Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
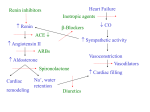
J Appl Physiol 92: 18–24, 2002. . Effects of moderate heart failure and functional overload on rat plantaris muscle ESPEN E. SPANGENBURG,1 SIMON J. LEES,1 JEFF S. OTIS,1 TIMOTHY I. MUSCH,2 ROBERT J. TALMADGE,1 AND JAY H. WILLIAMS1 Muscular Function Laboratory, Department of Human Nutrition, Foods, and Exercise, Virginia Polytechnic Institute and State University, Blacksburg, Virginia 24061; and 2Departments of Kinesiology and Anatomy and Physiology, Kansas State University, Manhattan, Kansas 66506 Received 19 April 2001; accepted in final form 17 August 2001 MANY INVESTIGATIONS HAVE DEMONSTRATED that, during congestive heart failure (CHF), reductions in exercise capacity are not solely due to altered myocardial function (2, 15, 16, 29). In fact, various indicators of cardiac function (e.g., ejection fraction) do not correlate well with overall exercise capacity in CHF patients, even though there are strong correlations in healthy subjects (5, 29). For example, Franciosa et al. (5) showed that measurements of left ventricular performance (i.e., ejection fraction) were unrelated to exercise duration (r ⫽ ⫺0.06) in CHF patients. Exercise capacity does seem to correlate well with indexes of peripheral muscular function such as muscular strength and endurance (16, 29). For example, Minotti et al. (16) demonstrated that knee extensor muscular endurance and peak oxygen consumption were closely correlated (r ⫽ 0.90) in CHF patients. Interestingly, the same correlation was not significant in healthy subjects (r ⫽ 0.37). The most obvious skeletal muscle functional changes include decreased strength and endurance (2). Specifically, CHF patients performing voluntary leg extensions all show decreased maximal force and are less able to maintain the force output during repetitive contractions compared with control subjects. Using animal models of CHF, Williams and Ward (31) and Perrault et al. (20) showed decreases in maximal force production. These changes persisted even when forces were normalized by wet muscle mass or cross-sectional area (CSA), suggesting that factors other than muscle atrophy must contribute to the reduced force generation. Intracellular free Ca2⫹ transients in skeletal muscle are tightly regulated by various proteins associated with the sarcoplasmic reticulum (SR) (4). Ultimately, it is the control of these free Ca2⫹ transients that regulates skeletal muscle contractility (4). Alterations in the function of these proteins are associated with alterations in skeletal muscle force production (30, 32). Perrault et al. (20) showed that, in severe CHF, there is a slowing of intracellular Ca2⫹ transients in isolated extensor digitorum longus (EDL) fibers. Furthermore, Williams and Ward (31) found that moderate CHF induces increases in both SR Ca2⫹ uptake and release from isolated SR vesicles. One key difference between these published studies is the severity of CHF exhibited within the animal. Therefore, it is possible that severity of CHF affects the function of skeletal muscle SR. Recent investigations suggest that changes in SR Ca2⫹-ATPase activity may result from changes in sarco(endo)plasmic reticulum Ca2⫹-ATPase (SERCA) isoform expression (for review, see Ref. 4). SERCA1, Address for reprint requests and other correspondence: E. E. Spangenburg, Univ. of Missouri, Dept. of Biomedical Sciences, E102 Veterinary Medical Bldg., 1600 East Rollins, Columbia, MO 65211 (E-mail: [email protected]). The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. calcium-adenosinetriphosophatase; hypertrophy; sarcoplasmic reticulum; chronic heart failure 18 8750-7587/02 $5.00 Copyright © 2002 the American Physiological Society http://www.jap.org Downloaded from http://jap.physiology.org/ by 10.220.33.5 on May 2, 2017 Spangenburg, Espen E., Simon J. Lees, Jeff S. Otis, Timothy I. Musch, Robert J. Talmadge, and Jay H. Williams. Effects of moderate heart failure and functional overload on rat plantaris muscle. J Appl Physiol 92: 18–24, 2002.