Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
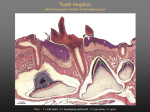
Genetic Interactions in Tooth Development and Agenesis Karen Harding Evo-Devo Fall, 2006 Abstract The human dentition is a continuously evolving system. The development of teeth is regulated by a variety of genes and transcription factors in a complex series of interactions. Although the genes involved in odontogenesis are highly conserved sequences, in the history of the dentition, changes have taken place and the human dentition continues to evolve. Among modern humans, there are variations in the number and size of teeth, with tooth agenesis affecting up to 20% of today’s population. The most common variation in the number of teeth is agenesis of the third molars, followed by the mandibular second premolars and maxillary lateral incisors. Less common are specific patterns of oligodontia, in which six or more teeth are missing. These differences reflect variations in the genes that are involved in tooth development. This paper will address tooth development and some of the mutations that can cause tooth agenesis. Primary tooth development begins prenatally. In humans, formation of the face and mouth begins in the fourth week of embryonic development. From the first brachial arch, or mandibular arch, develop the upper and lower jaws. Between the sixth and seventh weeks of prenatal development, odontogenesis of the primary dentition begins. There are four major stages in tooth development: initiation, bud, cap and bell stages. (4) The ectoderm-lined primitive mouth gives rise to the oral epithelium, a horseshoe-shaped tissue band at the surface of the stomodeum. A basement membrane separates the oral Page -2- epithelium from the mesenchyme. The oral epithelium gives rise to the dental lamina, which penetrates the mesenchyme during the bud stage. The dental lamina then develop into buds, which penetrate the underlying mesenchyme. From these structures, tooth buds develop into tooth germs and their surrounding structures. During the late bud stage, primary enamel knots form.(3, 4, 6 ) The enamel knot is made up of a population of transient epithelial cells that appear at the location of the primary tooth cusps. The primary enamel knots are epithelial signaling centers that mediate pattern formation and morphogenesis. Over 50 different genes are expressed in the knots. Twelve of those genes are signaling molecules that belong to the Bmp, Fgf, Shh and Wnt families. Their expression domains predict future cusp patterns. (5) During this stage, the capacity for induction of tooth morphogenesis resides in the mesenchyme. (4) It is during the next two stages of development, the cap and bell stages, that differentiation and morphogenesis occur. Cusp formation begins here, as the tooth crown begins to develop after the epithelium begins to grow into the underlying mesenchyme. The enamel knots secrete activators, including Bmps, which are expressed until a threshold is reached. At the threshold point, inhibitors, including Fgf’s and Shh, which counteract secretion of the activator, also inhibit formation of the second cusp. Activator and inhibitor modulate tissue growth, making developing shape itself have a causal role in the placement of new knots. Cusp sharpness influences the distance between knots, because sharpness modifies the volume of mesenchymal tissue into which the activator and inhibitor diffuse. During all stages, a tremendous number of genetic interactions occur. (4, 5 ) Patterning of the dentition is under tight control. In all mammals each group of teeth (incisors, canines premolars and molars) have their own shapes and sizes related to their dental Page -3- arch position. According to Butler (1939) there are 3 morphogenetic fields in tooth development, the incisors, canines and premolars. Butler suggests that all of the primordial teeth are initially equivalent, but develop differently depending on concentrations of local morphogens. Butler hypothesized that within each field there would be one key tooth, and other teeth within the field would be less evolutionarily stable. Clayton (1956), in a study of 3557 humans, found that the most posterior or terminal tooth in a series was most often missing. He hypothesized that these missing teeth were vestigal organs and have been lost in modern man due to having little value and no selective advantage. According to Osborn (1978), primordial teeth are specified by migrating neural crest cells. (3, 9 ) Msx and Dsx are two classes of homeobox genes which are expressed in migrating neural crest cells. Before odontogenesis, Msx1 and Msx 2are expressed mesenchymally in horseshoeshaped