Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
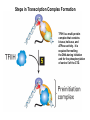
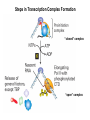
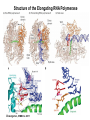
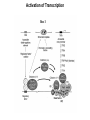
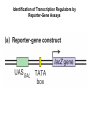
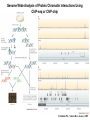
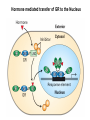
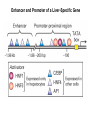
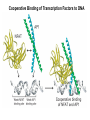
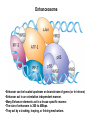
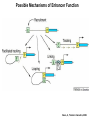
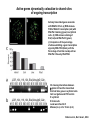
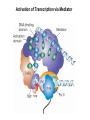
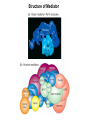
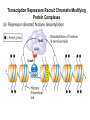
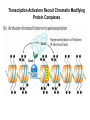
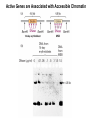
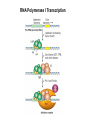
Transcription in Eukaryotes I and II Jörg Bungert, PhD 352-273-8098 [email protected] Objectives • • • • • • • • • • Know the basal promoter elements and the basal transcription factors. Know the RNA polymerase II CTD phosphorylation cycle. Understand reporter-gene assays. Know and understand enhancer function. Understand Chromosome Conformation Capture (3C). Know the structure and function of the mediator. Know histone modifications and how they impact gene expression. Know the different chromatin modifying activities (HAT, HDAC, HMT, chromatin remodeling complexes). Know what a DNAseI hypersensitive (HS) site is. Know RNA polymerase I and III transcription complex formation. Reading: Lodish 6th edition, chapter 7 (pp. 276-317). Transcription in Eukaryotes - Overview Eukaryotic RNA Polymerases 1. Pol I - Transcribes rRNA genes (28S, 18S, 5.8S RNA genes); accounts for 80-90% of total cellular RNA mass 2. Pol II – Transcribes protein-encoding mRNA’s, and several non-protein encoding genes; consists of 12 subunits that are conserved among diverse eukaryotic organisms 3. Pol III - Transcribes tRNA, 5S RNA, 7S RNA (component of signal recognition particle genes, and other small RNA-encoding genes (including genes encoding RNA’s involved in splicing; tRNA and 5S genes have internal promoter (i.e., within body of gene) 4. Each RNA Pol can be distinguished by differences in sensitivity to α-amanitin 5. Each RNA Pol has multiple subunits, is similar from yeast to mammals, and is more complex than the E. coli RNA polymerase 6. Some subunits of each RNA Pol share some sequence similarity to E. coli RNA pol, and all three eukaryotic RNA Polymerases share come common subunits 7. The C-terminal end of the largest subunit of pol II subunit contains a repeated 7 a.a. motif 1. This repeated region is termed the C-terminal domain (CTD) 2. Active transcription is correlated with phosphorylation of CTD 3. CTD is associated with several RNA processing factors 8. All three RNA Pols use TBP (TATA binding protein) in assembly of pre-initiation complex 9. New report (2005) of previously unknown mitochondrial RNA pol (spRNAP-IV) transcribing some nuclear mRNA-encoding genes with promoters different from those using pol II Subunit Structure of RNA Polymerases The TATA-Box Basal Promoter Elements in Higher Eukaryotes Housekeeping, constitutively expressed, genes are transcribed from multiple start sites. Tissue-specific and regulated genes are transcribed from a single start site. The mechanism of dispersed transcription initiation is not well characterized. Focused transcription initiation takes place at genes harboring specific basal promoter elements (e.g. TATA, INR, DPE). TATA and DPE are present in about 15 to 20% of genes transcribed by Pol II, the INR is found in about 50% of the genes. Juven-Gershon and Tjian, Developmental Biology, 2009 RNA Polymerase II Transcription The pre-initiation complex (PIC): PIC assembles at the basal promoter region of a gene PIC consists of RNA pol II, 5 general transcription factors (GTF’s), and promoter DNA GTF’s = TFII D, TFIIB, TFIIF, TFIIE, TFIIH (and TFIIA?) (e.g., HeLa cells) Identified by in vitro transcription assay (Run Off) using as a DNA template the adenovirus major late promoter (Ad2MLP). E-Box TATA-INR-DPE Steps in Transcription Complex Formation IIA TBP binds to the minor groove and bends