—It is thought that changes in sarco(endo)plasmic reticulum Ca2⫹-ATPase (SERCA) of skeletal muscle contribute to alterations in skeletal muscle function during congestive heart failure (CHF). It is well established that exercise training can improve muscle function. However, it is unclear whether similar adaptations will result from exercise training in a CHF patient. Therefore, the purpose of this study was to determine whether skeletal muscle during moderate CHF adapts to increased activity, utilizing the functional overload (FO) model. Significant increases in plantaris mass of the CHF-FO and sham-FO groups compared with the CHF and control (sham) groups were observed. Ca2⫹ uptake rates were significantly elevated in the CHF group compared with all other groups. No differences were detected in Ca2⫹ uptake rates between the CHF-FO, sham, and sham-FO groups. Increases in Ca2⫹ uptake rates in moderate-CHF rats were not due to changes in SERCA isoform proportions; however, FO may have attenuated the CHF-induced increases through alterations in SERCA isoform expression. Therefore, changes in skeletal muscle Ca2⫹ handling during moderate CHF may be due to alterations in regulatory mechanisms, which exercise may override, by possibly altering SERCA isoform expression. CHF AND EXERCISE ALTER SR CA2⫹ HANDLING MATERIALS AND METHODS Animal model. For all experiments, the rat coronary artery ligation model was used to induce CHF according to the methods of Musch et al. (17, 18). Twenty-three SpragueDawley rats (⬃200 g) female rats were subjected to surgically induced myocardial infarctions (CHF group), and thirteen additional rats were subjected to sham surgeries (sham group). Briefly, the rats were anesthetized with 2% halothane and then placed on a rodent ventilator, and a left lateral thoracotomy was performed. For the CHF animals, the heart was isolated and the left coronary artery was ligated with 6-0 suture. The sham animals underwent the same procedures except the artery was not ligated. The heart was checked for damage other than the effects of ligation and was gently placed back into the chest. The intercostal muscles and the incision in the intercostal space were closed with 3-0 suture. The animals were checked daily to assess and respond to any postsurgical complications. The ablation surgeries were performed 3 wk postinfarction. Sixteen CHF and eight sham animals were subjected to unilateral FO surgeries. A unilateral FO model was chosen because the myocardial infarction model results in variable degrees of myocardial injury. This design allows each animal to serve as its own internal control, having both non-FO and FO muscles. This resulted in four groups of muscles: sham, sham-FO, CHF, and CHF-FO. The remaining animals did not undergo FO and were used to ensure that the non-FO leg of the FO animals was indicative of a nonoperated control limb. The FO surgeries were performed according to the methodology of Sugiura et al. (25). The animals were anesthetized with 1.5% halothane. Under aseptic conditions, an incision was made through the skin on the posterior aspect of the animal’s lower leg. The distal tendon of the gastrocnemius was transected, and the muscle was removed from the surrounding tissue, with care taken not to disturb the vasculature and innervation leading to the soleus and plantaris muscles. The skin and hamstrings were then sutured closed with 3-0 silk suture. The animals were then returned to their cages for 9 wk, which have been shown to be ample time to J Appl Physiol • VOL allow for adaptation of both the soleus and plantaris (25). The animals were observed daily for sudden changes in health and/or monitoring of body weight. Tissue processing. Nine weeks postablation, the animals were anesthetized with an intraperitoneal injection of pentobarbital sodium (60 mg/kg). The plantaris muscles of both limbs were then subsequently removed, blotted dry, and weighed. The muscles were then cut in half, with one-half used for SR function and SERCA blot measurements and the other half placed in melting isopentane cooled by liquid nitrogen. The first half of the muscle was homogenized in 10 vol of buffer consisting of 20 mM HEPES (pH 7.4), 250 mM sucrose, 0.2% sodium azide, and 0.2 mM phenylmethylsulfonyl fluoride (30). The homogenate was then centrifuged for 10 min at 1,600 g at 4°C. The supernatant was removed and stored at ⫺80°C. A portion of this supernatant was used for SERCA isoform determination, and the other portion of the supernatant was used for SR Ca2⫹ uptake measurements (see SR Ca2⫹ uptake). Protein concentrations were determined via the Bradford methodology (1). The frozen portion of the muscle was stored at ⫺80°C for immunohistochemistry. The heart and lungs were removed, trimmed free of connective tissue, weighed, and placed into 10% buffered formalin. Determination of infarct size and ventricular mass. Determinations of the changes in heart mass and infarct size were determined according to the methods of Pfeffer et al. (22). The right and left ventricles were carefully dissected, weighed, and placed in 10% buffered formalin. The left ventricle was cut into three transverse sections and processed for histological studies. Sections (10 m thick) were cut, mounted, and stained with Masson’s trichrome. The sections were then projected onto a monitor where the percentage of the left ventricle circumference containing the infarcted region was determined. The