Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
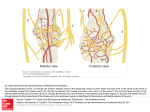
THE JOURNAL OF COMPARATIVE NEUROLOGY 384:483–500 (1997) Peripheral Development of Cranial Nerves in a Cyclostome, Lampetra japonica: Morphological Distribution of Nerve Branches and the Vertebrate Body Plan SHIGERU KURATANI,1* TATSUYA UEKI,1 SHINICHI AIZAWA,1 AND SHIGEKI HIRANO2 1Department of Morphogenesis, Institute of Molecular Embryology and Genetics (IMEG), Kumamoto University School of Medicine, Kumamoto 860, Japan 2Department of Anatomy, Niigata University School of Medicine, Niigata 951, Japan ABSTRACT The development of peripheral nerves was studied in a Japanese marine lamprey, Lampetra japonica, in whole-mount and sectioned embryos from hatching until the earliest ammocoete. Nerve fibers were immunohistochemically stained with a monoclonal antibody against acetylated tubulin. Branchiomeric nerves first developed in a simple metamerical pattern, each associated with a single pharyngeal arch. Of those, the ophthalmicus profundus, maxillomandibular, and facial nerves later developed a highly modified branching pattern, whereas postotic nerves were less specialized and showed the stereotypical branching pattern of post-trematic nerves. The early distribution of melanocytes in myotome-free space largely overlapped with the morphology of the cranial nerve and ganglion anlage, and resembled the cephalic crest cell distribution pattern in the early chick embryo. It was suggested that the cephalic crest cell distribution, which is also inhibited by myotomes in the lamprey, would be the common basis for branchiomeric nerve patterning. In later development of the lamprey embryo, myotomes 1 through 3, which had originated in the postotic region, grew rostrally into the preotic region, laterally covering all of the branchiomeric nerves. This results in a deep position of the cranial nerves, which is not observed in gnathostomes. J. Comp. Neurol. 384:483–500, 1997. r 1997 Wiley-Liss, Inc. Indexing terms: branchial region; neural crest; lampreys; embryo; mesoderm The gnathostome embryonic head is characterized by possession of two metamerisms, branchiomerism and somitomerism. The evolutionary origin of this body plan is still enigmatic because these two repetitions are not clearly separated in the nervous system of the amphioxus (e.g., Bone, 1959). To obtain insight into the primitive patterns of head metamerism, it seemed valuable as a first step to examine the developmental sequence of the pharyngeal regions in a sister group of gnathostomes. As a candidate for such animals, we chose Lampetra japonica, a marine lamprey available during their mating season in northern rivers of Japan. Lampreys used to be regarded as primitive representatives of a state between acraniate and gnathostome vertebrates (Neal, 1918a; de Beer, 1931; reviewed by Janvier, 1996. The presence of myotomes in the head region and the presence of the endostyle in the larval state were viewed r 1997 WILEY-LISS, INC. especially as plesiomorphic traits. The pre-ammocoete developmental process of the peripheral nervous system (PNS) should give us a clue to the plan of the vertebrate head morphology, but no detailed embryological studies have been performed. The description by Johnston (1905) of the peripheral morphology of the cranial nerve in Petromyzon was based on analysis of a feeding stage larva. Grant sponsor: NICHD; Grant number: N01-HD-2-3144; Grant sponsor: Ministry of Education, Science and Culture of Japan; Grant sponsor: Science and Technology Agency of Japan. *Correspondence to: Shigeru Kuratani, Department of Morphogenesis, Institute of Molecular Embryology and Genetics (IMEG), Kumamoto University School of Medicine, 4-24-1 Kuhonji, Kumamoto, Kumamoto 860, Japan. E-mail: [email protected] Received 17 September 1996; Revised 22 January 1997; Accepted 12 February 1997 484 S. KURATANI ET AL. Although spatiotemporal development of the cranial ganglia in Lampetra was also reported (Damas, 1944), the initial nerve branching pattern, which would be correlated with the primary branchiomere, has not been reported. Among the recent advances in embryology, whole-mount immunostaining has made it possible to observe the three-dimensional morphology of the nervous system (Ishikawa et al., 1986). Especially in early embryonic stages, this technique can be used to visualize the PNS, which cannot be accurately reconstructed from histological sections. Here we describe cranial nerve development in embryos of L. japonica based on whole-mount and sectioned specimens stained with several monoclonal antibodies. This is the first description of the peripheral morphology of developing cyclostome cranial nerves. At the same time, it is intended to evaluate the relevance of cyclostome morphology to understanding vertebrate head development. MATERIALS AND METHODS Embryos Adult male and female lampreys (Lampetra japonica) were collected in the Miomote River, Niigata, Japan, during the breeding season (late May through June) in 1995 and 1996. They were brought into the laboratory, where eggs were artificially fertilized and kept in fresh water at 18–23°C. Embryos were fixed twice a day with two kinds of fixatives, each for a different purpose (see below): 4% paraformaldehyde in 0.1 M phosphate-buffered TABLE 1. Comparison of Developmental Stages L. reissneri by Tahara (1988) Stage 21 Stage 22 Stage 23 Stage 24 Stage 25 Stage 26 Stage 27 Stage 28 Stage 29 Body length –3mm 3–3.5mm 4mm 5mm 6–7mm 7.5–8mm L. fluviatilis by Damas (1944) P. marinus by Piavis (1971) III–IV V VI VII–VIII IX–X XI XII–XIII XIV XV 12 13 13 13–14 14 15 15 16 18 saline (PFA/PBS) and Bouin’s fixative. Because development of the embryos was significantly affected by temperature, they were staged morphologically. The developmental sequence of the L. japonica embryos is similar to that of the brook species, L. reissneri, so that staging reported by Tahara (1988) for the latter was used in the present description. The comparison of developmental sequences between L. reissneri, L. fluviatilis (Damas, 1944), and Petromyzon marinus (Piavis, 1971) is listed in Table 1. For nervous elements, the terminology of Alcock (1898), Johnston (1905), Lindström (1949), Ronan and Northcutt (1987), and Koyama et al. (1990) was used. Whole-mount immunostaining For whole-mount immunostaining, embryos were prepared as described (Kuratani and Eichele, 1993) with minor modifications. After fixation with PFA/PBS at 4°C Abbreviations a3 bc bv Cc dlt dsr eb ebm ebt ep es eso fb fg gldix gldx1 glf glv2,3 hb hbm hm hy int io llp m ma mb mbc mc me mp mvl n olep ot p1–p8 a6 pharyngeal arches branchial arch cartilage branchial arch vein circumpharyngeal crest cells in the chick embryo dorsolateral fasciculus dorsal spinal nerves eyeball epibranchial muscles epibranchial tract of the vagus nerve epiphysis