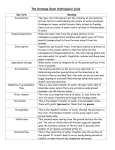
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Coupled Microbial and Transport Processes in Soils M. L. Rockhold,* R. R. Yarwood, and J. S. Selker Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. ABSTRACT 1992b; MacDonald et al., 1999a, 1999b). Almost all previous work on this topic has focused on liquid-saturated porous media systems, with relatively few attempts at addressing more complicated unsaturated or variably saturated systems. This lack of attention to variably saturated systems may be due in part to their added complexity, or may simply be due to the fact that low nutrient availability and competition and predation by other microorganisms limits the growth of many soil microorganisms so that they may only occupy a small or negligible volume of the pore space. Under the high nutrient loading conditions that might occur in applications such as wastewater treatment or bioremediation, however, bacteria and other microorganisms can proliferate, and the consequent changes in soil hydraulic properties may be significant (Rockhold et al., 2002). Many models have been developed for describing water flow, solute transport, and biodegradation processes in porous media. Most of these models were developed for one space dimension and are strictly applicable to saturated porous media representative of aquifer sediments. Examples include the numerical models described by Molz et al. (1986), Widdowson et al. (1988), Celia et al. (1989), Zysset et al. (1994a, 1994b), and Clement et al. (1997). Several multidimensional models have also been developed for simulating transport and biogeochemical reactions in saturated porous media. Noteworthy examples include the RAFT model described by Chilakapati (1995), and the coupling of the HYDROGEOCHEM (Yeh and Tripathi, 1991) and BIOKEMOD models, described by Salvage and Yeh (1998). Very few models account for possible reductions in porosity and permeability that result from biomass accumulation, although it is well known that such effects can sometimes be substantial, especially in the vicinity of the nutrient injection wells used in bioremediation applications (MacDonald et al., 1999a, 1999b). The models that have been developed for simulating water flow and solute transport processes in unsaturated or variably saturated porous media systems such as soils are often less sophisticated in their representations of biodegradation processes than their saturated-zone counterparts. Most of these models account for processes such as biodegradation in terms of simple, first-order solute decay reactions, without actually considering cell growth or substrate (nutrient) limitations on biodegradation rates. Examples include the HYDRUS-2D model (Šimůnek et al., 1999) and the STOMP model (White and Oostrom, 1996). Models that do not account for cell growth obviously cannot be used to access the significance of biomassinduced changes in the hydraulic properties of unsaturated porous media. Very few attempts have been made to directly measure or model biomass-induced changes in the hydraulic properties of unsaturated porous media. However, such changes can have a significant impact on This paper reviews methods for modeling coupled microbial and transport processes in variably saturated porous media. Of special interest in this work are interactions between active microbial growth and other transport processes such as gas diffusion and interphase exchange of O2 and other constituents that partition between the aqueous and gas phases. The role of gas–liquid interfaces on microbial transport is also discussed, and various possible kinetic and equilibrium formulations for bacterial cell attachment and detachment are reviewed. The primary objective of this paper is to highlight areas in which additional research may be needed—both experimental and numerical—to elucidate mechanisms associated with the complex interactions that take place between microbial processes and flow and transport processes in soils. In addition to their general ecological significance, these interactions have global-scale implications for C cycling in the environment and the related issue of climate change. S oils sustain life on Earth. They are important not only from an agronomic standpoint for supporting the growth of plants, but also from an environmental standpoint for mitigating many of the potentially adverse affects of surface-applied contaminants on the quality of groundwater resources. The filtering ability of soils is due to many processes, including physical filtration, sorption of contaminants on the surfaces of mineral grains and soil organic matter, geochemical reactions, and biodegradation by soil microorganisms. Improved understanding of coupled microbial and transport processes in soils is important to a variety of disciplines. In addition to their general ecological significance, these interactions have global-scale implications for C cycling in the environment and the related issue of climate change. As microbial biomass accumulates in porous media, it may change the physical and hydraulic properties of the media. These changes may then alter solute flow paths, gas exchange, microbial growth and redistribution, and other processes. Biomass-induced changes in the hydraulic properties of porous media have been studied for various applications, such as enhanced oil recovery (Jenneman et al., 1984; Raiders et al., 1986), water and wastewater treatment (McCalla, 1946; Nevo and Mitchell, 1967; Loehr, 1977; Overcash and Pal, 1979), and bioremediation of contaminated aquifer sediments (Taylor and Jaffe, 1990a, 1990b, 1990c; Taylor et al., 1990; Cunningham et al., 1991; Vandevivere and Baveye, 1992a, M.L. Rockhold, Pacific Northwest National Lab., P.O. Box 999/MS K9-36, Richland, WA 99352; R.R. Yarwood, Dep. of Crop and Soil Sciences, Oregon State Univ., Corvallis, OR 97331; J.S. Selker, Dep. of Bioengineering, Oregon State Univ., Corvallis, OR 97331. Received 20 June 2003. Special Section: Colloids and Colloid-Facilitated Transport of Contaminants in Soils. *Corresponding author (Mark. [email protected]). Published in Vadose Zone Journal 3:368–383 (2004). Soil Science Society of America 677 S. Segoe Rd., Madison, WI 53711 USA 368 369 Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. www.vadosezonejournal.org water flow and solute transport (Rockhold et al., 2002; Yarwood et al., 2002). The presence of a second fluid phase, air, in an unsaturated porous medium creates an important pathway for gas exchange with the atmosphere. This gas exchange, also known as soil respiration, is crucial for the maintenance of healthy plant roots and microbial activity. Leffelaar (1987) developed a one-dimensional numerical model to study interactions among water flow, solute transport, microbial activity, and respiration in soil aggregates. His emphasis was on multinary gas diffusion and anaerobiosis in soil aggregates. Hence, he did not consider the possibility of changes in the physical and hydraulic properties of the porous media due to biomass accumulation. Concerns over global CO2 emissions have generated a great deal of interest by the scientific community in the topic of C sequestration. These concerns have also motivated the development of a number of computer models to describe CO2 production in soils. For example, Ouyang and Boersma (1992) developed a onedimensional numerical model to simulate the coupled processes of water flow, heat transport, and O2 and CO2 exchange in unsaturated porous media. Their model did not consider the effects of microbial growth or the transport of O2 and CO2 in the aqueous phase. Šimůnek and Suarez (1993) developed a one-dimensional numerical model called SOILCO2 to simulate water flow, heat transport, and CO2 production and transport in soils. Although this model contains source terms to account for CO2 production by plant roots and microbial activity, it neglects microbial growth, as well as the O2 transport and consumption required to support the growth of aerobic microorganisms and plant roots. Several multifluid flow and transport simulators have recently been developed or extended for use in C sequestration studies to account for multicomponent liquid- and gas-phase transport in variably saturated porous media systems using a more rigorous, fully coupled approach (Battistelli et al., 1997; Oldenburg et al., 2001; White and Oostrom, 2003). Most of these simulators were designed principally for high temperature and pressure applications in deep geologic formations. Consequently, they do not consider microbial processes. Travis and Rosenberg (1997) and Battistelli (2003) described two threedimensional numerical simulators that consider coupled, multifluid flow as well as biologically reactive transport processes in variably saturated porous media. While these simulators arguably represent the current state of the art, they assume that microbes are strictly immobile and do not account for pore clogging or other biomass-induced changes in fluid-media properties. It seems that most of the models that have been developed for simulating fluid flow and solute transport in unsaturated or variably saturated porous media either do not account for microbial activity at all, or they do so in an incomplete way, by not actually accounting for microbial growth and transport, and/or by not accounting for the possible effects of microbial growth on the physical and hydraulic properties of the porous media. The primary objectives of this paper, therefore, are to review the status of activity on this research topic and to highlight areas in which both experimental and numerical research may be needed to elucidate mechanisms associated with the complex interactions that take place between microbial processes and transport processes in soils. This paper is organized as follows. Mass balance equations for water flow and solute and microbial transport in variably saturated porous media are first reviewed. Special emphasis is given to issues related to using a single-phase flow approximation (i.e., the Richards equation) in conjunction with advection–dispersion–reaction equations to model water flow and transport of biologically reactive solutes and bacteria in soils. Work related to microbial transport in porous media is then reviewed, and equations representing reaction rate source–sink terms for cell attachment–detachment, growth, accumulation, substrate depletion, and by-product formation are discussed. The role of gas–liquid interfaces is emphasized. The paper concludes with suggestions of topics for future research. FLOW AND TRANSPORT MODELING Soils are three-dimensional, multiphase, multicomponent, nonisothermal, biogeochemical systems that contain a dynamic consortium of microorganisms. The reaction rates and fluid properties in soil systems are well known to be temperature-dependent and hysteretic, with these factors intensively studied and simulated in previous work. For the purposes of this review, we will focus on variably saturated porous media systems, containing a single species of bacterium, under isothermal, nonhysteretic conditions. Our primary interests are the processes of water flow, gas diffusion, solute and microbial transport, and other microbial processes, including substrate consumption, cell growth, and attachment and detachment to and from surfaces. Governing mass balance equations, constitutive relations, and other assumptions necessary for the development of numerical models that describe these processes are reviewed in the following sections. Water Flow The mass conservation of water in porous media can be expressed as t 冤 兺 (n S )冥 ⫽ ⫺ 兺 (䉮F w ␥⫽ᐉ,g ␥ ␥ ␥ ␥⫽ᐉ,g w ␥ ⫹ 䉮J␥w) ⫹ ⍀ w k k F␥w ⫽ ⫺ ␥ ␥ r␥ (䉮P␥ ⫹ ␥gzg) for ␥ ⫽ ᐉ, g ␥ J␥w ⫽ ⫺␥n␥S␥ Mw w w D␥䉮␥ for ␥ ⫽ ᐉ, g M␥ [1] where n is the porosity, w␥ is the mass fraction of water in phase ␥, ␥ is the fluid phase density, S␥ is the phase saturation, Fw␥ is the advective flux, and Jw␥ is the diffusive-dispersive flux, and ⍀ is a source–sink term. The superscript w denotes water, and the subscripts ᐉ and g denote the liquid and gas phases, respectively. The term Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. 