Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
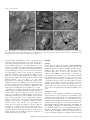
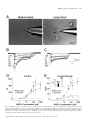
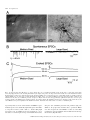
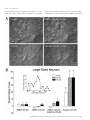
European Journal of Neuroscience, Vol. 14, pp. 1577±1589, 2001 ã Federation of European Neuroscience Societies Differential sensitivity of medium- and large-sized striatal neurons to NMDA but not kainate receptor activation in the rat Carlos Cepeda, Jason N. Itri, Jorge Flores-HernaÂndez, Raymond S. Hurst,* Christopher R. Calvert and Michael S. Levine Mental Retardation Research Center and Brain Research Institute, 760 Westwood Plaza NPI Room 58±258, UCLA School of Medicine, Los Angeles, CA 90024, USA Keywords: brain slices, electrophysiology, excitatory amino acid receptors, excitotoxicity, Huntington's disease Abstract Infrared videomicroscopy and differential interference contrast optics were used to identify medium- and large-sized neurons in striatal slices from young rats. Whole-cell patch-clamp recordings were obtained to compare membrane currents evoked by application of N-methyl-D-aspartate (NMDA) and kainate. Inward currents and current densities induced by NMDA were signi®cantly smaller in large- than in medium-sized striatal neurons. The negative slope conductance for NMDA currents was greater in medium- than in large-sized neurons and more depolarization was required to remove the Mg2+ blockade. In contrast, currents induced by kainate were signi®cantly greater in large-sized neurons whilst current densities were approximately equal in both cell types. Spontaneous excitatory postsynaptic currents occurred frequently in medium-sized neurons but were relatively infrequent in large-sized neurons. Excitatory postsynaptic currents evoked by electrical stimulation were smaller in large- than in medium-sized neurons. A ®nal set of experiments assessed a functional consequence of the differential sensitivity of mediumand large-sized neurons to NMDA. Cell swelling was used to examine changes in somatic area in both neuronal types after prolonged application of NMDA or kainate. NMDA produced a time-dependent increase in somatic area in medium-sized neurons whilst it produced only minimal changes in large interneurons. In contrast, application of kainate produced signi®cant swelling in both medium- and large-sized cells. We hypothesize that reduced sensitivity to NMDA may be due to variations in receptor subunit composition and/or the relative density of receptors in the two cell types. These ®ndings help de®ne the conditions that put neurons at risk for excitotoxic damage in neurological disorders. Introduction The striatum is composed primarily of medium-sized spiny projection neurons (Jiang & North, 1991; Gerfen, 1992). These cells make up > 95% of the total number of striatal neurons and they have been studied extensively. The other neuronal subtypes, medium- and largesized interneurons, are less numerous and because of their scarcity they have not been extensively studied from a functional perspective. Whilst there are several subpopulations of interneurons, one subtype, the large cholinergic neuron, has been the subject of much interest because it is the source of acetylcholine in the striatum (Bolam et al., 1984) and it has been implicated in neurological disorders (Vonsattel et al., 1985; Di Chiara et al., 1994). Until recently it has been dif®cult to study large cholinergic neurons electrophysiologically because they are scarce (Graveland & DiFiglia, 1985) and rarely sampled with standard in vivo or in vitro techniques (Wilson et al., 1990). The advent of infrared videomicroscopy and differential interference contrast optics (IR-DIC), allowing visual identi®cation of different cell types during brain slice Correspondence: Dr Michael S. Levine, as above. E-mail: [email protected] *Present address: Pharmacia Corporation, 7250±209±216, 301 Henrietta Street, Kalamazoo, MI 49007, USA Received 27 June 2001, revised 8 September 2001, accepted 26 September 2001 experiments (Dodt & ZieglgaÈnsberger, 1994), makes recording from these neurons an easier task (Kawaguchi, 1993; Bennett & Wilson, 1998, 1999). Brie¯y, in vivo these neurons display slow spontaneous ®ring and low amplitude short latency excitatory postsynaptic potentials (EPSPs) to cortical and thalamic stimulation. In vivo and in vitro large cholinergic interneurons are more depolarized than medium-sized projection neurons and show timedependent recti®cation (Wilson et al., 1990; Jiang & North, 1991; Kawaguchi, 1993). Neuronal inputs from the cerebral cortex appear to be considerably reduced compared to those on medium-sized neurons, whereas a prominent thalamic projection which is likely to be excitatory has been demonstrated (Lapper & Bolam, 1992). These large interneurons express multiple subtypes of glutamate receptors (Chen et al., 1996; Standaert et al., 1996). Excitatory postsynaptic currents (EPSCs) induced by local intrastriatal stimulation can be mediated by activation of both N-methyl-D-aspartate (NMDA) and non-NMDA receptors (Kawaguchi, 1992; Bennet & Wilson, 1998). Large cholinergic interneurons also express dopamine receptors (Bergson et al., 1995; Yan et al., 1997). Dopamine, via activation of D1 family receptors, produces an enhancement of excitability by suppression of K+ conductances and by modulation of the afterhyperpolarization (Aosaki et al., 1998; Bennet & Wilson, 1998). One important property of large cholinergic interneurons is that, unlike medium-sized spiny striatal neurons, they are spared from 1578 C. Cepeda et al. FIG. 1. (A) An IR image of a patch pipette attached to a large-sized striatal neuron (white arrow). A medium-sized striatal neuron is next to the large-sized neuron (white double arrows). (B and D) IR images of a medium- (B) and a large-sized (D) neuron. The same cells are shown in C and E after biocytin processing. Scale bar in A also applies to B and D; in E also applies to C. neurodegeneration in Huntington's disease, a genetically based neurodegenerative disorder (Ferrante et al., 1985; Kowall et al., 1987). Large cholinergic interneurons also may be less vulnerable to ischemic challenge (Chesselet et al., 1990; Calabresi et al., 1997; Centonze et al., 2001). It has been hypothesized that glutamate receptor excitotoxicity is responsible for cell death of medium-sized neurons in Huntington's disease because some of the neurochemical and neuropathological effects of this disorder are mimicked by application of glutamate receptor agonists (McGeer & McGeer, 1976; Beal et al., 1991). Potential explanations for sparing of large interneurons have implicated altered expression of normal or mutant huntingtin (Ferrante et al., 1997; Fusco et al., 1999) and/or hyporesponsiveness to glutamate-containing inputs compared to the medium-sized spiny neuron (DiFiglia, 1990). A previous study has provided some support for this explanation by demonstrating that large-sized striatal neurons are less responsive to glutamate receptor agonists than are medium-sized neurons (Calabresi et al., 1998). Our laboratory has studied extensively electrophysiological and synaptic responses of medium-sized striatal neurons to activation of excitatory amino acid receptors (Cepeda et al., 1991, 1993, 1998a; Levine et al., 1996; Hurst et al., 2001). In addition, in our studies of medium-sized neurons we have used cell swelling, an index of early signs of excitotoxicity (Choi, 1992), to examine functional characteristics of excitatory amino acid receptors and their modulation by dopamine and activation of metabotropic glutamate receptors (Colwell & Levine, 1996; Colwell et al., 1996; Cepeda et al., 1998b). The present study was designed to directly compare the responsiveness of medium- and large-sized striatal neurons to activation of excitatory amino acid receptors using several approaches, whole-cell voltage-clamp analysis of current responses, synaptic activation and cell swelling in response to application of excitatory amino acid receptor agonists. Methods Animals All procedures were carried out in accordance with the USPHS Guide for Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee at UCLA. SpragueDawley rat pups (12±18 