endostyle esophagus forebrain foregut epibranchial ganglion of the glossopharyngeal nerve epibranchial ganglion of the first vagus nerve facial ganglion maxillomandibular nerve ganglion hindbrain hypobranchial muscles hyomandibular nerve hyoid arch intestinal ramus of the vagus nerve infraoptic nerve lower lip myotomes adductor muscle of the branchial arch midbrain buccal muscle constrictor muscle external mandibular ramus muscle plate of the branchial arch ventral lip muscle notochord olfactory epithelium otocyst pharyngeal pouches pc ph pllg plln plsp pnd prtx1 ps4 rb rbc rdix rc rm rmp rpvii rrec rthyr rv rvel sc so s1–s8 tb ulp vel vsr ws II V V1 V2,3 VII IX X x1–x6 XII posterior commissure pharynx posterior lateral line ganglion posterior lateral line nerve spinal nerve plexus pronephric duct pre-trematic ramus of X1 fourth pharyngeal slit buccal ramus of the facial nerve buccinator ramus of the facial nerve dorsal ramus of the glossopharyngeal nerve communicating branches of vagus nerves maxillary ramus of the maxillomandibular nerve posterior muscular rami of the pharyngeal ramus pharyngeal ramus of facial nerve recurrent nerve thyroid ramus of the facial nerve visceral rami of vagus nerves velar ramus of the maxillomandibular nerve sclerotome supraoptic nerve somites taste bud upper lip velum ventral spinal nerves ciliary string of the branchial arch optic nerve trigeminal nerve anlage in gnathostome embryos ophthalmicus profundus nerve maxillomandibular nerve facial nerve glossopharyngeal nerve vagus nerve first to sixth vagal branchial branches hypoglossal nerve CRANIAL NERVE DEVELOPMENT IN THE LAMPREY 485 Fig. 1. Microphotographs of the whole-mount stage 25 embryos stained with anti-acetylated tubulin antibody. A: All the branchiomeric nerve roots have developed showing metamerical organization. Note the presence of spinal motor nerves (arrowheads). Pronephric ducts (pnd) are also stained with the antibody. B: Enlargement of a head portion of A. Position of the otocyst is marked by a circle. Each branchiomeric nerve trunk is oriented rostrally toward the dorsal tip of the pharyngeal pouches (p1–4). Proximally, the nerves are arising from the dorsolateral fasciculus (dlt) of the hindbrain. Arrowheads indicate the independent development of ophthalmic profundus nerve root. The vagus nerve (X) passes between myotomes 2 (m2) and 3 (m3). The arrow points to the communication between the facial and maxillomandibular nerves. Scale bars 5 100 µm. for 1 day, embryos were washed in 0.9% NaCl/distilled water, dehydrated in a graded methanol series (50%, 80%, 100%), and stored at 220°C. The samples to be stained were placed on ice in 2 ml dimethyl sulfoxide (DMSO) methanol (1:1) until they sank. Five-tenths milliliter of 10% Triton X-100/distilled water was added, and the embryos were further incubated for 30 minutes at room temperature. After washing in TST (TST is Tris-HCl-buffered saline: 20 mM Tris-HCl, pH 8.0, 150 mM NaCl, 0.1% Triton X-100), the samples were sequen- 486 S. KURATANI ET AL. Fig. 2. Morphological pattern of myotomes and cranial nerves. Myotomes (numbered) are superimposed over a stage 25 embryo. The glossopharyngeal nerve root develops medial to myotome 1. The vagus nerve (X) passes between the second and third myotomes. Scale bar 5 100 µm. Fig. 3. Graphical illustration of the neurite development at stage 26 of L. japonica based on a whole-mount embryo. Ventral (vsr) and dorsal spinal nerve (dsr) anlagen are developing in an alternating segmental pattern. The distal portion of the maxillomandibular nerve (V2,3) has been bifurcated into two portions, one innervating the upper lip (ulp) and the other (rvel) the velum. The main branch of the facial nerve (VII), the hyomandibular nerve (hm), innervates the lower lip (llp), showing the latter structure is a derivative of the hyoid arch. The vagus nerve (X) has branched to the pre-trematic ramus (prtx1) in the first branchial arch and a post-trematic nerve (x1) in the second branchial arch. Caudally, the vagus nerve develops a bundle of nerves, the epibranchial trunk (ebt). Dorsally, the posterior lateral line nerve (plln) is independent of the vagus nerve anlage. Note the development of independent bundles of fibers (asterisks) found close to the surface ectoderm. One of these is the earliest anlage of the dorsal ramus (rdix) of the glossopharyngeal nerve. Scale bar 5 100 µm. tially blocked with aqueous 1% periodic acid and with 5% nonfat dry milk in TST (TSTM). For whole-mount immunostaining of the cranial nerves, a monoclonal antibody raised against acetylated tubulin (monoclonal anti-acetylated tubulin, No. T-6793, Sigma Chemical Company, St. Louis, MO) was found to be most suitable. Embryos were incubated in the primary antibody (diluted 1/1,000 in spin-clarified TSTM containing 0.1% sodium azide) for 2 to 4 days at room temperature while being gently agitated on a shaking platform. The secondary antibody used was horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (ZYMED Lab. Inc., San Francisco, CA) diluted 1/200 in TSTM. After a final wash in TST, the embryos were preincubated with peroxidase substrates 3,38-diaminobenzidine (DAB, 100 mg/ml) in TS (20 mM Tris-HCl, pH 8.0, 150 mM NaCl) for 1 hour and allowed to react in TS with the same concentration of DAB with 0.01% hydrogen peroxide for 20 to 40 minutes at 0°C. The reaction was stopped and simultaneously embryos were clarified with 30% glycerol in 0.5% KOH. The stained embryos were transferred to a 60% glycerol/water mixture and mounted on glass depression slides for observation. CH-1 (purchased from the Developmental Studies Hybridoma Bank, Iowa City, IA), which recognizes tropomyosin, was used to detect the development of myotomes. Hardisty and Rovainen (1971) was referred to for the anatomical identification of muscles. Fig. 4. Photomicrographs of whole-mount embryos of L. japonica at stage 26. A,C: Lateral aspects of the embryos. The embryo in C is slightly older than those in A. B,D: Enlargement of the boxes in A and C, respectively. In B and C, the myotomes are illuminated by polarized light. Arrowheads in B indicate the rostral and caudal surface of myotome 2. The vagus nerve (X) develops an arch-shaped nerve trunk (B,C). In slightly older specimens (D), late-forming vagal nerve fibers can develop along the boundaries of myotomes (arrows), indicating that the myotomes inhibit the neurite growth. Scale bars 5 100 µm. Fig. 5. Sectioned specimens of a stage 26 embryo stained with HNK-1 (an antibody that stains the nervous system). A: At the level of the first arch. The maxillomandibular ganglion (glv2,3) is situated beneath the surface ectoderm. B: At the level of the otocyst. Myotomes (m) cover the otocyst (ot) and the petrosal ganglion (gldix). C: Sclerotomes (sc) and myotomes (m) are being separated at this level. The root and the epibranchial