370 VADOSE ZONE J., VOL. 3, MAY 2004 kr␥ is the fluid relative permeability, k is the intrinsic permeability tensor, ␥ is the kinematic viscosity, P␥ is the phase pressure, g is the acceleration of gravity, and zg is a unit gravitational direction vector. The term ␥ is the phase tortuosity, M is the molecular weight, Dw␥ is the diffusion coefficient, and w␥ is the mole fraction of water in phase ␥. (For a complete list of symbols see the Appendix.) It is generally assumed that the principal directions of anisotropy are aligned with the coordinate system so that the cross components of the k tensor are zero. If coupled, multifluid flow is considered, equations similar to Eq. [1] can also be written to describe the mass conservation of air and nonaqueous phase liquids (White and Oostrom, 1996). In the soil physics literature, water flow has traditionally been modeled using a simplified, single (aqueous)phase version of Eq. [1], known as the Richards equation (Richards, 1931). Use of the Richards equation implies the assumption of a continuous gas phase at constant, atmospheric pressure. This assumption is typically justified based on the fact that the viscosity of air is about 50 times lower than that of water (Lide, 1996). Therefore, if the gas phase in an unsaturated porous medium is continuous, significant air flow can be caused by very small pressure gradients, and air is assumed to maintain atmospheric pressure, or to quickly reequilibrate to atmospheric pressure under most conditions, due to the relatively small (assumed negligible) resistance to flow. If atmospheric pressure is maintained, then the compressibility of air can be neglected. This assumption is thought to be reasonable most of the time for nearsurface conditions, except when the soil becomes watersaturated, and air contained in the pore space under the saturated region can become compressed under a wetting front. As liquid saturation increases, the airfilled porosity of a porous medium and its relative permeability to air become reduced. Therefore, resistance to air flow may become significant at higher water contents, and in fine-grained porous media. Numerical solutions to Eq. [1] or to the Richards equation require constitutive relations for the volumetric water content, ᐉ ⫽ nSᐉ, as a function of capillary pressure, and for the hydraulic conductivity, K ⫽ kr␥k, as a function of water content or aqueous-phase saturation. These constitutive relations are frequently represented using the well-known models of van Genuchten (1980), Mualem (1976), Brooks and Corey (1964), and Burdine (1953). Accumulation of bacterial cells and associated by-products of cellular metabolism during active growth may alter the hydraulic and transport properties of the porous media, and these changes may need to be considered in numerical modeling (Vandevivere, 1995; De Beer and Schramm, 1999; Rockhold et al., 2002). Solute Transport Transport of dilute concentrations of mobile, linearly partitioning constituents in variably saturated porous media system is generally modeled using equations of the following form (Rf␥Ck,␥) ⫽ 䉮 · (␥D · 䉮Ck,␥) ⫺ (q␥Ck,␥) ⫹ ⌳k,␥ t [2] where Rf is a dimensionless retardation factor, C is the mass of a particular constituent per volume of pore fluid, D is the hydrodynamic dispersion tensor, q␥ is the Darcian flux vector, and ⌳ represents a reaction-rate source– sink term. The subscript k is used here to refer to the constituent (i.e., k ⫽ ed for the electron donor or growth substrate, ea for the terminal electron acceptor, O2 for oxygen under aerobic conditions, CO2 for carbon dioxide, and m for microbes), and the subscript ␥ again refers to the phase. If the Richards equation is used to model water flow, then modeling the transport of constituents that partition between the aqueous and gas phases requires additional considerations, which will be described later. Note that bacterial chemotaxis is not accounted for in Eq. [2] and is not considered here. This important and fascinating topic is discussed in detail by Corapcioglu and Haridas (1984), Ford (1992), Berg (2000), Nelson and Ginn (2001), and others. The hydrodynamic dispersion tensor in Eq. [2] is commonly expressed as (Scheidegger, 1961; Bear, 1972) Dij ⫽ ␦ij␣T|v| ⫹ (␣L ⫺ ␣T) vivj ⫹ ␦ijDeff k,␥ |v| [3] where ␣L and ␣T are the longitudinal and transverse dispersivities, v ⫽ q␥/␥ is the fluid velocity, and Dk,eff␥ is an effective diffusion coefficient for phase ␥. Effective diffusion coefficients have been defined in various ways, such as mol Deff k,␥ ⫽ Dk,␥ 冢 n 冣 a ␥ b [4] where Dk,mol␥ is the coefficient of molecular diffusion, and a and b are empirical parameters. The second term on the right side of Eq. [4] accounts for the tortuosity of the diffusion path through a porous medium. This form of the tortuosity term has been used by numerous researchers for modeling both aqueous- and gas-phase diffusion in porous media, but with different values used for the parameters a and b. For example, Millington and Quirk (1960) used a ⫽ 2 and b ⫽ 2/3, while Millington and Quirk (1961) used a ⫽ 10/3 and b ⫽ 2, and Šimůnek and Suarez (1993) used a ⫽ 7/3 and b ⫽ 2. Moldrup et al. (2000) evaluated several data sets for gas diffusion in repacked soils and determined that overall the data were best represented using a ⫽ 2.5 and b ⫽ 1. Models for effective diffusion coefficients that include residual water content or irreducible saturation terms may be more accurate for modeling aqueous-phase diffusion at low liquid saturations (Olesen et al., 1996). Equations of the form of Eq. [2] through [4] can be used to model the transport of solutes and microbes in the liquid phase, as well as the transport of constituents that partition between the liquid and gas phases, such as O2 and CO2. With the assumption of equilibrium partitioning, the retardation factor, the effective dispersion coefficient, and the Darcian flux used in Eq. [2] can be defined in a manner analogous to that used by Šimůnek and Suarez (1993): 371 www.vadosezonejournal.org 冢KMRT 冣冢 冣 M ⫽D ⫹冢 D K RT 冣 M ⫽q ⫹冢 q K RT 冣 Rf ⫽ 1 ⫹ k g ᐉDE ᐉ [5] ᐉ H k ij g eff k,g [6] H Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. qE k ᐉ g [7] H where Mk is the molecular weight, KH is the Henry’s Law constant, R is the ideal gas constant, and T is absolute temperature. If the single-phase Richards equation is used to model water flow, dispersion in the gas phase is assumed to be negligible, and additional assumptions regarding gas fluxes are required. Following Šimůnek and Suarez (1993), it may be reasonable to assume that the gas flux is zero at the lower soil boundary, L, as well as at the lateral boundaries of a modeled domain. Changes in liquid volume can then be assumed to be immediately matched by corresponding changes in gas volume, such that gas is allowed to enter or exit only through the upper boundary. For one-dimensional, vertically oriented systems, the gas flux into or out of a profile during a time step can be calculated from L V dᐉ qg(0) ⫽ ⫺ 冮 dz [8] A dt z⫽0 where V is the volume of porous media represented by a model grid block, and A is the cross-sectional area of the grid block normal to the direction of flow. The vertical gas flux at any location within a profile can be estimated in a similar fashion. For multidimensional systems, estimation of gas fluxes will generally require the solution of fully coupled sets of equations for the flow of both water and air, rather than the single-phase Richards equation. Diffusion is usually the dominant mechanism for gas-phase transport in unsaturated porous media under most natural conditions, however, so it may not always be necessary to explicitly consider gas advection. Gas advection may be significant during rapid changes in barometric pressure, especially in the vicinity of wells (Massman and Farrier, 1992), and in any type of forced pumping scenario such as air sparging or soil vapor extraction. Furthermore, if gas-phase constituents are significantly denser than air, then a single-phase flow approximation will no longer be adequate, and fully coupled multifluid flow equations will be necessary to accurately model densitydriven gas advection (Lenhard et al., 1995). Even if diffusion is the dominant mechanism for transport in the gas phase, consumption of O2 and concomitant production of CO2 by soil microorganisms may create counter-current movement of these gases that will influence their rates of diffusion. Jaynes and Rogowski (1983) estimated coefficients of molecular diffusion for O2 and CO2 in air from DO2⫺CO2,gDO2⫺N2,g DOmol2,g ⫽ DO2⫺N2,gXCO2,g ⫹ DO2⫺CO2,gXN2,g ⫹ rDO2⫺N2,gXO2,g [9] and Fig. 1. Effective diffusion coefficients for O2 in the gas phase of a sand porous medium as a function of aqueous-phase saturation for different values of the respiration quotient, r. mol DCO ⫽ 2,g DO2⫺CO2,gDCO2⫺N2,g DCO2⫺N2,gXO2,g ⫹ DO2⫺CO2,gXN2,g ⫹ (DCO2⫺N2,gXCO2,g/r) [10] respectively. The terms containing D on the right sides of Eq. [9] and [10] are binary diffusion coefficients for each gas pair, which are assumed to be independent of composition. The terms XO2,g, XCO2,g, and XN2,g are the mole fractions of each constituent in the gas phase. The respiration coefficient, r (moles of CO2 produced/moles of O2 consumed), can be calculated from the stoichiometry of the biologically mediated oxidation–reduction reaction. Equations [9] and [10] were derived from the Stefan–Maxwell equations for steady-state, counter-current diffusion of O2 and CO2 in the ternary system O2– CO2–N2, in which the molar flux of N2 was assumed to be zero (Jaynes and Rogowski, 1983). In the Stefan– Maxwell approximations, it is assumed that the effects of Knudsen diffusion and viscous or pressure-induced flow are negligible (Massman and Farrier, 1992). Additional assumptions associated with the development of the Stefan–Maxwell approximations are discussed by Hirshfelder et al. (1964) and Whitaker (1986). Note that Eq. [9] and [10] are strictly applicable to an unbounded gas phase and should therefore be used in conjunction with Eq. [4] or a similar expression when used to represent gas diffusion in porous media. Figure 1 shows an example of Deff O2,g as a function of aqueous phase saturation calculated using Eq. [4] and [9] for a uniform sand with three different values of the Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. 