Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
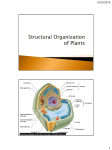
PLANT STRUCTURE AND GROWTH Chapter 35 There are about 280,000 species of flowering plants or angiosperms. This is the dominant group in terrestrial habitats. Our survival depends on them. Food, medicine, lumber, etc. The name angiosperm is derived from the Greek angeion, means vessel, and sperma means seed. The form of plants is controlled both by the environment and by their genetic makeup. Plants show plasticity, the ability to alter or mold itself in response to local environmental conditions. Plants also have a morphology, external form or external anatomy, controlled by their genotype. THE PLANT BODY A plant's structure reflects its interaction with the environment. Plants have become adapted through natural selection to specific problems posed by terrestrial environments. Plants have three basic organs: roots, stems and leaves. Differences between dicots and monocots. MONOCOTS Flower parts usually in multiples of three. Pollen with single furrow or pore. Embryo with single cotyledon. Major leaf veins parallel Stem vascular bundles scattered Roots are adventitious, fibrous system. Secondary growth absent DICOTS Flower parts usually in multiples of four or five. Pollen with three furrows or pores. Embryo with two cotyledons. Major leaf veins reticulated Stem vascular bundles in a ring Roots, taproot system. Secondary growth commonly present The plant body is divided into two main systems: a subterranean root system and an aerial shoot system. ROOT SYSTEMS Taproot system. Produced by gymnosperms and dicots. It consists of one large vertical root that grows downward and is called then a taproot. It produces lateral roots. This system generally penetrates deep into the soil. Fibrous root system. It is found in monocots. Adventitious roots develop from the stem. This system is usually shallow. The tip of the roots in both monocots and dicots is covered with root hairs. Root hairs are extension of the epidermal cells and increase the absorbing area of the root. Some plants have roots arising above ground from stems or leaves. These roots are said to be adventitious. FUNCTIONS OF THE ROOT 1. 2. 3. 4. 5. Anchorage. Absorption of water and minerals. Storage of food. Production of some hormones and secondary metabolites. Conduction of absorbed water and ions, and hormones and secondary metabolites. SHOOT SYSTEM: STEM AND LEAVES Shoot systems may be vegetative or reproductive. Stem FUNCTIONS 1. Support of the photosynthetic, reproductive and storage parts. 2. Conduction of water and metabolites. 3. Production of new stem tissues. EXTERNAL MORPHOLOGY (see fig. 35.13) 1. Terminal and lateral buds. Covered with bud scales while dormant. Contain meristem and produce primary tissues. Lateral buds are associated with leaf axils. Bud scale scars. 2. Nodes and internodes. Nodes are the regions of leaf attachment. Internodes are the space between two nodes. 3. Leaf scars and bundle scars. 4. Lenticels. Loosely arranged cells that allow gas exchange. Broken epidermis. The apical bud is partially responsible for inhibiting the growth of the lateral buds. This is called apical dominance. MODIFIED STEMS Stolons are slender stems growing along the surface of the ground that root and produce a plantlet, e.g. strawberry, Boston fern. Tubers are swollen ends of underground stems used in storage and reproduction, e.g. potato. Rhizomes are elongated horizontal underground stems bearing buds in the axils of reduced scale leaves, e.g. iris. Bulbs are small conical stems with numerous storage leaves attached to it, e.g. onion, garlic. Corms are thickened rounded storage stems covered with thin papery scales, e.g. crocus, gladiolus. LEAVES Leaves normally consist of a blade or lamina and a petiole or stalk. Leaves may have a simple or compound blade. Compound leaves have the lamina divided into leaflets. Most monocots have parallel veins that run the length of the leaf blade. Dicots have a network of veins. Most leaves are specialized for photosynthesis. CELL TYPES Parenchyma cells Living cells at maturity. Have thin primary walls. Function in storage, secretion and photosynthesis. Found throughout the body of the plant. Collenchyma cells Living cells at maturity. Have unevenly thickened primary cell walls. Function in support in flexible parts of the plant. Found in petioles, leaf veins and other parts of the plant that must be flexible. Sclerenchyma cells Have both primary and thickened secondary cell walls. Cells are often dead at maturity. Secondary wall with pits. Provide structural support. There are two types of sclerenchyma cells: sclereids and fibers. Sclereids may be living or dead at maturity. Short, cubical cells. Sclereid rich tissue may be hard and