ganglion of the vagus nerve (X) are located medial to the myotome. D: Trunk level. The myotome is not separated from the sclerotome at this level. The vagal epibranchial ganglion (gldix) lies ventral to the myotome. The posterior lateral line nerve (plln) is located medial to the dorsomedial edge of the myotome (arrow). Scale bar 5 100 µm. CRANIAL NERVE DEVELOPMENT IN THE LAMPREY 489 Fig. 6. Schematic diagram to show the topographical relationships between the nervous system and mesodermal components at stage 26. Oral cavity and pharyngeal pouches are indicated by dashed lines. Owing to the growth of the myotomes, the postotic nerve trunks and ganglia are covered by myotomes. Based on the same embryo shown in Figure 5 (A–D indicate the planes in which the sections were cut). Scale bar 5 100 µm. Immunostaining on sectioned specimens RESULTS Specimens fixed in Bouin’s were embedded in paraffin and sectioned at 5 µm. They were deparaffinized and treated with 1% periodic acid for 5 minutes at room temperature, followed by washing in TST. Monoclonal antibodies T-6793, as well as HNK-1 (Leu-7, Becton Dickinson, San Jose, CA), were used to label the developing nervous system on the paraffin-sectioned specimens. Primary antibodies were diluted in TSTM and applied to the sections for 1.5 hours at room temperature. After washing with TST, secondary antibodies, HRP–anti-mouse IgG (for those labeled with T-6793) and IgM (for those labeled with HNK-1) (ZYMED Lab. Inc.), were diluted 1/200 in TSTM and applied to the specimens for 40 minutes. The sections were counterstained either with cresyl violet or hematoxylin after the peroxidase reaction. The following description of our results deals with the peripheral nervous system (PNS) morphology after stage 25. The earlier development of the nerves will be described elsewhere (Kuratani et al., unpublished observation). Briefly, the facial nerve root anlage first develops at early stage 23 (stage 23-), followed by the maxillomandibular nerve root that appears at stage 23. The ophthalmicus profundus nerve root is first recognized late in stage 23 (stage 231) to stage 24. Morphology of branchiomeric nerves Stage 25 (Figs. 1, 2). By stage 25, the four anteriormost pharyngeal pouches (pharyngeal pouches 1 through 4) are visible, and the otic vesicle has formed. The ophthalmicus profundus and maxillomandibular nerves arise 490 S. KURATANI ET AL. Fig. 7. Developing peripheral nerves in stage 27 embryos. A: Graphical illustration of the neurite development based on a wholemount embryo. All the major branches have appeared by this stage. The dorsal ramus of IX (rdix) is connected with the main trunk of the glossopharyngeal nerve (IX). The thyroid ramus (rthyr) that passes caudally in the pharyngeal floor has branched from the facial nerve. The overall configuration of the peripheral nervous system shows separation between the branchiomeric and somitomeric series of nerves. Note that the main trunk of the vagus (X) and hypoglossal nerves (XII) passes along the interface between the somitic and branchiomeric regions, the circumbranchial region. B: Photomicrograph of a different embryo at the same stage. Scale bars 5 100 µm. from the hindbrain with separate roots from the area corresponding to rhombomere 2 at the preceding stage (Fig. 1). The maxillomandibular nerve extends several branches in front of the first pharyngeal pouch or within the mandibular arch. The facial nerve arises from the hindbrain, medial to the otocyst and is directed rostrally toward the dorsal end of the first pharyngeal pouch. Dorsal to the pouch, the facial nerve makes a connection with the maxillomandibular nerve. Caudal to the otocyst, the glossopharyngeal nerve arises from the hindbrain at the level of the third pharyngeal pouch (Fig. 1). Curiously, this nerve lies medial to myotome 1 (Fig. 2). More caudally, the vagus nerve trunk has developed and passes between myotomes 2 and 3 (Figs. 1B, 2). The posterior lateral line nerve primordium is present, lateral to the dorsolateral fasciculus of the hindbrain and caudal to the otocyst, with its rostral end close to the vagus nerve root. In the trunk region, this nerve is found on the dorsomedial edge of myotomes (see Fig. 4D). Stage 26 (Figs. 3–6). Pharyngeal pouch 8 appears in some embryos by stage 26. The ophthalmicus profundus nerve extends its processes along the lateral aspect of the forebrain. The maxillomandibular nerve possesses two obvious branches by this stage, one innervating the upper lip (maxillary ramus) and the other the velum (velar ramus). Lateral to the facial nerve, there develops a fine bundle of fibers, the buccal nerve, that belongs to the anterior lateral line nerve. Rostrally, the buccal nerve extends lateral to the maxillomandibular nerve and ventral to the ophthalmicus profundus nerve. The main branch of the facial nerve has formed the ramus hyoideus within the second pharyngeal arch. The main branch of the glossopharyngeal nerve, the post-trematicus nerve, has grown into the first branchial CRANIAL NERVE DEVELOPMENT IN THE LAMPREY 491 Fig. 8. Semi-diagrammatic illustration of a stage 28 embryo. A: The whole head of the embryo. Terminal branches of facial (me), glossopharyngeal, and vagus (x1–x6) nerves are innervating lateral line organs which are shown by circles. Note the extensive plexus formed of rostral spinal nerve roots (plsp). From the plexus two branches (supraoptic nerve 5 so and infraoptic nerve 5 io) arise and pass rostrally to innervate myotomes 1 through 3 (see Fig. 10). B: Enlargement of the branchiomeric nerve roots, showing the passage of putative motor fibers within the ganglion. Note the development of the recurrent nerve (rrec) connecting the facial (glf) and posterior lateral line (pllg) ganglia. Arrow indicates the communication with the spinal nerve plexus (plsp in A) which is cut. Scale bars 5 100 µm. arch by stage 26. A lateral line nerve is often found close to the glossopharyngeal nerve, provisionally called the dorsal ramus (Fig. 3). The epibranchial ganglion of the glossopharyngeus lies medial to the myotome (Figs. 5, 6). The vagus nerve is an arch-shaped bundle of fibers, consisting of a dorsal part representing the proximal root and a ventral part representing the epibranchial tract (Fig. 3). As a whole, this nerve forms an arch that overhangs slightly rostral to the myotome 2⁄3 border (Fig. 4). Late-forming nerve fibers can develop along this border (Fig. 4D). No nerve branches are found on the surface of the myotome. The proximal root passes through the posterior lateral line ganglion medial to the dorsolaterally expanding myotomes (Fig. 6). Along the epibranchial tract of the vagus, sensory ganglion primordia are found between each pharyngeal pouch, ventrolateral to the myo- tome (Figs. 5, 6). Ventrally the vagus nerve possesses the post-trematic ramus of x1 within the second