372 VADOSE ZONE J., VOL. 3, MAY 2004 Fig. 2. Effective diffusion coefficients for O2 in the aqueous and gas phases of a sand porous medium as a function of aqueous-phase saturation. respiration coefficient. Binary diffusion coefficients reported by Jaynes and Rogowski (1983) are used, and tortuosity coefficients of a ⫽ 2.5 and b ⫽ 1 are assumed (Moldrup et al., 2000). In Fig. 1, mole fractions (or partial pressures) of XO2,g ⫽ 0.21, XCO2,g ⫽ 0.000 34, and XN2,g ⫽ 0.789 66 are represented, which are typical atmospheric concentrations of these gases. Figure 1 indicates that the effective diffusion coefficient for O2 in the gas phase decreases rapidly as the aqueous phase saturation increases, due to increased tortuosity. As expected, the effective gas phase diffusion coefficient reaches its maximum value for air-dry conditions, or at residual aqueous-phase saturation. Figure 2 shows effective diffusion coefficients for O2 in the aqueous and gas phases (both with a ⫽ 2.5, b ⫽ 1) for the same porous medium as a function of aqueous phase saturation. Gas-phase diffusion clearly dominates over aqueous-phase diffusion for this porous medium until the aqueous phase saturation reaches approximately 98%. This behavior is expected since coefficients of molecular diffusion in air are typically about four orders of magnitude greater than in water. It should be noted, however, that the exponents a and b in Eq. [4] may not be the same for the aqueous and gas phases, and may also vary with soil type. Equations [9] and [10] were derived based on the assumption of steady-state diffusion with N2 stagnant (XN2,g constant). If diffusion coefficients are calculated using Eq. [9] and [10], and used in advection–dispersion–reaction equations in conjunction with the Richards equation, for consistency with the isobaric assumption (which is implied by the use of the Richards equation), it is necessary to assume that XO2,g ⫹ XCO2,g ⫹ XN2,g ⫽ 1. This assumption neglects any contributions from water vapor and trace gases. If nonequimolar respiration occurs, such that XN2,g ⬆ 1 ⫺ (XO2,g ⫹ XCO2,g), changes in total gas pressure will induce advective gas fluxes in a porous medium so that gas transport is not strictly a diffusion process. Leffelaar (1987) and Freijer and Leffelaar (1996) used the single-phase Richards equation to model water flow, in conjunction with advection–dispersion–reaction equations that contained “pressure adjustment flux” terms to ensure that isobaric equilibrium was maintained and to compensate for possible errors associated with using Fick’s Law to model gas transport. Their calculation of pressure adjustment flux is strictly applicable to onedimensional systems, however, and cannot be generalized to multidimensional problems. As noted previously, accurate estimation of gas fluxes in multidimensional porous media systems requires the solution of fully coupled multifluid flow equations (Pruess and Battistelli, 2002; White and Oostrom, 2003). In some cases, however, such as in relatively coarse porous media at low to moderate liquid saturations, and with respiration coefficients close to 1, Eq. [1] through [10] should provide reasonably good approximations. Thorstenson and Pollock (1989) provided a detailed discussion of multicomponent gas transport in unsaturated porous media and further commentary on the adequacy of Fick’s Law. Bacterial cells are known to secrete various surfaceactive compounds that can accumulate at gas–liquid interfaces. This accumulation can result in the lowering of surface tension and increasing gas–liquid interfacial areas. Accumulation of these compounds at gas–liquid interfaces can also increase the mass transfer resistance across the interfaces. The net effect is usually a reduction of gas–liquid mass transfer rates (Bailey and Ollis, 1986; Schügerl, 1982). Bailey and Ollis (1986) noted that for a variety of sparingly soluble gases, surfactant adsorption at gas–liquid interfaces resulted in an average reduction in the interphase mass transfer coefficient of 60%. Accounting for this type of mass-transfer resistance requires a kinetic modeling approach rather than the equilibrium partitioning described above. Kinetic interphase (gas–liquid) mass transfer can be described using traditional film models (Bailey and Ollis, 1986), or in a similar form 冢 冣 dCk,ᐉ Dkmol ⬀ dt r 1/2 Agᐉ(C*k,ᐉ ⫺ Ck,ᐉ) [11] where Dmol is the harmonic mean of the coefficients of k molecular diffusion in the liquid and gas phases, r is a characteristic time scale for diffusion across the gas– liquid interface, which depends on its mass-transfer resistance, and Agᐉ is the specific gas–liquid interfacial surface area (Eq. [21]). The equilibrium concentration in the liquid phase is C*k,ᐉ ⫽ KHpk, where again KH is the Henry’s law constant, and pk is the partial pressure of the gas. The combination of the first two terms on the right side of Eq. [11] effectively represents a mass transfer coefficient. Eq. [11] is of a form originally proposed www.vadosezonejournal.org by Higbie (1935), and later adapted by Danckwerts (1970), but modified here for interphase mass transfer in variably saturated porous media. Equations such as [11] can be incorporated into reaction rate source–sink terms used in Eq. [2]. Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. Bacterial Cell Attachment and Detachment The processes governing bacterial cell attachment to and detachment from surfaces are complex. Attachment has been attributed to various physicochemical forces including van der Waals’ forces, electrostatic interactions, hydrophobic effects, and specific adhesion (Daeschel and McGuire, 1998). Attachment of bacteria can also be caused by physical filtration or straining (Tien et al., 1979; O’Melia 1985; Logan et al., 1995). Bos et al. (1999) provided an excellent review of various mechanisms for bacterial adhesion. Numerous attempts have been made to relate various properties of bacteria and physicochemical properties of fluid-media systems to bacterial transport. For example, Gannon et al. (1991) characterized the hydrophobicity, net surface electrostatic charge, cell size, and presence of flagella and capsule (extracellular polymeric substances or EPS) for 19 strains of bacteria, and attempted to correlate these properties with transport of the bacteria through a water-saturated loam soil. They noted a positive correlation between the size of bacteria and the percentage of bacteria retained by the soil, but no statistically significant relationships were evident between cell hydrophobicity and retention. All the strains tested had a net negative surface electrostatic charge, but no pattern was evident between surface charge and cell retention by the soil. Mixed results were obtained for relationships between retention of bacterial cells, capsule formation, and presence of flagella. Gannon et al. (1991) concluded that several physiological or morphological properties of bacteria, interacting with properties of the surfaces of soil particles, determine the occurrence and extent of bacterial movement through soil. They suggested that further work would be required to define these properties. Mills et al. (1994) conducted batch experiments to study the sorption of two bacterial strains with different cell surface hydrophobicities to water-saturated, clean quartz sand, to iron oxyhydroxide–coated quartz sand, and to mixtures of the clean and iron oxide–coated sand. Their data for clean quartz sand yielded linear, equilibrium adsorption isotherms whose slopes varied with the bacterial strain used and with the ionic strength of the aqueous solution. The greatest sorption was observed for the highest ionic strength solutions, which is consistent with the interpretation that the electrical double layer is compressed at higher ionic strengths, resulting in stronger adsorption. When the iron oxyhydroxide– coated sand was used, all of the bacteria were adsorbed up to a threshold concentration, above which no more bacteria were adsorbed. This irreversible, threshold adsorption was attributed to strong electrostatic attraction between the Fe coatings and the bacterial cells, and was modeled using a linear adsorption isotherm with a non- 373 zero intercept (Mills et al., 1994). Their results for mixtures of clean and iron oxide–coated sands were described well by a simple additive model for sorption on the two types of surfaces. Bacterial cell attachment to and detachment from porous media surfaces and fluid interfaces have been represented using both equilibrium and kinetic models. The applicability of a local equilibrium assumption depends on environmental conditions, the type and physiological status of the cells, the mineralogical composition of the porous media, the ionic strength of the liquid, and the time-scale of experimental observations. For example, Bengtsson and Lindqvist (1995) conducted stirred flow chamber and column experiments and determined that dispersal of bacteria in their soil was controlled by rate-limited, nonequilibrium sorption rather than instantaneous equilibrium. In contrast, Yee et al. (2000) conducted batch experiments to study the adsorption of a Bacillus subtilus bacterium onto the surfaces of the minerals corundum (␣-Al2O3) and quartz as a function of time, pH, ionic strength, and bacteria/mineral mass ratio. Their experimental data demonstrated that adsorption of B. subtilus onto the mineral corundum is a fully reversible equilibrium process, with equilibrium occurring within 1 h. Adsorption of B. subtilus on quartz was very weak, however, within the two standard deviation error bounds of their control experiments. They assumed that cell attachment and detachment are controlled by the chemical speciation of bacterial and mineral surfaces, and successfully modeled the adsorption of the B. subtilis on corundum using a chemical equilibrium model. In an attempt to provide a more rigorous theoretical basis for explaining bacterial cell attachment and transport behavior, Chen and Strevett (2001) characterized the surface thermodynamic properties of two types of porous media (silica gel and sand) and three types of bacteria (Escherichia coli, Pseudomonas fluorescens, and B. subtilis) in different physiological states using contact angle measurements with liquids of different surface tensions. They found a strong correlation between the total Gibbs free energies of surface interaction, calculated from contact angle measurements, and deposition (or attachment) coefficients (Bolster et al., 1998) that were calculated from the fraction of bacteria recovered during column experiments in the water-saturated porous media. They also showed that deposition is correlated with physiological growth state, with stationary-phase cells being the most strongly adsorbed. In addition, Chen and Strevett (2001) used infrared spectroscopy to show that increased deposition is correlated with an increase in the H-bonding functional groups on bacterial cell surfaces. Although their calculated Gibbs free energies of surface interaction were correlated with first-order attachment coefficients, Chen and Strevett (2002) showed that an advection–dispersion equation with simple first-order attachment did not provide adequate descriptions of their bacterial breakthrough curve data. Chen et al. (2003) simulated the experiments using a model based on a two-site (equilibrium and kinetically controlled) advec- Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. 