Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
structure and function of the digestive t r a c t of the grasscarp
cover photograph: Pharyngealteethofthegrasscarp.
Usually,awelldevelopedpharyngealmasticatoryapparatusispresentinstomachlessfish.Thefoodisfragmentedbetweentheventralteeth(photograph)
andadorsalmasticatoryplate.Scanningelectron.micrographx45.
0000
Promotor: dr. L.P.M. Timmermans, hoogleraar in de algemene dierkunde
Co-Promotor: dr. J.W.M. Osse , hoogleraar in de algemene dierkunde
/uiuoYzo? k
HENRI W.J. STROBAND
STRUCTURE AND FUNCTION OF THE DIGESTIVE
TRACT OF THE GRASSCARP
Proefschrift
t e r v e r k r i j g i n g van de graad van
doctor in de landbouwwetenschappen,
op gezag van de rector magnificus,
dr.H.C. van der Plas,
hoogleraarindeorganische scheikunde,
in het openbaarteverdedigen
opwoensdag 10september 1980
des namiddags tehalfdrieindeaula
vandeLandbouwhogeschool teWageningen
»i
CONTENTS/INHOUD
VOORWOORD
6
GENERAL INTRODUCTION
9
A.
Kaderstelling
9
B.
Structure and function of the digestive t r a c t of f i s h e s . . . 1 1
C.
Objectives of the experiments
24
GENERAL DISCUSSION AND SUMMARY
26
Conclusions
32
REFERENCES
34
SAMENVATTING (Zieookbijlage)
38
PUBLICATIONS:
A.
Stroband , H.W.J. Growth and d i e t dependant s t r u c t u r a l
adaptations of the digestive t r a c t i n j u v e n i l e grasscarp ( Ctenopharyngodon idella,
Val.) J . Fish B i o l . 11,
167-174 (1977)
B.
^Ê
Stroband , H.W.J. & F.M.H. Debets. The u l t r a s t r u c t u r e and
renewal of the i n t e s t i n a l epithelium of the j u v e n i l e
grasscarp, Ctenopharyngodon idella
(Val.) Cell Tiss. Res.
187, 181-200 (1978)
C.
£
Stroband , H.W.J. , H. v . d . Meer & L.P.M. Timmermans.
Regional functional d i f f e r e n t i a t i o n i n the gut of the
grasscarp, Ctenopharyngodon idella
(Val.) Histochemistry
64, 235-249 (1979)
D.
Stroband , H.W.J. & F.H. van der Veen. The l o c a l i z a t i o n
f
STELLINGEN
1. Er z i j n geen argumenten voor de veronderstelling dat b i j juveniele en adulte maagloze vissen de e i w i t v e r t e r i n g in het darmlurnen minder e f f e k t i e f zou z i j n
dan b i j maaghoudende soorten.
Dit proefschrift.
2. De opvatting van Trevisan, dat vooral het caudale deel van de graskarperdarm betrokken is b i j de resorptie van nutriënten, is o n j u i s t en komt voort u i t
het trekken van voorbarige konklusies u i t alleen morfologische informatie.
Trevisan, P. : Anat. Anz. 145, 237-248 (1979).
3. Het voorstel van Green & P h i l l i p s , graanmutanten te kweken met verminderde
feedbackinhibitie van aspartaatkinase door lysine en threonine, om zo het gehalte aan één of meer aminozuren u i t de aspartaat-familie te vergroten, verdient
s t e l l i g waardering maar l i j k t geen praktische bijdrage t o t de oplossing van het
wereldvoedselvraagstuk te kunnen leveren.
Green, C E . &P h i l l i p s , R.L. : Crop Science 14, 827-829 (1974).
4. De circadische f l u k t u a t i e s in gevoeligheid van diverse weefsels voor bepaalde hormonen suggereert dat hormoonreceptoren een korte levensduur hebben.
Meier, A.H. ; John, T.M. &Joseph, M.M. : Comp. Biochem. P h y s i o l . 40A,
459-465 (1971).
Meier, A.H. ; Trobec, T.N. ; Joseph, M.M. &John, T.M. : P r o c . Soc. Exp.
B i o l . and Med. 37, 408-415 (1971).
5. De i s o l a t i e van zowel één g l y c o p r o t e i n e r i j k - als één glycoproteinearm gonaidotroop hormoon u i t hypofyses van beenvissen ondersteunt de opvatting dat "glob u l a i r e " en " v e s i c u l a i r e " P.A.S.- positieve gonadotrope cellen t o t hetzelfde
(celtype behoren, maar s l u i t n i e t u i t dat er een tweede gonadotroop celtype, mog e l i j k niet P.A.S.- p o s i t i e f , aanwezig i s .
Ng, T.B. &I d l e r , D.R. : Gen. Comp. Endocrinol. 38, 410-420 (1979).
Peute, J. ; Goos, H.J.Th. ; de Bruin, M.G.A. &van Oordt, P.G.W.J. : Ann.
B i o l . Anim. Bioch. Biophys. 13, 905-910 (1978).
Ueda, H. : Bull. Fac. F i s h . Hokkaido Univ. 31, 1-15 (1980).
6. Het zou van groot belang voor de wereldvrede kunnen z i j n , onderzoek te verrichten naar de oorzaken van het verschijnsel dat achteruitgang van de levensstandaard veelal l e i d t t o t toename van onredelijk vijand-denken en stijgende wapenproduktie, en in d i t verband onder andere aandacht te schenken aan de mogel i j k e rol van het " M i l i t a i r Industrieë'el Komplex" .
7. De boycot van de Olympische Spelen i n Moskou, waarvoor de verschillende redenen op zichzelf terecht werden aangevoerd, doet vermoeden dat w i j , westerl i n g e n , het restant van de boterberg die aan de Sovjetunie is verkocht op ons
hoofd hebben gesmeerd in plaats van ook onszelf boter op het brood te geven.
8. De "vertrossing" van de omroepen l e i d t er toe dat de a f l e i d i n g die wordt
gebracht de bevolking a f l e i d t van de meest wezenlijke problemen van onze samenleving.
9. Het f e i t , dat de vervoersorganisaties bezwaar maken tegen het gebruik van
"hun" belastinggelden ten behoeve van de aanleg van fietspaden betekent dat de
wegvervoerders te weinig oog hebben voor de fietsende medemens en onderstreept
derhalve de noodzaak van genoemde wielerpaden.
10.De "brede maatschappelijke discussie" over kernenergie z a l , wanneer het kabinet zich n i e t onthoudt van voorbarige uitspraken, vergelijkbaar b l i j k e n met
een golfstroom : oeverloos en met voorspelbare bestemming.
Proefschrift vanH.W.J.Stroband
Structure and function of thedigestive tract of thegrasscarp.
Wageningen, 10september 1980.
of protein absorption during the transport of food i n
the i n t e s t i n e of the grasscarp, Ctenopharyngodon
idella
(Val.) Submitted to J . Fish Biol
Stroband, H.W.J. & Annet G. Kroon. The development of
the stomach i n Clarias lazera and the absorption of
protein macromolecules. Submitted to Cell Tiss.Res
URRICULUM VITAE
IJLAGE :
Stroband , H.W.J., J.H.W.M. Rombout & J.H.M. Davina.
Maagloze vissen; Bouw en functie van het darmkanaal.
Natuur en Techniek 48, 38-55 (1980)
Vooo-u^ocyi-cL
De.
ncduuuA-
kvdt
op
mu
<xl£u.oL
e&n.
astoti.
cum
JttZM/ekAv'rL uoyn. Z&n. c m o W o « ^ in. eé-n. 7)uIusa^t<feJ&nÇ>cAc<.pp
sùjAe.
Z/cÀtin^,t/Hl. yôle.clt
g&&ig&>n^rie-<.ol
cnn.cnnTTULT
e*>',#A.
diep,
AÀ&.CAZ.ej&n.
ec^ï.
Q£JCagjesri^fte^.oL
rruW
ç*iia«.
cOep
«CHOJ« hvxjeu
(e, *4h*- fw^eé-n. AJbin. speojzx. uoyn. to-n.
X«jf £ > t o / e
Geßcovun. ^cLout ote. TZocéuu^. sreMiq. lo . CJui mü. £etre/é Äe&ßt
<JUL ozJóislcL tot een. toe*tt*tuisnjoCe. t&L&'tAcd. Votyt. etedliusrit. ÏJCL.
uoort- W « ds, ScAefv/og.
/'s , op t</e£»fe UJ'CLTJL <U&K£. ocrK éot
ITbjfa. %&ruuo7n cutting, mut ujere^r>cJicLPpeÂu^
on.J&vru>eÂ.
huât
nvt kesÂ. sjt&i cù- ^te£cL.ti{/i^eM:
vast. da?yruw>z£âkekw
rus doen. tsi.vaA£*i. en. heoM- yniL Im. uzejP. opzJcÂtfrn. \MM£iae.uMyr-pirn, tot ksi. TL)Vo uasn. zJesi. , nor&n., uoeJ&n., i.u,ike^rL. est- utA.bc
Z£sri-. ü e f o t t t n O UOUTL. U*AJüüonrLclsi/li«ri.cL. , SZA.bie.oi- 6*1. oùcunÂbcux
r\£J<A- trtjuùmop da. Oocrta^-orusL &n~hejt JÜfn. otCxjg.. a&ssoeJibn.b
UJcLeuL-cucun. \J( i/fL de. ee/xote. pXcœdb) uJrcUuJ&Âinq. u/il aju/esi. in. out
1/ocyiMJoeytoL. , cLouL [/o&ixui- e&ru oiovnJçuJoo^Uit- ia.
KJ^Lcuu-OL UJil ÙK OCUTL. C/JL. WÜ. rux. qjzJjooi&m. at&flAn. njüoL at&ruAÀk maAesn.
om. igjjLtAJüe^n. Z&. / W o t o n ^ w i . a^e~, CÜA&C
ot- hmx/JsiiLcjt , e&n. zo-6. ÄJIBJ.6 çp^peii&t.
SV'Ù. mijn. \/arvntÀsruL,
-htU.ciUjL f o£ AsJt rot STconJC Jrvrnxsn- Uovn. otit UJtnJrie. . / a t u 77'u
aXlU. ^a^nftyi. Jçasn. IÂ 'un. out JstóteJ; voornam-. , mou**-. 'tA
hoop deut
Z.U OÜJL. kei. cLosno<tjaL.t z-icÄ. a.Gvn.atàtrij&rçen. u/e.
U-n. .
O n . oCe. ee/idUL. p^Ccuodo
Ârtsn. ïJL müm. ouu/t/vn ott&viJi
v&rs>chujŒ.<UjiLoL , l/ocn.c(sC ocr& omota-t ze. "ma Ucun. Lemgo a/^
cuwi.
ez<ri- asuotiL. trT-ayfleJoi. uasn. o6t**Jt&n- &n. ncunoù^te^rt- eux.
Vts/i- , uJcLoiA-ctoo^~ man. sfçcvtv) op onZp<;ooi/na £>'&>%• u/eA.cL vtn
a/lx>ot . \Zk/LdU/u njL&Jc rLusn. OCÓT£JCJ'£\-are.unrL. un. u;o<ri*£ &nTHicùù&vL.
fnü. otm ^ c e t / e ^ . ae?»/*//V-ee-^o£ 6^>- -*»%*>• &rucUe. n
<£xitrLci£.h 7n.ün. o n & ^ t o tx/LC UK nmCtn.&c4oort-OLLc/ksis>
cA-ovnJwsn- VtxrT- 06e. JOü^jm-oleA^.
uj'ànje. U/a.a*.op *cji CL£JÜCL in&t-
bj>icma&rL &ÖM- cnn. ~mä? cmaLcunÀb fud. JjuJen.
in. QJvn. £si.q. W^fyxe/xJn., de. ojL^axjikbLoL
Cfe. a e v e / t . opéiiv)a.aJL a.
-btuoùiAJim..
Van. oui. o-txjt. \yCui#ifU&n. 7ivi£ u/te. iJc vJcLctA.ua.i/u^_
tn,
yLcuuA-^cvmj.. fcrri-raJcf&n- kxJlr THoqgm. rieJlr^n(jJiK. L& hm*. CAL.
JLa*mi&itL XjZ&JuisnJ>*s>-Qs 7>o4<*n£sn- cli*-, olocn- AcLavt. CLttsjTuxüfieJcL en.
pÜbCuaoLe.-iin.
ort.œéTujiy£e£.oL OT.óle- \sn.\/-Coe.cL. op Wwn. orj-ujikht.'*\- *-ü"i- fooi. ynjL uctsn. cpioói
JteJeJvbn.lo.
~T&n- CLam/ziin. L>um. kut.
rtLzn- JirtocJvuu/e*L.
omcù^u"sxJr
AJ&rt- un
un. ctz. te/uite. P£.CLCL£O cLasmJr
ueyu>c/ujAcùa^xxvn.
müm. P^crrfoTcTL. , Lu.cy Tùm*nin--y»cufts> , ß-n. co~
Pio-moro-T-,
"Itua. Os»se_ . Z ii rudl&tvL. e t . m&z. (ptou. in^La-t ubvx. g&.ü\/&u>L
e&rt- Ucuflç CfrMrtAD 02. yßfrVyrttAJlsn. C£LL- , TWLclJL. oLtxn- cZe. XsyxjyvMyi.
fa/n.
vnJüb^otJr&ia
op Ir^divioCu. , u/e&he£.
es*- aiX v)/\so,ç&n.
'i'tLouaJS) ynoj^i. *-üii. LU<Z.CIA— oO'K ZBsn. on-caMu?*-*
CL***> hxT cm. Wlnsfiau/ioJZ- XJn.t/o<- srCcvn— U/aruaCe^n- uÂ.i.aeA^o<jx-oC. . l/e*c&t_ oicurJv
 rutm^ UT&T- oU- cptote. Tnctte. ucen- w.ü.neJcL eile. mü. C.ot. is.
XILU aruÂA/iLoeJx
a&J-cuUbn. uJt^.cL , e^n. uoan- <Ue_ zjuesi- A~cm^ bT-yi.ocJrtle.y/e- vutui.
uicccuioP use. UCUTL. a&c/at.oAZii*t.
/i^KrO^n-
J)e. snoKJxcpeL.'b
vcuYL. de.
uaJraftotj>
Ç^pesu?rtiUru'e£a~
")'uiA^yncr^o\.ùaÀjt. &7L. GeJ2Joio£ccple. , cfae. oCoxn^ /LUW- sursitriZ>CJU&.
lin. Zbn- Po4//le.\/e.
^>ü.ooi.a.a£- hjlJb&tsrt- QXJJU/ZA-GL eta^t- heJl
•m-clisvJLoeJç, u/iJ- Isfa n.ixyL- oJco Qkzo^p &e.oCa*idresi—. <£e^i_
WiAeMß- UJ'/j.on.ol&AJsriq. mouccJc LÀ i/ocrt- >«fn_ fKowoou-Û : u/ij.
Chu*.