inflexible. Form part of the shell and pits of fruits, e.g. coconuts, walnuts, cherries, etc. Fibers are often dead at maturity. Long, tapered cells often in clumps. Have few pits in their secondary wall. Provide strength and elasticity. Found throughout the plant body, common in stems and some leaves. Water-conducting cells Sugar conducting cells TISSUES Each organ, - stem, leaf and root - has three tissue systems. The principal tissues of plants are grouped together into larger units called tissue systems based on their continuity throughout the plant body. Tissues are group of cells that are structurally and/or functionally distinct. Plant tissues composed of only one type of cell are called simple tissues, and those made of two or more types of cell are called complex tissues. Each tissue system is continuous throughout the plant body, but their arrangement varies in the different organs. 1. DERMAL TISSUE SYSTEM Composed of two complex tissues. Epidermis parenchyma cells guard cells Periderm cork cells cork cambium cork parenchyma Dermal tissue system is the outer protective covering of herbaceous plants and the young tender parts of woody plants. Epidermis usually consists of a single layer of parenchyma cells with guard cells and trichomes; secretes the waxy cuticle; gas exchange occurs through the stomata. It is made of parenchyma type cells. 2. VASCULAR TISSUE SYSTEM Composed of two complex tissues. Xylem tracheids vessel elements parenchyma cells fibers Phloem sieve tube members with sieve plates companion cells parenchyma cells fibers Vascular tissue system conducts materials throughout the plant body and provides support. Xylem conducts water and minerals from the roots to all parts of the plant; tracheids and vessel elements are the conducting cells and are dead at maturity; both have wall pits for lateral transport. Phloem transports sugars in solution to all plant parts; sieve tube members are the conducting cells; companion cells regulate the metabolism of the sieve tube members; both remain alive at maturity; sieve tube members lack nucleus and other organelles. 3. GROUND TISSUE SYSTEM Composed of three simple tissues found in the pith and cortex. Parenchyma tissue Collenchyma tissue Sclerenchyma tissue parenchyma cells collenchyma cells sclereids, fibers Ground tissue performs many functions: Photosynthesis, storage, secretion, flexible and rigid structural support Parenchyma cells have thin walls; polyhedral cells; function in photosynthesis, storage and secretion; remain alive at maturity. Collenchyma cells have an unevenly thickened wall; provide flexible structural support in soft non-woody organs; cells elongated; remain alive at maturity. Sclerenchyma cells have thick walls; provide rigid support to organs; cells usually die at maturity; fibers are long and tapered; sclereids are short and cubical. THE PROCESS OF PLANT GROWTH AND DEVELOPMENT Growth is irreversible increase in mass that results from cell division and cell expansion. Development is the sum of all of the changes that progressively elaborate an organism's body; all the changes from conception to maturity. MERISTEM Annuals are plants that complete their life cycle in one year or less. Biennials complete their life cycle in two years. Perennials live for many years. Growth in plants is localized in regions called meristems. Plants have indeterminate growth; they grow for as long as they live. Plants have perpetually embryonic cells capable of cell division. Growth involves cell division, cell elongation and cell differentiation. Some cells remain as part of the meristem and continue to divide. 1. Primary growth causes the roots and stems to elongate. It occurs in all plants. The apical meristem at the tip of roots and stems is responsible. 2. Secondary growth is an increase in stem and root girth. It occurs in woody plants only. It is due to the activity of the lateral meristems: vascular cambium and cork cambium. Vascular cambium forms a cylinder along the length of roots and stems, between the xylem and phloem; it produces more xylem and phloem. Cork cambium is located in the outer bark. Primary growth of roots The tip of the root is covered by a rootcap that produces mucigel. The rootcap is a thimblelike mass of living parenchyma cells that covers the apical meristem behind it and helps the root to penetrate the soil. As the root grows it pushed the rootcap forward and the peripheral cells are sloughed off. The sloughed off cells and the cap is covered with a slimy mucigel that lubricates the path of the root through the soil. New cells produced by the apical meristem are added to the cap to replace the ones that have been sloughed off. The apical meristem and the region around it where most or the cell division occurs is called the region or zone