374 VADOSE ZONE J., VOL. 3, MAY 2004 tion–dispersion equation (Šimůnek et al., 1999; van Genuchten and Wagenet, 1989). Significantly improved results were obtained relative to using an advection– dispersion equation with simple first-order attachment. Chen et al. (2003) showed that the parameters in the two-site model were also strongly correlated with independently measured thermodynamic properties of the bacterial cell and porous media surfaces. The thermodynamic approach used by Chen and Strevett (2001, 2002) was based on the extended DLVO theory of van Oss (1994) that considers Lewis acid–base interactions. One of the criticisms of applying surface thermodynamic approaches and the DLVO theory in general to describe bacterial adhesion in porous media is that these approaches are typically based on the assumption of perfectly smooth, uniform, spherical particles attaching to smooth flat surfaces (Hermansson, 1999). Many bacteria and other soil microorganisms have various heterogeneous surface structures (e.g., fimbriae, flagella) that can facilitate attachment (Atlas and Bartha, 1993; Madigan et al., 1997; Hermansson, 1999). Hence it could be argued that DLVO theory and its modifications are not strictly applicable to modeling attachment during microbial transport in porous media. Nevertheless, as shown by Chen and Strevett (2001, 2002), Chen et al. (2003), and others, surface thermodynamics have provided a useful framework for evaluating microbial attachment in porous media. The dynamic and adaptive nature of soil microorganisms may confound the quantification of specific mechanisms of attachment. For example, bacteria are known to produce extracellular polymeric substances, including polysaccharides, and other types of conditioning films that can buffer cells from desiccation in unsaturated porous media and also promote adhesion (Williams and Fletcher, 1996; Vandevivere and Kirchman, 1993; Chenu, 1993; Roberson and Firestone, 1992). Changes in soil water potential are known to trigger physiological changes in bacterial cells and to alter their metabolic processes (Kral, 1981). Some bacteria are also known to produce surface-active agents, or biosurfactants, during active metabolism that can adsorb to gas–liquid interfaces, resulting in lower surface tensions (Kosaric, 1993; Déziel et al., 1996). Lawrence et al. (1987) studied the surface colonization behavior of a P. fluorescens bacterium and determined that it could be subdivided into the following sequential phases: motile attachment phase, reversible attachment phase, irreversible attachment phase, growth phase, and recolonization phase. Harvey and Garabedian (1991) and Hendry et al. (1997, 1999) found that it was necessary to account for bacteria that were both reversibly and irreversibly attached at solid–liquid interfaces to match observed and simulated breakthrough curves for bacteria in saturated porous media systems. Ginn et al. (2002) suggested that in saturated porous media systems, bacterial cell attachment and detachment are related to both residence time and growth processes. Most of the work on bacterial transport in porous media has been done under no-growth and liquid-saturated conditions, in which bacteria adhere strictly to solid– Fig. 3. Conceptual model of possible sorption sites for bacterial cells in variably saturated porous media and corresponding rate coefficients. liquid interfaces. A number of studies have also been conducted in unsaturated porous media, however, which have shown that some bacteria may preferentially adsorb to gas–liquid interfaces (Wan et al., 1994; Powelson and Mills, 1998; Schäfer et al., 1998; Jewett et al., 1999). Wan et al. (1994) suggested that sorption of bacterial cells to gas–liquid interfaces is essentially irreversible due to cell surface hydrophobicity. This conclusion should be dependent, however, on the surface properties of the bacteria and fluid-media system (Docoslis et al., 2000). Given the adaptive nature of bacteria, and the difficulty in isolating and quantifying specific mechanisms, unstructured models and phenomenological approaches are generally used to represent bacterial cell attachment and detachment processes in porous media. Given the literature cited above, in kinetic modeling approaches the possibility of sorption at three different types of sites may need to be considered. These are (i) reversible sorption at solid–liquid interfaces (and/or on other attached cells), referred to here as s1 sites; (ii) irreversible sorption at solid–liquid interfaces (e.g., on ironoxide coatings), referred to as s2 sites; and (iii) irreversible sorption at gas–liquid interfaces. These possible sorption sites and corresponding rate coefficients are depicted in Fig. 3. An alternative, equilibrium approach for modeling cell sorption at gas–liquid interfaces is described later. Reaction Rate Source–Sink Terms General (kinetic) forms of the reaction rate source– sink terms that could be used in Eq. [2] are given by ⌳ed,ᐉ ⫽ ⫺Yed/m(ᐉCm,ᐉ ⫹ bCm,s1 ⫹ bCm,s2 ⫹ gCm,g) [12] ⌳ea,ᐉ ⫽ ⫺Yea/m(ᐉCm,ᐉ ⫹ bCm,s1 ⫹ bCm,s2 ⫹ gCm,g) [13] ⌳m,ᐉ ⫽ ᐉCm,ᐉ ⫺ k1ᐉCm,ᐉ ⫹ k2bCm,s1 ⫺ k3FᐉCm,ᐉ ⫹ k4gCm,g ⫺ k5GᐉCm,ᐉ ⫹ k6bCm,s2 [14] where is a metabolic lag function; Yed/m and Yea/m are yield coefficients representing the mass of electron donor or substrate (e.g., glucose) consumed and the mass of terminal electron acceptor (e.g., oxygen) consumed, respectively, per mass of cells generated; is the specific growth rate of the bacterium; k1 through k6 are attachment–detachment rate coefficients; F and G are blocking 375 Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. www.vadosezonejournal.org functions; and b is the bulk density. The terms Cm,ᐉ, Cm,s1, Cm,s2, and Cm,g represent the mass of cells in the aqueous phase per volume of pore liquid, the mass of cells reversibly attached to solids per mass of porous media, the mass of cells irreversibly attached to solids per mass of porous media, and the mass of cells attached to gas– liquid interfaces per volume of gas phase, respectively. Note that in Eq. [12] through [14] the same growth rate is applied to bacteria everywhere. This does not necessarily have to be the case. Different growth rates could be applied to bacteria in different regions if, for example, bacteria at air–water interfaces are thought to have greater access to O2, and hence higher growth rates, than those associated with solid–water interfaces. The metabolic lag function, , is an empirical means of incorporating the time lag that is sometimes observed before cells start to metabolize a substrate after their initial exposure to it. This function may take a very simple form, such as ⫽0 ⫽ t ⱕ L ⫺ L E ⫺ L L ⬍ ⱕ E ⫽1 [15] t ⬎ E where is the length of time that the cells at any given location have been exposed to some minimum threshold concentration of a substrate, Cmin k,ᐉ , L is the observed time lag before the cells start to metabolize the substrate, and E is the time required for the cells to reach their exponential growth phase (Wood et al., 1995). Strictly speaking, Eq. [15] should only be applied to bacteria that are irreversibly attached. The specific growth rate is usually represented by a multiplicative Monod-type kinetics model (Megee et al., 1970): 冢K ⫽ max ea 冣冢 Cea,ᐉ Ced,ᐉ ⫹ Cea,ᐉ Ked ⫹ Ced,ᐉ 冣 [16] where max is the maximum specific growth rate (h⫺1), and Kea and Ked are half-velocity (or half-saturation) coefficients (mg L⫺1) for the terminal electron acceptor (e.g., oxygen) and the electron donor or C source (e.g., glucose), respectively. Note that Eq. [16] can be easily modified to include additional constituents that might limit growth, such as N or P, and to account for various types of substrate inhibition effects (Bailey and Ollis, 1986; Barry et al., 2002). Mass balance equations for attached biomass can be written as (bCm,s1) ⫽ bCm,s1 ⫹ k1ᐉCm,ᐉ ⫺ k2bCm,s1 t [17] (gCm,g) ⫽ gCm,g ⫹ k3FᐉCm,ᐉ ⫺ k4gCm,g t [18] (bCm,s2) ⫽ bCm,s2 ⫹ k5GᐉCm,ᐉ ⫺ k6bCm,s2 t [19] The maximum extent of cell sorption at gas–liquid interfaces is assumed to correspond to monolayer cover- Fig. 4. Gas–liquid interfacial surface area per bulk volume of a 40/50 grade of quartz sand as a function of effective aqueous-phase saturation, calculated using the Brooks–Corey water retention model with n ⫽ 0.332, bc ⫽ 8.85, hb ⫽ 26.5 cm, ⫽ 0.993 g cm⫺3, g ⫽ 981 cm s⫺2, and ⫽ 72.8 mN m⫺1. age. In a two-phase (air–water) system, the fraction of the gas–liquid interfacial surface area available for sorption, F, can be represented using a Langmuir-type blocking function: C A F ⫽ 1 ⫺ g m,g cp Agᐉ ⬎ 0 mcAgᐉ F⫽0 Agᐉ ⫽ 0 [20] where Acp is the maximum projection area of a cell, mc is the average mass of a fully hydrated cell, and Agᐉ is the gas–liquid interfacial surface area per volume of porous media, which can be estimated from (Niemet et al., 2002) Agᐉ(Se) ⫽ 冢 3gnhb 1 1⫺ 2 bc ⫺1 冣冢 1⫺ 1 ⫺ Se 1 bc 冣 h ⱖ hb h ⬍ hb [21] Agᐉ(Se) ⫽ 0 where Se ⫽ (ᐉ ⫺ r)/(n ⫺ r), and where r is the residual or irreducible water content, is the mass density of the water, g is the gravitational constant, is the interfacial tension at the gas–liquid interface, and hb and bc are parameters in the Brooks and Corey (1964) water retention function. Note that a number of other similar expressions have also been developed for describing interfacial surface areas in variably saturated porous media (Leverett, 1941; Skopp, 1985; Cary, 1994; Oostrom et al., 2001; and others). Figure 4 shows the gas–liquid interfacial surface area Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. 376 VADOSE ZONE J., VOL. 3, MAY 2004 as a function of effective liquid saturation, given by Eq. [21], for a uniform, 40/50 grade of quartz sand. Although Fig. 4 indicates a nearly linear relationship, it should be noted that Eq. [21] applies strictly to conditions following drainage from a fully water-saturated state, and does not account for the effects of hysteresis and air entrapment, which may create much more complicated relationships between gas–liquid interfacial surface area and liquid saturation. The maximum extent of cell sorption at the s2 sites is assumed here to correspond to monolayer coverage of the fractional area of solid–water interfaces consisting of s2 sites. This can be represented by the function C A G ⫽ 1 ⫺ b m,s2 cp mcAsᐉ [22] where 0 ⬍ ⱕ 1 is the fractional area of solid–liquid interfaces consisting of s2 sites, and Asᐉ is the solid–liquid interfacial surface area per volume of porous media. The parameter Asᐉ can be estimated from the product of the first two terms on the right side of Eq. [21]. The fractional area consisting of s1 sites does not necessarily have to be explicitly accounted for in Eq. [22] if it is assumed that bacterial cells can continue to accumulate on top of one another, even after monolayer coverage of all the s1 and s2 sites has been reached. Transport equations with kinetic rate coefficients similar to the ones described here have been proposed for modeling the effects of sorption at gas–liquid (air– water) interfaces on the transport of viruses (Chu et al., 2001), bacteria (under nongrowth conditions) (Schäfer et al., 1998; Jin et al., 2002), and colloids (Corapcioglu and Choi, 1996). The experimental results of Wan and Wilson (1994) and Wan et al. (1994) have been cited in all of these studies as justification for explicitly considering sorption at gas–liquid interfaces. Most of the previously reported bacterial transport studies only considered steady flow and nongrowth conditions, however, so complications associated with modeling attachment– detachment and simultaneous growth at dynamic gas– liquid interfaces were avoided. Furthermore, as noted previously, the relative propensity for cells to attach to gas–liquid vs. solid–liquid interfaces will depend on the surface properties of the bacteria and fluid-media system. The nature of gas–liquid interfaces in unsaturated porous media is also subject to some debate, as to whether they are more representative of no-slip boundaries (e.g., the static wall of a capillary tube), or slip boundaries (e.g., the surface of a flowing river), or some combination of these extremes (e.g., trapped gas bubbles as well as a continuous, mobile gas phase). The nature of gas–liquid interfaces in unsaturated porous media obviously has some bearing on how sorption at these interfaces should be modeled. An alternative to the kinetic approach described above for modeling cell sorption at gas–liquid interfaces is to assume equilibrium adsorption. Several studies have measured bacterial cell surface energies against air and have found them to be less than the surface energies (or surface tensions) at air–water interfaces (Absolom et al., 1983; Gerson, 1993). Sorption of hydrophobic cells and associated by-products of cell metabolism at gas– liquid interfaces could therefore be expected to result in some degree of surface tension lowering (Rockhold et al., 2002). One way of accounting for sorption of surfactants and/or cells at gas–liquid interfaces and concomitant changes in surface tension is to the use the Gibbs adsorption equation (Adamson and Gast, 1997): M dlg ⌫⫽⫺ k RT d ln C [23] where ⌫ is the surface “excess” (mg cm⫺2) adsorbed to gas–liquid interfaces, Mk is the molecular weight of the adsorbing molecule (or cell), R is the gas constant, T is the absolute temperature, lg is the surface tension at the liquid–gas interface, and C is the aqueous-phase concentration. Eq. [23] can also be expressed as ⌫⫽ Mk d␥sp C RT dC [24] where the equilibrium spreading pressure is ␥sp ⫽ o ⫺ lg, and where o is the surface tension at the liquid–gas interface for the aqueous solution in the absence of bacteria or other surface-active compounds. The equilibrium spreading pressure can be expressed in terms of the familiar Langmuir- or Freundlich-type isotherm models as (Adamson and Gast, 1997) ␥sp ⫽ ABC 1 ⫹ AC [25] and ␥sp ⫽ AC B [26] respectively, where A and B are empirical parameters. For the Langmuir-type model, B also represents a maximum spreading pressure, B ⫽ o ⫺ min, where min is the minimum liquid–gas interfacial tension that would presumably be obtained at monolayer coverage of the interfaces by bacterial cells (or near the critical micelle concentration for surfactants). Use of the Langmuirtype model for sorption at gas–liquid interfaces implies a finite sorption capacity that can be estimated using Eq. [20] and [21]. If the Langmuir-type model is used to represent sorption of cells (or surfactants) at gas–liquid interfaces, Eq. [24] and [25] can be combined to determine a retardation factor, Rf, used in Eq. [2] as Rf ⫽ 1 ⫹ M 冢 A 冣冢RT 冣冤(1 ⫹ABAC) 冥冤1 ⫺ 冢1 2AC ⫹ AC 冣冥 gᐉ k 2 ᐉ [27] Similarly, if the Freundlich-type model is used, Eq. [24] and [26] can be combined to determine a retardation factor as Rf ⫽ 1 ⫹ M 冢 A 冣冢RT 冣AB C gᐉ k 2 B⫺1 [28] ᐉ Equations [25] through [28] are nonlinear, unless B in Eq. [26] and [28] equals one, which corresponds to the classical linear Freundlich isotherm model. Equations [27] and [28] are strictly applicable to equi- Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. www.vadosezonejournal.org librium adsorption at gas–liquid interfaces. If equilibrium sorption at solid–liquid interfaces also occurs, retardation effects due to sorption at these interfaces can be accounted for in the usual way, and used in conjunction with Eq. [27] and [28]. However, it is difficult to account for the simultaneous growth of bacteria at interfaces where equilibrium sorption has been assumed due to the mixing of equilibrium and coupled kinetic reactions. Nevertheless, Eq. [25] through [28] can be readily used with Eq. [2] to model microbial transport and adsorption at gas–liquid interfaces under nongrowth conditions, and/or surfactant adsorption at gas–liquid interfaces with concomitant surface-tension lowering effects. It should be noted that both the kinetic and equilibrium descriptions of sorption at gas–liquid interfaces given above implicitly assume that these interfaces are effectively static, no-slip boundaries. However, gas–liquid interfacial areas may change as a function of liquid saturation, following Eq. [21]. Surface tension lowering in porous media can result in decreases in liquid saturation and changes in the gradients of capillary pressure. Both of these factors affect water flow and hence solute and microbial transport. These effects can be accounted for by scaling of capillary pressure–saturation relations (Rockhold et al., 2002), followed by simple updating of capillary pressures at the end of each transport step. Changes in capillary pressure will then create changes in liquid saturation, permeabilities, and pressure gradients that will control the magnitude and direction of flow velocities at the next time step. In this loosely coupled approach, concentration-dependent changes in capillary pressure due to surface active agents, and concomitant changes in liquid saturation and permeability, are lagged by one time step. This approach requires minimal additional computational effort, but may not be as accurate as some other methods. Smith and Gillham (1994) described an alternative, iterative method that was used to simulate the effects of surfactants on water flow and solute transport in laboratory column experiments. Their method required time step sizes on the order of milliseconds. Hence, albeit more accurate, this approach may be impractical for simulation of field-scale problems. An alternative method, involving the solution of fully coupled multifluid flow equations, with surfactant as one of the fluids, is described by White and Oostrom (1998). Rate Coefficients As noted above, most of the models that have been developed for simulating microbial transport in porous media have used kinetic modeling approaches. Harvey and Garabedian (1991), Hornberger et al. (1992), Deshpande and Shonnard (1999), and others assumed that bacterial cell attachment in saturated porous media could be represented by a first-order kinetic model and used the model of Tien et al. (1979) to estimate attachment coefficients. In the model of Tien et al. (1979) the attachment coefficient, k1, is estimated from k1 ⫽ 冢 冣 3 qᐉ (1 ⫺ ᐉ) ␣c 2 ᐉ dg [29] 377 where dg is the median grain diameter of the porous medium (or collector), and and ␣c are the so-called collector and collision (or sticking) efficiencies (Logan et al., 1995; Deshpande and Shonnard, 1999). Equation [29] is based on particle filtration theory and the earlier work of Rajagopalan and Tien (1976). The expression for the collector efficiency is essentially an empirical correlation function that contains a number of dimensionless terms to describe London–van der Waals interactions, interception, sedimentation, and diffusion. The collector efficiency is calculated from the physical properties of the porous media (e.g., porosity and median grain diameter), fluid properties (e.g., density, viscosity, velocity, and temperature), and the size and density of the suspended particles (Tien et al., 1979; Logan et al., 1995). The sticking efficiency, ␣c, is an empirical parameter. Chiang and Tien (1985a, 1985b) developed another empirical correlation function that modifies the model of Tien et al. (1979) to account for a so-called “filter ripening” effect that is frequently observed in particle filtration systems, where the collector efficiency increases with time as more suspended particles become attached to the filter media. The model of Tien et al. (1979) and the empirical correlation function by Chiang and Tien (1985a, 1985b) do not account for detachment (reentrainment) of particles, or surface interactions resulting from differences in charge, the presence of electrolytes, or changes in the ionic strength of the suspending liquid. Such surface interactions might include both particle– particle interactions, such as flocculation or electrostatic repulsion, and particle–collector interactions that might occur, for example, if the electrostatic double-layer thickness is reduced by an increase in the ionic strength of the pore water. In spite of the fact that Eq. [29] does not explicitly consider the surface interactions noted above, Deshpande and Shonnard (1999) used it to study the effects of systematic increases in ionic strength on the attachment kinetics of a P. fluorescens bacterium in saturated sand columns. They varied the and ␣c parameters, as well as a third parameter, c, in the filter ripening model of Chiang and Tien (1985a, 1985b), to optimize the fit between simulated and observed bacteria breakthrough curves for different ionic strengths of the suspending fluid. Reasonably good results were obtained for all cases, but somewhat better results were obtained when the additional fitting parameter, c, was used. McDowell-Boyer et al. (1986) provide a review of various factors that may affect particle transport through porous media and different modeling approaches. Note that the term particle has been used rather loosely to refer to objects of many different sizes, shapes, and compositions, including latex colloids, bacteria, and viruses. However, particle filtration models are usually based on the assumption of smooth, inert, and perfectly spherical particles. Although these models account for a number of physical factors, they generally neglect electrostatic interactions and hydrophobic effects. Given the heterogeneous surface characteristics and nonspherical nature of most soil microorganisms, one could question the validity of using particle filtration models (Eq. [29]) in Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. 378 VADOSE ZONE J., VOL. 3, MAY 2004 conjunction with advection–dispersion equations to model microbial transport in porous media. Nevertheless, these equations have been applied successfully in numerous cases to describe microbial transport in porous media. Murphy and Ginn (2000) and Ginn et al. (2002) provided further discussion on this topic and suggested alternative modeling approaches. Rittmann (1982) developed an equation to describe detachment as a function of fluid shear stress, using data for a P. aeruginosa bacterium that was obtained using a centrifuge (Trulear and Characklis, 1982). Speitel and DiGiano (1987) suggested, however, that detachment is much more closely related to biomass growth rate than to the amount of biomass present, and modified Rittmann’s model with an additional term to account for the specific growth rate. MacDonald et al. (1999a) used Rittmann’s model to account for detachment as a function of fluid velocity and shear in the vicinity of well screens for a hypothetical bioremediation scenario involving the injection of nutrients into an aquifer. Alternative models for bacterial cell detachment have been proposed by Johnson et al. (1995), Ginn (1999), and Ginn et al. (2002), who considered detachment to be a residence time–dependent process. They did not ascribe this time dependence to any particular mechanism. Hence these models are essentially empirical, but provide more flexibility in fitting breakthrough curve data. The models proposed by Johnson et al. (1995) and Ginn (1999) require the use of particle-tracking algorithms to track the time history of individual particles (or cells). Scheibe and Wood (2003) described a novel approach, also based on particle tracking, to describe pore-scale exclusion processes and their effects on bacterial transport. Using particle tracking methods to model bacterial transport is made somewhat more complicated if cell growth is considered, but growth was not considered in any of the aforementioned papers. A simplified