Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
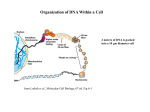
Eukaryotic cells evolved special DNA packaging. Chromosome structure = dynamic 48 million bp in chromosome 22 – laid out long double helix – 1.5cm if stretched. During mitosis = 2 micrometres (compaction ratio = 10,000 fold). Interphase still tightly packed – 500-fold Human cell contains 2m DNA. Tightly packed to form chromatin – hierarchal process – DNA associated with histones – nucleosomes – basic subunit of chromatin. Folded into higher and higher levels of organisation Histone tails extend from these structures – site of extensive post-translational modification – methylation, acetylation, phosphorylation Paragraph 1: Proteins that bind to DNA divided into two classes – histones & non-histone chromosomal proteins. Complex of both with DNA = chromatin. Total mass of histones = same as DNA (huge amount) Histones = most basic level of chromosome packaging Examination with electron microscope of interphase DNA showed chromatin present in form of 30nm diameter fibre. If treated so it unfolds partially it looks like “beads on a sting” – beads = nucleosome core particles (DNA would around octomer core of histone proteins). String = DNA (linking DNA). Nucleosome formation produces a fibre approximately 10nm in diameter and condenses the DNA about six-fold. Biochemical experiments – nuclear core particle separated from isolated chromatin by using nucleases to digest and degrade linker DNA (cannot attack DNA around nucleosome core). Once dissociated from nucleosome into protein core and DNA measure length of DNA. Disc-shaped nucleosome core particle composed of histones: H2A, H2B, H3 AND H4 (all small and share histone fold structural motif). DNA strand 147 nucleotide pairs long (wrapped 1.7 times around histone core), held by 142 H bonds between amino acid histone backbone and phosphodiester backbone of DNA. Hydrophobic interactions and salt linkages also hold them together. Assembly of nucleosome – H3-H4 and H2A-H2B dimer complexes formed by binding. H3-H4 dimers bind to form tetramers. H3-H4 tetramer combines with two H2A-H2B dimers to form compact octomer core. Assembly mediated by specific histone chaperone proteins. Path of DNA around core not smooth – kinks in DNA – this compresses DNA substantially. Some DNA binds core more strongly. Paragraph 2: Histones each have N-terminal amino acid “tail” which extends from core. Chromatin structure and function modified by post-translational modifications – methylation, acetylation, phosphorylation. Histones are highly evolutionarily conserved eukaryotic proteins – reflects how fundamental they are in determining chromatin structure. Suggests that histone functions involve nearly all amino acids Any change in position = deleterious = lethal. Those that aren’t lethal cause abnormalities in normal pattern of gene expression. Tested by mutating histone gene in vitro and introducing into yeast in place of normal copy of gene Eukaryotes produce small amounts of variable histones – vary in amino acid sequence. Variants + covalent modification to tails give rise to large variation in chromatin composition needed for DNA function in higher organisms. Paragraph 3: Access to DNA needed by transcription and replication machinery. Therefore cannot be permanently bound by tight association between histone and DNA In an isolated nucleosome, DNA partially unwraps from each end. Rate = 4 times per second. Exposed for 10-50 milliseconds, then recloses. This allows other proteins to bind. Eukaryotes have variety of chromatin remodelling complexes. These are ATP-dependent. Bind to histone core and to double helix of DNA wound around it. Using energy from hydrolysis of ATP, remodelling complex changes conformation of nucleosome, making association weaker. Cyclical – repeated hydrolysis pulls nucleosome core along the double helix – decreases strength of association – allows other proteins to bind. Remodelling complex can also remove all or part of nucleosome core. Different ATP-dependent chromatin remodelling complexes specialised for different roles. Switching on and off of genes causes specific complexes to bind to specific regions of DNA – determine chromatin structure. Nucleosome position depends mostly on positioning of tightly-bound proteins on the DNA. Some promote adjacent nucleosome formation. Some act as obstacles. Remodelling complex give rise to dynamic arrangement of nucleosomes. Along with histone chaperones repositions cores, arranged with different histones or completely removed. Adaptable to needs of the cell. Paragraph 4: DNA usually undergoes further compression by interactions between histone tails. (increases compaction ~ 40-fold Nucleosomes in ‘beads on a string’ conformation are stacked on top of each other – 30nm diameter fibre – wider. Two methods proposed for how compressed (still debated). X-ray crystallography experiments - high-resolution structural analyses support zig-zag structure. Cryoelectron microscopy supports solenoidal structure (intercalated nucleosomes) How this is achieved = uncertain. May involve long-range interactions between fibres. Or may use scaffold attachment regions (interactions between these regions and loops of AT-rich DNA facilitate condensation). Histone H1 protein (larger than individual core histones) links DNA and nucleosomes and determines eist path of DNA – involved in creating interlocked structure of 30nm fibre. Less conserved. Linker DNA between nucleosomes can vary in length; introducing local perturbations into the structure. DNA-binding and histone-binding proteins also change structural arrangement of nucleosome array. Conclusion: Chromosomes condense lots during mitotic phase of cell cycle. Formation of chromatin is a hierarchal process. Enormous interest is where specific regions of interphase chromosomes de-condense as cells gain access to specific DNA sequences for gene expression, DNA replication and repair. Then re-condense once completed. Packing allows rapid, localised, on-demand access to DNA. Reversible, changes in chromatin structure Repeated arrays of DNA-protein nucleosome paricles. Higher order chromatin structure is still mysterious. Studies in amphibian egg cells have suggested that the overall stability of condensed chromatin and the independent condensation of separate chromosomes require topoisomerases and the proteins condensin I and condensing II.