fields in the anterior region of the first brachial arch. In the posterior border, Msx1 expression extends farther than Msx2. This region corresponds to the position of future primary epithelial thickening. Msx1 and Msx2 both become localized as tooth development progresses in mesenchymal cells of the papilla and dental follicle, but Msx2 also is expressed strongly in the enamel organ. In studies of mice with targeted mutations in Msx1, incisors failed to develop and molar development was arrested at the late bud stage. In mice with targeted mutations in both Dlx1 and Dlx2, maxillary molars were absent. Dlx1 and Dlx2 are expressed only in the proximal regions of the maxillary and mandibular arch where the molars will develop. Barx-1 is also a homeobox containing transcription factor that is expressed in the region of future molar development. It is not expressed in anterior regions. (3) Page -4- Each region of the developing jaw contains a unique combination of homeobox gene expression which gives rise to specific types of teeth. This is due to differential expression of coded homeobox nuclear proteins responsible for regulating downstream gene transcription. (3) Bone morphogenetic proteins (Bmps) are involved in signaling functions mediating tissue interactions during development. During early tooth development, Bmp2, 4, and 7 are expressed in the dental epithelium. Only Bmp-4 is shifts in its expression pattern from epithelium to mesenchyme at the same time that the inductive potential for odontogenesis is shifting from epithelium to mesenchyme. Studies of mice have shown that Msx1 is necessary to mediate this shift in the expression of Bmp-4 from epithelium to mesenchyme. In experiments with noggin, which is an antagonist of Bmp-4, it was found that Bmp-4 provides a positive feedback loop that maintains expression of Bmp-4 and Msx-1 in the mesenchyme. In this manner, the expression of these genes is restricted to the future regions of tooth formation. (3) Also important in tooth development are the fibroblast growth factors (Fgfs). Fgf-4,Fgf8, and Fgf-9 expression is found in the epithelial cells of the tooth germ when epithelialmesenchymal interactions regulating tooth morphogenesis are occurring. Fgf-8 and Fgf-9 have mediating roles in initiation of tooth development, and Fgf-4 and Fgf-9 have roles in determination of coronal morphology. (3, 5) Sonic hedgehog (Shh) is also known to have a role in dental patterning. It is expressed in the epithelial thickenings that will become regions of tooth formation and in the enamel knot. Shh signaling is closely controlled during early tooth development, and Shh signaling is antagonized in areas where teeth do not develop (1,3) Prx1 proteins are associated with signaling pathways that are important in normal molar tooth development. Prx1 and Prx2 are paired-related homeobox genes that code for transcription Page -5- factors necessary for orofacial development. They are expressed in the mesenchyme of tissues that develop into cartilage, bone and teeth. Prx1 and Prx2 transcripts are widely expressed in undifferentiated mesenchyme throughout the development of the mandible and maxilla preceding tooth development. They are necessary for mesenchymal-epithelial signaling that occur during tooth morphogenesis. In a study by Mitchell et al (2006), in mutant mice that were lacking Prx1 and Prx2 genes, it was found that there were severe craniofacial defects and mandibular hypoplasia. Also resulting were missing or fused incisors at the midline of the mandible. In addition, in the anterior mandible there was altered expression of Shh, Pax9, Dlx2 and Alx3, all of which affect tooth morphogenesis. In studying mesenchymal expression and morphological defects, it was found that Prx genes are involved in morphogenesis of the maxillary and mandibular second molars. The Prx null embryos exhibited epithelial malformations such as ectopic epithelial projections, cuspal patterning defects and cervical loop hypoplasia. (11) Three genes have been associated with the nonsyndromic form of tooth agenesis in humans; they are AXIN2, MSX1, and PAX9. Pax9 has been studied most intensely. (2) The PAX gene family is known to code for transcription factors. The PAX proteins contain a 128-aa DNA binding domain called the paired domain, which is highly conserved. There have been nine PAX genes found in mammals (PAX1-PAX9). PAX 9 is involved in palate and tooth formation. It is expressed in the developing vertebral column, the pharyngeal pouches, and the neural-crest-derived mesenchyme of mandibular and maxillary arches. (2) Pax9 is an early mesenchymal marker of tooth development. It is necessary for tooth