approach for representing time-dependent detachment rates would be to use k2 ⫽ f exp(⫺tgc) [30] where f and gc are empirical parameters, and t is equal to the value of in Eq. [15], or to the time since any location in the modeled domain is first exposed to some minimum threshold concentration of growth substrate. In essence, this amounts to keeping track of the time in which local environmental conditions have been conducive to optimal growth, rather than keeping track of the time history of individual particles (or bacterial cells). Note that Eq. [30] has the same affect as the filter ripening model of Chiang and Tien (1985a, 1985b). Time-dependent rate coefficients have also been used for modeling irreversible sorption of proteins at solid– water interfaces. For example, Lee et al. (1999) modeled observed decreases in protein adsorption rate using an adsorption (or attachment) coefficient that was a power function of time, rather than a blocking function, as used in Eq. [18] and [19], or an exponential function of time, of the type given by Eq. [30]. Decreases in protein adsorption rate with time were attributed to an increasing energy barrier associated with the increase in ad- sorbed mass as irreversible sorption sites approached monolayer coverage (Lee et al., 1999). Changes in protein adsorption rates with time have also been attributed to conformational or structural changes that occur in protein molecules as they “unfold” on the surfaces to which they attach (Daeschel and McGuire, 1998). Further discussion on protein adsorption as it relates to virus transport in porous media is given by Thompson et al. (1998) and Jin and Flury (2002). In kinetic modeling approaches, adsorption of bacteria at gas–liquid interfaces has typically been assumed, based on the results of Wan et al. (1994), to be an essentially irreversible process. It may be reasonable to assume that sorption at gas–liquid interfaces is also a function of both the aqueous phase concentrations of bacteria, as well as the gas–liquid interfacial area that is available for sorption, as suggested by Eq. [20] and [21]. The attachment coefficient for bacteria at gas– liquid interfaces can be represented by the parameter, k3, which is assumed to be a constant. Detachment of bacteria from gas–liquid interfaces can then be assumed to be negligible until these interfaces reach monolayer coverage, or until increases in aqueous phase saturation reduce the gas–liquid interfacial area, causing the mobilization of cells that were previously attached. After monolayer coverage is reached, additional growth of cells at gas–liquid interfaces will presumably result in instantaneous detachment and resuspension in the aqueous phase. This condition can be represented by k4 ⫽ 0 F⬎0 k4 ⫽ F⫽0 [31] The possibility for irreversible attachment of bacterial cells at solid–liquid interfaces was also considered by Harvey and Garabedian (1991), Hendry et al. (1997, 1999), and others. The rate of irreversible attachment may be assumed to be proportional to the concentration of bacteria in the aqueous phase and to the fraction of surface area available for sorption at s2 sites, as given by Eq. [19] and [22], where the k5 parameter is assumed to be a constant, and the k6 parameter is represented the same way as k4 in Eq. [31], but based on the value of G rather than F. This treatment is consistent with the observations of Mills et al. (1994), cited previously, that suggest that some bacteria may become instantaneously and irreversibly adsorbed on iron-oxide coatings. Lawrence et al. (1987) suggested, however, that cells first become reversibly attached before they become irreversibly attached, which could possibly represent a different mechanism, such as the production of conditioning films by the bacteria. By inspection of Eq. [18] and [19], it can be seen that the rate parameters k3 and k4 are somewhat redundant with the rate parameters k5 and k6, and may therefore be unnecessary. Although they can potentially be used to represent different mechanisms, their net effect is indistinguishable under most conditions, unless sorption of cells at gas–liquid interfaces also causes other effects, such as surface tension lowering. If this is the case, then sorption at gas–liquid interfaces might be better 379 Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. www.vadosezonejournal.org represented using the Gibbs adsorption equation, as described above. It may be of interest to note that some earlier solute transport studies also observed more apparent sorption or incomplete solute displacement at lower liquid saturations (Nielsen and Biggar, 1961). However, this behavior was attributed to a larger fraction of water in the porous media being contained in relatively slow moving or stagnant regions that acted as sinks to ionic diffusion. These observations led to the development of the mobile–immobile water concept that is frequently used in solute transport modeling (Coats and Smith, 1964; van Genuchten and Cleary, 1979; van Genuchten and Wagenet, 1989). Use of the mobile–immobile water concept may lead to reductions in simulated peak concentration values that may be similar to the results obtained by Wan et al. (1994) for bacteria when liquid saturation was reduced in their glass micromodel and column experiments. However, the mechanisms to which this phenomenon is attributed are different. The mobile– immobile water model generally leads to extended tailing of breakthrough curves, due to slow mass transfer of solutes out of immobile water regions after the main solute pulse has passed, while irreversible sorption tends to simply reduce peak concentrations without significantly affecting the higher moments of a breakthrough curve. SUMMARY AND CONCLUSIONS We have discussed coupled microbial and transport processes in soils. These coupled processes and interactions include (i) the effects of substrate limitations, biomass growth and accumulation, and by-product formation on the physical and hydraulic properties of variably saturated porous media, and the associated effects on water flow and solute and microbial transport and (ii) the ability of gas-phase diffusion, which is strongly dependent on aqueous-phase saturation as well as biomass concentration distributions, to provide O2 as a terminal electron acceptor in aerobic, biologically mediated oxidation–reduction reactions in soils. For one reason or another, such interactions have been largely neglected in both experimental work and numerical modeling of transport processes in soils. The lack of consideration of these types of interactions in many experimental and modeling studies may be partly responsible for the difficulty in reconciling some lab and field data, and in reproducing field data for reactive transport problems using model parameters obtained from lab studies. Two main topics emerge as warranting further research: (i) the development of novel experimental methods for observing and measuring coupled biogeochemical processes in variably saturated porous media systems, to elucidate underlying mechanisms and emergent phenomena that may be associated with these process interactions, particularly in heterogeneous systems, and (ii) the development of more computationally efficient and accurate numerical methods for simulating these coupled processes, their interactions, and feedback mechanisms. Novel experimental methods involving biolumi- nescent bacteria, translucent porous media, and lighttransmission chambers, such as those described by Yarwood et al. (2002) and Weisbrod et al. (2003), may provide new insights into the interactions between microbial processes and transport processes in unsaturated porous media. Although the Richards and advection– dispersion–reaction equations may be applicable under some conditions, the strongly coupled nature of the processes, and various feedback mechanisms, will ultimately require the use of fully coupled, multifluid flow and multicomponent reactive transport equations, such as those described by Pruess and Battistelli (2002) and Battistelli (2003), that allow for more accurate representations of the coupled processes for a wider range of conditions. Ginn et al. (2002) provided additional suggestions for future research related to microbial transport in the subsurface. This work has focused primarily on processes associated with the growth and transport of a single bacterial species in variably saturated porous media, and methods for modeling these processes. It should be recognized, however, that all naturally occurring biogeochemical processes in the subsurface are influenced by interacting microbial consortiums, rather than single species (Atlas and Bartha, 1993; Barry et al., 2002; Battistelli, 2003). Further research is warranted toward developing a better understanding of the complex processes and interactions associated with the spatial and temporal dynamics of microbial communities under variable fluxes of water, energy, and nutrients, especially in heterogeneous, unsaturated porous media. Topics of special interest include the dynamics of mixed microbial populations growing on multiple substrates (e.g., diauxic growth), and chemotaxis in response to substrate limitations. Interdisciplinary research that combines the traditional fields of study of soil physics, chemistry, and microbiology (or microbial ecology) should prove to be especially fascinating. APPENDIX a A Acp Agᐉ b B C dg D Dk,mol␥ Dk,eff␥ f F Fw␥ g gc G exponent in numerator of tortuosity function empirical parameter in isotherm models average cell projection area (m2) gas–liquid interfacial surface area/bulk volume (m2 m⫺3) exponent in denominator of tortuosity function empirical parameter is isotherm model solute or bacterial concentration (kg m⫺3) median grain diameter (m) diffusion–dispersion tensor (m2 s⫺1) coefficient of molecular diffusion, (m2 s⫺1) effective diffusion coefficient, (m2 s⫺1) empirical parameter in model for detachment coefficient Langmuir-type blocking function for air-water interfaces advective flux of water in phase ␥ [kg/(m2 s)] gravitational constant (m s⫺2) empirical parameter in model for detachment coefficient blocking function for solid–liquid interfaces 380 h hb Jw␥ Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. K k1–k6 kr KH Kea Ked mc Mk N pk qᐉ, qg r R Rf S␥, Se t T v Xk Y ␣c ␣L, ␣T ␦ ␥sp ⌫ ᐉ, g bc ⌳ , max f , 0 L E r ⍀ VADOSE ZONE J., VOL. 3, MAY 2004 soil water pressure head (m) air-entry pressure (m) diffusive–dispersive flux of water in phase ␥ [kg/ (m2 s)] intrinsic permeability (m2) attachment–detachment rate coefficients (s⫺1) relative permeability or hydraulic conductivity (m s⫺1) Henry’s Law constant [kg/(m3–atm)] half-velocity coefficient for terminal e-acceptor (kg m⫺3) half-velocity coefficient for electron donor (kg m⫺3) average mass of bacterial cell (kg) molecular weight of adsorbing molecule or cell (kg mol⫺1) porosity (m3 m⫺3) partial pressure of gas component k, (atm) Darcian fluxes of liquid and gas phases (m s⫺1) respiration coefficient (mol mol⫺1) gas constant [(L atm)/(mol K)] solute retardation factor absolute and effective fluid saturations (m3 m⫺3) time (s) absolute temperature (K) fluid velocity (m s⫺1) mole fraction (partial pressure) of gas component k yield coefficient (kg kg⫺1) collision or sticking efficiency longitudinal and transverse dispersivities, (m) Kronecker delta function equilibrium spreading pressure (mN m⫺1) surface “excess” adsorbed to gas-liquid interfaces (kg m⫺2) collector efficiency volumetric liquid (water) and gas (air) contents (m3 m⫺3) microbial metabolic lag, Brooks and Corey function parameter reaction rate source–sink term (kg s⫺1) specific and maximum specific growth rates, (s⫺1) fluid viscosity [kg/(m s)] fractional area of solid-liquid interfaces consisting of s2 sites density of aqueous solution (kg m⫺3) gas–liquid interfacial tension with and without cells or surfactants (mN m⫺1) exposure time of cells to substrate (s) lag time before cells start to metabolize substrate (s) time required to reach exponential growth stage (s) time-scale for diffusion across gas-liquid interface (s) fluid source sink term (kg s⫺1) ACKNOWLEDGMENTS We would like to thank Markus Flury, Brian Wood, and an anonymous reviewer for their helpful critiques of this paper. REFERENCES Absolom, D.R., F.V. Lamberti, Z. Policova, W. Zingg, C.J. van Oss, and A.W. Neumann. 