days old; n = 60) were used in these experiments. The choice of this age range was based on our experience with NMDA receptor function as determined by electrophysiological experiments or by cell-swelling experiments from visualized cells in striatal slices (Cepeda et al., 1995, 1998a; Colwell et al., 1996, 1998; Cepeda et al., 1998b). This age represents a compromise between the presence of excitatory amino acid-evoked responses, and the ability to visualize cells which becomes more dif®cult in tissue from older rats. It is approximately the same age range used in recent studies analysing electrophysiological responsiveness of large-sized striatal interneurons (Bennett & Wilson, 1998, 1999). Preparation of slices Procedures for tissue preparation, visualization, electrophysiology and measurement of cross-sectional areas in cell-swelling experiments have been described (Cepeda et al., 1995; Colwell & Levine, 1996; Colwell et al., 1996; Cepeda et al., 1998b). Brie¯y, rats were anaesthetized with halothane and then killed by decapitation. After dissection, brains were placed in cold oxygenated arti®cial cerebrospinal ¯uid (ACSF1) containing (in mM) NaCl, 130; NaHCO3, 26; KCl, 3; MgCl2, 5; NaH2PO4, 1.25; CaCl2, 1.0; glucose, 10 (pH 7.2± 7.4). Transverse striatal sections were cut (350 mm) and placed in oxygenated (95%O2±5% CO2) ACSF2 (differing from ACSF1 as follows: CaCl2, 2 mM; MgCl2, 2 mM; lactate, 4 mM) at 25±27 °C for at least 1 h, and then transferred to a perfusion chamber attached to ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 NMDA responses in striatal neurons 1579 the ®xed-stage of an upright microscope (Zeiss Axioskop, Thornwood, New York, USA) in which the slice was submerged in continuously ¯owing oxygenated ACSF2 (25 °C, 4 mL/min, lactate removed). Cells were visualized with a 403 water-immersion objective, illuminated with near IR light (790 nm, Ealing Optics, Holston, MA, USA) and the image detected with an IR-sensitive CCD camera. Digital images were stored for subsequent analysis when necessary. Cells were typically visualized from 30 to 100 mm below the surface of the slice. It was relatively easy to distinguish medium- and large-sized cells. The somatic area of medium-sized cells was » 100 mm2 whereas the somatic area of large cells was typically > 150 mm2 (Fig. 1). Whole-cell voltage-clamp Patch electrodes (3±6 MW) were ®lled with one of the following internal solutions, depending on the purpose of the experiment, (in mM): K-gluconate, 140; HEPES, 10; MgCl2, 2; CaCl2, 0.1; ethylene glycol-bis(b-aminoethyl ether)-N,N,N¢,N¢-tetraacetic acid (EGTA), 1.1; and K2ATP 2 or Cs-methanesulphonate, 125; NaCl, 4; KCl, 3; MgCl2, 1; MgATP, 5; EGTA, 9; HEPES, 8; GTP, 1; phosphocreatine, 10 and leupeptin, 0.1 (pH 7.25±7.3; osmolality 280±290 mOsm). Unless otherwise noted, tetrodotoxin (1 mM) was added to the external solution to block Na+ currents after the whole-cell con®guration was obtained. In experiments in which responses to bath application of excitatory amino acid receptor agonists during injection of ramp command voltages were examined, a CsF-based solution was used in order to block voltage-gated Ca2+ currents. It contained (in mM): CsF, 125; NaCl, 4; KCl, 3; MgCl2, 1; EGTA, 9; HEPES, 8; and K2ATP, 5. To block K+ currents, tetraethylammonium (20 mM) was in the external solution. Axopatch 200A or 1D ampli®ers were used for voltage-clamp recordings. A 3-M KCl±agar bridge was inserted between the extracellular solution and the Ag±AgCl indifferent electrode. Tight seals (2±10 GW) from visualized large- and medium-sized cells were obtained by applying negative pressure. The membrane was disrupted with additional suction and the whole-cell con®guration was obtained. The access resistances ranged from 8 to 15 MW. Series resistances were compensated 60±85%. Cell capacitance measurements as well as somatic area were used to distinguish between large- and medium-sized neurons. Once the whole-cell con®guration was obtained, the capacitance of the cell was calculated by using a 10-mV hyperpolarizing pulse (40 ms duration). In experiments in which excitatory amino acid receptor agonists were bath-applied, the magnitude of the current was examined in response to a ramp. The ramp consisted of a voltage command from ±70 to ±90 mV over 0.4 s, followed by a ramp to +40 mV over 6.4 s to ensure that unblocked or poorly blocked Ca2+ channels were inactivated. At +40 mV, the holding potential was followed by a ramp command to ±90 mV over 0.9 s. Current responses in the absence of excitatory amino acid receptor agonists were subtracted from responses in the presence of bath-applied NMDA or kainate to isolate speci®c agonist-induced currents. Isolated currents were plotted against voltage to determine current±voltage relationships (Figs 2 and 3). Currents were converted to current density by dividing by cell capacitance to normalize responses between the different cell sizes to better permit comparison of data across experimental conditions (Alzheimer et al., 1993). Both upward (±90 to +40 mV) and downward (+40 to ±90 mV) ramps were used to generate current±voltage plots. Currents obtained from the upward and downward ramps were similar and the downward ramp was used when data were quanti®ed (Burgard & Hablitz, 1994). NMDA (100 mM) and kainate (100 mM) were bath-applied for 3 min prior to applying the ramp protocol. Each application of either NMDA or kainate was followed by a 5±10 min washout. We used kainate instead of a-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) because large interneurons have been shown to express kainate but not AMPA receptor subunits (Chen et al., 1996), although a more recent single-cell analysis indicated that AMPA receptor subunit proteins are expressed (Richardson et al., 2000). Whole-cell voltage-clamp data, especially quantitative estimates of currents from neurons in slices, should be interpreted with caution because the currents measured at the soma are undoubtedly distorted as a result of the nonisopotentiality over the neuronal surface due to space-clamp limitations (Armstrong & Gilly, 1992). To partially control for this problem we also examined responses in acutely dissociated neurons. Electrical stimulation A glass pipette ®lled with the external solution was placed » 200 mm from the recording pipette. Single pulses (200 msec duration) were delivered every 10±20 s. The stimulus intensity was adjusted to 23 the threshold to evoke synaptic responses. Four to six pulses were applied at different holding potentials and averaged. Excitatory postsynaptic currents (EPSCs) mediated by activation of glutamate receptors were isolated by adding bicuculline methiodide (10 mM), a GABAA receptor blocker, to the bath solution. Acute neuron dissociation Some slices also were used for acute dissociation of neurons. Slices were incubated for 1±6 h at room temperature in NaHCO3-buffered saline bubbled with 95% O2/5% CO2 (in mM except where otherwise noted): NaCl, 126; KCl, 2.5; MgCl2, 2; CaCl2, 2; NaHCO3, 26; Na2HPO4, 1; pyruvic acid, 1; glutathione 5 mM; NG-nitro-L-arginine, 1; kynurenic acid, 1; glucose, 10; HEPES, 15; pH 7.4 with NaOH, 300±305 mOsm/L. After 1 h incubation, a slice was placed in lowCa2+±isethionate solution (in mM): Na isethionate, 140; KCl, 2; MgCl2, 4; CaCl2, 0.1, glucose, 23; HEPES, 15; pH 7.4, 300± 305 mOsm/L, and the dorsal striatum was dissected and placed in an oxygenated cell-stir chamber (Wheaton, Inc., Millville, NJ USA) containing papain (Calbiochem, La Jolla, CA USA; papain, 1±2 mg/ mL) in HEPES-buffered Hanks' balanced salt solution (HBSS, Sigma Chemical Co., St. Louis, MO, USA) at 35 °C. After 20±40 min of enzyme digestion, tissue was rinsed three times with the low-Ca2+± isethionate solution and mechanically dissociated with a graded series of ®re-polished Pasteur pipettes. The cell suspension was plated into a 35-mm NUNCLON dish containing HEPES-buffered HBSS saline on the microscope stage. Whole-cell recordings from dissociated neurons used standard techniques (Hamill et al., 1981; Bargas et al., 1994). The internal solution consisted of (in mM): N-methyl-D-glucamine, 180; HEPES, 40; MgCl2, 2; EGTA, 10; phosphocreatine, 12; Na2ATP, 2; Na3GTP, 