SH. 1987. Pseudocopulation of an orchid by male ants: a test of two hypotheses accounting for the rarity of ant pollination. Oecologia 73: 522–524. Peter CI, Johnson SD. 2006. Doing the twist: a test of Darwin’s cross-pollination hypothesis for pollinarium reconfiguration. Biology Letters 2: 65–68. Peter CI, Johnson SD. 2009. Reproductive biology of Acrolophia cochlearis (Orchidaceae): estimating rates of cross-pollination in epidendroid orchids. Annals of Botany 104: 573–581. Pouyanne A. 1917. La fécondation des Ophrys par les insectes. Bulletin de la Société d’Histoire Naturelle de l’Afrique du Nord 8: 6–7. Proctor M, Yeo P, Lack A. 1996. The natural history of pollination. Portland: Timber Press. Pyke GH, Waser NM. 1981. The production of dilute nectars by hummingbird and honeyeater flowers. Biotropica 13: 260–270. Ramírez SR, Gravendeel B, Singer RB, Marshall CR, Pierce NE. 2007. Dating the origin of the Orchidaceae from a fossil orchid with its pollinator. Nature 448: 1042–1045. Roberts DL. 2003. Pollination biology: the role of sexual reproduction in orchid conservation. In: Dixon KW, Barrett RL, Cribb P, eds. Orchid conservation. Kota Kinabalu: Natural History Publications (Borneo), 113–136. Rodríguez-Robles JA, Meléndez EJ, Ackerman JD. 1992. Effects of display size, flowering phenology, and nectar availability on effective visitation frequency in Comparettia falcata (Orchidaceae). American Journal of Botany 79: 1009–1017. Romero GA, Nelson CE. 1986. Sexual dimorphism in Catasetum orchids: forcible pollen emplacement and male flower competition. Science 232: 1538–1540. Rothschild LW, Jordan K. 1903. A revision of the lepidopterous family Sphingidae. Novitates Zoologicae 9 (Suppl.): 1–972. Salazar GA, Cabrera LI, Madriñán S, Chase MW. 2009. Phylogenetic relationships of Cranichidinae and Prescottiinae (Orchidaceae, Cranichideae) inferred from plastid and nuclear DNA sequences. Annals of Botany 104: 403–416. Salguero-Faría JA, Ackerman JD. 1999. A nectar reward: is more better? Biotropica 31: 303–311. Schiestl FP, Ayasse M, Paulus HF, Lofstedt C, Hansson BS, Ibarra F, Francke W. 1999. Orchid pollination by sexual swindle. Nature 399: 421–421. Schiestl FP, Peakall R, Mant JG, Ibarra F, Schulz C, Franke S, Francke W. 2003. The chemistry of sexual deception in an orchid–wasp pollination system. Science 302: 437–438. Schlechter R. 1926. Das System der Orchidaceen. Notizblatt der Botanischer Gartens zu Berlin 9: 590–591. Schlüter PM, Ruas PM, Kohl G, Ruas CF, Stuessy TF, Paulus HF. 2009. Genetic patterns and pollination in Ophrys iricolor and O. mesaritica (Orchidaceae): sympatric evolution by pollinator shift. Botanical Journal of the Linnean Society 159: 583–598. Singer RB. 2001. Pollination biology of Habenaria parviflora (Orchidacae: Habernariinae) in Southeastern Brazil. Darwiniana 39: 201–207. © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19 ORCHID POLLINATION Singer RB. 2002. The pollination mechanism in Trigonidium obtusum Lindl. (Orchidaceae: Maxillariinae): sexual mimicry and Trap-flowers. Annals of Botany 89: 157–163. Singer RB, Cocucci AA. 1999. Pollination mechanisms in southern Brazilian orchids which are exclusively or mainly pollinated by halictid bees. Plant Systematics and Evolution 217: 101–117. Singer RB, Flach A, Koehler S, Marsaioli AJ, MdCE A. 2004. Sexual mimicry in Mormolyca ringens (Lindl.) Schltr. (Orchidaceae: Maxillariinae). Annals of Botany 93: 755–762. Singer RB, Sazima M. 2000. The pollination of Stenorrhynchos lanceolatus (Aublet) L.C.Rich. (Orchidaceae: Spiranthinae) by hummingbirds in southern Brazil. Plant Systematics and Evolution 223: 221–227. Slikas B, Jones IB, Derrickson SR, Fleischer RC. 