Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
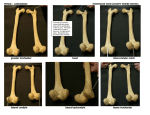
AN UNUSUAL THEROPOD FROM THE UPPER CRETACEOUS OF MONGOLlA by S. M. Kurzanov In: Iskopayemyye pozvonochnyye Mongolii (Fossil Vertebrates of Mongolia). Trudy Sovmestnay Sovetsko-Mongolskay Paleontologiyeskay Ekspeditsiy (Joint Soviet-Mongolian Paleontological Expedition) 15:39-49. Nauka Moscow, 1981 Translated by Catherine Siskron and Samuel P. Welles minor editing by Matthew Carrano, January 2000 During the exploration of the Joint Soviet-Mongolian paleontological expedition in the summer of 1973, at the Udan-Sayr (southern Gobi) location, was found a fairly complete skeleton of a bird-like theropod, which belonged to the new genus and species Avimimus portentosus. With the exception of a crushed skull fragment, all the remaining bones are very well preserved. The location of Udan-Sayr is 75 km. south of Hovd-somon Ubur-Hangayskaymak, in the foothills of the Gurvan-Sayhan mountain range. The osseous layers of this location are in the red colored sands and sandstone with rare interlayers of clay and conglomerates. The visible thickness of the deposits is approximately 15 m. The age of the deposits can be established as Nemegetinsk. This is indicated by the presence in it of teeth of Tarbosaurus, a carnosaur, known from the deposits of that age from various locations in the Southern Gobi. Besides the Udan-Sayr location, Avimimus remains were also found in the Shara-Tsav location, 7 km north from the Bayshin-Tsav in the southeastern Gobi. Besides this, here was also found an almost complete skeleton of the ornithomimid Gallimimus (classification by Barsbold) and the skull of the sauropod Nemegtosaurus, which are also highly characteristic for the Nemegetinsk deposits of the southern Gobi. The fauna found in this region clearly indicates that the deposits in this location are of the Nemegetsk suite. The discovery of Avimimus is of particular interest. On one hand this is the first dinosaur that has such clearly expressed bird features, in such large numbers, on the other it is also the first among the theropods with such unusual structure of the pelvis. Such a combination of unique features places Avimimus in a category all its own among the dinosaurs and places it in a new family, Avimimidae, fam. nov. SUBORDER THEROPODA SUPERFAMILY COELUROIDEA FAMlLY AVIMIMIDAE KURZANOV, FAM. NOV. Diagnosis. Small theropods, 1-1.5 m in length. The skull is small with relatively large orbits. The frontal and parietal bones are fused. The cervical vertebrae increase noticeably in site as they approach the trunk. On the last two cervical and the first three dorsal vertebrae, the hypapophyses are developed. There are no fewer than 11 cervfcal vertebrae. The pelvis is wide. The ilium, ischium and pubis are fused around the acetabular (orig. “vertlugian”) cavity. The posterior branches of the ilium are directed almost horizontally. On the femora are developed special additional condyles for the articulation with the fibula. The latter are highly reduced and are fused with the tibia at their distal ends. The bones of the distal row of the tarsus are fused with the metatarsus. The second and fourth metatarsals fuse proximarly with each other, the fifth fuses fully with the fourth. the phalanges of the external toes are highly shortened. Family composition. Single genus Avimimus gen. nov. Comparison. The closest to avimimids are members of the family Ornithomimidae. The number of cervical vertebrae in ornithomimids is the same as that of other theropods, and does not rise above 10, while Avimimus has no fewer than 11. Until now hypapophyses were not observed on cervical and dorsal vertebrae among theropods, but they are present in the case of Avimimus. The same is true of the fusion of the parietal and frontal bones. One of the main features, which differentiates avimimids not only from ornithomimids, but also from other theropod families, is the structure of the pelvis and the forelimbs. The rather wide pelvis of Avimimus, if one is to judge by the distance between the acetabular cavities – in relative dimensions, it is almost twice as wide as that of ornithomimids. Practically all theropods have an ischium that is directed posteriorly, and only in Avimimus is it projected anteriorly and ventrally, parallel to the pubis. Also, all theropods, with the exception of Avimimus, have pubic bones which participate in the formation of the acetabular cavity. Another first among theropods is that Avimimus has an additional third condyle on its femur. Undoubtedly, among features peculiar to Avimimus is the full fusion of the proximal row of bones of the metatarsus, the proximal fusion of metatarsals II, IV and V, and the highly shortened external toes, Such features are also completely unknown among theropods. Genus Avimimus Kurzanov, gen. nov. Name of genus. From avis, Latin, bird and mimus, Latin, to mimic (?), Type species. Avimimus portentosus, Upper Cretaceous, Nemegetinsk suite, Mongolian National Republic. Species composition. Monotypic genus. Distribution. Upper Cretaceous, MNR. Avimimus portentosus Kurzanov, sp. nov. Name of species. From portentosus, Latin, unusual. Holotype. PIN, #3907/1, incomplete