Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
REVIEW ARTICLE Mutations of the Red Blood Cell Membrane Proteins: From Clinical Evaluation to Detection of the Underlying Genetic Defect By Jiri Palek and Kenneth E. Sahr S E V E W RECENT publications have reviewed the principal structural features of the membrane proteins, the organization of their genes, and the recently described membrane protein The objective of this review is to apply this rapidly expanding knowledge to the understanding of the pathobiology of red blood cell (RBC) lesions and the detection of the underlying membrane protein defects in inherited disorders of the RBC membrane proteins. MACROMOLECULAR ORGANIZATION OF THE MEMBRANE SKELETON: IMPLICATIONS FOR MEMBRANE STABILITY, DEFORMABILITY, AND CELL SHAPE Electron microscopic images of uniformly extended membrane skeletal preparations show a relatively uniform network of hexagons with occasional pentagons and heptagons (Fig 1). The hexagon corners contain oligomeric actin together with proteins 4.1 and adducin, which bind to both spectrin and actin, thereby facilitating contact between the two proteins, and protein 4.9.1° The hexagon arms are composed of fibers of spectrin tetramers (SpT) and, occasionally, double-stranded tetramers or higher oligomers. Near the center of the SpT, where the two a.p spectrin heterodimers (SpD) assemble head to head to form SpT, the SpT are decorated by small globular structures representing ankyrin, the principal attachment site of the skeleton to the major transmembrane protein band 3, the anion exchange protein." Destabilization of the lipid bilayer, microvesiculatwn, and spherocytosk. In the intact erythrocyte membrane, the skeletal network is in a "condensed" configuration with the spectrin subunits randomly folded in a close apposition12 (Fig 2). The skeletal proteins cover approximately 60% of the inner membrane surface and bind weakly to the lipids of the inner leaflet of the bilayer.14-18 Consequently, the skeleton confers stability to the fluid lipid bilayer. A decreased density of the skeletal network, such as seen in spectrin-deficient RBCs in hereditary spherocytosis (HS), is thought to facilitate uncoupling of parts of the lipid bilayer from the underlying skeleton (Fig 2). An enhanced lipid bilayer-skeleton uncoupling in HS RBCs is readily demonstrable under a variety of conditions in vitro.' It is From the Department of Biomedical Research and the Diviswn of HematologylOncology, St Elizabeth's Hospital of Boston, Tufs University School of Medicine, Boston, MA. Submitted July 9,1991; accepted April 7,1992. Supported by National Institutes of Health Grants No. HL-37462 and HL-27215. Address reprint requests to Jiri Palek, MD, Department of Biomedical Research, St Elizabeth's Hospital of Boston, 736 Cambridge St, Boston, MA 02135. 0 1992 by The American Society of Hematology. 0006-4971l92/8002-OO31$3.00l0 308 also likely to take place in vivo, thus providing a plausible explanation for the surface area deficiency observed in hereditary spherocytosis.' Membrane fragmentation. The condensed configuration of the skeleton in intact cells also underlies the high degree of cell membrane deformability by permitting a considerable extension of the skeletal network followed by elastic recovery of the biconcave shape after the deforming force has been discontinued (Fig 3). An example is the transformation of RBCs into extended ellipsoids induced by a shear ~ t r e s s . ' ~Such J ~ elongated ellipsoids resume a biconcave shape after the shear stress has ceased. They fragment only if the length of the deformed ellipsoids exceeds the length of the maximally unidirectionally extended membrane skeleton. In subjects with hereditary elliptocytosis (HE) carrying a. spectrin or p spectrin mutations that weaken the SpD-SpD contact, the hexagonal skeletal lattice is disrupted (Fig 4), leading to RBC fragmentation and poikilocytosis.1,20In severely affected subjects, such as homozygotes or double heterozygotes for one or two spectrin mutations, or patients with hereditary pyropoikilocytosis (HPP; see below), fragmentation is readily seen during examination of the peripheral blood film.' In mildly affected subjects (ie, simple heterozygotes carrying a mildly dysfunctional spectrin) marked RBC fragmentation is seen only in conjunction with another factor such as a mechanical insult to the cells, exemplified by disseminated intravascular coagulation,2l or another unrelated skeletal defect. For example, neonates carrying elliptocytogenic spectrin mutations have a considerably more severe hemolysis, fragmentation, and poikilocytosisthan a d ~ l t sbecause ~ ~ , ~of~ the additive effect of a weakened spectrin-protein 4.1 contact induced by increased concentrations of free 2,3diphosphoglycerate; the elevation of this compound is due to its weak binding to fetal h e m o g l ~ b i n ? ~ - ~ ~ Plastic deformation and formation of elliptocytes. When normal RBCs are deformed for prolonged periods of time, the proteins of the deformed skeleton undergo an active rearrangement involvinga disconnection of existing proteinprotein contacts and a formation of new associations that permanently stabilize the cells in the deformed shapeZ7sz (Fig 3). It is possible that the elliptical shape of HE RBCs is a consequence of such permanent plastic deformation. Cells that flow through microcirculation or cells subjected to shear stress have either a parachute-like or elliptical ~ h a p e ~ in ~ .HE, ~ ~ ;such shape may become permanent because the weakened protein-protein contacts (such as the weakened SpD-SpD association) facilitate skeletal reorganization.28 Indeed, HE-nucleated RBC precursors are round' and the elliptical deformation progressively increases with increase in cell density and, presumably, cell age.31 Blood, VOI 80, NO2 (July 15). 1992: pp 308-330 RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS 309 - . ... i A Fig 1. RBC membrane skeleton prepared by removing membrane lipids and integral proteins by extraction with Triton-X 100. The skeleton has lmen artificially extended so that the individual structural components can be seen. Low magnification (a) shows a highly ordered, relatively uniform structure. High magnification images (b through e) show that this network is assembled into hexagons (b and c) or heptagons and pentagons (d and e). The arms of each polygon are formed by spectrin tetramers (Sp4) and, occasionally, double tetramers (2Sp4) or hexamers (Sp6). as highlighted In (b) and (c). The tetramer arms are interconnected by junctional complexes containing actin and protein 4.1. In the middle of the SpT are globular structures representing ankyrin (b and c). This composite figure is compiled in part from previously published work’with permission. BIOGENESIS OF THE RBC MEMBRANE SKELETON: RELEVANCE TO CLINICAL EXPRESSION OF MEMBRANE PROTEIN MUTATIONS Whereas the synthesis of both spectrin and ankyrin can be readily detected at the earliest stages of erythroid differentiation,3*-” only a small fraction of the newly synthesized spectrin and ankyrin is assembled on the RBC membrane, and the membrane-associated spectrin and ankyrin turn over rapidly. At the proerythroblast stage, band 3 synthesis is initiatcd and, together with the synthesis of the protein 4.1, increases up to the late erythroblast stage?2.3s.MDuring this time, the messenger RNA (mRNA) C levels and the synthesis of spectrin and ankyrin decline. In a striking contrast, the fraction of the newly assembled spcctrin and ankyrin on the membrane and their net amounts progressively and the turnover of these proteins on the membrane decline^.^"^.^ This paradoxical finding is related to a progressive increase in the synthesis of band 3 and 4.1 proteins that serve as the principle membrane anchors of the s k e l e t ~ n . ~ ~ . ~ ~ , ~ An important feature of membrane skeletal biogenesis, common to both mammalian and avian RBCs. is a seemingly wasteful synthesis of both spectrin and ankyrin and, to a lesser extent, the 4.1 protein, with a relatively small 310 PALEK AND SAHR Fig 2 Ulhmuctun of the membrane skeleton in intact normal RBC ghosts (a) and in ghosts from a patient with severe HS associated with a deficiency of spectrin (b). Hemoglobin-free ghosts were sonicated t o open a "window" that allows the visualization of the intact skeleton at the inner membrane surface. The ghosts were fired and negatively stained. The skeleton is preferentially stained under these conditions. In ghosts from normal RBCs (a), the individual skeletal proteins are in close apposition t o each other, forming a monomolecular layer. In contrast, the HS skeleton (b) is considerably less dense with the individual proteins separated by skeletonfree areas. (c) Separation of the lipid bilayer from the underlying skeleton in normal RBC ghosts. RBC ghosts were suspended in hypotonic sodium phosphate buffer, and, after restoring isotonicity, w e n examined after 10 minutes of incubation. Arrows at the neck region of the spicules show the separation of the membrane lipidsfrom the filamentous skeleton. Arrowheads indicate the vesicles released from the echinocytic ghosts. (c) deplctl this process in normal cells; the release of such vesicles is enhanced in HS.'Z Compiled from previously published work'," with permission. fraction of thc ncwly synthcsizcd a spcctrin, f3 spcctrin, and ankyrin asscmblcd into thc stablc nctwork.3L3H-'"Particularly striking arc thc diffcrcnccs in thc synthcsis of a spcctrin and p spcctrin subunits,"' with thc synthesis of a spcctrin bcing about four timcsgrcater than thc synthesisof p spcctrin. Howcvcr. thc two chains assemble on thc mcmbranc in stoichiomctric amounts as stablc hctcrodimcrs via a high-affinity binding of p spectrin to ankyrin. Thus, thc amount of spcctrin asscmblcd on thc membrane is principally dctcrmincd by the amounts of ncwly synthcsizcd p spcctrin and thc numbcr of availablc ankyrin binding sitcs. Band 3 protcin, thc prcdominant RBC transmembrane protein, appears to determine the ovcrall stoichiometry of the components of the membrane skeleton by providing a high-affinity binding site for a n k ~ r i n . ~Thc . ~ ' intcraction behvcen ankyrin and band 3 proteins also appears to be the key determinant of the pcriphcral localization of the membrane skeIet0n.~3~ The cxccssivc a spcctrin synthesis may explain why a spectrin dcfccts that lcad to a reduced synthesis of a spectrin are clinically apparcnt only when prcscnt in a homozygous or doubly heterozygous form, such as in recessively inherited HS associatcd with a severe spectrin defi~iency.~.~' In a simplc hetcrozygous carricr of such a defect, thc amount of RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS 311 In situ Extended ( lipid-free membrane skeleton) (intact membrane) A Flg 3. Uniaxialelastlc deformation without distention. (A) A schematic diagram of an isolated membrane skeleton that can be uniformly extended t o cover a surface about seven times larger than the surface of the normal RBC membrane. In the intact membrane, the hydrophobicity of the lipid bilayer precludes an Increase in membrane surface area (~3% t o 4’/0) without rupture. However, the membrane can undergo a large deformation under a constant area because of the large exstensibility of the membrane skeleton. (6) During uniaxial extension, the skeleton undergoes an isometric uniaxial stretching from a square t o a long rectangle. After a cessation of an external force, a square surface area is resumed because the elastic skeletal network remains intact, retaining a memory. (C) Prolonged uniaxial extension leads t o a skeletal protein rearrangement, resulting in a formation of new bonds. This leads t o a permanent plastic deformation. A weakening of protein-protein contacts (particularly the SpD-SpD contacts), such as found in common HE, facilitates skeletal protein rearrangement, thus permanently stabilizing the cells in elliptical shape, which is similar t o a shape of RBCs deformed by a shear force. - Uniaxial skeletal extension y= 1 y = 0.5 B x = l x = 2 Extension is no lonaer evident. Skeletal protein reahangement produced a permanent (plastic) deformation. y=l C x = l x = 2 (3) abnormalities involving defective protein-protein intcractions. This last group includes abnormal spectrin-protein 4.1 interaction in HS,XZ.x3a rcduccd binding of ankyrin to the ankyrin stripped inside-out membrane a OVERVIEW OF MOLECULAR DEFECTS OF THE RBC defect that was later found in association with and, possibly, MEMBRANE PROTEINS secondary to spectrin a1/6s mutationx5(see below), and defects Characterized by reduced assembly of SpD to SpT In analogy to other inherited defects, the number of (Table 1). detected membrane protein mutations does not coincide The latter group of mutations, involving defective selfwith their natural prevalence.4? The detection of these association of SpD to SpT, represent the most common mutations is facilitated by their hematologic presentation group of elliptocytogcnicmutations. This functional defect and the underlying abnormality of membrane protein is readily dctcctcd by estimating the relative amounts of composition and function. Consequently, three groups of SpD and SpT in spectrin extracts by nondenaturing gel defects are readily detected either by polyacrylamide gel electrophoresis.”.”.~ In the majority of patients, these electrophoresis (PAGE) of membrane proteins solubilized abnormal spectrins can be further characterized by limited is sodium dodecyl sulfate (SDS) or by protein association tryptic digestion of spectrin: some of the tryptic peptides, as studies (Fig 5): (1) in HS moderate to severe deficiencies of by two dimcnsional electrophoresis, corrcspond to the proteins spectrin,u4 ankyrir1,4~anion e x c h a n g ~ r , ~ . ~resolved ~ distinct structural and functional domains of sp~ctrin!~.~ protein 4.2,sh56 and, in HE, protein 4.1 defi~iency~’-~; (2) Among these peptides, the 80-Kd a1 domain peptide, defects characterized by an abnormal protein migration, such as faster migrating a spectrins6’.“ and f3 ~pectrins~~7-~’ representing the SpD self-association site of a spectrin, is (most ofwhich are associated with HE and exhibit defective the most prominent peptide. Abnormal digestion of this self-association of SpD to SpT), a fast migrating ankyrin in peptide, resulting either from increased susceptibility of the HS,74 and a fast or a slow migrating 4.1 protein in HE62-7s’x1; existing cleavage sites to proteolysis or a formation of new the newly synthesized normal a spectrin, produced by the single normal a spectrin allele, may still exceed the total amount of the newly synthesized f3 spectrin. 312 PALEK AND SAHR Fig 4. Representative electron micrographs of uniformlyspread membrane skeletons from normal RBCs and cells from patients with HS, HE, and HPP. (a) Normal membrane skeleton showing a primarily hexagonal lattice. (b) Skeleton from HS RBCs with a mild rpechin deficiency. The membrane skeletal network appears to be only slightly less intact than in normal cells. (c) Membrane skeleton from a patient with mild heterozygous HE Sp d'", containing dysfunctional spectrin but having normal spectrin content. The integrity of the skeleton is moderately disrupted because of the reduced ability of SpD to self-associate into SpT. (d) Membrane skeletonfrom a patient with HPP. HPP RBCs are not only spectrin-deficient but they also contain dysfunctional spectrin, causing a defective self-association of spectrin dimers into tetramers. Note a striking disruption of the skeleton with a complete loss of skeletal lattice. The magnification of all micrographs is identical. (Adapted from a previous publication for the author's laboratorymwith permission.) cleavage sites, has been detected in the majority of patients with weakened SpD-SpT self-association.'