Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
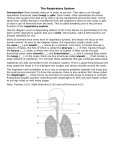
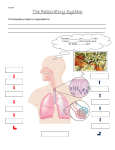
The Respiratory & Circulatory Systems Ch. 11-12 Introduction • To metabolize effectively and survive, cells within the body must replenish the oxygen they consume and remove the carbon dioxide they produce. • These are the responsibility of the respiratory and circulatory systems. Introduction • Most of the body’s energy needs are met by cellular oxidation of ingested foods. – In small animals the oxygen needed for this and the carbon dioxide produced can be carried between the external environment and the cells by diffusion, but this is a slow process and efficient only over short distances. • Diffusion still plays a role in the movement of gasses in larger animals, but a system is needed for the bulk flow, or movement by a muscular pump, of the medium containing the gases. • Some sort of a pump must move the medium containing the gases across a thin, moist and vascular membrane through which the gases can diffuse and be exchanged with those in the blood. – A membrane of this type is called a respiratory membrane. • Another pump, the heart, must move the blood through vessels that extend between the respiratory membrane and the vicinity of the cells, where diffusion again takes place. • An efficient respiratory system optimizes the diffusion and exchange of gases between the body and external environment, but many factors affect the nature of the respiratory membrane and the way it is ventilated. • High diffusion rates require a large surface area in the respiratory membrane and a short diffusion distance between the external medium and the blood. – Flow rates of the medium across the membrane, and of blood, must be slow enough for diffusion to occur; but fast enough to maintain the concentration gradients of the gases in the 2 mediums. • Ventilation rates or the amount of membrane in use, or both, are adjusted so that gas exchange matches the needs of the animal as the activity level varies. • The quantity of gas in the water or air also affects the size of the membrane and its rate of ventilation. • The density and viscosity of the medium affect the amount of energy required to move it across the respiratory membrane. • The movements of other diffusible molecules, such as water, salts, and nitrogenous wastes across the membrane may need to be reduced or enhanced. • Body size is an important factor because of its effect on surface-volume relationships. The respiratory System of Fishes • A fish’s respiratory system must be adapted to 2 major constraints of life in water. • First, the amount of O2 dissolved in water is much less than in air. • Second, water is much denser and more viscous than air. – Because of these two constraints, a fish must have a rather large respiratory surface and move large volumes of water across it at a considerable energy expense. • Any design feature of the respiratory system that reduces these energetic costs would be to the fish’s advantage. • Unloading of CO2 is less of a problem because it is highly soluble in water. – It does combine with water to form carbonic acid, but most fishes live in a large enough body of neutral (pH 7) water that this is of no consequence. • Several other problems arise from the close proximity of blood and water across the respiratory membrane. – Heat is quickly exchanged – Water, salts, and nitrogenous waste also diffuse across the membrane. Gills • As ancestral craniates became larger and more active gills, which provide a large surface area, evolved where a water current could ventilate them. – The larvae of many fishes have external gills, which are highly vascularized, filamentous processes with large surface areas attached to the lateral surface of the head between certain gill slits. – Adult fishes have internal gills, which consist of a large number of vascularized plates, the primary gill lamellae, attached to the walls of the gill or brachial pouches or to the gill arches. The Structure and Development of Internal Gills • The pharyngeal pouches develop from a lateral portion of the archenteron that will become the pharynx. – The plates of tissues between successive pouches, to which the primary gill lamellae attach, are called interbranchial septa; a skeletal visceral arch lies in each septa. • Skeletal, supporting gill rays usually extend peripherally from the visceral arches into the interbranchial septa or primary gill lamellae. – Muscles and nerves are associated with the skeletal elements • Each of the first 6 septa of a jawed fish contains an embryonic artery, the aortic arch, that supplies the gill. • Gill pouches were numerous in primitive fishes, some had up to 15 pairs. – Most