skeleton; MNR, Udan-Sayr, Upper Cretaceous, Nemgetinsk suite. Material. Besides the holotype, paratype #3906/1 from the location Shara-Tsav, individual fragments of the postcranial skeleton; MNR, Upper Cretaceous, Nemgetinsk suite. Description. Almost all the vertebrae, girdle of the forelimbs, and pelvis were found separated, while the rest of the bones were found articulated. They were found in the perimeters of a rather small area, and undoubtedtly belong to the same specimen, the most characteristic in size. At the Udan-Sayr location, Shara-Tsav, were discovered at the same time a large quantity of individual bones from other specimens of Avimimus, practically all of similar dimensions. Skull (fig. 1). Only the badly crushed skull roof was preserved along with part of the occipit and the lower part of the left wall of the cranium. Some bones were displaced. However, the fact that they mostly preserved the shape and surface, allowed a partial reconstruction of the skull of Avimimus. The skull is small, its length from the top of the upper occipit to the begining of the conjectural nasal openings is approximately 50 mnm. In addition to this, the possible size of the orbit is no less than 25 mn, that is the orbit, as in the case of birds, is proportionally very large, and started also as in the case of birds almost immediately behind the nasal opening. The upper temporal cavities are very small, their greatest diameter was not above 10 mm. Judging by the preserved posterior edge, the nasal openings were narrow and elongated in length. The frontal and temporal bones are fully fused with each other, sutures with adjoining bones are partially obliterated. They form the convex skull cupola and the medial parts of the upper temporal fenestrae. The single fronto-temporal bone extended anteriorly almost to the nasal openings. The postorbital bones form the posterior half of the upper edge of the orbit. Medially they become quite thick. Between them and the fronto-parietal on both sides there is a bend. Anteriorly they rest on the posterior part of the lacrimals. Lacrimals comprise the anterior half of the upper part of the orbit. apparently, with a descending process. Anteriorly they rest on the nasals. Narrow, short, From the nasals were preserved only the proximal part of the left nasal with the surrounding smooth cavity, gradually passing, apparently, into the nasal opening. It is important to note that the supposedly long, narrow nostrils were located not so much on the lateral as on the dorsal part of the skull, in a position very close to that of the birds. The upper occipital bone occupied almost the whole upper half of the occipital curface. In the middle of its upper edge is located a small, symmetrically located swelling. As a whole the bone is wide and flat, and forming a considerable part of the occipit, makes it also flat and even, as in birds. Ventrally the supraoccipital fuses firmly with the exoccipitals, anteriorly and laterally with the prootic. The highly fragmented prootic, which forms the left wall of the endocranial cavity, is visible only from the medial side. A considerable part of it is occupied by the lower half of the large circular cavity, located directly inside the osseous labyrinth (along its edge pass the semicircular canals) and the corresponding recessus intracucticus of some theropods (Kurzanov, 1976). An analogous formation is noted also in the cranium of the Upper Cretaceous sea bird Hesperornis regalis (Marsh, 1880), in which the endocranium as a whole is similar to that of theropods from the famny Itemiridae. This structure is present also in contemporary birds, although it is relatively smaller in size, and accommodates special excrescence of the cerebelum – flocculi. Apparently, an analogous function was performed by the otic cavity of Avimimus, and there is no basis to associate its existance with the lymphatic or venous systems, as was supposed earlier (Kurzanov, 1976). Immediately under the otic cavity is located a horizontally elongated cavity, on the bottom of which are located two openings for the otic nerve, through the anterior of which passes the vestibular, and posteriorly the cochlear nerve. Directly anteriorly and somewhat above it is located the opening for the facial nerve. Posteriorly from the otic capsule, the endocranial cavity remains equally high, and the medial curvature of the otic bone indicates that it becomes narrower. Ventrally, immediately behind the otic capsule is located a narrow high fissure of the jugular opening, and directly in front of it a snlall opening for the hypoglossal nerve. The shape and relative dimensions of the preotic bones, and also of the fronto-temporal allow to speculatively judge the shape of the brain. The cupola-shaped and wide roof of the skull, and the lateral curvature of the anterior part of the preotic bones, indicate an unusual development of the large hemispheres of the brain. If one is to judge by the height of the preotic bone, the endocranial cavity is relatively large and high behind the otic capsule, as is the cerebellum. Such a structure of this part of the cranium suggests the possibility that the height of the