the DNA. Bending may facilitate melting of the DNA or assembly of the transcription complex. The binding of TBP is facilitated by TFIIA Steps in Transcription Complex Formation TFIIB bridges the TBP-TATA box complex with the RNA polymerase. Steps in Transcription Complex Formation TFIIF interacts with the RNA Polymerase and is required for preventing unspecific initiation at non-promoter sites as well as for efficient transcription elongation. Steps in Transcription Complex Formation TFIIE stimulates the helicase and ATPase activities of TFIIH Steps in Transcription Complex Formation TFIIH is a multi-protein complex that contains kinase, helicase, and ATPase activity. It is required for melting the DNA during initiation and for the phosphorylation of serine 5 of the CTD. Steps in Transcription Complex Formation “closed” complex “open” complex TBP/TATA box complex and TFIID TFII D composed of:: •TBP – TATA-Binding Protein •TAF’s – TBP-associated factors Also, evidence for: - Cell type-specific TBP’s - TAF complexes w/out TBP that can functionally replace TFIID Structure of the Preinitiation Complex TBP/TFIIA/TATA-Box TBP/TFIIA/TFIIB/TATA-Box Structure of the Elongating RNA Polymerase Cheung et al., EMBO J. 2011 Phosphorylation of RNA Pol II CTD Correlates with Gene Activation CTD Heptapeptide: Thr Ser Pro Thr Ser Pro Ser (yeast, 26X) Tyr Ser Pro Thr Ser Pro Ser (mammals 52X) Red: Antibody for phosphorylated CTD Green: Antibody for unphosphorylated CTD 74/EF/75b: ecdysone induced high level transcription in “puff” regions. Transcription Elongation Factors Transcriptional pause is a frequent event during elongation caused by slight misalignment of the RNA 3’OH and the active site. It is self-reversible and regulated by numerous cellular factors. Transcriptionally arrested complexes, in which the RNA 3’OH and active site are irreversibly misaligned, resume elongation after Pol II mediated transcript cleavage and realignment of the active site and the RNA 3’OH end, processes that are stimulated by TFIIS. Modification of RNA Pol II CTD During Transcription Elongation CTD phosphorylation cycle: TFIIH subunit Kin 28 phosphorylates Ser 5. Transcription elongation complex (TEC) is arrested at checkpoint for pre-mRNA capping. P-TEFb (Bur1/2 or Ctk1 in yeast) then phosphorylates Ser 2, which allows further elongation. Specific phosphatases (possibly Ssu72 and FCP1, which are required for Pol II Recycling) dephosphorylate the CTD at Ser 5 and Ser2. Activation of Transcription Tissue-specific Gene Expression Simple gene expression array analysis. cDNAs are spotted onto nitrocellulose or nylon membranes and hybridized to radio-labeled RNA from liver, kidney, or brain. For identifying tissue-specific DNA control elements the genomic regions of the liver-specific genes can be isolated from genomic libraries. The upstream promoter regions can then be ligated in front of reporter genes and analyzed for tissuespecific expression. Identification of Regulatory DNA Elements by Progressive Deletions and Reporter Gene Assays Assume TTR is a liver specific gene identified in the previous experiment. The upstream regulatory region of this gene was cloned in front of a reporter gene (Reporter Gene: e.g. LacZ, luciferase, GFP). The reporter gene constructs were transiently transfected into a liver cell line. The data demonstrate that a tissue-specific cis-regulatory DNA element, required for activated expression of the reporter gene is located between the 5’end of fragment 2 and the 5’end of fragment 3. In addition, a basal promoter element is located between the 5’end of fragment 4 and the 5’ end of fragment 5. The regulatory DNA element that is located between the 5’end of fragment 2 and the 5’end of fragment 3 can be further analyzed for the presence of transcription factor binding sites. A. B. C. D. In silico analysis for known factors using databases. Electrophoretic Mobility Shift Assays (EMSA). DNA Footprinting. Chromatin Immunoprecipitation. Modular Structure of Transcription factors -Plus: Nuclear Localization Sequences (NLS), Protein/Protein Interaction Domains, Sites of Modification. Identification of Transcription