development to continue beyond the bud stage. In mouse embryos that are Pax9 deficient, development of teeth stops at the bud stage, where in normal embryos, tooth mesenchyme produces signals that are Page -6- necessary for the dental ectoderm to further develop. In humans, nine different mutations of Pax9 have been found, on exons 2 and 4. These mutations are associated with autosomal dominant forms of oligodontia and nonsyndromic hypodontia. Selective tooth agenesis, especially of the molars, was found to be caused by these mutations or deletion of the Pax 9 allele. Families with Pax9 mutations had tooth agenesis as the only clinical sign. (2) The expression of Pax9 marks the future sites of tooth development prior to any morphological manifestation of odonotgenesis. Induction of mesenchymal Pax9 expression is under control of signals produced in the ectoderm. FGF8 has been identified as an ectodermal inducer of Pax9 expression, and BMP2 and BMP4 have been shown to function as antagonists of the FGF8 inductive signal. Therefore, tooth formation in the mandibular arch is not initiated by a local inducer, but by the combination of different signals that are produced in wide, overlapping domains. (2) (See Figure 1) After Pax9 expression has been induced in the mesenchyme, and after thickening of the dental ectoderm, BMP4 has a different function in odontogenesis. It is expressed transiently in thickened dental ectoderm and acts as a signal, mediating inductive tissue interactions in developing teeth. (2) In a study of Pax9 variation, Pereira et al. analyzed the DNA sequence of the coding regions and the exon-intron boundaries of Pax9 in eighty-six people. The people were divided into three geographic groups: Asian, Native American, and European. Also, two groups from Poland were investigated for a mutation in exon 3; the first a group of 144 healthy individuals and the second, 206 individuals with facial/dental anomalies. Also analyzed were four species of New World monkeys, a gorilla, and orthogolous Pax9 sequences from public databases. From this information, Pereira et al. came up with new insights about the evolution of Pax 9 and Page -7- whether it contributed to the variation of dental phenotype within humans and variations between humans and other primate species.(2) In each of the 86 people, 172 chromosomes were sequenced. Of those, only 12% showed variations. However, all of those variations were represented in only four distinct mutations, which were all located in exon 3. Three of these had to do with amino acid replacements, one of which was a G->C transversion at nucleotide 718, causing a substitution at residue 240 of Ala for Pro(GCG->CCG, Ala240Pro). The Ala at codon 240 is highly conserved in several species, and this polymorphism of Ala240Pro is in a region homologous to one in the zebrafish, where a potent transactivating domain has been reported. (2) Pereira et al. found in humans a greater genetic diversity in Pax9 coding regions than in non-coding regions. The Pax9 paired domain, which is located in exon 2, has been highly conserved, as it is identical in humans and mice at the amino acid level. Only in the Pax9 exon 3 were variants found, so this exon was examined in other species as well as in humans. They found a striking conservation. At the nucleotide level, humans and gorillas are identical, and both are identical to the mouse at the amino acid level. There has been no amino acid replacement at this exon in the approximately 64-74 million years that human and mouse lineages are separated by. In contrast, five differences have occurred in the 35-48 year-old lineage separating apes/humans and New World monkeys. In measuring the nonsynonomous divergence/ synonomous divergence ratios of the exon 3 ortholog regions, it was found that this region is under selective pressure and is therefore likely to be functional. They concluded that nonsynomous substitutions in exon 3 are under strong negative selection. Looking at the combined data, they concluded that he evolution of Pax 9 exon 3 ortholg regions is driven by intensely purifying selection(2). Page -8- Msx1 is a non-clustered homeobox protein which plays a role in tooth development. In experiments on mice, it was found that Msx1 deficient mice have cleft palate, complete agenesis of incisors and bud-stage arrest of development of molars. Mutations in Msx1 have been found to be responsible for specific patterns of inherited tooth agenesis in humans. Specifically, an Arg196Pro missense mutation was found in a large family with severe autosomal-dominant agenesis. (Vastardis, 1996) (See Figure 2) A Ser105Stop mutation was found in another family with a similar