endocranium may be connected with the double curvature of the elongated brain, which is characteristic for birds. The small pieces preserved of the orbitosphenoid bones carry large foramina for the optic nerves. The orbitosphenods are almost at a right angle and meet in the middle, where they contact the unpaired ethmoid. The latter at this point becomes thin, forming the fissura craniofacialis, which separates the facial part of the skull from the cranium. Further forward the ethmoid suddenly thickens. The sutures between the orbitosphenoids and the ethmoid (?) are obliterated. Vertebral column (fig. 2. Only seven cervical vertebrae have been preserved (2, 4, 6, 8-11), and also nine dorsal vertebrae, in sequence. The vertebrae of both sectors are platycoelous, and the articular surfaces are slightly concave. The dimensions of the cervical vertebrae slightly increase toward the trunk. The length of the axis is 17 mm, that of the ninth vertebrae is 24 mm. The change in the size of the vertebrae allowed us to establish their approximate number: a minimrirn of 11 cervical vertebrae. The body of the vertebra is elongated. In the middle is located a fairly large oval opening, which leads to the cavity inside the body. On vertebrae from 6th to the ninth an additional opening is located on the anterior edge. The lower surface of the bodies, starting with the axis, with a pointed crest in the middle, which gradually flattens, and then on the tenth vertebrae again becomes pointed and remains pointed on all the dorsal vertebrae. On the anterior edge of the supposedly last (11th) cervical vertebra this crest immediately becomes the hypapophysis, fairly thick, with a rounded end. Judging by the site of this hypapophysis, it is perfectly logical to suppose that a somewhat smaller one was found also on the tenth vertebrae, the ventral surface of which was destroyed. The arches are low, with very short transversal processes, immediately under which is found a cavity with three sections. The articular surfaces of the majority of the cervical vertebrae are not displaced in relation to one another; and only the fourth (possibly, that this was true also of the 3rd and 5th which were not preserved) the anterior articular surface is raised dorsally in relation to the posterior, creating the necessary natural curvature of the neck. Dorsal vertebrae, in differentiation from the cervicals, are of equal dimensions. The body of the first vertebrae in cross-section have an elongated-oval shape, while close to the tail they become rounded. Up to the 9th dorsal vertebra a pointed crest is present on the ventral surface. It is especially well expressed on the dorsal vertebrae, where on the first and third it turns into a hypapophysis on the anterior edge, a very large one on the first. The articular facets of the rib head are well expressed and gradually move from the anterior edge to the middle of the body (first vertebra) to the level of the anterior articular processes, to the arch (starting with the seventh). In the articulated state the dorsal vertebrae form a gently sloping arch. The lateral sides of the body at first are almost flat, then become more and more curved in the anteroposterior direction. The neural canal is relatively large, its diameter is approximately two times smaller than than the body af the vertebra. The anterior articular processes are short, barely project beyond the surface of the body. As is apparent from the description, the most characteristic features of the vertebra of Avimimus are: considerable increase in the length of the cervical vertebrae and the first dorsal vertebrae, the appearance of which is probably connected with the strenghtening of the m. rectus capitis. Pectoral girdle (fig. 3). The scapula and the coracoid are fused without a visible suture. Only a wide strip, slightly roughened, which passes vertically from the middle of the upper edge of the glenoid cavity, indicates that place (where the suture ought to be). The scapular part, just as is the case in all theropods, is wide ventrally and slightly more sharply narrowed in the dorsal part. What is important is that there are no sharply expressed boundaries between these parts, which is most characteristic of birds and theropods from the family Dromeosauridae. The glenoid cavity is fairly narrow, elongated, directed straight down, thus indicating the position of forelimbs vertically below, directly under the body. The supraglenoidal thickening is not very well expressed. The most characteristic peculiarity of the coracoidal part is, apparently, its elongated shape. Thanks to this it is possible to approximately measure the angle between the scapula and the coracoid – a blunt angle of approximatety 160° which is quite characteristic for running birds, which is also true of the fact that the coracoid is elongated. In front and below the coracoidal opening is located a large pointed tubercle, the existence of which is probably connected with the attachment of m. biceps brachii, the most powerful flexor of the antebrachium. Directly below this tubercle is located a large concave surface – most likely the area of attachment of m. coracohumeralis. Humerus (fig. 3). The humerus of