Regulators by Reporter-Gene Assays The previous experiment identified a cis-regulatory element of the TTR gene that mediates high-level expression in a liver cell line. To identify a transcription factor that binds to the cis-regulatory element (X) and activates transcription, cotransfection experiments can be performed. Plasmid 1 carries the gene encoding the transcription factor of interest, while plasmid 2 carries regulatory sequence X linked to a reporter gene. A control experiment would be performed with plasmid 2 lacking sequence X. Identification of Transcription Regulators by Reporter-Gene Assays Identification of DNA binding and Transcription Regulation Domains Genome Wide Analysis of Protein Chromatin Interactions Using ChIP-seq or ChIP-chip Farnham P.J., Nature Rev. Genet., 2009 Hormone mediated transfer of GR to the Nucleus Enhancer and Promoter of a Liver-Specific Gene Cooperative Binding of Transcription Factors to DNA Enhanceosome •Enhancer can be located upstream or downstream of genes (or in introns) •Enhancer act in an orientation independent manner. •Many Enhancer elements act in a tissue-specific manner. •The size of enhancers is 200 to 400bps. •They act by a tracking, looping, or linking mechanism. Activation of the stripe 2 enhancer by a gradient of transcription factors in the developing Drosophila embryo The drosophila eve gene is regulated by many enhancer elements that confer Expression of the eve gene in specific stripes of the developing embryo. The stripe 2 enhancer is composed of binding sites for activating proteins (Bicoid: Bcd, Hunchback, Hb) and repressing proteins (Kreisler, Kr, and Giant, Gnt). During early embryonic development, a gradient of transcription factors is set up in the multi-nucleated embryo (syncytium). At this stage, the concentration of bicoid and hunchback is high in stripe 2, whereas the concentration of kreisler and giant is low. Ptashne and Gann, 2002 Possible Mechanisms of Enhancer Function Dean, A., Trends in Genetics, 2006 Transcription and Transcription Factories Model of dynamic associations of genes with transcription factories. Chromatin loops (black) extruding from chromosome territories (gray). Transcribed genes (white) in RNA Pol II factories (black circles). Potentiated genes (free loops) that are not associated with Pol II factories are temporarily not transcribed and can migrate to a limited number of preassembled Pol II factories (dotted arrows) Szentimayr and Sawadogo, Nucl. Acids Res.. 2002 Osborne et al., Nat. Genet., 2004 Chromosome conformation capture Dekker, Nat. Methods 2006 Active genes dynamically colocalize to shared sites of ongoing transcription Actively transcribed genes associate with RNA Pol II foci. (a) RNA immunoFISH of Hbb-b1 transcription (red) with RNA Pol II staining (green) in erythroid cells. (b) DNA immuno staining of Eraf (red) and RNA Poll II (green). (c) Comparison of the percentage of alleles exhibiting a gene transcription signal by RNA FISH (black), with the Percentage of loci that overlap with an RNA Pol II focus by DNA FISH. 3C showing interactions between b -globin LCR and the transcribed Eraf and Uros genes in erythroid cells. Calr is a ligation and PCR control. E: erythroid B: brain cells nuclei were fixed for 5 Minutes (in a) or for 10 min (in b) Osborne et al., Nat. Genet., 2004 Transcription Factories Model of a transcription factory (diameter 70 nm) containing 8 polymerases (green crescents). Genes are reeled through these factories (one polymerase per gene). Heat shock gene activation and formation of a transcription factory that facilitates reinitiation. Sutherland, H., and W.A. Bickmore, Nat. Rev. Genet., 2009 Activation of Transcription via Mediator Structure of Mediator Transcription Repressors Recruit Chromatin Modifying Protein Complexes Silencing of Chromatin Domains by Polycomb Repressor Complexes The PRC complex mediates the silencing of genomic loci. It is recruited to specific sites in chromatin by the PRC responsive elements (PREs) and proteins that interact with PREs. There are two PRC complexes. PRC2 methylates H3K27 and represses gene expression. PRC1 recognizes