Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
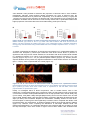
Author version | Pathogens and Disease (2016) 74(2) ftv125 Ângela França and Nuno Cerca* Plasma is the main regulator of S. epidermidis biofilms virulence genes transcription in human blood Centre of Biological Engineering, LIBRO – Laboratory of Research in Biofilms Rosário Oliveira, University of Minho, Campus de Gualtar, 4710-057, Braga, Portugal. * Corresponding author: Nuno Cerca, Centre of Biological Engineering, University of Minho, Campus de Gualtar, 4710-057, Braga, Portugal. Phone: (+351) 253604423. Fax: (+351) 253504400. E-mail: [email protected] Abstract Staphylococcus epidermidis is frequently associated with the emergence of medical deviceassociated bloodstream infections, due to its ability to form biofilms on the surface of vascular catheters. Although these biofilms may be in continuous contact with human blood, how S. epidermidis biofilm cells interact with blood and its cellular and soluble components is poorly understood. Herein, we evaluated biofilm structure, biofilm cells culturability and viability, and the transcription of a panel of genes associated with S. epidermidis biofilms virulence, upon interaction with whole human blood or plasma. Our results showed that although whole human blood caused significant alterations in biofilm structure and in the number of culturable and viable cells, plasma was the main regulator of the transcription of genes with central role in biofilm formation, maturation and immune evasion. These findings highlight the urgent need to intensify studies aiming to evaluate the impact of host soluble factors on S. epidermidis biofilms fitness and persistence. Main text Biofilms present a significant challenge to current clinical practice due to their increased tolerance to antibiotics and host immune response being, consequently, commonly associated with the development of chronic and recalcitrant infections (Costerton 1999). S. epidermidis, a skin- and mucosae-colonizing bacterium, has a remarkable ability to form biofilms on the surface of indwelling medical devices (Vadyvaloo 2005), particularly on peripheral and central vascular catheters. In fact, it was reported that 22% of bloodstream infections diagnosed among patients in intensive care units, in the USA, are due to S. epidermidis biofilm formation on these catheters (Otto 2009). Nevertheless, although medical device-related bloodstream infections are one of the most frequent infections caused by S. epidermidis biofilms, the interaction between biofilm cells and human blood is not well understood. Recently, we have shown that the incubation with human blood led to an extensive reprogramming of S. epidermidis biofilm cells transcriptome (Franca 2014). Nevertheless, since human blood is a complex mixture of cells and soluble molecules, both involved in bacteria targeting, it was not possible to identify which of these fractions were causing the transcriptional alterations observed in that study. Therefore, we became interested in differentiating the response of S. epidermidis biofilms to blood circulating cells versus soluble factors. Hence, herein, S. epidermidis biofilms were incubated, respectively, with whole human blood or human plasma and the abundance of transcripts encoding S. epidermidis biofilm virulence factors was assessed. In addition, biofilm structure and biofilm cells culturability and viability were also evaluated. Human blood was collected, into BD Vacutainer® lithium heparin tubes (NJ, USA), from healthy adults volunteers and under a protocol approved by the Institutional Review Board of the University of Minho (SECVS 002/2014), which is in strict accordance with the Declaration of Helsinki and Oviedo Convention. Informed written consent was signed by all volunteers. Plasma was obtained by centrifugation of whole blood at 300 g, for 20 minutes, at 4 oC. Biofilms of S. epidermidis strain RP62A (ATCC 35984) were formed, in 24-well plates (Orange Scientific, Braine-l’Alleud, Belgium), by adding1.6 µL of overnight culture into 1 mL of tryptic soy broth (Liofilchem, Teramo, Italy) supplemented with 1% glucose (v/v) and by