Avimimus is noticeably differentiated in structure from that of small theropods. The deltopectoral crest is well developed and its length is greater than 1/3 the length of the humerus. On its dorsolateral surface, along the entire length of the thickening, are noted the beginnings of m. deltoideus, which performed the function of pronation and supination of the shoulder. On the distal end of the deltopectoral crest remained a small swelling from the m. pectoralis. On the proximal end of the humerus dorsally was preserved a rough, slanted, wide strip of m. coracohumeralis, which passed from the upper edge of the deltopectoral crest in the direction of the tuberculum mediale. On the dorsolateral surface, exactly in the middle, is located a slanted seam – an approximate line for the attachment of m. triceps brachii, and quite possibly also of m. latissimus dorsi. It is possible that the most important feature of the Avimimus humerus is the characteristic three-segmented distal epiphysis, which is divided into three unequal parts. The largest of them is the one in the center, and the one that projects the most is the medial. Most likely the middle condyle, being the largest, corresponds to the ulnar, while the lateral corresponds to the radial, which is indicated by its position in relation to the ulnar condyle and also its elongated shape, which allows a minor displacement of the proximal end of the radius during pronation and supination. The deve!opment of the third, medial part of the distal epiphysis, which cannot be named a condyle and does not exist in other theropods, possibly is connected with the attachment of muscles, which accomplish pronation of the antebrachium – m. pronator and and flexor – flexor carpi and metacarpi. A quite similar construction of this sector of the humerus can be found among birds. Immediately above the central condyle, the distal joint the humerus is noticeably indented, which apparently considerably increased the flexing possibilities of the radial joint. Imnediately above was found the beginning of m. brachialis anticus, the most powerful flexor of the antebrachium. The external edge directly above the radial condyle is greatly thickened and rough, indicating the attachment of m. extensor digitorum comnunis. Immediately below, directly to the lateral condyle, is attached a supinator, simultaneously also the flexor of the antebrachium. As can be gathered from the above information, the clearest muscle imprints are connected mostly with the flexors of the antebrachium, flexors and extensors of the hand. The approximate, due to the incomplete preservation of the forelimbs, analysis of the shoulder muscles indicates a definite tendency to the amplification and possibly predominance of the flexing-extending motion of the limb. The ulna, of which we have only the compressed proximal epiphysis with a relatively low olecranon process. On the external side, closer to the anterior edge, near the joint is located a triangular rough surface for the m. brachialis anticus. From the internal side, adjoining the radius, the ulna is slightly concave. Pelvic girdle. The hind limbs of Avimimus are long and slender, with a tibia considerably enlarged relation to other sectors. The length of the femur has the ratio to the tibia and to the length of the metatarsus of 1:1.37:0.81. The distal parts of the limb sectors are noticeably thinner than the proximal, just as the whole limb becomes thinner distally. All articular surfaces are very smooth, The femur (fig. 4) is insignificantly curved, with very well defined joints. Its length is 188 mm. The head is ball shaped, with a rounded cross-section thinner than the diameter of the head and neck. It is located at a right angle to the axis of the femur, so that just like the shape of the head, it is highly characteristic for the majority of birds. A little above the middle of the medial surface of the head is located a small indentation for the attachment of ligamentum teres. A very large greater trochanter surpasses the head 1.5 times in width. It is separated by a deep, narrow fissure from the lesser trochanter, which is a high, subquadrate in cross-section, projection, which does not project up beyond the surface of the greater trochanter. On the lateral side of the latter is located a triangular outgrowth, apparently connected with m. ilio-femoralis. The distal condyles of the femur are large, the external is more developed, thus establishing the position sector of the limb – somewhat medially, under an obtuse angle to the femur. The intercondyle furrow is wide, with a flat bottom, forming a large area for the cruciate ligaments of the knee joint, analogous to that of birds. A remarkable feature of Avimimus, which is found for the first time.among dinosaurs in general, is the development of a special additional supracondyle on the external condyle of the distal epiphysis of the femur, which forms the joint for the reduced fibula. The proximal head of the latter moved in the furrow between the additional supracondyle and the lateral condyle. Apparently, this is connected with the reduction of necessity in the rotary motions and the strenghtening of the joint at the expense of the full