branchial arch and sometimes grows the post-trematic ramus of x2 and other posterior branches in slightly older embryos (Fig. 3). A pre-trematic ramus can accompany the nerve x1 (Fig. 3). Stage 27 (Fig. 7). The ophthalmicus profundus nerve passes dorsal to the eye by stage 27, and the ophthalmic ganglion is separated from the maxillomandibular ganglion as is the root. The maxillary ramus of the maxillomandibular nerve innervates the anterior buccal muscle in the upper lip (not shown). Associated with the facial nerve, a sensory ganglion primordium is visible in the whole-mount (Fig. 7). In its relation to the buccal nerve, this seems to represent the anlage of the anterior lateral line ganglion in the adult 492 S. KURATANI ET AL. Fig. 9. Photomicrographs of the same stage 28 embryo as in Figure 8. A: Trigeminal innervation area. Ganglia are marked by asterisks. The ophthalmic profundus (V1) and maxillomandibular (V2,3) nerves still possess two separate nerve roots on the hindbrain. Note that in the maxillomandibular nerve the neurite bundle bifurcates within the maxillomandibular ganglion (arrows) and passes into the maxillary (rm) and velar (rvel) branches. B: Nerve plexus formed of several rostralmost ventral spinal nerves (arrowheads). C: The intestinal ramus (int) of the vagus nerve. The intestinal tract is indicated by broken lines. Note the dorsal position of the ramus in relation to the esophagus (eso) and foregut (fg). Scale bar 5 100 µm. (Ronan and Northcutt, 1987; Koyama et al., 1990). The intracapsular or otic ganglion was not clearly identified in the present study; the ganglion associated with the facial nerve is, therefore, simply referred to as the facial ganglion in the discussion below. The ventral part of the hyomandibular ramus has extended several smaller branches into the lower lip. They correspond to the external mandibular ramus anlage that primarily belongs to the lateral line nerve. The distal portion of the hyomandibular nerve, or the thyroid ramus, grows caudally traversing the ventral aspect of the pharynx. As the rostral vagus nerve root develops, it becomes continuous rostrally with the glossopharyngeal nerve root. The posterior part of the root remains separated from the glossopharyngeo-vagal root. In many embryos the vagus nerve possesses six post-trematic branches (x1–x6). The pre-trematic ramus of x1 has disappeared by this stage. Stage 28 (Figs. 8–12). Because the monoclonal antibody T-6793 does not stain the perikarya at stage 28, it is often possible in the whole-mount embryo to identify rami-containing motor fibers as a non-bifurcating bundle penetrating the ganglion (Figs. 8B, 9A). The ophthalmicus profundus nerve appears to be composed of sensory fibers. In the maxillomandibular nerve, the maxillary nerve has grown a medial branch, the buccinator ramus (Fig. 8A), that passes caudally into the buccal floor. The buccal ramus of the facial nerve appears not to contain any motor fibers, consistent with the adult anatomy (Koyama et al., 1990). Our observations suggest that the glossopharyngeal sensory fibers participate in this ramus (Fig. 8B). Most of the motor fibers passing through the facial ganglion originate from a more ventral level of the hindbrain than that of the sensory root (Fig. 8B), and they pass mainly into the thyroid branch, more or less separated from the external mandibular ramus (Fig. 8B). The latter ramus arises directly from the facial ganglion and would also be sensory; the nerve bifurcates distally and some of the nerve bundles terminate on the lateral line organs (Fig. 8A). From the more proximal portion of the hyomandibular CRANIAL NERVE DEVELOPMENT IN THE LAMPREY 493 Fig. 10. Morphology of the myotomes at stage 28. Myotomes (numbered) are superimposed over a stage 28 embryo. Note that rostral myotomes (1 to 3) which developed postotically have grown rostrally into the preotic region. The hypobranchial muscle (hbm) is developing along the course of the hypoglossal nerve. Scale bar 5 100 µm. ramus, a posteriorly oriented ramus, provisionally called the posterior pharyngeal ramus, arises and grows along the dorsal aspect of the pharynx (Fig. 8B). The recurrent nerve is constantly present; the nerve communicates between the facial and the posterior lateral line ganglion, and also communicates with the dorsal ramus of the glossopharyngeal nerve lateral to the otocyst (Fig. 8A,B). Within the posterior lateral line ganglion, the vagus nerve rootlets pass caudal to the glossopharyngeal nerve root. The latter is also located within the ganglion (Fig. 8B). The intestinal ramus arises from the caudal portion of the epibranchial tract of the vagus and passes caudally along the dorsal aspect of the esophagus and foregut (Fig. 9C). The epibranchial tract of the vagus has established an extensive communication with the hypoglossal nerve by this stage. The latter nerve has invaded the ventrolateral aspect of the pharyngeal wall. Simultaneously, the hypobranchial muscle anlage appears in the ventrolateral aspect of the pharyngeal wall which later divides into repeating segments (Fig. 10). Myotomes 1 and 2 grow rostrally, covering laterally all of the cranial nerves in the preotic region (Figs. 10, 11). The rostral group of ventral spinal nerves form an extensive nerve plexus (Fig. 9B) and send out two nerve branches to innervate myotomes 1 through 3, one dorsal to the eye (the supraoptic nerve), and the other ventral to the eye (the infraoptic nerve) (Figs. 8A, 10). Due to the position of the muscles they innervate, these spinal nerve branches are located lateral to the dorsal ramus of IX (Fig. 8A). The distribution pattern of melanocytes, probably of neural crest origin, first becomes apparent at this stage and largely overlaps with the PNS elements (Fig. 12). The melanocytes generally fall into two categories: those surrounding the central nervous system (CNS), and more compact cell populations in the pharyngeal area. The latter is further divided into three major cell populations, i.e., the anterior population within the upper lip and in the prechordal region, the second within the hyoid arch, and the posterior in the postotic branchial arch region (Fig. 12). The posterior cell population is stacked between epibranchial myotomes and the pharyngeal arches forming a compact longitudinal cell strand corresponding to the epibranchial tract of the vagus nerve. At the level of epibranchial ganglia, this cell strand shows local swellings (Fig. 12B). Slightly later, streams of melanocytes from this cell strand extend ventrally to fill each branchial arch (Fig. 12B). It could not be determined, however, whether this was due to the migration of the cells or delayed differentiation of the ectomesenchyme that already populated the arches. Morphology of branchial branches (Fig. 13) Caudal to the glossopharyngeal nerve, the branchiomeric nerve