Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
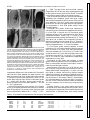
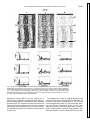
Fetal tissue containing the suprachiasmatic nucleus restores multiple circadian rhythms in old rats HUA LI1 AND EVELYN SATINOFF1,2 of Psychology, University of Illinois at Urbana-Champaign, Champaign, Illinois 61820; and 2Department of Psychology and Program in Neuroscience, University of Delaware, Newark, Deleware 19716-2577 1Department neural transplantation; body temperature; activity; drinking; aging; vasoactive intestinal polypeptide AGING AFFECTS THE PERIOD, amplitude, and phase of circadian rhythms (for review, see Ref. 25). The most commonly reported change is diminished amplitude of, for instance, the circadian temperature rhythm (24), of sleep (20) and of wheel running and drinking (23). Aging also affects cellular, metabolic, and electrophysiological properties of the suprachiasmatic nucleus (SCN). Using 2-deoxy-D-glucose uptake as a marker, Wise et al. (38) found a substantial decrease in metabolic activity in the SCN of aged rats. Sutin et al. (33) found a decreased response of the immediate-early gene c-fos in the SCN of old rats after photic stimulation. The firing rate of SCN cells in a slice preparation is of lower amplitude in aged animals than in young ones (28). There is a decrease in the number of arginine vasopressin (AVP) and vasoactive intestinal polypeptide (VIP) neurons in the SCN of old rats, as revealed by immunocytochemical staining (7, 26). Partial SCN lesion studies show that the size of the SCN is crucial for the expression of its pacemaker properties (27). The decrease in SCN volume and cell number might be one factor underlying circadian rhythm disturbances in old rats. The SCN in the hypothalamus is the major circadian pacemaker in mammals. Perhaps the strongest evidence for this is that circadian rhythmicity is restored in SCN-lesioned arrhythmic hosts after transplantation of fetal tissue containing the SCN (e.g., Ref. 18). There has been very little behavioral work done on fetal SCN transplants in aged rats. When fetal SCN tissue was transplanted into old hamsters that had had their own SCN ablated, the free-running period of these animals lengthened to resemble that of younger hamsters (36). When grafted aged hamsters with intact SCN were injected with triazolam, a phase-shifting drug, activity rhythms phase-shifted more quickly than did those of young hamsters (35). Recently, Cai et al. (3) reported that fetal SCN grafts into middle-aged female rats with intact SCN enabled the aged hosts to regain their ability to show diurnal patterns of fos expression. The present study is based on the fact that the circadian rhythms of many aged rats become aperiodic. In this respect they resemble the circadian rhythms of young rats with partial SCN lesions (29). In the present experiments we measured three circadian rhythms: body temperature (Tb ), drinking, and activity. Our hypothesis was that fetal transplants of SCN tissue into intact but aperiodic aged hosts might improve circadian rhythms just as they do in young animals with SCN lesions. The SCN contains several different neuropeptides and neurotransmitters (for review, see Ref. 14). For example, there are large populations of VIP cells in the ventrolateral SCN (5). This area receives dense projections of neuropeptide Y (NPY)-ergic fibers from the intergeniculate leaflet (6). There are large populations of AVP cells in the dorsomedial SCN (34). The codistribution of dense fiber plexi characterized by VIP, AVP, and NPY immunoreactivity in the SCN provides a distinct neuropeptide profile for distinguishing SCN from adjacent hypothalamic tissue and from other brain tissues (17). A similar cytoarchitecture is preserved in fetal SCN tissue grafts (1, 17, 18). Immunocytochemical staining of these three peptides is frequently used to identify the SCN both in intact animals and in grafted hypothalamic tissues. Therefore we used these peptide markers in the present experiments. 0363-6119/98 $5.00 Copyright r 1998 the American Physiological Society R1735 Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 Li, Hua, and Evelyn Satinoff. Fetal tissue containing the suprachiasmatic nucleus restores multiple circadian rhythms in old rats. Am. J. Physiol. 275 (Regulatory Integrative Comp. Physiol. 44): R1735–R1744, 1998.—The suprachiasmatic nucleus (SCN) is the major circadian pacemaker in mammals. When fetal tissue containing the SCN is transplanted into young rats whose circadian rhythms have been abolished by SCN lesions, the rhythms gradually reappear. Circadian rhythms in many rats deteriorate or disappear with age. The rationale of the present study was that old rats with poor circadian rhythms resemble young rats with SCN lesions. If there is a similar mechanism underlying this resemblance, then fetal tissue containing the SCN should restore rhythms in old rats. Therefore, we implanted such tissue into the third ventricle of intact aged rats with poor circadian rhythms. Body temperature, locomotor activity, and/or drinking were measured simultaneously within subjects. Grafts and hosts were stained immunocytochemically for vasoactive intestinal polypeptide (VIP), arginine vasopressin (AVP), and neuropeptide Y (NPY). Of 23 SCN grafts, 14 were viable (cells observable with Nissl or peptide staining). In 7 of the 14 aged hosts, up to three circadian rhythms were improved or restored. VIP cells were always observable, which was not the case for AVP cells or NPY fibers. In the other seven hosts, no circadian rhythm was improved. Compared with the successful grafts, these unsuccessful grafts had similar amounts of AVP and NPY staining but significantly less VIP cell and/or fiber staining. Fetal cerebellar grafts, which do not contain any of the three peptides, did not improve or restore