methylated H3K27 and methylates CpGs. This leads to a stably repressed chromatin configuration that can be transmitted to daughter cells after mitosis. Transcription Activators Recruit Chromatin Modifying Protein Complexes Histone Modifications In Euchromatin and Heterochromatin “Histone Code” A histone mark associated with sites of Pol II recruitment is trimethylated H3K4. This modification has been shown to assist in the recruitment of TFIID. Dimethylated H3K4 is associated with active chromatin domains H3K27 and H3K9 trimethylation is associated with repressed chromatin. H3K36 methylation is associated with transcription elongation. Histone modifications Histone acetylation • Occurs on lysine side chains of N-terminal tails of histones • Changes charge of histone tails – makes them less basic (neutralizes positive charge) • May weaken histone-DNA interactions and “open” the nucleosome • May alter histone-histone interactions • May alter interactions between histones and regulatory proteins • May facilitate binding of regulatory proteins to cis-acting elements in DNA • Effects may be on individual nucleosomes and/or higher order chromatin structure • Makes histone tails more -helical • Catalyzed by histone acetyltransferases (HAT’s) • Reversible by histone deacetylases (HDAC’s) Histone methylation • Does not alter net charge of histone tails • Mono-, di-, tri-methylation of lysine side chain • Catalyzed by histone methyltransferases (HMT’s) • Methylation of histone tails thought to be stable • Histone demethylase isolated in 2005 – LSD1, lysine-specific demethylase, removes a methyl group from one particular residue (H3, K4), probably others ATP-Dependent Chromatin Remodeling Complexes Steps in Gene Regulation Genes are normally embedded in inaccessible chromatin. The first step in the activation of gene expression is often the recruitment of chromatin remodeling complexes (Swi/SNF) by pioneer transcription factors that recognize their respective binding site in the context of nucleosomes or higher order structure. In the example shown here, the yeast DNA binding factor SWI5 recruits the SWI/SNF complex to its binding site in chromatin. In an ATP hydrolysis dependent manner Swi/SNF mobilizes nucleosomes, rendering a short segment of chromatin accessible (nuclease sensitive). Subsequently, SWI5 and other DNA binding proteins recruit histone acetyltransferases (HATs) like yeast GCN5. Steps in Gene Regulation HATs acetylate H3 and H4 N-terminal tails and provide docking sites for other co-regulatory protein complexes. For example, acetylated histones recruit additional HATs that acetylate neighboring nucleosomes leading to the spreading of accessible chromatin. Steps in Gene Regulation The partial opening of chromatin structure allows other DNA binding transcription factors to access their binding sites. Steps in Gene Regulation The binding of additional transcription factors is followed by recruitment of mediator… Steps in Gene Regulation … and the transcription preinitiation complex. This leads to a segment of DNA that is completely devoid of a nucleosome, thus forming a DNAseI hypersensitive site. The example shown above is just one way genes can be activated. In other cases, like heat shock genes, the transcription complex is already bound at the promoter but stalled. Gene activation leads to a change in the transcription complex that renders it elongation competent. Recent evidence suggests that highly expressed genes are recruited to transcription factories in the nucleus. Transcription factories are domains in the nucleus that contain multiple aggregated transcription complexes. Active Genes are Associated with Accessible Chromatin Mapping DNase I Hypersensitive Sites [ DNase I ] 0 1) Treat chromatin/cells with DNase I 2) Purify genomic DNA 3) Digest DNA with restriction enzyme (BamHI) 4) Fractionate DNA on agarose gel 5) Blot gel to membrane (Southern blot) 6) Hybridize blot with probe 7) Expose gel to film for autoradiogram - 15 kb DHS1 - 3.1 kb DHS2 - 1.5 kb Southern blot DHS1 DHS2 B B Exon 1 Probe 1.5kb 3.1kb 15kb RNA Polymerase I Transcription RNA Polymerase III Transcription U6 Gene