1983. Surface thermodynamics of bacterial adhesion. Appl. Environ. Microbiol. 46:90–97. Adamson, A.W., and A.P. Gast. 1997. Physical chemistry of surfaces. John Wiley and Sons, New York. Atlas, R.M., and R. Bartha. 1993. Microbial ecology—Fundamentals and applications. 3rd ed. Benjamin/Cummings, Redwood City, CA. Bailey, J.E., and D.F. Ollis. 1986. Biochemical engineering fundamentals. McGraw-Hill, New York. Barry, D.A., H. Prommer, C.T. Miller, P. Engesgaard, A. Brun, and C. Zheng. 2002. Modelling the fate of oxidisable organic contaminants in groundwater. Adv. Water Resour. 25:945–983. Battistelli, A. 2003. Modeling the biodegradation of organic contaminants with TMVOCBIO. In Proceedings of the TOUGH Symposium 2003 Lawrence Berkeley Laboratory, Berkeley, CA. 12–14 May 2003. LBNL, Berkeley, CA. Battistelli, A., C. Calore, and K. Pruess. 1997. The simulator TOUGH2/ EWASG for modeling geothermal reservoirs with brines and noncondensible gas. Geothermics 26:437–464. Bear, J. 1972. Dynamics of fluids in porous media. Dover Publications, New York. Bengtsson, G., and R. Lindqvist. 1995. Transport of soil bacteria controlled by density-dependent sorption kinetics. Water Resour. Res. 31:1247–1256. Berg, H.C. 2000. Motile behavior of bacteria. Phys. Today 1:24–29. Bolster, C.H., G.M. Hornberger, A.L. Mills, and J.L. Wilson. 1998. A method for calculating bacterial deposition coefficients using the fraction of bacteria recovered from laboratory columns. Environ. Sci. Technol. 32:1329–1332. Bos, R., H.C. van der Mei, and H.J. Busscher. 1999. Physico-chemistry of initial microbial adhesive interactions– its mechanisms and methods for study. FEMS Microbiol. Rev. 23:179–230. Brooks, R.H., and A.T. Corey. 1964. Hydraulic properties of porous media. Hydrol. Paper 3. Colorado State University, Fort Collins. Burdine, N.T. 1953. Relative permeability calculations from size distribution data. Trans. Am. Inst. Mining Metall. Pet. Eng. 198:71–77. Cary, J.W. 1994. Estimating the surface area of fluid phase interfaces in porous media. J. Contam. Hydrol. 15:243–248. Celia, M.A., J.S. Kindred, and I. Herrera. 1989. Contaminant transport and biodegradation. 1. A numerical model for reactive transport in porous media. Water Resour. Res. 25:1141–1104. Chen, G., M. Rockhold, and K.A. Strevett. 2003. Equilibrium and kinetic controlled adsorption of bacteria on alluvial sand and surface thermodynamic interpretation. Res. Microbiol. 154:175–181. Chen, G., and K.A. Strevett. 2001. Impact of surface thermodynamics on bacterial transport. Environ. Microbiol. 3:237–245. Chen, G., and K.A. Strevett. 2002. Surface free energy relationships used to evaluate microbial transport. J. Environ. Eng. 125:445–453. Chenu, C. 1993. Clay- or sand-polysaccharide associations as models for the interface between micro-organisms and soil: Water related properties and microstructure. Geoderma 56:143–156. Chiang, H.-W., and C. Tien. 1985a. Dynamics of deep-bed filtration. Part I: Analysis of two limiting situations. AIChE J. 8:1349–1359. Chiang, H.-W., and C. Tien. 1985b. Dynamics of deep-bed filtration. Part II: Experiment. AIChE J. 8:1360–1371. Chilakapati, A. 1995. RAFT: A simulator for reactive flow and transport of groundwater contaminants. PNNL-10636. Pacific Northwest Natl. Lab., Richland, WA. Chu, Y., Y. Jin, M. Flury, and M.V. Yates. 2001. Mechanisms for virus removal during transport in unsaturated porous media. Water Resour. Res. 37:253–263. Clement, T.P., B.M. Peyton, R.S. Skeen, D.A. Jennings, and J.N. Petersen. 1997. Microbial growth and transport under denitrification conditions: Experiments and simulations. J. Contam. Hydrol. 24:269–285. Coats, K.H., and B.D. Smith. 1964. Dead-end pore volume and dispersion in porous media. SPE J. 4:73–84. Corapcioglu, M.Y., and A. Haridas. 1984. Microbial transport in soils and groundwater: A numerical model. Adv. Water Resour. 8:188–200. Corapcioglu, M.Y., and H. Choi. 1996. Modeling colloid transport in unsaturated porous media and validation with laboratory column data. Water Resour. Res. 12:3437–3449. Cunningham, A.B., W.G. Characklis, F. Abedeen, and D. Crawford. Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. www.vadosezonejournal.org 1991. Influence of biofilm accumulation on porous media hydrodynamics. Environ. Sci. Technol. 25:1305–1311. Daeschel, M.A., and J. McGuire. 1998. Interrelationships between protein surface adsorption and bacterial adhesion. Biotechnol. Genet. Eng. Rev. 15:413–438. Danckwerts, P.V. 1970. Gas–liquid reactions. McGraw-Hill, New York. De Beer, D., and A. Schramm. 1999. Micro-environments and mass transfer phenomena in biofilms studied with micro-sensors. Water Sci. Technol. 7:173–178. Deshpande, P.A., and D.R. Shonnard. 1999. Modeling the effects of systematic variation in ionic strength on the attachment kinetics of Pseudomonas fluorescens UPER-1 in saturated sand columns. Water Resour. Res. 5:1619–1627. Déziel, P., G. Paquette, R. Villemur, F. Lepine, and J. Bisaillon. 1996. Biosurfactant production by a soil Pseudomonas strain growing on polycylic aromatic hydrocarbons. Appl. Environ. Microbiol. 62: 1908–1912. Docoslis, A., R.F. Giese, and C.J. van Oss. 2000. Influence of the water–air interface on the apparent surface tension of aqueous solutions of hydrophilic solutes. Colloids Surf. B 19:147–162. Ford, R.M. 1992. Mathematical modeling and quantitative characterization of bacterial motility and chemotaxis. p. 177–215. In C.J. Hurst (ed.) Modeling the metabolic and physiologic activities of microorganisms. John Wiley and Sons, New York. Freijer, J.I., and P.A. Leffelaar. 1996. Adapted Fick’s law applied to soil respiration. Water Resour. Res. 32:791–800. Gannon, J.T., V.B. Manilal, and M. Alexander. 1991. Relationship between cell surface properties and transport of bacteria through soil. Appl. Environ. Microbiol. 57:190–193. Gerson, D.F. 1993. The biophysics of microbial surfactants: Growth on insoluble substrates. p. 269–286. In N. Kosaric (ed.) Biosurfactants. Marcel-Dekker, New York. Ginn, T.R. 1999. On the distribution of multi-component mixtures over generalized exposure time in subsurface flow and reactive transport: Foundations, and formulations for groundwater age, chemical heterogeneity, and biodegradation. Water Resour. Res. 35:1395–1407. Ginn, T.R., B.D. Wood, K.E. Nelson, T.D. Scheibe, E.M. Murphy, and T.P. Clement. 2002. Processes in microbial transport in the natural subsurface. Adv. Water Resour. 25:1017–1042. Harvey, R.W., and S.P. Garabedian. 1991. Use of colloid filtration theory in modeling movement of bacteria through a contaminated sandy aquifer. Environ. Sci. Technol. 25:178–185. Hendry, M.J., J.R. Lawrence, and P. Maloszewski. 1997. The role of sorption in the transport of Klebsiella oxytoca through saturated silica sand. Ground Water 35:574–584. Hendry, M.J., J.R. Lawrence, and P. Maloszewski. 1999. Effects of velocity on the transport of two bacteria through saturated sand. Ground Water 1:103–112. Hermansson, M. 1999. The DLVO theory in microbial adhesion. Colloids Surf. B 14:105–119. Higbie, R. 1935. The rate of absorption of a pure gas into a still liquid during short periods of exposure. Trans. AIChE 31:365–389. Hirshfelder, J.O., C.F. Curtiss, and R.B. Bird. 1964. Molecular theory of gases and liquids. Wiley, New York. Hornberger, G.M., A.L. Mills, and J.S. Herman. 1992. Bacterial transport in porous media: Evaluation of a model using laboratory observations. Water Resour. Res. 28:915–938. Jaynes, D.B., and A.S. Rogowski. 1983. Applicability of Fick’s Law to gas diffusion. Soil Sci. Soc. Am. J. 47:425–430. Jenneman, G.E., R.M. Knapp, M.J. McInerney, D.E. Menzie, and D.E. Revus. 1984. Experimental studies of in situ microbial enhanced oil recovery. SPE J. 24:33–37. Jewett, D.G., B.E. Logan, R.G. Arnold, and R.C. Bales. 1999. Transport of Pseudomonas fluorescens through quartz sand as a function of water content. J. Contam. Hydrol. 36:73–89. Jin, Y., and M. Flury. 2002. Fate and transport of viruses in porous media. Adv. Agron. 77:39–102. Jin, Y., M.V. Yates, and S.R. Yates. 2002. Microbial transport. p. 1482–1509. In J.H. Dane and G.C. Topp (ed.) Methods of soil analysis. Part 4. SSSA Book Ser. 5. SSSA, Madison, WI. Johnson, W.P., K.A. Blue, B.E. Logan, and R.G. Arnold. 1995. Modeling bacterial detachment during transport as a residence-timedependent process. Water Resour. Res. 31:2649–2658. 381 Kosaric, N. (ed.) 1993. Biosurfactants. Marcel Dekker, New York. Kral, D.M. (ed.) 1981. Water potential relations in soil microbiology. SSSA Spec. Publ. 9. SSSA, Madison, WI. Lawrence, J.R., P.J. Delaquis, D.R. Korber, and D.E. Caldwell. 1987. Behavior of Pseudomonas fluorescens within the hydrodynamic boundary layers of surface microenvironments. Microb. Ecol. 14: 1–14. Lee, W.-K., J. McGuire, and M.K. Bothwell. 1999. A mechanistic approach to modeling protein adsorption at solid–water interfaces. J. Colloid Interface Sci. 213:265–267. Leffelaar, P.A. 1987. Dynamics of partial anaerobiosis, denitrification, and water in a soil aggregate. Simulation. Soil Sci. 146:427–444. Lenhard, R.J., M. Oostrom, C.S. Simmons, and M.D. White. 1995. Investigation of density-dependent gas advection of trichloroethylene: Experiment and a model validation exercise. J. Contam. Hydrol. 19:47–67. Leverett, M.C. 1941. Capillary behavior in porous solids. Trans. Am. Inst. Min. Metall. Pet. Eng. 142:152–169. Lide, D.R. (ed.) 1996. Handbook of chemistry and physics. CRC Press, Inc., Boca Raton, FL. Loehr, R.C. (ed.) 1977. Land as a waste management alternative. Proceedings of the 1976 Cornell Agricultural Waste Management Conference. Ann Arbor Science Publ., Ann Arbor, MI. Logan, B.E., D.G. Jewett, R.G. Arnold, E.J. Bouwer, and C.R. O’Melia. 1995. Clarification of clean-bed filtration models. J. Environ. Eng. 121:869–873. MacDonald, T.R., P.K. Kitanidis, P.L. McCarty, and P.V. Roberts. 1999a. Effects of shear detachment on biomass growth and in situ bioremediation. Ground Water 4:555–563. MacDonald, T.R., P.K. Kitanidis, P.L. McCarty, and P.V. Roberts. 1999b. Mass-transfer limitations for macroscale bioremediation modeling and implications on aquifer clogging. Ground Water 4: 523–531. Madigan, M.T., J.M. Martinko, and J. Parker. 1997. Brock biology of microorganisms. 8th ed. Prentice Hall, Upper Saddle River, NJ. Massman, J., and D.F. Farrier. 1992. Effects of atmospheric pressures on gas transport in the vadose zone. Water Resour. Res. 28:777–791. McCalla, T.M. 1946. The influence of microorganisms and some organic substances on water percolation through a layer of Peorian loess. Soil Sci. Soc. Am. Proc. 10:175–179. McDowell-Boyer, L.M., J.R. Hunt, and N. Sitar. 