any rhythms. Thus the degeneration of circadian rhythms in aged rats may be due, at least in part, to deterioration of the aged SCN and in particular, to a loss of function of VIP-containing neurons. R1736 SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS MATERIALS AND METHODS Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 Subjects and housing. Subjects were young (3–6 mo) and old (19.5–29.6 mo) male and female Long-Evans rats bred in the animal colonies of the University of Illinois and the University of Delaware. Both colonies were pathogen-free, and regular serological testing was performed to ensure that condition. Rats with visible lesions or tumors were not used. Food and water were available ad libitum. The rats were maintained from birth at an ambient temperature of 23 6 1°C on a 12:12-h light-dark cycle (LD). At the beginning of the screening process for grafting candidates, all rats (except 3) were maintained in constant darkness (DD). This was done because we were mainly interested in endogenous circadian rhythms, which can only be observed under constant conditions. The effect of SCN grafting on entrained rhythms was of lesser interest. Experimental design. Using periodogram analysis and gray scales (see Data analysis), we have found over the years that if a circadian rhythm of an old rat is poor or nonexistent, i.e., showing very low or no circadian power (power at 24 h), for at least 2 wk, it does not become rhythmic again under either LD or DD. In the present study we identified 28 old rats in which at least one of the rhythms we measured had no power for at least 1 mo; they became candidates for grafting. Twenty-five of these rats were placed in DD at least 1 mo before grafting. (One of these, OF119, was shifted from DD to LD 2 mo postgrating.) Three other rats were housed and grafted in LD. Tb, locomotor activity, and drinking were continuously monitored. Fetal tissue containing either the SCN (n 5 23) or a comparable amount of cerebellar tissue (controls, n 5 5) was implanted into the third ventricle of those animals. Two other control groups received fetal SCN grafts. They were included to examine possible interactions between the healthy host SCN and the grafted SCN. These groups were old rats with good rhythms (n 5 4) and 3- to 6-mo-old intact rats (n 5 6). Measurements continued for 1–2 mo after the graft operation. The animals were then killed and their brains removed, cut, stained, and examined microscopically. Grafts and host SCN were stained for the presence of VIP and AVP cells and NPY fibers and also stained with thionin. Temperature telemeter placement. Young rats were anesthetized with ketamine-HCl (90 mg/kg) and xylazine (13 mg/kg). Two-thirds of this dose was used for old rats. A temperature telemeter (model VM; Mini-Mitter, Sunriver, OR) was implanted in the peritoneal cavity, and the wound was sutured. The cylinder-shaped telemeter (diameter 1.2 cm, length 1.7 cm, weight 3 g) was calibrated at 30 and 42°C in a temperature-controlled water bath. The telemeter was accurate to 0.1°C. Fetal tissue transplantation. Anesthetized rats were placed in a stereotaxic device. Fetal tissue containing the SCN was lowered to the third ventricle using the following coordinates: anterior-posterior 21.0 mm from bregma, dorsoventral 29.0 mm from the dura, and laterally at the midline. Implants (volume 0.5–1 mm3 ) were obtained from the brains of rat embryos at day 17 (E17) of gestation (day 1 being the first day after mating) to E20. Pregnant female rats were anesthetized, and an incision was made at the level of the lower abdomen to expose the uterus. One pup at a time was taken from the uterus. The head of the pup was removed and placed in a sterile petri dish on ice. Under a stereoscopic zoom microscope, the brain of the pup was dissected out. The tissue block containing the SCN was cut out from the ventral surface of the hypothalamus with coronal cuts just rostral and caudal to the optic chiasm and with lateral cuts equidis- tant from the midline. Embryonic segments from two donors were pooled in sterile saline and injected into each host animal using a modified 20-gauge needle. The time between microdissection and implantation of the fetal tissue never exceeded 1 h. Usually, 2–6 rats were grafted with pups from one mother in one session. At the end of a grafting session, the pregnant female was euthanized with an overdose of ketamine. The grafted rats were returned to their home cages to recover. Histology. The rats were perfused with 4% paraformaldehyde in 0.1 M phosphate buffer, pH 7.3. After overnight postfixation, 50-µm sections were cut on a vibratome or a microtome. Sections through the host SCN and the graft were saved in series. One or two series of sections were Nisslstained with thionin. The other serial sections were stained immunocytochemically for the presence of VIP, AVP, and NPY according to the avidin-biotin peroxidase complex method. Sections were washed for 10 min in phosphate-buffered saline to remove excess aldehydes and then incubated with goat serum for 1 h to block nonspecific binding. Next, sections were incubated overnight at 4°C in primary antisera to either AVP (vasopressin-neurophysin II, Incstar), VIP (Incstar), or NPY (Incstar). Sections were then incubated with biotinylated secondary antibodies, followed by avidin-biotinylated peroxidase complex (Vectastain, Vector Laboratories). Diaminobenzidine was the chromogen substrate oxidized by horseradish peroxidase to form an insoluble precipitate. A mixture of glucose and glucose oxidase was used to generate the hydrogen peroxide substrate. Sections were mounted on gelatincoated slides and dried overnight. They were then dehydrated in graded alcohols, cleared in xylene, and put on coverslips with Permount. Control sections in which the primary antiserum was omitted completely