Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
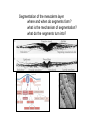
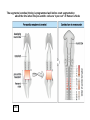
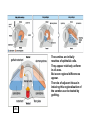
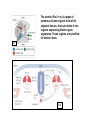
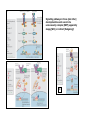
What’s going on….at cellular level. G10 W8 Changes in individual cell shape (apical constriction) added together produce tissue deformation (dimpling, invagination)…. But there’s more….. W8 ECM (fibronectin) migration substrate Cell migration leads cells to crawl over the underside of the blastocoel roof, on an adhesive carpet of fibronectin matrix. But there’s more…… W3,8 brachyury The new ability of mesoderm cells to migrate, with a net direction toward the midline, plus a general loosening (but not loss) of cellcell adhesion, allows cells of lateral origin to intercalate between more medial cells. As a mass, the tissue becomes narrower but lengthens: this is called convergent extension. But there’s more……. Gastrulation stage mitotic marker G10 W8 The cells of the animal hemisphere—fated to be ectoderm– increase in number by mitosis and decrease in number of cell layers (by radial intercalation). They may also become individually more flattened. The net result of this is this layer increases in area. This also drives the ectoderm to “take over” all the superficial layer, and helps drive internalisation of other cell types (mesoderm, endoderm). This type of morphogenetic movememt is called epiboly. The total outcome of invagination+cell migration+convergent extension+epiboly internalises the cells defined as endoderm and mesoderm, with the latter surrounding the former. These two are completely surrounded by the ectodermal cell layer. The relative importance of these processes seems to vary between species, but the overall outcome (3 layers) is in principal similar. This then places tissues together in new ways so that new inductive interactions are possible. EARLY AMNIOTE VERTEBRATE DEVELOPMENT The chick embryo starts as a thin disk on top of an immense yolk sphere This thin disk of “normal” oocyte cytoplasm houses the oocyte pronucleus, and after fertilization, the zygotic nuclei. W2 Initially, complete cell division lags behind nuclear proliferation, but soon catches up. The single layer (epiblast) seeds cells to form another layer beneath (hypoblast), separated by a fluid-filled space….just like a squashed version of a frog blastula. G11 W2 Gastrulation: Some epiblast cells now invaginate. Process is very similar to gastrulation in frogs, but the role of ‘individual” cell migration is clearer. End result: specific epiblast cells are internalised to produce a middle layer (mesoderm). Gastrulation in birds and mammals is an example of the Epithelial-Mesenchymal Transition (EMT). EMT is used repeatedly in development where cells start to move around. It also occurs when carcinoma cells start to invade. G11 The site of invagination– the primitive streak – persists for some hours but the cells in it change continuously. The primitive streak gets longer then shorter. The end –Hensen’s Node--becomes a major signalling centre equivalent to the Spemann Organizer/ dorsal lip of the blastopore. G11 At the end of gastrulation, a large number of cells have been lost from the epiblast layer: the depleted top layer is then called the ectoderm. The invaginated cells form the mesoderm layer, but some go further and intercalate into the hypoblast (now called endoderm) Patterning genes expressed in frog blastulas (hollow spherical) are also found in positionally relevant places in the blastoderms of birds (flat blastoderm) and mice (flat but cup-shaped blastoderm). And the relevant regions (eg. chick Hensen’s Node) can signal across species barriers (eg. Like frog DLB), indicating that function is conserved, as well as gene/protein sequence and spatiotemporal expression pattern. W4 Segmentation of the mesoderm layer where and when do segments form? what is the mechanism of segmentation? what do the segments turn into? The segments (somites) timing is programmed well before overt segmentation about the time when the pre-somitic cells are “spun out” of Hensen’s Node W4 One segment carved off every 90 min….. Pattern of Hairy1 mRNA expression W4 Scheme in Pre-Somitic Mesoderm: G6 •FGF-8 at Hensen’s node represses Lunatic Fringe •As cells displaced further anterior to HN, Lun Fringe activated •Lunatic fringe potentiates Notch signalling •Notch signals induce a repressor of Notch gene •Repressor has a short half-life. •Notch appears in an oscillating pattern. •Notch signals to form new boundary, presumably by…... •Notch regulates many genes inc. Hairy Candidate genes for transcriptional control by hairy1 is Ephrin and Eph receptors. The Eph-Ephrin system are cell surface molecules signalling cellular repulsion. Given a segmental striping of expression of these genes, his would be an ideal mechanism for separating tissue blocks. The segments (somites) type is programmed well before overt segmentation about the time when the pre-somitic cells are “spun out” of Hensen’s Node W4 The somites are initially rosettes of epithelial cells. They appear relatively uniform in all axes. But soon regional differences appear. The role of adjacent tissue in inducing this regionalisation of the somites can be tested by grafting. W4 The somite (Pax3 +ve) is target of numerous known signals from all its adjacent tissues, that sub-divide it into regions expressing distinct gene signatures. These regions are specified for distinct fates. W4 G14 Signalling pathways in these [and other] developmental events seem to be unnecessarily complex [BMP], apparently sloppy [Wnt], or indirect [Hedgehog]! G6 L-R Patterning--Not included 2007