branches of L. japonica have similar morphology in each branchial arch. Morphology of the branches is closely associated with the branchial arch. In histological sections of a stage 26 embryo, a typical branchial arch of L. japonica is first seen as a simple, dorsoventral plate of the pharyngeal wall, successively delineated by endodermal indentations which are the pharyngeal pouches (Fig. 13A). Each arch contains a muscle plate that occupies the caudal aspect and a cartilage bar in the rostralateral portion. Each post-trematic nerve passes between the muscle plate and the cartilage (Fig. 13A). The branchial arch grows in the rostromedial direction at stage 27 and the muscle plate also grows in the same direction (Fig. 13B, C). All of the arches now contain the post-trematic nerve, which extends small branches posteriorly and also produces ventral communicating branches (Fig. 7). The posterior ramus innervates the constrictor muscle, the derivative of the posterior part of the muscle plate (Fig. 13D). There is also a rostrally oriented visceral ramus issuing from the post-trematic nerve (Figs. 8A, 13D). 494 S. KURATANI ET AL. Fig. 11. Development of the myotome in late embryos. A: Photomicrograph of a stage 28 embryo. A whole-mount embryo stained with anti-acetylated tubulin antibody. Myotomes are simultaneously illuminated with polarized light. Note the rostral growth of rostral myotomes (m2 and m3) that have divided into ventral and dorsal halves on dorsal and ventral sides of the eye. The epibranchial tract (ebt) with epibranchial ganglia develops between the myotomes and pharyngeal arches, and the posterior lateral line nerve (plln) along the dorsal edge of the myotomes. B: Stage 30 embryo stained with CH-1 (tropomyosin) antibody. The hypobranchial muscle (hbm) has been divided longitudinally into pseudometamerical segments, that are associated neither with true branchiomerism nor with epibranchial myotomes (ebm). Note the muscles that are present in the upper lip (buccal muscle, mbc) and in the lower lip (mvl). Scale bars 5 500 µm. Perforation of the pharyngeal slits is completed by stage 29 (Fig. 13E). The visceral ramus innervates the rostromedial subdivision of the branchial arch, the ciliary string (Fig. 13E), where the nerve fibers are associated with taste buds (Fig. 13E). The anterior subdivision of the muscle plate, the adductor muscle, is also associated with the visceral ramus (Fig. 13D). By stage 29 the post-trematic trunk has shifted to the lateral aspect of the branchial arch cartilage. The main trunk of the post-trematic nerve grows farther ventrally and bifurcates into several small branches which are mostly innervating lateral line organs in the ventral aspect of the pharynx (Fig. 8A). DISCUSSION Branchiomeric nerve morphology Through the developmental stages examined, the ophthalmicus profundus nerve of L. japonica develops independently from the maxillomandibular nerve (Figs. 1, 3, 7, 8). In the adult L. japonica, however, both of the roots are fused to form a single root as seen in many gnathostomes (Koyama et al., 1987). This indicates that the separate profundus nerve configuration is an embryonic feature, which has also been regarded as a plesiomorphic character (reviewed by Goodrich, 1930). CRANIAL NERVE DEVELOPMENT IN THE LAMPREY 495 Fig. 12. Distribution pattern of melanocytes in L. japonica. A: The lateral view of a stage 28 embryo. Anterior is to the left. Melanocytes form three separate populations, the rostral (arrowheads), the hyoid arch (hy), and the glossopharyngeo-vagal or caudal cell populations. A small number of cells surround the neural tube. The caudal cell population is accumulated along the interface between myotomes (illuminated with polarized light) and pharyngeal arches. B: Stage 29. Anterior is to the right. From the accumulation of melanocytes along the epibranchial tract, some subpopulations migrated into each pha- ryngeal arch (a3–a6). Arrows indicate the position of vagal epibranchial ganglia surrounded by melanocytes. The arrowhead indicates the accumulation of melanocytes between the maxillomandibular ganglion and facial ganglion. C: HNK-1-stained section of an embryo at stage 29 at the level of the second pharyngeal arch. Dark cells along the arch are melanocytes (arrows). Note that the cells surround the pharynx (ph) and the hindbrain (hb) as well as the notochord (n). Scale bar 5 200 µm for A, B, 100 µm for C. As found in other aquatic forms (reviewed by Haller v. Hallerstein, 1934), branches of glossopharyngeal and vagus nerves of the lamprey appear to be in a low state of specialization, each being similar to the others (Fig. 8A). The main ramus of a typical branchiomeric nerve is the post-trematic nerve. Unlike some aquatic gnathostomes (Tanaka, 1976, 1979; Tanaka and Nakao, 1979), however, pharyngeal and pre-trematic nerves are not found in all the stages examined, except for a temporary pre-trematic branch associated with x1 (Fig. 3). Each branchial arch of L. japonica is subdivided into anteromedial and posterolateral portions and so is the branching pattern of the post-trematic nerve. The visceral ramus innervating the anteromedial part, or the ciliary string, presumably contains gustatory as well as motor fibers. Morphologically, no equivalent branching was found in the maxillomandibular or facial nerves, although gustatory fibers have been reported in this nerve of the lamprey by Fritzsch and Northcutt (1993). The posterolateral portion of the branchial arch is innervated by the posterior ramus. A ramus of equivalent nature would be found in the velar ramus of the maxillomandibular and in the thyroid ramus of the facial nerve (Fig. 8A). The post-trematic ramus bifurcates distally to terminate on lateral line organs (Fig. 8A). The external mandibular ramus of the facial nerve, which primarily belongs to the anterior lateral line nerve in gnathostomes, may be of a similar nature (Fig. 8A,B). Each epibranchial ganglion of the vagus nerve in Petromyzon planeri sends out a minor ramus dorsal to each gill opening, innervating epibranchial sense organs (Dohrn, 1888; Alcock, 1898). In L. japonica, similar rami were found to arise from the epibranchial tract and terminate on the lateral line organs by stage 29 (not shown). These rami were barely visible in younger embryos. The posterior ramus of the maxillomandibular nerve that innervates the velothyroid muscle in the velum has been termed the velar ramus (Figs. 3, 7, 8). This nerve apparently resembles the mandibular ramus of the maxillomandibular nerve of gnathostomes. However, the other ramus of the maxillomandibular nerve of L. japonica, the maxillary branch, also contains motor fibers and is distributed in the upper lip. Johnston (1905) thought the maxillary branch contained a part of the mandibular ramus in other fishes; i.e., both the rami appear to correspond to