1986. Particle transport through porous media. Water Resour. Res. 22:1901–1921. Megee, R.D. III, S. Kinoshita, A.G. Fredrickson, and H.M. Tsuchiya. 1970. Differentiation and product formation in molds. Biotechnol. Bioeng. 12:771–801. Millington, R.J., and J.M. Quirk. 1960. Transport in porous media. p. 97–106. In F.A. Van Beren et al. (ed.) Trans. 7th Int. Congr. Soil Sci., Vol. 1., Madison, WI. 14–24 Aug. 1960. Elsevier, Amsterdam. Millington, R.J., and J.M. Quirk. 1961. Permeability of porous solids. Trans. Faraday Soc. 57:1200–1207. Mills, A.L., J.S. Herman, G.M. Hornberger, and T.H. DeJesús. 1994. Effect of solution ionic strength and iron coatings on mineral grains on the sorption of bacterial cells to quartz sand. Appl. Environ. Microbiol. 60:3300–3306. Moldrup, P., T. Olesen, J. Gamst, P. Schjønning, T. Yamaguchi, and D.E. Rolston. 2000. Predicting the gas diffusion coefficient in repacked soil: Water-induced linear reduction model. Soil Sci. Soc. Am. J. 64:1588–1594. Molz, F.J., M.A. Widdowson, and L.D. Benefield. 1986. Simulation of microbial growth dynamics coupled to nutrient and oxygen transport in porous media. Water Resour. Res. 22:1207–1216. Mualem, Y. 1976. A new model for predicting the hydraulic conductivity of unsaturated porous media. Water Resour. Res. 12:513–522. Murphy, E.M., and T.R. Ginn. 2000. Modeling microbial processes in porous media. Hydrogeol. J. 8:142–158. Nelson, K.E., and T.R. Ginn. 2001. Theoretical investigation of bacterial chemotaxis in porous media. Langmuir 17:5636–5645. Nevo, Z., and R. Mitchell. 1967. Factors affecting biological clogging of sand associated with ground water recharge. Water Res. 1:231–236. Nielsen, D.R., and J.W. Biggar. 1961. Miscible displacement in soils: I. Experimental information. Soil Sci. Soc. Am. Proc. 25:1–5. Niemet, M.R., M.L. Rockhold, N. Weisbrod, and J.S. Selker. 2002. Relationships between gas–liquid interfacial surface area, liquid Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. 382 VADOSE ZONE J., VOL. 3, MAY 2004 saturation, and light transmission in variably saturated porous media. Water Resour. Res. 38. doi: 38:10.1029/2001WR000785. Oldenburg, C.M., K. Pruess, and S.M. Benson. 2001. Process modeling of CO2 injection into natural gas reservoirs for carbon sequestration and enhanced gas recovery. Energy Fuels 15:293–298. Olesen, T., P. Moldrup, K. Henriksen, and L.W. Petersen. 1996. Modeling diffusion and reaction in soils: IV. New models for predicting ion diffusivity. Soil Sci. 161:633–645. Oostrom, M., M.D. White, and M.L. Brusseau. 2001. Theoretical estimation of free and entrapped nonwetting–wetting fluid interfacial areas in porous media. Adv. Water Resour. 24:887–898. O’Melia, C.R. 1985. Particles, pretreatment, and performance in water filtration. J. Environ. Eng. 6:874–890. Ouyang, Y., and L. Boersma. 1992. Dynamic oxygen and carbon dioxide exchange between soil and atmosphere. I. Model development. Soil Sci. Soc. Am. J. 56:1695–1702. Overcash, M.R., and D. Pal. 1979. Design of land treatment systems for industrial wastes—Theory and practice. Ann Arbor Science Publ., Ann Arbor, Michigan. Powelson, D.K., and A.L. Mills. 1998. Water saturation and surfactant effects on bacterial transport in sand columns. Soil Sci. 163:694–704. Pruess, K., and A. Battistelli. 2002. TMVOC, a numerical simulator for three-phase non-isothermal flows of multicomponent hydrocarbon mixtures in saturated-unsaturated heterogeneous media. LBNL-49375. Lawrence Berkeley Natl. Lab., Berkeley, CA. Raiders, R.A., M.J. McInerney, D.E. Revus, H.M. Torbati, R.M. Knapp, and G.E. Jenneman. 1986. Selectivity and depth of microbial plugging in Berea sandstone cores. J. Ind. Microbiol. 1:195–203. Rajagopalan, R., and C. Tien. 1976. Trajectory analysis of deep-bed filtration with the sphere-in-cell porous media model. AIChE J. 3:523–533. Richards, L.A. 1931. Capillary conduction of liquids through porous mediums. Physics 1:318–333. Rittmann, B.E. 1982. The effect of shear stress on biofilm loss rate. Biotechnol. Bioeng. 24:501–506. Roberson, E.M., and M.K. Firestone. 1992. Relationship between desiccation and exopolysaccharide production in a soil Pseudomonas sp. Appl. Environ. Microbiol. 58:1284–1291. Rockhold, M.L., R.R. Yarwood, M.R. Niemet, P.J. Bottomley, and J.S. Selker. 2002. Considerations for modeling bacterial-induced changes in hydraulic properties of variably saturated porous media. Adv. Water Resour. 25:477–495. Salvage, K.M., and G.-T. Yeh. 1998. Development and application of a numerical model of kinetic and equilibrium microbiological and geochemical reactions (BIOKEMOD). J. Hydrol. (Amsterdam) 209:27–52. Schäfer, A., P. Ustohal, H. Harms, F. Stauffer, T. Dracos, and A.J.B. Zehnder. 1998. Transport of bacteria in unsaturated porous media. J. Contam. Hydrol. 33:149–169. Scheibe, T.D., and B.D. Wood. 2003. A particle-based model of size or anion exclusion with application to microbial transport in porous media. Water Resour. Res. 39(4):1080. doi:0.1029/2001WR001223. Scheidegger, A.E. 1961. General theory of dispersion in porous media. J. Geophys. Res. 66:3273–3278. Schügerl, K. 1982. Characterization and performance of single- and multi-stage tower reactors with outer loop for cell mass production. p. 94–217. In A. Fiechter (ed.) Advances in biochemical engineering. 22A. Springer-Verlag, New York. Šimůnek, J., M. Šejna, and M.Th. van Genuchten. 1999. The HYDRUS-2D software package for simulating two-dimensional movement of water, heat, and multiple solutes in variably saturated porous media, Version 2.0, Rep. IGWMC-TPS-53. Int. Ground Water Model. Ctr., Colorado School of Mines, Golden, CO. Šimůnek, J., and D.L. Suarez. 1993. Modeling of carbon dioxide transport and production in soil. 1. Model development. Water Resour. Res. 29:487–497. Skopp, J. 1985. Oxygen uptake and transport in soils: Analysis of the air–water interfacial area. Soil Sci. Soc. Am. J. 49:1327–1331. Smith, J.E., and R.E. Gillham. 1994. The effect of concentrationdependent surface tension on the flow of water and transport of dissolved organic compounds: A pressure head-based formulation and numerical model. Water Resour. Res. 30:343–354. Speitel, G.E., and F.A. DiGiano. 1987. Biofilm shearing under dynamic conditions. J. Environ. Eng. 3:464–475. Taylor, S.W., and P.R. Jaffe. 1990a. Biofilm growth and the related changes in the physical properties of a porous medium. 1. Experimental investigation. Water Resour. Res. 26:2153–2159. Taylor, S.W., and P.R. Jaffe. 1990b. Biofilm growth and the related changes in the physical properties of a porous medium. 3. Dispersivity and model verification. Water Resour. Res. 26:2171–2180. Taylor, S.W., and P.R. Jaffe. 1990c. Substrate and biomass transport in a porous medium. Water Resour. Res. 26:2181–2194. Taylor, S.W., P.C.D. Milly, and P.R. Jaffe. 1990. Biofilm growth and related changes in the physical properties of a porous medium. 2. Permeability. Water Resour. Res. 26:2161–2169. Thompson, S.W., M. Flury, M.V. Yates, and W.A. Jury. 1998. Role of air–water–solid interface in bacteriophage sorption experiments. Appl. Environ. Microbiol. 64:304–309. Thorstenson, D.C., and D.P. Pollock. 1989. Gas transport in unsaturated zones: Multicomponent systems and the adequacy of Fick’s Law. Water Resour. Res. 25:477–507. Tien, C., R.M. Turian, and H. Pendse. 1979. Simulation of the dynamic behavior of deep bed filters. AIChE J. 3:385–395. Travis, B.J., and N.D. Rosenberg. 1997. Modeling in situ bioremediation of TCE at Savannah River: Effects of product toxicity and microbial interactions on TCE degradation. Environ. Sci. Technol. 31:3093–3102. Trulear, M.G., and W.G. Characklis. 1982. Dynamics of biofilm processes. J. Water Pollut. Control Fed. 54:1288. Vandevivere, P. 1995. Bacterial clogging of porous media: A new modeling approach. Biofouling 8:281–291. Vandevivere, P., and P. Baveye. 1992a. Relationship between transport of bacteria and their clogging efficiency in sand columns. Appl. Environ. Microbiol. 58:2523–2530. Vandevivere, P., and P. Baveye. 1992b. Saturated hydraulic conductivity reduction caused by aerobic bacteria in sand columns. Soil Sci. Soc. Am. J. 56:1–13. Vandevivere, P., and D.L. Kirchman. 1993. Attachment stimulates exopolysaccharide synthesis by a bacterium. Appl. Environ. Microbiol. 59:3280–3286. van Genuchten, M.Th. 1980. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 44:892–898. van Genuchten, M.Th., and R.W. Cleary. 1979. Movement of solutes in soils: Computer-simulated and laboratory results. p. 349–386. In G.H. Bolt (ed.) Soil chemistry. B. Physico-chemical models. Elsevier, New York. van Genuchten, M.Th., and M. Wagenet. 1989. Two-site/two-region models for pesticide transport and degradation: Theoretical development and analytical solutions. Soil Sci. 53:1303–1310. van Oss, C.J. 1994. Interfacial forces in aqueous media. Marcel Dekker, New York. Wan, J., and J.L. Wilson. 1994. Colloid transport in unsaturated porous media. Water Resour. Res. 30:857–864. Wan, J., J.L. Wilson, and T.L. Kieft. 1994. Influence of gas-water interface on transport of microorganisms through unsaturated porous media. Appl. Environ. Microbiol. 60:509–516. Weisbrod, N., M.R. Niemet, and J.S. Selker. 2003. Light transmission technique for the evaluation of colloidal transport and dynamics in porous media. Environ. Sci. Technol. 37:3694–3700. Whitaker, S. 1986. Transient diffusion, adsorption, and reaction in a porous catalyst. Chem. Eng. Sci. 41:3015–3022. White, M.D., and M. Oostrom. 1996. STOMP—Subsurface Transport Over Multiple Phases. Theory guide. PNNL-11217. Pacific Northwest Natl. Lab., Richland, WA. White, M.D., and M. Oostrom. 1998. Modeling surfactant-enhanced nonaqueous-phase liquid remediation of porous media. Soil Sci. 163:931–940. White, M.D., and M. Oostrom. 2003. STOMP—Subsurface Transport Over Multiple Phases, Version 3.0 user’s guide. PNNL-14286. Pacific Northwest Natl. Lab., Richland, WA. Widdowson, M.A., F.J. Molz, and L.D. Benefield. 1988. A numerical transport model for oxygen- and nitrate-based respiration linked to substrate and nutrient availability in porous media. Water Resour. Res. 24:1553–1565. Williams, V., and M. Fletcher. 1996. Pseudomonas fluorescens adhesion and transport through porous media are affected by lipopolysaccharide composition. Appl. Environ. Microbiol. 62:100–104. Reproduced from Vadose Zone Journal. Published by Soil Science Society of America. All copyrights reserved. www.vadosezonejournal.org Wood, B.D., T.R. Ginn, and C.N. Dawson. 1995. Effects of microbial metabolic lag in contaminant transport and biodegradation modeling. Water Resour. Res. 31:553–563. Yarwood, R.R., M.L. Rockhold, M.R. Niemet, J.S. Selker, and P.J. Bottomley. 2002. Bioluminescense as a nondestructive measure of microbial cell density and distribution in unsaturated porous media. Appl. Environ. Microbiol. 68:3597–3605. Yee, N., J.B. Fein, and C.J. Daughney. 2000. Experimental study of the pH, ionic strength, and reversibility behavior of bacteria–mineral adsorption. Geochim. Cosmochim. Acta 64:609–617. 383 Yeh, G.T., and V.S. Tripathi. 1991. HYDROGEOCHEM: A coupled model of hydrologic transport and geochemical equilibria in reactive multi-component systems. ORNL-6371. Oak Ridge Natl. Lab., Oak Ridge, TN. Zysset, A., F. Stauffer, and T. Dracos. 1994a. Modeling of chemically reactive groundwater transport. Water Resour. Res. 30:2217–2228. Zysset, A., F. Stauffer, and T. Dracos. 1994b. Modeling of reactive groundwater transport governed by biodegradation. Water Resour. Res. 30:2423–2434.