.'2.2~ Several underlying DNA and primary structure defects have been subsequently detected in the vicinity of the abnormal cleavage site (Table 1). These mutations are designated according to size of the largest peptide derived from the abnormal a1 Below, we summarize the major structural, functional, and clinical features of the welldefined mutations of the membrane proteins. Specrrin mururions. This group of elliptocytogenic spectrin mutations is associated with the formation of a 74-Kd peptide resulting from cleavage of arginine or lysine at positions 45 and 48, respectively'3.w.y1 (see Table 1 for the numbering of amino acids, as based on the cDNA sequence). The Lys 48 cleavage site is present in normal spcctrin?' The normal 80-Kd peptide strongly inhibits SpD self-association; the 74-Kd peptide derived from normal spectrin by a more extensivc proteolysis has lost this function,Rgimplying that the critical site involved in SpDSpD self-association is within the first 48 amino acids, largely residing in the first helix of a spectrin. This helical segment has bcen proposed to be a part of a combined a,p spcctrin triple helical unit representing the SpD-SpD contact site4? the first two helices are contributed by the C-terminus of f3 spectrin, while the helix three is the first helical segment of a spectrin (Fig 5). The Sp mutations are of considerable interest because they appear to be the most common membrane protein mutations reported to date, occurring in people of diverse ethnic backgrounds, including whites, blacks, Arabs, and Melanesian~.'.~~ Therefore, it is not surprising that many different mutations are associated with this phenotype (Table 1). Two of these mutations are of particular interest: (1) the mutations of codon 28, representing the most common group of mutations detected to date, involve a substitution of arginine 28 by either histidine, cysteine, leucine, or serincw.y3.w; (2) the mutations of p spectrin that lead to a destabilization of the 48 lysine cleavage site of a spectrin that gives rise to the 74-Kd p e p t i d ~ . 4 ~ Detection . ~ ~ - ~ ~of one of these mutations provided a basis for the above noted model of spectrin self-association in which the last repeat unit of p spectrin lies in direct apposition to the first helix of a spectrin.4' Consequently, mutations within this region lead to a destabilization of the lysine 48 cleavage site (giving rise to the 74-Kd peptide) and a marked weakening of the SpD-SpD self-association. It is also of interest that in the majority, but not in all of these mutations, the self-association of spectrin is severely impaired and the patients have a symptomatic hemolytic anemia. This is particularly the case in the f3 spectrin mutations leading to the Sp p h e n ~ t y p e ~and, ~ . ~ "to a lesser extent, mutations involving a spectrin codon 28.9' However, additional presently unknown genetic factors may modify clinical expression of these mutations: in the case of the 28 Arg to His mutation, a simple heterozygous state for this mutation may present either as a dominantly inherited elliptocytic hemolytic anemia or as an asymptomatic carrier state, an offspring of which has HPP.9' RED BLOOD CELL MEMBRANE PROTEiN MUTATiONS IC 313 t NH2 ,COOH B N Fig 5. A schematic diagram illustrating molecular assembly of membrane skeleton and the approximate location of the molecular defects in HS, HE, and HPP. Known skeletal protein deficiencies in HS and HE include those of spectrin, ankyrin (both in HS), protein 4.1, and glycophorin C (both in HE). The principalskeletal protein dysfunctions (mutations)include (1) defects of the head region of either a or p spectrin, which lead to a defective self-association of SpD into tetramers (arrows, all in HE and/or HPP); (2) defective p spectrin, which binds poorly to protein 4.1 (in HS); and (3) abnormal protein 4.1, which binds weakly to spectrin, thereby destabilizing spectrin4.1-actin interaction (in HE). (A) A schematic illustration of the skeletal protein constituentsand their assembly. Spectrin is composed of two chains, a and p, that are twisted along each other into heterodimers (SpD). At the head region (arrows), SpD are assembled into tetramers (SpT). At their distal end, SpT binds to the junctional complexes of oligomeric actin with the aid of protein 4.1 and adducin. Spectrin is attached to transmembrane proteins by linkage of p spectrin to ankyrin (band 2.1). which binds to the cytoplasmic domain of the anion transport protein. Protein4.2 is likely to be located at this site as well. (B) Schematic illustration of the protein constituentsof the junctional complex. Each SpT end is attached to oligomeric actin by the spectrin binding protein 4.1 and by adducin that binds to two SpT ends. Actin oligomers are stabilized by tropomyosin that lies in the groove of F-actin. Other actin-associated proteins include dematin (an actin-binding protein) and myosin (not shown). The 4.1 protein also binds to glycophorinC. (C) The model of spectrin structure. The u and p chains of spectrin are arranged in an antiparallel fashion with the N-terminus of u chain and the C-terminus of the p chain forming the SpD-SpD contact site. Both spectrin chains are composed of multiple repeat units. The open blocks are highly homologous; the repeat 10 (stippled)of a spectrin is nonhomologous. The repeat units' structure breaks down at the distal end of spectrin tetramers, sharing homology with analogous segments of a actinin and dystrophin (solid rectangles).mal ID) Model of the contact site between the u and p spectrin chains of the opposite heterodimer. Each repeat unit is composed of three u helical segments, connected by short nonhelical regions. The proposed contact site is a combined triple helical unit containing2 u helical segments from repeat 17 of the C-terminus of p spectrin (hatched area) and the first u helical segment of u spectrin near the N4erminus.e Mutations of 0 and p spectrin involving a defective assembly of SpD to SpT are the most common membrane protein defects reported to date (Table 1). The abnormal cleavage sites giving rise to the most commonly seen abnormal tryptic peptides in these mutationsare shown by solid lines; the sites of the mutations, most of which are in the vicinity of the cleavage sites, are designated. Mutations of p spectrin giving rise to a truncated protein are also depicted in this diagram. They are spectrins LePuy.'23 Gottingen.126 Rouen.'z4 Tandil," and Nice.'a (Adapted and reprinted with permission.l2u3) The mutations involving a spectrin codons 46 and 49 appear to be associated with a milder clinical expression,91,99although the percentage of unassembled SpD in the crude spectrin extract is increased from approximately 6% (control value) to values exceeding 50% of the SpD + SpT p 0 0 1 ? ~ ,The ~ reason for this discrepancy between the apparently severe spectrin dysfunction and the relatively mild clinical presentation is not known. The high percentage of SpD could be an artifact of extractions8; in contrast to the normal SpD self-association, which is kinetically immobilized at a temperature of near 0°C (the temperature at which spectrin is extracted from the membrane),lOOJO1 the mutant spectrin self-association may not be kinetically frozen at this temperature, thus permitting some of the mutant SpT to dissociate into SpD during the low ionic strength extraction. Spectrin a1/78 mutations. Although rare, these mutations are of interest because they highlight the complexity of SpD-SpD contact Lower molecular weight and reduced phosphorylation p Sp Gottingen SpD-SpD p Sp Rouen Lower molecular weight and absent phosphorylation SpD-SpD contact p Sp Nice contact Lower molecular weight and absent phosphorylation SpD-SpD contact Lower molecular weight and absent phosphorylation Four abnormal spots (31. 26,21.18 Kd) and a reciprocal decrease of four normal a11 peptides) 78-Kd peptide cleaved at al 16 Lys $ Sp LePuy p Spectrin Sp all Not defined SpD-SpD contact and another unknown defect sp a11131 (Sp Jendouba) SpD-SpD Sp all* Truncated C terminus due to absence of sequences of exon Y and a premature chain termination GAT G to T substitution (+3) in the 5’ splice site causing skipping of exon Y and aframeshift with a premature chain termination T to A substitution in the 5’ splice site following exon X leading to deletion of exon X cDNA sequences and a frameshift leading to premature termination of translation AG duplication at codons 2045/2046 in axon X leading to frameshift and premature chain termination A to G mutation in +4 position of the 5’ donor consensus splice site of the intron following exon X, leading to deletion of exon X cDNA sequences and a frame shift leading to premature termination of translation GCT (Continuedon following page) Truncated C terminus due to absence of sequences encoded by exon X and a premature chain termination Truncated C terminus due to premature chain termination Truncated C terminus due to absence of sequences encoded by exon X and a premature chain termination all Ala + Arg in repeat a9 all 748 Asp + Glu in repeat a7 GAC + GAA 45 AGG + AGT in exon 2 of a Sp a145 Arg + Ser in repeat a1 41 CGG + TGG in exon 2 of a Sp - 46 GGT + G l T in exon 2 of a Sp 49 (37- I T in exon 2 of a Sp 48 AAG + AGG in exon 2 of a Sp 28 CGT TGT in exon 2 of a Sp 28 CGT + C l T in exon 2 of a Sp 28 CGT + AGT in exon 2 of a Sp 28 CGT + CAT in exon 2 of a Sp 2053 GCT CCT in exon X of p Sp a1 28 Arg Cys in repeat a1 a1 28 Arg Leu in repeat a1 aI 28 Arg Ser in repeat a1 a 128 Arg -* His in repeat al pl2053 Ala + Pro in repeat 1317 a1 41 Arg Trp in repeat a1 a1 46 Gly Val in repeat a1 aI 49 Leu -* Phe in repeat a1 aI 48 Lys Arg in repeat al --- 74-Kd peptide cleaved at aI 48 Lys andlor aI 45 Arg SpD-SpD contact a1 471 Glu + Pro in repeat a5 a1469 His + Arg in repeat a5 50-Kd peptide cleaved at a1 468 Arg or 470 Arg SpD-SpD contact Sp a1/-b Sp 471 CAG -* CCG in exon 11 of a Sp 469 CAT -* CGT in exon 11 of a Sp a1 260 Leu + Pro in repeat a3 a1 261 Ser + Pro in repeat a3 aI 207 Leu +Pro in repeat a2 50-Kd peptide cleaved at aI 256 Arg o r a l 258 Lys SpD-SpD contact Sp allm - 260 CTG + CCG in exon 6 of a Sp 261 T C + CCC in exon 6 of a Sp CTG CCG in exon 5 of a Sp aI 154 Leu duplication in repeat a2 154 T G duplication in exon 4 of a Sp DNA Defect 65-Kd peptide cleaved at al 137 Arg Primary Structure. SpD-SpD contact Function Sp a Spectrin Provisional Designation Protein Defect Limited Proteolysis or Other Study Mild to moderately severe common HE Asymptomatic carrier Mild common HE Mild HE Autosomal recessive HS HPP A rare group of p spectrin mutations Probably rare A common group of a Sp mutations, heterogeneousat the primary structure and DNA levels, found in whites, blacks, arabs, and melanesians 124 123-126 41,119 120 102,103 90.91.93.94 104,107,112 Apparently rare Asymptomatic HPP carrier or mild to moderately severe common HE 104,107,115 Relativelycommon in blacks, heterogeneous at the primary structure1 DNA levels HPP Asymptomatic HPP carrier or mild, common HE References Relatively common in 98,104-107 blacks, homogeneous, at protein and DNA levels Prevalence and Comments HE of moderate severity Heterozygote Double Heterozygote or Homozygote Clinical Expression Mild HE Table 1. Membrane Protein Mutations in HS, HE, and HPP W $ x Fm 0 P 2 Incompletely studied Incompletely studied Membrane instability resulting from weakened specwin-actin interaction Truncated C terminus due to 7-bp insertion causing a frameshift and premature termination Primary Structure* - Absence of glycophorin C Partial deficiency of the 4.1 band Deletion of exons 3 and 4 resulting in frameshift and premature stop codon TGGCCG +lTGCG in codons 44.45 resulting in frame shift and a stop codon (nucleotides 166-168) Mutations in codons 44 and 45 Asymptomatic Mild HE Mild common HE Deletion of three exons (including the erythroid initiation codon) precluding mRNA translation Probably rare About 10% of HE cases in North Africa the molecular basis of the defect may be heterogeneous, probably rare Probably rare Mild common HE 240 nucleotide deletion spanning the codons for Lys 407-Gly 486 due to deletion of two exons encoding the spectrin binding Rare Probablyabout 10% of kindred with autosomal dominant HS Prevalence and Comments Probably rare HE with severe hemolysis Double Heterozygote or Homozygote Mild common HE HS of moderate severity Dominantly inherited HS Heterozygote 369 nucleotide duplication of codons for Lys 407-Glu 529 due to a duplication of three exons encoding the spectrin binding domain Deletion of one copy of ankyrin gene due to heterozygousdeletion of chromosome8[de1(8) ( p l l - p Z l . l ) ] TGG + CGG 7-bp deletion in exon X leading to a frameshift and premature chain termination DNA Defect Deletion of exon 3 and 4 15-Kd insertion adjacentto New 44-Kd and 60-Kd chymotryptic peptides with the binding domain the same isoelectric point as the normal 34-and 50-Kd peptides New -40-Kd chymotryptic Lack of the entire spectrin binding domain peptides presumably resulting from fusion of peptides adjacent to the C and N terminus of the deleted sequence About 40% decrease in ankyrin and spectrin content Abnormal digestion pattern Trp + Arg substitution in POsition 202 of p Sp of p Sp N-terminus Lower molecular weight and absent phosphorylation Limited Proteolysis or Other Study Clinical Expression 161 154-156 80.81 80.81 139 130 73 References Putative mutations, which are yet to be characterized at the DNA level, are not included. They are tabulated in Palek and Lambert.' Asymptomatic variants are not included in this table. *Note that the numbering of amino acids and the respective codons of a spectrin are based on the cDNA sequence.220 thus including the first six amino acids that were not detected by protein sequence.92 The repeat numbering of (Y and B spectrin proteins are based on the protein sequences derived from cDNA sequencing.220~227 Glycophorin C deficiency Glycophorin C 4.1 deficiency Low molecular Membrane instability weight 4.1 resulting from (4.168'ffi) weakened spectrin-actin interaction High molecular weight 4.1 (4.1") 4.1 protein Ankyrin deficiency Reduced binding of spectrin to the membrane leading to a secondary deficiency of spectrin Sp-4.1 contact, formation of Sp-4.1actin complex p Sp-4.1 Ankyrin SpD-SpD contact Function p Sp Tandil Provisional Designation Protein Defect Table 1. Membrane Protein Mutations in HS, HE, and HPP (Cont'd) 4z 0 3 -u m g m 2 8 Rr r 5 U m n 316 PALEK AND SAHR regulation of the SpD-SpD self-association as well as the an A to G substitution in position 3 of codon 362, creating a susceptibility of the spectrin a1 domain to proteolysis. The 5' splice site that produces an inframe splicing out of cleavage site giving rise to the 78-Kd peptide involves a codons 363 to 371. As in the majority of other spectrin mutations, the change involves helix 3. Several other spectrin lysine 16 in helix 3 of the combined cup spectrin defects