fraction of the left ventricle that was infarcted vs. the amount noninfarcted was then calculated from these measurements. Western blotting for SERCA. SERCA isoform determination was performed according to Kandarian et al. (12). The proteins were electrophoretically separated on 7.5% SDSpolyacrylamide gels (13). They were then transferred to nitrocellulose membranes according to Towbin et al. (28). The membranes were then blocked for 1 h with 5% nonfat dry milk and incubated overnight with the primary antibody against slow/cardiac Ca2⫹ pump (2A7-A1, Affinity Bioreagents; 1:500) or the fast skeletal Ca2⫹ pump (IIH11, Affinity Bioreagents; 1:2,500) at 5°C. The blots were incubated in secondary antibody, an anti-mouse IgG conjugated to alkaline phosphatase (A9316, Sigma Chemical; 1:30,000). Color development was performed using bromochloroindolyl phosphatase-nitro blue tetrazolium as a substrate for alkaline phosphatase. The bands were scanned using video densitometry (Alpha Innotech Image Analysis System). Each blot also contained a homogenized EDL sample, determined to express only SERCA1, or a homogenized heart sample, determined to express to SERCA2 (33). This allowed plantaris samples to be normalized by the heart or EDL sample, providing semiquantification of protein content. Furthermore, neither SERCA antibody showed any cross-reactivity with other isoform of SERCA (data not shown). SR Ca2⫹ uptake. The SR Ca2⫹ uptake measurements were measured using a homogenate fraction according to the methods of Ward et al. (30). Briefly, 250 g of homogenate protein were added to 1 ml of buffer containing 100 mM KCl, 20 mM HEPES, 5.0 mM MgCl2, 2.5 mM ATP, and 5.0 mM oxalate (pH 7.0). Temperature was maintained at 37°C, and the buffer was constantly stirred with a micro stir bar. 92 • JANUARY 2002 • www.jap.org Downloaded from http://jap.physiology.org/ by 10.220.33.5 on May 2, 2017 the fast isoform, is typically present predominately fast skeletal muscle, whereas SERCA2, the slow isoform, is present in slow and cardiac muscle (33). A number of studies show that altered activity as well as various disease states can affect both SERCA isoform expression and SR function (10, 11, 21, 23). Therefore, it may be possible that the changes in muscle SERCA expression may account for changes in SR function in CHF. In fact, two reports suggest that such is the case (21, 23). We hypothesized that a form of increased skeletal muscle activation may attenuate and/or prevent intracellular changes in the SR induced by moderate CHF in the skeletal muscle. In this study, we combined the rat myocardial infarction model of CHF with a skeletal muscle functional overload (FO) model. The FO model was chosen to ensure increased activation of the skeletal muscle during CHF. The interesting aspect of these two models is that they produce opposing effects on SR function. For example, moderate CHF increases the rate of Ca2⫹ uptake by the SR (31), whereas Ca2⫹ uptake is reduced in FO (12). Here we show that FO increases muscle mass and also attenuates CHF-induced increases in SR function by altering SERCA isoform expression. 19 20 CHF AND EXERCISE ALTER SR CA2⫹ HANDLING Table 1. Effects of CHF on heart and lung morphology Parameter Sham Group (n ⫽ 12) CHF Group (n ⫽ 24) Body mass, g Heart mass, g Heart mass, mg/g Right ventricle mass, g Right ventricle mass, mg/g Left ventricle mass, g Left ventricle mass, mg/g Lung mass, g Lung mass, mg/g 271.5 ⫾ 5.3 1.04 ⫾ 0.02 3.84 ⫾ 0.09 0.14 ⫾ 0.01 0.51 ⫾ 0.02 0.78 ⫾ 0.02 2.87 ⫾ 0.05 1.35 ⫾ 0.03 4.99 ⫾ 0.08 283.6 ⫾ 3.73 1.35 ⫾ 0.05* 4.78 ⫾ 0.18* 0.23 ⫾ 0.01* 0.80 ⫾ 0.05* 0.98 ⫾ 0.02* 3.47 ⫾ 0.08* 1.78 ⫾ 0.18 6.33 ⫾ 0.66 Values are means ⫾ SE; n, no. of animals. CHF, congestive heart failure; Sham, control. Animals that did and did not undergo ablation surgery within the same condition have been pooled. * P ⬍ 0.05 between conditions. J Appl Physiol • VOL RESULTS Body mass, heart mass, and lung morphology. No differences in mean body weights between the CHF and sham groups were detected (Table 1). The CHF animals demonstrated a 33 ⫾ 1% infarction of the left ventricular endocardial circumference, whereas no infarctions were detected in any of the sham animals. There was a 24% increase in relative heart mass as well as 21 and 60% increases in relative left and right ventricular masses, respectively, in the CHF group compared with sham (Table 1). There were no significant changes in absolute or relative lung mass between groups. This suggests that these animals were in a moderate form of heart failure based on the tissue and hemodynamic characteristics previously published by Delp et al. (3), who used the same model to induce CHF. Skeletal muscle mass. There was no significant effect of condition on plantaris muscle mass (Fig. 1). However, FO resulted in similar increases in both absolute and relative mass in both the sham and CHF groups Fig. 1. Effects of congestive heart failure (CHF) and functional overload (FO) on plantaris muscle mass. A: absolute mass. B: relative mass. Values are means ⫾ SE; n ⫽ 8 for sham and 16 for CHF groups. *P ⬍ 0.05 vs. non-FO muscle within each condition. 