of cell division. Near the center of the apical meristem is the quiescent center, a group of cells that divide much more slowly that the other meristematic cells. These cells are a reserve to restore the meristem in case of damaged Above the apical meristem, cells form three concentric cylinders of cells that continue to divide for some time. These are the primary meristems that will produce the three primary tissue systems of the root: Protoderm: dermal tissue. Procambium: vascular tissue. Ground meristem: ground tissues. See fig. 35.14. The region of cell elongation is immediately behind the region of cell division. It usually s a few millimeters in length. The elongation of cells results in most of the increase in root length. There is no elongation beyond this area. Only one small portion of the root is pushed through the soil at a given time. In the region of maturation or differentiation is where most of the primary tissues mature. Root hairs are produced here. This region is also called the "root hair zone". There is gradual transition from one zone to the next. Some cells may begin to elongate and differentiate in the region of cell division whereas others do it the region of elongation. Primary tissues of the root. There are three tissue systems in the root: epidermis (dermal tissue system), cortex (ground tissue system) and vascular bundle or stele (vascular tissue system). The innermost layer of the cortex is the endodermis. The vascular cylinder consists of vascular tissue and the nonvascular pericycle. The pericycle originates from the procambium. The pericycle is made of parenchyma, which may develop secondary cell wall as they age. Lateral roots originate in the pericycle. Primary tissue of stems a) The shoot. The apical meristem of the shoot gives rise to protoderm, procambium, and ground meristem, which in turn develop into the three tissue stems. Axillary buds develop from groups of meristematic cells left by the apical meristem at the bases of the leaf primordia Growth of the shoot is due to the elongation of the cells below the apex within the internode. b) The stem There are three basic stem patterns: 1. Woody dicots form a continuous ring around the pith. The vascular bundle is surrounded by the cortex, e.g. Tilia, basswood 2. Herbaceous dicots for a ring of discrete vascular bundles around the pith. Gaps filled with ground tissue join the pith and the cortex, e.g. Sambucus, elderberry 3. Monocots have many vascular bundles scattered throughout the ground tissue, e.g. Zea, corn. These are the primary meristems that will produce the three primary tissue systems of the stem (same as in the root): Protoderm: dermal tissue. Procambium: vascular tissue. Ground meristem: ground tissues. LEAF TISSUES 1. Epidermis There are upper and lower epidermises that form the surface of the leaf. Made of living parenchyma cells. Lack chloroplasts. Covered with a waxy layer, the cuticle. Have small openings for gas exchange called stomata (sing. stoma). Guard cells flank each stoma. Trichomes or hairs may be present. 2. Mesophyll The photosynthetic tissue found in between the two epidermises is called mesophyll. It consist of... Made of living parenchyma cells. Abundant chloroplasts. Usually loosely arranged with many air spaces. Often arrange in two regions: palisade and spongy mesophyll. 3. Venation Veins and diffusion cooperate in the movement of materials in veins. Veins or vascular bundles extend through the mesophyll. Each vein contains xylem and phloem tissue. Xylem is usually restricted to the upper side of the vein, and phloem to the lower side of the vein. A non-vascular parenchymatous layer of cells called the bundle sheath surrounds veins. SECONDARY GROWTH Primary and secondary growth occurs simultaneously but in different regions of the plant. Secondary growth begins in portions of the root that are not elongating, in the area between the xylem and phloem. Woody dicot plants undergo secondary growth, an annual increase in width of stems and roots. It is the result of the activity of the vascular cambium and cork cambium. Secondary growth in monocots is very rare. Woody monocots increase their diameter by the enlargement of the primary tissue behind the apical meristem, e.g. palms. Vascular cambium is located between the xylem and the phloem. Secondary xylem (wood) is produced to the inside. Secondary phloem (inner bark) to the outside. Ray initials are cambium cells that produce rays of parenchyma cells. Fusiform initials produce new xylem and phloem. Rays are chains of parenchyma cells that radiate out of the center of the stem and transport materials laterally between the secondary xylem and secondary phloem Cork cambium is located near the surface of the stem. Cork cells are produced toward the outside. Cork