TA(jz£e.
tri-
Wee*?) uwn.
o&t_ 7 % * ^ -
»recé
LA
CLrL-
dcA./'PP&u
f
G«A.-f-J^cum~ U&. G-T-GLaß-, (Zwner
hieran-
e^u
&-jk
Vcvn.
e - a/ncuCistèsn.
'•>Ta.ae.
"0
oCU. i/oox. sn~cn.ze. ry^C
O
oucon-
n&-i emo«^_-
TjytÄ
k&ß&fn-
mjvexuMj&iJït.
7)'ULL TrtoojeJŒfftr azMjwïb
t-üt-
Ooo*-icpyno XJOU. £1*.
e™. cuttele^.
XJOTI-OUA- cue.. uTrtnheJßßäJft.
Z.(yLeLtnA.oiXe. amtuL. *&iotecÄvii/rus>
5u.rJtg-
uet 10
Lie^rLohxcc. </*- f */-
~
[/cur*- oU. s*r(&£.a<*J'S xVu-i*/erî_ cfe. Ucunrcpi*cp »Ut bÖLvUot-fL&n- tdt keJL cm.d^unLoeJ\ uf'^C 'ußr rio&mem. olt nxAJun. 7.ocun.
CAL»X.éio€4K ( f-ys/o&aie.
cfoi. Dinsw**- ) , otie. X-tf™- ïiarof**- sCa&oTa.ro-iiu*r*t- a<tAr\AUL. Uooz. oru) op€*isy&éu£. • S.A. HuAsivneUL
oüe. cu&> mibUejbuesisrct^. ua/n. où.
Ch.a.<v*t)<>a,LL£.iiuu [/t^-b^he. ,
luneL Ucuru oCt. sôiwrU*LV i'Y>&iti
de. -nvuvne. Q/las>itcLn.p&is> -&uKnjoül. IAJCUUICLWI. rit^t UJVuA is> ve^UcJuC- ;
\ZcnA (C-A.6.0.)
oLCe., ti>e>n.a£o où AJUZSL. A.W.b.M.
vcun. C^^-CUSLO
(*>?/cne>6/o/ o * / ' e J FOL OTVXJL. asiole. oCcurun^cuuL neJe/- yO&uù.oL u/as> oùe.
LrwL CLwin<rJLu*iA—<xsr>aiJ!.y$&> uit
te. \/o*WTtsn. ù^vy^orr-e.
cù. AWLC . l l RicJLr&t. f[/Cn^ee£.T e*u i / À ^ - e t y J oùie. orv>CÙL
Cx<t/t-/<is> sZaLt^Mvle. ucrox. cmcùsi.z.crt*rc TtCLct*. où. cmTnJ'unJreJzünjL
(/CUn. où. mcLoLOr.
H&i iyeJU. uitAsk otaX
Ujiiw \/a£e^i. i^uxicÀ.é'fe.
t^n.
Jfe-noe-l/C. x/asn. où- Lcùuv-n.etJ'ùr) un. oLtT p^toe/oonyii^
U/CLOULCÙÉ*'uk bua.rn.oun..
De. JUZ&L W.6. C&A-i (T.F.O.L.)
oLt*Jc ik uvcru
cù. JOuLcrrci^rig, Ucvn. AûJ. S.M. ^orontcut^-i'OUL£.
.
JDasnJr Js&n. (sn crtrK u<AOcsnujÇ.&ùi<pc/- ouun. Ae^t. c&*.
Aa.t ueze. ^ypeu/e^Jr
Uoon. hjuun. zeJa*u*i<L, TJcunuvn. ;
m,£Hy
Ta.pi'A.cuna. , HcuM^y \AnJsu.cLZ , G toucla. 0-0*1. 7Pcuê»6~t- , 7h<~
UCVTL. CouppeJc^e.
frru >rui.tsiou->*JP. IhjwzXAS) {TeJrsT o4smJ&nsn/\ i»q.
Ûe.
VTL&pcvn^ts*<L&>ri. Ucvn. <*e. riM/l. L.. ^£>o(mr.a.cut>iP
tin. JçonA&Jrtùe.
U<vn. nvüsL. (sOfLoo^-)
"esi.ae£o " AMX- 'un bjÀ0
Z<m.oUn. op fztf
ppvÄcJcjvL .
TilOA-'à^rLe. , -nÓLÓLvt J^OUM/ zcrZ. cu&i -or&usn. e^i_ /o£</-*t_JLoLaX. ruiAr Jjt. où. sVCWl*ir>Utez&^t- , QL&paj*A.oC g.cut*n.t/e. -yruLC AAJ
toc £>fa#>.o£ sfrovnjwvcuru oùlt p^ofJ^cJuLiJt/
un. Aocpi.-nretÉe.
Jltfp&isnt
cCocn. j^)u.ny s&4AßslcljLeJUo^L e&wït. pcieut. v**ve*
Ucun. oU. \s<McJfa)'CZ&>rieU. m cvr)us>'TCA.ipr&rL. uÀt OL "rlJUresi.. J«.
VUL£L Tt'utX. ctccé
LA J*. oCeteuiwocn. Oec/<w£,
Tffocovi. wü ypvi-
ke>n.
cJUreuvi-
Trog w&£ -rrudest- .
(fL&ru*--^
?
GENERALINTRODUCTION
A:KADERSTELLING
Hetinditproefschrift beschreven onderzoek werd gestartin1973 binnen
de toenmalige afdeling DierkundevandeLandbouwhogeschool waarvandeleiding berustte bij prof. dr.J.W.M.Osseenmevr. dr.L.P.M.Timmermans.In
die periode werdeenaantal jonge biologen aangetrokken inverband metde in
1970 gestarte studierichting Biologie aandeLandbouwhogeschool.Het
isdanookniet verwonderlijk dat het onderzoek vanmeetafaan beïnvloed
werd door drie faktoren:
1.Het moest toegankelijk zijn voor doctoraal studenten,niet alleeninde
biologie maar ookinbijvoorbeeld voedingen(vee)teeltwetenschappen.
Daarom diendehetvoldoende breedteworden opgezet.
2.Het diendetepassen binnenhet kadervanhet onderzoek vandeLandbouwhogeschool,datwil zeggendat"landbouwkundige" aspekten o.i. gewenst
warenenhet onderzoek daarom nietaltezeer zuiverwetenschappelijkvan
karakter moest zijn.
3.Develeoptezettenprédoctorale onderwijs elementen eisten relatief
veel tijd op,indeeerste jaren zelfs praktisch 100%.zodat het onderzoek
een trage start ondervond.
De derde faktor leiddeertoe datergedurendeeenlange periode konworden nagedacht overdeinvulling vandeonderzoekstaak vandeafdeling,de
eerste twee faktoren bepaalden medederichting waarin gedacht werd.
Bijenkelevandebetrokken medewerkers bestond ervaring inhet onderzoek
metvissen.Dewetenschap dat visseneenbelangrijke voedselbron (kunnen gaan)
vormeninvele landen naasthet feit dat fundamenteel onderzoek aandeze lagere gewervelde dierenaandeLandbouwhogeschool ontbrak hebbenertoegeleid
datvanmeetafaan vissenalsproefdieren het meestinaanmerking lekente
komen.
Tenslotte werd gekozen voor o.a. onderzoek naardevoedselopnameen-verwerking bij vissen,waarbij morfologisch onderzoek t.a.v.devoedsel verwerving werd uitgevoerd binnen desectie Functionele Morfologie terwijl binnende
sectie Histologie/Ontwikkelingsbiologie onderzoek werd verricht naar bouwen
funktievanhetdarmkanaal. Dit laatste onderzoek bestond uiteen tweetal
projecten, waarin r e s p e c t i e v e l i j k vorm en functie van het darmepitheel en
aspecten van endocriene r e g u l a t i e , met name structuur en funktie van hormoon
producerende cellen,de aandacht hadden (Rombout).
Van de diverse takken van onderzoek die zich l a t e r (met name na de s p l i t sing van de afdeling Dierkunde in een voorlopige vakgroep Experimentele Diermorfologie en Celbiologie en een voorlopige vakgroep Dierkunde) binnen de
vakgroep E.D.C, ontwikkeld hebben is er één zeer d u i d e l i j k aan het darmonderzoek gerelateerd, namelijk
het onderzoek naar de ontwikkeling van het
immuun-apparaat b i j vissen (v. Muiswinkel). De r e l a t i e met d i t binnen de
sectie Celbiologie bewerkte projekt l i g t vooral in de w a a r s c h i j n l i j k n i e t
onaanzienlijke rol die afweercellen in het darmepitheel spelen b i j het voorkomen van i n f e k t i e s via het (maagloze) spijsverteringskanaal van de door
ons onderzochte proefdieren.
10
GENERAL INTRODUCTION
B :FORMANDFUNCTION OFTHEDIGESTIVE TRACT INTELEOSTS.
1. Morphology
a. Mouth cavity, pharynx and
esophagus.
The digestive system of teleosts is not basically d i f f e r e n t from that of
other vertebrates including mammals. In a number of respects, however, i t s
structure is a s i m p l i f i e d one. Digestive and absorptive functions seem to
be carried out by a lesser d i v e r s i f i e d morphological system.
Mouth cavity and pharynx in aquatic vertebrates are f o r continuous resp i r a t o r y and discontinuous feeding functions, more so than in t e r r e s t r i a l
vertebrates. G i l l s and associated structures occupy nearly the whole pharynx
( f i g . 3). Taste buds in the pharyngeal epithelium are numerous in many f i s h
species (Curry, 1939; Mc Vay & Kaan, 1940; G i r g i s , 1952; Kapoor et a l . ,
1975a; Sinha, 1976 a ; Sinha & Moitra, 1975 a ; f i g . 1, 2 ) . No salivary glands
have been found (Fahrenholz, 1937). The position of the mouth, the presence
or absence of teeth on the jaws, vomer, palatines and pharyngeal bones, the
morphology of the g i l l rakers, and other characteristics are closely related
to the feeding habits. Reviews on this subject have been presented by
Suyehiro (1942) and Kapoor et a l . (1975 b ).
The esophagus is usually lined with a squamous epithelium, j u s t as the
pharynx ( f i g . 2). I t s surface shows concentric microridges, j u s t as e p i t h e l i al c e l l s in the skin of teleosts (Merrilees, 1974; Reutter et a l . , 1974).
In the epithelium, mucous c e l l s are abundant and tastebuds may be present
(Chitray, 1965; Sinha, 1976 b ; Sinha & Moitra, 1975 a ; Verigina, 1976; Moitra
& Ray, 1977). Esophageal m u l t i c e l l u l a r glands are not common (Kapoor, 1975 ).
b.
Stomach.
In about85%ofallteleostspecies theesophagus leads intothestomach.
The other 15%ofthebony fishesdonothaveastomach (Jacobshagen, 1937);
the esophagus enters theintestine directly.Thesame applies tothelarval
stagesofmost speciesoffish (Balon, 1975),when they take exogenous food
although their stomach hasnotyetdeveloped.
11
^'•^,"•''.'.
12
The walls of the stomach and the i n t e s t i n e consist of s i m i l a r layers of
tissue as found in higher vertebrates ,but the i n t e s t i n a l
mucosa lacks a
muscularis mucosae ( C i u l l o , 1975; Korovina, 1976).
The stomach usually shows two d i s t i n c t sections: a corpus part with a
l i n i n g of mucous-producing c e l l s with underlying gastric glands, and a
p y l o r i c part without gastric glands (Mohsin, 1962; Kapoor et a l . , 1975 ;
Moitra & Ray, 1977). The glands are formed by only one type of c e l l ; no
d i s t i n c t i o n can be made between pepsin producing c e l l s and oxyntic c e l l s .
I t has been shown that corpus gland cells produce pepsin as well as hydroc h l o r i c acid in bony fishes (Blake, 1936; Barrington, 1957; Bucke, 1970;
Verma & Tyagi, 1974; Moitra & Ray, 1977; Noaillac Depeyre & Gas, 1978). The
same applies to a l l nonmammalian Vertebrates (Smit, 1968).
e. Pyloric
aaeaae
Pyloric caecae are found in many teleosts with a stomach. The histology
is not d i f f e r e n t from that of the i n t e s t i n e and this suggests that t h e i r
primary role is to enlarge the intestinal area (Moitra & Ray, 1977).
i.
Intestine.
In stomachless f i s h and in f i s h larvae, the f i r s t part of the i n t e s t i n e
is a widened tube, called i n t e s t i n a l bulb. I t is assumed to have a storage
function (Babkin & Bowie, 1928; Mc Vay & Kaan, 1940; Berry & Low, 1970;
/ e r i g i n a , 1978 ). In the bigmouth b u f f a l o , Verigina (1976) found an i n t e s t i lal bulb with a very thick p a r t l y s t r i a t e d muscularis. This may be related to
;he mechanical processing of food, and might be seen as an adaptation to the
loorly developed pharyngeal teeth and the absence of a pharyngeal plate in
;his species.
The b i l e and pancreatic ducts, generally located closely t o g e t h e r , j o i n
he i n t e s t i n a l bulb at a short distance from the entrance of the esophagus
Rogick, 1931; Curry, 1939; G i r g i s , 1952; Noaillac-Depeyre & Gas, 1976;
e r i g i n a , 1978 b ).
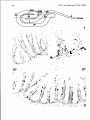
ig. ].Scanning electronmicrograph of the rostral part of thebuccal
cavity of a6-monthsold grasscarp (x 105).L = dorsal lip;
V =valve prohibiting outflow ofwater during the expiratory phase
of respiration. R=roof of thebuccal cavitywith concentrations
of tastebuds (arrows), also present in the areadirectly behind the
lip.
13
fig. 2.Scanning electronmicrograph ofpart of the roof of thepharynx.
Note thepresence of concentric microridges onepithelial cells,
possibly facilitating gas exchange of the cells and/or
holdingmucous at the cell surface(Reutter et al., 1974).
In the left lower corner,a tastebud (x 5250).
14
Morphologically,no clear regional differentiation was madeinthe intestineofteleostswithastomach.Some species havearectum,set apartfrom
the restofgutbysomekindofvalveorfolds (Jacobshagen,1937;Ciullo,
1975).
e. Histology of the intestinal
mucosa
Themucosaofthe teleostintestine showsamoreorless complex pattern
of folds (fig.4 ) .Villi haveneverbeen observed,multicellular glandsand
cryptsofLieberkühn,presentwithin thewallofpartsofthe mammalian
intestine,areabsentinteleosts,exceptinthefamily Gadidae (Jacobshagen,
1937; Klust,1940;Bishop&Odense, 1966). Theepitheliumisasimple
columnaroneandcontains threeepithelial cell types.Absorptiveenterocytesbearmicrovilli (fig.6)andciliahavebeen foundinafewspecies
(Barrington,1957;Iwai, 1967 a 'b ;Bucke,1970;Verigina,1978 a ). Furthermore
theepithelium contains mucous goblet cells (fig.3,4)andenteroendocrine
cells.