eliminated the reaction product. The specificity of the antibodies was assessed by preabsorbing each antibody with a 10-µm solution of the appropriate antigen (AVP and VIP, Sigma; NPY, Bachem), which was sufficient in each case to block all immunoreactivity. For thionin staining, the cut sections were mounted on slides and air-dried overnight. Next they were hydrated through xylene, graded alcohols, and 0.1 M acetate buffer, and then stained in 0.1% thionin (Sigma) solution for 5–10 min. Then the sections were dehydrated, cleaned, and put on coverslips. All sections were examined under bright- and dark-field microscopy. The graft was considered nonviable if no cells were stained or only gliosis was present. A viable graft could have Nissl-stained cells but lack peptide staining. Data collection. Tb was recorded continuously through the implanted temperature telemeter, which emits a broad-band radio frequency pulse at a rate proportional to its temperature. This signal was converted to a Tb value by a microcomputer and stored on disk every 10 min. Activity was measured by monitoring cage movement. The movement of the rat displaced a piece of transparent plastic with an opaque dot pattern on it. This piece of plastic was located inside a photocell. The displacement triggered electrical pulses. The number of pulses was summed, stored, and sent to disk every 10 min. Drinking was measured using a standard drinkometer circuit. Wires from the water bottle and cage floor closed a circuit when the rat licked the drinking spout. The number of licks was summed and stored every 10 min. Data analysis. To examine the data for rhythmicity, computer-generated plots were drawn for Tb, activity, and drinking. Each 10-min data point was plotted as a black line if its value was above the daily mean of the entire day or a blank if its value was below the daily mean. The black line was half-length if the value of the data point was within 0.1°C of R1737 SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS Table 1. Summary of experimental conditions 15 Controls 6 Young rats with SCN grafts, no changes (M, n 5 6) 4 Old with good CRs, SCN grafts, no changes (M, n 5 3; F, n 5 1) 5 Old with poor CRs, CBL grafts, no improvements (M, n 5 3; F, n 5 2) 23 Old, poor CRs 9 Nonviable or absent grafts; no improvements (M, n 5 6; F, n 5 3) 14 Viable grafts 7 Improved (M, n 5 3; F, n 5 4) 7 Not improved (M, n 5 4; F, n 5 3) CR, circadian rhythm; CBL, cerebellar; M, male; F, female; n, sample size (total 5 38). RESULTS General findings. Of the 23 animals in the experimental group, 14 had viable grafts verified histologically (Table 1). Transplants restored or improved the freerunning (n 5 6) or entrained (n 5 1) rhythms in seven of them within 2–4 wk. Transplants of cerebellar tissue were ineffective in improving any rhythms (n 5 4; one graft was not viable). The improvements postgrafting occurred in one or more rhythms. Table 2 lists those rhythms that were restored or improved postgraft. To be accepted as ‘‘improved,’’ the rhythms had to have power at 24 h that was significantly greater than the white noise level and greater than the power pregraft (P , 0.01 in at least one rhythm that was poor or nonexistent pregraft). Well-developed grafts of various sizes were located within the third ventricle. The grafts were always attached to the host. Indeed, parts of the grafts often merged with the host tissue so well that it was difficult to determine the host-graft border (e.g., Fig. 1). The rostral-caudal position of the grafts varied from the level of the medial preoptic nucleus to the level of the median eminence (Table 2). To determine whether the size of the graft affected functional recovery, we compared the size of the successful grafts (ranging from 0.5 3 0.25 mm to 1.75 3 0.63 mm, n 5 7, Table 2) to that of the unsuccessful grafts (ranging from 0.6 3 0.5 mm to 1.7 3 0.75 mm, n 5 7, Table 3) using a binomial logit link regression analysis. The test revealed no differences in the sizes of successful and unsuccessful grafts (P , 0.14). Six of the seven successful grafts stained for both VIP and AVP. The seventh graft showed only VIP staining. Peptidergic organization of the host SCN of control and experimental animals was also examined. Although no systematic quantification was performed, there were Table 2. Successful transplantation: host and graft characteristics Pre-G Post-G Rat Sex Age, mo Lighting T A D T A D Latency, days G Level G Size, mm VIP AVP NPY Tumor, mm OF122 OF119 OM64 OF117 OM44 OF112 OM82 F F M F M F M 19.5 23 24 24 25 27 29.6 DD DD DD DD LD DD DD w s w w s w s w w w w w w w w a a a a w a >* s w >* s >* s <* w w >* >* w w <* >* >* >* >* >* >* 10 12 14 10 15 15 30 ME MPO/ME SCN SCN-RCH ME SCN-RCH SCN/RCH 1.73 3 0.63 0.60 3 0.50 0.50 3 0.25 1.00 3 0.25 0.75 3 0.25 0.70 3 0.40 0.50 3 0.25 11 11 1 11 1 11 1 11 11 1 2 1 11 11 2 11 1 2 2 11 11 53534 53433 Age, age of host at time of grafting; LD, 12:12-h light-dark cycle; DD, constant darkness; Pre-G, pregraft strength of body temperature (T), activity (A), and drinking (D) rhythms as determined by gray scales and periodograms; w, weak; s, strong; a, absent; post-G, postgraft; >*, stronger than pregraft (P , 0.05); <*, weaker than pregraft (P , 0.05); Latency, days until improvement of rhythm was apparent; G level, level of graft location in host brain; ME, median eminence; RCH, retrochiasmatic area; MPO, medial preoptic nucleus; SCN, suprachiasmatic nucleus; SCN-RCH, graft extending rostrocaudally from the level of SCN to the level of RCH; SCN/RCH, graft with separate pieces at level of the SCN and at level of RCH; G size, size of graft (length 3 width); 11, rich in peptide cells and/or fibers; 1, presence of peptide cells and/or fibers; 2, minimal or no peptide immunoreactivity; VIP, vasoactive intestinal polypeptide; AVP, arginine vasopressin; NPY, neuropeptide Y. All grafts were from embryonic day 18 (E18) or E17 fetal tissue. Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 the daily mean Tb, or within 5% of daily mean licks and activity measures. Consecutive days were plotted from top to bottom. For ease of visualization, two consecutive days were plotted on each line. The basic tool we used to analyze the rhythmic data is the periodogram, which describes how the variance in a time series may be accounted for by cyclic components at different frequencies. Periodograms reveal not only significant frequencies but also their relative strength, or power. Ten days of continuous data were used for periodogram analysis to determine the strength of the circadian rhythm. To compare relative power between individuals, or in the same individual at different times, the periodograms were normalized by equating the variance present in the data to one unit. For any periodicity to be significant at P , 0.01, the lower limit of the 99% confidence interval of the power estimate must exceed the white noise level. For ease of visualization, the lower limit of the 99% confidence interval of the power estimate was plotted instead of the actual power. To test for significant differences between the power at the circadian range pre- and postgrafting, we used a test procedure described by Diggle (8). It relies on the fact that the ratio of the two power values (spectral estimates) follows an F distribution. A graft was defined as successful when one or more circadian rhythms showed significantly increased power at 24 h postgraft compared with pregraft. We compared the histological data between successful and unsuccessful grafts using the nonparametric Wilcoxon rank sum test. To determine the effect of gender on functional recovery rate, we used the Pearson x2 test to examine the association of two variables expressed in a two-way contingency table. To determine the effect of age and graft size on the functional recovery rate, a binomial logit link regression analysis (22) was performed using SAS procedure PROC GENMOD. R1738 SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS no apparent differences in the numbers of VIP and AVP cells and NPY fibers between the aged rhythmic and aged arrhythmic host SCN. Grafts that were successful in restoring rhythms had significantly more VIP cell and/or fiber staining than did the unsuccessful grafts (P , 0.001). AVP and NPY staining was similar between successful and unsuccessful grafts (P , 0.1 and P , 0.17, respectively). Of the 23 old hosts with poor rhythms, 3 of 13 males and 4 of 10 females demonstrated functional recovery after SCN grafting. The recovery rates of the two sexes were not significantly different at P 5 0.05 (x2 5 0.30 , Table 3. Unsuccessful transplantation: host and graft characteristics Rat Sex Age, mo Lighting G Level G Size, mm VIP AVP NPY Tumor, mm OF118 OM113 OM93 OM106 OM91 OF116 OF113 F M M M M F F 23.5 24 24.3 24.3 25 25 26 DD DD DD DD LD DD DD SCN-RCH SCN SCN-RCH RCH-ME SCN-RCH SCN SCN 1.75 3 0.63 0.87 3 0.50 1.25 3 0.63 0.75 3 0.38 1.00 3 0.50 1.70 3 0.75 0.60 3 0.50 2 2 2 2 2 1 2 1 2 2 2 2 11 2 1 1 2 11 2 11 1 43632 12 3 8 3 5 73837 For explanation of symbols, see Table 2. All grafts were from E17 fetal tissue except for that of OM113, which was E19. Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 Fig. 1. Localization of neuropeptides in a functional SCN graft (OF112). Serial sections stained for vasoactive intestinal polypeptide (VIP; A), arginine vasopressin (VP; B), and neuropeptide Y (NPY; C). D and E: magnified views of cell clusters in A and B, respectively. F shows arginine vasopressin staining of a section through anterior portion of graft at level of host suprachiasmatic nucleus (SCN). There is an arginine vasopressin efferent fiber (large arrowhead) crossing the graft-host border (small arrowheads). VIP cell cluster is ventral to arginine vasopressin cell cluster (notched arrows in A and B). VIP and arginine vasopressin cells (arrows in D and E) have spherical or slightly elongated shapes with diameters of 5–15 µm. Fibers from these VIP and arginine vasopressin cells ramified within clusters and extended into other parts of the graft (arrows in A and B). NPY fibers (A) are rich in entire graft, including the region of the arginine vasopressin and VIP cell clusters. Calibration bar 5 200 µm (A–C) or 50 µm (D–F). G, graft; III, third ventricle; PVN, paraventricular nucleus; RCH, retrochiasmatic area. x21 5 3.84). The age of rats with functional recovery ranged from 19.5 to 29.6 mo, which overlapped the age of rats without recovery (from 23.5 to 26 mo). There was no significant impact of age on functional recovery. Four of five old rats with poor rhythms that had been implanted with cerebellar grafts had large, viable, well-vascularized grafts that did not stain for any of the three peptides. Four of six young intact controls and three of four old rats with good rhythms had peptidergic organization in their SCN grafts similar to the successful grafts. Functional SCN grafts. Grafts that improved or restored rhythms contained neuropeptides characteristic of the SCN in normal rats. All functional grafts contained groups of VIP-immunoreactive cells, which sometimes formed a cluster (Fig. 1, A and D) and sometimes were more loosely distributed (Fig. 2). These cells gave rise to fibers that occasionally formed a plexus surrounding the cell cluster (Fig. 1). These VIP cells, like those seen in an intact SCN, had spherical or slightly elongated somas, ,5–15 µm in diameter. In six of seven grafts, sections adjacent to those containing VIP cell clusters had AVP-positive neurons that, like those in an intact SCN, were parvocellular cells with diameters of 5–15 µm (Fig. 1). The AVP cells clustered close to VIP cells and their fibers also ramified within the graft (Fig. 1, B and E). This