92 • JANUARY 2002 • www.jap.org Downloaded from http://jap.physiology.org/ by 10.220.33.5 on May 2, 2017 Uptake was initiated by the addition of CaCl2 (⬃1 M free Ca2⫹) and continued until no change in extravesicular free Ca2⫹ is observed. Extravesicular free Ca2⫹ was measured using the fluorescent Ca2⫹ indicator fura 2 and a Jasco CAF 110 dual-wavelength fluorometer. Excitation light was filtered at 340 and 380 nm, and emission fluorescence was detected at 500 nm. Fluorescence ratios were used to calculate free Ca2⫹ concentration ([Ca2⫹]) in the incubation medium according to the methods of Grynkiewicz et al. (9). The peak rate of Ca2⫹ uptake was calculated from the steepest portion of the negative slope of the total [Ca2⫹] vs. time curve and then normalized to the sample protein concentration. Immunohistochemistry. Serial sections from the deep region of the plantaris were cut in a cryostat (Microm HM 505 N) maintained at ⫺20°C. The 10-m-thick sections were placed on gelatin-coated slides. The immunohistochemistry was preformed according to the techniques of Talmadge et al. (27). The sections were exposed to mouse monoclonal antibodies specific for SERCA1 (VE121G9, Novocastra Laboratories) and SERCA2 (IID8, Novocastra Laboratories) of the rat. For determination of myosin heavy chain expression (MHC), the fibers were exposed to mouse monoclonal antibodies specific to the various MHC isoforms of the rat. The antibodies used were as follows: BF-D5 (anti-type I MHC), SC-71 (anti-type IIa MHC), BF-35 (anti-all MHCs except type IIx), BF-F3 (anti-type IIb MHC), BF-13 (anti-type II MHC), and BF-G6 (anti-embryonic MHC). The sections were incubated in the primary antibody for 1.5 h at 37°C. The avidin-biotin procedure was used to amplify the antigen-antibody complex (Vectastain ABC kits, Vector Laboratories). After staining, a single region containing ⬃100 fibers from each muscle were randomly selected from the deep region of the plantaris. The fibers were followed through the serial sections using video images generated off a microscope (model E400, Nikon) and the Scion image program on a microcomputer. A fiber that showed a positive reaction was considered to be positive for that particular primary antibody. Individual muscle fibers staining positive for different isoforms of SERCA were labeled as SERCA isoform-positive fibers. Fiber size or CSA was measured according to the techniques of Grossman et al. (8). Briefly, all of the fibers in the randomly selected region were manually traced with the aid of an image-processing system (Scion image). The calibrated system allows for determination of the CSA of individual fibers. Statistics. A two-way ANOVA (split-plot ANOVA) with repeated measures was used to compare the effects of condi- tion (sham, CHF) and treatment (FO, non-FO). If ANOVA revealed significant effects, a Tukey’s post hoc test was applied. Differences were considered statistically significant at the P ⬍ 0.05 level. CHF AND EXERCISE ALTER SR CA2⫹ HANDLING 21 Fig. 2. Effects of CHF and FO on sarcoplasmic reticulum Ca2⫹ uptake. Values are means ⫾ SE; n ⫽ 8 for sham and 16 for CHF groups. A: Ca2⫹ uptake. Rates were normalized to sample protein content. *P ⬍ 0.05 vs. all other groups. B: Ca2⫹ concentration that elicited 50% of peak uptake rates ([Ca2⫹]50). *P ⬍ 0.05 vs. CHF-FO group. J Appl Physiol • VOL subsequent increase in type I fibers of the sham-FO group compared with the sham group. None of the groups had any significant changes in type IIa fibers. Some groups did demonstrate the appearance of coex- Fig. 3. Sarco(endo)plasmic reticulum Ca2⫹-ATPase (SERCA) isoform expression of the plantaris muscle as determined by Western blotting procedure. A: SERCA1 and SERCA2 expression. Lane 1, sham-FO; lane 2, sham; lane 3, CHF-FO; lane 4, CHF; lane 5, control extensor digitorum longus for SERCA1 and control heart tissue for SERCA2. SERCA1 gels were loaded with 10 g of total protein, whereas SERCA2 lanes were loaded with 50 g of total protein. B: results for SERCA1 and SERCA2 expression. Values are means ⫾ SE; n ⫽ 8 for sham and 16 for CHF groups. *Significantly different from CHF group, P ⬍ 0.05. 