Betweenthethree epithelial cell types,migrating cellsmaybepresent
(Rogick, 1931;Girgis,1952;Bullock,1963;Hale,1965;Smit, 1968;Bucke,
1970; Krementz&Chapman,1974;Weinberg, 1975;Davinaetal. 1980), These
areprobably lymphocytes,macrophagesandgranular leucocytes. The
presenceofsocalled "pear-shaped cells"or"rodletcells"intheepithelium
lasbeen reported formany teleostspecies.Someauthors areofthe opinion
thatthese cellsareinfactprotozoan parasites (Rhabdospora
thelohani)
b
bannister, 1966; Iwai,1968). Al Hussaini (1949 , 1964)mentionedthe
lossibilityofrodlet cells being developing mucous cells,but thisisnot
:orrect (Hirji&Courtney, 1979). Catton (1957)suggested thatthe rodlet
:ellsaregranulocytic leucocytes,but concentrationsinbloodcelliroducingorgans were never found.Many recent authors considertherodlet
:ellsasunicellular glands (Leino, 1974;Grünberg&Hager, 1978;Matteyet
1., 1979).
Cytology of absorptive
cells.
The general morphologyofthe absorptive enterocytesofbony fishes
howsastriking resemblancewith thatofhigher vertebrates.Similarorgaellesarefound.Minor differencesarethelackofcomplex interdigitations
15
fig. 3.Lateral part of thepharynx floor,with twobranchial arches,gi
filaments (F)and gill lamellae (L).On thebranchial arches the
gill rakers (R) (x 100).Inset: Tastebuds (arrows)on the gill
rakers (x 700).
16
of the l a t e r a l plasma membranes of adjacent c e l l s , and the presence
of lamellar infoldings of the plasma membrane in the basal part of the
c e l l s in teleosts (Yamamoto, 1966). These infoldings appear to be s i m i l a r
to those described f o r the "basal l a b y r i n t h " of proximal tubule c e l l s of the
kidney in mammals. A function in osmoregulation has been suggested (Yamamoto,
1966; Noaillac-Depeyre & Gas, 1973 b ).
In stomachless f i s h three segments can be distinguished, on the strength
of the morphology of the absorptive c e l l s , and thi smorphology is
directly
related to the absorption of food. Therefore, some information about the
digestive enzymes and absorption in teleosts should be discussed before
dealing with the regional d i f f e r e n t i a t i o n .
2.
Physiology
a) Digestive
enzymes
Theenzymesintheintestinal lumenofbony fishes are essentially
similartothose foundinmammals.Intheory theyareproducedinthe pancreas,
the gastric mucosa orthe intestinal mucosa (including pyloric caecae).
Productionbythe intestineisdoubtful (Kenyon, 1925;Jany, 1976), although
Kapooretal.,(1975)areoftheopinion that themain protein-,carbolydrate-andfatdigesting enzymesare also producedbythe pyloric caecae
îndintestinal mucosa.Itismore likely,however,thatenzymemolecules,
derived from the pancreas,havethetendencytoaccumulate inthe glycocalix
)ftheenterocytes (Fänge&Grove, 1979). Thismayalso explainthepresence
)fproteinases,lipaseandamylaseinextracts from carp intestine (Al
ïussaini, 1949 ).Onlyanenterokinaseandprobablyanamino peptidaseare
)roducedbythemucosaofthe fishgut(Creach, 1963;Ishida, 1936;Bondi
1Spandorf, 1954).
In fishwithastomachthecorpus glands produce hydrochloric acidand
>epsinogen.When activated theenzyme showsaoptimumpHofabout 2.5,
/hichiscommoninvertebrates.Since more thanonepHoptimum has often been
oundinacid protease activity (Alliotetal., 1974;Creach, 1963),asecond
iroteolyticenzymeislikelytobepresent,probably cathepsin withan
iptimumpHof3-3.5. Thishasthe samequantitative proteolytic activityas
lepsininpikeandtrout (Buchs, 1954).
Therearenoimportant differencesintheproductionofenzymesbythe
17
18
pancreas for f i s h with or without a stomach. In both cases the pH in the
i n t e s t i n e is neutral to s l i g h t l y alkaline (Shcherbina & Kazlauskene, 1971;
Creach, 1963; A l l i o t et a l . , 1974). Consequently there is no peptic a c t i v i t y
in the digestive t r a c t of stomachless f i s h (Kenyon, 1925; Babkin & Bowie, 1928;
Smit, 1968; Ishida, 1936; Kawai & Ikeda, 1972; Jany, 1976). The p r o t e o l y t i c
enzymes produced by the pancreas are t r y p s i n , chymotrypsin and carboxy peptidases (Creach, 1963; A l l i o t et a l . , 1974; Fänge & Grove, 1979). Among the
other enzymes in the i n t e s t i n a l lumen of f i s h are the carbohydrases. Amylase,
maltase, glycogenase, sucrase and saccharase a c t i v i t i e s have been found in
some stomachless teleosts by Ishida (1936), Sarbahi (1951), and Kawai &
Ikeda (1971), invertase by Ishida (1936) and Dhaliwal (1975). Cellulase could
not be detected in the teleost i n t e s t i n e ( I s h i d a , 1936; Migita & Hashimoto,
1949). The pancreatic j u i c e and the i n t e s t i n a l lumen of most of the studied
teleosts contain also lipase (Babkin & Bowie, 1928; Agrawal et a l . , 1975;
Goei, 1974; Sastry, 1974 a ' b ; Kapoor et a l . , 1975b; Falge & Shpannkhof,1976).
Patton et al.1975, suggested the presence of another fat-hydrolysing enzyme
in f i s h that may compete e f f e c t i v e l y with lipase as a major f a t digesting
enzyme.
Apart from endogenous enzymes, exogenous substances might be of importance
f o r the digestion of food. The possible role of microorganisms in digestion
has been studied f o r carp, grasscarp and tench by Jankevicius and colleagues
(Syvokiene et a l . , 1974; Syvokiene & Jankevicius, 1976, 1977; Lubyanskiene
et a l . , 1977). Other studies were made by Paris et a l . (1977) and Sacquet et
a l . (1979)for carp, grasscarp and t r o u t . The results show that microorganisms
nay play a role in the fermentation of carbohydrates and the digestion of
Droteins, but only the l a t t e r may be physiologically relevant in stomachless
f i s h . According to Dabrowski & Glogowski (1977), a u t o l y t i c enzymes in food
night play an important role in the digestion of proteins in f i s h larvae.
Other enzymes, probably produced by i n t e s t i n a l c e l l s , and located in the
nembranes of the microvillous border of the absorptive enterocytes might be of
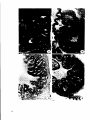
Eig.4. Scanning electron micrograph of somemucosal folds in the intestinal
hulb.Note themany mucous goblet cells (arrows) (x 150).
:ig. 5.As fig. 4,x 1800.B=bacteria.
fig.6.As fig.4, x 8750. Note thepresence ofmany microvilli on the
epithelial cells.
19
significance at least in mammals (Ugolev, 1971; Gossrau, 1975). Only a few
studies on this subject have been made for t e l e o s t s . Of interest are the
findings of Fänge &Grove (1979) in white grunt. A dipeptidase activity was
noticed, especially in the epithelium of the anterior i n t e s t i n e . In mammals
proteins are likely to be broken down to oligopeptides, and these are
absorbed (Smyth, 1971; Crampton, 1972).
The presence of a dipeptidase, especially in the anterior gut musoca
is in accordance with Babkin &Bowie (1928), Hickling (1966), Alliot et a l . ,
(1974) and Cockson &Bourn (1973), who found maximum proteolytic activity in
the anterior intestine of the studied fish species. Similar results were
obtained for lipase and amylase a c t i v i t i e s (Hickling, 1966; Al Hussaini, 1949 )
but Cockson &Bourn (1973) found similar amylase activity in anterior and
posterior intestine of Barbus paludinosus.
b. Absorption of
nutrients.
The expectation that the localization of most digestive enzyme a c t i v i t i e s
in the anterior part of the intestine might lead to a proximal to distal
gradient in absorption of nutrients has not yet been confirmed. There are a
number of studies, however, that suggest this to be true for many fish.
Alkaline phosphatase activity is higher in the anterior than in the posterior
part of the gut of several teleosts (Al Hussaini, 1949 ; Arvey, 1960; Sastry,
1975; Srivastava, 1966). Khalilov (1969) found an increase in the size of the
Golgi apparatus and in alkaline phosphatase activity in the anterior intestine
of tench after a fatty meal. The same was found after feeding starch. Broussy
&Serfaty (1958), Sivadas (1964), Iwai (1968, 1969), Tanaka (1972), Gauthier&
Landis (1972), and Noaillac-Depeyre &Gas (1974, 1976) by applying morphological techniques found that most of the lipid was absorbed in the anterior intestine of carp, goldfish, Tilapia, and in a number of fish larvae. This was
confirmed by physiological experiments (Shcherbina, 1973, concerning l i p i d s ,
Shcherbina & Sorvatchev, 1969 and Shcherbina et a l . , 1976, for proteins;
Farmanfarmaian et a l . , 1972, Sastry en Garg, 1976 and Shcherbina et a l . , 1977
for absorption of sugar). Sastry en Garg (1976),however, found absorption of
lipids all over the intestine of Ophiocephalus and Heteropneustus by the
application of histochemical techniques.
20
3. Regional differentiation
of the
intestine.
In stomachless teleosts and in f i s h larvae two i n t e s t i n a l segments can
be often distinguished: an a n t e r i o r segment with enterocytes loaded with
l i p i d p a r t i c l e s a f t e r a f a t t y d i e t , and a posterior segment with many pinoc y t o t i c vesicles in the apical part of the c e l l s (Yamamoto, 1966; Iwai,
1969; Gauthier & Landis, 1972; Tanaka, 1971; Noaillac Depeyre & Gas, 1973 a ,
1974, 1976). In some cases a t h i r d segment (rectum) has been described.
The caudal segment contains many lamellar i n f o l d i n g s , which according to
Noaillac-Depeyre & Gas (1973 ) might have a specialized function in osmoregulation.
Orally administered horseradish peroxidase was absorbed by c e l l s of the
second segment in adult stomachless f i s h (Gauthier & Landis, 1972; NoaillacDepeyre & Gas, 1973 ). This capacity to absorb protein macromolecules is
also known f o r suckling mammals, in which the digestive system i s s t i l l not
f u l l y developed (Clark, 1959; Leissring et a l . , 1962; Kraehenbuhl & Campiche
1969; Staley et a l . , 1972). Some authors suggested that the presence of a
"second segment" i s to be correlated to the lack of a stomach and peptic
digestion in order to account f o r the i n e f f i c i ë n t protein digestion in the
gut lumen (Yamamoto, 1966; Gauthier & Landis, 1972; Noaillac-Depeyre & Gas,
1976). I t is of i n t e r e s t that Shcherbina & Sorvatchev (1969) found a protein
absorption of 78 %in carp. In f i s h with a stomach, even higher percentages
j f ingested protein may be absorbed (Kapoor et a l . , 1975 ). I t should be
realized, however, that protein d i g e s t i b i l i t y can be correlated to the
;ame extend with the composition of the d i e t (Inaba et a l . , 1962, 1963).
(itamikado & Morishita (1965) found a protein d i g e s t i b i l i t y of 10% i f t r o u t
vere fed with soybeans. Aoe et a l . (1974) noticed that f o r carp hydrolysates
) f casein (amino acids and peptides) are far i n f e r i o r in n u t r i t i v e value to
intact p r o t e i n . Some native proteins seem d i f f i c u l t to be digestid by carp
[Jany, 1976).
K Relations between food preference and digestive
tract
Many authors t r i e d to correlate the morphology and physiological characters t i e s of the digestive t r a c t to the feeding habits of certain species,
teviews were presented by Suyehiro (1942) and Kapoor et a l . (1975 ). The
lossible relationships seem to be very complex. This applies in p a r t i c u l a r
21
to the lack of a stomach i n a number of t e l e o s t s : stomachless f i s h are supposed to be descendant from f i s h with a stomach and many
of them are believed to have adapted t h e i r d i e t s , as a r e s u l t of which there
are herbivorous, omnivorous and carnivorous stomachless fishes (Rogick, 1931;
Klust, 1940; G i r g i s , 1952; Kapoor et a l . , 1975 b ; Kafuku, 1977).
Many ecological studies indicate an apparent preference f o r one or several
types of food, but others point to the opportunistic feeding behaviour of
f i s h in periods of food s c a r c i t y .
Carnivorous, herbivorous, e t c . only
indicate general tendencies in feeding h a b i t s , and not c h a r a c t e r i s t i c
habits.
Only the length of the i n t e s t i n e seems to be c l e a r l y associated with the
feeding habits of a p a r t i c u l a r species. Some herbivorous and microphagous
stomachless f i s h have a r e l a t i v e l y long gut: 7 x to 24 x standard body
length (Rogick, 1931; G i r g i s , 1952; Das & Nath, 1965; Sinha & Moitra, 1975
'
; Kafuku, 1977). The gut length of omnivorous species l i k e Cyprinus
cavpio and Barbus aonahonius is 2-3 times the body length (Curry, 1939; Das
& Nath, 1965). Carnivorous species have the shortest gut lengths, approximately 0,5 - 0,8 times the body length (Klust, 1940; Das & Nath, 1965).
Khanna & Mehrotra(1971) found a r e l a t i v e l y short gut in 5 species of
carnivorous teleosts. A large v a r i a b i l i t y may be found w i t h i n a given
species, f o r example in the cyprinid Carasaius auratus (Vickers, 1962).
This seems to be related to the kind of food administered during the ontogeny.
The same is known from some cyprinodonts (Hykes & Moravek, 1933).
Therefore, the v a r i a b i l i t y is evidently less pronounced in stenophags
(Kapoor e t a l . , 1975 ). An additional problem i s that seasonal factors
(quantity of food?) may a f f e c t the gut length (Ciborowska, unpublished
results).
In some studies a r e l a t i o n is described between the d i e t and the a c t i v i t y
of several digestive enzymes. According to Sarbahi (1951) and Kitamikado &
Tachino (1961), carbohydrases are more abundant in the stomachless goldfish
than in the largemouth bass. Carbohydrases show the highest a c t i v i t i e s in
herbivorous f i s h (Vonk, 1927; Al Hussaini, 1949 ; Agrawal e t a l . , 1975).
Turpayev (1941) found t r y p t i c a c t i v i t y to be dominant i n a carnivorous
cyprinid (Asp-ius) , and in the herbivorous Soavdinus the amylase a c t i v i t y
was evident. Agrawal et a l . (1974) noticed a higher peptidase a c t i v i t y in
omnivorous than in herbivorous species. Kawai & Ikeda (1972) found a r e l a t i o n
22
of the a c t i v i t y of amylase and protease in carp to the composition of the
food. According to Sinha (1978), protease and lipase are the major digestive
enzymes in Cirrhinus
mrigala larvae, which are zooplankton feeders. In juve-
niles (omnivorous) and adults (herbivorous) strong amylase and weak protease
a c t i v i t i e s have been observed.