juxtaposition of VIP and AVP cells resembles the peptidergic organization in intact SCN. Innervation by NPY fibers was variable. In some grafts, the entire graft was richly innervated with NPY fibers in some grafts, whereas in others NPY immunoreactivity was minimal or absent. Nonfunctional SCN grafts. In 9 of 23 old rats with poor rhythms, the graft could not be located or was not viable (determined histologically). In seven old rats that failed to resume free-running rhythms despite viable SCN implants, the pattern of peptide immunostaining within the graft was completely lacking in two rats and sparse in the others (Table 3). Parvocellular VIP cells were absent in all but one graft. The graft in that rat, OF116, contained a small number of scattered VIP cells. However, this rat had a very large pituitary tumor. Pituitary tumors. In our colony, about one-third of rats age 24 mo or older develop pituitary tumors, which can grow quite large and at times severely depress the ventral brain. In extreme cases, such tumors caused SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS the animal to stop eating and drinking, resulting in rapid weight loss and death. In the present study, pituitary tumors were found in 8 of 32 old rats. Two old rats with small tumors showed improvements after SCN grafting (Table 2). From the presence of VIP cells in the graft, we would have predicted that OF116 (Table 3) should have recovered some rhythms. However, this rat had an extremely large pituitary tumor, ,480 mm3, which depressed the ventral brain severely. This tumor growth might have been responsible for the lack of recovery in this rat. Examples of improved rhythms in selected old rats. All three rhythms were improved in an old female rat (OF117) in which E17 SCN tissue was grafted when the rat was 24 mo old (Fig. 3). Tb and activity rhythms were poor before grafting and there was no drinking rhythm. About 10 days postgrafting, all three rhythms became stronger and their circadian power increased significantly over the pregraft level (P , 0.05). The graft in this rat was richly innervated with VIP cells. There was no evidence of AVP cells or NPY fibers. The drinking rhythm was also improved significantly (P , 0.05) after an E17 SCN graft at 23 mo in a female rat, OF119 (Fig. 4). Pregraft the Tb and activity rhythms showed significant 24-h periodicities. About 12 days postgraft, improvement in drinking started to become evident. The circadian power of drinking increased significantly (P , 0.05). The period of all three freerunning rhythms lengthened slightly postgraft. When the rat was switched to LD at 25 mo, all three rhythms synchronized to the LD, but this looks more like masking than entrainment (see Ref. 21). The graft of this rat was richly innervated with VIP and NPY (Fig. 2). Another old rat that was kept in LD throughout the experiment demonstrates that poor entrained rhythms may also be improved by an SCN graft. Figure 5 shows results in OM44, in which E18 SCN tissue was grafted at the age of 25 mo. Pregraft, only the Tb rhythm was robust, and it remained so for the duration of the measurements. The activity rhythm (poor pregraft) and the drinking rhythm (nonexistent pregraft) were both significantly improved postgraft. Two other rats maintained in LD showed no improvement. In one the graft was necrotic, and in the other no peptide staining was discernible. There were no improvements in circadian rhythms in old control rats bearing cerebellar grafts. No significant changes were seen in the host rhythms after SCN transplants in young intact controls or in old rats with good rhythms. DISCUSSION Our results demonstrate that fetal SCN grafts are able to improve or restore poor or absent circadian rhythms in aged rats that retain their own SCN. When all three rhythms were poor, as in OF117, all three were significantly improved by the graft. When only two rhythms were poor, as were activity and drinking in OM44, they were improved or restored and the Tb rhythm remained the same. In general, the circadian drinking rhythm was the worst of the three rhythms measured, and it was improved in six of seven old rats. One old rat, OF122, was unusual in that its Tb rhythm was the weakest pregraft. It became much stronger after the graft, while activity and drinking rhythms became worse. The worsening of some rhythms postoperatively was seen in old rats in all groups, including rats with cerebellar grafts and unsuccessful SCN grafts. We attribute this to the trauma of the anesthesia and operation and the poorer recovery of old rats. Our data are consistent with previous graft studies using adult rats with SCN lesions. Two such studies demonstrated that multiple circadian rhythms can be restored in the same individual. Boer et al. (2) found that fetal SCN grafts restored circadian rhythms of drinking, feeding, and wheel running in the same SCN-lesioned rats. In an abstract, Edgar et al. (9) reported that SCN grafts restored rhythms of sleepwake, Tb, and activity in concert in adult rats with SCN Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 Fig. 2. Neuropeptide profile of a functional SCN graft (OF119) consisting of 3 pieces. One piece (D) was at the level of medial preoptic nucleus. The other 2 pieces attached to the top and the bottom of the 3rd ventricle, at the level of the median eminence (ME; A). Adjacent sections stained for Nissl (A), NPY (B), and arginine vasopressin (C). D is a section through the anterior piece of the graft stained for VIP. There were both a VIP cell and/or fiber cluster (arrowhead in D) and an arginine vasopressin cell and/or fiber cluster (arrow in C). Magnocellular arginine vasopressin fibers (arrowhead in C) and NPY fibers (arrowhead in B) ramified extensively throughout the graft. Calibration bar 5 100 µm (C and D) or 200 µm (B) or 500 µm (A). AHP, anterior