transfer of support to the tibia. From the internal condyle, along the anterior side of the femur, passes a low, but clearly defined, crest, apparently one that marked the line of attachment of m. femorotibialis. On the posterior side,.immediately above the distal joint is located a relatively deep fossa, which is also found in birds – fossa poplitea. A moderately well developed fourth trochanter is displaced proximally. The tibia (see fig. 4), 257 mm long, is in every sense of the word a tibiotarsus due to the full fusion with proximal elements of the tarsus. Such a structure was previously observed only among birds. There is no condyle between them, and they fuse smoothly. The cnemial crest is large, laterally triangular in shape, suggesting strong development of the knee flexor (partly m. femorotibialis), which is confinned by the clearly expressed traces of flexors on the femur. Slightly below the cnemial crest on the external side is developed an elongated right-angled processus fibularis. The trunk of the tibia is absolutely straight, semi-circular in cross-section. The distal end with two almost equal in size condyles, is slightly turned on its long axis inward. There are hardly any signs left of the fusion between the crus and tarsus, although on the external side of the tibia is clearly visible the distal end of the fused fibula. The articular surface of the intertarsal joint is enlarged, allowing the turn of the metatarsus to at least 120°. The distal condyles project forward to almost half the thickeness of the bone, and above them is formed an articular area, which is also reminiscent of that of birds. Their joint width is considerably greater than the width of the trunk, which is reminiscent of the same region in birds. The fibula, as was mentioned, is highly reduced. Approximately along one-third of its length is fused to the tibia. The proximal end is flat, elongated – triangular in shape with a small process for the attachment of the fibula. The metatarsus (fig. 5) is almost analogous to that of birds, for the fusion of distal elements of the tarsus and metatarsus occurs, and as well as the fusion of the elements of the latter with each other, but only proximally. This, it is true, does not affect the rudiment of the 5th tarsal, which fuses fully, and the 3rd which is proximally reduced and ends approximately in the middle of the tarsus. The posteriar proximal surface of the tarsus due to the fusion of the metatarsals II and IV, forms a monolithic area with a rough shagreene from the attached m. gastrocnemius and short toe flexors. Although a process similar to that of ground birds is not formed here, nevertheless the articular surface for the attachment of muscles for the flexing of toes, the crurotarsal and the knee joints are noticeably enlarged in comparison with ornithomimids. The length of the tarsus is 153 mm. The toes (see fig. 5) are shortened at the expense of the length of the distal phalanges (this is especially true of the outside toes). Compared to the ornithomimids the basic phalanges are also shortened, which is most visible in the second toe. The opposite is true of the middle toe, where the basic phalanx is considerably elongated, and the others remain of about the same length as those of the ornithomimids. This is why the third toe is considerably longer than the external ones, especially if one is not to take into account the claw phalanges. In this case, with the normally articulated metapodials, the second toe reaches only the middle of the first phalanx of the third, and the fourth did not reach to the articular fossa on it. The articular surfaces of the phalanges of the external toes are peculiarly arranged. For example, on the fourth toe of the right foot the external part of the articular surface of each phalanx is slightly more developed than the internal, allowing the toe as a whole to have some curvature to the inside. An analogous situation can be seen in the case of the second toe of the left hind limb. It develops that the foot as a whole, due to the curvature of the external toe, narrows toward the middle one. With the exception that in the shape and development of articular surfaces the phalanges of Avimimus are quite similar to those of ornithomimids. But this does not apply to the claw phalanges of the outside toes. The latter are differentiated by a ventral suiface that is pointed laterally and from such a peculiarity are reminiscent of claws. Besides that, they are relatively compressed laterally and above the claw phalanx of the third toe. The latter has a rather common shape – it is smaller than the external, symmetrical, with a flat lower edge, hardly curved at all. Also the claw phalanges of the external toes are only slightly curved. Pelvis (fig. 6). Only the posterior parts of the ilia, starting at the edge of the acetabular cavity, remain. The edge of the acetabular cavity is thick at the expense of the development in this place of a thick antitrochanter which is triangular in shape if viewed laterally, with a slightly convex surface. On deep bending of the femur the antitrochanter articutated with the greater trochanter and the neck, giving support for the femur, for in this case