2000. Phylogenetic relationships of Micronesian white-eyes based on mitochondrial sequence data. The Auk 117: 355–365. Smithson A. 2002. The consequences of rewardlessness in orchids: reward–supplementation experiments with Anacamptis morio (Orchidaceae). American Journal of Botany 89: 1579–1587. Smithson A, Gigord LDB. 2001. Are there fitness advantages in being a rewardless orchid? Reward supplementation experiments with Barlia robertiana. Proceedings of the Royal Society of London. Series B, Biological Sciences 268: 1435–1441. Stebbins GL. 1970. Adaptative radiation of reproductive characteristics in angiosperms. I: pollination mechanisms. Annual Review of Ecology and Systematics 1: 307–326. Steiner KE, Withehead VB, Johnson SD. 1994. Floral and pollinator divergence in two sexually deceptive South African orchids. American Journal of Botany 81: 185–194. Stiles FG. 1981. Geographical aspects of bird–flower coevolution, with particular reference to Central America. Annals of the Missouri Botanical Garden 68: 323–351. Stpiczyńska M, Davies KL, Gregg A. 2007. Elaiophore diversity in three contrasting members of Oncidiinae (Orchidaceae). Botanical Journal of the Linnean Society 155: 135–148. Svensson MGE, Rydell J, Töve J. 1998. Deep flowers for long tongues. Trends in Ecology and Evolution 13: 460–460. Swarts ND, Dixon KW. 2009. Terrestrial orchid conservation in the age of extinction. Annals of Botany 104: 543–556. Tremblay R, Zimmerman JK, Lebron L, Bayman P, Sastre I, Alers-Garcia A. 1998. Host specificity and low 19 reproductive success in the rare endemic Puerto Rican orchid Lepanthes caritensis. Biological Conservation 85: 297–304. Tremblay RL. 1992. Trends in the pollination ecology of the Orchidaceae: evolution and systematics. Botany 70: 642– 650. Tremblay RL, Ackerman JD, Zimmerman JK, Calvo RN. 2005. Variation in sexual reproduction in orchids and its evolutionary consequences: a spasmodic journey to diversification. Biological Journal of the Linnean Society 84: 1–54. van den Berg C, Higgins WE, Dressler RL, Whitten WM, Soto-Arenas MA, Chase MW. 2009. A phylogenetic study of Laeliinae (Orchidaceae) based on combined nuclear and plastid DNA sequences. Annals of Botany 104: 417–430. van der Cingel NA. 1995. An atlas of orchid pollination: European orchids. Rotterdam: A. A. Balkema Publishers. van der Cingel NA. 2001. An atlas of orchid pollination: America, Africa, Asia and Australia. Rotterdam: A. A. Balkema Publishers. van der Pijl L, Dodson CH. 1966. Orchid flowers, their pollination and evolution. Coral Gables: University of Miami Press. Wallace AR. 1867. Creation by law. Quarterly Journal of Science 4: 470–488. Wallace AR. 1871. Contributions to the theory of natural selection, 2nd edn. London: Macmillan. Wasserthal LT. 1997. The pollinators of the Malagasy star orchids Angraecum sesquipedale, A. sororium and A. compactum and the evolution of extremely long spurs by pollinator shift. Botanica Acta 110: 343–359. Wasserthal LT. 1998. Deep flowers for long tongues. Trends in Ecology and Evolution 13: 459–460. West DA. 2003. Fritz Müller, a naturalist in Brazil. Blacksburg: Pocahontas Press. Whittall JB, Hodges SA. 2007. Pollinator shifts drive increasingly long nectar spurs in columbine flowers. Nature 447: 706–709. Willson MF. 1983. Plant reproductive ecology. New York: John Wiley & Sons. Wolf LL, Hainsworth FR, Gill FB. 1975. Foraging efficiencies and time budgets in nectar-feeding birds. Ecology 56: 117–128. Zimmerman JK, Aide TM. 1989. Patterns of fruit production in a neotropical orchid: pollinator vs. resource limitation. American Journal of Botany 76: 67–73. © 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 1–19