0.2; leupeptin, 0.1; pH 7.2±7.3 with H2SO4, 265±270 mOsm/L. The external solution consisted of (in mM): NaCl, 135; CsCl, 20; BaCl2, 5; glucose, 10; HEPES, 10; tetrodotoxin, 0.001; glycine, 0.02; pH 7.3 with NaOH, 300±305 mOsm/L. Recordings from acutely dissociated neurons were obtained with an Axopatch 200A ampli®er (Axon Instruments, Foster City, CA, USA). Electrode resistance was 2±4 MW in the bath. After seal rupture, series resistance (4±10 MW) was compensated (70±90%) and periodically monitored. Recordings were made only from medium- or large-sized cells that had short (< 75 mm) proximal dendrites. NMDA (1, 10, 50, 100, 200 or 1000 mM, 3 s duration every 20 s) was applied with a gravity-fed `two-pipe' system. The array of application capillaries (» 150 mm i.d.) was positioned a few hundred mm from the ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 1580 C. Cepeda et al. FIG. 2. Responses of medium- and large-sized neurons to NMDA application (100 mM). (A) Current±voltage plots for typical medium- and large-sized neurons. A ramp command was used to change voltage from ±90 to +40 mV and back to ±90 mV. The plotted data were obtained from the downward ramp. Control current±voltage plots were subtracted from plots in the presence of NMDA. (B) Current±voltage plots (mean 6 SEM) for medium- and large-sized neurons. (C) Current density±voltage plots for medium- and large sized neurons. (D) Conductance±voltage plots for medium- and large-sized neurons. Drawn curves represent best ®t functions according to the formula in the Results. In B, C and D asterisks indicate group differences were statistically signi®cant (P < 0.05). See Results for further details. cell under study. Solution changes were effected by changing the position of the array with a DC drive system, controlled by a SF-77B perfusion system (Warner Instruments Co., Hamden, CT, USA) synchronized by pClamp. Solutions could be delivered within 5± 10 ms with this system. Cell identi®cation In most experiments, electrodes were ®lled with 0.2% Biocytin (Sigma, St Louis, MO, USA) in the internal solution. After the experiment, the slice was ®xed in 4% paraformaldehyde overnight, then processed according to published protocols (Horikawa & Armstrong, 1988). Cell swelling Preparation of brain slices for these experiments were the same as those described for electrophysiology except tetrodotoxin and tetraethylammonium were omitted from the bath. After equilibration for 10 min, a baseline image of the cells was obtained and stored. The slice was then exposed to different solutions depending upon the purpose of the speci®c experiment. Images were obtained and stored at 5-min intervals for the duration of each experiment (15 min exposure to experimental treatment). In order to quantify changes in response to excitatory amino acid receptor agonists, cross-sectional somatic area was measured at each 5-min time point, before and during experimental treatments. Each measurement was made two or three times on each image and the average value recorded for each cell at each time point. For each experimental group, data were obtained from several animals. Typically, 3±7 medium-sized cells were visualized within each slice whilst large cells were sparse so that usually only one cell could be analysed in each slice. In slices containing large-sized neurons, neighbouring medium-sized cells were also measured. Cross-sectional areas at each time point were converted to percentage change with respect to the baseline area for each cell. For statistical analyses, data were pooled for all cells in experimental and control groups. Concentrations of NMDA (50 and 100 mM) or kainate (100 mM) were based on our previously published studies (Colwell & Levine, 1996; Cepeda et al., 1998b). Drugs were applied in the bath and freshly prepared each day. Statistics Values in the tables, ®gures and text are presented as means 6 SEM. Differences among group means were assessed with appropriate ttests or analyses of variance (ANOVAs). Welch's approximation to the t-test for unequal variances was used when group variances were not homogeneous (Welch, 1947). For post hoc evaluations using ANOVAs, the Bonferroni t-test was used because this test is one of the more conservative approaches using multiple comparisons. In the text, only P-values and the type of test used are reported. ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 NMDA responses in striatal neurons 1581 Results Neuron identi®cation Both medium- and large-sized neurons could be visualized easily in the slices (Fig. 1). Cross-sectional areas were signi®cantly smaller for the medium-sized neurons (Fig. 1, compare B vs. D and C vs. E) [103 6 5 mm2 (n = 15) vs. 327 6 13 mm2 (n = 21) for medium- and large-sized neurons, respectively; t-test, P < 0.001]. Dendritic processes were examined after ®lling the neuron with biocytin. Mediumsized neurons typically displayed dendritic ®elds with spine-like varicosities (Fig. 1C), although spines were not abundant due to the young age of the rats (Hattori & McGeer, 1973; Tepper et al., 1998). Large-sized neurons exhibited dendritic branches with or without scarce varicosities (Fig. 1E). Although several types of large neurons have been described in striatum, the present study targeted the very large, putative cholinergic, interneurons (Kawaguchi, 1993). These cells more probably correspond to the so-called `Type 1' large neurons (Bolam et al., 1984). Because we did not identify the large cells immunohistochemically, we use the term large-sized neurons rather than cholinergic interneurons to describe these cells. NMDA- and kainate-induced currents during ramp voltage commands Whole-cell voltage-clamp recordings were obtained from 30 neurons (15 medium- and 15 large-sized) exposed to bath application of NMDA (100 mM). For both large- and medium-sized neurons ramp voltage commands produced a typical current±voltage function to NMDA application (Fig. 2A). Lower amplitude currents were produced in large- compared to medium-sized neurons (Fig. 2A and B). These differences were statistically signi®cant from ±40 to ±25 mV (ANOVA, post hoc t-tests, P < 0.05; Fig. 2B). Capacitance measurements for medium- (48.3 6 3.1 pF) and large-sized (83.8 6 5.7 pF) neurons re¯ected the difference in cross-sectional area and were used to calculate current densities. It is important to take into account cell size or capacitance when evaluating ionic conductances in neurons of different sizes because the absolute magnitude of the current can be a function of neuronal size and not necessarily the density of channels (Alzheimer et al., 1993). The differences between mean current densities for large- and mediumsized neurons were proportionately much greater than the differences between current amplitudes (Fig. 2C). These differences were statistically signi®cant from ±60 to ±10 mV (ANOVA, post hoc ttests, P < 0.05). In addition, the voltages at which peak currents were obtained were signi®cantly shifted to more depolarized potentials in the large-sized neurons (±27.0 6 1.0 mV vs. ±30.4 6 1.2 mV for large- and medium-sized cells, respectively; t-test, P < 0.025; Fig. 2A±C). Because the slope of the current±voltage plots of the NMDAinduced currents in large- and medium-sized neurons appeared different, we examined more closely the voltage-dependence of NMDA receptor activation. The current±voltage relationship was converted to a conductance±voltage relationship (G±V) according to the formula G = I/(Vm ± Vrev), where Vm is the membrane potential and Vrev is the potential where the current was observed to reverse polarity. The midpoint potential (V0.5) and slope factor (b) of the G±V relationship were calculated by ®tting the data obtained from individual neurons by G/Gmax = 1/{1 + exp[(V0.5 ± Vm)/b]}, where V0.5 is the potential at which G/Gmax = 0.5 and b is the number of mV required to cause an e-fold change in conductance. According to this function, the smaller the value of b, the fewer mV are needed to cause an e-fold change in conductance. In other words, the smaller the value of b, the steeper the slope of the G±V relationship. Mean conductances (G) were signi®cantly greater for medium- than for large-sized neurons