chondrichthyans and early bony fish have 6 pairs of pouches, the first is reduced to a pair of spiracles. – Teleosts have lost the spiracle and now have only 5 pair of gill pouches. • Interbranchial septa that bear gill lamellae on both surfaces constitute a complete gill, or holobranch. – Jawed fishes usually have 4 of these • Only a small, gill-like structure lies in the spiracle, because it receives oxygenated blood from other gills it is called the pseudobranch. • Gill lamellae are seldom present on the posterior surface of the last gill pouch, because no aortic arteries exist here to supply them Lamprey Gills • Internal gills are arranged in different ways among fishes, and different patterns of ventilation exist. • Jawless fishes have large, saccular branchial pouches that are lined with primary gill lamellae. – These are called pouched gills. • Water is drawn into the pharynx through the mouth, enters the branchial pouches through pore shaped internal gill slits, and exits the pouches through pore shaped external gill slits. Elasmobranch Gills • The branchial pouches of chondrichthyan fishes are narrow chambers, and the gill lamellae are borne on the interbranchial septa, which continue to the body surface. – These are septal gills. • A vertically elongated internal gill slit leads from the pharynx into each branchial chamber. • Gill rakers at the base of the interbranchial septa keep food in the pharynx. • Blood and water flow in opposite directions through and across the secondary lamellae. – This countercurrent flow affords a considerably more efficient gas exchange than blood and water moving in the same direction, concurrently. Gills of Bony Fishes • Bony fishes developed a branchial apparatus that is somewhat different from that of elasmobranchs. – A flap of body wall supported by bones, known as the operculum, extends from the hyoid arch region of the head laterally and caudally over the gills. – There is a large, common opercular cavity for all the gills, and one valved, external gill slit. – The interbranchial septa are reduced, so the primary gill lamellae extend freely into the opercular cavity. • The gills are described a aseptal. • With the emergence of the opercular system, teleosts have a respiratory cycle that produces a continuous flow of water in one direction from the oropharynx to the opercular cavity over the gills. – The cycle constitutes 2 phases; the suction pump stage and force pump stage. • This produces an uninterrupted unidirectional flow of respiratory water that runs in the opposite direction of the blood flow on an optimized countercurrent system. Accessory Respiratory Organs • The gills of fishes are efficient respiratory organs that remove 80-90% of the available O2 from the water. • But in some situations the gills are not enough to meet the respiratory demands of the fish. – O2 levels can be very low in shallow, warm pools or in swamps where the decay of vegetation decreases O2 levels. • Many teleost fishes can supplement gill respiration with accessory respiratory organs. – Some have even become obligate air breathers – Virtually all bimodal breathers retain gills and simultaneously evolve a special air breathing organ. Bimodal Breathers • Some use vascular skin as a respiratory surface. • Others possess modifications of many parts of the gut: the lining of the mouth, pharynx, esophagus, intestine, and rectum. • There are also those that develop an outpocketing above the gills to form a suprabranchial air chamber, which can be filled with air and functions like a lung. Lungs • Many bony fishes have either lungs or swim bladders. – The organ is most lung-like in the primitive members of the group, so lungs seem to have appeared early in the evolution of fishes. • Actinopterygian fishes ventilate the lung with a characteristic 4 stroke buccal pump: – The buccal cavity expands so that spent air can pass into it. – Once filled, it compresses to expel the air. – The empty buccal cavity then expands again to take in fresh air – Finally it compresses to force the air into the lungs. • Lungs develop embryologically in lungfishes and amphibians as a ventral evagination of the floor of the digestive tract just caudal to the last pair of pharyngeal pouches • Suggests a homology between these structures. • All vertebrates with lungs have specialized cells that produce a surface film of lipoprotein known as surfactant. – The composition of the surfactant is such that it acts more as an “anti-glue,” preventing adhesion of adjacent epithelial surfaces during low lung volume. • When breathing air lungfishes come to the surface and inhales fresh air by expanding the buccal cavity. • The lungfish’s ventilation pattern is typically a 2 stroke pump, the buccal cavity is expanded