the extensors developed a considerable lateral motion, which aimed to push out the head from the acetabulum, which was resisted by the antitrochanter. The lower posterior edge of the femur is practically straight, only in the middle can be noted a small tubercule, from which starts a large triangular roughness of the flexor of the tibia, m. flexor tibialis. Slightly toward the anterior, on the inner side are preserved two fully fused processes, which connected with the sacral vertebrae. In the most posterior part of the lower edge is visible the roughness of the supinator of the tibia, m. ilio-fibularis. All of the lower part behind the acetabulum is considerably thickened in relation with the majority of theropods, suggesting an intensive development of the extensors of the femur. The ischium is rather common for theropods, with a very well expressed surface of the acetabulum. The posterior process, for the articulation with the ilium, and the anterior are developed practically to the same extent and jointly form slightly less than half of the acetabular cavity. From the lateral side, is proximally devetoped a wide, short and rough crest, which marks the area of attachment of the m. puboischiofemoralis internus. The distal parts of the ischium were not preserved, but if one is to judge from the character of the break, apparently became flat and turned into flat lamellae, which contaced the posterior distal surface of the pubis. The pubis has a short, for theropods, but wide "boot". Distally, the pubic bones diverge very widely, relatively wider than other theropods with the exception of Segnosauridae. For example, wider than in the ornithomimid Gallimimus bullatus, specimen PIN, #551/54, which surpasses Avimimus in size almost double. Approximately from the middle the right and left pubic bones fuse. Directly from this spot they become elongated, triangular in cross-section, whereas the posterior edges are turned outward and the pubic become "open" to the posterior. In this manner is formed a peculiar receptacle for the distal ends of the ischia. The anterior edges, to the contrary, in differentiation from other theropods, are closely approximated and between them remains only a smarl depression. On the break it is quite visible that the proximal pubic bones become much thinner, however their ends were not preserved. In spite of the fact that the pelvic bones were not found in an articulated state, and were incomplete, their position in relation to one another can be properly reestablished. In as much as the ischial process of the ilium and the iliac process of the ischium were preserved, then they sufficiently clearly establish the position of these bones, as well as the maximal posterior position of the ilium. If one is to take into consideration of the posterior part of the ilium approximately parallel to the spine, then the direction of the ischium to it is at almost a 90° angle, and if one is to take into consideration that there is no precise contact between the pelvic bones, then the angle may have been greater. Such a situation is completely uncharacteristic for theropods, where the angle between the spine and the ischium ranges from 30° to 50°. It turns out that the ischia of Avimimus are turned anteriorly by approximately 50°, that is in a positlon that is more characteristic for pubes. The latter were located anteriorly, in their usual place, more or less parallel to the ischia. In as much as in specimen #3907/1 from Udan-Sayr were not preserved proximal parts of of the pubes, then it was only possible to establish aproximately the character of their articulation with the pelvis after examining additional material from the location of Shar-Tsav. Among the bone fragments which belonged to different Avimimus skeletons, was discovered the proximal part of an ischium, which was fully fused with another pelvic bone. Jointly they formed the upper anterior of the acetabulum. After the comparison of this fragment with those of specimen #3907/1, it became clear that the pelvic bone in question was a small piece of the ilium. In this manner the acetabular cavity was formed by the ischium and ilium, while the pubis had no participation in its formation. In the same manner that the anterior position of the ischium, the exclusion of the pubis from the acetabular cavity has no analogues among the dinosaurs in general. Due to this fact, and to the absence of proximal ends of the pubes, the manner of its articulation with the pelvis is not very clear. For the time being, the most likely supposition is the following: approximately at the point of contact of the ischium and the ilium on the external side is located a small protuberance, to which possibly was attached, with the help of ligaments, the pubis. Indirectly this type of attachment is supported by the rather sudden decrease in the thickness of the pubes. At the present time the above described pelvic structure appears likely and doubts can be had only in relation to the articulation of the pubes. Geologic Age. Upper Cretaceous, Nemegetinsk suite, Maastrichtian. Location. Mongolian National Republic, southern and southeastern Gobi. As is apparent from description, the pelvis of Avimimus