pattern of missing teeth. In the latter family, some of the affected individuals also had cleft lip or cleft palate. A Ser202Stop mutation was found to be present in individuals with Witkop syndrome, which includes nail dysgenesis and tooth agenesis. (10) In a study of 92 individuals from 82 families, all of whom were missing at least one secondary tooth, Lidral and Reising (2002) found a Msx1 Met61Lys substitution in two siblings. The siblings were from a large family in which it was found that the mutation was consistent with a specific pattern of tooth agenesis in the extended family. This specific pattern, in which third molars, second premolars, and lateral incisors are missing, is consistent with other mutations of Msx1, and suggests that Msx1 mutations are responsible for this pattern of tooth agenesis. No mutations were found in individuals with the more common cases of incisor or premolar agenesis, suggesting a different etiology for those cases. (See figures 2, 3) (5, 10) An Axin2 mutation, a nonsense Arg656Stop, has also been implicated in cases of oligodontia. Axin2 is a Wnt signaling regulator. Both Axin2 and Wnt are necessary for normal tooth development. Wnt genes are also known to be involved in carcinogenesis. In a study of a four-generation Finnish family with this mutation, 11 members were found to be missing at least eight secondary teeth, and two members had developed only three secondary teeth. Most were lacking secondary premolars, molars, maxillary lateral incisors, and lower incisors. Three were Page -9- lacking canines. The tooth phenotypes in this family were different from those associated with other mutations. The records revealed that two patients from the oldest generation who had oligodontia also had a history of colorectal cancer. During this study, it was discovered that six other family member affected with oligodontia also had colorectal cancer, while none of the family members with a normal dentition did. In this family, it seems that oligodontia was associated with colorectal cancer. (7) (See Figure 4) It is apparent that the process of tooth development is a very complex one in which there are many genetic interactions occurring. The study of tooth development provides insight into the developmental control mechanisms occurring during embryogenesis. A better understanding of induction and patterning mechanisms can be gained by studying the developing tooth. The discovery that a certain form of oligodontia has been linked to colorectal cancer is an important one, and the possibility that similar connections exist should be studied further. Future research will undoubtedly expand our understanding of these complex interactions. Page -10- Figure 1 (8) Page -11- Figure 2 (9) Figure 3 (10) Page -12- Figure 4 (11) Page -13- Bibliography 1. Cobourne, Martyn T., I. Miletichz, P.T. Sharpe. 2004 Restriction of Sonic Hedgehog During Early Tooth Development. Development 131, 2875-2885. 2. Pereira, Tiago, F.M. Salzano, A. Mostowska, WH Trzeciak, A.Ruiz-Linares, J.A. Chies, C. Saavedra, C. Nagamachi, A.M, Hurtado, K. Hill, D. Castro-de -Guerra, W.A. Silva-Junior, and M.C. Bortolini. (2006) Natural selection and molecular evolution in primate PAX9 gene, a major determinant of tooth development. PNAS vol. 103 no. 15. 5676-5681 3. Cobourne, Martyn. T. The Genetic Control of Early Odontogenesis. March, 1999 4. Bath-Balogh, Mary and Fehrenbaach, Margaret. 1997. Illustrated Dental Embryology, Histology, and Anatomy. W.B. Saunders, Philadelphia, Pa. 5. Salazar-Ciudad, Issac, and J. Jernvall. 2002. A gene network model accounting for development and evolution of mammalian teeth. PNAS vol 99 no. 12 . 81168120. 6. Http://8e.devbio.com/article.php?ch+13&id=274 7. Mitchell, J.M., D.M. Hicklin, P.M. Doughty, J.H. Hicklin, J.W. Dickert, S.M. Tolbert, R. Peterkova, M.J.Kern. (2006) The Prx1 Homeobox Gene is Critical for Molar Tooth Morphogenesis. J Dent Res 85 (10) 888-893. 8. Stock, David W. 2001. The genetic basis of modularity in the development and evolution of the vertebrate dentition. The Royal Society. Phil Trans R. Soc. Lond. B (2001) 356, 1633-1653. 9. Vastardis, Helini. 2000. The genetics of human tooth agenesis: New discoveries for understanding dental anomalies. American Journal of Orthodontics and Dentofacial Orthopedics. June, 2000. 650-656. 10. Lidral, A.C. and Reising, B.C. 2002. The Role of MSX1 in Human Tooth Agenesis. Journal of Dental Rest. 81(4) 274-277. 11. Lammi, Laura, S. Arte, M. Somer, H. Jarvinen, P. Lahermo, I. Thesleff, S. Pirinen, P. Nieminen. 2004. Mutations in Axin2 Cause Familial Tooth Agenesis and Predispose to Colorectal Cancer. Am. J. Human Genetics. 74:1043-1050. Page -15-