92 • JANUARY 2002 • www.jap.org Downloaded from http://jap.physiology.org/ by 10.220.33.5 on May 2, 2017 (33 and 29% increases, respectively in relative mass). Furthermore, the plantaris masses of the sham and CHF animals that did not undergo ablation surgery (1.41 ⫾ 0.05 and 1.33 ⫾ 0.03 mg/g, respectively) were similar to that of the non-FO leg of those that did experience FO surgery. Thus, with respect to muscle mass the non-FO limb did serve as a “true control limb.” Ca2⫹ uptake rates. The Ca2⫹ uptake rate was significantly elevated by 44% in the non-FO muscles of the CHF animals muscles compared with those of the sham animals (Fig. 2). FO reduced the Ca2⫹ uptake rate in the CHF animals to a level that was not different from the sham. FO, however, did not significantly alter Ca2⫹ uptake in the sham animals. Similarly, the [Ca2⫹] required to evoke 50% of peak uptake rates ([Ca2⫹]50) of the CHF non-FO muscles was significantly lower than the other three muscle groups. SERCA isoform expression. Western blotting showed that there was no effect of CHF on whole muscle SERCA isoform expression (Fig. 3). There was, however, a significant effect of FO in that SERCA2 expression was increased in the FO muscles of both the sham and CHF animals. Immunohistochemical analysis revealed that there was no effect of CHF on the percentage of fibers expressing SERCA1 or, SERCA2 or coexpressing SERCA1 and SERCA2 (Fig. 4). FO, however, significantly decreased the SERCA1 fiber percent in both CHF and sham animals (9 and 12%, respectively) and increased the percentage of SERCA2 fibers in the sham animals only (to 27%). MHC-based fiber-type expression. The CHF group contained 14% more type IIx fibers compared with the sham group. Figure 5 indicates that the CHF-FO group demonstrated a significant increase in the percentage of fibers expressing type I MHC compared with the CHF group. Furthermore, CHF-FO had a significant 19% reduction in the fibers expressing type IIX MHC compared with the CHF group. As expected, there were also significant reductions in the type IIx fibers and a 22 CHF AND EXERCISE ALTER SR CA2⫹ HANDLING Fig. 6. Individual-fiber cross-sectional area (CSA) from the deep medial region of the plantaris muscle. Values are means ⫾ SE; n ⫽ 8 for sham and 16 for CHF groups. *Significantly different from CHF group, P ⬍ 0.05. # Significantly different from sham group, P ⬍ 0.05. of fibers coexpressing both SERCA isoforms were not affected by either CHF or FO. DISCUSSION pressing fibers (i.e., fibers containing multiple MHC isoforms); however, none of the differences was significant. Fiber CSA. There was no effect of condition on CSA of individual fibers (Fig. 6). This was true for fibers expressing SERCA1, SERCA2, or SERCA1 and SERCA2. There were, however, significant effects of the treatment. FO increased CSA of fibers expressing SERCA1 in both CHF and sham animals by 25 and 14%, respectively. In addition, FO increased the CSA of SERCA2 fibers by 56% in the CHF animals only. CSA Fig. 5. Myosin heavy chain-based fiber-type expression of the deep medial region of the plantaris. Values are means ⫾ SE; n ⫽ 8 for sham and 16 for CHF group. * Significantly different from CHF group, P ⬍ 0.05. # Significantly different from sham group, P ⬍ 0.05. J Appl Physiol • VOL The major finding of this study was that skeletal muscle retains its ability to adapt to increased loading in the presence of moderate CHF. This was evidenced by the plantaris muscle hypertrophy and normalization of Ca2⫹ uptake kinetics in the CHF-FO muscles. The changes induced by the FO also occurred without significant alterations in cardiac morphology. In addition, the skeletal muscle hypertrophy that occurred was similar for the CHF-FO and sham-FO groups. Collectively, these data suggest that moderate CHF did not inhibit or attenuate skeletal muscle adaptations resulting from increased activity and load bearing. Surprisingly, the 44% increase in Ca2⫹ uptake rate due to moderate CHF was not associated with changes in SERCA isoform expression. Similarly, Williams and Ward (31) found a 30% increase in Ca2⫹ uptake rates of isolated SR from moderate CHF rats without notable changes in SERCA expression. Williams and Ward also found that that this increase in Ca2⫹ uptake was associated with increased rates of muscle relaxation and decreased force production. On the other hand, Simonini et al. (23) reported a significant 16% reduction in SERCA2 expression in the rat soleus muscle during severe CHF. Their study did not report functional measures, so it is unclear whether such a reduction in SERCA2 translated into alterations in Ca2⫹ pump activity. Also, these investigators found a reduction in soleus muscle mass, whereas our plantaris muscles showed no such change in response to moderate CHF. It is possible that the SR of the plantar flexor muscles respond differently to CHF. Our laboratory has previously shown that MHC expression is altered in some, but not all plantar flexor muscles during CHF (24). 