from ±35 to ±20 mV (Fig. 2D) (ANOVA, post hoc t-tests P < 0.05). The slope also was signi®cantly greater in mediumthan in large-sized neurons (13.1 6 0.8 vs. 11.0 6 0.7, respectively; t-test, P < 0.05). The midpoint of the G±V relationship was » 10 mV more negative in medium- than in large-sized neurons (±38.3 6 1.85 vs. ±30.5 6 2.1 mV, respectively, t-test, P < 0.05) indicating that NMDA receptors of large-sized neurons required more depolarization to remove the Mg2+ block. Kainate (100 mM)-induced currents were measured in 10 large- and 10 medium-sized cells. For both large- and medium-sized neurons, ramp voltage commands produced a typical linear current±voltage function to kainate application (Fig. 3A and B). In contrast to NMDA application, higher amplitude currents were produced in large- vs. medium-sized neurons. These differences were statistically signi®cant from ±90 to ±30 mV (ANOVA, post hoc t-tests, P < 0.05; Fig. 3B). Capacitance measurements for medium- (53.4 6 4.3 pF) and large-sized (107.3 6 6.3 pF) neurons were used to calculate current densities. The differences between mean current densities for large- and medium-sized neurons were proportionately smaller and were not statistically signi®cant (Fig. 3C). Acutely dissociated neurons Concentration±response relationships to activation of NMDA receptors were examined in more detail in acutely dissociated large- and medium-sized neurons using a rapid perfusion system so that neurons could be tested with a series of concentrations of NMDA. Data were obtained from 12 medium- and 12 large-sized neurons (Fig. 4A). All neurons were held at ±40 mV in Mg2+-free ACSF to remove the voltage-dependence of the response. Neurons were exposed to a series of ascending concentrations of NMDA (1, 10, 50, 100, 200 and 1000 mM, 3 s duration). Medium-sized neurons displayed a rapid peak current followed by a steady-state response (Fig. 4B). Largesized neurons displayed a smaller peak response than did mediumsized neurons, and this was also followed by a steady-state response (Fig. 4C). Concentration±current response relationships were sigmoidal and medium-sized neurons displayed greater peak currents at 50, 100, 200 and 1000 mM concentrations (Fig. 4D). However, because not all neurons were tested at each concentration the difference was only statistically signi®cant at the 100-mM concentration (t-test, P < 0.05). The respective EC50s were 49.5 6 9.8 and 38.2 6 22.4 mM for medium- vs. large-sized neurons. Current densities for peak responses were also computed by dividing by cell capacitances (Fig. 4E, inset). There were much greater increases in concentration±current density responses for medium- than for largesized neurons at 50, 100, 200 and 1000 mM concentrations (Fig. 4E). The differences were statistically signi®cant for each of these concentrations (t-tests, P < 0.05 to P < 0.01). The respective EC50s were 33.3 6 14.6 and 14.7 6 14.1 mM for medium- and large-sized neurons, respectively. Mean steady-state currents at the 100-mM concentration were greater for medium- than for large-sized neurons (±329 6 60 vs. ±196 6 26 pA, respectively), but the difference was not statistically signi®cant. In contrast, steady-state current densities at the 100-mM concentration were signi®cantly greater for mediumvs. large-sized neurons (±54 6 10 vs. ±12 6 2 pA/pF, respectively; t-test, P < 0.001). Spontaneous and evoked excitatory postsynaptic currents Spontaneous EPSCs were examined in large- and medium-sized neurons in the presence of bicuculline (10 mM), a GABAA receptor blocker. Spontaneous currents were sampled from 30 s up to 15 min in individual neurons at a holding potential of ±60 mV. These ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 1582 C. Cepeda et al. currents (> 5 pA in amplitude) occurred frequently in medium-sized neurons (Fig. 5B, left trace). In marked contrast, few spontaneous events were present in large-sized neurons (Fig. 5B, right trace). The difference in mean frequency was statistically signi®cant (1.64 6 0.42 vs. 0.05 6 0.03 events/s in medium- and large-sized neurons, respectively; t-test, P < 0.05). Average amplitudes of events were similar in medium- and large-sized neurons (11.5 6 0.89 and 9.2 6 0.58 pA, respectively; difference not statistically signi®cant). The decrease in frequency of occurrence of spontaneous currents probably re¯ects less afferent input to large- compared to the medium-sized neurons (Bolam et al., 1984). Electrical stimulation in close proximity to the cell was used to evoke EPSCs in medium- and large-sized neurons. The mean currents required to evoke EPSCs at 23 threshold in large- and medium-sized neurons were similar (29.5 6 9.0 and 20.0 6 1.43 mA, respectively; difference not statistically signi®cant). EPSCs were evoked at three holding potentials: ±60, ±20 and +30 mV. To analyse the NMDA receptor-mediated component 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX, 50 mM) to block non-NMDA receptors was added and the membrane was held at ±20 mV. Peak currents at ±60 mV holding potentials (in the absence of CNQX and mediated primarily by activation of non-NMDA receptors) did not differ signi®cantly between medium- and large-sized neurons (±169 6 42 and ±110 6 21 pA, respectively). There was a statistically signi®cant difference between cell capacitances for medium- and large-sized neurons (44 6 7.1 vs. 94 6 13.9 pF, respectively; t-test, P < 0.005). To evaluate peak current densities, peak currents were divided by cell capacitance. There was a statistically signi®cant decrease in current density in the large- compared to the medium-sized neurons (±1.4 6 0.3 vs. ±3.9 6 0.6 pA/pF, respectively; t-test, P < 0.001). EPSCs mediated by activation of NMDA receptors (peak currents measured at ±20 mV holding potential in the presence of CNQX) in medium-sized neurons were not signi®cantly greater in amplitude than those in large-sized neurons (±30.4 6 7.8 vs. ±16.4 6 3.3 pA, FIG. 3. Responses of medium- and large-sized neurons to kainate (100 mM) application. (A) Current±voltage plots for typical medium- and large-sized neurons. A ramp command was used to change voltage from ±90 to +40 mV and back to ±90 mV. The plotted data were obtained from the downward ramp. Control current±voltage plots were subtracted from plots in the presence of kainate. (B) Current±voltage plots (mean 6 SEM) for medium- and large-sized neurons. (C) Current density±voltage plots for medium- and large sized neurons. In B asterisks indicate groups differences were statistically signi®cant (P < 0.05). See Results for further details. ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 NMDA responses in striatal neurons 1583 FIG. 4. (A) Medium- and large-sized striatal neurons after acute dissociation and with the patch electrodes attached. Calibration in right panel refers to both panels. (B and C) Current traces evoked by 3-s application of NMDA at 1, 10, 50, 100, 200 and 1000 mM concentrations in (B) medium- and (C) large-sized neurons. (D) Concentration±peak current curves for medium- and large-sized neurons. (E) Concentration±peak current density curves for medium- and largesized neurons. Inset shows mean cell capacitances (6 SEM). In D and E, asterisks indicate groups differences were statistically signi®cant (P < 0.05 to P < 0.01). See Results for further details. Scale bars in C also apply to B. ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 1584 C. Cepeda et al. FIG. 5. (A) Top left panel shows IR images of a pair of striatal cells; one is a medium-sized (double arrow) and the other is a large-sized neuron (single arrow). Top right panel shows cells after being ®lled with biocytin. Single arrow points to the large neuron and the double arrow to the medium-sized neuron. (B) Patch-clamp recordings were obtained from both cells. Spontaneous EPSCs were recorded in the presence of bicuculline (10 mM) at a holding potential of ±60 mV. Middle traces are slow-speed recordings illustrating the marked absence of spontaneous EPSCs in the large-sized neuron (right) compared to the medium-sized cell (left). (C) A stimulating electrode was placed » 200 mm from the cells, thus approximately equidistant. Bottom traces are faster speed recordings showing evoked EPSCs at three holding potentials: ±60, ±20 and +30 mV. The stimulation parameters were identical for both cells. In all cases the evoked EPSCs were of greater amplitudes in the medium-sized neuron. Downward arrows indicate time of occurrence of stimulation pulse. respectively). However, mean current densities for NMDA receptormediated responses were statistically signi®cantly greater in mediumvs. large-sized neurons (±0.64 6 0.10 vs. ±0.20 6 0.04 pA/pF, respectively; t-test, P < 0.001). In selected experiments (n = 3) we were able to record from pairs of large- and medium-sized neurons, using the same stimulation placement and parameters (Fig. 5A). Similar to the grouped data described above, spontaneous EPSCs were more frequent and evoked EPSCs recorded at each holding potential were of greater amplitude in the medium- than in the largesized neurons (Fig. 5B and C). ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 NMDA responses in striatal neurons 1585 Cell swelling Another approach to examining excitatory amino acid receptor function in large- and medium-sized striatal neurons is to expose slices to agonists and assess cell swelling. Cell swelling, induced by activation of excitatory amino acid receptors, is presumably the ®rst step in a cascade that may ultimately lead to cell death (Choi, 1992). Previously, we showed that bath application of NMDA or kainate produced dose- and time-dependent swelling of medium-sized striatal cells (Colwell & Levine, 1996; Cepeda et al., 1998b). In the present experiments we examined the ability of large-sized striatal neurons to swell in the presence of NMDA or kainate. Modal cross-sectional areas for large cells were between 250 and 350 mm2 (Fig. 6B, inset). NMDA (50 or 100 mM) induced almost no change in cross-sectional area in large-sized striatal neurons (Fig. 6A, top panels). Average percentage increases in area were < 5% over the 15-min assessment period (Fig. 6B). When a group of medium-sized neurons (n = 15) was exposed to the same concentration of NMDA, statistically signi®cant cell swelling occurred. After 5 min exposure to NMDA there was a 19 6 3% increase in somatic cross-sectional area (ANOVA, post hoc t-test, P < 0.001). By 15 min the somatic crosssectional area increased by 43 6 6% (ANOVA, post hoc t-test, P < 0.001). Because it is possible that large-sized cells may be unable to increase somatic area, we tested the response to kainate in another group of large-sized neurons. Kainate (100 mM) produced a statistically signi®cant increase (» 40%) in largesized cell cross-sectional area 5, 10 and 15 min after application (ANOVA, P < 0.003) (Fig. 6A, bottom panels, and 6B). Thus, the lack of response to NMDA was not due to an inability of largesized cells to swell in the presence of excitatory amino acid receptor agonists. In medium-sized neurons we have shown previously that 100 mM kainate produces about a 30% increase in cross-sectional area (Colwell et al., 1998). We have shown previously that dopamine via activation of D1family receptors increases cell swelling induced by excitatory amino acid receptor agonists in medium-sized striatal neurons (Cepeda et al., 1998b). Thus, we examined whether NMDA in combination with dopamine could produce cell swelling in large-sized neurons (n = 5 cells). In the presence of dopamine (50 mM), cell swelling was not signi®cantly increased (Fig. 6B). Discussion The present experiments demonstrate, using a number of electrophysiological approaches, that large-sized striatal neurons were less responsive to NMDA receptor activation than were medium-sized neurons. First, whole-cell currents and to an even greater extent current densities, induced by exogenous application of NMDA, were decreased in large- compared to medium-sized neurons. Comparable ®ndings occurred in acutely dissociated neurons. Peak and steadystate current densities to NMDA application were reduced in largecompared to medium-sized neurons and this effect was consistent across a series of concentrations. There was also a shift in the voltage-dependence toward less negative values in large- compared to medium-sized neurons and a less steep negative slope conductance. In contrast, kainate-induced currents were greater in large- than in medium-sized neurons whilst kainate-induced current densities were similar in both cell types. In addition to decreased responsiveness to exogenous application of NMDA, large-sized neurons received less excitatory synaptic input than medium-sized neurons. Spontaneous EPSCs were markedly reduced in frequency and evoked EPSCs mediated by activation of NMDA or non-NMDA receptors were reduced in amplitude. Finally, NMDA-induced cell swelling, a ®rst step in an excitotoxic cascade, does not occur or is minimal in largesized neurons. This was true even in the presence of dopamine, a treatment that enhances NMDA-induced cell swelling in mediumsized neurons (Cepeda et al., 1998b). In contrast, kainate-induced swelling was robust in large-sized neurons. It is clear from the present ®ndings as well as previous analyses (Calabresi et al., 1998) that large-sized cholinergic neurons display reduced responsiveness to activation of NMDA receptors compared to medium-sized neurons. There are a number of mechanisms that could underlie such reduced responsiveness. These include decreased density of receptors and/or altered NMDA receptor subunit composition. The results of the present study clearly demonstrate that current density, which is a measure of current per unit of membrane clamped, is signi®cantly reduced in large-sized interneurons compared to the medium-sized cells, suggesting lower density of receptors. Whilst these values will be affected by differential space-clamp limitations to a certain extent, the magnitude of the difference cannot be accounted for by space-clamp errors. Of at least equal importance is the difference in NMDA receptor subunit composition between large- and medium-sized neurons (Landwehrmeyer et al., 1995). Differences in subunit composition will affect the agonist af®nity, Mg2+ sensitivity, voltage-dependence and the kinetics of the responses. For example, the lack of a large rapid peak response to NMDA application in dissociated large-sized neurons is probably due to differences in subunit composition (Huganir & Greengard, 1990). Large-sized cholinergic interneurons express lower levels of NMDA-R1 and NMDA-R2B subunit mRNA than do enkephalin-positive medium-sized neurons, and preferentially express the NMDA-R1 splice variant forms lacking one alternatively spliced carboxy-terminal region (Landwehrmeyer et al., 1995), which would change the sensitivity of protein phosphorylation (Tingley et al., 1993). The large-sized cholinergic interneurons also do not express NMDA-R2A subunits (Standaert et al., 1996). This ®nding is relevant because NMDA-R2 subunits impart different pharmacological pro®les to NMDA receptors (Monaghan & Larsen, 1997), and residues on the NMDA-R2A subunit control glutamate potency (Anson et al., 1998). Finally, the NMDA-R2D subunit is selectively expressed in striatal interneurons and absent in mediumsized projection neurons (Standaert et al., 1996, 1999). Channels expressing this subunit are unique in that they have a very long offset decay and an altered Mg2+ blockade (Monyer et al., 1994) which may explain, in part, the reduced negative slope conductance of the NMDA current. Furthermore, cells expressing NMDA-R2D subunits may have lower af®nity for agonists (Buller et al., 1994) which could partially explain sparing of the large neurons after quinolinic acid lesions. Recent studies using single-cell PCR have corroborated and extended the initial analyses of NMDA receptor subunit composition in large-sized cholinergic interneurons (Richardson et al., 2000). Interestingly, the NMDA-R2A subunit was detected when the sensitivity of the single cell assay was increased, suggesting low levels of expression. There are also fewer excitatory synaptic inputs to large- than to medium-sized neurons. The present data con®rm and extend the initial analyses demonstrating that these cells receive less excitatory input than do medium-sized spiny neurons (Wilson et al., 1990; Bennet & Wilson, 