Of special i n t e r e s t is the development of the digestive t r a c t and i t s
functions during ontogeny, as most f i s h larvae are f i r s t carnivorous, feeding
on zooplankton. The food regime of omnivorous and herbivorous species changes
during ontogeny. This may be correlated with changes in the morphology and
physiology of the digestive apparatus.
23
GENERAL INTRODUCTION
C :Objectivesofour investigations.
Fromthestartofthis studywehave realized that some fish change their
feeding habits during development. Grasscarp larvaewere supposed tobe
zooplankton feeders,justasmost fish larvae,whereasthe adults were
called herbivorous (Gidumal, 1958;Aliyev&Bessmertnaya, 1968;Fisher,
1968). The grasscarp mightbeofspecial importancetoagriculture becauseit
feedsonandthus controls water weeds (Cross,1969;Stott &.Robson,1970;
Opuszynsky, 1972;Michewiczetal., 1972).Thespeciesmightbealso valuable
for consumption,asithasahigh growth rate (Hickling,1960;Anderson, 1970).
The selectionofgrasscarpforthe present study wasnotbased primarilyonthe
absenceofastomach.
The first objective wastofind morphological evidencefor theadaptationof
the intestinetothe changing feeding habits during ontogeny.Forthis purpose
the experiment described inpaperAhas been carried out.Theresultswerenot
encouraging,butour attention was attracted tothedistinct regional differentiationofthe intestineinthis stomachless species.
Because,justasinmammals,theintestinal epithelium incyprinidsisregularly renewed (Vickers,1962;Hyodo,1965;Gas&Noaillac-Depeyre, 1974),the
speciesmightbeusedfor studying thedifferentiationofenterocytes withinth
successive segments. PaperBdealsmainly withtherenewal ofthe gutepitheliu
and themorphologyofabsorptiveanddifferentiating cells. Many interesting
results have been obtained,butthe unexpected discoveryoffunctional cellsin
the proliferative area indicated thattheintestinal epithelium isnotthemost
useful tissuefor studyingthe differentiationofabsorptive cells.
Itappeared thatjustasinother cyprinidsasegmentwiththe abilityof
pinocytosisispresentinthe caudal partofthe gut,andthismightberelated
tothelackofastomach. Firstithadtobeascertained whether active absorption takes placeintherostral intestinal segmentandabsorptionofmacromoleculesbypinocytosismayberestricted tothesecond partofthegut.Inadditi
the absorptive activity along themucosal foldshadtobeinvestigated. Therefc
the experiments described inpaperChave been carried out.Totestwhetherthe
abilitytoabsorp protein macromoleculesisrelated tothelackofastomach,t
locationofprotein absorption inthe grasscarp intestinehadtobedetermined
(paperD ) .
24
The development of the i n t e s t i n e in a teleost (Clarias
lazera) in which
a stomach is developed at the end of the larval stage i s described in
paper E. I t was investigated whether the development of the stomach might
be related to the disappearance of pinocytosis in the second gut segment.
25
GENERAL DISCUSSION AND SUMMARY
General morphology of the digestive
tract
Structurallyandfunctionally the digestive tractoffishes hasmuchin
common with thatofother vertebrates,butinfishesthesystemissimplified.
Salivary glands areabsent,justasesophageal glands.Approximately 15%ofthe
species doesnothaveastomach.Theintestineisless complexduetothe absenceoffoldsofKerkringandreal villi. Furthermore,most fishes lackthecrypts
of Lieberkühn,the glandsofBrunnerandamuscularismucosae,thereisno
differentiation insmallandlarge intestine,andinmany speciesarectum
cannotberecognized.
For these reasons fishes appeartooffer several advantagesforstudying morphological andphysiological aspects,ofthedigestive system.
Food,preference
and the need for animal
protein
Another advantageisthatmany species change their food preference during
their life.Anexampleisthechinese grasscarp, Ctenopharyngodon idella, the
animal used forour experiments. PaperAdealsmainly with changesinthemorphologyoftheintestineinrelationtogrowthandchanging dietofthe young
grasscarp.
Animal food was foundtostimulate rapid growth,also after theanimals becom
capableofingesting plantmaterial.The composition ofthefood protein,especiallyastotheessential aminoacids maybethemain factor determiningthe
effectivenessofthe food.Many plantmaterialsareknowntocontain only small
amountsofmethionineandlysineandmuch vegetable material inthefood might
causeanimproper amino acid balance (Shcherbinaetal. 1964,quotedbyPhillips
1969)andconsequentlyalowgrowth rate.Grasscarp,keptinponds forweed
control,mayformaseriousthreattoother speciesasapredatoraswell asa
competitorforfood.
Length and regional
differentiation
of the gut
The digestionand absorptionofplant foodingrasscarp ismainly facilitated
byanincreaseofthe relative lengthofthe intestine from + 0,7toabout twic
the bodylength.Thelatterwas foundin6monthsoldspecimensandalsoinadult
(paper D ) ,andislowforapresumably herbivorous species.
26
Inmany other fish species alsoachangeinfeeding habits takes placeduring
evelopment.Thisisalways relatedtoanincreaseingutlength from+0,5x
odylengthinthecarnivorous larvaetomuch highervaluesinomnivorousand
erbivorous juvenilesandadults.Infig.7thegutlengthsofseveral Cyprinids
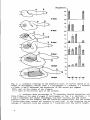
regivenatanumberofstagesofdevelopment.Intheherbivorous Catla oatla
hemost rapid increaseinrelativegutlengthwas found,whilethegrasscarp
howsaslower increaseingutlength than themirror-carp, Cyprinus
oarpio,
presumably omnivorousspecies.
5-,
^
X
I-
LU
z
>
—I
UJ
* /
AO
10
BODY LENGTH (cm)
7. GraphshowingtherelativelengthofthegutofsomeCyprinidlarvaeat
severalages.FromStroband,H.W.J.&Dabrowski,K.R.,in:"Nutrition
desPoissons"C.N.E.R.N.A.Paris (inpress).
O-commoncarp,»-grasscarp (Ctenopharyngodon idella) ,A- silvercarp
(Hypophthalmichthys molitrix) ,A-bighead (Aristiohthys nob-ilia)(after
Ciborowska,unpublished data),*- common carp (after Klust, 1940),
•-grass carp (afterStroband,1977)jjCatla aatla (after Kafuku, 1977).
27
Themorphology oftheintestinal epithelium ofthestomachless grasscarpis
described inpaperAand,inmoredetail,inpaperB.Thegutshowsasimilar
regional differentiation asinother cyprinids (Yamamoto,1966;Gauthier&
Landis,1972;Noaillac-Depeyre&Gas,1974,1976).Theanterior segment shows
thecharacteristics oflipid absorption;inthe"second segmenfenterocytescon
tainmany pinocytotic vesiclesandoneormore supranuclear vacuoles probably
representing large secundary lysosomes.Athird caudal segment contains entere
cyteswith thecharacteristics ofwateroriontransport.
The length oftheintestine isalso affected bythediet during early life.
Vegetable food causesaslight increase ingutlength,whichwasalso foundir
other species.Ourresults indicate thattheanterior segmentisinvolved int
growth,andthismight well berelated tothemain absorptive function ofthis
partoftheintestine (paperD ) .
The intestinal
epithelium as a cell renewal system
The regional differentiation andtherelatively lesscomplex structureof1
mucosa seem tomakethefish intestine auseful model forstudying thediffère
tiationofepithelial cells.InpaperBthecell renewal systemofthegutepi
theliumofthegrasscarp isdescribed with light-microscopic radioautography,
3
using Hthymidine.Thesystem appeared tobecomparable tothatinthemammalian small intestine;proliferation takes place inthebasal partsofthemucc
sal folds infishes,andinthecryptsofLieberkühn inthesmall intestineo1
mammals.Therenewal time inthegrasscarp isrelatively long:10-15daysat
20°C.InpaperBtheultrastructureoftheintestinal epithelium isdescribee
for starved andfedspecimens.
Functional absorptive cells proved tobepresent intheproliferative area
whereas undifferentiated cells could notbeidentified. This isamajordifferenceinrespecttomammals,inwhich undifferentiated proliferative cellsin
the crypts only become functional aftermigration towards theintestinal villi
(Rijke, 1977).
A comparison ofradioautoqraphs andelectronmicrographsforlarvaeof Barbus
aonchonius (Romboutetal.,1980)and Clavias
lazera (paperE)shows thatfuni
tional absorptive cellsarecapableofproliferation. Similar resultswerefoi
inthelarvaeoftheamphibian Xenopus laevis (Marshall &Dixon, 1978).The
difference ofthecell renewal system inmammals andfishes isalso reflected
the presenceofalkaline phosphatase activity inthemicrovillous borderof
enterocytes intheproliferative area ofthegrasscarp (paper C ) ,which also
28
ndicatesthecellstobecapableofabsorption.
Intracellular factors,relatedtothestateofdifferentiation,and extraellular factors (chalones,hormones,stimuli from the food)are possiblyinolvedindeterminingtherateofproliferationofvertebrate cells. Inhibition
f proliferationbyintracellular factors possibly develops onlywhen thecells
ave reachedarelatively high stateofdifferentiationinfishes.Inthis
onnectionitisinteresting thatinthecolonofmammals functional cellsand
ndifferentiated proliferative cellsareintermingled (Lipkin, 1973). This imliesamajor effectofthe intracellular factorsinblocking cell divisionin
nedifferentiated cells.
he digest-ion
moment
and absorption
of protein
and the function
of the second gut
An important factoristheregional differentiationofthe grasscarp intestine,
idespecially the presenceofa"second segment"with pinocytotic activity.This
isalso found aftertheingestionofexogenous foodinother stomachless fishes
idinfish larvae,which usually are initially stomachless (Tanaka,1969;
»perE ) .
PaperCdealswith the activityofalkaline phosphataseandtheuptakeof
'allyadministered horseradish peroxidaseintheintestineofgrasscarp larvae
idjuveniles.Aproximo-distal gradientinalkaline phosphatase activitycan
;demonstrated.This suggests that themain active absorption takes placein
ieanterior gut segment. Peroxidase was demonstrated histochemicallyinentero'tesofonly the second segment.This implies thatthe enzymemust have been
»sorbedbypinocytosis.Similar results have been found inother Cyprinids
loaillac-Depeyre&Gas,1973)andinClarias lazera larvae (paperE ) .
The enterocytesinthesecond segmentofthe grasscarp have also theability
»pinocytosisoflarge ferritin particles (paper D). Since pinocytosisofmacro»leculestakes place alsointheintestineofsuckling mammals before the stolenfunctionsarefully developed, itiswidely believed thatthepresenceof
"second segment"instomachless fishesisrelatedtothe lackofpepticdiistion.Recently,however,a"second segment"was also found intheintestine
r
the perch,afish withastomach (Noaillac-Depeyre&Gas,1979): this segment,
wever,isrelatively small (+11 %ofthegutlengthand+20%infish larvae
d stomachless fishes).
ShcherbinaandSorvatchev (1969)andShcherbinaetal. (1976),whousedthe
ertmarker technique,discovered thatmost proteinisabsorbed intheanterior
29
60 %ofthe carp intestine,i.e.inthe anterior gut segment.Intheirexperiments Shcherbinaetal. pooledthegut contentsof6specimens.Therefore, the
locationofprotein absorption inthe grasscarp intestine has been determined
alsoinindividual test animals.Theresultsaredescribed inpaperD.Itwill
beproved that proteins arealmost completely absorbed within the anterior
40-50 %ofthegut i.e.intherostral two thirdofthe first segment.This
implies adequate digestionofproteininthegutlumen,sincenonoticeable
pinocytosisof macromoleculeswas found intheanterior segment.Ourresults
also indicate thatessential amino acids are preferentially absorbed inthe
grasscarp,justasinmammals (Orten,1963;Ben-Ghedaliaetal., 1974).The
rapid disappearanceoflysineandarginine from the chyme suggestsatryptic
breakdownofproteins.
In their paperonthe perch,Noaillac-Depeyre&Gas (1979)suggested that
protein digestion inthegut lumenoffishmightbeless efficient thanin
mammals. They suggest that completionofhydrolysis takes placebyintracellular enzymesinthesecond segment.Our results prove thistobeincorrect
proteins are digestedandabsorbed adequately withinthe anterior gut segment,
butanexception should possiblybemadefor fish larvae,inwhich food reache
the distal partofthe intestine very shortly after feeding (Fänge&Grove,
1979).
Under physiological circumstances pinocytosisofafewmacromolecules undoub
ly takes place.Thismayberelevantinaqualitative sense.Itmaybecompara
tothesituation inadultmammals,inwhich macromolecules areabsorbed insma
quantities andmaybeantigenicorbiologically active (Walker&Isselbacher,
1974; Warshawetal., 1974).
Similarity between the anterior gut segment in fish and the small
in mammals
intestine
Astoitsabsorptive functions,the anterior intestinal segmentoffishesY
muchincommon with the small intestineofmammals.Mammals also showaproxin
distal gradientinalkaline phosphatase activity,andinboth classesofverte
brates sugars,lipidsandprotein aremerely absorbed inthe anterior partof
thegut(Belletal., 1972;Crampton,1972;Farmanfarmaianetal., 1972;Shche
binaetal., 1973;1976;1977;BenGhedaliaetal., 1974;Noaillac-Depeyre&
Gas, 1976;paper B ) .Themorphological characteristicsoflipid absorptionare
similarinmammalsandfishes,andinboththetransportofsugarsandaminoacidsissodium-dependent (Cartieretal., 1979).
30
The development
of the stomach in Glorias
lazera
The digestive systemoffish larvae resembles thatofstomachless teleosts
inmany respects:a"secondgutsegment"mayberecognized immediately afterthe
beginningofexogenous feeding.Thisisalso trueforfish species that develop
a stomachattheendofthelarval period.
In paperEthe resultsare described ofanother attempttoestablishapossible relation betweenthe presenceofa"second segment"andthelackofastomach.The developmentofthe stomachinclarias lazera isdescribed,andits
ultrastructureiscompared tothatofadult specimens.Itisofinterest that
the stomach corpus glands contain only one cell type,which apparently is involvedinproducing hydrochloric acidandpepsinogen. From theultrastructure
3
of the glands,the H-thymidine labeling index,andthepHindicator tests,it
isconcluded thatafunctional stomach ispresent from approximately 12days
after fertilization. However,inlarvaeaswell asinjuvenile Cl. lazera a
"second segment"ofapproximately 20%ofgut length shows theabilitytopinocytosisofhorseradish peroxidase.Thesamemay applytoadult specimens,since
they also possessa"second segment"of20%ofgut length,asproved with lightand electron-microscopic studies.Therefore,the conclusion seems justified,
thatthe abilitytopinocytosisofmacromoleculesisnotpositively correlated
tothelackofafunctional stomach.Our results also indicate thataregional
differentiationofthe intestine,includingagutsegmentwith pinocytosis,is
a general featureofthedigestive tractinteleosts.
Function of the stomach in
vertebrates
Since proteins aredigested adequately intheintestineofstomachless fishes
-although food passes throughthegutinnotmore than about7hinthegrasscarp (Hickling, 1966)-thequestion ariseswhether peptic digestion inanacid
environment,asfound inother vertebrates,isinfactofminor importance.