has a rather unusual construction, which until now has not been observed in theropods, or dinosaurs in general. This first of all is true of the up and down direction of the ischia, which together with the ilia formed the acetabulum. To a lesser degree this also applies to the width of the pelvis. Approximately measured along the distance between the centers of the acetabular cavities, it seemed equal to 10 cm. For an animal of such small proportions as Avimimus, the pelvis is relhtively much wider than in the majority of theropods, with the exception of the representatives of a new family, Segnosauridae (Perle, 1979). Such a shape of the pelvis unquestionably adds stability to the body during bipedal locomotion, and is characteristic, continuing the comparison with birds, of running and climbing animals. In as much as there are no climbing features found in Avimimus, the majority of the other peculiarities of the sketeton indicate it to be a running form. It is known (Gambaryan, 1972) that the greatest development in running is achieved in the extensors of the pelvo-femoral, knee and tibia-foot joints. It is precisely this type of development that we find in Avimimus: an unusually thick, massive wing on the ilium, a large cnemial crest, a large area for the posterior part of the metapodials. The strenghtening of the extensor musculature of the femur occurred at the expense of the increase of the area of its attachment on the posterior branch of the ilium, for which the latter occupied at most a horizontal position. The place for the attachment of the extensors also has great significance for increasing the speed of locomotion. The closer it is to the proximal sector of the linb, and the shorter are these proximal sectors, the greater will be the gain in speed, and the greater the loss in strength. This applies to both the relative shortening of the femur of Avimimus, as well as the proximal displacement on it of the fourth trochanter. However due to the turn of the ischia forward occurred some weakening of the extending capability of m. puboischiofemoralis. With such an arrangement the posterior muscles change the direction of their motion and the extensor-adductor becomes mainly an adductor. But, as was already mentioned, a considerable strengthening in the femur extensors compensated a similar weakening. Otherwise the turn of the ischia forward should not introduce basic changes in the mechanics of the pelvo-femoral muscles. One of the possible explanations of the unusual arrangement of the Avimimus pelvis makes it similar to the open pelvis of birds, which is adapted to the laying of relatively large eggs. Approximately the same types of tendencies for adaptation to running are also observed in the structure of the foot, the metatarsal bones of which have the thinnest, most slender features, which is characteristic for the distal sector of the limbs of running birds. The adaptation to running is also reflected in the relative elongation of the first phalanges of the third toes and the shortening of the toes themselves, which is particularly noticeable on the second. Undoubtedly, this cannot yet be called a reduction, but apparently it is already beginning to take place, in as much as the second metatarsal is smaller and thinner than the fourth. It was already noted that the reduction of the toes went quite far. Thus, if in the forward position of the foot the support fell on all the phalanges, with the exception of the first exterior toes, then in the back position – only an the 2nd-4th-phalanges of the third, while the external, most likely, did not touch the ground at all. If one is to take into consideration some curvature of the external toes toward the middle one, then the support in the forward position of the foot was very narrow. All this to some degree is reminiscent of the foot arrangement of the ostrich. It is interesting to note that in differentiation from running birds the claw phalanges of the hind limbs of Avimimus are practically straight, and the flexor tubercles are not developed on them, but there is no viable explanation for this at the present time. However in spite of the similarities that were stressed between Avimimus and running birds, neither Avimimus, and even less so ornithomimids, are not running animals in the same sense as the contemporary Gradientes. Even in Avimimus, the closest to the latter, the limb proportions are far from being the same, which is also true of the development of the joints and musculature of the knee and crurotarsus, which are not as well developed. For example, the ratio of length of the femur to the length of the tibia and to the metatarsus in Avimimus is 1:1.37:0.81, while in rheas it is 1:1.84:1.70. In ornithomimids, which we are used to considering running animals, as in the case of ostriches, all the features related to this are expressed more weakly than in Avimimus. One one hand, is clear the conditional aspect of the definition of “running” animals, on the other would it not be productive to look for analogies of a different type to compare the way of life of the two groups. Especially since in all the previous views of manner of life of the ornithomimids, only an insignificant