incubating at 37 ºC in an orbital shaker at 120 rpm. Twenty-four hours-old biofilms were incubated for 2 hours, under o static conditions, at 37 C and 5% CO2 with either 1 mL of whole blood or plasma. Biofilms were then washed twice with 0.9% of NaCl and immediately suspended in 1 mL of the same solution, for the evaluation of biofilms biomass and biofilm cells culturability and viability, or in 1 mL RNA ProtectTM bacteria reagent (QIAGEN, Hilden, Germany), for the quantification of gene expression levels. Biofilms biomass was determined by optical density (OD) (Freitas 2014) at 640 nm, the number of culturable cells by the standard CFU counting method, and the number of viable cells by flow cytometry (EC800 flow cytometer, Sony, CA, USA) using SYBR Green and propidium iodide staining as described before (Cerca 2011b). All the quantification analyses were performed using blood/plasma of four different donors (two males and two females), with triplicates for each condition. For confocal laser scanning microscopy (CLSM), 24 hours-old biofilms were formed on NuncTM ThermanoxTM coverslips (Thermo Scientific, MA, USA), which were placed inside the wells of the 24-well plates used. After incubation with blood or plasma, the biofilms formed on the coverslips were washed twice with ultrapure water, the coverslips removed from the well and placed onto a glass slide. Then, polysaccharides were stained with Wheat Germ Agglutinin (WGA) Oregon Green 488 (Life Technologies, CA, USA) and nucleic acids with DAPI (Sigma, MO, USA), as detailed elsewhere (Cerca 2012). Biofilm images were acquired using an OlympusTM FluoView FV1000 (Lisboa, Portugal) microscope. Biofilms were observed using a 60 × water-immersion objective (60 ×/ 1.2 W). Images were taken, with 1024 × 1024 resolution, in at least four different regions of each of the surfaces analyzed. This experiment was performed using blood/plasma of three different donors (two males and one female), with duplicates for each condition. For the quantification of gene expression, the abundance of target transcripts was determined as optimized before (Franca 2014). In brief, total RNA was isolated from biofilms incubated with blood/plasma of five different donors (three males and two females) by combining silica beads lysis with RNeasy mini kit (QIAGEN purification column system. Genomic DNA was degraded by DNase I (Thermo Scientific) and complementary (c) DNA synthesized, from 300 ng of total RNA, by RevertAid H minus Reverse Transcriptase enzyme (Thermo Scientific) in the presence of random primers (NZYTech, Lisboa, Portugal), and in a 10 µL reaction volume. Finally, qPCR run was performed by mixing 2 μL of 1:400 diluted cDNA with 5 μL of Q™ SYBR® Green Supermix (Bio-Rad, CA, USA), 0.5 μL of each forward and reverse primers at 10 µM and 2 μL of nuclease-free water. The run was completed in a CFX96TM (Bio-Rad) thermal cycler with the following parameters: o o o o 94 C 10 minutes and 35 cycles of 94 C 15 seconds, 20 seconds at 58 C and 72 C for 25 seconds. The abundance of transcripts was determined by applying the Pfaffl equation (Pfaffl 2004) using 16 rRNA as reference gene. The sequence of the primers used can be found in previous publications (Franca 2012, Sousa 2014), with the exception of the primers used to detect sepA (FW: TCTTAAGGCATCTCCGCCTA and RV: GTCTGGTGCGAATGATGTTG, 194 bp amplicon) and capA (FW: TGATTATGCGAACCTTTCCAG and RV: TAGCGAATGCTGGTGCAG, 175 bp amplicon) transcripts. The sequences of target genes were downloaded, for primer design, using GeneBank accession number NC_002976. The heat map of relative transcription was created using matrix2png interface (Pavlidis and Noble 2003). Statistical analysis was carried out using GraphPad Prism (CA, USA). Kruskal-Wallis and Dunn’s multiple comparison tests were applied. Accordingly, data are depicted in median with range. Differences among groups were considered significant when P was less than 0.05. As expected, biofilms biomass (Figure 1A) and biofilm cells culturability (Figure 1B) and viability (Figure 1C) were significantly reduced in the presence of whole human blood. A more pronounced reduction, however, was observed in the culturability