1999). We showed both reduced frequency of spontaneous EPSCs and reduced amplitudes of evoked EPSCs. Morphological evidence indicates fewer cortical excitatory synaptic inputs on large-sized interneurons than on other striatal cell types, although a thalamostriatal projection which is probably excitatory has ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 1586 C. Cepeda et al. been demonstrated (Lapper & Bolam, 1992). Based on electron microscopic studies, synaptic input to the perikarya and proximal dendrites of large cholinergic interneurons is very sparse and consists predominantly of boutons forming putative inhibitory symmetrical ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 NMDA responses in striatal neurons 1587 synaptic contacts. Asymmetric excitatory synaptic contacts are infrequent near the soma, although their number increases in distal dendrites (Bolam et al., 1984). A previous study described a generalized decrease in responsiveness of large cholinergic interneurons to NMDA, AMPA and kainate (Calabresi et al., 1998). Paradoxically, a large NMDA receptormediated EPSP was expressed in the cholinergic interneurons in the presence of Mg2+ in the external solution, even at relatively hyperpolarized membrane potentials (±75 mV). The electrophysiological data in the present study do not support an indiscriminate hyposensitivity of large-sized neurons, at least for kainate receptor activation. Neither do they support the presence of a large NMDA receptor-mediated synaptic component at hyperpolarized potentials. In fact, our results indicate that more depolarization is required to remove the Mg2+ blockade in large interneurons and the NMDA receptor-mediated synaptic current is reduced. Calabresi et al. (1998) only examined peak voltages or currents and did not include measurements of current density; this could signi®cantly alter the interpretation of the outcomes because large- and medium-sized neurons have markedly different surface areas. The present study has examined, in more detail and more extensively, current density and current±voltage relationships as well as directly comparing synaptic responses using voltage-clamp techniques in which changes in other variables would contribute less to the overall currents than in the synaptic responses in the previously reported current-clamp study. Calabresi et al. (1998) also did not initially block GABAA receptors when examining the excitatory amino acid receptor-mediated EPSPs, and consequently the electrical stimulation produced a mixed depolarizing potential consisting of multiple components due to activation of ligand- and voltage-gated channels. As pointed out above, other studies have shown that, relative to medium-sized spiny neurons, the EPSP in large interneurons is smaller (Wilson et al., 1990) and, although it is mediated by activation of both NMDA and non-NMDA receptors, the NMDA component is negligible at hyperpolarized potentials (Kawaguchi, 1992). Although differences in maturation of large- vs. medium-sized neurons may have contributed to the disparity between the present ®ndings and those of Calabresi et al. (1998), we believe this is unlikely because our ®ndings on large-sized neurons are in general agreement for NMDA receptor function but contrast on kainate receptor function. We have shown previously that non-NMDA receptor function actually develops earlier in the striatum than do NMDA receptors (Colwell et al., 1998; Nansen et al., 2000; Hurst et al., 2001). Thus, at the ages used in the present study, kainate receptor function would be at least as mature as or more mature than NMDA receptor function. Some of the unique properties of large-sized neurons are relevant for understanding their resistance to excitotoxic challenge, ischemia and their preservation in Huntington's disease. For example, c-fos induction after NMDA receptor activation is absent in large interneurons but not in medium-sized neurons, suggesting less coupling of NMDA receptor function to gene expression in the large cells (Aronin et al., 1991). Large interneurons express kainate receptor subunit proteins (Chen et al., 1996) and the present results indicate that they respond to kainate application. It was originally reported that large interneurons do not express AMPA receptor subunits (Chen et al., 1996) although they respond to AMPA application (Calabresi et al., 1998). More recently, single-cell PCR analyses have demonstrated the presence of AMPA receptor subunit proteins (Richardson et al., 2000). Unfortunately, little is known about the subtypes of glutamate receptors activated by the thalamostriatal pathway. Differential effects of anoxia±hypoglycemia occur between medium- and large-sized neurons in the striatum (Calabresi et al., 1997). Cholinergic interneurons hyperpolarize in response to anoxia± hypoglycemia whilst medium-sized neurons depolarize. Thus, they may be protected from the deleterious effects of depolarization induced by ischemic challenge. Whilst it was ®rst hypothesized that activation of a K+ current that is insensitive to tolbutamide mediated the hyperpolarization (Calabresi et al., 1997), subsequent studies have demonstrated an ATP-sensitive K+ channel (Lee et al., 1997, 1998) that may provide a protective role in anoxia±hypoglycemia. There are a number of reasons why large-sized cholinergic neurons may be spared in Huntington's disease. First, there may be differences in expression of normal or mutant huntingtin in largesized cholinergic interneurons (Ferrante et al., 1997). Selective vulnerability of medium-sized spiny striatal neurons observed in Huntington's disease has been associated with higher levels of huntingtin expression, whereas the relative resistance of large- and medium-sized aspiny neurons has been associated with low levels of huntingtin expression (Ferrante et al., 1997; Kosinski et al., 1997). However, a more recent study using a different antibody to identify huntingtin expression indicates that large cholinergic interneurons have higher levels of huntingtin than medium-sized neurons and suggests that it is the local environment that ultimately decides whether a neuron will be vulnerable to degeneration in Huntington's disease (Fusco et al., 1999). Early animal models of Huntington's disease produced excitotoxic lesions with kainate to duplicate human striatal cell loss (Coyle & Schwarcz, 1976; McGeer & McGeer, 1976). However, soon thereafter it was found that the excitotoxic model using quinolinic acid, which has more selectivity for NMDA receptors, better replicated the human ®ndings (Schwarcz et al., 1983; Beal et al., 1986; Ferrante et al., 1993; Roberts et al., 1993). If it is true that cholinergic interneurons are less sensitive to exogenous application of NMDA but not kainate as demonstrated by the present electrophysiological and cell swelling ®ndings, how can we explain the sparing of these cells after kainate application? One possibility is that excitotoxic lesions in the striatum require two conditions, activation of NMDA receptors and a signi®cant excitatory input (McGeer et al., 1978). The paucity of excitatory inputs impinging upon cholinergic interneurons may explain their relative resistance to kainate, whereas their resistance to NMDA could be explained by the combination of reduced receptor expression and different receptor subunit assembly, as well as paucity of excitatory inputs (Ferrante et al., 1987, 1993). FIG. 6. (A) IR images. Top left and right panels show NMDA application (100 mM, 15 min duration) failed to produce cell swelling in the large neurons (white arrows). In the same slice, the medium-sized cells displayed signi®cant cell swelling (white arrowheads). Bottom left and right panels show that, in contrast to the lack of swelling after NMDA, large neurons showed signi®cant increases in somatic area after kainate (100 mM, 15 min duration) (white arrows). (B) Bar graphs showing lack of cell swelling in large-sized striatal neurons at 5, 10 and 15 min during application of NMDA at either 50- or 100-mM concentrations. In the presence of dopamine, there was a slight increase in cell swelling to 100 mM NMDA. In contrast, kainate (100 mM, 15 min duration) produced signi®cant cell swelling after 5-, 10- and 15-min applications in large neurons. Inset shows a histogram of cross-sectional areas of large neurons. Asterisks in B indicate groups differences were statistically signi®cant (P < 0.003). See Results for further details. ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 1588 C. Cepeda et al. How are these results relevant for the understanding of Huntington's disease or other neurodegenerative disorders? We believe that the present results emphasize that multiple conditions must be met before neurons are at risk for excitotoxic degeneration. Large-sized cholinergic interneurons do not meet these conditions and thus are spared both in excitotoxic models of Huntington's disease and in the disease itself. In contrast, medium-sized spiny neurons meet both conditions and are at risk in both the models and the disorder. In conclusion, the present study demonstrates that large interneurons in the striatum display unique responsiveness to NMDA receptor activation, differing from medium-sized neurons. Inward currents are signi®cantly smaller and a stronger membrane depolarization is required to relieve the Mg2+ blockade. Cell swelling to NMDA, a ®rst step in an excitoxic cascade, is practically nonexistent. In contrast, large-sized neurons respond to kainate receptor activation in a manner similar to medium-sized neurons. We also found that the distribution of excitatory inputs onto large interneurons is relatively sparse compared to medium-sized neurons. Thus, the reduced responsiveness coupled with the altered NMDA receptor composition or reduced receptor density may help explain the relative resistance of large interneurons to neurodegeneration in neurological diseases. Taken together the present ®ndings help de®ne the conditions that put neurons at risk for damage in neurological disorders and they will aid in the design of therapies that can reduce or prevent neurodegeneration. Acknowledgements The authors greatly acknowledge Donna Crandall and Carol Gray for the preparation of the illustrations. This work was supported by USPHS grants NS33538 and NS35649. Abbreviations ACSF, arti®cial cerebrospinal ¯uid; AMPA, a-amino-3-hydroxy-5-methyl-4isoxazole propionic acid; CNQX, 6-cyano-7-nitroquinoxaline-2,3-dione; EGTA, ethylene glycol-bis(b-aminoethyl ether)-N,N,N',N¢-tetraacetic acid; EPSC, excitatory postsynaptic current; EPSP, excitatory postsynaptic potential; HBSS, Hanks' balanced salt solution; IR-DIC, infrared-differential interference contrast; NMDA, N-methyl-D-aspartate. References Alzheimer, C., Schwindt, P.C. & Crill, W.E. (1993) Postnatal development of a persistent Na+ current in pyramidal neurons from rat sensorimotor cortex. J. Neurophysiol., 69, 290±292. Anson, L.C., Chen, P.E., Wyllie, D.J., Colquhoun, D. & Schoepfer, R. (1998) Identi®cation of amino acid residues of the NR2A subunit that control glutamate potency in recombinant NR1/NR2A NMDA receptors. J. Neurosci., 18, 581±589. Aosaki, T., Kiuchi, K. & Kawaguchi, Y. (1998) Dopamine D1-like receptor activation excites rat striatal large aspiny neurons in vitro. J. Neurosci., 18, 5180±5190. Armstrong, C.M. & Gilly, W.F. (1992) Access resistance and space clamp problems associated with whole-cell patch clamping. Meth. Enzymol., 207, 100±122. Aronin, N., Chase, K., Sagar, S.M., Sharp, F.R. & DiFiglia, M. (1991) N-methyl-D-aspartate receptor activation in the neostriatum increases c-fos and fos-related antigens selectively in medium-sized neurons. Neuroscience, 44, 409±420. Bargas, J., Howe, A., Eberwine, J., Cao, Y. & Surmeier, D.J. (1994) Cellular and molecular characterization of Ca2+ currents in acutely isolated, adult rat neostriatal neurons. J. Neurosci., 14, 6667±6686. Beal, M.F., Ferrante, R.J., Swartz, K.J. & Kowall, N.W. (1991) Chronic quinolinic acid lesions in rats closely resemble Huntington's disease. J. Neurosci., 11, 1649±1659. Beal, M.F., Kowall, N.W., Ellison, D.W., Mazurek, M.F., Swartz, K.J. & Martin, J.B. (1986) Replication of the neurochemical characteristics of Huntington's disease by quinolinic acid. Nature, 321, 168±171. Bennett, B.D. & Wilson, C.J. (1998) Synaptic regulation of action potential timing in neostriatal cholinergic interneurons. J. Neurosci., 18, 8539±8549. Bennett, B.D. & Wilson, C.J. (1999) Spontaneous activity of neostriatal cholinergic interneurons in vitro. J. Neurosci., 19, 5586±5596. Bergson, C., Mrzljak, L., Smiley, J.F., Pappy, M., Levenson, R. & GoldmanRakic, P.S. (1995) Regional, cellular, and subcellular variations in the distribution of D1 and D5 dopamine receptors in primate brain. J. Neurosci., 15, 7821±7836. Bolam, J.P., Wainer, B.H. & Smith, A.D. (1984) Characterization of cholinergic neurons in the rat neostriatum. A combination of choline acetyltransferase immunocytochemistry, Golgi-impregnation and electron microscopy. Neuroscience, 12, 711±718. Buller, A.L., Larson, H.C., Schneider, B.E., Beaton, J.A., Morrisett, R.A. & Monaghan, D.T. (1994) The molecular basis of NMDA receptor subtypes: native receptor diversity is predicted by subunit composition. J. Neurosci., 14, 5471±5484. Burgard, E.C. & Hablitz, J.J. (1994) Developmental changes in the voltagedependence of neocortical NMDA responses. Brain Res. Dev. Brain Res., 80, 275±278. Calabresi, P., Ascone, C.M., Centonze, D., Pisani, A., Sancesario, G., D'Angelo, V. & Bernardi, G. (1997) Opposite membrane potential changes induced by glucose deprivation in striatal spiny neurons and in large aspiny interneurons. J. Neurosci., 17, 1940±1949. Calabresi, P., Centonze, D., Pisani, A., Sancesario, G., Gubellini, P., Mar®a, G.A. & Bernardi, G. (1998) Striatal spiny neurons and cholinergic interneurons express differential ionotropic glutamatergic responses and vulnerability: implications for ischemia and Huntington's disease. Ann. Neurol., 43, 586±597. Centonze, D., Mar®a, G.A., Pisani, A., Picconi, B., Giacomini, P., Bernardi, G. & Calabresi, P. (2001) Ionic mechanisms underlying differential vulnerability to ischemia in striatal neurons. Prog. Neurobiol., 63, 687±696. Cepeda, C., Peacock, W., Levine, M.S. & Buchwald, N.A. (1991) Iontophoretic application of NMDA produces different types of excitatory responses in developing human cortical and caudate neurons. Neurosci. Lett., 126, 167±171. Cepeda, C., Buchwald, N.A. & Levine, M.S. (1993) Neuromodulatory actions of dopamine in the neostriatum are dependent upon the excitatory amino acid receptor subtypes activated. Proc. Natl Acad. Sci. USA, 90, 9576±9580. Cepeda, C., Chandler, S.H., Shumate, L.W. & Levine, M.S. (1995) Persistent Na+ conductance in medium-sized neostriatal neurons: characterization using infrared videomicroscopy and whole cell patch-clamp recordings. J. Neurophysiol., 74, 1343±1348. Cepeda, C., Colwell, C.S., Itri, J.N., Chandler, S.H. & Levine, M.S. (1998a) Dopaminergic modulation of NMDA-induced whole cell currents in neostriatal neurons in slices: contribution of calcium conductances. J. Neurophysiol., 79, 82±94. Cepeda, C., Colwell, C.S., Itri, J.N., Gruen, E. & Levine, M.S. (1998b) Dopaminergic modulation of early signs of excitotoxicity in visualized rat neostriatal neurons. Eur. J. Neurosci., 10, 3491±3497. Chen, Q., Veenman, C.L. & Reiner, A. (1996) Cellular expression of ionotropic glutamate receptor subunits on speci®c striatal neuron types and its implication for striatal vulnerability in glutamate receptor-mediated excitotoxicity. Neuroscience, 73, 715±731. Chesselet, M.F., Gonzales, C., Lin, C.S., Polsky, K. & Jin, B.K. (1990) Ischemic damage in the striatum of adult gerbils: relative sparing of somatostatinergic and cholinergic interneurons contrasts with loss of efferent neurons. Exp. Neurol., 110, 209±218. Choi, D.W. (1992) Excitotoxic cell death. J. Neurobiol., 23, 1261±1276. Colwell, C.S., Altemus, K.L., Cepeda, C. & Levine, M.S. (1996) Regulation of N-methyl-D-aspartate-induced toxicity in the neostriatum: a role for metabotropic glutamate receptors? Proc. Natl Acad. Sci. USA, 93, 1200±1204. Colwell, C.S., Cepeda, C., Crawford, C. & Levine, M.S. (1998) Postnatal development of glutamate receptor-mediated responses in the neostriatum. Dev. Neurosci., 20, 154±163. Colwell, C.S. & Levine, M.S. (1996) Glutamate receptor-induced toxicity