hypothalamus, posterior part; VMH, ventromedial hypothalamic nucleus; DA, dorsal hypothalamic area. R1739 R1740 SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS lesions. Lehman et al. (18) reported that although circadian locomotor rhythms were restored in SCNlesioned hamsters, reproductive responses to photoperiod and the ability to entrain to light at intensities to which SCN-intact hamsters respond were not. Taken together, these data indicate that the requirements for restoring individual rhythms may be different. Improvements in circadian rhythms were evident from 10 to 30 days after transplantation. This time course is comparable to the latency to recovery after grafting in young rats and hamsters, which ranges from several days to 2 mo (1, 18). Postgraft improvements were seen in both male (3 of 13 rats) and female (4 of 10 rats) old hosts, at ages ranging from 19.5 to 29.6 mo. Functional grafts differed widely in size, ranging from 0.5 3 0.25 mm to 1.75 3 0.63 mm in cross section. There were no differences between successful and unsuccessful grafts in these three parameters. In other words, neither host gender, age, nor graft size were crucial in predicting graft success. There was no relation between graft placement in the third ventricle and improvements in circadian rhythmicity. In two successfully transplanted animals, the grafts were located in the third ventricle at the level of the median eminence, distant from the host SCN. Conversely, six of seven of the unsuccessful grafts were located at the level of the host SCN (Table 3). This finding agrees with previous work in young SCNlesioned hamsters (37). We noted sparse and limited neural connections between the functional grafts and the host brain. Even in cases where the grafts maintained physical contact with the host SCN region, fiber connections with the host SCN or with the SCN target areas were rarely observed. Similar observations have been reported in studies of grafting in SCN-lesioned animals using Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 Fig. 3. Gray scales (top) and periodograms pregrafting (middle) and postgrafting (bottom) of body temperature (Tb; T), activity (A), and drinking (D) of a successfully transplanted old female rat, OF117 in constant darkness. Embryonic day 17 (E17) SCN tissue grafted at 24 mo (G). All 3 rhythms were poor before grafting. Periodograms show significant but low power at 24 h for Tb and activity; there is no significant 24-h power for drinking. About 10 days postgrafting, all 3 rhythms became stronger and power at 24 h increased significantly over pregraft levels (* P , 0.05). Dark bars at top indicate darkness. Vertical bars on left of Tb plots indicate data segment used for computing the periodograms. Numbers on right indicate age (mo) of the host. In the periodograms the power value plotted is the lower limit of the 99% confidence interval of the actual power. Thin horizontal line above x-axis of each periodogram is the white noise level. Any power value higher than the white noise level is significant at P , 0.01. SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS R1741 Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 Fig. 4. Same as Fig. 3 for a successfully transplanted old female rat, OF119. E17 SCN tissue grafted at 23 mo. Pregraft there was significant 24 h periodicity in Tb and activity but none in drinking. About 12 days postgrafting, improvement started to show in drinking. Circadian power of drinking began to improve and increased significantly (* P , 0.05) compared with the pregrafting level. Activity did not improve. When the rat was 25 mo old the photoperiod was switched to a 12:12-h light-dark cycle (LD). All rhythms became synchronized with LD in ,2 wk. a, b, and c indicate data segments for the 3 periodograms. peptidergic staining (18) or dil tract tracing (4). In studies using xenografts and donor-specific markers, more extensive anterior hypothalamic graft efferents were found (16, 31). However, even with the use of such sensitive techniques, efferents from anterior hypothalamic grafts are variable and there is no clear correlation between the pattern of innervation and recovery of function (31). The presence of VIP cells in the graft appeared to be critical for functional improvements in aged rats. All functional SCN grafts showed positive VIP staining and had significantly more VIP cell and/or fiber staining than did the unsuccessful grafts. On the other hand, AVP and NPY staining in grafts were not consistently correlated with functional improvements: AVP cells were absent in one functional graft and NPY was R1742 SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS absent in three (Table 2). In the aged host SCN, VIP-positive cells were frequently observed but it seemed that the presence or absence of VIP in the host SCN did not determine the preservation of rhythmicity in the aged rats. Possibly, differences in characteristics other than cell numbers, such as biochemical, electrophysiological, or connectivity properties of VIP, alone or in combination with other peptide cells or trophic factors, may underlie the differences in ability to sustain circadian rhythms in old age. The importance of VIP neurons in young animals has been noted in the transplantation literature. Sollars and Pickard (32) found dense aggregations of VIP neurons in SCN-lesioned hamsters in which circadian locomotor rhythms were restored by SCN transplants. This was true for hamster-to-hamster and mouse- or rat-to-hamster grafts. In hamsters the presence of VIP cells and fibers is the most consistent marker in functional SCN grafts, whether whole tissue grafts (12) or dispersed cell suspensions (30). Kaufman and Menaker (15) reported a high correlation between the presence of VIP cells in the SCN graft and restoration of running-wheel rhythms in SCN-lesioned