92 • JANUARY 2002 • www.jap.org Downloaded from http://jap.physiology.org/ by 10.220.33.5 on May 2, 2017 Fig. 4. Percentage of individual fibers expressing an isoform of SERCA, as determined via immunohistochemistry, from the deep medial region of the plantaris. SERCA1 & 2, fibers coexpressing both isoforms of SERCA. Values are means ⫾ SE; n ⫽ 8 for sham and 16 for CHF groups. *Significantly different from CHF group, P ⬍ 0.05. # Significantly different from sham group, P ⬍ 0.05. $ Significantly different from CHF-FO group, P ⬍ 0.05. CHF AND EXERCISE ALTER SR CA2⫹ HANDLING J Appl Physiol • VOL SERCA expression found here with the results of Kandarian et al. (12). They found that FO in healthy rats caused an 80% increase in plantaris mass and a small (15%) reduction in Ca2⫹ uptake that were accompanied by a large (130%) increase in SERCA2 expression. Furthermore, Kandarian et al. also found a subsequent 25% decrease in SERCA1 and total SERCA protein expression. In the present study, the increase SERCA2 expression was smaller, only 24%, and no change occurred in SERCA1 expression. This suggests that, in the study of Kandarian et al., the reduction SERCA expression may account for the changes in Ca2⫹ATPase activity. This differs from our study because SERCA expression did not decline; however, this difference may be due to the amount of hypertrophy exhibited in the plantaris muscle. Kandarian et al. found an 80% increase in muscle mass by ablating both legs and removing the soleus muscle, whereas we chose a unilateral ablation and left the soleus intact and only found 33% increase in plantaris muscle mass. Despite this, our FO model completely reduced the rather large (44%) increase in uptake rate caused by moderate CHF. Apparently, in our case, SR Ca2⫹ uptake is a more complex process than can be explained by measures of SERCA isoform expression. As expected, FO in the sham animals caused a 48% increase in the percentage of fibers expressing SERCA2, which was nearly identical to that found by Talmadge et al. (26). FO in the moderate-CHF animals, however, did not induce an increase in the proportion of fibers expressing the SERCA2 protein in the deep region of the plantaris. This was surprising considering the overall increase in SERCA2 expression, when determined by Western blotting. However, it should be noted that immunohistochemistry is not a quantitative measure (i.e., cannot quantify the amount of protein in a fiber), and it is possible that some individual fibers had increased SERCA2 levels. Therefore, it is possible that number of fibers expressing SERCA2 increased, without an increase in total SERCA2 expression. Furthermore, the same reasoning can be used to explain why there is not less SERCA1 in the whole muscle, whereas the percentage of fibers expressing SERCA1 is less in the FO animals. Furthermore, it may be possible that we are detecting a regional adaptation in the muscle fiber SERCA1 expression that is masked in the whole muscle measurements. It should be noted that, during CHF and/or increased loading, there are alterations in SR protein function and expression and it is unclear whether these changes translate into functional adaptations of the skeletal muscle. Therefore, it is imperative that the effects of resistance training during CHF on skeletal muscle function be characterized. This will allow one to truly comprehend the physiological significance of these findings. The data presented here suggest that moderate CHF affects skeletal muscle, particularly the ability of its SR to resequester Ca2⫹. Our results also indicate that this change is independent of alterations in SERCA isoform and reflects changes in other SERCA regula- 92 • JANUARY 2002 • www.jap.org Downloaded from http://jap.physiology.org/ by 10.220.33.5 on May 2, 2017 Here, we confirm that the fiber-type shifts are occurring in the plantaris muscles of animals inflicted with moderate CHF. The present data do not agree with those of Peters et al. (21), who found reductions in both SERCA1 expression and Ca2⫹-ATPase activity. However, this may be due to their use of a genetic rat model of CHF having a severity of CHF that was likely much greater than that in the present study (S. C. Kandarian, personal communication). This model is also susceptible to other conditions that could affect SERCA expression, such as hypertension, cyanosis, and lung hyperemia (21). In addition, Perrault et al. (20), using the same CHF model, found prolonged Ca2⫹ transients in isolated muscle fibers during twitch contractions. However, it should be noted that they obtained a much more severe form of CHF and also they did not measure SERCA expression, so it is unclear whether they had any changes in protein expression accompanying