of biofilm cells rather than in viability. This difference was not surprising since biofilms are highly heterogeneous communities comprising cells in different metabolic states including, among others, dormant cells (Stewart 2008). In addition to the intrinsic biofilm heterogeneity, the high concentration of glucose (1%) used for the growth of these biofilms is known to further stimulate dormancy within S. epidermidis biofilms (Cerca 2011a). By comparing the number of CFU and the number of viable cells obtained it was possible to determine the proportion of dormant cells in each condition. Interestingly, although control biofilms already presented 0.5 Log10 of dormant cells, the incubation with either blood or plasma led, respectively, to an increase of 2-fold (1 Log10) and 1.5-fold (0.75 Log10) in the proportion of dormant cells. This phenomenon may contribute to the persistence of biofilm cells in the host as it was recently shown that S. epidermidis biofilms with higher proportion of dormant cells have lower inflammatory profile (Cerca 2014). Figure 1A,1B,1C. The influence of whole human blood and plasma on S. epidermidis biofilms. (A) Biofilms biomass, (B) number of CFU or (C) viable cells after incubation with whole human blood or plasma. The bars represent the median with range of four independent assays. Statistical significance among groups was determined using Kruskal-Wallis and Dunn’s multiple comparison tests. **P ≤ 0.01, *** P ≤ 0.001. To further characterize the influence of human blood and plasma on S. epidermidis biofilms, its structure was analyzed by CSLM upon interaction with whole human blood or plasma. In agreement with the previous results obtained, the incubation with whole human blood resulted in a significant reduction (P ≤ 0.001) of the maximum biofilm thickness, from 35.6 ± 13.3 µm (control) (Figure 1D) to 8.2 ± 4.3 µm (blood-exposed biofilms) (Figure 1E). Human plasmaexposed biofilms did not present any significant alteration in either biofilm structure or thickness (22.1 ± 4.5 µm), when compared with the control (Figure 1F). Figure 1D, 1E, 1F. The influence of whole human blood and plasma on S. epidermidis biofilms. Representative images of the three-dimensional structure of control biofilms (D), biofilms exposed to blood (E) or plasma (F). DAPI, which stain nucleic acids, is represented in red and WGA, which stains polysaccharides, is represented in green. Finally, to investigate which of blood components, cells or soluble factors, were in fact influencing S. epidermidis biofilms gene expression profile, the transcription levels of genes with a central role in biofilm development/maturation such as aap (Hussain 1997, Rohde 2005), bhp (Tormo 2005), icaA (Mack 1996) and lrgB (Bayles 2007) and in immune evasion such as icaA (Cerca 2006, Kristian 2008), capA (Kocianova 2005), mprF/fmtC (Gill 2005, Richards 2015) and sepA (Lai 2007) were measured. Despite the characteristic variability inherent to each donor, the transcription of these genes was significantly increased, when compared with biofilm cells harvested before any manipulation, in biofilms exposed to either whole human blood or plasma (Figure 1G). However, when comparing the abundance of transcripts between blood- and plasma-exposed biofilms, no meaningful differences were observed with the exception of sepA and aap transcripts, which were found in higher amount within biofilm cells incubated with human plasma (Figure 1H). SepA is a protease that is involved in bacterial immune evasion through the degradation of antimicrobial peptides produced by the host (Lai 2007). As it could be expected, as a defense mechanism, the transcription of sepA would be increased in the presence of either blood or plasma. However, the increased transcription observed in plasmaexposed biofilms may be related to the overrepresentation of host proteins and due to the absence of leukocytes, which may interact with biofilm cells and selectively trim the transcription of particular genes. Similarly, the higher levels of aap transcripts detected in plasma-exposed biofilms were probably due to the same reasons. Interestingly, it was shown that host proteases are capable of activating