of spectrin, such as the spectrin a1/61 repeat unit.102J03 However, the mutations are 25 and 29 117and spectrin amino acids downstream at positions 41 or 45, the latter a1/43118 have been reported, but the underlying primary being next to one of the amino acid mutations that give rise structure and DNA defects remain to be defined. The spectrin a1/61,first detected in a doubly heterozygous to the Sp a1/74 phenotype?l Spectrin a1165 mutation. This HE mutation is unique individual for this particular mutation, and spectrin ~ $ among the spectrin mutations in that it is homogeneous illustrate some of the technical limitations of two-dimenboth at the protein and DNA levels, being confined to the sional tryptic peptide mapping. Because the amount of the black race?8J04-107This feature is of interest in light of abnormal 61-Kd peptide is very small, the defect is difficult to discern on Coumassie blue-stained gels and it can be recent reports of the relatively high prevalence (0.6%) of HE in equatorial Africa.lo8 Furthermore, RBCs containing detected only on Western blots with polyclonal antibodies mutant spectrin are somewhat resistant to invasion by directed against the a1 domain. malaria and exhibit a reduced parasite growthl@J'lO; thus, it Mutations of the spectrin dZ domain. These rare mutais possible that this mutant spectrin and, possibly, other tions have been detected because of an abnormal migration similar spectrin mutations provide some degree of protecof the a11 domain peptides as shown by two-dimensional tion to malaria parasite invasion. The Sp a1/65 mutation tryptic peptide mapping.41J19-122In contrast to the a1 appears to be the mildest among all a1 domain mutations: domain mutations, in which the disease pathobiology can the spectrin SpD self-association is only mildly impaired be explained by the disruption of the SpD-SpD selfand the clinical expression is very mild; even homozygous association, the link between the a11 mutations and disease subjects carrying this mutation have a relatively mild pathobiology is not clear. For example, the mutation hemolytic anemia that does not require s p l e n e ~ t o m y . ~ ~ J ~ ~involving helix 3 of the repeat 7 of a spectrin is expressed as Spectrin mutations. The cleavage sites leading to a mild dominantly inherited HE with a mild increase in SpD the generation of the 50-Kd tryptic peptide involves either in the crude 0°C spectrin extract," while a replacement of 256 arginine or 258 lysine due to a1260 Leu + Pro or a1261 alanine (GCT) codon by arginine (GAT) in the ninth Ser + Pro substitutions, r e s p e c t i ~ e l y . ~This ~ J ~relatively ~ repeat unit of spectrin is asymptomatic in a simple heterozycommon mutation should be distinguished from a rare gous state; in a homozygous form it is associated with a mutation that is a consequence of either 471 Glu --f Pro severe recessively inherited ~pherocytosis.4~J~~ In another substitution or 469 His + Arg substitution in the fourth defect, the abnormality was located in the C-terminus of the repeat unit (the Sp a1/50b mutation).104J07J12 The group of a11 domain.lZ1The underlying genetic defect involves an Sp a1/50a mutations appears identical to defects designated inframe skipping of exon 18, due to G to A substitution in as Sp a1I6 by another l a b ~ r a t o r y , the ~ ~ differences ~J~~ in the the acceptor splice site of intron 17 (ag/GA + aa/GA). size of the peptides reflecting the difference in estimation of The ensuing protein is shortened by 41 amino acids, the apparent molecular mass. Although apparently conproducing an abnormal tryptic cleavage site after Arg 890 in fined to the black population, the Sp a1/46defects are helix 3 of repeat a9 (J. Delauney et al, personal communicaheterogeneou~.~~ OfJ ~particular ~ J ~ ~ interest is a recently tion, 1992). The phenotype is that of recessively inherited reported Leu + Pro mutation,l15 the site of which is at a severe hemolytic anemia with microspherocytes, poikiloconsiderable distance from the cleavage site, residing 51 cytes, thermal instability of RBCs, and a mild defect in SpD amino acid to the N-terminus in helix 2 of the second repeat self-association. Although a11 domain defects may have unit, thereby destabilizing the cleavage site located in the some influence on the SpD-SpD self-association, it is apposed helix 3. apparent that the underlying pathobiology may be related It is of interest that in the spectrin mutants discussed to other mechanisms that are yet to be defined. above, most of the amino acid substitutions are located in Mutations involving the C-terminus of p spectrin. With the exception of the p spectrin defects leading to increased helix 3 of a given repeat unit, near the connecting segment among the individual repeats. Thus, this region of the susceptibility of the lysine 48 of a spectrin to proteolysis mutations, see above), all known p spectrin spectrin molecule is important in regulating spectrin func(the Sp tion. mutations have been identified on the basis of their increased mobility on SDS-PAGE.67-73 Some of the shortened Other d domain mutations. While the abnormal spectrins giving rise to abnormal a1peptides of 74,65, and 50 Kd p spectrins also enhance the susceptibility to proteolysis of (the Sp a1/74, Sp and Sp (YI/~O, respectively) appear to the 48 lysine of a spectrin, thus causing increased formation represent the most common group of spectrin mutations, of the 74-Kd peptide derived from a ~ p e c t r i n In . ~ ~most of several other defects have been reported, all leading to a the reported truncated p spectrins, the underlying defect defect of SpD-SpD self-association and all creating new has now been defined. In three of them, it involves skipping tryptic cleavage sites. In spectrin Sfax (Sp the first of an exon located near the 3' end of the p spectrin gene reported mutation of repeat 014, the abnormal 36-Kd (designated exon X or exon Y), due to mutations of the 5' fragment starts at Ala 357 and contains a deletion involving donor consensus splice site sequence of the intron following amino acid 363 to 371.116 This deletion is a consequence of the particular exon; the resulting mRNA lacking sequences 1 ~ ~ RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS from exon X or Y contains a frameshift and premature stop ~ o d o n . In~ two ~ ~other - ~ ~reports, the truncated p spectrin is a result of a premature chain termination because of a change in the reading frame due either to a 2-bp insertion in codon 2O46lz6or a 7-bp deletion following codon 2041.73 These mutations resulting in a truncated p spectrin protein also result in a decreased phosphorylation of the C-terminus of p spectrin. Indeed, in spectrin LEPuY,'~~ spectrin Gottigen,lZ and spectrin phosphorylation is absent. This group of mutations is also characterized by defective SpD self-association. Spectrin Rouen is of special interest in this regard because it lacks only the very C-terminal64 residues of p spectrin that are not part of the combined ap triple helical structure, suggesting that this sequence is also involved in SpD self-association. Mutations of the pIVdomain of spectrin. This domain of p spectrin is involved in interaction with protein 4.1.loJz7A recently reported mutation underlies a markedly weakened binding of spectrin to protein 4.1. It is associated with a clinical phenotype of dominantly inherited HS with some acanthocytes on the peripheral blood film.82J28The same functional defect of p spectrin was previously reported by another but the underlying mutation has not been yet defined. The pathophysiologic mechanism whereby this mutation leads to the HS phenotype is unclear. For unexplained reasons, these patients are also partially deficient in spectrin as are nearly all other patients with HS.lZyAs discussed above in the section on the macromolecular assembly of the skeleton, the spectrin deficiency is likely responsible for the HS phenotype in these subjects. HS cells carrying this defect are less mechanically stable than cells from other HS patients30; this is possibly related to the weakened p spectrin-4.1 interaction. In contrast, a disruption of the spectrin-protein 4.1 interaction due to a reciprocal defect involving the spectrin binding domain of the 4.1 protein leads to a phenotype of mild to moderately severe elliptocytosis (see below). Because the abnormal function of the above p spectrin mutant is normalized by reducing agents in vitro,130it is possible that in circulating RBCs the p Sp-protein 4.1 binding is near normal owing to the relatively high concentrations of reducing compounds in the cells. Spectrin polymorphisms. Tryptic peptide mapping has shown several asymptomatic variants involving the aII, aIII, and a V domains of a spectrin, as well as the pIV domain of p ~ p e c t r i n . ' , ~InJ ~the ~ common a11 domain polymorphism, characterized by four distinct phenotypes of different size and isolectric point,132the underlying genetic basis is four haplotypes involving substitutions of Arg 701 to His, Ile 809 to Val, and Thr 853 to Arg in exons 6, 17, and 18, re~pective1y.l~~ Additional reported polymorphisms should be useful in linkage studies attempting to compare such polymorphism with the disease ~ h e n o t y p e . ' ~ ~ - ' ~ ~ Deficiency of ankyrin. In contrast to the rapidly increasing list of the mutations of spectrin, our knowledge of the molecular defects of human ankyrin is incomplete. The observation that the deficiency of ankyrin may represent the primary defect underlying spectrin deficiency in inherited spherocytosis was first made in spherocytic mice 317 carrying the nb m ~ t a t i 0 n . lThe ~ ~ description of ankyrin deficiency in humans followed.46Subsequent studies of one of the originally reported subjects showed a profound decrease of ankyrin synthesis due to an as yet unclear defect leading to a marked decrease in ankyrin mRNA expression. Consequently, assembly of spectrin on the membrane is reduced despite normal spectrin synthesis.138Furthermore, a complete deletion of the ankyrin gene resulting from an interstitial deletion within a short arm of chromosome 8 is associated with the phenotype of moderately severe HS,139 thereby explaining the known association between HS and abnormalities of chromosome 8.140-14 Furthermore, recent studies of linkage of the inheritance of HS with a restriction fragment length polymorphism (RFLP) of the ankyrin gene implicate ankyrin as the primary molecular lesion in a large HS kindred.145 It is possible, but is yet to be firmly established, that ankyrin deficiency in dominantly inherited HS is quite common.146J47 Abnormally migrating ankyrin. Recently, a fast migrating ankyrin (Ankyrin Prague)74 was detected due to a yet undefined defect within the regulatory domain of ankyrin; it is dominantly coinherited with mild HS. Recent cloning of the ankyrin C D N A , ~ ~progress * J ~ ~ in characterization of the detection of a new spectrin binding site of ankyrin,5J50-152 ankyrin gene polymorphism,153as well as new advances in the detection of DNA defects are likely to open this potentially exciting field. Deficiency of 4.1 protein. Partial deficiency of the 4.1 protein, although rare in the United States, has been detected more frequently in HE subjects in Southern France and North A f r i ~ a . ~In~ a, simple ~ ~ , ~heterozygous ~~ state, it is expressed as a mild, common HE; the homozygous state is associated with a severe hemolytic anemia that is partially corrected by splenectomy.6°The molecular basis of 4.1 deficiency is heterogeneous. Detailed studies of the index case showed a deletion of 318 nucleotides that included the translation initiation site that gives rise to the major erythroid 4.1 In a recent study (N.D. Venezia and J. Delaunay, personal communication), a Spanish patient with homozygous 4.1(-) HE was found to have a point mutation in the downstream erythroid initiation codon (AUG -+ AGG), resulting in a complete absence of RBC 4.1 protein. Although the protein 4.1 gene is expressed in erythroid and nonerythroid tissues, protein 4.1 deficiency in these individuals is erythroid specific.lS6This is related to the fact that the protein 4.1 pre-mRNA is subject to extensive alternative splicing that affects the function of the protein157J58and that nonerythroid tissues express high molecular weight isoforms through the use of an upstream 5' translation initiation site that is not affected by the above noted mutations. In another kindred, protein 4.1 mRNA was lacking approximately 2 kb at the 5' end, possibly reflecting aberrant mRNA sp1i~ing.l~~ Abnormally migrating 4.1 protein. Two other protein 4.1 defects have been reported, each involving the spectrin binding domain of the protein as shown by enzymatic and chemical digestion of the 4.1 protein and amplification of appropriate regions of the CDNA.~O,~~ In an elongated form of 4.1 protein (95 Kd), the Lys 407 -+ Glu 529 sequence 318 PALEK AND SAHR spanning the spectrin binding domain is duplicated due to a Plasmodium falciparum m e r o z o i t e ~ , ~and ~ ~the J ~ Ge ~ Gerbich blood group phenotype is common in certain parts of duplication of three consecutive exons of the protein 4.1 malaria infested M e l a n e ~ i a . ' ~ ~ , ' ~ ~ gene?' In a shortened 4.1 protein, the Lys 407 + Glu 486 Protein 4.2 defects. Many case reports of patients having sequence of the spectrin binding domain is deleted as a either a reduced or absent 4.2 protein have been reresult of a deletion of two adjacent exons of the 4.1 protein gene.s1As expected, the patient with the shortened protein p ~ r t e d ,but ~ ~the - ~molecular ~ basis of the defect is unknown and the function of the 4.2 protein is not fully understood.' 4.1 had a more severe hemolytic anemia than the patient In two reports, protein 4.2 deficiency appeared to result with a high molecular 4.1 protein, presumably because the latter protein retains some if not most of its ability to bind p from a defective binding of the 4.2 protein to the band 3 p r ~ t e i n , ' ~as~evidenced J~ by a weak binding of the normal spectrin. A second shortened 4.1 variant (4.1 Presles) has been described." This defect appears to result from a 4.2 protein to inside out membrane vesicles from the preferential splicing of protein 4.1 pre-mRNA to generate a patient. In one of these cases, the underlying genetic defect relatively abundant RBC transcript lacking motif I1 (S. was reported to represent substitution of 327 Pro by Arg in Feddal and J. Delaunay, personal communication, 1992). a highly conserved region of the band 3 protein.181 In Other abnormalities of the 4.1 protein, characterized by another report, a mutation of the 4.2 protein involving abnormal mobility, have been reported,' but detailed studcodon 142 from GCT (Ala) to ACT (Thr) was detected.ls2 ies of the molecular basis of these defects are not available. However, it remains to be established whether this mutaDeficiency of glycophorin C. Deficiencies of each of the tion is coinherited with protein 4.2 deficiency, thus causing RBC membrane glycoproteins have been reported.' Gethe clinical phenotype of hemolytic anemia with spheroovalnetic abnormalities of glycophorins A and B, the genes of ocytes. which are physically linked on the q28-q31 region of Defects of the anion exchanger. While several functional chromosome 8, including deficiencies of both glycophorin A and structural defects of the anion exchanger have been and B and the formation of fusion proteins, are clinically reported, only a few have been characterized at the level of asymptomatic. primary structure and DNA. Two interesting defects have Glycophorin C deficiency is clinically expressed as a mild, been found in association with acanthocytosis. In one, recessively inherited HE with no hemolysis and mild associated with hereditary acanthocytosis without obvious e1liptocytosis.l The molecular basis of the deficiency is hemolysis, band 3 had a larger molecular mass, a restricted heterogeneous, including deletions of two exons (exons 3 rotational diffusion, and a decreased number of highand 4) and alterations leading to a frameshift and premaaffinity binding sites for ankyrin.ls3At the structural level, ture codon termination.161J62 Except for elliptocytosis, these two-dimensional peptide mapping showed changes in a subjects are clinically asymptomatic. RBC membrane vis17-Kd anion transport segment consistent with the addition coelastic properties, as measured by micropipette, are of tyrosines or tyrosine-containing peptides. A second normal,31 suggesting that the diminished deformability of defect, reported in a patient with choreacanthocytosis glycophorin C-deficient elliptocytes during shear, as desyndrome, involved structural abnormalities of the C-termitected by the ectocytometer, is likely to reflect abnormal nus of the anion exchange protein without obvious funcflow of elliptocytes in the shear stress field rather than tional alterations.1s4 However, the linkage of these two changes of intrinsic membrane mechanical proper tie^.