Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
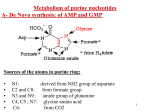
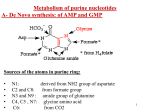
Nucleotide Metabolism Pyrimidine Met. Purine Met. Learning Objectives 1. How Are Purines Synthesized? 2. How Are Purines Catabolized? 3. How Are Pyrimidines Synthesized and Catabolized? 4. How Are Ribonucleotides Converted to Deoxyribonucleotides? 5. How Is dUTP Converted to dTTP? 6. Abnormal metabolism of uric acid. 7. Anticancer drugs targets Two types of pathways lead to nucleotides: the de novo pathways and the salvage pathways. De novo synthesis of nucleotides begins with their metabolic precursors: amino acids, ribose 5-phosphate, CO2, and NH3. Salvage pathways recycle the free bases and nucleosides released from nucleic acid breakdown. Salvage and de Novo Pathways The purine ring structure is built up one or a few atoms at a time, attached to ribose throughout the process. The pyrimidine ring is synthesized as orotate, attached to ribose phosphate, and then converted to the common pyrimidine nucleotides required in nucleic acid synthesis. Although the free bases are not intermediates in the de novo pathways, they are intermediates in some of the salvage pathways. de Novo Pathway for Purine Nucleotide Synthesis The synthesis of the purine ring is more complex. The only major component is glycine, which donates C-4 and C-5, as well as N-7. All of the other atoms in the ring are incorporated individually. C-6 comes from HCO3–. Amide groups from glutamine provide the atoms N-3 and N-9. The amino group donor for the inclusion of N-1 is aspartate, which is converted into fumarate in the process, in the same way as in the urea cycle . Finally, the carbon atoms C-2 and C-8 are derived from formyl groups in N10- formyl-tetrahydrofolate The Purine Ring System Is Assembled on Ribose Phosphate Glutamine phosphoribosyl amidotransferase catalyzes this reaction. De novo purine biosynthesis, like pyrimidine biosynthesis, requires PRPP, but for purines, PRPP provides the foundation on which the bases are constructed step by step. The initial committed step is the displacement of pyrophosphate by ammonia, rather than by a preassembled base, to produce 5-phosphoribosyl-1-amine, with the amine in the β configuration. de Novo Purine Biosynthesis Inosinate Formation Generating AMP and GMP Salvage Pathways Economize Intracellular Energy Expenditure Two salvage enzymes with different specificities recover purine bases. Adenine phosphoribosyltransferase catalyzes the formation of adenylate whereas hypoxanthine-guanine phosphoribosyltransferase (HGPRT) catalyzes the formation of guanylate as well as inosinate (inosine monophosphate, IMP), a precursor of guanylate and adenylate Pyrimidine Nucleotides Are Made from Aspartate, PRPP, and Carbamoyl Phosphate The common pyrimidine ribonucleotides are cytidine 5-monophosphate (CMP; cytidylate) and uridine 5-monophosphate (UMP; uridylate), which contain the pyrimidines cytosine and uracil. De novo pyrimidine nucleotide Biosynthesis proceeds in a somewhat different manner from purine nucleotide synthesis; the sixmembered pyrimidine ring is made first and then attached to ribose 5-phosphate. In the first step of the carbamoyl phosphate synthesis pathway, bicarbonate is phosphorylated by ATP to form carboxyphosphate and ADP. Ammonia then reacts with carboxyphosphate to form carbamic acid and inorganic phosphate. Carbamoyl phosphate reacts with aspartate to form carbamoylaspartate in a reaction catalyzed by aspartate Transcarbamoylase . Carbamoylaspartate then cyclizes to form dihydroorotate which is then oxidized by NAD+ to form orotate. Orotate reacts with PRPP to form orotidylate, a pyrimidine nucleotide. This reaction is driven by the hydrolysis of pyrophosphate. The enzyme that catalyzes this addition, pyrimidine phosphoribosyltransferase, Pyrimidine synthesis Carbamoyl phosphate reacts with aspartate to yield N-carbamoylaspartate in the first committed step of pyrimidine biosynthesis . This reaction is catalyzed by aspartate transcarbamoylase By removal of water from N-carbamoylaspartate, a reaction catalyzed by dihydroorotase, the pyrimidine ring is closed to form L-dihydroorotate. This compound is oxidized to the pyrimidine derivative orotate, a reaction in which NAD is the ultimate electron acceptor. Once orotate is formed, the ribose 5-phosphate side chain, provided once again by PRPP, is attached to yield orotidylate . Orotidylate is then decarboxylated to uridylate, which is phosphorylated to UTP. CTP is formed from UTP by the action of cytidylate synthetase. de Novo Pathway for Pyrimidine Nucleotide Synthesis. The C-2 and N-3 atoms in the pyrimidine ring come from carbamoyl phosphate, where as the other atoms of the ring come from aspartate. The pyrimidine ring is made up of three components: the nitrogen atom N-1 