gnathostome mandibular ramus. Immunocytochemical data are consistent with these results. Engrailed proteinlike immunoreactivity has been detected in the mandibular arch muscle of gnathostome embryos (Hatta et al., 1990). In the lamprey, similar immunoreactivity has been detected in the muscle anlage of the upper lip and of the velum, both cheek process (mandibular process) derivatives (Holland et al., 1993). The cheek process which contains muscle anlage is probably equivalent to the mandibular process of gnathostome embryos. This further implies that the maxillary portion (or prominence) of the mandibular arch is an apomorphic structure of gnathostomes. Figure 13 CRANIAL NERVE DEVELOPMENT IN THE LAMPREY 497 Branchiomeric and somitomeric nerves Before the addition of the anterior lateral line nerves, initial branchiomeric nerve anlagen of L. japonica clearly develop on the segmental scheme of branchial arches (Fig. 1). At these stages, the placodal contribution was apparent in the epibranchial ganglia of the vagus and glossopharyngeus, and also of the facial nerves (not shown). These ganglia appear to also follow the branchiomeric developmental scheme. A more detailed study of cranial gangliogenesis in L. japonica is now in progress. Since Balfour (1878), it has been argued that mesodermal segments exist in the vertebrate head region, similar to somites in the trunk. The heads of amniote embryos show incomplete mesodermal segments called somitomeres (reviewed by Jacobson, 1993; also see Freund et al., 1996) whose metamerical and developmental correlation with the head cavities of chondrichthys remains unknown. This line of ideas has primarily been based on the hypothesis that there should exist the same number of mesodermal segments, each being associated with a set of single pharyngeal arch-derivatives (van Wijhe, 1882; Goodrich, 1930; Jarvik, 1980). However, the mesodermal metamerism, the somitomerism, does not always coincide with metamerism of gills, the branchiomerism. It has been observed during development of several chordate species, including the lamprey embryo, that branchial arches and myotomes develop at different rates and intervals (Damas, 1944; reviewed by Jefferies, 1986); this was also ascertained in the present study (see below). A group of morphologists thus proposed that somitomerism and branchiomerism are two separate metamerisms, which leads to the dual segmental theory of the vertebrate body (reviewed by Romer, 1972). Metamerism of the vertebrate PNS is prefigured by the distribution pattern of crest cells (Noden, 1975; Loring and Erickson, 1987). Primarily based on observations in the chick embryo, the migration pathways of amniote crest cells have been classified into two major categories, the dorsolateral and the ventral (reviewed by Le Douarin et al., 1984). The ventral pathway is dominant in the trunk region of gnathostome embryos and provides the basis for somitomeric development of spinal nerve formation (Detwiler, 1934; Keynes and Stern, 1984); both the motor root and gangliogenic crest cells are metamerically patterned by somites. Thus, the different nature of branchiomerism Fig. 13. Branching pattern of post-trematic nerves. A: Horizontal section of the stage 26 pharynx stained with the anti-acetylated tubulin antibody. The post-trematic ramus (arrows) grows between the branchial arch cartilage (bc) and the muscle plate (mp). B,C: Stage 27 pharynx at the vagal level. Post-trematic nerves are marked by large arrows in C. Developing rami are labeled with small arrows in C and arrows in B. Note that two groups of rami, rostral and caudal, are issuing from the post-trematic nerve. D: Whole-mount embryo at stage 28, showing the branching pattern of vagal postotic nerves. Muscle fibers are illuminated with polarized light. The posterior ramus (rmp) innervates the constrictor muscle (mc) and the visceral ramus (rv), the adductor muscle (ma) developing within the ciliary string (ws in E). Asterisks indicate vagal epibranchial ganglia. E: Horizontal section of the pharynx at the first vagal segment in a stage 29 embryo. Immunochemically stained nerve fibers are marked with arrowheads and the post-trematic nerve of X1 by an arrow. Note the taste bud (tb) developing at the rostral edge of the ciliary string (ws). Scale bars 5 50 µm in C for A–C and E, 50 µm in D. Fig. 14. Comparative morphology of the cranial nerve anlage in vertebrate embryos. Ganglion primordia of two gnathostome species are illustrated. For comparison with lamprey embryos, see Figure 2. Note in both that somites inhibit development of the cranial nerve anlage. As a result, the initial anlage of the vagus nerve forms an arch along one of the anterior somites. A few anteriormost somites (first to fifth somites in the shark, the first somite in the chick) allow the development of the vagus nerve lateral to themselves. Note the similar distribution pattern of melanocytes in the lamprey (compare with Fig. 12). The circumpharyngeal crest cells (Cc) in the chick occupy the postotic region beneath epibranchial myotomes, which corresponds to the site of the epibranchial tract in the lamprey. The embryos are not drawn to scale. Modified from Goodrich (1930) and Kuratani (1997). and somitomerism is exemplified by developmental organization of the PNS. Metamerical morphology of the lamprey spinal nerves implies somite-dependent regulation, although the developmental process would be very different from that of the chick. Ventral spinal nerves of the lamprey develop at mid-somitic levels and dorsal spinal nerves at intersomitic levels (reviewed by Goodrich, 1937). The dorsolateral pathway characterizes the cephalic crest cell distribution patterns; the cells pass beneath the surface ectoderm and fill the pharyngeal arch ventrally (reviewed by Bronner-Fraser, 1995). Sensory ganglia of branchiomeric nerves are formed along this pathway (Noden, 1975). Description of the crest cell migration is very limited in lampreys (e.g., Nakao and Ishizawa, 1987). Most of the classical works on this issue are based on graphic reconstruction of histological sections (e.g., Veit, 1939; Damas, 1944), and no lineage markers have been utilized. Demonstrations of the detailed distribution pattern of crest cells in early embryos would necessitate vital labeling, which belongs to a future project. At this time, several pieces of circumstantial evidence suggest the close relationship between the crest cells and PNS morphology. Fig. 15. Evolution of the peripheral nervous system (PNS) developmental pattern—a hypothesis. Both in lampreys and gnathostomes, postotic myotomes inhibit the cranial nerve development. In the lamprey, the vagus nerve first develops between myotomes 2 and 3 (the glossopharyngeal nerve exceptionally develops medial to the first myotome). In later development, the secondary rostral growth of myotomes 1 