and compressed only once. – Because mixed air is delivered to the lung, this pattern is called the mixed-air buccal pump system. • Lungfishes hold air in the lungs for a considerable time as the O2 is slowly used. – Long periods of breath holding, or apnea, alternate with short periods of lung ventilation. Swim Bladders • The lungs of actinopterygian fishes have transformed into swim bladders in neopterygian species and coelacanths. – Neopterygian fishes live in O2 rich waters where evolution would favor the conversion of lungs into a hydrostatic organ that would endow the fishes with neutral buoyancy. • The swim bladder of primitive actinopterygian fishes continues to be an important site of respiratory exchange, but it has also acquired a hydrostatic function. • In a teleost it is primarily a hydrostat that can be regulated, allowing the fish to attain neutral buoyancy and maintain position in the water with minimal energy expense. • Problems arise when fish change depth. – As it goes deeper the increased pressure will compress the swim bladder making the fish sink faster. – As the fish rises the bladder would expand, so air must be removed to maintain neutral buoyancy. • The gas in the swim bladder is 80% O2 and it is secreted into the swim bladder by a gas gland on one surface of the organ. – Often gas must be excreted against considerable concentration gradients because pressure increases rapidly at depth. • Oxygen is kept in the swim bladder by a rete mirabile – A set of long, parallel capillaries located just before the gas gland; together these form the red body. Respiration in Early Tetrapods • Because dry air at sea level contains 210mL/L O2, adult terrestrial vertebrates have far more oxygen available to them than do fishes, but they need a moist respiratory membrane because O2 must be in solution to diffuse into the blood. • Major problems for terrestrial verts are: 1. how to expose and ventilate the respiratory organ without an excessive loss of body water, and 2. how to prevent the collapse of the respiratory organ in air. • Lungs, which were inherited from fishes, are well adapted to meet these problems. • Air within the lungs contains a great deal of water vapor. • The respiratory surface is kept moist without a great deal of water loss because: – the rate of ventilation is low, – the air is conditioned by mucous glands in the airways prior to entering the lungs, – and not all the air in the lungs is exchanged in each breathing cycle. Amphibian Respiratory Organs • Amphibian larvae are aquatic and possess gills. • Salamanders retain external gills throughout their larval period and neotonic species retain them as adults. • The external gills of young tadpoles become covered by an opercular fold that opens to the body surface through a single pore, the spiracle. – The operculum is extensive, even covering the developing forelimbs. • Lungs develop and begin to function late in larval life. – Gills are lost at metamorphosis, the forelimbs push through the operculum, and the remains of the operculum fuse with the body wall. • Air passes through the buccopharyngeal cavity and enters the glottis. – The glottis leads to a small, triangular laryngotracheal chamber, from which the lungs emerge. • All of these passages are lined by cilia and by mucous secreting cells: – these secretions keep air moist and trap dirt particles and carry them away from the lungs. • Lungs in frogs are sac shaped organs, in salamanders they are elongated. • Ventilation of the lung in amphibians resembles that in lungfishes, except that air enters and leaved through the nares and nasal cavity. • The glottis then opens and air is expelled by elastic recoil of the lungs and by the contraction of flank muscles. • As in lungfish, amphibians can tolerate long periods of apnea alternated with short periods of ventilation. – Because their metabolic needs are low a single breath of air can last an amphibian for several minutes. • In addition to lungs, the thin, moist, vascular skin of most amphibians acts as a respiratory membrane and considerable cutaneous gas exchange occurs. – In species with both lungs and skin respiration, lungs are more important in O2 uptake and the skin is more important in the removal of CO2. • Cutaneous gas exchange exposes amphibians to considerable water loss. – Amphibians mitigate this effect by living in moist habitats or being most active in the more humid early morning and evening hours. The Reptile Respiratory System • Neck length is longer is reptiles and the primitive laryngotracheal chamber becomes divided into a larynx and trachea. • Cartilaginous rings or partial rings in the tracheal wall keep the trachea open for the free flow of air. • The horny skin of most reptiles makes the