in neostriatal cells. Brain Res., 724, 205±212. Coyle, J.T. & Schwarcz, R. (1976) Lesion of striatal neurones with kainic acid provides a model for Huntington's chorea. Nature, 263, 244±246. Di Chiara, G., Morelli, M. & Consolo, S. (1994) Modulatory functions of ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589 NMDA responses in striatal neurons 1589 neurotransmitters in the striatum: ACh/dopamine/NMDA interactions. Trends Neurosci., 17, 228±233. DiFiglia, M. (1990) Excitotoxic injury of the neostriatum: a model for Huntington's disease. Trends Neurosci., 13, 286±289. Dodt, H.U. & ZieglgaÈnsberger, W. (1994) Infrared videomicroscopy: a new look at neuronal structure and function. Trends Neurosci., 17, 453±458. Ferrante, R.J., Beal, M.F., Kowall, N.W., Richardson, E.P. Jr & Martin, J.B. (1987) Sparing of acetylcholinesterase-containing striatal neurons in Huntington's disease. Brain Res., 411, 162±166. Ferrante, R.J., Gutekunst, C.A., Persichetti, F., McNeil, S.M., Kowall, N.W., Gusella, J.F., MacDonald, M.E., Beal, M.F. & Hersch, S.M. (1997) Heterogeneous topographic and cellular distribution of huntingtin expression in the normal human neostriatum. J. Neurosci., 17, 3052±3063. Ferrante, R.J., Kowall, N.W., Beal, M.F., Richardson, E.P. Jr, Bird, E.D. & Martin, J.B. (1985) Selective sparing of a class of striatal neurons in Huntington's disease. Science, 230, 561±563. Ferrante, R.J., Kowall, N.W., Cipolloni, P.B., Storey, E. & Beal, M.F. (1993) Excitotoxin lesions in primates as a model for Huntington's disease: histopathologic and neurochemical characterization. Exp. Neurol., 119, 46± 71. Fusco, F.R., Chen, Q., Lamoreaux, W.J., Figueredo-Cardenas, G., Jiao, Y., Coffman, J.A., Surmeier, D.J., Honig, M.G., Carlock, L.R. & Reiner, A. (1999) Cellular localization of huntingtin in striatal and cortical neurons in rats: lack of correlation with neuronal vulnerability in Huntington's disease. J. Neurosci., 19, 1189±1202. Gerfen, C.R. (1992) The neostriatal mosaic: multiple levels of compartmental organization in the basal ganglia. Annu. Rev. Neurosci., 15, 285±320. Graveland, G.A. & DiFiglia, M. (1985) The frequency and distribution of medium-sized neurons with indented nuclei in the primate and rodent neostriatum. Brain Res., 327, 307±311. Hamill, O.P., Marty, A., Neher, E., Sakmann, B. & Sigworth, F.J. (1981) Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. P¯ugers Arch., 391, 85±100. Hattori, T. & McGeer, P.L. (1973) Synaptogenesis in the corpus striatum of infant rat. Exp. Neurol., 38, 70±79. Horikawa, K. & Armstrong, W.E. (1988) A versatile means of intracellular labeling: injection of biocytin and its detection with avidin conjugates. J. Neurosci. Meth, 25, 1±11. Huganir, R.L. & Greengard, P. (1990) Regulation of receptor desensitization by protein phosphorylation. Neuron, 5, 555±567. Hurst, R.S., Cepeda, C., Shumate, L.W. & Levine, M.S. (2001) Delayed postnatal development of NMDA receptor function in medium-sized neurons of the rat striatum. Dev. Neurosci., 23, 122±134. Jiang, Z.G. & North, R.A. (1991) Membrane properties and synaptic responses of rat striatal neurones in vitro. J. Physiol. (Lond.), 443, 533±553. Kawaguchi, Y. (1992) Large aspiny cells in the matrix of the rat neostriatum in vitro: physiological identi®cation, relation to the compartments and excitatory postsynaptic currents. J. Neurophysiol., 67, 1669±1682. Kawaguchi, Y. (1993) Physiological, morphological, and histochemical characterization of three classes of interneurons in rat neostriatum. J. Neurosci., 13, 4908±4923. Kosinski, C.M., Cha, J.H., Young, A.B., Persichetti, F., MacDonald, M., Gusella, J.F., Penney, J.B. Jr & Standaert, D.G. (1997) Huntingtin immunoreactivity in the rat neostriatum: differential accumulation in projection and interneurons. Exp. Neurol., 144, 239±247. Kowall, N.W., Ferrante, R.J. & Martin, J.B. (1987) Pattern of cell loss in Huntington's disease. Trends Neurosci., 10, 24±29. Landwehrmeyer, G.B., Standaert, D.G., Testa, C.M., Penney, J.B. Jr & Young, A.B. (1995) NMDA receptor subunit mRNA expression by projection neurons and interneurons in rat striatum. J. Neurosci., 15, 5297±5307. Lapper, S.R. & Bolam, J.P. (1992) Input from the frontal cortex and the parafascicular nucleus to cholinergic interneurons in the dorsal striatum of the rat. Neuroscience, 51, 533±545. Lee, K., Brownhill, V. & Richardson, P.J. (1997) Antidiabetic sulphonylureas stimulate acetylcholine release from striatal cholinergic interneurones through inhibition of K (ATP) channel activity. J. Neurochem., 69, 1774± 1776. Lee, K., Dixon, A.K., Freeman, T.C., & Richardson, P. J. (1998) Identi®cation of an ATP-sensitive potassium channel current in rat striatal cholinergic interneurones. J. Physiol. (Lond.), 510, 441±453. Levine, M.S., Li, Z., Cepeda, C., Cromwell, H.C. & Altemus, K.L. (1996) Neuromodulatory actions of dopamine on synaptically-evoked neostriatal responses in slices. Synapse, 24, 65±78. McGeer, E.G. & McGeer, P.L. (1976) Duplication of biochemical changes of Huntington's chorea by intrastriatal injections of glutamic and kainic acids. Nature, 263, 517±519. McGeer, E.G., McGeer, P.L. & Singh, K. (1978) Kainate-induced degeneration of neostriatal neurons: dependency upon corticostriatal tract. Brain Res., 139, 381±383. Monaghan, D.T. & Larsen, H. (1997) NR1 and NR2 subunit contributions to N-methyl-D-aspartate receptor channel blocker pharmacology. J. Pharmacol. Exp. Ther., 280, 614±620. Monyer, H., Burnashev, N., Laurie, D.J., Sakmann, B. & Seeburg, P.H. (1994) Developmental and regional expression in the rat brain and functional properties of four NMDA receptors. Neuron, 12, 529±540. Nansen, E.A., Jokel, E.S., Lobo, M.K., Micevych, P.E., Ariano, M.A. & Levine, M.S. (2000) Striatal ionotropic glutamate receptor ontogeny in the rat. Dev. Neurosci., 22, 329±340. Richardson, P.J., Dixon, A.K., Lee, K., Bell, M.I., Cox, P.J., Williams, R., Pinnock, R.D. & Freeman, T.C. (2000) Correlating physiology with gene expression in striatal cholinergic neurones. J. Neurochem., 74, 839±846. Roberts, R.C., Ahn, A., Swartz, K.J., Beal, M.F. & DiFiglia, M. (1993) Intrastriatal injections of quinolinic acid or kainic acid: differential patterns of cell survival and the effects of data analysis on outcome. Exp. Neurol., 124, 274±282. Schwarcz, R., Whetsell, W.O. Jr & Mangano, R.M. (1983) Quinolinic acid: an endogenous metabolite that produces axon-sparing lesions in rat brain. Science, 219, 316±318. Standaert, D.G., Friberg, I.K., Landwehrmeyer, G.B., Young, A.B. & Penney, J.B. Jr (1999) Expression of NMDA glutamate receptor subunit mRNAs in neurochemically identi®ed projection and interneurons in the striatum of the rat. Brain Res. Mol. Brain Res., 64, 11±23. Standaert, D.G., Landwehrmeyer, G.B., Kerner, J.A., Penney, J.B. Jr & Young, A.B. (1996) Expression of NMDAR2D glutamate receptor subunit mRNA in neurochemically identi®ed interneurons in the rat neostriatum, neocortex and hippocampus. Brain Res. Mol. Brain Res., 42, 89±102. Tepper, J.M., Sharpe, N.A., Koos, T.Z. & Trent, F. (1998) Postnatal development of the rat neostriatum: electrophysiological, light- and electron-microscopic studies. Dev. Neurosci., 20, 125±145. Tingley, W.G., Roche, K.W., Thompson, A.K. & Hunganir, R.L. (1993) Regulation of NMDA receptor phosphorylation by alternative splicing of the C-terminal domain. Nature, 364, 70±73. Vonsattel, J.P., Myers, R.H., Stevens, T.J., Ferrante, R.J., Bird, E.D. & Richardson, E.P. Jr (1985) Neuropathological classi®cation of Huntington's disease. J. Neuropathol. Exp. Neurol., 44, 559±577. Welch, B.L. (1947) The generalization of Student's problem when several different population variances are involved. Biometrika, 3, 28±35. Wilson, C.J., Chang, H.T. & Kitai, S.T. (1990) Firing patterns and synaptic potentials of identi®ed giant aspiny interneurons in the rat neostriatum. J. Neurosci., 10, 508±519. Yan, Z., Song, W.J. & Surmeier, J. (1997) D2 dopamine receptors reduce N-type Ca2+currents in rat neostriatal cholinergic interneurons through a membrane-delimited, protein-kinase-C-insensitive pathway. J. Neurophysiol., 77, 1003±1015. ã 2001 Federation of European Neuroscience Societies, European Journal of Neuroscience, 14, 1577±1589