AccordingtoBertin (1958),most stomachless teleosts possess some kindofmasticatory apparatus,likethepharyngeal teethinCyprinids. Fishes without this
apparatusaresupposedtoneedastomach withalowpHtofractionate the food.
Botharetherefore believedtofacilitate feeding with relatively largeorganisms.
Itiswidely believed thatthestomach evolved togetherwiththe jawsbutprimarilyasastorage organ.Acid productionmayhave followed,andacted infractioning large food elementsandpossiblyinprotectingthestored food against
bacterial digestion.The productionofpepsinfor thedigestionofproteinsin
31
the acid environment might have been the last important step in the evolution
of the stomach (Barrington, 1957;Romer,1962;Waterman et al., 1971). From
phylogenetic studies it isgenerally believed that the absence of astomach in
teleosts isa secundary feature (Barrington, 1942).
CONCLUSIONS
1.The grasscarp isastomachless teleost.The intestine does not containmulticellular glands.
2.The relative length of the grasscarp intestine increases from 0,7 x body
length inyoung larvae to 2xbody length inadults.The gut length is the
onlymorphological characteristic to changewhen the grasscarp turns from
carnivorous to herbivorous feeding.
3.After feeding thegrasscarp with vegetable food,the increase ingut length
is higher than in fishes fed with animal food.
4.Animal food stimulates rapid growth in the grasscarp,also after they are
able to ingest plantmaterial. Grasscarp may represent a serious threat
to other species as a predator aswell as acompetitor for food.
5.The intestine of the grasscarp shows a similar regional differentiation as
found inother Cyprinids,with aproximo-distal gradient inalkaline phosphatase activity.
6.The anterior gut segment is involved in the absorption of lipids and proteins
(and probably also of sugars),which are absorbed inenterocytes after hydrolysis in the lumen.Also in regard to themorphological characteristics of
lipid absorption,the epithelium shows resemblance with the epithelium in
the small intestine ofmammals.
7. A"second gut segment",running from +60-85 %of gut length, is characterized
by enterocytes with a high pinocytotic activity. These cells are capable of
absorbing protein macromolecules like horseradish peroxidase and ferritin.
8. Protein digestion isefficient despite the absence of a stomach. Quantitatively relevant amounts of protein are not absorbed by the"second segment".
9. The "second segment" isnot related to the lack of peptic activity in stomachless fishes,since it isalso present in clavias lazera after the stomach
has developed and has become functionally active.
10. The stomach of Clavias lazera contains only one type of corpus glandular
cellswith themorphological characteristics of chief-aswell as parietal
cells.
32
1.The intestinal epithelium ofthegrasscarp represents acell renewal system
and iscompletely renewed within 10-15 days.
2. Incontrast tomammals the proliferative pool ofcells consists of functional cells ingrasscarp larvae andjuveniles and in Clavias lazera larvae.
33
REFERENCES
Agrawal,V.P., Sastry, K.V. &Kaushab, S.K.S. (1975).Acta Physiol.
Acad. Sei. Hung 46, 93-98.
Al Hussaini,A.H. (1949). Quart. J. Micr. Sei. 90, 323-354.
Al Hussaini,H.A. (1964). Bull. Inst. Egypt. 40, 23-32
Aliyev, D.S. &Bessmertnaya, R. Ye. (1968).
Amer. Fish. Soc. 8, 319-321.
Alliot, E.Febvre,A. &Métailler,R. (1974). Ann. Biol. anim.
Biophys. 14, 229-237.
Anderson, E.N.Jr. (1970). J. Trop. Geograph. 30, 11-16.
Aoe,H., Ikeda,K. & Saito, T. (1974) Bull. Jap. Soc. Sei. Fish.
40, 375-379.
Arvy, L. (1960). C R . Soc. Biol. (Paris) 154, 313-315.
Babkin, B.P. &Bowie, D.J. (1928). Biol. Bull. 54, 254-277.
Balon,E.K. (1975). J. Fish Res.Bd. Can. 32, 1663-1670.
Bannister, L.H. (1966). Parasitology, 56, 633-688.
Barrington, E.J.W. (1942). Biol.Revs.17 , 1-27.
Barrington, E.J.W. (1957). In: "The Physiology of fishes" (M.E. Brown,ed.).
Vol. I, 109-161.Acad. Press.New York.
Bell,G.H., Davidson, J.N., Emslie-Smith,D. (1972):"Textbook of physiology
and biochemistry".Churchill Livingstone, Edinburgh, London.
Ben-Ghedalia, D., Tagaris H., Bondi,H., Tadmor,A. (1974).Br. J. Nutr. 32,
125-142
Berry, P.Y. &Low,M.P. (1970). Copeia, 1970, 708-726.
Bertin, L. (1958). In: "Traité de Zoologie" (Grasse, P.P. ed.)vol. XIII 1248.
Masson, Paris.
Bishop, C. &Odense, P.H. (1966). J. Fish.Res. Bd. Can. 23, 1607-1617.
Blake,I.H. (1936). J. Morphol. 60, 77-102.
Bondi,A. &Spandorf, A. (1954). Brit. J. Nutr. 8, 240-246.
Broussy, J. &Serfaty, A. (1958). Bull. Soc.Hist.Nat. Toulouse 93,
81-85.
Buchs, S. (1954). Z. Vergl. Physiol. 36, 165-175.
Bucke,D. (1970). J. Fish. Biol. 3, 421-423.
Bullock, W.L. (1963). J. Morphol. 112, 23-44.
Cartier, M., Buclon,M., Robinson, J.W.L. (1979). Comp. Biochem. Physiol.
62A, 363-370.
Catton, W.T. (1957). Blood 6, 39-60.
Chitray, B.B. (1965). Ichthyologica IV (i),53-62.
Ciullo, R.H. (1975). In: :The pathology of fishes" (W.E.Ribelin and G.
Migaki eds.). Univ. Wisconsin Press, Wisconsin.
Clark, S.L. (1959) J. Biophys. Biochem. Cytol. 5,41-50.
Cockson, A. &Bourn, D. (1973). Hydrobiologia 43, 357-363.
Crampton, R.F. (1972). In: "Peptide transport inbacteria and mammalian gut".
Ciba Foundation Symposium 11nov. 1971.Ass. Publ., Amsterdam.
Creach, P.V. (1963).Ann. Nutr.Alim. 17, 375-471.
Cross,D.G. (1969). J. Fish. Biol. 1, 27-30.
Curry, E. (1939). J. Morphol. 65, 53-78.
Dabrowski, K. &Glogowski, J. (1977). Hydrobiologia 52, 171-174.
Das, S.M. &Nath, S. (1965). Ichthyologia 1-2, 63-79.
Davina, J.H.M.,Rijkers, G.T., Rombout, J.H.W.M.,Timmermans, L.P.M.,
Muiswinkel van,W.B. (1980). In: "Development and Differentiation of
Vertebrate lymphocytes", J.D. Horton (ed.). Elsevier/North Holland
Biomedical Press.
Dhaliwal, R. (1975). Sei. Cult. 41, 523-524.
34
Fahrenholz, C. (1937). In: "Handbuch der vergleichenden Anatomie der Wirbeltiere" (L.Bolk, E.Goppert, E.Kallius &Lubosch eds.) Vol.III.
Urban und Schwärzenberg, Berlin und Wien.
Falge, R. &Shpannkhof, L. (1976). J. Ichthyol. 16, 672-677.
Fänge, R. &Grove, D. (1979). In:"Fish Physiology" (W.S.Hoar, D.J.
Randall and J.R. Brett eds.) Vol. VIII, 161-260.Acad. Press,New York.
Farmanfarmaian, A., Ross,A. &Mazal, D. (1972). Biol. Bull. 742,427-445.
Fisher, Z. (1968).Pol.Arch. Hydrobiol. 15, 1-8.
Gas,N. &Noaillac Depeyre, J. (1974).C R . Acad. Sei. 279, 1085-1088.
Gauthier, G.F. &Landis, S.C. (1972). Anat. Ree. 172, 675-701.
Gidumal, J.C. (1958). Hong Kong Univ. Fish. J. 2, 1-6.
Girgis, S. (1952). J. Morphol. 90, 317-362.
Goel,K.A. (1974).Acta Histochem. 51, 13-17.
Gossrau.R. (1975). Acta Histochem. Cytochem. 8, 153-163.
Grünberg, W. &Hager, G. (1978). Anat. knz.143 ,277-290.
Haie, P.A. (1965). J. Zool. 146, 132-149.
Hickling, C F . (1960). Malay Agric. J. 42, 49-53.
Hickling, C F . (1966). J. Zool. 140, 408-419.
Hirji,K.N. &Courtney, W.A.M. (1979). J. Fish. Biol. 15, 469-472.
Hykes,0.V.&Moravek,Fr.(1933).C R . Soc. Biol.Paris 113, 1239-1241.
Hyodo,Y. (1965). Rad. Res. 26, 383-394.
Inaba, D.,Ogino, C , Takamatsu, C , Sugano, S. &Hâta,H. (1962).
Bull. Jap. Soc. Sei. Fish.20 ,367-371.
Inaba, D., Ogino, C , Takamatsu, C , Ueda, T. &Kurokawa,K. (1963).
Bull. Jap. Soc. Sei. Fish. 29, 242-244.
Ishida, J. (1936). Annot. Zool. Jap. 15, 263-284.
Iwai, T. (1967*). Bull. Jap. Soc. Sei. Fish. 33, 1116-1119.
Iwai, T. (1967 ) .Bull. Jap. Soc. Sei.Fish. 33, 489-496.
Iwai, T. (1968). Bull. Jap. Soc. Sei. Fish. 34, 973-978.
Iwai, T. (1969). Arch. Histol. Jap. 30, 183-189.
Jacobshagen, E. (1937). In: "Handbuch der Vergleichende Anatomie der Wirbeltiere" (L. Bolk., E. Göppert,E. Kallius and W. Lubosch. eds.). Vol. III,
563-724. Urban und Schwarzenberg, Berlin und Wien.
Jany, K.D. (1976). Comp. Biochem. Physiol. 53 B, 31-38.
Kafuku, T. (1977). Bull. Freshwater Fish. Res.Lab. (Tokyo) 27, 1-20.
Kapoor, B.G., Evans,H.E., & Peuzner, R.A. (1975 ) .In: "Advances in Marine
Biology Vol. 13 (F.S.Russell and M. Yonge, eds.). pp. 53-108
Acad. Press., London.
Kapoor, B.G., Smit,H. &Verigina, I.A. (1975 ) .In:"Advances inMarine BiologyVol. 13 (F.S.Russell and M.Yonge,eds.)pp. 109-239.Acad.Press.London.
Kenyon,W.A. (1925). Bull.U.S. Bur. Fish.,Washington, 4J, 181-200.
Kawai, S. &Ikeda, S. (1971). Bull. Jap. Soc. Sei. Fish. 37, 333-337.
Kawai, S. & Ikeda, S. (1972). Bull. Jap. Soc. Sei. Fish. 38, 265-270.
Khalilov, F. Kh. (1969). "Materials onmorphology and Histochemistry of the
digestive system in teleosts"- Maktop Alma Ata (InRussian).
Quoted by Kapoor et al., 1975) .
Khanna, S.S. &Mehrotra, B.K. (1971). Anat. Anz. 129, 1-18.
Kitamikado, M. &Tachino, S. (1961). Chem. abstr. 55, 5789 a,b, c.
Kitamikado, M. &Morishita T. (1965). Chem. abstr. 62, 15129 d, e.
Klust, G. (1940). Int. Rev. Hydrobiol. Hydrograph. 40, 88-173.
Korovina, V.M. (1976). J. Ichthyol. 16, 617-624.
Kraehenbuhl, J.P. (1969). J. Cell Biol. 42, 345-365.
Krementz, A.B. &Chapman, G.B. (1974). J. Morphol. 145, 441-482.
Leino, R.L. (1974). Cell Tiss. Res. 155, 367-381.
Leissring, J . C , Anderson, J.W. & Smith, D.W. (1962). Amer. J. dis. Children
103, 160-165
35
Lipkin,M. (1973). Physiol. Rev. 53, 891-915.
Lubyanskiene, V., Jankevicius,K., Trepsiene, 0. &Zableckis, J. (1977).
Liet. TSR.MoksluAkad. Darb. Ser. C.Biol.Mokslai
1 (77), 87-92.
Marshall, J.A.,Dixon,K.E. (1978) J. Exp. Zool. 203, 31-40
Mattey, D.L., Morgan,M. &Wright, D.E. (1979). J. Fish. Biol. 15, 363-370
McVay,J.A.&Kaan,H.W. (1940). Biol.Bull. 78, 53-67.
Merrillees, M.J. (1974). J. Ultrastr. Res. 47, 272-283.
Michewicz, J.E., Sutton, D.L. &Blackburn,R.D. (1972). Weed Science 20, 106110.
Migita, M. &Hashimoto, Y. (1949). Bull. Jap. Soc. Sei. Fish. 15, 259-261.
Mohsin, S.M. (1962). Acta Zool. (Stockholm). 43, 79-133.
Moitra, S.K. &Ray,A.K. (1977).Anat.Anz. 141, 37-58.
Noaillac-Depeyre, J. &Gas,N. (1973^ .Z. Zeilforsch 146, 525-541.
Noaillac-Depeyre, J. &Gas,N. (1973 ) .C R . Acad. Sei. (Paris) 276, 773-776.
Noaillac-Depeyre, J. &Gas,N. (1974). Cell. Tiss.Res. 153, 353-365.
Noaillac-Depeyre, J. &Gas,N. (1976). Tissue & Cell.S, 511-530.
Noaillac-Depeyre, J. &Gas,N. (1978). Tissue &Cell. 10, 23-37.
Noaillac-Depeyre, J. &Gas,N. (1979). Anat. Rec. 195,
621-640.
Opuszynski, K. (1972). Aquaculture 1, 61-74.
Orten, A.U. (1963). Fed. Proc. 22, 1103-1109.
Paris,H. Murat, J.C. &Castilla, C. (1977). C R . Acad. Sei. (Paris) 171,
1297-1301.
Patton, J.S. (1975).Lipids, 10, 575-583.
Philips,A.M. Jr. (1969). In: "Fish Physiology". (Hoar,W.S. &Randall, D.J.,
eds.)vol.1, 391-432.Academic Press,New York and London.
Reutter, K., Breipohl,W., Bijvank, G.J. (1974). Cell Tiss. Res. 153, 151-165.
Rogick, M.D. (1931). J.Morphol. 52, 1-25.
Rombout, J.H.W.M.,Stroband, H.W.J.,Verstijnen, C.P.H.J.:Proliferation and
differentiation of intestinal epithelial cells during development of
Barbus oonohonius
{Teleostei,
Cyprinidae).
Submitted toJ.Exp.Zool.
June 1980.
Romer,A.S. (1962). The Vertebrate body. Saunders, Philadelphia, London.