hamsters. Our experiments were not designed to determine whether the improved rhythms in aged rats are generated solely by the grafts or are rejuvenating the host SCN, or some combination. Using senescent mutant and wild-type hamsters, Hurd et al. (13) observed various interactions between grafted and host SCN. Because our grafts were homografts, it is not surprising Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 Fig. 5. Same as Fig. 3 for a successfully transplanted old male rat, OM44. E18 SCN tissue grafted at 25 mo. This rat was maintained in LD throughout the experiment. Pregraft there was significant circadian periodicity in Tb and activity (* P , 0.05). About 15 days after grafting, the drinking rhythm was restored, the activity rhythm was significantly increased, and the Tb rhythm did not change. SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS to find that the improved rhythms had periods similar to the pregrafting periods. In summary, we have demonstrated that fetal tissue containing the SCN can restore multiple circadian rhythms to aged animals whose rhythms are poor or absent. Control cerebellar grafts did not improve any rhythms in any aged rats. One major difference between the successful grafts and unsuccessful grafts was that the former had VIP cell and/or fiber staining whereas the latter did not. Thus the degeneration of circadian rhythms in aged rats may be due, at least in part, to deterioration of the aged SCN and in particular to a loss of function of VIP-containing neurons. The present study is one of the few studies examining the functional recovery of multiple circadian rhythms simultaneously in the same individuals after implantation of fetal SCN tissue. This technique is potentially a powerful tool for mechanistic investigations of the circadian system and of the aging process, but much remains to be explored. For example, many studies in grafted adult rodents with SCN lesions show that the presence of VIP-staining cells is an important indicator of recovery. But does VIP play an important role in the functional recovery following grafting in SCN-lesioned animals or in rhythm-degenerated aged animals, or is it simply a marker? By manipulating the material being grafted, e.g., cultured VIP neurons, or by varying the host brain milieu, it should be possible to determine whether VIP is indeed a regulator of the circadian system. The improvement or restoration of circadian rhythms in intact aged animals following fetal SCN grafting strongly implies that changes in the SCN are, at least in part, the cause of age-associated deterioration in behavior. Our results add another dimension to the accumulating data base showing that neural transplants can ameliorate neurological, behavioral, and cognitive deficits associated with aging. For example, fetal substantia nigra grafts alleviated motor deficits in old rats (11), septal grafts improved spatial learning ability (10), and raphe grafts improved specific electrophysiological impairments (19) in aged rats. Neural transplantation is both a powerful tool to investigate the aging processes and a promising therapeutic strategy in treating afflictions associated with human aging like Parkinson’s disease and Alzheimer’s disease. We thank Dr. JianMing Ding for teaching us immunocytochemistry and how to evaluate histological materials; Dr. Rae Silver for teaching us fetal tissue transplantation technique; Elizabeth Peloso and Dr. Maciek Wachulec for their generous help with the surgical operations, data line maintenance, animal care, and data analysis; Robert Peloso for consultation with time-series analysis; Esther Williams for using her time-series software; and Dr. Michael Price for writing data analysis software. This work was supported by National Institute of Mental Health Grant RO1-MH-41138 and an Alzheimer’s Association/Willard and Rachael Olsen Pilot Research Grant to E. Satinoff. Address reprint requests to E. Satinoff. Received 5 May 1997; accepted in final form 7 August 1998. REFERENCES 1. Aguilar-Roblero, R., S. Shibata, J. C. Speh, R. DruckerColı́n, and R. Y. Moore. Morphological and functional development of the suprachiasmatic nucleus in transplanted fetal hypothalamus. Brain Res. 580: 288–296, 1992. 2. Boer, G. J., H. A. Griffioen, H. Duindam, T. P. van der Woude, and W. J. Rietveld. Light/dark-induced effects on behavioral rhythms in suprachiasmatic nucleus-lesioned rats irrespective of the presence of functional suprachiasmatic nucleus brain implants. J. Interdiscip. Cycle Res. 24: 118–136, 1993. 3. Cai, A., M. N. Lehman, J. M. Lloyd, and P. M. Wise. Transplantation of fetal suprachiasmatic nuclei into middle-aged rats restores diurnal Fos expression in host. Am. J. Physiol. 272 (Regulatory Integrative Comp. Physiol. 41): R422–R428, 1997. 4. Canbeyli, R. S., M. N. Lehman, and R. Silver. Tracing SCN graft efferents of with Dil. Brain Res. 554: 15–19, 1991. 5. Card, J. P., N. Brecha, H. J. Karten, and R. Y. Moore. Immunocytochemical localization of vasoactive intestinal polypeptide-containing cells and processes in the suprachiasmatic nucleus of the rat: light and electron microscopic analysis. J. Neurosci. 11: 1289–1303, 1981. 6. Card, J. P., and R. Y. Moore. Ventral lateral geniculate nucleus efferents to the rat suprachiasmatic nucleus exhibit avian pancreatic polypeptide-like immunoreactivity. J. Comp. Neurol. 206: 390–396, 1982. 7. Chee, C. A., B. Roozendaal, D. F. Swaab, E. Goudsmit, and M. Mirmiran. Vasoactive intestinal polypeptide neuron changes in the senile rat suprachiasmatic nucleus. Neurobiol. Aging 9: 307–312, 1988. 8. Diggle, P. J. Time Series: A Biostatistical Introduction. New York: Oxford University Press, 1990. 9. Edgar, D. M., M. R. Ralph, W. F. Seidel, L. K. Lee, and W. C. Dement. Fetal SCN-transplants restore sleep-wake and body temperature circadian rhythms in SCN-lesioned rats (Abstract). Sleep Res. 21: 371, 1992. 