^' abnormalities with the inheritance acanthocytosis is yet to Deficiency of glycophorin C is also accompanied by the loss be established, and the contribution of these abnormalities of the expression of all antigens of the Gerbich blood group to the acanthocytic shape is likewise unknown. (Ge:l, Ge:2 and Ge:3), the so called Leach p h e n ~ t y p e . * ~ ~ J ~A partial deficiency of the band 3 protein has recently In contrast, the loss of expression of only one or two Ge been detected in a subset of patients with dominantly inherited HS.48,49The deficiency is linked with the inheriantigens (Ge-2,+3, the Yus phenotype, and Ge-2,-3, the tance of HS. The nature of the defect remains unknown, Gerbich phenotype) is asymptomatic because these RBCs carry an abnormal glycophorin C-related glycoprotein.164-168 but RFLP linkage studies in one large family assigned the primary defect to the band 3 gene.185The synthesis of band The underlying genetic defect involves a deletion of 2 exons 3 protein has been studied in erythroblasts of one patient (exons 3 and 4) in the Leach phenotype or a deletion of one and was found normal.ls6 In several patients, separation of of the duplicated domains encoded by exon 3 (the Gerbich RBCs on density gradients showed that band 3 deficiency is phenotype) or exon 2 (the Yus p h e n ~ t y p e ) . ' ~ ~ - ' ~ ~ Several in vitro studies have suggested that both glycomore profound in the high-density Although the phorin A and glycophorin C provide an attachment point of increase in RBC density may not be directly proportional to increasing RBC age,187 the data, nevertheless, raise a the skeletal protein 4.1 to the membrane.173J74The experiments of nature described above, however, show that possibility of a release of the band 3 protein from the complete deficiency of glycophorin A in vivo has no membrane of circulating RBCs, possibly resulting from apparent effects on cell function. Likewise, complete defiweakened binding of the band 3 protein to ankyrin. This ciency of glycophorin C, although producing a mild elliptolatter defect may reside in the ankyrin binding domain of cytosis, does not lead to significant alterations of membrane the cytoplasmic segment of the band 3 protein or, conproperties?l On the other hand, both glycophorin A and versely, the band 3 binding domain of ankyrin. glycophorin C may be involved in the attachment of Several groups have recently detected abnormalities of RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS 319 ASSESSMENT OF RBC MORPHOLOGY, MEMBRANE the band 3 protein in RBCs in Southeast Asian ovalocytosis PROTEIN ANALYSIS, AND DETECTION OF THE (SAO),188-192a condition highly prevalent in malaria inUNDERLYING GEN ETlC DEFECT fected regions of Malaysia, the Philippines, and Papua New Guinea. S A 0 RBCs are rigid, resist invasion by various When we attempt to define the underlying molecular strains of malaria parasites, and exhibit reduced agglutindefect in a patient with HE, HPP, or HS, we begin with a ability to antisera against several blood group a n t i g e n ~ . l ~ ~ - lcareful ~~ assessment of RBC morphology. In a patient The underlying genetic defect involves an intra-exon delecontaining elliptocytes on the peripheral blood film, it is tion of codons 400-408 at the boundary of the cytoplasmic essential that a clinical diagnosis of HE be unequivocally segment and the first transmembrane segment of the band 3 established (1) by finding of an autosomal dominant pattern p r ~ t e i n . ~This ~ ~ mutation , ’ ~ ~ is tightly linked with a substituof inheritance in the family, and (2) by excluding one of the tion of lysine 56 by glutamic acid.189The latter mutation common causes of acquired elliptocytosis, including iron and B12 deficiencies, myelodysplasia, myofibrosis, and other alters the mobility of the cytoplasmic segment of the band 3 conditions.1~22~23 The percentage of elliptocytes or the deprotein or its proteolytic fragments and was originally gree of elliptical deformation as shown by elliptocyte length designated band 3 One group reported that the ovalocyte band 3 contains an altered N - t e r m i n ~ s . ’ ~ ~to width ratio, previously used as one of the diagnostic criteria, have no clinical use, as the underlying defect can be However, this is contrary to the recent sequencing data detected even in patients with elliptocytes not exceeding indicating that, except for the 56 Lys + Glu mutation, the 20% (normal range, 0% to 5%).’ The second step is to cDNA encoding the first 110 amino acids, including 150 bp categorize HE into one of the previously described morphoof the 5’ untranslated sequence is n01mal.l~~ Furthermore, logical subtypes1J3: (1) common HE, characterized by the sequence of the “extended” terminus192is, with the biconcave elliptocytes, rod-shaped cells, and a varying exception of one amino acid, homologous to the sequence number of poikilocytes; (2) HPP, a disorder closely related of a region of the ring-infected erythrocyte surface antigen to HE characterized by a striking micropoikilocytosis and (RESA),lWsuggesting that the sequence of the “extended microspherocytosis; (3) spherocytic HE, a phenotypic hyN terminus” is derived from a protein other than band 3. brid of HE and HS; and (4),stomatocytic elliptocytosis, also S A 0 band 3 also exhibits a markedly reduced lateral called Southeast Asian or Melanesian ovalocytosis (Fig 6). mobility in the membrane.’%Ym It is also excessively phosCommon hereditary elliptocytosis. In common HE, the ph0rylated~~~,~01; it is presently unknown whether or not this underlying defect can be detected in about 60% to 70% of abnormality is responsible for the abnormal function of the cases by using the following screening approaches (Fig 7). band 3 protein. The membrane domain of S A 0 band 3 also The first step involves a standard SDS-PAGE, which in appears to have an altered structure based on its abnormal about 10% of cases may show a partial deficiency or an binding of some anion transport inhibitors, and to have a absence of the 4.1 protein, or abnormally migrating 4.1, a reduced capacity for anion transportJo2 The mechanism spectrin, or p spectrin proteins. The second step involves an whereby this mutation leads to increased membrane rigidity electrophoretic separation of SpD and SpT from the crude and malaria resistance is currently under study. spectrin extract under nondenaturing conditions at 0°C. Involvement of nonerythroid tissues in hereditary defects of This temperature kinetically immobilizes the SpD-T equilibthe RBC membrane skeleton. Although spectrin, ankyrin, rium so that the fraction of unassembled SpD in the extract and the band 3, 4.1, and 4.2 proteins are present in many approximates the fraction of the assembled SpD in the nonerythroid tissue^,^,^^^ clinically significant symptoms inmembrane in ~ ~ v o . ~ J ~ This J O ~ fraction does not exceed 10% volving nonerythroid tissues are rare. One of the possible of the SpD + SpT pool in healthy subjects. An increase in explanations of the lack of a multisystem involvement is the the fraction of unassembled SpD reflects a defective SpD to fact that nonerythroid isoforms of spectrin and ankyrin are SpT self-association that is typically due to a mutation of a encoded by distinct genes.203However, both the erythroid f3 or f3 spectrin involving the spectrin heterodimer contact spectrin and ankyrin genes are expressed in skeletal muscle site. The next essential step involves a limited proteolysis of and some neurons as tissue-specific isoforms by alternative spectrin, followed by a separation of the resulting peptides pre-mRNA s p l i ~ i n g . 2This ~ ~ may possibly account for the by electrophoresis in two dimensions, involving isoelectric findings of mental retardation in two reports of HS associfocusing in the first dimension and SDS-PAGE in the ated with ankyrin d e f i ~ i e n c y 4 ~and J ~ ~an earlier report of a second dimension.22~u~87~89 Under carefully controlled condispinal cord disease in an HS family in which membrane tions, the tryptic peptide maps are highly reproducible, and protein studies are not available.m The importance of the 80-Kd peptide, corresponding to the a1 domain and the these clinical observations is highlighted by recent reports SpD-SpD self-association site of a spectrin, is the most that distinct isoforms of ankyrin are associated with specific prominent spot. Mutations of (Y spectrin involving the a1 structures of mouse brain and that the erythroid ankyrin domain, as well as some of the mutations of f3 spectrin, give isoforms are missing in ankyrin-deficient (nb/nb) spherorise to abnormally migrating peptides that are reactive with cytic mice.2o5One of the structures containing erythroid polyclonal antibodies raised against the normal a1 domain. ankyrin isoforms are cerebellar Purkinje cells; the progresAt the present time, this step is essential before the sive loss of these cells in ankyrin deficient (nb/nb) mice characterization of the defect at the DNA level. In most, coincides with a development of a psychomotor disorder.2M but not all cases, the site of the mutation is in the vicinity of 320 Fig 8. Abnormalities of RBC shape in inherited disorders of RBC membrane proteins. (1) Typical HS. Although spherocytes are prominent, many cells retain some degree of biconcave shape. (2) HS with pincered (mushroom shaped) cells (arrow). Plncered RBCs are found in about 80% of HS patients with band 3 deficiency, whereas they are rare in other forms of HS (D. Arnold, P. Jarolim, and J. Palek, recent unpublished data). (3) HS with acanthocytes, as reported in HS containing mutant p spectrin that binds poorly t o the 4.1 protein.u.lz* (4) Severe hemolytic HS associated with a combined deficiency of spectrin and ankyrin. In addition t o spherocytosis, many ceils have an irregular contour; some of the cells resemble RBCs in HPP, shown in (8). 15) Spherocytic elliptocytosis (hemolytic ovalocytosis). Many cells have an oval shape. These patients often have hemolytic anemia despite minimal changes in RBC morphology. (6) Common mild HE associated with Sp d I4 mutation. In addition t o elliptocytes, occasional rod-shaped ceils (arrow) are present. (7) "Homozygous" common HE in a compound heterozygote for Sp dt74and Sp d's. Note prominent elliptocytosis with numerous fragments and poikilocytes. (8) HPP Sp d'". Note prominent microspherocytosis and marked RBC fragmentation and poikilocytosis. but only occasional elliptocytes. Some poikilocytes are in the process of "budding" (snow). In addition t o the presence of spectrin mutation, these cells are also possibly deficient in s p e ~ t r i n . (9) ~ ~SA0 ~ ~ (stometocytic ~.~~ elliptocytosis). The cells often contain a longitudal slit or a transverse ridge (arrow). (Reprinted with pewnission.l) the abnormal cleavage site that gives rise to the abnormally migrating peptide. In some of the previously studied mutations, the site of cleavage has been mapped by transferring the tryptic peptides to immobilon membrane, followed by PALEK AND SAHR direct N-terminal sequencing of the peptide of interest.*Jm Occasionally, the mutation can be detected during this step."J'' More often, the mutation is subsequently found by analyzing exon or cDNA sequences encoding both the N-terminal and C-terminal sequences adjacent to the cleavage site (Tablc 1). This can be readily accomplished using the polymerase chain reaction (PCR) technique2m.mto amplify genomic DNA or cDNA. In two spectrin mutations, this step can now be bypassed. The HE Sp u116s mutation appears to be homogeneous involving a duplication of the leucine codon at position 154, thus permitting the use of appropriate allele-specificoligonucleotide (ASO) probes to verify or exclude this m u t a t i ~ n . ~In~8-of~ ~ 12' patients ~ with the spectrin u1/74mutation examined so far, the defect involves codon 28ms99'.w and eliminates an Aha 11 restriction site9'; this change can be used as diagnostic of this mutation. However, because several different changes can result in ablation of a restriction enzyme site, this type of change must be confirmed by sequence analysis or use of AS0 hybridization probes. As discussed above, in some patients with the Sp u1174 defect, the mutation may involve p spectrin. Likewise, in one of the spectrin ailMdefects, the mutation is not adjacent to the abnormal cleavage site.Ils The majority of HE patients who are simple heterozygotes for a spectrin mutation are asymptomatic, presenting either without detectable hemolysis or with a minimal hemolysis only. However, rare HE patients present with moderate to severe hemolysis. This clinical picture is typically accompanied by the presence of poikilocytes and fragments on the peripheral blood film. In some patients, this clinical syndrome is dominantly transmitted, and coinherited with a mutant spectrin (such as Sp c P 4 ) that is grossly dysfunctional, as also reflected by a marked increase in unassembled dimeric spectrin. reaching 50% of the total SpD + SpT p ~ o l . ~Other . ~ . ~rare ~ patients are either homozygous or doubly heterozygous for one or two spectrin mutations involving defective SpD self-association?* In some patients, the augmentation of hemolysis together with poikilocytosis reflects an interaction of the elliptocytogenic spectrin mutation with another factor. This other factor can be either inherited or acquired. An example of an inherited factor is a silent genetic factor, in trans, ie, linked to the normal spectrin allele as detected either by analysis of tryptic peptide maps (the Sp uv/4ipolymorphism)131 or by studies of linkage with a spectrin RFLP.