Rijke, R.P.C. (1977). Control mechanisms of cell proliferation in intestinal
epithelium. Thesis, Erasmus University, Rotterdam.
Sacquet, E.,Lesel, R., Mejean, C Riottot,M. &Leprince, C (1979). Ann.
Biol. anim. Biochem. Biophys. 19, 285-391.
Sarbahi, D.S. (1951). Biol. Bull. 100, 244-257.
Sastry, K.V. (1974H. Acta Histochem. 48, 320-325.
Sastry, K.V. (1974 ) .Acta Histochem. 51, 18-23.
Sastry, K.V. (1975). Anat. Anz. 137, 159-165.
Sastry, K.V. &Garg,V.K. (1976). Z.Mikrosk. Anat. Forsch. Leipzig 90,
1032-1040.
Shcherbina, M.A. (1973). J. Ichthyol. 13, 104-111.
Shcherbina, M.A. &Kazlauskene,0.P.(1971).Vopr. Ichtiol. 11, 103-108.
Shcherbina, M.A. &Sorvatchev, K. (1969).Vopr. Prud. Rubov. 16, 315-322.
Shcherbina, M.A., Trofimova, L.N. &Kazlauskene, 0. (1976). J. Ichthyol.
16, 632-636.
Shcherhina, M.A., Shcherbina, T.V., Kazlauskene, O.P. (1977). J. Ichthyol.
17, 327-331.
Sinha,C M . (1976*). Z.Mikrosk. Anat. Forsch. Leipzig 89, 294-304.
Sinha, C M . (1976 ) .Anat. Anz. 139, 348-362.
Sinha,C M . (1978). Zool. Beitr. 24, 349-358.
36
Sinha,G.M. &Moitra, S.K. (1975,). Anat.Anz. 138, 222-239.
Sinha, G.M. &Moitra, S.K. (1975 ) .Anat. Anz. 137, 395-407.
Sivadas, P. (1964). J. Cell, and Comp. Physiol. 65, 249-254.
Smit,H. (1968). In: "Handbook of Physiology sect. 6Alimentary canal"
(F.C. Code, ed.).Vol.V., 2791-2805. Am. Physiol. Soc. Washington D.C.
Smyth, D.H. (1971). In:"Peptide transport inbacteria and mammalian gut"
Ciba Foundation symposium 11nov. 1971,ass. Sei.Publ. Amsterdam.
Srivastava, A.K. (1966). Curr. Sei. 35, 154-155.
Staley, T.E., Corley, L.D. Bush, L.J. &Jones,E.W. (1972). Anat. Rec. 172,
559-579.
Stott, B. &Robson, T.O. (1970). Nature, 226, 870.
Suyehiro, Y. (1942). Jap. J. Zool. 10, 1-303.
Syvokiene, J. Jankevicius,K., Lesauskiene, L. &Antanyniene, A. (1974).
Liet. TSR.Mokslu Akad. Darb. Ser. C. Biol.Mokslai 1 (65) 143-150.
Syvokiene, J. &Jankevicius, K. (1976). Liet. TSR. Mokslu Akad. Darb. Ser.
C. Biol.Mokslai 2(74), 143-150.
Syvokiene, J. & Jankevicius, K. (1977).
Liet. TSR. Mokslu Acad. Darb. Ser.
C. Biol.Mokslai 2 ( ) 79-86.
Tanaka, M. (1969) Jap. J. Ichthyol. 16, 141-149.
Tanaka, M. (1971). Jap. J. Ichthyol. 18, 164-174.
Tanaka, M. (1972). Jap. J. Ichthyol. 19, 15-25.
Turpayev, T.M. (1941). Bull. Exp. Biol. Med. Moscow 12
46-48.(in Russian)
quoted by Kapoor et al., 1975) .
Ugolev,A.M. (1971). In: "Peptide transport inbacteria andmammalian gut"
Ciba Foundation symposium 11nov. 1971,Ass. Sei. Publ. Amsterdam.
Verigina, I.A. (1976). J. Ichthyol. 16,
-95.
Verigina, I.A. ( 1 9 7 8 ) . J. Ichthyol. 17, 424-431.
Verigina, I.A. (1978 ) .J. Ichthyol. 17, 964-968.
Verma, S.R. &Tyagi,M.P. (1974). Morph.
Jahrb. 120, 244-253.
Vickers, T. (1962). Quart. J. Micr. Sei.
103, 93.
Vonk, H.J. (1927). Physiologie 5, 445-546.
Walker, W.A., Isselbacher, K.J. (1974). Gastroenterol. 67, 531-550.
Warshaw, A.I.,Walker, W.A., Isselbacher, K.J. (1974). Gastroenterology 66,
987-992.
Waterman, A.J., Frye,B.E., Johanssen, K., Kluge,A.G.,Moss,M.L., Noback,
C R . , Olsen, I.D., Zug, G.R. (1971) "Chordate Structure and Function".
Mac Millan Company, New York.
Weinberg, S. (1975). Bijdr. Dierk. 45, 195-204.
Weisel, G.F. (1973). J. Morphol. 140, 243-256.
Yamamoto, T. (1966). Z. Zeilforsch. 72, 66-87.
The
37
SAMENVATTING
De bouw van het spijsverteringskanaal van vissen is in het algemeen
minder complex dan b i j hogere vertebraten het geval i s . Zo ontbreken speeks e l k l i e r e n en gewoonlijk ook slokdarmklieren. Ongeveer 15% der vissoorten,
waaronder de karperachtigen, is maagloos. Wat de darm b e t r e f t v a l t vooral het
ontbreken van plooien van Kerkring, v i l l i met crypten en meercellige klieren
i n de darmwand op. Bovendien kan er geen onderscheid worden gemaakt i n dunneen dikke darm. Eén en ander heeft er toe geleid dat vissen een steeds belangr i j k e r plaats innemen i n het fundamenteel onderzoek naar vorm en f u n k t i e van
elementen van het s p i j s v e r t e r i n g s s t e l s e l .
De graskarper is als larve carnivoor en voedt zich met zoöplankton.
Tijdens z i j n ontwikkeling, w a a r s c h i j n l i j k al gedurende de eerste maanden,
verandert het dieet en aangenomen wordt dat de volwassen graskarper herbivoor
i s . Deze verandering l i j k t deze vissoort bijzonder geschikt te maken voor
onderzoek naar de r e l a t i e tussen de struktuur van de darm en het d i e e t .
U i t a r t i k e l A b l i j k t echter dat graskarpers, ook lang nadat ze voor het
eerst i n staat z i j n plantaardig voedsel te n u t t i g e n , het snelst groeien
wanneer ze met d i e r l i j k materiaal worden gevoed. De enige morfologische
verandering die werd waargenomen gedurende de periode waarin het dieet zich
zou wijzigen was een toename van de relatieve darmlengte van 0,7 t o t 2 maal
de lichaamslengte. Dit is tamelijk kort voor een vis die verondersteld wordt
herbivoor te z i j n , en w i j s t als zodanig meer op een omnivore levenswijze.
Dit kan betekenen dat graskarpers, wanneer ze massaal worden uitgezet ter bes t r i j d i n g van waterplanten, een bedreiging worden voor andere soorten, zowel
als predator alsook als konkurrent wat het voedsel b e t r e f t .
In p u b l i k a t i e A, en meer gedetailleerd i n a r t i k e l B, wordt de morfologie
van het darm epitheel beschreven. Er z i j n 3 darmsegmenten te onderscheiden.
Het meest rostral e segment, dat + 60% van de darm beslaat, bezit resorberende
cellen die dezelfde morfologische kenmerken van vetopname vertonen die ook i n
enterocyten i n de dunne darm van zoogdieren worden aangetroffen. In het middelste of tweede segment (+ 25% van de darmlengte) komen resorberende cellen
voor die veel pinocytotische a k t i v i t e i t vertonen. Het caudale of 3e segment
(+ 15%) speelt w a a r s c h i j n l i j k een rol b i j de osmoregulatie.
38
Wegens deze vormverschillen, die ook een regionale d i f f e r e n t i a t i e t . a . v .
•esorptie van bepaalde voedingsstoffen l i j k e n i n te houden (zo kunnen e i w i t :en in het tweede segment worden gepinocyteerd), werd de graskarper een ge;chikt object geacht om de d i f f e r e n t i a t i e van verschillende typen resorptie
:ellen te bestuderen.
In a r t i k e l Bwordt de vernieuwing van het darm epitheel beschreven. Deze
ili j k t r e l a t i e f traag te verlopen: 10 à 15 dagen b i j 20°C tegenover 2 à 3
lagen b i j de meeste zoogdieren. In de dunne darm van zoogdieren is de proifererende "pool" van cellen gelokaliseerd i n de crypten van Lieberkühn.
e bestaat u i t ongedifferentieerde c e l l e n , die zich d i f f e r e n t i e r e n tijdens
e migratie naar de v i l l u s , alwaar zich funktionele cellen bevinden. De
esultaten b i j de graskarper wijken in belangrijke mate af van d i t beeld.
aar crypten ontbreken, bevinden zich de prolifererende cellen aan de basis
3
an de darmplooien, zoals b l i j k t u i t de H-thymidine labeling van epitheel e l l e n , die alleen i n d i t gebied optreedt. Er worden hier echter alleen
unktionele cellen aangetroffen. In p u b l i k a t i e Ewordt aangetoond dat ook
i j Clarias lazera larven de p r o l i f e r a t i v e cel "pool" i n het darm epitheel
i t funktionele cellen bestaat. Bovendien b l i j k e n b i j deze vissoort f u n k t i o ele cellen u i t het maag epitheel i n staat te z i j n zich te delen. Terwijl
oogdiercellen al vroeg i n het d i f f e r e n t i a t i e proces, lang vóórdat ze
unktioneel worden, hun delingsvermogen verliezen, l i j k t de blokkering
an de mogelijkheid t o t p r o l i f e r a t i e b i j vissen pas i n een veel l a t e r staium van de d i f f e r e n t i a t i e op te treden. Eén en ander houdt in dat de grasarper zich minder goed leent voor het bestuderen van morfologische aspecten
an het d i f f e r e n t i a t i e proces van darm epitheel c e l l e n .
In a r t i k e l Cwordt aangetoond dat min of meer komplete protéine macroDleculen (mierikswortel peroxidase) i n het tweede darmsegment kunnen worden
=pinocyteerd. Terwijl i n het eerste segment de m i c r o v i l l i der resorptie
ï l l e n een hoge alkalische fosfatase a k t i v i t e i t vertonen, ontbreekt deze
-ijwel geheel in het tweede segment. Dit w i j s t er op dat a k t i e f transport
)or de apicale celmembraan vooral plaatsvindt i n het eerste segment.
Omdat ook pasgeboren zoogdieren de mogelijkheid t o t pinocytose van macro)leculen bezitten - vóórdat a l l e maagfunkties t o t ontwikkeling gekomen z i j n
wordt wel aangenomen dat deze mogelijkheid in verband staat met het onteken van een maag en vooral van peptische e i w i t v e r t e r i n g . Deze hypothese
lakte het wenselijk de e i w i t v e r t e r i n g en opname nader te onderzoeken.
ertoe werden de experimenten zoals beschreven in p u b l i k a t i e D uitgevoerd.
39
B i j de graskarper b l i j k t , dat de 75% van het voedsel e i w i t die worden opgenomen vrijwel volledig door de r o s t r a l e 40% van de darm worden geresorbeerd. Een k w a n t i t a t i e f belangrijke rol kan dus, wat de e i w i t opname b e t r e f t ,
n i e t aan het 2e segment worden toegeschreven. U i t de experimenten b l i j k t
tevens dat er een preferentie bestaat voor essentiële aminozuren, zoals dat
ook b i j zoogdieren het geval i s . De r e l a t i e f snelle resorptie van arginine
en lysine w i j s t op het belang van vooral tryptische vertering b i j de graskarper.
Een tweede manier om na te gaan of de aanwezigheid van een tweede segment
met pinocytotische a k t i v i t e i t i n r e l a t i e staat t o t het ontbreken van een
maag is het bestuderen van de ontwikkeling van het spijsverteringskanaal
van een maaghoudende v i s . B i j vislarven is de maag nog n i e t aangelegd, en
de darm vertoont dezelfde regionale d i f f e r e n t i a t i e als die van maagloze
vissen. In a r t i k e l Ewordt de vorming van de maag beschreven b i j de tropische
meerval Clarias
lazeva.
Een bijzonderheid ten aanzien van de struktuur van de maag van vissen i s ,
dat - i n tegenstelling t o t zoogdieren - slechts één type k l i e r c e l voorkomt
waarin zowel pepsinogeen als HCl wordt geproduceerd. Het tweede segment
b l i j k t na de vorming en het funktioneel worden van de maag te b l i j v e n bestaan. B i j larven én b i j adulte specimen van clarias
lazeva beslaat het
+ 23% van de darmlengte. Er bestaat dus geen r e l a t i e tussen de mogelijkheid
macromoleculen op te nemen en het ontbreken van een maag. U i t a r t i k e l D
b l i j k t bovendien dat de afwezigheid van een maag n i e t of nauwelijks van
invloed i s op de e f f i c i ë n t i e van de e i w i t v e r t e r i n g .
De funktie van het 2e segment b l i j f t dan ook o n d u i d e l i j k . B i j v i s l a r v e n ,
waarbij het voedsel zéér snel het achterste deel van de darm b e r e i k t , kan
het een k w a n t i t a t i e f belangrijke rol spelen b i j de eiwitopname, maar b i j
oudere vissen heeft de pinocytose van macromoleculen vermoedelijk alleen
kwalitatieve betekenis. Ook b i j zoogdieren worden kleine hoeveelheden e i w i t
als macromoleculen opgenomen. Deze hebben geen betekenis voor de voeding,
maar z i j n w e l l i c h t op één of andere wijze biologisch a k t i e f (bijvoorbeeld
als anti geen).
U i t het f e i t dat b i j de graskarper ondanks het ontbreken van een maag
en een v r i j snelle passage van het voedsel door de darm (+ 7 uur)toch een
zeer e f f i c i ë n t e e i w i t v e r t e r i n g plaatsvindt v a l t af te leiden dat de maag
met betrekking t o t de e i w i t v e r t e r i n g slechts van secundaire betekenis i s ,
of althans dat de productie van pepsine en/of zuur geen voorwaarde vormt
voor een adequate vertering van voedseleiwitten.
40
Reprinted from J. Fish Biol. (1977) 11,167-174
Growthanddiet dependant structural adaptations ofthe
digestivetract injuvenile grasscarp {Ctenopharyngodon
idella,Val.)
H. W. J. STROBAND
Agricultural University, Department of Experimental Animal
Morphology and Cell Biology, Hollandseweg 13, Wageningen
The Netherlands
{Accepted 8 June 1976)
The morphology of the intestinal epithelium is described and compared with published
data. In the gut of this stomachless fish, three zones can be distinguished based on light
and electron microscopic observations of absorptive cells: a rostral zone with numerous
fat droplets, a second zone with large P.A.S.-positive supranuclear vacuoles and pinocytoticvesicles,andathirdzonewithoutthesecharacteristics. Inthesezonesdeepinvaginations of the cell membrane is a feature of the basal part of the cells. The presence of
pinocytotic vesicles in the second zone isconsidered evidence of the uptake of undigested
protein-like substances.