cambium and cork make the periderm. Bark refers to all tissues external to the vascular cambium: secondary phloem, cork cambium and cork (phloem plus periderm). MECHANISMS OF PLANT GROWTH AND DEVELOPMENT Plants have a tremendous developmental plasticity. The plant body form - height, branching pattern, reproductive output - is influenced by environmental factors. A broad range of body forms can arise from the same genotype. Three processes responsible for development: growth, morphogenesis and differentiation. Morphogenesis: the development of body form and organization. Differentiation: modification in structure and function of the cells resulting in increased specialization. Growth The plane (direction) of cell division is an important determinant of plant form. The plane in which a cell will divide is determined during late interphase. The cytoskeleton takes certain orientation that will determine the plane of cell division. Microtubules in the outer cytoplasm become concentrated into a ring called the preprophase band. The band disappeared before metaphase but it has already set the future plane of cell division. An ordered array of actin microfilaments remains after the microtubules of the preprophase disappeared. These actin microfilaments hold the nucleus in a fixed orientation until the spindle forms. The microfilaments also direct the movement of the vesicles that will form the cell plate. The wall separating the two daughter cells form in the plane defined earlier by the preprophase band. The orientation of cell expansion: Growing plant cells expand through water intake. Small vacuoles that form in the intake of water coalesce to form the large central vacuole. Enzymes weaken the cross-links in the cell wall. The orientation of cell expansion is perpendicular to the orientation of the cellulose microfibrils in the wall. Cellulose microfibril correct orientation is essential in plant cell differentiation. The cellulose microfibrils are embedded in a matrix of polysaccharides, some of which form the cross-links between microfibrils. The cell secretes hydrogen ions that weaken the cross-links between polymers in the wall. The pattern of microfibrils mirrors the orientation of microtubules across the plasma membrane in the cytoplasm side of the membrane. According to one hypothesis... The plasma membrane has cellulose-synthesizing enzymes that can move in the fluid membrane. The microtubules direct the flow of these membranes in a given direction as they synthesized cellulose microfibrils. The microfibrils become locked in place by cross-linking to other microfibrils. Plant growth is directed by cortical microtubules located in the outer region of the cytoplasm. The entire process is genetically controlled. Morphogenesis Morphogenesis depends on pattern formation, the development of specific structures in specific locations. Development of tissues and organs in specific locations depends on the ability of cells to detect and respond to positional information. How cells detect their location is not known. One hypothesis states that cells detect a diffusing chemical from the outside and/or apical cells. This substance creates a gradient as it diffuses away from its source. The plant establishes its axial polarity in the zygote's first cell division. This is an asymmetrical division that establishes the position of the shoot and the root at both ends of the axis. Plants have master regulatory genes called homeotic genes that mediate many of the major events in an individual's development, such as the initiation of an organ. Differentiation Cellular differentiation depends on the control of gene expression. Specific genes are switched on or off at critical times in a cell's development. Different proteins are synthesized in different cells. The position of the cell in a developing organ determines how is going to differentiate. Internal or environmental cues may cause a plant to switch from one developmental phase to another, e. g. from development of juvenile leaves to development of mature leaves. Flower formation involves a phase change from vegetative growth to reproductive growth. Meristem identity genes control the transition from a vegetative shoot to a floral shoot. Once a shoot meristem is induced to flower, position information commits each primordium tat arises on the flanks of the shoot tip to develop into an organ of specific structure and function. Organ identity genes are regulated by positional information and produce transcription factors. They function in the development of the floral pattern, e. g. sepals form the first whorl, petals the next whorl, then anthers, etc. The ABC model of flower formation identifies how three classes of organ identity genes control formation of sepals, petals, stamens, and carpels.