skin a poor respiratory surface and most gases are exchanged in the lungs. • The lungs of most reptiles are more compartmentalized and larger than those of amphibians. • A wide, central bronchus leads from the trachea into each lung, secondary bronchi branch from it, and alveolar sacs of varying size bud off them. • The structure of the lung varies greatly among different species and larger reptiles have greater compartmentalization than do the smaller reptiles. • Reptiles use an aspiration pump; during inhalation: – Contraction of intercostal muscles enlarge the pleuroperitoneal cavity in which the lungs lie. – Pressure within the cavity decreases to below atmospheric pressure and, as the nares are opened, the lung expands and air is sucked into them. – The glottis is then closed and the air is held until the next respiratory cycle. • Long periods of apnea occur. • Expiration occurs by the contraction of the rib muscles and smooth muscles of the lung wall. Respiration in Birds • Birds need an exceptionally efficient and compact respiratory system that will sustain a high level of metabolism and not significantly increase weight. • Bird lungs are relatively small organs that adhere to the dorsal wall of the pleural cavity and do not change appreciable in size during ventilation. • The lungs are small, but connect to a system of air sacs that pass among the viscera and even extend into many of the bones, this gives them a volume 2-3 times that of a comparable mammal. • The walls of the air sacs are not highly vascularized, so they do not participate in gas exchange. • Instead, the air sacs, together with a unique pattern of airways within the lung, make possible a unidirectional flow of air through the lungs. • This one-way flow carries relatively fresh air, with more O2 and less CO2 than in animals with a two way air flow. • The morphology of the avian respiratory system is complex. – The numerous air sacs can be grouped by function into a anterior and posterior set. – The trachea bifurcates into a pair of primary bronchi, each of which passes thorough the center of the lung in a mesobronchus, which connects with the posterior air sacs. – Air in the posterior sacs returns through mediodorsal bronchi, which connect with thousands of small parabronchi. – Innumerable short air capillaries bud off the parabronchi and connect with adjacent air capillaries – The air capillaries are interwoven into a respiratory labyrinth in which gas exchange occurs. • The lungs are ventilated by rocking movements of the sternum, which alternately expands and contracts the bellows-like air sack. • The glottis is held open most of the time, because the bird lung is continually ventilated. • Two separate inspirations and expirations are required to fully ventilate the lung, and no long periods of apnea occur. • Blood in the vascular capillaries flows transversely to the air flow in the parabronchi and capillaries in a pattern called cross-current flow. • Because air and blood flow across each other a gradient for the transfer of O2 exists at each crossing of the parabranchi and vascular capillaries. • The net result of this system, as in countercurrent flow, is that most of the available O2 is removed from the inspired air. Respiration in Mammals • The evolution of the secondary palate in mammals made it possible to separate food and respiratory passages. • The paired nasal cavities, which lie dorsal to the hard palate, are relatively larger than found in other verts. • Each contains 3 folds, or scrolls of bone called turbinates which increase their surface area. • The mucous membranes that cover the turbinates and line other respiratory passages are very vascular, ciliated, and contains mucous producing cells. • Considerable conditioning of the air occurs as it passes across these surfaces on its way to the lungs. – The air is warmed and moistened, and dirt is trapped in the mucous and carried to the throat to be swallowed or expectorated. • The nasal cavities connect through the choanae with the nasopharynx, which is separated from the oropharynx by the soft palate. • Food and air passages cross in the laryngopharynx – Food normally does not enter the respiratory passage because the larynx and hyoid apparatus are pulled forward to the base of the tongue during swallowing. • A trough-like fold, the epiglottis, flips back over the glottis or deflects food around it to the esophagus. • Except during swallowing, the glottis is held open because the mammalian lung is constantly ventilated. • The trachea continues down the neck and gives rise to the primary bronchi, which enter the lobes of the lungs. • The internal passages of the mammalian lung are more compartmentalized than those of amphibians and reptiles. • The airways