*I" Examples of acquired factors are an altered intracellular environment such as found in neonates whose RBCs contain high hemoglobin F (neonatal poikilocytosis, see section on membrane fragmentation), microcirculatory damage to RBCs during disseminated intravascular coagulation or thrombotic thrombocytopenia purpura,1u certain infections, or other conditions such as superimposed BIZdeficiency.L3 The presence of a second interacting factor is suggested by the finding of a relatively mild defect of spcctrin, as reflected by a mild increase in unassembled dimeric spectrin in conjunction with a symptomatic hemolytic disorder.'-' Hereditary pyropoikilqtosis. The diagnosis of HPP is suggested by the presence of a severe recessively inherited RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS 321 1A 1 3 2 .band 3’ Fig 7. Evaluation of a membrane proteln defect In HE and HPP. (A) Identification of f3 speetrin mutant with abnormal mobility (Spectrin Nice, gel 2) and the deficiency of protein 4.1 (gel 4) in HE. Membrane proteins from control RBCs (gels 1and 3) and RBCs of the two patients (gels 2 and 4) were solubilized in SDS and electrophoresed in a Fairbanks nonlinear 3.5% t o 17% polyacrylamide gradient gel‘“ (first two gels) and Laemmli gel’.’ (last two gels). Note the excellent resolution of a spectrin, f3 spectrin, and ankyrin bands, as well as the f3 spectrin mutant (Spectrin Nice) in the modified Fairbanks system. In the Laemmli system, f3 spectrin and ankyrin bands overlap. However, the Laemmli system resolves band 4.1 into two bands (4.la and 4.lb). Both bands are absent in the HE 4.1 (gel 4). (B) Nondenaturing polyacrylamide gels of crude spectrin extracts. Lane 1 is a control, 2 is one of the two parents who both have common HE, and 3 is their homozygous HE offspring. RBCa of the HE parent, who is heterozygous for the Sp contain both dimeric (SpD) and tetrameric ( S p n Sp. In the homozygous offspring, spectrin is almost exclusively in a dimeric state. (C) Two-dimensional peptide map (isoelectric focusing followed by SDS-PAGE [IEFISDS-PAGE]) of limited tryptic digest of spectrin from a control and HE subject with the Sp d Mdefect. The positions of the normal d domain and those of the abnormal d peptides of molecular weight of 65 Kd are indicated by arrows. (Modified and reprinted with permission.’) Fairbanks 4 B 1 2 3 1 Laemmli Nondenaturing gel c 6 -1 P 65kd’- hemolytic anemia, associated with the presence of numerous poikilocytcs, microsphcrocytcs, and fragments on the periphcral blood film. The fragmcntation often leads to a decrcase in mcan corpuscular volume and a marked increase in the RBC size distribution width. Incubation of RBCs at tempcratures of 45 to 47°C shows a gross thcrmal instability in the majority of cases?’ further supporting the diagnosis of HPP. It should be pointed out, howevcr, that varying dcgrccs of thermal instability of RBCs are noted in the majority of elliptocytogenic spectrin mutations rcgardless of whether thc clinical phenotype is that of HE or HPP. Although the molccular basis of HPP is heterogcncous, the common feature of HPP is the presence of one of several spcctrin mutations involving a defective SpD selfassociation togethcr with a partial dcficicncy of spcctrin.22J’.2”’ Some patients are homozygous for the mutant spectrin and their parcnts may both have mild HEvn;others are cithcr doubly hctcrozygous for two spcctrin mutations (one of which may often present with HE in one of thc parents) or, more commonly, thcy are compound hetcrozygotcs for a spcctrin mutation charactcrizcd by dcfcctivc SpD self-association and anothcr unknown genetic defect that augments the clinical cxprcssion of the mutant spcctrin and produccs a concomitant spcctrin deficiency, such as a thalassemia-likc dcfcct in spcctrin synthesis.” Thc prcsencc of such a second genetic defect was recently verified (1) by findings of rcduccd a spcctrin synthesis in an in vitro culturc systcm (both in thc HPP offspring carrying the elliptogenic spcctrin mutation, and her fully asymptomatic father) and (2) by findings of rcduccd amounts of mRNA produccd by thc structurally normal spcctrin allclc.21i.212 In HPP patients who are cithcr homozygous or doubly het- PALEK AND SAHR 322 erozygous for one or two spectrin mutations involving a defective SpD contact, the spectrin deficiency results from an instability of spectrin leading to accelerated spectrin degradation before assembly on the membrane.w7,213In RBCs of homozygotes for a severely dysfunctional spectrin that leads to a marked increase in the fraction of unassembled dimeric spectrin, the spectrin deficiency may be related to the binding stoichiometry of one copy of ankyrin per one spectrin tetramer; thus, in patients with a marked increase of unassembled SpD, the number of ankyrin copies may not be sufficient to bind all SpD to the RBC membrar~e.',~~ SoutheastAsian ovalocytosis. In contrast to common HE and HPP that is heterogeneous in molecular terms, SA0 appears h o m o g e n e ~ u s , ~ presumably ~~J~ because its prevalence is related to a selective pressure resulting from a resistance of ovalocytes to invasion by malaria parasifes.195J96,214 In addition to characteristic RBC morphology, the presence of this defect is suggested by the finding of resistance of ovalocytes or their ghosts to changes in ~ h a p e , ' ~ ~and , * l by ~ analysis of limited proteolytic digests of whole cells or ghosts (showing the "band 3 Memphis" polymorphism, which is tightly linked with SAO),188or by PCR amplification of the region of cDNA or genomic DNA that contains the deletion of eight c o d o n ~ . ' ~ ~ J ~ ~ In our experience, the strategy outlined above detects the underlying defect in about 70% of patients with HE and in all cases of HPP and SAO. In a rare patient with a mild recessively inherited elliptocytosis, staining of membrane proteins with periodic acid (PAS) may show a lack of glycophorin C'; the heterozygous carriers of this defect are asymptomatic. Hereditary spherocytosis. The principal cellular lesion of hereditary spherocytosis is a surface area deficiency reflecting a loss of membrane lipids. In most cases, the cause of the lipid loss is a partial deficiency of spectrin.' As a result of the deficiency of spectrin, the density of the submembrane skeletal network in the HS RBC membrane is reduced, presumably underlying the lipid bilayer destabi- lization and microvesiculation.' Indeed, the degree of spectrin deficiency is proportional to the increase of osmotic fragility, a crude indicator of cell surface area deficiency, and to the loss of membrane material from the cells in vitro.",216 Consequently, clinical severity is proportional to the amounts of RBC spectrin with either a life-threatening44 or even fatal disease217in subjects with severe spectrin deficiency. In the latter report of a severe spectrin deficiency (26% of normal) with a normal content of ankyrin and band 3,217 progenitor-derived erythroblasts in culture exhibited a strikingly reduced synthesis of a spectrin and a markedly reduced assembly of a and p spectrin on the membrane. A moderate to severe deficiency of spectrin can be detected by protein separation by SDS-PAGE, with densitometric estimation of spectrin content by measuring a ratio of spectrin and band 3 peaks. It should be noted, however, that this technique is too insensitive to detect mild spectrin deficiencies. Furthermore, the loss of membrane lipids from the HS RBC membrane is accompanied by a loss of the band 3 protein, producing a mild deficiency of this protein that can spuriously normalize the spectrin to band 3 ratio.' Thus, a direct quantitation of the number of spectrin copies per cell by radioimmunoassay represents the definitive method to measure spectrin content per cell. Although varying degrees of spectrin deficiency are detected in the majority of patients with HS, it should be emphasized that the underlying molecular basis of HS is likely to be heterogeneous and that the primary molecular lesion is likely to involve several membrane proteins, including spectrin, ankynn, band 3, and band 4.2 proteins (see above). This biochemical heterogeneity of HS is evident by analysis of membrane protein composition by SDS-PAGE (Table 2). This approach shows abnormalities in about 60%of HS and segregates the disorder into at least four categories: (1) isolated spectrin deficiency; (2) a combined deficiency of spectrin and ankyrin; (3) a deficiency of the band 3 protein as reflected by a marked increase in the ratios of spectrin, actin, 4.1, and 4.2 protein Table 2. Membrane ProteinAbnormalities in HereditarySpherocytosisas Shown by SDS-PAGE Membrane Protein Abnormalities Severe ( - 50%) spectrin deficiency Mild to moderate (20% to 30%) spectrin deficiency Severe ( - 50%) deficiency of both spectrin and ankyrin Mild to moderate (20% to 30%) deficiency of spectrin and ankyrin Deficiency of band 3 protein Deficiency of protein 4.2 Inheritanceand Clinical Presentation Autosomal recessive HS with severe hemolysis Autosomal dominant HS with mild to moderate hemolysis HS with severe hemolysis, inheritance unknown Autosomal dominant HS with mild to moderate hemolysis Autosomal dominant HS with mild to moderate hemolysis Autosomal recessive hemolytic anemia with sphero-ovalocytes Underlying Molecular Defects* Prevalence References a spectrin Probably rare 41.1 19.217 p spectrin Ankyrin' Ankyrin About 10% of patients Unknown Probably rare 82.83.1 28 Ankyrin' 20% or more of kindred 219.t Band 3 protein About 15% to 20% of kindred 48,49,185,186 Band 4.2 protein* Band 3 protein* Relatively common in Japan 47.138 50-56.181 Although less sensitive than direct quantitation of the number of copies of spectrin, ankyrin, or band 3 protein per cell, SDS-PAGE followed by densitometrictracing and determination of the ratio of spectrin and ankyrinto band 3 shows abnormalitiesin about 70% of patients with HS. *Presumed primary defect, the DNA defect is likely to be heterogeneous. tJarolim P, Palek J, unpublished observations, 1992. RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS 323 to the band 3 protein; and (4) a deficiencyof the 4.2 protein. verified by a direct immunoassay and the characterization of this defect is currently in progress. However,subsequent detailed characterizationof the underThe last group of defects shown by the above approach is lying molecular defect is difficult, as there is no simple a partial or complete deficiency of the 4.2 protein. Both functional or structural test that would provide further partial and complete deficiencies have been reported, and clues in regard to the nature of the underlying molecular the hematologicphenotype is either that of HS and, in some defect. patients, ovalo-stomatocytosis.50-55 It remains to be estabIn a patient with a severe spectrin deficiency and a lished whether the 4.2 deficiency represents the primary recessively inherited HS, the primary molecular defect has defect or simply a marker of another yet unknown defect. been recently assigned to ci spectrin by protein and RFLP In a striking contrast to HE, HPP, and SAO, the linkage studies; in a group of recently reported patients, the underlying cDNA or genomic DNA defect remains unclear underlying defect involves a mutation that leads to spectrin in the majority of patients with HS. In many patients with deficiencyby a yet undefined molecular me~hanism.4~9~~~ In HE or HPP, the approximate site of the genetic defect dominantly inherited HS, isolated spectrin deficiency is could be deduced from studies of protein self-association likely to be heterogeneous. As noted above, a subset of (ie, involving SpD-SpD contact) or from limited proteolytic patients may have an abnormal p spectrin that binds poorly peptide maps of spectrin. In HS, a similar strategy has been to the 4.1 pr0tein.82~~ It is yet to be determined whether a useful in characterizing one defect involving a weakened p subset of patients with spectrin deficiency carries a defect spectrin-protein 4.1 c0ntact.8~,*~ In the majority of HS involving a mutant ankyrin, which.has a reduced ability to patients, however, the assignment of the tentative site of bind spectrin to the membrane. Another possible defect the mutation to a specific region of the protein has not been may involve spectrin that binds weakly to ankyrin; one such made. For several reasons this area of investigation will defect has been noted in an asymptomatic parent of a likely open up in the near future. Firstly, the cDNA patient manifesting with HPP,Z18 but the nature of the sequences of human a n k ~ r i n , ' ~ciJ ~spectrin,m ~ specdefect remains unclear. trin,221protein 4.1,2u protein 4.2,2',224 and band 3,2253226 and In patients with combined spectrin and ankyrin deficienthe gene organization of human ci ~ p e c t r i n land ~ , ~mouse ~~ cies, the defect underlying a deficiency of ankyrin has been band 3228have been recently described. Secondly, several detected in several p a t i e n t ~ ! ~ J ~In ~ Jone ~ ~ report, the entire rapid and sensitive techniques have been developed to ankyrin gene was found absent due to an interstitial detect mutations in cDNA and genomic DNA. These deletion within a short arm of chromosome 8.139In another scanning techniquesu9 include chemical cleavage of mispatient with severe spectrin and ankyrin deficiency, the matched base pairs,m RNase protection,B1 denaturing mRNA levels and the synthesis of ankyrin in reticulocytes gradient gel electrophoresis (DGGE),U2-u5and singlewere reduced.138Spectrin was synthesized normally but the stranded conformational polymorphism analysis assembly of spectrin on the membrane was reduced be(SSCP).u6,u7These methods can be used in conjunction cause of the underlying defect involving the synthesis of with the PCR technique to detect small deletions or ankyrin. However, in the majority of patients with domiinsertions and single base substitutions. If by one of these nantly inherited HS, the nature of combined spectrin and methods the amplified DNA appears to be abnormal, the ankyrin deficiencies remains unknown. The detection of the specific change can then be determined by subsequent underlying defects is of considerable interest as this biosequence analysis. Although neither RNase protection, chemical phenotype is very common, found in up to DGGE, or SSCP techniques have been applied to the two-thirds of all patients219(Table 2). It is possible, but analysis of gene defects in HE or HS, their potential has presently not proven, that some of these subjects could have been illustrated in experiments to analyze mutations of the a thalassemia-like defect of ankyrin synthesis. type I collagen gene by RNase protection238and mutations Analysis of membrane skeleton proteins by SDS-PAGE in p-thalassemia by DGGE.B9 has recently shown a unique subset of patients with domiACKNOWLEDGMENT nantly inherited HS with high ratios of spectrin, ankyrin, 4.1, and actin to the band 3 protein, implying a partial The authors are indebted to Gabriella Maitino and Loretta deficiency of the band 3 pr0tein.4~This deficiency has been Wencis for typing the manuscript and to Joan Joss for the art work. REFERENCES 1. Palek J, Lambert S: Genetics of the red cell membrane Blood Cell Membranes: Structure, Function, Clinical Implications skeleton. Semin Hematol27:290,1990 (vol 11). New York, NY, Dekker, 1989, p 77 2. Conboy JG, Mohandas N Characterization of the gene 4. Winkelmann JC, Let0 TL, Forget BG: Spectrin genes, in coding for human erythrocyte protein 4.1: Implications for underAgre P, Parker JC (eds): Red Blood Cell Membranes: Structure, standing hereditary elliptocytosis, in Agre P, Parker JC (eds): Red Function, Clinical Implications (vol 11). New York, NY, Dekker, Blood Cell Membranes: Structure, Function, Clinical Implications 1989, p 111 (vol 11). New York, NY,Dekker, 1989, p 167 5. Delaunay J, Alloisio N, Morle L, Pothier B: The red cell 3. Marchesi S L The erythrocyte cytoskeleton in hereditary skeleton and its genetic disorders. Mol Aspects Med 11:161,1990 6. Gallagher PG, Tse WT, Forget BG: Clinical and molecular elliptocytosis and spherocytosis, in Agre P, Parker JC (eds): Red 324 aspects of disorders of the erythrocyte membrane skeleton. Semin Perinatoll4351,1990 7. Agre P Partial deficiencies of erythrocyte spectrin in hereditary spherocytosis, in Agre P, Parker JC (eds): Red Blood Cell Membranes: Structure, Functions, Clinical Implications (vol 11). New York, NY,Dekker, 1989, p 707 8. Dhermy D: The spectrin super-family. Biol Cell 71:249, 1991 9. Liu SC, Derick LH, Palek J: Visualization of the hexagonal lattice in the erythrocyte membrane skeleton. J Cell Biol 104:527, 1987 10. Bennett V: The spectrin-actin junction of erythrocyte membrane skeletons. Biochim Biophys Acta 988107,1989 11. Bennett V The molecular basis for membrane-cytoskeleton association in human erythrocytes. J Cell Biochem 18:49,1982 12. Liu SC, Derick LH, Palek J: Molecular anatomy of erythrocyte membrane skeleton in health and disease, in Cohen CM, Palek J (eds): Cellular and Molecular Biology of Normal and Abnormal Erythroid Membranes, UCLA Symposia on Cellular and Molecular Biology, New Series (~01118).New York, NY,Liss, 1990, p 171 13. Liu SC, Derick LH, Duquette MA, Palek J: Separation of the lipid bilayer from the membrane skeleton during discocyteechinocyte transformation of human erythrocyte ghosts. Eur J Cell Biol49:358,1989 14. Bitbol M, Dempsey C, Watts A, Devaux PF: Weak interaction of spectrin with phosphatidylcholine-phosphatidylserine multilayers: A 2H and 31PNMR study. FEBS Lett 244:217,1989 15. Cohen AM, Liu SC, Lawler J, Derick LH, Palek J: Identification of the protein 4.1 binding site to phosphatidylserine vesicles. Biochemistry 27:614, 1988 16. Cohen AM, Liu SC, Derick LH, Palek J: Ultrastructural studies of the interaction of spectrin with phosphatidylserine liposomes. Blood 68:920,1986 17. Maksymiw R, Sui SF, Gaub H, Sackmann E: Electrostatic coupling of spectrin dimers to phosphatidylserine containing lipid lamellae. Biochemistry 26:2983, 1987 18. Rybicki AC, Heath R, Lubin B, Schwartz RS: Human erythrocyte protein 4.1 is a phosphatidylserine binding protein. J Clin Invest 81:255,1988 19. Sutera SP, Mehrjardi M, Mohandas N: Deformation of erythrocytes under shear. Blood Cells 1:369,1975 20. Liu SC, Derick LH, Agre P, Palek J: Alteration of the erythrocyte membrane skeletal ultrastructure in hereditary spherocytosis, hereditary elliptocytosis and pyropoikilocytosis. Blood 76:198,1990 21. Jarolim P, Palek J, Coetzer TL, Lawler J, Velez-Garcia E, Fradera J, Charles A, Jacob H, Moake J: Severe hemolysis and red cell fragmentation due to the combination of a spectrin mutation with a thrombotic microangiopathy. Am J Hematol3250,1989 22. Palek J: Hereditary elliptocytosis, spherocytosis and related disorder: Consequences of a deficiency or a mutation of membrane skeletal proteins. Blood Rev 1:147,1987 23. Palek J: Hereditary elliptocytosis and related disorders. Clin Haematol14:45,1985 24. Mentzer WC Jr, Iarocci TA, Mohandas N: Modulation of erythrocyte membrane mechanical fragility by 2,3-diphosphoglycerate in the neonatal poikilocytosis/elliptocytosissyndrome. J Clin Invest 79:943,1987 25. Sheetz MP, Casaly J: 2,3-Diphosphoglycerate and ATP dissociate erythrocyte membrane skeletons. J Biol Chem 255:9955, 1980 26. Wolfe LC, Lux SE, Ohanian V: Regulation of spectrin-actin binding by protein 4.1 and polyphosphates. J Cell Biol 87203a, 1980 (suppl) 27. Liu SC, Derick LH, Zhai S, Palek J: Ultrastructural anatomy of the red cell membrane lesion in sickle cells: Penetration of the PALEK AND SAHR hemoglobin S polymers through the membrane skeleton and the reorganization of the skeletal lattice. Blood 74:44a, 1989 (abstr, SUPPI 1) 28. Liu SC, Derick LH, Palek J: Permanent deformation of normal erythrocyte membrane is abolished by inhibition of spectrin tetramer-dimer transformation. Blood 76:12a, 1990 (abstr, suppl 1) 29. Bessis M: Living Blood Cells and Their Ultrastructure. New York, NY,Springer-Verlag, 1973, p 140 30. Mohandas N, Chasis JA, Shohet SB: The influence of membrane skeleton on red cell deformability membrane material properties and shape. Semin Hematol20225,1983 31. Nash GB, Parmar J, Reid M E Effects of deficiencies of glycophorins C and D on the physical properties of the red cell. Br J Haematol76282,1990 32. Hanspal M, Hanspal J, Kalraiya R, Howard D, Gao X, Palek J: Asynchronous synthesis and assembly of membrane skeletal proteins during terminal differentiation of Friend virus infected murine erythroblasts. Blood 769a, 1990 (abstr, suppl 1) 33. Koury MJ, Bondurant MC, Rana SS: Changes in erythroid membrane proteins during erythropoietin-mediated terminal differentiation. J Cell Physiol133:438,1987 34. Lazarides E: From genes to structural morphogenesis: The genesis and epigenesis of a red blood cell. Cell 51:345,1987 35. Cox JV,Stack JH, Lazarides E Erythroid anion transporter assembly is mediated by a developmentally regulated recruitment onto a preassembled membrane cytoskeleton. J Cell Biol105:1405, 1987 36. Woods CM, Boyer B, Vogt PK, Lazarides E: Control of erythroid differentiation: Asynchronous expression of the anion transporter and the peripheral components of the membrane skeleton in AEV- and S13-transformed cells. J Cell Biol 103:1789, 1986 37. Lazarides E, Moon R T Assembly and topogenesis of the spectrin-based membrane skeleton in erythroid development. Cell 37:354,1984 38. Moon RT, Lazarides E: Biogenesis of the avian erythroid membrane skeleton: Receptor-mediated assembly and stabilization of ankyrin (goblin) and spectrin. J Cell Biol98:1899,1984 39. Blikstad I, Nelson WJ, Moon RT, Lazarides E: Synthesis and assembly of spectrin during avian erythropoiesis: Stoichiometric assembly but unequal synthesis of (Y and p spectrin. Cell 321081,1983 40. Hanspal M, Palek J: Synthesis and assembly of membrane skeletal proteins in mammalian red cell precursors. J Cell Biol 105:1417,1987 41. Marchesi SL, Agre PA, Speicher DW, Tse WT, Forget BG: Mutant spectrin a11 domain in recessively inherited spherocytosis. Blood 74:182a, 1989 (abstr, suppl 1) 42. Furie B, Furie B C Molecular basis of hemophilia. Semin Hematol27:270,1990 43. Tse WT, Lecomte MC, Costa FF, Garbarz M, Fto C, Boivin P, Dhermy D, Forget BG: Point mutation in the B-spectrin gene associated with hereditary elliptocytosis. J Clin Invest 86:909, 1990 44. Agre P, Orringer EP, Bennett V: Deficient red-cell spectrin in severe, recessively inherited spherocytosis. N Engl J Med 306:1155,1982 45. Agre P, Asimos A, Casella JF, McMillan C Inheritance pattern and clinical response to splenectomy as a reflection of erythrocyte spectrin deficiency in hereditary spherocytosis. N Engl J Med 315:1579,1986 46. Agre P, Casella JF, Zinkham WH, McMillan C, Bennett V: Partial deficiency of erythrocyte spectrin in hereditary spherocytosis. Nature 314:380, 1985 RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS 47. Coetzer TL, Lawler J, Liu SC, Prchal JT, Gualtieri R, Brain MC, Dacie JV, Palek J: Partial ankyrin and spectrin deficiency in severe, atypical hereditary spherocytosis. N Engl J Med 318:230, 1988 48. Lux S, Bedrosian C, Shalev 0,Morris M, Chasis J, Davies K, Sawides P, Telen M: Deficiency of band 3 in dominant hereditary spherocytosis with normal spectrin content. Clin Res 38:300A, 1990 (abstr) 49. Jarolim P, Ruff P, Coetzer TL, Prchal JT, Ballas SK, Poon M-C, Brabec V, Palek J: A subset of patients with dominantly inherited hereditary spherocytosis has a marked deficiency of the band 3 protein. Blood 76:37a, 1990 (abstr, suppl 1) 50. Burke BE, Shotton DM: Erythrocyte membrane skeleton abnormalities in hereditary spherocytosis. Br J Haematol 54:173, 1983 51. Hayashi S, Koomoto R, Yano A, Ishigami S, Tsujino G, Saeki S, Tanaka T Abnormality in a specific protein of the erythrocyte membrane in hereditary spherocytosis. Biochim Biophys Res Commun 57:1038,1974 52. Nozawa Y, Hoguchi T, Fukushima H, Sekiya T, It0 Y Erythrocyte membrane of hereditary spherocytosis: Alterations in surface ultrastructure and membrane proteins as inferred by scanning electron microscopy and SDS disc gel electrophoresis. Clin Chim Acta 55231,1974 53. Rybicki AC, Heath R, Wolf JL, Lubin B, Schwartz RS: Deficiency of protein 4.2 in erythrocytes from a patient with a Coombs negative hemolytic anemia. Evidence for a role of protein 4.2 in stabilizing ankyrin on the membrane. J Clin Invest 81:893, 1988 54. Yawata Y, Ata K, Kanzaki A, Wada H, Ikoma K, Higo I, Yamada 0: Membrane characteristics of membrane protein band 4.2 deficiency with congenital hemolytic anemia. Blood 76:21a, 1990 (abstr, suppl 1) 55. Ideguchi H, Nishimura J, Nawata H, Hamasaki N: A genetic defect of erythrocyte band 4.2 protein associated with hereditary spherocytosis. Br J Haematol74:347,1990 56. Ghanem A, Pothier B, Marechal J, Ducleuzeau T, Morle L, Alloisio N, Feo C, Abdeladhim AB, Fattoum S, Delaunay J: A haemolytic syndrome associated with the complete absence of red cell membrane protein 4.2 in two Tunisian siblings. Br J Haematol 75:414,1990 57. Alloisio N, Mor16 L, DorlCac E, Gentilhomme 0, Bachir D, Guetarni D, Colonna P, Bost M, Zouaoui Z, Roda L The heterozygous form of 4.1 (-) hereditary elliptocytosis [the 4.1 (-) trait]. Blood 65:46, 1985 58. Alloisio N, Dorleac E, Girot R, Delauney J: Analysis of the red cell membrane in a family with hereditary elliptocytosis-totalor partial absence of protein 4.1. Hum Genet 59:68,1981 59. Dhermy D, Garbarz M, Lecomte MC, FCo C, Bournier 0, Chaveroche I, Gautero H, Galand C, Boivin P: Hereditary elliptocytosis: Clinical, morphological and biochemical studies of 38 cases. Nouv Rev Fr Hematol28:129,1986 60. Fko CJ,Fischer S, Piau JP, Grange MJ, Tchernia G: Premier observation de I’absence d’une proteine de la membrane erythrocytaire (bande 41) dans un cas d’anemie elliptocytaire familiale. Nouv Rev Fr Hematol22:315,1980 61. Lambert S, Zail S: Partial deficiency of protein 4.1 in hereditary elliptocytosis.Am J Hematol26:263,1987 62. McGuire M, Smith BL, Agre P Distinct variants of erythrocyte protein 4.1 inherited in linkage with elliptocytosisand Rh type in three white families. Blood 72287, 1988 63. Tchernia G, Mohandas N, Shohet SB: Deficiency of skeletal membrane protein band in homozygous hereditary elliptocytosis: Implications for erythrocyte membrane stability. J Clin Invest 68:454,1981 325 64. Mueller TJ, Morrison M: Cytoskeletal alterations in hereditary elliptocytosis.J Supramolec Struct 5:131,1981 65. Dhermy D, Lecomte MC, Garbarz M, Fto C, Galand C, Bournier 0, Gautero H, Boivin P A new kindred of hereditary elliptocytosis (HE) with a shortened spectrin (Y chain. Blood 7052a, 1987 (abstr, suppl 1) 66. Lane PA, Shew RL, Iarocci TA, Mohandas N, Hays T, Mentzer W C Unique (Y spectrin mutant in a kindred with common hereditary elliptocytosis.J Clin Invest 79:989,1987 67. Lecomte MC, Lahary A, Vannier JP, Dhermy D, Boivin P: Spectrin Rouen (P220-218). A new shortened spectrin (Sp) P chain variant in a kindred of hereditary elliptocytosis (HE). Xe CongrCs SOCFranc HCmat, Rouen, France, 1989 68. Ohanian V, Evans JP, Gratzer WB: A case of elliptocytosis associated with a truncated spectrin chain. Br J Haematol 61:31, 1985 69. Pothier B, MorlC L, Alloisio N, Ducluzeau MT, Caldani C, F t o C, Garbarz M, Chaveroche I, Dhermy D, Lecomte MC, Boivin P, Delaunay J: Spectrin Nice (P 220/216): A shortened p chain variant associated with an increase of the fragment in a case of elliptocytosis.Blood 691759,1987 70. Dhermy D, Lecomte MC, Garbarz M, Bournier 0, Garland C, Gautero H, Feo C, Alloisio N, Delaunay J, Boivin P: Spectrin P-chain variant associated with hereditary elliptocytosis. J Clin Invest 70:707,1982 71. Eber SW, Morris SA, Schroter W, Gratzer WB: Interactions of spectrin in hereditary elliptocytes containing truncated spectrin P-chains.J Clin Invest 81:523,1988 72. Yawata Y, Kanzaki A, Wada H, Ata K, Inone T, Akatsuka J, Delaunay J A trait of beta spectrin variant HE (SpB220/216) with hereditary elliptocytosis.Blood 76:22a, 1990 (abstr, suppl 1) 73. Garbarz M, Feldman L, Boulanger L, Lecomte MC, Gautero H, Galand C, Boivin P, Dhermy D: Molecular basis of spectrin Tandil, a shortened p spectrin variant associated with hereditary elliptocytosis.Blood 78:84a, 1991(abstr, suppl 1) 74. Jarolim P, Brabec V, Lambert S, Liu SC, Zhou Z, Palek J: Ankyrin Prague: A dominantly inherited mutation of the regulatory domain of ankyrin associated with hereditary spherocytosis. Blood 76:37a, 1990 (abstr, suppl 1) 75. Conboy J, Kan YW, Agre P, Mohandas N: Molecular characterization of hereditary elliptocytosis due to an elongated protein 4.1. Blood 68:34a, 1986 (abstr, suppl 1) 76. Garbarz M, Dhermy D, Lecomte MC, FCo C, Chavaroche I, Galand C, Bournier 0, Bertrand 0, Boivin P: A variant of erythrocyte membrane skeletal protein band 4.1 associated with hereditary elliptocytosis.Blood 641006,1984 77. Letsinger JT, Agre P, Marchesi S L High molecular weight protein 4.1 in the cytoskeletons of hereditary elliptocytes. Blood 68:38a, 1986 (abstr, suppl 1) 78. Marchesi SL, Letsinger JT, Agre PA, McGuire M, Speicher D W Characterization of high and low molecular weight variants of protein 4.1 associated with elliptocytosis. Blood 72:31a, 1988 (abstr, suppl 1) 79. Mor16 L, Garbarz M, Alloisio N, Girot R, Chaveroche I, Boivin P, Delaunay J The characterization of protein 4.1 Presles, a shortened variant of RBC membrane protein 4.1. Blood 65:1511, 1985 80. Marchesi SL, Conboy J, Agre P, Letsinger JT, Marchesi VT, Speicher DW, Mohandas N Molecular analysis of insertion/ deletion mutations in protein 4.1 elliptocytosis. I. Biochemical identification of rearrangements in the spectrinlactin binding and functional characterizations. J Clin Invest 86516, 1990 81. Conboy J, Marchesi S, Kim R, Agre P, Kan YW, Mohandas N Molecular analysis of insertion/deletion mutations in protein PALEK AND SAHR 326 98. Coetzer T, Palek J, Lawler J, Liu SC, Jarolim P, Lahav M, 4.1 in elliptocytosis.11. Determination of molecular genetic origins Prchal JT, Wang W, Alter BP, Schewitz G, Mankad V, Gallanello of rearrangements. J Clin Invest 86524,1990 R, Cao A Structural and functional heterogeneity of a spectrin 82. Wove LC, John KM, Falcone JC, Byrne AM, Lux SE: A mutations involving the spectrin heterodimer self-association site: genetic defect in the binding of protein 4.1 to spectrin in a kindred Relationshipsto hematologic expression of homozygous hereditary with hereditary spherocytosis.N Engl J Med 307:1367,1982 elliptocytosis and hereditary pyropoikilocytosis. Blood 752235, 83. Goodman SR, Shiffer KA, Casoria LA, Eyster ME: Identifi1990 cation of the molecular defect in the erythrocyte membrane 99. Mor16 L, R o w AF, Alloisio N, Pothier B, Starck J, Denoroy skeleton of some kindreds with hereditary spherocytosis. Blood L, Mor16 F, Rudigoz RC, Forget BG, Delaunay J, Godet J: Two 60:772,1982 elliptocytogenic~ $ variants 1 ~ ~of the spectrin a I domain spectrin 84. Agre