Aap by proteolysis, which in turn will help to establish a more cohesive biofilm with increased resistance to host factors (Rodhe 2005). Figure 1D, 1E, 1F. The influence of whole human blood and plasma on S. epidermidis biofilms. (G) Heat map of the relative transcription levels (comparing to T0h) of the selected virulence genes upon 2 hours of interaction with whole human blood or plasma. (H) Relative transcription levels of the selected virulence genes in whole human blood (comparing to plasma). The vertical bars represent the median of 5 independent assays. Log2 fold change equal to 0 indicates no alteration in the transcription, Log2 fold change below 0 indicates increase in transcription in plasma-exposed biofilms and Log2 fold change above 0 indicates increase in transcription in blood-exposed biofilms. Each symbol represents a single donor. Statistical significance among groups was determined using Kruskal-Wallis and Dunn’s multiple comparison tests. **P ≤ 0.01, *** P ≤ 0.001. The ability of S. epidermidis biofilm cells to evade bacteria-targeting molecules present in human plasma provides an exceptional competence to survive in a wide variety of human fluids. Apart from constituting approximately 55% of whole human blood, plasma is present, at varying concentrations, in several other important body fluids such as lymphatic and synovial fluids, acute soft tissue wounds and burn wounds exudates (Cardile 2014), fluids where S. epidermidis biofilm infections may have special incidence. Overall these results indicate that, although plasma is less capable of reducing the number of biofilm cells or biomass, is the main regulator of the transcription of genes encoding key biofilm virulence factors (Figure 1 G, H). In addition, this finding highlights the urgent need to study, in more detail, the interaction between the components of human plasma and S. epidermidis biofilms in order to better understand the capacity of these bacterial cells to persist in the host. It is however important to highlight that the proteins that are secreted during bacterial infections (acute-phase proteins, such as C-reactive protein), which may polarize biofilm response (Malachowa 2011), are not present in healthy individuals such the ones used for the experiments herein reported. Hence, in the future, to better comprehend the interplay between S. epidermidis biofilm cells and human plasma components, it would be important to repeat these experiments using plasma of infected patients. In addition, other S. epidermidis strains need to be tested since strain-to-strain variability may occur, as we have previously shown (Franca 2013). Acknowledgements This work was supported by European Union funds (FEDER/COMPETE) and by national funds (FCT) under the project with reference FCOMP-01-0124-FEDER-041246 (EXPL/BIAMIC/0101/2013). The authors thank the FCT Strategic Project of UID/BIO/04469/2013 unit and the project FCOMP-01-0124-FEDER-027462 (RECI/BBB-EBI/0179/2012). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. AF acknowledges the financial support of an individual grant SFRH/BPD/99961/2014. NC is an Investigador FCT. The authors would like to thank Sandra Maia and Filomena Costa for human blood collection procedure. The authors declare no conflict of interest. References Bayles KW. The biological role of death and lysis in biofilm development. Nat Rev Microbiol 2007; 5: 721-6. Cardile AP, Sanchez CJ, Jr., Samberg ME et al. Human plasma enhances the expression of Staphylococcal microbial surface components recognizing adhesive matrix molecules promoting biofilm formation and increases antimicrobial tolerance In Vitro. BMC Res Notes 2014; 7: 457. Cerca F, Andrade F, Franca A et al. Staphylococcus epidermidis biofilms with higher proportions of dormant bacteria induce a lower activation of murine macrophages. J Med Microbiol 2011a; 60: 1717-24. Cerca F, Franca A, Perez-Cabezas B et al. Dormant bacteria within Staphylococcus epidermidis biofilms have low inflammatory properties and maintain tolerance to vancomycin and penicillin after entering planktonic growth. J Med Microbiol 2014; 63: 1274-83. Cerca F, Trigo G, Correia A et al. SYBR green as a fluorescent probe to evaluate the biofilm physiological state of Staphylococcus epidermidis, using flow cytometry. Can J Microbiol 2011b; 57: 850-6. Cerca N, Gomes F, Pereira S et al. Confocal laser scanning microscopy analysis of S. epidermidis biofilms exposed to farnesol, vancomycin and rifampicin. BMC Res Notes 2012; 5: 244. Cerca N, Jefferson KK, Oliveira R et al. Comparative antibody-mediated phagocytosis of Staphylococcus epidermidis cells grown in a biofilm or in the planktonic state. Infect Immun 2006; 74: 4849-55. Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science 1999; 284: 1318-22. Franca A, Carvalhais V, Maira-Litran T et al. Alterations in the Staphylococcus epidermidis biofilm transcriptome following interaction with whole human blood. Pathog Dis 2014; 70: 444-8. Franca A, Freitas AI, Henriques AF et al. Optimizing a qPCR gene expression quantification assay for S. epidermidis biofilms: a comparison between commercial kits and a customized protocol. PLoS One 2012; 7: e37480. Franca A, Vilanova M, Cerca N et al. Monoclonal antibody raised against PNAG has variable effects on static S. epidermidis biofilm accumulation in vitro. Int J Biol Sci 2013; 9: 518-20. Freitas AI, Vasconcelos C, Vilanova M et al. Optimization of an automatic counting system for the quantification of Staphylococcus epidermidis cells in biofilms. J Basic Microbiol 2014; 54: 750-7. Gill SR, Fouts DE, Archer GL et al. Insights on evolution of virulence and resistance from the complete genome analysis of an early methicillin-resistant Staphylococcus aureus strain and a biofilm-producing methicillin-resistant Staphylococcus epidermidis strain. J Bacteriol 2005; 187: 2426-38. Hussain M, Herrmann M, von Eiff C et al. A 140-kilodalton extracellular protein is essential for the accumulation of Staphylococcus epidermidis strains on surfaces. Infect Immun 1997; 65: 519-24. Kocianova S, Vuong C, Yao Y et al. Key role of poly-gamma-DL-glutamic acid in immune evasion and virulence of Staphylococcus epidermidis. Journal of Clinical Investigations 2005; 115: 688-94. Kristian SA, Birkenstock TA, Sauder U et al. Biofilm formation induces C3a release and protects Staphylococcus epidermidis from IgG and complement deposition and from neutrophildependent killing. J Infect Dis 2008; 197: 1028-35. Lai Y, Villaruz AE, Li M et al. The human anionic antimicrobial peptide dermcidin induces proteolytic defence mechanisms in staphylococci. Mol Microbiol 2007; 63: 497-506. Mack D, Fischer W, Krokotshc A et al. The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a liner beta-1.6-linked glucosaminoglycan: purification and structural analysis. J Bacteriol 1996; 178: 175-83. Malachowa N, Whitney AR, Kobayashi SD et al. Global changes in Staphylococcus aureus gene expression in human blood. PLoS One 2011; 6: e18617. Otto M. Staphylococcus epidermidis--the 'accidental' pathogen. Nat Rev Microbiol 2009; 7: 55567. Pfaffl MW. Quantification strategies in real-time PCR. In: Bustin, S. A. La Jolla, CA, USA: International University Line, 2004, 87-112. Pavlidis P, Noble WS. Matrix2png: a utility for visualizing matrix data. Bioinformatics 2003; 19: 295-6. Richards RL, Haigh RD, Pascoe B et al. Persistent Staphylococcus aureus Isolates from Two Independent Cases of Bacteremia Display Increased Bacterial Fitness and Novel Immune Evasion Phenotypes. Infect Immun 2015; 83: 3311-24. Rohde H, Burdelski C, Bartscht K et al. Induction of Staphylococcus epidermidis biofilm formation via proteolytic processing of the accumulation-associated protein by staphylococcal and host proteases. Mol Microbiol 2005; 55: 1883-95. Sousa C, Franca A, Cerca N. Assessing and reducing sources of gene expression variability in Staphylococcus epidermidis biofilms. Biotechniques 2014; 57: 295-301. Stewart PS, Franklin MJ. Physiological heterogeneity in biofilms. Nat Rev Microbiol 2008; 6: 199-210. Tormo MA, Knecht E, Gotz F et al. Bap-dependent biofilm formation by pathogenic species of Staphylococcus: evidence of horizontal gene transfer? Microbiology 2005; 151: 2465-75. Vadyvaloo V,Otto M. Molecular genetics of Staphylococcus epidermidis biofilms on indwelling medical devices. Int J Artif Organs 2005; 28: 1069-78.