Juvenile grass carp from 15to 335 days, fed on either animal or vegetable food, were
measured. The growth rates indicate that animal food stimulates rapid growth. The data
also show a vegetable diet causes a slight increase in relative gut length, mainly in the
length of thefirst zone.
I. INTRODUCTION
TheadultWhiteAmur or grasscarp(Ctenopharyngodon idellaVal.)eatslargequantitiesofaquaticplantsand asaresultiseffective inweedcontrol (Lin, 1935;Penzes&
Tolg, 1966;Fischer, 1968;Cross, 1969; Stott &Robson, 1970). Thefish also has a
high growth rate (Hickling, 1960; Anderson, 1970) which increases its economic
significance, and consequently the grasscarp has beenthe subject ofmany investigations.
Several authors have described the histology of the alimentary tract in Cyprinids
(Rogick, 1931;Curry, 1939;Klust, 1940;McVay &Kaan, 1940;AlHussaini,1949a,
b; Girgis, 1952). It appears to be difficult to relate histological and morphological
adaptations to the types of food. According to Nikolsky (1956, cited by Hickling,
1966)theyounggrasscarp iscarnivorous until reachinga length of 2-5cm;whenits
feedinghabitschangerapidly,anditreachestheherbivorousstageatalengthof3cm.
InseveralCyprinids,fat andproteinappear tobeabsorbed indifferent partsofthe
gut(Yamamoto, 1966;Iwai, 1968,1969;Gauthier &Landis, 1972; NoaillacDepeyre
& Gas, 1973). The location, relative length and histological features of these parts
werethe subject of our investigation.
II. MATERIALS AND METHODS
Anumberofgrasscarpfry(15 daysafter hatching)wereprovidedbythe'Organisatieter
Verbetering van deBinnenvisserij', Holland in February 1975. Onarrivalthefisheswere
167
168
H. W. J. STROBAND
dividedintothreegroups,allwerefed onTubificidae(theirownbodyweight aday). Group2
received additional plant material (young darnel). From the28th day after hatching(when
thedarnel wasconsumedfor thefirsttime)thefishingroup2receivedtubifex and anexcessiveamount ofdarnelandthethirdgroupwasfedanexcessiveamount ofdarnelonly. After
50days the darnel wasreplaced by grass pellets and the daily amounts of tubifex werereduced to 100ga day per 30animals. Eachgroup of 30animals waskept in a 2001 tank at
23°C±1° C,withlightfor 14haday. Standardlengthandweightwereregularly measured.
Forhistologicalexamination,theanimalsweresacrificed at 15,27,48 days,3and 7months,
each time 2-4h after feeding. The wholefish(up to 2 months old; the ventral body wall
removed) orsmallpiecesof tissue(fish 2months or older)werefixedin Flemmingsosmium
tetroxidefixative(Romeis, 1968). Thetissueswereembedded inparaffin waxandcut at 1-4
urn.
Azonaldifferentiation ofthegutwasmadehistologically. Todeterminethelengthofthe
subdivisions,thenumberofhistologicalsectionsofeachzonewascounted. Infishlargerthan
50mm,thelengthofthetotalalimentarytractwasmeasuredbeforefixation,andpartsofthe
gutwereexaminedhistologicallytolocatethetransitionofeachzone.
For electron microscopical examination, the tissues were prefixed in a mixture of 1%
osmium-tetroxideand2%glutaraldehydebuffered with0 1 M sodiumcacolylatebuffer pH7-2
for 15 min,and postfixed for45mininthesamemixturetowhichl%potassium bichromate
was added. Fixation was carried out at 0°C. The tissues were embedded in epoxy resin.
Ultrathin sectionswerecut on aLKBHuxleyor LKBIII Ultramicrotome and photographs
weremadeonaPhilipsE.M.300electronmicroscope.
TABLEI. Growth rate of grasscarp fed on animal or vegetable food
Standardlength(mm)
r
Age(days)
15
28
48
57
70
110
206
335
Group1
(animal food)
—
—
43±4-7(8)*
49±7(3)
61±5-6(2)
95±7(2)
113±9-4(10)
120±8(10)
Group2
(control animals)
16-3±1-4(6)*
25-7±2-8(9)
42-2±3-9(5)
—
55± 7(2)
88± 5(2)
—
110±13(10)
Group 3
(plant food)
—
31-9±3-2(8)*
45± 4(3)
56±5-4(4)
66-7±5-2(4)
76±8-l (10)
75± 6(10)
*Thenumber offishexamined isindicated between brackets.
III. GROWTH RATE
A difference ingrowth ratewas observed betweengrasscarpfed ontubifex (group 1)
and those fed on grass (group 3) (see Table I). Between the groups 1(animal food)
and 2 (animal+plant food) no significant differences were found. Tubifex-fed fish
grewabout0-8mm/day to an ageof 110days;thenthegrowthdiminished. A reduced
growth rate for plant fed animals was noticed from the start of the experiment. The
reduced growth of tubifex fed fish between 110 and 335 days may be partly due to
crowding.
The meanweight of specimensfrom the two groups is also different: at 30days this
was 370mg, at 7 months the mean weight of plant fed animals was 7-3±2-6 g, and
that oftubifex fed specimenswas24-7±7 g(for ten animalsper group).
DIGESTIVE TRACT OF GRASS CARP
169
IV. MORPHOLOGY AND REGIONAL DIFFERENTIATION OF THE GUT
In Cyprinids, the stomach is absent. The oesophagus opens directly into the
intestine a few mm anterior to the entrance of the ductus choledochus. Thefirst
limb or intestinal bulb (intestinal swelling) is wide. Its rostral part has complex
branching mucosalfolds; at theend oftheintestinal swelling,thesefolds areat right
angles to the long axis of the gut. From this area the diameter of the intestine
decreasestowardstheanus.
15Days
cp^
t
o
27Days
48Days
I-
5mm
FIG. 1. A schematic drawing of the digestive tract of grass carp at three stages of development.
Ventralview:cp,caudal pharynx;t,position ofpharyngeal teeth; o,oesophagus;ib,intestinal
bulb; ip, intestine proper; a, anus. A and B,boundaries of the second zone of the intestine.
Based onhistological features theintestinecanbedividedintothreezones(Fig.1).
Table II showsthe relative length of the total alimentary canal, the intestine and its
three zones in specimens of different ages and fed on different kinds of food. The
relativelengthofthegutincreasesfrom approximately 1xstandard bodylengthin15
daysto 2xbody length in 210days. This increase was mainly due to growth of the
firstzone. The relative length of the gutislargerin grass fed animals than in those
fed on tubifex. The difference can be attributed to growth of thefirstzone of the
intestine.
170
H. W. J. STROBAND
TABLEII. Ratioofstandard bodylengthtolengthofthegutanditssubdivisions
Intestine
A
Age(days)
Totalcanal
r
Zonel
Zone2
Zone3
0-4
0-4
0-6
0-6
0-3
0-3
0-3
0-3
005
01
0-2
0-2
10
11
1-4
1-4
Total
07
0-8
1-1
11
210(1)
210(1)
210(1)
210(1)
1-8
1-8
1-7
1-9
1-5
1-5
1-5
1-6
0-9
0-9
0-9
0-9
0-3
0-4
0-3
0-4
0-3
0-2
0-3
0-3
210(3)
210(3)
210(3)
210(3)
210(3)
2-4
2-2
20
1-9
20
20
1-9
1-7
1-6
1-7
1-3
1-2
10
0-9
0-9
0-4
0-4
0-3
0-3
15
27
48(1)*
48(3)
0-7
0-4
0-4
0-3
0-3
* (1)= group1fedonanimalfood.
(3)= group3fedonplantfood.
V. HISTOLOGY
HISTOLOGY OF THE INTESTINAL EPITHELIUM
The gut does not possess villiform projections, but the surface contains mucosal
foldswhichgraduallydecreaseinheightfromtheanteriortotheposteriorpartsofthe
intestine. The mucosa islinedwithacolumnar epithelium consistingoftwotypesof
cells:absorptive cellswithmicrovilliand,lessfrequently, glandular cellsofthegoblet
typewithP.A.S.positivecontents. The latter cellsare moreabundant inthe caudal
part of the gut. Other celltypes found between the epithelial cellsinclude:lymphocytes (Plate I), cells with rod-like inclusions [probably Rhabdospora thelohani
(Laguesse)], and a few acidophilic granular cells. At the base of the folds, mitotic
stagescanbeobserved (PlateI). Specialattention waspaidto the appearance ofthe
absorptive cellsthroughout the intestinal canal. These cells are high columnar with
nuclei situated in the lower third, the nuclei have clear nucleoli, the cytoplasm is
granularandhasa subapicalbasophilicband andabasalbasophilicarea.
Theintestinewasdivided intothree distinct zonesonthebasisoftheappearanceof
the absorptive cells. In the first zone (including the intestinal bulb and the rostral
part of the intestine proper), many cells in the apical region of the mucosal folds
showed a number of clear vacuoles between nucleus and the subapical basophilic
cytoplasm(PlateI). Thevacuolesafter Bouinsfixationappearedemptybutshowedan
osmiophilic substance afterfixationin Flemmingsfluid(Plate II). The nucleiin this
zonebecomelargerandmoreroundedtowardstheapicalregionofthemucosalfolds.
The cells contained small P.A.S. positive granules in the perinuclear cytoplasm.
Thesecondzoneofthegutischaracterized bythepresence oflarge' supra-nuclear
bodies' and vacuoleswith basophilic and P.A.S.positive contents (Plate III). Cells
withvacuoleswere observed alloverthe mucosal folds. Thevolume ofthevacuoles
PLATEI. Cross section through mucosal fold inthefirstzoneof intestine ofan animal of48days,fed
on tubifex and plant material. Note the presence of clear vacuoles in absorptive cells at the
apex of the fold. Mitotic figures can be seen at the base of the fold. A few goblet cells are
present(arrows). Bouinfixative,Azan stain.
DIGESTIVE TRACT OF GRASS CARP
171
increasedfrom rostraltocaudalinthesecondzone. Theapicalbasophilicareaofthe
cellsisdistinct in this zone (Plate III). Finally, a third zoneisfound near theanus.
Inthiszonetheabsorptivecellsdonotcontainvacuoles.
This description is based on control and tubifex fed animals, between which no
difference could bedetected. Theepithelium ofplantfedfishisnearlysimilarto that
of animal fed specimens, the absorptive cells of the first zone of the gut do not,
however,containvacuoles,andthecellsinallthreezonesareshorter.
ULTRASTRUCTURAL FEATURES OF ABSORPTIVE CELLS* OF CONTROL
ANIMALS
The absorptive cells in the first zone of the intestine are typically columnar and
contain well-developed microvilli (Plate IV). In the terminal web area beneath the
microvillous border, cellular organelles are absent and the cytoplasmic matrix
consists of a dense fibrillar meshwork. Between the terminal web and the nucleus
(which is situated at one third of cell height) the cytoplasm is packed with vesicles
varying in diameter. They are lessnumerous in the cytoplasm between nucleus and
basal membrane. The vesicles contain small particles with a diameter of approximately 70nm (Plate IV). Similar particles were found in the numerous intercellular
spaces. Mitochondria andpolyribosomes arepresent throughout thecell,the former
aremorenumerousinthebasalregion (PlatesIV,V). In somecellsstrandsofrough
endoplasmaticreticulum(RER)havebeendetected. Inthebasalpart ofthecellsand
occasionally in apical positions lamellar structures are present, generally running
paralleltothe longaxisofthecells(PlateV). Onlycellsattheapicesofthemucosal
folds contain largevacuoleswith electron lucent contents (PlateIV).
In the second zoneofthe gut the absorptive cellsshowinvaginations ofthe apical
cell membrane into the terminal web area (Plate VI). One or more large supranuclear vacuoles, mostly surrounded by mitochondria, contain an electron lucent
mass and numerous fibrillar structures (Plate VI). A distinct Golgi apparatus, a
number of mitochondria and scattered strands of RER are present between these
vacuoles and the nucleus. Many mitochondria, RER and lamellar structures are
presentinthebasalcytoplasm.
Thecellsofthethirdzoneoftheintestinearecharacterizedbytheirregulararrangement of the microvilli, which are short and few in number (Plate VII). The cells
contain many mitochondria, especially in apical and basal position. A perinuclear
Golgi apparatus is always present. Many vesicles and tubules of smooth endoplasmatic reticulum can be observed in the cytoplasm. Lamellar structures similar to
thosedescribedforfirstzonecellsarealsopresentnearthebasalmembrane.
VI. DISCUSSION
Theobservation that thegrasscarpjuveniles growfaster onanimalthan vegetable
food suggeststhefishesarenotherbivorousinthefirst7monthsafter birth. Thisisin
accordancewith Fischer (1972)whostressed theneedfor animalproteinsfor normal
growthofCtenopharyngodon idella. Thelengthof theintestinalcanalof 7month old
specimensdoesnotindicate aherbivorouswayoflife sinceotherpresumed herbivorousCyprinidshaveanintestineofsixtimesthebodylength(Campostomasp.Rogick,
*Details of ultrastructure are given as far as they underline the characteristics of the absorptive
cells in the three zones, and to compare our results with experimental ultrastructural data in the
literature. Adetailed description oftheintestinalultrastructure willbepublishedelsewhere.
172
H. W. J. STROBAND
1931)oreven15timesthebodylength(Labeohorie, Girgis,1952). Ourresultsdonot
allowconclusionsastothetimeintervalinwhichthechangefrom acarnivoroustoa
strictly herbivorouswayoflife takesplace. Olderjuveniles andevenadultgrasscarp
have the samerelative gut length as our 7months oldfish(Inaba &Nomura, 1956;
Berry &Low, 1970); consequently the suggestion of Inaba &Nomura (1956) that
grass carp gradually acquires omnivorous habits after the first month and actually
neverbecomesrealherbivorous seemsplausible. Hickling(1960)reported veryrapid
growthofgrasscarponNapier-grass. Howeverthefishwerekeptinlargepondsand
the stomach contents were not analysed, it cannot be excluded therefore that small
amounts of animal food were swallowed with thegrass.