role was assigned to the fore limbs, which both in ornithomimids and in Avimimus are well developed and must have served some function. It is possible to say that, up to the present, Avimimus is the most bird-like dinosaur of all the theropods and of all the known dinosaurs as a whole. Such a degree of of development of bird-like characteristics in a theropod allows one to speak not only about the accepted kinship of theropods and birds, but, what is more important, about the possible formation of bird anatomy among the running forms of dinosaurs. Such thoughts were expressed a long time ago (Furbringer, 1888; Broom, 1913) and, it seems, find confirmation in the skeletal structure of Avimimus. However, it is important to note that a considerable similarity is observed between Avimimus and birds in general, and with running birds only in some aspects. This is why it would seem that such similatities in structure would indicate kinship ties, rather than analogues in manner of life. Apparently, supposition concerning the origin of birds from running, bipedal theropods is correctly made. Avimimus is not an ancestor of birds, not even probably the running birds, in as much as in the structure of the pelvis are noted features which are not characteristic for them (the exclusion of pubes from the acetabulum, the turning of the ischia forward). Nevertheless the very fact of the formation of numerous typical bird features among dinosaurs suggests another proof of the possible origin of bitds both from the running bipedal theropods as well as the climbing forms, which in recent years is the dominant point of view. Among these features we should mention the cupolashaped skull, formed by fused frontal and parietal bones, the close positioning of orbits and nasal openings, the presence of the third additional condyle on the femur for articulation with with the fibula, a wide pelvis, fusion of the tarsus and the metatarsus, the presence of the hypapophyses on the last cervical vertebrae and the first dorsal, reduction and fusion of the fibula with the tibia, and also a series of other, less significant features. The formation of such features is hard to explain as parallel development, but rather seems to indicate kinship wfth birds. In addition, the more abundant are these types of features, the closer in the bunch of parallel lines of development will stand the forms being compared. It is possible to find more bird features in Avimimus than in the well-known Archaeopteryx, and if it were not for the time factor, then there would be less basis for including the latter in the ancestral group for birds. At the present time it is still premature to talk of some definite kinship ties of Avimimus with other theropods, since the comparison even with the closest of those ornithomimids is expressed mostly in the structure of hind limbs, and carries, most likely, a clearly converging character. Much light could be thrown on this question by a find of a more complete skull and pelvis. The non-reptilian structure, and possibly manner of life, of Avimimus is suggested by both the high level of exchange of matter, characteristic only for animals with a constant body temperature, although in the present case are absent facts indicating a temperature-conserving cover, which are crucial in the resolurion of this question. Figure Captions Fig. 1. Avimimus portentosus sp. nov.; holotype, PIN, #3907/1. Skull fragment: a – side view, b – top view; Udan-Sayr, Upper Cretaceous, Nemegetinsk suite. fcr – fissura craniofacialis; fj – jugular opening; Fr – fronto-patietal; La – lacrimal; Na – nasal; Osph – orbito-sphenoid; Po – postorbital; Pro – preotic; rin – recessus intracusticus; So – supraoccipital; VII, VIII, IX – openings for cranial nerves. Fig. 2. Avimimus portentosus sp. nov.; holotype, PIN, #3907/1. Last cervical and the first three dorsal vertebrae, lateral view; Udan-Sayr, Upper Cretaceou Nemegetinsk suite. Fig. 3. Avimimus portentosus sp. nov.; holotype, PIN, #3907/1. Front shoulder girdle: a – humerus, front view, b – scapulocoracoid, side view; Udan Sayr, Upper Cretaceous; cr – radial supracondyle; cu – ulnar supracondyle; dp – deltopectoral crest; gl – glenoid cavity; tbb – articular tubercle for m. biceps brachii. Fig. 4. Avimimus portentosus sp. nov.: holotype, PIN, #3907/1. Left hind limb, front view: a – femur, b – tibia, Udan-Sayr, Upper Cretaceous, Nemegetinsk suite. cf – head of femur; cfib – additional condyle for articulation with fibula; f – fused distal end of the fibula; pcn – cnemial crest; pf – processus fibularis; tm – greater trochanter; tmn – lesser trochanter. Fig. 5. Avimimus portentosus sp. nov.; holotype, PIN, #3907/1. Left foot, front view. Udan-Sayr, Upper Cretaceous, Nemegetinsk suite. Fig, 6. Avimimus portentosus sp. nov.; holotype, PIN, #3907/1 (a), paratype PIN, #3906/1 (b). Left half of the pelvis, side view: a – reconstruction of the position of pelvic bones, b – fused parts of ilium and ischium in front of the acetabulum. Udan-Sayr (a), Shara-Tsav (b), Upper Cretaceous, Nemegetinsk suite. ac – acetabulum; ant – antitrochanter; il – ilium; is – fschium; p – pubis; pifi – crest for the attachment of m. puboischiofemoralis internus.