through 3 and of hypobranchial muscle covers the entire PNS laterally. This does not take place in gnathostome development. The origin of the lamprey hypobranchial muscle does not actually reflect the number of segments. In terms of primary arrangement of the paraxial mesodermal component, the pattern of cranial nerve development in gnathostomes is less deviated than in the lamprey. CRANIAL NERVE DEVELOPMENT IN THE LAMPREY 499 The distribution pattern of the melanocytes in L. japonica implies that cephalic crest cells distribute in a pattern similar to that in gnathostomes; the three melanocyte populations observed at stage 28 are strikingly reminiscent of the crest cell distribution pattern in early chick embryos (Fig. 14). Interestingly, such a pattern of melanocyte distribution partly coincides with the morphology of the cranial nerves; the anterior cell population overlaps the ophthalmicus profundus and maxillomandibular nerve innervating region, the middle coincides with the facial nerve, and the caudal cell strand colocalizes with the glossopharyngeo-vagus as well as the hypoglossal nerves. The caudal melanocyte population is very similar to the caudalmost cephalic crest cell population called circumpharyngeal crest cells in the chick (Kuratani and Kirby, 1991, 1992). In both animals, the postotic epibranchial ganglia develop in this location ventral to epibranchial somites. In the fully grown ammocoetes larvae of Petromyzon, Johnston (1905) described a composite nerve bundle containing the epibranchial tract and spinal nerve elements. However, the vagus and hypoglossal nerves of L. japonica develop primarily independently from each other (Fig. 7), and an extensive communication between the two nerves is only secondarily established (Fig. 8). The hypoglossal nerve of L. japonica is solely composed of spinal motor nerves as in other vertebrates, and it innervates the hypobranchial muscles derived from somites (Neal, 1897), i.e., belonging to the somitomeric part of the body. Both the hypoglossal nerve and muscle circumnavigate the pharynx dorsocaudally, thereby making an anteriorly opened arch (Fig. 8A) representing the head/trunk interface. This morphology is common to all gnathostome embryos at the pharyngular stage of development (Kuratani, 1997 and references therein; Fig. 15). topographical relationships with nerves, somite 2 of the chick embryo resembles myotome 3 of L. japonica. The similarity of the cranial nerve configuration between lampreys and gnathostomes is seen only at the early stages of development, before the secondary modification by myotomes. Such morphology can be characterized as the coexistence of branchiomerism and somitomerism in the postotic region; both make an S-shaped interface. From an evolutionary point of view, cranial nerve and mesodermal development in gnathostome embryos may maintain a plesiomorphic or less-derived configuration than the lamprey embryo (Fig. 15). The arched shape of the vagus and hypoglossal nerve exemplifies the head/trunk interface, or the interface between two distinctive metamerisms, each of which has an independent patterning mechanism. The alternating metamerical pattern of the peripheral nerve (spinal dorsal nerve) in amphioxus is suggestive of myotome-dependent axonal growth (reviewed by Franz, 1927). However, they have not attained the arched shape in any of the nerves, i.e., the amphioxus probably lacks the distinction between the branchiomerism and somitomerism in the nervous system. The lamprey does not represent the intermediate state between the acraniates and gnathostomes: Preotic pharyngeal arches are highly modified, and dual metamerism is completely established. Possession of S-shaped nerve morphology, therefore, serves as a synapomorphy that establishes either Vertebrata or Craniata. It now remains to examine whether or not myxinoids, the sister group of vertebrates, show PNS morphogenesis reflecting the most basic plan of the vertebrate body—the dual metamerism. Myotomes and cranial nerves We are grateful to Nobutoshi Maeda and Miyuki Yamamoto in the Institute for Laboratory Animals, Niigata University School of Medicine, for maintenance of animals. We also thank Dr. Hiromichi Koyama for critical reading of the manuscript. Sincere gratitude is extended to Dr. Glenn Northcutt for his valuable comments on the manuscript. The CH-1 antibody developed by Dr. J. Lin was obtained from the Developmental Studies Hybridoma Bank maintained by the Department of Pharmacology and Molecular Sciences, Johns Hopkins University School of Medicine, Baltimore, MD, and the Department of Biological Sciences, University of Iowa, Iowa City, under contract N01-HD-2-3144 from the NICHD. This work was supported by grants-in-aid from the Ministry of Education, Science and Culture of Japan (Specially Promoted Research) and the Science and Technology Agency of Japan. ACKNOWLEDGMENTS The most intriguing difference in the peripheral morphology of cranial nerves between lampreys and gnathostomes is the primary topographical relationship between the nerves and myotomes (Figs. 2, 13). In gnathostome development, branchiomeric nerve anlagen are essentially located superficially. This condition lasts well beyond the state of pharyngula (Kuratani, 1997 and references therein). In the lamprey, in contrast, some rostralmost myotomes grow rostrally and entirely cover the cranial nerves laterally by stage 28 (Figs. 2, 10, 11). Obviously, the myotomes found in the preotic region at stage 28 originate from postotic somites that have nothing to do with the preotic head cavities or head somites illustrated by Neal (1918b). Earlier development of myotomes in relation to the cranial nerves will be described elsewhere. Dermomyotomes with the exception of several rostralmost ones, have been shown in the chick embryo to inhibit the dorsolateral migration of crest cells, and subsequently cranial nerve formation (Oakley et al., 1994; Kuratani and Aizawa, 1995), suggesting that a somite-free environment is a prerequisite for cranial nerve patterning. The vagus nerve trunk consistently circumnavigates the rostral aspect of somite 2 (Kuratani and Kirby, 1992; Shigetani et al., 1995). The same seems to be true for the lamprey embryo; the early vagus nerve passes between myotomes 2 and 3, and no nerve fibers were formed lateral to myotomes at early stages (Figs. 3C, 4). In terms of the LITERATURE CITED Alcock, R. (1898) The peripheral distribution of the cranial nerves of Ammocoetes. J. Anat. Physiol. 33:131–153. Balfour, F.M. (1878) The development of the elasmobranchial fishes. J. Anat. Physiol. 11:405–706. de Beer, G.R. (1931) On the nature of the trabecula cranii. Q. J. Microsc. Sci. 74:701–731. Bone, Q. (1959) The central nervous system in larval acraniates. Q. J. Microsc. Sci. 100:509–527. Bronner-Fraser, M. (1995) Origins and developmental potential of the neural crest. Exp. Cell Res. 218:405–417. 