the decreases in Ca2⫹ transients. Thus it is possible that the decrease in SERCA1 expression observed by Peters et al. (21) as well as the decrease in SERCA2 expression demonstrated by Simonini et al. (23) and the decrease in Ca2⫹ transients (20) may only occur in animals with severe CHF. In addition, pilot data our laboratory has collected indicated that Ca2⫹ uptake and release rates are also reduced in cases of severe CHF compared with moderate CHF (J. H. Williams and C. W. Ward, unpublished observations), and our laboratory has also shown that changes in MHC expression are dependent on the severity of failure (24). Our finding that moderate CHF causes increased SR Ca2⫹ uptake without altering SERCA isoform expression is rather unique because chronic exercise conditions are thought to alter SR Ca2⫹ handling by altering the expression of SERCA1 and SERCA2 (10, 12). The present results and those of Williams and Ward (31) suggest that during moderate CHF some unidentified regulatory mechanism is responsible for the increased Ca2⫹ pump activity. There are a number of possibilities that could affect overall Ca2⫹ uptake rates in the absence of altered SERCA expression. Our results also show increased Ca2⫹ sensitivity of the Ca2⫹ pump in the non-FO muscles of CHF animals, a change that is also reversed or prevented by FO. A decrease in the [Ca2⫹]50 would allow higher Ca2⫹ uptake rates at lowto-moderate free [Ca2⫹]. Protein-protein interactions play important roles in the Ca2⫹ handling process (7). It is quite possible that changes in the various proteins that are associated with Ca2⫹ uptake, such as calsequestrin, sarcalumenin, or sarcolipin, may alter the rate of uptake (6, 14, 19). Interestingly, a decrease in sarcolipin content could possibly increase both SERCA1 Ca2⫹ affinity and maximal activity (19). In addition, chemical modification of the Ca2⫹-ATPase, such as phosphorylation status (4) or oxidation of various domains of the Ca2⫹ pump (10), can affect overall pump function. Therefore, it is possible that, during moderate CHF, there are modifications in the Ca2⫹ transport process that are independent of SERCA expression that could affect Ca2⫹ uptake. This notion is emphasized by comparing changes in Ca2⫹ uptake and 23 24 CHF AND EXERCISE ALTER SR CA2⫹ HANDLING tory mechanisms. However, FO can reverse or attenuate at least some of the moderate CHF-induced alterations in the SR, possibly by changing SERCA isoform expression. This leads to the speculation that resistance exercise in the form of strength training may be a beneficial treatment for CHF patients and may ultimately help to improve their exercise capacity. This study was supported in part by National Institute of Arthritis and Musculoskeletal and Skin Diseases Grant AR-41727 (to J. H. Williams) and a National Aeronautics and Space Administration Space Physiology Research Grant from the American College of Sports Medicine Foundation (to E. E. Spangenburg). Present address of E. E. Spangenburg: Veterinary Biomedical Sciences, University of Missouri, E102 Veterinary Medical Bldg., Columbia, MO 65211. REFERENCES J Appl Physiol • VOL 92 • JANUARY 2002 • www.jap.org Downloaded from http://jap.physiology.org/ by 10.220.33.5 on May 2, 2017 1. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–254, 1976. 2. Buller NP, Jones D, and Poole-Wilson PA. Direct measurement of skeletal muscle fatigue in patients with chronic heart failure. Br Heart J 65: 20–24, 1991. 3. Delp MD, Duan C, Mattson JP, and Musch TI. Changes in skeletal muscle biochemistry and histology relative to fiber type in rats with heart failure. J Appl Physiol 83: 1291–1299, 1997. 4. Dux L. Muscle relaxation and sarcoplasmic reticulum function in different muscle types. Rev Physiol Biochem Pharmacol 122: 69–147, 1993. 5. Franciosa JA, Park M, and Levine TB. Lack of correlation between exercise capacity and indexes of resting left ventricular performance in heart failure. Am J Cardiol 47: 33–39, 1981. 6. Froemming GR, Murray BE, Harmon S, Pette D, and Ohlendieck K. Comparative analysis of the isoform expression pattern of Ca2⫹-regulatory membrane proteins in fast-twitch, slow-twitch, cardiac, neonatal and chronic low-frequency stimulated muscle fibers. Biochim Biophys Acta 1466: 151–168, 2000. 8. Grossman EJ, Roy RR, Talmadge RJ, Zhong H, and Edgerton VR. Effects of inactivity on myosin heavy chain composition and size of rat soleus fibers. Muscle Nerve 21: 375–389, 1998. 7. Froemming GR and Ohlendieck K. Oligomerization of Ca2⫹regulatory membrane components involved in the excitationcontraction-relaxation cycle during postnatal development of rabbit skeletal muscle. Biochim Biophys Acta 1387: 226–238, 1998. 