10. Gage, F. H., A. Bjorklund, U. Stenevi, S. B. Dunnett, and P. A. T. Kelly. Intrahippocampal septal grafts ameliorate learning impairments in aged rats. Science 225: 533–536, 1984. 11. Gage, F. H., S. B. Dunnett, U. Stenevi, and A. Bjorklund. Aged rats: recovery of motor impairments by intrastriatal nigral grafts. Science 221: 966–969, 1983. 12. Griffioen, H. A., H. Duindam, T. P. van der Woude, W. J. Rietveld, and G. J. Boer. Functional development of fetal suprachiasmatic nucleus grafts in suprachiasmatic nucleuslesioned rats. Brain Res. Bull. 31: 145–160, 1993. 13. Hurd, M. W., K. A. Zimmer, M. N. Lehman, and M. R. Ralph. Circadian locomotor rhythms in aged hamsters following suprachiasmatic transplant. Am. J. Physiol. 269 (Regulatory Integrative Comp. Physiol. 38): R958–R968, 1995. 14. Kalsbek, A., and R. M. Buijs. Peptidergic transmitters of the suprachiasmatic nuclei and the control of circadian rhythmicity. Prog. Brain Res. 92: 321–333, 1992. 15. Kaufman, C. M., and M. Menaker. Effect of transplanting suprachiasmatic nuclei from donors of different ages into completely SCN-lesioned hamsters. J. Neural Transplant. Plast. 4: 257–265, 1993. 16. Lehman, M. N., J. LeSauter, C. Kim, S. J. Berriman, P. A. Tresco, and R. Silver. How do fetal grafts of the suprachiasmatic nucleus communicate with the host brain? Cell Transplant. 4: 75–81, 1995. 17. Lehman, M. N., R. Silver, and E. L. Bittman. Anatomy of suprachiasmatic nucleus grafts. In: Suprachiasmatic Nucleus: The Mind’s Clock, edited by D. C. Klein, R. Y. Moore, and S. M. Reppert. New York: Oxford University Press, 1991, chapt. 19, p. 349–374. 18. Lehman, M. N., R. Silver, W. R. Gladstone, R. M. Kahn, M. Gibson, and E. L. Bittman. Circadian rhythmicity restored by neural transplant. Immunocytochemical characterization of the graft and its integration with the host brain. J. Neurosci. 7: 1626–1638, 1987. 19. Levkovitz, Y., V. Greenberger, and M. Segal. The effects of raphe grafts on hippocampal electrophysiology in aged rats. Brain Res. 719: 234–238, 1996. Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 Perspectives R1743 R1744 SCN TRANSPLANTS RESTORE CIRCADIAN RHYTHMS IN OLD RATS 31. 32. 33. 34. 35. 36. 37. 38. circadian rhythmicity in SCN-lesioned adult hamsters. Brain Res. 525: 45–58, 1990. Sollars, P. J., and G. E. Pickard. Neural heterografts as a model for the study of mammalian circadian behavior. In: Neural Transplantation, CNS Neuronal Injury/Regeneration, edited by J. Marwah, H. Teitelbaum, and K. N. Prasad. Boca Raton, FL: CRC, 1994 p. 161–181. Sollars, P. J., and G. E. Pickard. Vasoactive intestinal peptide efferent projections of the suprachiasmatic nuclei in anterior hypothalamic transplants: correlation with functional restoration of circadian behavior. Exp. Neurol. 136: 1–11, 1005. Sutin, E. L., W. C. Dement, H. C. Heller, and T. S. Kilduff. Light-induced gene expression in the suprachiasmatic nucleus of young and aging rats. Neurobiol. Aging 14: 441–446, 1993. Van den Pol, A. N., and K. L. Tsujimoto. Neurotransmitters of the hypothalamic suprachiasmatic nucleus: immunocytochemical analysis of 25 neuronal antigens. Neuroscience 15: 1049– 1086, 1985. Van Reeth, O., Y. Zhang, P. C. Zee, and F. W. Turek. Grafting fetal suprachiasmatic nuclei in the hypothalamus of old hamsters restores responsiveness of the circadian clock to a phase shifting stimulus. Brain Res. 643: 338–342, 1994. Viswanathan, N., and F. C. Davis. Suprachiasmatic nucleus grafts restore circadian function in aged hamsters. Brain Res. 686: 10–16, 1995. Vogelbaum, M. A., J. Galef, and M. Menaker. Factors determining the restoration of circadian behavior by hypothalamic transplants. J. Neural Transplant. Plast. 4: 239–256, 1993. Wise, P. M., I. R. Cohen, N. G. Weiland, and E. D. London. Aging alters the circadian rhythm of glucose utilization in the suprachiasmatic nucleus. Proc. Natl. Acad. Sci. USA 85: 5305– 5309, 1988. Downloaded from http://ajpregu.physiology.org/ by 10.220.32.247 on August 10, 2017 20. Li, H., and E. Satinoff. Changes in circadian rhythms of body temperature and sleep in old rats. Am. J. Physiol. 269 (Regulatory Integrative Comp. Physiol. 38): R208–R214, 1995. 21. Matsumoto, S., J. Basil, A. E. Jetton, M. N. Lehman, and E. L. Bittman. Regulation of the phase and period of circadian rhythms restored by suprachiasmatic transplants. J. Biol. Rhythms 11: 145–162, 1996. 22. McCullagh, P., and J. A. Nelder. Generalized Linear Models. New York: Chapman & Hall, 1989. 23. Mosko, S. S., G. F. Erickson, and R. Y. Moore. Dampened circadian rhythms in reproductively senescent female rats. Behav. Neural Biol. 28: 1–14, 1980. 24. Refinetti, R., H. Ma, and E. Satinoff. Body temperature rhythms, cold tolerance, and fever in young and old rats of both genders. Exp. Gerontol. 25: 533–543, 1990. 25. Richardson, G. S. Circadian rhythms and aging. In: Handbook of the Biology of Aging, edited by E. L. Schneider and J. W. Rowe. New York: Academic, 1990, p. 275–305. 26. Roozendaal, B., W. A. van Gool, D. F. Swaab, J. E. Hoogendijk, and M. Mirmiran. Changes in vasopressin cells of the rat suprachiasmatic nucleus with aging. Brain Res. 409: 259–264, 1987. 27. Rusak, B. The mammalian circadian system: models and physiology. J. Biol. Rhythms 4: 121–134, 1989. 28. Satinoff, E., H. Li, T. K. Tcheng, C. Liu, A. J. McArthur, M. Medanic, and M. U. Gillette. Do the suprachiasmatic nuclei oscillate in old rats as they do in young ones? Am. J. Physiol. 265 (Regulatory Integrative Comp. Physiol. 34): R1216–R1222, 1993. 29. Satinoff, E., and R. A. Prosser. Suprachiasmatic nuclear lesions eliminate circadian rhythms of drinking and activity, but not of body temperature in male rats. J. Biol. Rhythms 3: 1–22, 1988. 30. Silver, R., M. N. Lehman, M. Gibson, W. R. Gladstone, and E. L. Bittman. Dispersed cell suspensions of fetal SCN restore