and carbons C-4 to C-6 are derived from aspartate, carbon C-2 comes from HCO3-, and the second nitrogen (N-3) is taken from the amide group of glutamine. Degradation of Purines and Pyrimidines Produces Uric Acid and Urea, Respectively Purine nucleotides are degraded by a pathway in which they lose their phosphate through the action of 5-Nucleotidase . Adenylate yields adenosine, which is deaminated to inosine by adenosine deaminase, and inosine is hydrolyzed to hypoxanthine (its purine base) and D-ribose. Hypoxanthine is oxidized successively to xanthine and then uric acid by xanthine oxidase, a flavoenzyme with an atom of molybdenum and four iron-sulfur centers in its prosthetic group. Molecular oxygen is the electron acceptor in this complex reaction. Purine Catabolism Purine bases are converted first into xanthine and then into urate for excretion. Xanthine oxidase catalyzes two steps in this process. Urate Crystals. Micrograph of sodium urate crystals. Joints and kidneys are damaged by these crystals in gout. Uric acid is the excreted end product of purine catabolism in primates, birds, and some other animals. A healthy adult human excretes uric acid at a rate of about 0.6 g/24 h; the excreted product arises in part from ingested purines and in part from turnover of the purine nucleotides of nucleic acids. In most mammals and many other vertebrates, uric acid is further degraded to allantoin by the action of urate oxidase. Lesch-Nyhan syndrome A genetic lack of hypoxanthine-guanine phosphoribosyltransferase activity, seen almost exclusively in male children, results in a bizarre set of symptoms . Children with this genetic disorder, which becomes manifest by the age of 2 years, are sometimes poorly coordinated and mentally retarded. In addition, they are extremely hostile and show compulsive self-destructive tendencies: they mutilate themselves by biting off their fingers, toes, and lips. Excess Uric Acid Causes Gout Long thought, erroneously, to be due to “high living,” gout is a disease of the joints caused by an elevated concentration of uric acid in the blood and tissues. The joints become inflamed, painful, and arthritic, owing to the abnormal deposition of sodium urate crystals. The kidneys are also affected, as excess uric acid is deposited in the kidney tubules. Gout occurs predominantly in males. Its precise cause is not known, but it often involves an underexcretion of urate. A genetic deficiency of one or another enzyme of purine metabolism may also be a factor in some cases. Gout is effectively treated by a combination of nutritional and drug therapies. Foods especially rich in nucleotides and nucleic acids, such as liver or glandular products, are withheld from the diet. Major alleviation of the symptoms is provided by the drug allopurinol , which inhibits xanthine oxidase, the enzyme that catalyzes the conversion of purines to uric acid. multiple tophi on the hands (Panel A), feet, knees, Some of the tophi exuded a white, chalky material. Laboratory studies were notable for a serum uric acid level of 8.5 mg per deciliter (506 µmol per liter), Xray hand:soft tissue swelling and pararticular erosions • The Gout-By James Gilray-1799 Johnson and Rideout NEJM, 350 (11): 1071, Figure 1 March 11, 2004 Control of Purine Biosynthesis. Feedback inhibition controls both the overall rate of purine biosynthesis and the balance between AMP and GMP production Many Chemotherapeutic Agents Target Enzymes in the Nucleotide Biosynthetic Pathways The first set of agents includes compounds that inhibit glutamine amidotransferases. Recall that glutamine is a nitrogen donor in at least half a dozen separate reactions in nucleotide biosynthesis. The binding sites for glutamine and the mechanism by which NH4 is extracted are quite similar in many of these enzymes. Most are strongly inhibited by glutamine analogs such as azaserine and acivicin Several Valuable Anticancer Drugs Block the Synthesis of Thymidylate One inhibitor that acts on thymidylate synthase, fluorouracil, is an important chemotherapeutic agent. Fluorouracil itself is not the enzyme inhibitor. In the cell, salvage pathways convert it to the deoxynucleoside monophosphate FdUMP, which binds to and inactivates the enzyme. Inhibition by FdUMP is a classic example of mechanism-based enzyme inactivation. Another prominent chemotherapeutic agent, methotrexate, is an inhibitor of dihydrofolate reductase. This folate analog acts as a competitive inhibitor; the enzyme binds methotrexate with about 100 times higher affinity than dihydrofolate. Methotrexate is a valuable drug in the treatment of many rapidly growing tumors, such as those in acute leukemia and choriocarcinoma, a cancer derived from placental cells. However, methotrexate kills rapidly replicating cells whether they are malignant or not. Anticancer Drug Targets END L26