9. Grynkiewicz G, Poenie M, and Tsien RY. A new generation of Ca2⫹ indicators with greatly improved fluorescence properties. J Biol Chem 260: 3440–3450, 1985. 10. Harmon S, Froemming GR, Leisner E, Pette D, and Ohlendieck K. Low-frequency stimulation of fast muscle affects the abundance of the Ca2⫹-ATPase but not its oligomeric status. J Appl Physiol 90: 371–379, 2001. 11. Kandarian SC, Peters DG, Favero TG, Ward CW, and Williams JH. Adaptation of the skeletal muscle calcium release mechanism to weight-bearing condition. Am J Physiol Cell Physiol 270: C1588–C1594, 1996. 12. Kandarian SC, Peters DG, Taylor JA, and Williams JH. Skeletal muscle overload upregulates the sarcoplasmic reticulum slow calcium pump gene. Am J Physiol Cell Physiol 266: C1190–C1197, 1994. 13. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680–685, 1970. 14. Leberer E, Timms BG, Campbell KP, and MacLennan DH. Purification, calcium binding properties, and ultrastructural localization of the 53,000- and 160,000 (sarcalumenin)-dalton glycoproteins of the sarcoplasmic reticulum. J Biol Chem 265: 10118–10124, 1990. 15. Lunde PK, Verburg E, Vollestad NK, and Sejersted OM. Skeletal muscle fatigue in normal subjects and heart failure patients. Is there a common mechanism? Acta Physiol Scand 162: 215–228, 1998. 16. Minotti JR, Christoph I, Oka R, Weiner MW, Wells L, and Massie BM. Impaired skeletal muscle function in patients with congestive heart failure. Relationship to systemic exercise performance. J Clin Invest 88: 2077–2082, 1991. 17. Musch TI, Moore RL, Leathers DJ, Bruno A, and Zelis R. Endurance training in rats with chronic heart failure induced by myocardial infarction. Circulation 74: 431–441, 1986. 18. Musch TI and Terrell JA. Skeletal muscle blood flow abnormalities in rats with a chronic myocardial infarction: rest and exercise. Am J Physiol Heart Circ Physiol 262: H411–H419, 1992. 19. Odermatt A, Becker S, Khanna VK, Kurzydlowski K, Leisner E, Pette D, and MacLennan DH. Sarcolipin regulates the activity of SERCA1, the fast-twitch skeletal muscle sarcoplasmic reticulum Ca2⫹-ATPase. J Biol Chem 273: 12360–12369, 1998. 20. Perreault CL, Gonzalez-Serratos H, Litwin SE, Sun X, Franzini-Armstrong C, and Morgan JP. Alterations in contractility and intracellular Ca2⫹ transients in isolated bundles of skeletal muscle fibers from rats with chronic heart failure. Circ Res 73: 405–412, 1993. 21. Peters DG, Mitchell HL, McCune SA, Park S, Williams JH, and Kandarian SC. Skeletal muscle sarcoplasmic reticulum Ca(2⫹)-ATPase gene expression in congestive heart failure. Circ Res 81: 703–710, 1997. 22. Pfeffer MA, Pfeffer JM, Fishbein MC, Fletcher PJ, Spadaro J, Kloner RA, and Braunwald E. Myocardial infarct size and ventricular function in rats. Circ Res 44: 503–512, 1979. 23. Simonini A, Chang K, Yue P, Long CS, and Massie BM. Expression of skeletal muscle sarcoplasmic reticulum calciumATPase is reduced in rats with postinfarction heart failure. Heart 81: 303–307, 1999. 24. Spangenburg EE, Williams JH, Talmadge RJ, Musch TI, Pfeifer PC, and McAllister RM. Changes in skeletal muscle MHC expression from rats with varying degrees of heart failure (Abstract). FASEB J 14: A265.9, 2000. 25. Sugiura T, Miyata H, Kawai Y, Matoba H, and Murakami N. Changes in myosin heavy chain isoform expression of overloaded rat skeletal muscles. Int J Biochem 25: 1609–1613, 1993. 26. Talmadge RJ, Roy RR, Chalmers GR, and Edgerton VR. MHC and sarcoplasmic reticulum protein isoforms in functionally overloaded cat plantaris muscle fibers. J Appl Physiol 80: 1296–1303, 1996. 27. Talmadge RJ, Roy RR, and Edgerton VR. Persistence of hybrid fibers in rat soleus after spinal cord transection. Anat Rec 255: 188–201, 1999. 28. Towbin H, Staehelin T, and Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA 76: 4350–4354, 1979. 29. Volterrani M, Clark AL, Ludman PF, Swan JW, Adamopoulos S, Piepoli M, and Coats AJ. Predictors of exercise capacity in chronic heart failure. Eur Heart J 15: 801–809, 1994. 30. Ward CW, Spangenburg EE, Diss LM, and Williams JH. Effects of varied fatigue protocols on sarcoplasmic reticulum calcium uptake and release rates. Am J Physiol Regulatory Integrative Comp Physiol 275: R99–R104, 1998. 31. Williams JH and Ward CW. Changes in skeletal muscle sarcoplasmic reticulum function and force production following myocardial infarction in rats. Exp Physiol 83: 85–94, 1998. 32. Williams JH, Ward CW, Spangenburg EE, and Nelson RM. Functional aspects of skeletal muscle contractile apparatus and sarcoplasmic reticulum after fatigue. J Appl Physiol 85: 619– 626, 1998. 33. Wu KD and Lytton J. Molecular cloning and quantification of sarcoplasmic reticulum Ca2⫹-ATPase isoforms in rat muscles. Am J Physiol Cell Physiol 264: C333–C341, 1993. [Corrigenda. Am J Physiol Cell Physiol 267: July 1994, following table of contents.]