P, Orringer EP, Chui DHK, Bennett V: A molecular Culoz (GGT [61d] d G?T, a I 40 Gly + Val) and spectrin Lyon defect in two families with hemolytic poikilocytic anemia. Reduc(CTT + 'ITT, a I43 Leu + Phe). J Clin Invest 86:548,1990 tion of high affinity membrane binding sites for ankyrin. J Clin 100. Liu SC, Windisch P, Kim S, Palek J: Oligomeric states of Invest 68:1566,1981 spectrin in normal erythrocyte membranes: Biochemical and elec85. Marchesi S L The erythrocyte cytoskeleton in hereditary tron microscopic studies. Cell 37:587,1984 elliptocytosis and spherocytosis, in Agre P, Parker JC (eds): Red 101. Ungewickell E, Gratzer W Self-association of human Blood Cell Membranes: Structure, Function, Clinical Implications. spectrin. A thermodynamic and kinetic study. Eur J Biochem New York, NY,Dekker, 1989,p 77 88:379,1987 86. Palek J, Lux S: Red cell membrane skeletal defects in 102. Lecomte MC, Garbarz M, Grandchamp B, F6o C, Gautero hereditary and acquired hemolytic anemias. Semin Hematol 2 0 H, Devaux I, Bournier 0, Galand C, d'Auriol L, Galibert F, Sahr 189,1983 KE, Forget BG, Boivin P, Dhermy D: Sp A mutation of the a 87. Speicher DW, Morrow JS, Knowles WJ, Marchesi VT: A I spectrin domain in a white kindred with HE and HPP phenostructural model of human erythrocyte spectrin. Alignment of types. Blood 741126,1989 chemical and functional domains. J Biol Chem 257:9093,1982 103. Mor16 L, Mor16 F, Roux AF, Godet J, Forget BG, Denoroy 88. Speicher DW, Morrow JS, Knowles WJ, Marchesi VT: L, Garbarz M, Dhermy D, Kastally R, Delaunay J Spectrin Tunis Identification of proteolytically resistant domains of human eryth(Sp an elliptocytogenicvariant, is due to the CGG + TGG rocyte spectrin. Proc Natl Acad Sci USA 775673,1980 codon change (Arg + Trp) at position 35 of the a I domain. Blood 89. Morrow JS, Speicher DW, Knowles WJ, Hsu J, Marchesi 74828,1989 V T Identification of functional domains of human erythrocyte 104. Marchesi SL, Letsinger JT, Speicher DW, Marchesi VT, spectrin. Proc Natl Acad Sci USA 77:6592,1980 Agre P, Hyun B, Gulati G: Mutant forms of spectrin a-subunits in 90. Garbarz M, Lecomte MC, FCo C, Devaux I, Picat C, hereditary elliptocytosis.J Clin Invest 80191,1987 Lefebvre C, Galibert F, Gautero H, Bournier 0,Galand C, Forget 105. Mor16 L, Row A-F, Mor16 R, Alloisio N, Pothier B, Sahr BG, Boivin P, Dhermy D: Hereditary pyropoikilocytosis and KE, Forget BG, Godet J, Delaunay J: The diversity of hereditary elliptocytosis in a white French family with the spectrin a1/74 elliptocytosis in North Africa: Protein aspects and molecular variant related to a CGT to CAT codon change (Arg to His) at genetics, in Cohen CM, Palek J (eds): Cellular and Molecular position 22 of the spectrin a1 domain. Blood 75:1691,1990 Biology of Normal and Abnormal Erythroid Membranes. UCLA 91. Mor16 L, R o w A-F, Alloisio N, Pothier B, Starck J, Denoroy Symposium on Molecular and Cellular Biology, New Series 118. J, Mor16 F, Rudigoz R-C, Forget BG, Delaunay J, Godet J: Two New York, NY,Liss, 1990,p 223 elliptocytogenic~ $ variants 1 ~ ~of the spectrin a I domain. Spectrin 106. Roux AF, Mor16 F, Guetarni D, Colonna P, Sahr K, Forget Culoz and spectrin Lyon, mutated at position 40 (GGT G?T; BG, Delaunay J, Godet J: Molecular basis of Sp allfi hereditary LEU -+PHE), GLY -+ VAL) and at position 43 (CTT + elliptocytosisin North Africa: Insertion of a 'ITG triplet between respectively.Blood 74:60a, 1989 (abstr, suppl 1) codons 147 and 149 in the a-spectrin gene from five unrelated 92. Speicher DW, Davis G, Marchesi VT: Structure of human families. Blood 73:2196,1989 erythrocyte spectrin. 11. The sequence of the a-I domain. J Biol 107. Sahr KE, Tobe T, Scarpa A, Laughinghouse K, Marchesi Chem 258:14938,1983 SL, Agre P, Linnenbach AJ, Marchesi VT, Forget BG: Sequence 93. Coetzer T, Sahr K, Prchal J, Blacklock H, Peterson L, Koler and exon-intron organization of the DNA encoding the a1 domain R, Doyle J, Manaster J, Forget B, Palek J: Four different mutations of human spectrin: Application to the study of mutations causing in codon 28 of a spectrin are associated with structurally and hereditary elliptocytosis.J Clin Invest 841243,1989 functionally abnormal spectrin in hereditary elliptocytosis. J 108. Lecomte MC, Dhermy D, Gautero H, Bournier 0, Galand Clin Invest 88:743,1991 C, Boivin P: L'elliptocytose hereditaire en Afrique de I'Ouest: 94. Floyd PB, Gallagher PG, Valentino LA, Davis M, Marchesi Frequence et repartition des variants de la spectrine. C R Acad Sci SL, Forget BG: Heterogeneity of the molecular basis of hereditary [III] 306:43, 1988 pyropoikilocytosis and hereditary elliptocytosis associated with 109. Facer CA: Malaria, hereditary elliptocytosis and pyincreased levels of the spectrin aI/74-kilodalton tryptic peptide. ropoikilocytosis. Lancet 1:8643,1989 Blood 781364,1991 110. Schulman S, Roth EF Jr, Cheng B, Rybicki AC, Sussman 95. Coetzer T, Lawler J, Jarolim P, Sahr K, Forget BG, Palek J 11, Wong M, Wang W, Ranney HM, Nagel RL, Schwartz RS: Molecular heterogeneity of a spectrin mutants in hereditary Growth of plasmodium falciparum in human erythrocytescontainelliptocytosis/pyropoikilocytosis.J Cell Biochem 13B:212,1989 ing abnormal membrane proteins. Proc Natl Acad Sci USA 96. Lecomte MC, Gautero H, Garbarz M, Boivin P, Dhermy D: 87:7339,1990 Abnormal tryptic peptide from the spectrin a- or P-chain muta111. Boivin P, Lecomte MC, Garbarz M, F6o C, Boumier 0, tions: Two genetically distinct forms of the Sp a1/74 variant. Br J Devaux I, Galand C, Gautero H, Dhermy D: Clinical expression of Haematol76:406,1990 spectrin a I variants, in Cohen CM, Palek J (eds): Cellular and Molecular Biology of Normal and Abnormal Erythroid Mem97. Pothier B, Alloisio N, Mor16 L, Markcha1 J, Barth6lemy H, branes. UCLA Symposia on Molecular and Cellular Biology, New Ducluzeau MT, Dorier A, Delaunay J: Two distinct variants of Series 118.New York, NY,Liss, 1990, p 235 erythrocyte p IV domain. Hum Genet 83:373,1989 - RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS 112. Gallagher PG, Marchesi SL, Forget BG: A new point mutation associated with aI/SOb kDa hereditary elliptocytosis (HE) and hereditary pyropoikilocytosis(HPP). Clin Res 39313a, 1991 113. Lawler J, Liu SC, Palek J: Molecular defect of spectrin a subgroup of patients with hereditary elliptocytosis: Alterations in the a subunit domain involved in spectrin self-association. J Clin Invest 73:1688,1984 114. Lawler J, Palek J, Liu SC, Prchal J, Butler WM: Molecular heterogeneity of hereditary pyropoikilocytosis: Identification of a second variant of the spectrin a-subunit. Blood 62:1182,1983 115. Gallagher PG, Tse WT, Coetzer T, Zarkowsky HS, Lemmte MC, Garbarz M, Baruchel A, Ballas SK, Dhermy D, Palek J, Forget BG: A common type of the spectrin a1 46-50a kDa peptide abnormality in hereditary elliptocytosis and pyropoikilocytosis is associated with a mutation distant from the proteolytic cleavage site. Blood (in press) 116. Baklouti F, Marechal J, Wilmotte R, Alloisio N, Morle L, Ducluzeau MT, Denoroy L, Mrad A, Ben Aribia MH, Kastally R, Delaunay J: Elliptocytogenica1/36 spectrin Sfax lacks 9 amino acids in helix 3 of repeat 4. Evidence for the activation of a cryptic 5’-splice site in exon 8 of spectrin a-gene. Blood (in press) 117. Lawler J, Coetzer TL, Mankad VN,Moore RB, Prchal JT, Palek J Spectrin a1/61: A new structural variant of a spectrin in a double heterozygous form of hereditary pyropoikilocytosis. Blood 72:1412,1988 118. Lambert S, Zail S: A new variant of the a subunit of spectrin in hereditary elliptocytosis.Blood 69:473,1987 119. Marchesi SL, Agre PA, Speicher D W Abnormal spectrin a I1 domain in recessive spherocytosis. J Cell Biochem 213, 1989 (suppl13B) 120. Alloisio N, Baklouti F, Morle L, Wilmott R, Marechal J, Ducluzeau M-T, Denoroy L, Forget BG, Kastally R, Delaunay J: a11/31 Spectrin Jendouba: An elliptocytogenic a11 domain variant associated with the dI 112 ASP -+ GLU (GAC --f GAA) mutation. Blood 76:23a, 1990 (abstr, suppl 1) 121. Lecomte MC, FCo C, Gautero H, Bournier 0, Galand C, Garbarz M, Boivin P, Dhermy D: Severe recessive poikilocytic anaemia with a new spectrin a chain variant. Br J Haematol74:497, 1990 122. Alloisio N, Mor16 L, Pothier B, Row AF, Markcha], Ducluzeau T, Benhadji-Zouaoui Z, Delaunay J: Spectrin Oran ( C X I I / ~ ~ ) , a new spectrin variant concerning the a11 domain and causing severe elliptocytosis in the homozygous state. Blood 71:1039,1988 123. Gallagher PG, Tse WT, Costa F, Scarpa A, Boivin P, Delaunay J, Forget BG: A splice site mutation of the p-spectrin gene causing exon skipping in hereditary elliptocytosis associated with a truncated p-spectrin chain. J Biol Chem 266:23,1991 124. Garban M, Tse WT, Gallagher PG, Picat C, Lecomte MC, a Galibert F, Dhermy D, Forget BG: Spectrin Rouen (pzzo/z18), novel shortened P-chain variant in a kindred with hereditary elliptocytosis.J Clin Invest 88:76,1991 125. Yoon SH, Yu H, Eber S, Prchal JT: Molecular defect of truncated p-spectrin associated with hereditary elliptocytosis. P-Spectrin Gottingen. J Biol Chem 266:8490,1991 126. Tse WT, Gallagher PG, Pothier B, Costa FF, Scarpa A, Delaunay J, Forget BG: An insertional frameshift mutation of the p-spectrin gene associated with elliptocytosis in spectrin Nice (p2M/216).Blood 78517, 1991 127. Cohen CM: The molecular organization of the red cell membrane skeleton. Semin Hematol20:141,1983 128. Becker PS, Tse WT, Lux SE, Forget BG: Identification of the molecular defect of p spectrin in autosomal dominant heredi- 327 tary spherocytosis (HS) associated with defective binding of protein 4.1, HS (Sp-4.1). Blood 76:25a, 19W (abstr, suppl 1) 129. Becker PS, Lux SE: Hereditary spherocytosis and related disorders. Clin Haematol14:15,1985 130. Becker PS, Morrow JS, Lux SE: Abnormal oxidant sensitivity and p-chain structure of spectrin in hereditary spherocytosis associated with defective spectrin-protein 4.1 binding. J Clin Invest 8037,1987 131. Alloisio N, Mor16 L, MarCchal J, Foux AF, Ducluzeau MT, Guetarni D, Pothier B, Baklouti F, Ghanem A, Kastally R, Delaunay J: Sp aVI4l: A common spectrin polymorphism at the alv-avdomain junction. Relevance to the expression level of hereditary elliptocytosisdue to a-spectrin variants located in trans. Blood 76:3a, 1990 (abstr, suppl 1) 132. Knowles WJ, Bologna ML, Chasis JA, Marchesi SL, Marchesi VT:Common structural polymorphismin human erythrocyte spectrin. J Clin Invest 73:973,1984 133. Gallagher PG, Kotula L, DiPaulo B, Curtis P, Speicher D, Forget B G Polymorphismsof the a11 domain of human a spectrin and of the 3’ untranslated region of a spectrin mRNA. Blood 78:364a, 1991 (abstr, suppl 1) 134. Hoffman N, Stanislovitis P, Watkins PC, Linnenbach AJ, Forget BG: Three RFLP’s are detected by an alpha spectrin genomic clone. Nucleic Acids Res 15:4696,1987 135. Kotula L, Curtis P: XbuZ polymorphismsin the 5’ region of human erythrocyte alpha spectrin gene (Spna). Nucleic Acids Res 16:10950,1988 136. Tang TK, KO YL, Lien WP: A SfuZ RFLP in the human beta-spectrin gene (SPTB). Nucleic Acids Res 19:2515, 1991 137. Bodine DM IV, Birkenmeier CS, Barker JE: Spectrin deficient inherited hemolytic anemias in the mouse: Characterization by spectrin synthesis and mRNA activity in reticulocytes. Cell 37:721,1984 138. Hanspal M, Yoon SH, Yu H, Hanspal JS, Palek J, Prchal J T Molecular basis of spectrin and ankyrin deficiencies in severe hereditary spherocytosis: Evidence implicating a primary defect of ankyrin. Blood 77:165,1991 139. Lux SE, Tse WT, Menninger JC, John KM, Harris P, Shalev 0, Chilcote RR, Marchesi SL, Watkins PC, Bennett V, McIntosh S, Collins FS, Francke U, Ward DC, Forget BG: Hereditary spherocytosis associated with deletion of human erythrocyte ankyrin gene on chromosome 8. Nature 345:736,1990 140. Bass EB, Smith SW, Stevenson RE, Rose W F Further evidence of the location of the spherocytosis gene on chromosome 8. Ann Intern Med 99:192,1983 141. Chilcote RR, LeBeau MM, Dampier C, Pergament E, Verlinsky Y, Mohandas N, Frischer H, Roley JD: Association of red cell spherocytosis with deletion of the short arm of chromosome 8. Blood 69156,1987 142. Kimberling WJ, Fulbeck T, Dixon L, Lubs HA: Localization of spherocytosis to chromosome 8 or 12 and report of a family with spherocytosis and a reciprocal translocation. Am J Hum Genet 27586, 1975 143. Kimberling WJ, Taylor RA, Chapman RG, Lubs HA: Linkage and gene localization of hereditary spherocytosis (HS). Blood 52859,1978 144. Cohen H, Walker H, Delhanty JDA, Lucus SB, Huehns ER: Congenital spherocytosis, B19 parvovirus infection and inherited interstitial deletion of the short arm of chromosome 8. Br J Haematol78:251,1991 145. Costa FF, Agre P, Watkins PC, Winkelmann JC, Tang TK, John KM, Lux SE, Forget BG: Linkage of dominant hereditary spherocytosis to the gene for the erythrocyte membrane skeleton protein ankyrin. N Engl J Med 323:1046, 1990 PALEK AND SAHR 146. Iolascon A, DelGiudice EM, Camaschella C, Pinto L, Nobili B, Perrotta S, Cutillo S: Ankyrin deficiency in dominant hereditary spherocytosis: Report of three cases. Br J Haematol 78:551,1991 147. Sawides P, Shalev 0, John KM, Lux S E Combined spectrin and ankyrin deficiency is common in autosomal dominant hereditary spherocytosis.Clin Res 39:313A, 1991 148. Lambert S, Yu H, Prchal JT, Lawler J, Ruff P, Speicher D, Cheung MC, Kan YW, Palek J: The cDNA sequence for human erythrocyte ankyrin. Proc Natl Acad Sci USA 871730,1990 149. Lux SE, John KM, Bennett V: Analysis of cDNA for human erythrocyte ankyrin indicates a repeated structure with homology to tissue-differentiation and cell-cycle control proteins. Nature 34436,1990 150. Weaver DC, Pasternack GR, Marchesi V T The structural basis of ankyrin function. 