within the lung branch, and rebranch at least 20 times, forming a respiratory tree that terminates in the bronchioles, alveolar sacs, and individual alveoli. – The walls become progressively thinner at each branch terminating in the alveoli with walls only a few cells thick. • A costal ventilation pump ventilates the mammalian lung. • Air is moved in and out through movement of the pleural cavities, alternately increasing and decreasing the pressure within the lungs relative to atmospheric pressure. • The increase in size occurs primarily through the contraction of the diaphragm. • During stronger inspiration, the intercostal muscles pull the ribs rostrally and increase the dimensions of the thorax. • Expiration is largely a passive process that relies of elastic recoil of the respiratory muscles. Vertebrate Circulatory Systems • The principal differences in the blood-vascular system of the vertebrates involves the separation of the heart into two separate pumps as vertebrates evolved from an aquatic life to a fully terrestrial life. – In fishes blood circulates in a single circuit, beginning at the heart, traveling to the gills, and then continuing to the body. – In terrestrial vertebrates there is a distinct division of blood flow to the lungs (pulmonary respiration) and to the body (systemic respiration. Pumps • An adequate circulatory system depends on one or more pumps (hearts) and on channels or conduits in which the blood can flow. • By wrapping muscle around a tube or channel it is possible to achieve a reduction of volume. • Two different type of pumps can be designed this way: peristaltic pumps, found mostly in invertebrates, and chamber pumps. • A chamber pump may have contractile walls (vertebrate heart) or external pressure from body parts can cause the reduction in volume (venous system of the human leg). Evolution of the Circulatory System Patterns of Circulation Fish Heart • A fish heart contains two main chambers in series, an atrium and a ventricle. – The atrium is preceded by a enlarged chamber, the sinus venosus, which collects blood from the venous system and assures smooth delivery of blood. • Elasmobranchs have a fourth chamber, the conus arteriosus, which dampens blood pressure oscillations before they reach the delicate blood capillaries. – Teleosts have a bulbous arteriosus that serves the same function. Evolution of the Heart with the Evolution of the Lungs • With the evolution of the lungs vertebrates developed a high-pressure double circulation: – A systemic circuit that provides blood to the body organs. – A pulmonary circuit that serves the lungs. • The beginning of this major evolutionary change probably resembled the condition seen in the lungfishes and amphibians. Terrestrial Vertebrate Heart • In modern amphibians the atrium is completely separated by a partition into two atria. – The right atrium receives venous blood from the body while the left atrium receives oxygenated blood from the lungs and skin. • The ventricle is undivided but venous and arterial blood remain mostly separated by a spiral fold of the conus arteriosus. • A septum partially separates the ventricle in most reptiles. Amphibian Heart Terrestrial Vertebrate Hearts Cont. • In crocodiles, birds, and mammals the septum completely divides the ventricle, providing a true separation of the arterial and venous systems of circulation. • Birds and mammals are endothermic and require about 10 times more energy than similar sized ectotherm. – This means they require 10 times more fuel, food and oxygen. Mammalian Heart Arteries • All vessels leaving the heart are called arteries. • To withstand the pressure generated during ventricular systole (contraction) the largest arteries close to the heart are invested with large layers of elastic fibers, very little smooth muscle, and tough inelastic connective tissue. • Arteries further away from the heart possess more smooth muscle and less elastic fibers. Arteries • All vessels leaving the heart are called arteries. • To withstand the pressure generated during ventricular systole (contraction) the largest arteries close to the heart are invested with large layers of elastic fibers, very little smooth muscle, and tough inelastic connective tissue. • Arteries further away from the heart possess more smooth muscle and less elastic fibers. Veins • Veins into which capillary blood flows for its return to the heart are thinner walled, less elastic, and of considerably larger diameter than their corresponding arteries. • Venus blood pressure is very low and therefore it must get assistance from valves in the veins, body skeletal muscles, suction created during diastole (expansion), and the rhythmic action of the lungs. Structural Relationship of Blood Vessels END