500 Damas, H. (1944) Recherches sur la dévelopment de Lampetra fluviatilis L. Contribution à l’étude de la céphalogenèse des vertébrés. Arch. Biol. 55:5–284. Detwiler, S.R. (1934) An experimental study of spinal nerve segmentation in Amblystoma with reference to the plurisegmental contribution to the brachial plexus. J. Exp. Zool. 67:395–441. Dohrn, A. (1888) Studien zur Urgeschichte des Wirbeltierkörpers. Mitt. a. d. Zool. Stat. Neapel 8–13. Franz, V. (1927) Morphologie der Akranier. Z. Ges. Anat. 27:465–692. Freund, R., D. Dörfler, W. Popp, and F. Wachtler (1996) The metameric pattern of the head mesoderm—Does it exist? Anat. Embryol. (Berl.) 193:73–80. Fritzsch, B., and R.G. Northcutt (1993) Cranial and spinal nerve organization in amphioxus and lampreys: evidence for an ancestral craniate pattern. Acta. Anat. 148:96–109. Goodrich, E.S. (1930) Studies on the Structure and Development of Vertebrates. London: Macmillan. Goodrich, E.S. (1937) On the spinal nerves of the Myxinoidea. Q. J. Microsc. Sci. 80:153–158. Haller v. Hallerstein, V.G. (1934) Kranialnerven. In L. Bolk, E. Göppert, E. Kallius, and W. Lubosch (eds): Handbuch der vergleichenden Anatomie der Wirbeltiere. 2-1. Berlin und Wien: Urban & Schwarzenberg, pp. 541–840. Hardisty, M.W., and C.M. Rovainen (1971) Morphological and functional aspects of the muscular system. In M.W. Hardisty and I.C. Potter (eds): The Biology of Lampreys, Vol. 4. London: Academic Press, pp. 137–231. Hatta, K., T.F. Schilling, R.A. BreMiller, and C.B. Kimmel (1990) Specification of jaw muscle identity in zebrafish: correlation with engrailedhomeoprotein expression. Science 250:802–805. Holland, N.D., L.Z. Holland, Y. Honma, and T. Fujii (1993) Engrailed expression during development of a lamprey, Lampetra japonica: a possible clue to homologies between agnathan and gnathostome muscles of the mandibular arch. Dev. Growth Differ. 35:153–160. Ishikawa, Y., C. Zukeran, S. Kuratani, and S. Tanaka (1986) A staining procedure for nerve fibers in whole mount preparations of the medaka and chick embryos. Acta. Histochem. Cytochem. 19:775–783. Jacobson, A.G. (1993) Somitomeres: mesodermal segments of the head and trunk. In J. Hanken and B.K. Hall (eds): The Skull. Vol. 1, Chicago: University Chicago Press, pp. 42–76. Janvier, P. (1996) Early Vertebrates. New York: Oxford University Press. Jarvik, E. (1980) Basic Structure and Evolution of Vertebrates, Vol. 2. New York: Academic Press. Jefferies, R.P.S. (1986) The Ancestry of the Vertebrates. London: British Museum (Natural History). Johnston, J.B. (1905) The cranial nerve components of Petromyzon. Morphol. Jb. 34:149–203. Keynes, R.J., and C.D. Stern (1984) Segmentation in the vertebrate nervous system. Nature 310:786–789. Koyama, H., R. Kishida, R.C. Goris, and T. Kusunoki (1987) Organization of sensory and motor nuclei of the trigeminal nerve in lampreys. J. Comp. Neurol. 264:437–448. Koyama, H., R. Kishida, R.C. Goris, and T. Kusunoki (1990) Organization of the primary projections of the lateral line nerves in the lamprey Lampetra japonica. J. Comp. Neurol. 295:277–289. Kuratani, S. (1997) Spatial distribution of postotic crest cells defines the head/trunk interface of the vertebrate body: embryological interpretation of peripheral nerve morphology and evolution of the vertebrate head. Anat. Embryol. (Berl.) 195:1–13. Kuratani, S., and S. Aizawa (1995) Patterning of the cranial nerve in the S. KURATANI ET AL. chick embryo is dependent on cranial mesoderm and rhombomeric metamerism. Dev. Growth Differ. 37:717–731. Kuratani, S.C., and G. Eichele (1993) Rhombomere transplantation repatterns the segmental organization of cranial nerves and reveals cellautonomous expression of a homeodomain protein. Development 117: 105–117. Kuratani, S.C., and M.L. Kirby (1991) Initial migration and distribution of the cardiac neural crest in the avian embryo: an introduction to the concept of the circumpharyngeal crest. Am. J. Anat. 191:215–227. Kuratani, S.C., and M.L. Kirby (1992) Migration and distribution of the circumpharyngeal crest cells in the avian embryo. Anat. Rec. 234:263– 280. Le Douarin, N.M., P. Cochard, M. Vincent, J.L. Duband, G.C. Tucker, M.A. Teillet, and J.P. Thiery (1984) Nuclear, cytoplasmic, and membrane markers to follow neural crest cell migration: a comparative study. In R.L. Trelstad (ed): The Role of Extracellular Matrix in Development. New York: Alan R. Liss, Inc., pp. 373–398. Lindström, T. (1949) On the cranial nerves of the cyclostomes with special reference to n. trigeminus. Act. Zool. Stockh. 30:315–458. Loring, J.F., and C.A. Erickson (1987) Neural crest cell migratory pathways in the trunk of the chick embryo. Dev. Biol. 121:220–236. Nakao, T., and A. Ishizawa (1987) Development of the spinal nerves in the lamprey: II. Outflows from the spinal cord. J. Comp. Neurol. 256:356– 368. Neal, H.V. (1897) The development of the hypoglossus musculature in Petromyzon and Squalus. Anat. Anz. 13:441–463. Neal, H.V. (1918a) Neuromeres and metameres. J. Morphol. 31:293–315. Neal, H.V. (1918b) History of eye muscles. J. Morphol. 30:433–453. Noden, D.M. (1975) An analysis of the migratory behavior of avian cephalic neural crest cells. Dev. Biol. 42:106–130. Oakley, R.A., C.J. Lasky, C.A. Erickson, and K.W. Tosney (1994) Glycoconjugates mark a transient barrier to neural crest migration in the chick embryo. Development 120:103–114. Piavis, G.W. (1971) Embryology. In M.W. Hardisty and I.C. Potter (eds): The Biology of Lampreys, Vol. 1. London: Academic Press, pp. 361–400. Romer, A.S. (1972) The vertebrate as a dual animal—somatic and visceral. Evol. Biol. 6:121–156. Ronan, M., and R.G. Northcutt (1987) Primary projections of the lateral line nerves in adult lampreys. Brain Behav. Evol. 30:62–81. Shigetani, Y., S. Aizawa, and S. Kuratani (1995) Overlapping origins of pharyngeal arch crest cells on the postotic hindbrain. Dev. Growth Differ. 37:733–746. Tahara, Y. (1988) Normal stages of development in the lamprey, Lampetra reissneri (Dybowski). Zool. Sci. 5:109–118. Tanaka, S. (1976) Facial nerve of the shark (Mustelus manazo) and its comparison with the glossopharyngeal nerve. Acta. Anat. Nippon. 51:1–16. Tanaka, S. (1979) Makroscopische Untersuchung des Nervus glossopharyngeus der Haifische (Mustelus manazo). Anat. Anz. 146:456–469. Tanaka, S., and T. Nakao (1979) A macroscopical study of the vagal nerve of sharks (Mustelus manazo) with special reference to morphological features of its branchial branches (in Japanese). Acta. Anat. Nippon. 54:307–321. van Wijhe, J.W. (1882) Über die Mesodermsegmente und die Entwicklung der Nerven des Selachierkopfes. Ver. Akad. Wiss. Amsterdam: Groningen, pp. 1–50. Veit, O. (1939) Beiträge zur Kenntnis des Kopfes der Wirbeltiere. III. Beobachtungen zur Frühentwicklung des Kopfes von Petromyzon planeri. Morphol. Jb. 84:86–107.