11. Identification of two functional domains. J Biol Chem 259:6170, 1984 151. Davis LH, Bennett V Mapping the binding sites of human erythrocyte ankyrin for the anion exchanger and spectrin. J Biol Chem 265:10589,1990 152. Platt OS, Falcone JF, Lux S E A 12 kD tryptic peptide of ankyrin retains spectrin binding capacity. Blood 74:104a, 1989 (abstr, suppl 1) 153. Polymeropoulos MH, Rath DS, Xiao H, Merril CR: Dinucleotide repeat polymorphism at the human ankyrin gene (ANK1). Nucleic Acids Res 19969,1991 154. Conboy J, Kim R, Agre P, Tchernia G, Kan YW, Mohandas N: Novel alterations in erythroid protein 4.1 gene expression in patients with hereditary elliptocytosis.Blood 74:181a, 1989 (abstr, SUPPl1) 155. Conboy J, Mohandas N, Tchernia G, Kan YW: Molecular basis of hereditary elliptocytosis due to protein 4.1 deficiency. N Engl J Med 315:680,1986 156. Conboy JG, Chasis JA, Winardi R, Tchernia G, Kan YW, Mohandas N: Tissue-specific deficiency of protein 4.1 as a consequence of usage of an alternative translation initiation site. Blood 78:365a, 1991 (abstr, suppl 1) 157. Benz EJ Jr, Tang TK, Baklouti F, Huang H, Cho J, Marchesi V T Tissue specific selection of alternatively spliced exons of the protein 4.1 gene generates multiple isoforms with altered spectrin actin binding domains. Blood 78:365a, 1991 (abstr, SUPPI 1) 158. Discher D, Parra M, Conboy J, Mohandas N Functional characterization of alternatively spliced isoforms of the spectrinactin binding domain of protein 4.1. Blood 78:83a, 1991 (abstr, SUPPI 1) 159. Feddal S, Brunet G, Roda L, Chabanis S, Alloisio N, Mor16 L, Ducleuzeau M-T, Marechal J, Robert J-M, Benz EJ, Delaunay J, Baklouti F: Molecular analysis of a variety of 4.1 (-) hereditary elliptocytosis found in the French province of Savoy. Blood (in press) 160. Mor16 L, Garbarz M, Alloisio N, Girot N, Chaveroche I, Boivin P, Delaunay J: The characterizationof protein 4.1 Presles, a shortened variant of RBC membrane protein 4.1. Blood 65:1511, 1985 161. Telen MJ, Le Van Kim C, Chung A, Cartron J-P, Colin Y: Varying molecular basis of elliptocytosis due to glycophorin C deficiency. Blood 76:18a, 1990 (abstr, suppl 1) 162. Telen MJ, Le Van Kim C, Chung A, Cartron JP, Colin Y: Molecular basis for elliptocytosis associated with glycophorin C and D deficiency in the Leach phenotype. Blood 78:1603,1991 163. Anstee DJ, Parsons SF, Ridgwell K, Tanner MJ, Merry AH, Thomson EE, Judson PA, Johnson P, Bates S, Fraser ID: Two individuals with elliptocytic red cells apparently lack three minor erythrocyte membrane sialoglycoproteins.Biochem J 218:615,1984 164. Daniels GL, Shaw MA, Judson PA, Reid ME, Anstee DJ, Colpitts P, Comwall S, Moore BP, Lee S: A family demonstrating inheritance of the Leach phenotype: A Gerbich-negative phenotype associated with elliptocytosis.Vox Sang 50:117,1986 165. Le Van Kim C, Colin Y, Blanchard D, Dahr W, London J, Cartron JP: Gerbich blood group of the Ge:-1,-2,-3 types immunochemical study and genomic analysis with cDNA probes. Eur J Biochem 165:571,1987 166. Reid ME, Anstee DJ, Tanner MJA, Ridgwell K, Nurse G T Structural relationshipsbetween human erythrocyte sialoglycoproteins f3 and gamma and abnormal sialoglycoproteins found in certain rare human erythrocyte variants lacking the Gerbich blood-group antigen(s). Biochem J 244323,1987 167. Reid ME, Chasis JA, Mohandas N: Identification of a functional role for human erythrocyte sialoglycoproteins p and gamma. Blood 69:1068,1987 168. Reid ME, Anstee DJ, Jensen RH, Mohandas N Normal membrane function of abnormal B-related erythrocyte sialoglycoproteins. Br J Haematol67467,1987 169. Colin Y, Le Van Kim C, Tsapis A, Clerget M, d’Auriol L, London J, Galibert F, Cartron JP: Human erythrocyte glycophorin C. Gene structure and rearrangement in genetic variants. J Biol Chem 2643773,1989 170. High S, Tanner MJA, MacDonald EB, Anstee DJ: Rearrangementsof the red cell membrane glycophorin C (sialoglycoprotein f3) gene. A further study of alterations in the glycophorin C gene. Biochem J 26247,1989 171. Chang S, Reid ME, Conboy J, Kan YW, Mohandas N Molecular characterization of erythrocyte glycophorin C variants. Blood 77544,1991 172. Anstee DJ: Blood group-active surface molecules of the human red blood cell. Vox Sang 58:1,1990 173. Anderson RA, Lovrien RE: Glycophorin is linked by band 4.1 protein to the human erythrocyte membrane skeleton. Nature 307:655,1984 174. Mueller T, Morrison M: Glycoconnectin (PAS 2) a membrane attachment site for the human erythrocyte cytoskeleton. Prog Clin Biol Res 56:95,1981 175. Pasvol G: Receptors on red cells for Plasmodium falciparum and their interaction with merozoites. Phil Trans R SOC 133:189,1984 176. Pasvol G, Anstee DJ, Tanner MJA. Glycophorin C and the invasion of red cells by Plasmodium falciparum.Lancet 1:907,1984 177. Booth PB, Homabrook RW: The inheritance of the Gerbich blood groups: A n analysis of New Guinea families. Hum Biol Oceania 2:27,1973 178. Booth PB, McLoughlin K The Gerbich blood group system, especiallyin Melanesians. Vox Sang 2273,1972 179. Jarolim P, Korsgren C, Cohen CM, Coetzer TI.,, Prchal JT, Palek J: Molecular basis of partial deficiency of protein 4.2 in congenital spherocytic hemolytic anemia: A defect involving the cytoplasmic domain of the anion transport protein. Blood 74137a, 1989 (abstr, suppl 1) 180. Rybicki AC, Musto S, Nagel RL, Schwartz RS: Red cell protein 4.2 deficiency associated with a defect in the cytoplasmic domain of band 3. Blood 78:81a, 1991 (abstr, suppl 1) 181. Jarolim P, Palek J, Rubin HL, Prchal JT, Korsgren C, Cohen CM: Band 3 Tuscaloosa: PRO327+ ARC3” substitution in the cytoplasmic domain of erythrocyte band 3 protein associated with spherocytichemolytic anemia and partial deficiency of protein 4.2. Blood 78:252a, 1991 (abstr, suppl 1) 182. Schwartz RS, Yawata Y, Ata K, Nagel RL, Rybicki A C An alaanine to threonine substitution in protein 4.2 cDNA is associated with a japanese form of hereditary hemolyticanemia. Clin Res 39313a, 1991 RED BLOOD CELL MEMBRANE PROTEIN MUTATIONS 183. Kay MMB, Bosman GJCGM, Lawrence C Functional topography of band 3: Specific structural alteration linked to functional aberrations in human erythrocytes. Proc Natl Acad Sci USA 85:492,1988 184. Kay MMB, Bosman G, Lawrence C Neurologic disease with acanthocytosis is associated with erythrocyte membrane transport and band 3 abnormalities.Clin Res 37:547A, 1989 185. Prchal JT, Guan Y, Jarolim P, Palek J, Showe L, Bertoli L: Hereditary spherocytosis in a large family is linked with the band 3 gene and not with a-spectrin, f3-spectrin or ankyrin. Blood 78:81a, 1991 (abstr, suppl 1) 186. Saad STO, Liu SC, Golan D, Corbett JB, Thatte HS, Derick L, Hanspal M, Jarolim P, Fibach E, Palek J Mechanism underlying band 3 deficiency in a subset of patients with hereditary spherocytosis (HS). Blood 78:81a, 1991(abstr, suppl 1) 187. Dale GL, Norenberg S L Density fractionation of erythrocytes by Percoll/hypaque results in only a slight enrichment for aged cells. Biochim Biophys Acta 1036:183,1990 188. Liu SC, Zhai S, Palek J, Golan D, Amato D, Hassan K, Nurse G, Babona D, Coetzer T, Jarolim P, Zaik M, Borwein S: Molecular defect of the band 3 protein in Southeast Asian ovalocytosis. N Engl J Med 323:1530,1990 189. Jarolim P, Palek J, Amato D, Hassan K, Sapak P, Nurse GT, Rubin HL, Zhai S, Sahr KE, Liu S C Deletion in erythrocyte band 3 gene in malaria-resistant Southeast Asian ovalocytosis. Proc Natl Acad Sci USA 88:11022,1991 190. Mohandas N, Winardi R, Knowles D, Leung A, Parra M, George E, Conboy J, Chasis J: Molecular basis for membrane rigidity of hereditary ovalocytosis-A novel mechanism involving the cytoplasmicdomain of band 3. J Clin Invest 89:686,1992 191. Tanner MJA, Bruce L, Martin PG, Rearden DM, Jones G L Melanesian ovalocytes have a deletion in red cell band 3. Blood 78:2785,1991 192. Jones GL, Edmundson HM, Wesche D, Saul A Human erythrocyte band-3 has an altered N-terminus in malaria-resistant Melanesian ovalocytosis. Biochim Biophys Acta 1096:33, 1990 193. Mohandas N, Clark MR, Health BP, Rossi M, Wolfe L, Lux S, Shohet SB: A technique to detect reduced mechanical stability of red cell membrane: Relevance to elliptocytic disorders. Blood 59:768,1982 194. Kidson C, Lamont G, Saul A, Nurse G T Ovalocytic erythrocytes from Melanesians are resistant to invasion by malaria parasite in culture. Proc Natl Acad Sci USA 785829,1981 195. Hadley T, Saul A, Lamont G, Hudson DE, Miller LH, Kidson C: Resistance of melanesian elliptocytes (ovalocytes) to invasion by plasmodium knowlesi and plasmodium falciparum malaria parasites in vitro. J Clin Invest 71:780,1983 196. Booth PB, Serjeantson S, Woodfield DG, Amato D: Selective depression of blood group antigens associated with hereditary ovalocytosis among Melanesians. Vox Sang 32:99,1977 197. Jarolim P, Rubin HL, Mueller TJ, Palek J: Band 3 Memphis: Abnormality migrating erythrocyte band 3 protein due to the substitution AAG + GAG (Lys -+ Glu) in codon 56. Blood 78:82a, 1991 (abstr, suppl 1) 198. Yannoukakos D, Vasseur C, Driancourt C, Blouquit Y, Delaunay J, Wajcman H, Bursaux E: Human erythrocyte band 3 polymorphism (band 3 Memphis): Characterization of the structural modification (Lys -+ Glu) by protein chemistry methods. Blood 78:1117,1991 199. Husain-Chishti A, Ruff P: Malaria and ovalocytosisMolecular mimicry? Biochim Biophys Acta 1096:263,1991 200. Chasis JA, Knowles D, Winardi R, George E, Mohandas N: Conformational changes in cytoplasmic domains of band 3 and glycophorin-Aaffect red cell membrane properties. Blood 78:252a, 1991 (abstr, suppl 1) 329 201. Chishti AH, Andrabi K, Palek J, Amato D, Liu SC: Altered tyrosine phosphorylation of the red cell band 3 protein in malaria resistant southeast asian ovalocytosis (SAO). Blood 78:80a, 1991 (abstr, suppl 1) 202. SchofieldAE, Reardon DM, Tanner MJA Defective anion transport activity of the abnormal band 3 in hereditary ovalocytic red blood cells. Nature 355836,1992 203. Bennett V, Lambert S: The spectrin skeleton: From red cells to brain. J Clin Invest 87:1483,1991 204. McCann SR, Jacobs HS: Spinal cord disease in hereditary spherocytosis: Support of two cases with a hypothesized common mechanism for neurologic and red cell abnormalities. Science 48959,1976 205. Kordeli E, Bennett V: Distinct ankyrin isoforms at a neuron cell bodies and nodes of Ranvier resolved using erythrocyte ankyrin-deficientmice. J Cell Biolll41243,1991 206. Peters LL, Birkenmeier CS, Bronson RT, White RA, Lux SE, Otto E, Bennett V, Higgins A, Barker JE: Purkinje cell degeneration associated with erythroid ankyrin deficiency in nb/nb mice. J Cell Biol114:1233,1991 207. Coetzer TL, Palek J: Partial spectrin deficiency in hereditary pyropoikilocytosis. Blood 67:919, 1986 208. Mullis KB, Faloona F A Specific synthesis of DNA in vitro via a polymerase catalyzed chain reaction. Methods Enzymol 155:335,1987 209. Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, Horn GT, Mullis KT3, Erlich HA: Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239:487,1988 210. Guetarni D, R o w AF, Alloisio N, Mor16 F, Ducluzeau MT, Forget BG, Colonna P, Delaunay J, Godet J Evidence that expression of Sp hereditary elliptocytosis is compounded by a genetic factor that is linked to the homologous a-spectrin allele. Hum Genet 85627,1990 211. Gallagher PG, Tse WT, Marchesi SL, Zarkowsky HS, Forget BG: A defect in a-spectrin mRNA accumulation in hereditary pyropoikilocytosis(HPP). Clin Res 39:313a, 1991 212. Hanspal M, Hanspal J, Fibach E, Palek J: a Spectrin syntheticdefect underlies spectrin deficiency in a subset of patients with hereditary pyropoikilocytosis (HPP). Blood 78:84a, 1991 (abstr, suppl 1) 213. Hanspal M, Hanspal J, Palek J: Molecular basis of spectrin deficiency in hereditary pyropoikilocytosis(HPP) associated with mutant spectrin a'/&.Blood 72:43a, 1988 (abstr, suppl 1) 214. Cattani JA, Gibson FD, Alpers P, Crane GG: Hereditary ovalocytosis and reduced susceptibility to malaria in Papua New Guinea. Trans R SOCTrop Med Hyg 81:705,1987 215. Castelino D, Saul A, Myler P, Kidson C, Thomas H, Cooke R: Ovalocytosis in Papua New Guinea-Dominantly inherited resistance to malaria. Southeast Asian J Trop Med Public Health 12549, 1981 216. Chasis JA, Agre P, Mohandas N Decreased membrane mechanical stability and in vivo loss of surface area reflect spectrin deficiencies in hereditary spherocytosis. J Clin Invest 82617, 1988 217. Whitfield CF, Follweiler JB, Lopresti-Morrow L, Miller B A Deficiency of a-spectrin synthesis in burst-forming unitserythroid in lethal hereditary spherocytosis. Blood 78:11,1991 218. Zail SS, Coetzer T L Defective binding of spectrin to ankyrin in a kindred with recessively inherited hereditary elliptocytosis. J Clin Invest 74:753,1984 219. Shalev 0, John KM, Lux SE: Combined spectrin and ankyrin deficiency is common in autosomal dominant hereditary spherocytosis.Clin Res 39:313a, 1991 220. Sahr KE, Laurila P, Kotula L, Scarpa AL, Coupal E, Let0 TL, Linnenbach AJ, Winkelmann JC, Speicher DW, Marchesi VT, 330 Curtis PJ, Forget BG: The complete cDNA and polypeptide sequences of human erythroid a-spectrin. J Biol Chem 265:4434, 1990 221. Winkelmann JC, Chang JC, Tse WT, Marchesi VT,Forget BG: Full length sequence of the cDNA for human erythroid p spectrin. J Biol Chem 265:11827,1990 222. Conboy J, Kan YW, Shohet SB, Mohandas N Molecular cloning of protein 4.1, a major structural element of the human erythroid membrane skeleton. Proc Natl Acad Sci USA 83:9512, 1986 223. Korsgren C, Lawler J, Lambert S, Speicher D, Cohen CM: Complete amino acid sequence and homologies of human erythrocyte membrane protein band 4.2. Proc Natl Acad Sci USA 87:613, 1990 224. Sung LA, Chien S, Change LS, Lambert K, Bliss S, Bouhassira E, Nagle R, Schwarz R, Rybicki A. Molecular cloning of human protein 4.2: A major component of the erythrocyte membrane. Proc Natl Acad Sci USA 87955,1990 225. Tanner MJA, Martin PJ, High S: The complete amino acid sequence of the human erythrocyte membrane anion-transport protein deduced from the cDNA sequence. Biochem J 256:703, 1988 226. Lux SE, John KM, Kopito RR, Lodish HF: Cloning and characterization of band 3, the human erythrocyte anion exchange protein (AE1). Proc Natl Acad Sci USA 86:9089,1989 227. Kotula L, Laury-Kleintop LD, Showe L, Sahr K, Linnenbach AJ, Forget BG, Curtis PJ: The exon-intron organization of the human erythrocyte a-spectrin gene. Genomics 9:131,1991 228. Kopito RR, Andersson M, Lodish H F Structure and organization of the murine band 3 gene. J Biol Chem 2628035, 1987 229. Rossiter BJF, Caskey CI?Molecular scanning methods of mutation detection. J Biol Chem 265:12753,1990 230. Cotton RGH, Rodrigues NR, Campbell RD: Reactivity of cytosine and thymine in single-base-pairmismatcheswith hydroxylamine and osmium tetroxide and its application to the study of mutations. Proc Natl Acad Sci USA 854397,1988 PALEK AND SAHR 231. Myers RM, Larin Z, Maniatis T Detection of single base substitutionby ribonuclease cleavage at mismatches in RNADNA duplexes. Science 2301242,1985 232. Fischer SG, Lerman LS: DNA differing by a single basepair substitution are separated in denaturing gradient gels. Correspondence with meltin theory. Proc Natl Acad Sci USA 801579, 1983 233. Myers RM, Fischer SG, Maniatis T, Lerman LS: Modification of the melting properties of duplex DNA by attachment of a GC-rich DNA sequence as determined by denaturing gradient gel electrophoresis.Nucleic Acids Res 13:3111,1985 234. Sheffield Val C, Cox DR, Lerman LS, Myers RM: Attachment of a G + C-rich sequence (GC-clamp) to genomic DNA fragments by the polymerase chain reaction resulting in improved detection of single-base changes. Proc Natl Acad Sci USA 86:232, 1989 235. Myers RM, Fischer SG, Lerman LS, Maniatis T Nearly all single base substitutions in DNA fragmentsjoined to a GC-clamp can be detected by denaturing gradient gel electrophoresis.Nucleic Acids Res 13:3131,1985 236. Orita M, Iwahana H, Kanazawa H, Hayashi K, Sekiya T Detection of polymorphisms of human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc Natl Acad Sci USA 862766,1989 237. Orita M, Suzuki Y, Sekiya T, Hayashi K Rapid and sensitive detection of point mutations and DNA polymorphisms using the polymerase chain reaction. Genomics 5874,1989 238. Grange DK, Gottesman GS, Lewis MB, Marini J C Detection of point mutations in type I collagen by RNase digestion of RNA/RNA hybrids. Nucleic Acids Res 184227,1990 239. Cai S-P, Kan YW: Identification of the multiple P-thalassemia mutations by denaturing gradient gel electrophoresis. J Clin Invest 85:550,1990 240. Fairbanks G, Steck TL, Wallach D F Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry 102606,1971 241. Laemmli U K Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227680,1970

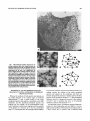
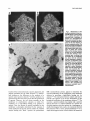










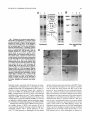










![A second allele of spectrin alpha-gene associated with the alpha... phenotype (allele alpha Ponte de Sor) [letter]](http://s1.studyres.com/store/data/007858793_1-cec18df0f8a6bd226cd9c76efa32daa1-150x150.png)