McVay &Kaan (1940)werethefirstauthors to describe a regional differentiation
ofthegutofaCyprinid. Accordingtotheseauthors,theanterior half ofthe goldfish
Carassiusauratus,intestineischaracterizedbythepresenceofabsorptivecellswithout
a single perinuclear vacuole, which was invariably found in cells of the posterior
intestine. This issupported by our results in plant fed animals. In tubifex fed grass
carp,however, osmiophilic droplets werefound in cellssituated inthe apical part of
themucosalfoldsinthefirstzoneoftheintestine. Thissuggeststhat absorption offat
takesplaceinthesecells. Gauthier &Landis(1972)found absorbed lipidsappearing
firstinthe apicalcellsand later inmorelateralregionsoffolds inthe anterior gutof
thegoldfish. Ourultrastructural examination indicatesthepresenceoflipid droplets,
possiblychylomicrons,invesiclesofdifferent sizesand inintercellular spaces of only
thefirstzoneoftheintestineofthegrasscarp. Thelargevacuolesfound attheapexof
the mucosalfoldspossiblyrepresent stored lipids,sinceabsorption seemsmost active
inthisarea. Theabsenceofpinocytoticinvaginationsinanterior intestine absorptive
cellswasalso observed byYamamoto (1966)and Gauthier &Landis (1972)ingoldfish and byIwai(1969)injuvenilecarp. Therefore, fat isprobably hydrolysed inthe
gutlumenand absorbed andresynthesized incellsofthefirstzoneofthegut. Thisis
in accordance with Al Hussaini (1949è), Sarbahi (1951) and Hickling (1966), who
statedthat lypolyticactivity islocated mainly inthe anterior intestine of stomachless
teleosts. This mechanism of fat absorption closely resembles that of the intestinal
triglycerideabsorption intherat(Cardelletal., 1967).
In the grass carp, the large supranuclear vacuoles in the second zone of the gut
containaP.A.S.-positive substance. ThisisconsistentwiththeresultsofGauthier &
Landis (1972) and Noaillac-Depeyre & Gas (1973a), obtained in goldfish and carp
respectively. The latter authors proved that digestion by amylase did not affect this
affinity for P.A.S. Onthebasisofthisobservation and ofthestainingreaction ofthe
substanceinthevacuoleswithtoluidineblue,theyconcludedthesubstanceisaneutral
or slightly anionic glycoprotein. The same authors showed that proteins are absorbed asintact molecules in the epithelial cellsof the posterior intestine of goldfish
and carp; in these cells they found indications of pinocytotic activity. Yamamoto
(1966)concluded that pinocytosistakesplaceinabsorptive cellsoftheposterior half
of the gut in the goldfish. Our results indicate that protein absorption in the grass
carptakesplacebypinocytosisinthesecondzoneoftheintestine,asinothercyprinids.
Studiesoftheactivitiesofseveraldigestiveenzymesmadeitclearthatpepticdigestion
isabsentinthesestomachlessteleosts(Sarbahi, 1951;Ishida, 1936). Trypticactivity,
however, was found in the posterior half of the intestine in grass carp (Hickling,
1965),goldfish (Sarbahi, 1951),carp,roach Rutilus rutilus and gudgeon Gobio gobio
(Al Hussaini, 19496). The incomplete digestion of proteins by the absence of a
DIGESTIVE TRACT OF GRASS CARP
173
stomach isprobably compensated by pinocytosis. A regional differentiation as in the
gutofadultCyprinidswasobserved in someteleost larvae (Iwai, 1968a,b, 1969;Iwai&
Tanaka, 1968a, b).
Most authors do not distinguish different zones in what they call the posterior
intestine of fishes. In the grass carp a caudal third zone can be clearly distinguished
histologically from the second zone. The presence of relatively few short microvilli
suggeststhat absorption isnot asimportant asinother zones ofthegut. The presence
of many lamellar structures in the basal part of the absorptive cells is in accordance
with the results of Noaillac-Depeyre & Gas (1973è) for the ' rectum ' of the goldfish.
These authors suggested that this part of the gut is specialized in ion transport.
Yamamoto (1966) did not pay special attention to the most posterior part of the
intestine ofthe goldfish, but described lamellar structures inthe absorptive cellsof the
whole intestine. If these structures are related to ion transport, our results indicate
that in the grass carp the first and the third zone of the gut are best adapted to this
function.
Our results indicate the more rapid increase in relative length of the intestine of
plant fed grass carp compared to that in tubifex fed specimens is merely a relative
increase in length of the lipid absorbing zone of the gut. This zone may be also
responsible for carbohydrate absorption, as a P.A.S. positive granulation was found
in the absorptive cells, and amylase is found chiefly in the anterior intestine of a
number of Cyprinids (Al Hussaini, 1949Ô,Sarbahi, 1951 ;Hickling, 1966). The cause
of the enlargement of the gut and its physiological significance needs further
investigation.
The distribution of absorptive cells in the intestine and mucosal folds differs from
that in mammals. The distinct regional differentiation in the carp intestine is considered useful for investigating the differentiation of epithelial cells. The cyprinid is a
useful model for examining fat and protein absorption pathways, the kinetics of the
epithelial cellsin different zones ofthe gut, and the influence of different kinds of food
on the latter.
Theauthorwishestothank Professor Dr J.W.M.Osseand Dr LucyP. M. Timmermans
for theirstimulatinginterest,MrW.Valenfor preparingtheillustrationsand MrEleriforhis
technical assistanceinelectron microscopy.
References
Al-Hussaini,A. H. (1949a). Onthefunctional morphology of the alimentary tract of some
fish inrelationtodifferences intheirfeedinghabits. I:AnatomyandHistology. Q.J.
microsc. Sei.90,109-139.
Al Hussaini,A. H. (1949A). Onthefunctional morphology of the alimentary tract of some
fishin relation to differences in their feeding habits. II: Cytology and Physiology.
Q.J.microsc.Sei.90,323-354.
Anderson, E. N. Jr. (1970). Traditional aqua culture in Hong Kong. J. Trop.Geogr. 30,
11-16.
Berry,P.Y. &Low,M.P.(1970). Comparativestudiesonsomeaspectsofthemorphology
and histology of Ctenopharyngodon idellus,Aristichthys nobilisand their hybrid
(Cyprinidae). Copeia1970,4,708-726.
Cardell,R.R.Jr.,Badenhausen,S.&Porter,K.R.(1967). Intestinaltriglyceride absorption
intherat. / . CellBiol.34,123-155.
Cross,D.G.(1969). Aquaticweedcontrolusinggrasscarp. J.Fish. Biol. 1,27-30.
Curry,E.(1939). Thehistologyofthedigestivetubeofthecarp(Cyprinus carpio communis).
J.Morph.65,53-78.
174
H. W. J. STROBAND
Fischer, Z.(1968). Foodselection in grasscarp(Ctenopharyngodon idellaVal)underexperimentalconditions. PolskieArchwm.Hydrobiol.15,1-8.
Fischer, Z. (1972). The elements of energy balance in grasscarp {Ctenopharyngodon idella
Val). Part.Ill, Assimilability of proteins, carbohydrates and lipids byfishfed with
plantandanimalfood. PolskieArchwm.Hydrobiol.19,83-95.
Gauthier, G.F. &Landis,S.C.(1972). Therelationshipofultrastructural andcytochemical
features to absorptive activityinthegoldfish intestine. Anat.Rec.172,675-701.
Girgis, S.(1952). On the anatomy and histology of the alimentary tract of an herbivorous
bottom-feeding cyprinoidfish,Labeohorie(Cuvier). J. Morph. 90,317-362.
Hickling, C. F. (1960). Observations on the growthrate of the chinese grasscarp,Ctenopharyngodonidellus(Cuv.etVal). MalayAgric.J.43,49-53.
Hickling,C.F.(1966). Onthefeeding processinthewhiteAmur, Ctenopharyngodon idella.
J.Zool.140,408-419.
Inaba,D.&Nomura,M.(1956). Onthedigestivesystemandfeedinghabitsofyoungchinese
carpcollectedintheRiverTone. J.TokyoUniv.Fish.42,17-25.
Ishida,J.(1936). Distribution ofthedigestiveenzymesinthedigestivesystemofstomachless
fish. AnnotnesZool.Jap.15,263-284.
Iwai, T. (1968a). Fine structure and absorption patterns of intestinal epithelial cells in
rainbowtroutalevins. Z.Zellforsch.Mikrosk.Anat.91,366-379.
Iwai, T. (19686). The comparative study of the digestive tract of teleost larvae V. Fat
absorption in thegut epithelium of goldfish larvae. Jap.Soc.Sei.Fish. 34,973-978.
Iwai, T. (1969). Fine structure of gut epithelial cells of larval and juvenile carp during
absorption offat andprotein. Arch,histol.Jap.30,183-189.
Iwai, T. & Tanaka, M. (1968a). The comparative study of the digestive tract of teleost
larvae. III. Epithelialcellsintheposteriorgutofhalfbeaklarvae. Jap.Soc.Sei.Fish.
34,44-48.
Iwai,T.&Tanaka,M.(1968b). Thecomparativestudyofthedigestivetractofteleostlarvae.
IV. Absorption offat bythegut ofhalfbeaklarvae. Jap.Soc.Sei.Fish.34,871-875.
Klust, G. (1940). Über Entwicklung, Bau und Funktion desDarmes beim Karpfen. (CyprinuscarpioL.). Int.Revueges.Hydrobiol.Hydrogr.40,88-173.
Lin,S.Y. (1935)Life history of Waan Ue, Ctenopharyngodon idellus (Cuv. &Val.). Lignan
Sei.J. 14,129-135.
McVay,J.A.&Kaan,H.W.(1940). ThedigestivetractofCarassiusauratus.Biol.Bull.Mar.
biol. Lab. Woods Hole.78,53-67.
Noaillac-Depeyre,J. & Gas, N. (1973a). Absorption of protein macromolecules by the
enterocytesofthecarp. (CyprinuscarpioL.). Z. Zellforsch. Mikrosk. Anat.146, 525541.
Noaillac-Depeyre,J. &Gas,N. (1973b). Miseenévidenced'unezoneadaptée au transport
des ions dans l'intestine de carpe commune (Cyprinus carpio L.). C.r.Hebd. Séanc.
Acad. Sei.(Paris) 276,773-776.
Penzes,B. &Tolg,I. (1966). Etude de la croissance et de l'alimentation de la ' grasscarp '
(Ctenopharyngodonidella)en Hongrie. Bull.fr. Piscic39,70-76.
Rogick, M.D. (1931). Studieson thecomparative histology ofthedigestivetubeof certain
teleost fishes. II. Aminnow (Campostoma anomalum). J. Morph.52,1-25.
Romeis, B. (1968). Mikroskopische Technik. München: Oldenbourg Verlag. Wien. 16
Auflage.
Sarbahi, D. S. (1951). Studies of the digestive tracts and the digestive enzymes of the
goldfish, Carassius auratus (1.)and thelargemouth black bass,Micropterussalmoides
(Lacépède). Biol. Bull. mar. biol. lab. WoodsHole,100,244-257.
Stott, B. &Robson, T. O.(1970). Efficiency ofgrasscarp (Ctenopharyngodon idella Val.) in
controllingsubmergedwaterweeds. Nature,Lond. 226,870.
Yamamoto, T. (1966). An electron microscope study of the columnar epithelial cellin the
intestineoffresh waterteleosts:goldfish (Carassiusauratus)andrainbowtrout(Salmo
irideus).Z. Zellforsch. Mikrosk. Anat.72,66-87.
Printed in Great Britain by Henry Ling Ltd., at the Dorset Press, Dorchester, Dorset
Cell Tiss. Res. 187, 181-200 (1978)
CellandTissue
Research
© by Springer-Verlag 1978
The Ultrastructure and Renewal
of the Intestinal Epithelium of the Juvenile Grasscarp,
Ctenopharyngodon idella (Val.)
H.W.J. Stroband and F.M.H. Debets*
Agricultural University, Department of Experimental Animal Morphology and Cell Biology,
Zodiac, Wageningen, The Netherlands
Summary.The intestinal absorptive epithelium of starved and fed fish has been
studied electron microscopically.After feeding, cellsoftheproximal segment of
theintestineshowmorphological characteristics oflipidabsorption. Absorptive
cellsinthemiddle segment contain manypinocytoticvesiclesinboth fasted and
fed specimens.Absorption of protein macromolecules issupposed to be one of
themainfunctions ofthispartofthegut.Inthemostcaudalpartoftheintestine,
absorptive cells carry relatively few and short microvilli. The proximal and
distal segments show structural indications of a function in osmoregulation.
The renewal of the epithelium has been studied with light microscopic
autoradiography, using tritiated thymidine. The intestinal mucosal fold
epithelium represents a cell renewal system. The cells proliferate at the base of
the fold and migrate towards the apex in 10-15 days at 20°C. The functional
absorptive cells proved to be generally present in the intestinal epithelium,
including theproliferative area. Undifferentiated cellshavenot been identified.
The results will be compared with data on absorption of lipid and protein
macromolecules in teleostean and mammalian intestines and with descriptions
of the cell renewal system in the mammalian intestine.
Key words: Intestine - Teleost - Epithelium - Renewal - Ultrastructure.
Introduction
A differentiation of the intestinal epithelium was found in a number of Cyprinids
(whichdonothaveastomach)(Yamamoto, 1966and Gauthier andLandis, 1972in
the goldfish; Iwai, 1969inthecarp; Noaillac-Depeyre and Gas, 1976in the tench).
Therostralpartofthegut,includingtheintestinalbulb,wasshowntoabsorblipids,
Send offprint requests to: Dr. H.WJ. Stroband, Agricultural University, Department of Experimental
Animal Morphology and Cell Biology, Zodiac, Prinses Marijkeweg 40, Wageningen, Niederlande
* The authors wish to thank Prof. Dr. J.W.M. Osse and Dr. Lucy P.M. Timmermans for their
stimulating interest, Mr. H.G. Eleri(T.F.D.L.) for his technical assistance in electron microscopy, and
Mr. W.J.A. Valen for making the drawings and light microscopic photographs
0302-766X/78/0187/0181/$04.00
182
H.W.J. Stroband and F.M.H. Debets
and the posterior part of the intestine absorbs protein by pinocytosis. NoaillacDepeyre and Gas(1973b) demonstrated that ion transport takes place in the most
caudalpartofthegutofthegoldfish. Inarecentmorphological study,the grasscarp
intestine was shown to consist of three segments (Stroband, 1977).
In the goldfish (Vickers, 1962; Hyodo, 1965) and the carp (Gas and NoaillacDepeyre, 1974), the intestinal epithelium represents a cell renewal system. Cell
proliferation takesplaceinthebasal part ofthemucosal folds, and thecellsmigrate
to the top of the folds where they are probably discarded.
The present study deals mainly with the renewal of the intestinal epithelium of
thegrasscarp.Autoradiography wasappliedafter injecting 3 H-thymidine,to locate
the supposedly present proliferative pool of cells and to trace the renewal of the
epithelium. The ultrastructure ofabsorptivecellshasbeenstudied inthree areas of
themucosalfold: thebase,thecentralpartandtheapex.Thepurposewasto obtain
general information on the ultrastructure of absorptive cells and on the
morphology of possibly differentiating cells,migrating from the proliferative pool
totheapex ofthefolds. To identify functional absorptive cells,thedifferences have
been studied between these cells in fed fish and in starved specimens.
Materials and Methods
Autoradiographic studies were made on 21 one year old juvenile grasscarp (Ctenopharyngodon idella,
Val.) provided by O.V.B. Lelystad, Holland. These were injected intraperitoneally with methyl- 3 Hthymidine (1uCi/gram body-weight; spec, activity 18 Ci/mmol; Radiochemical Centre, Amersham,
England). Before and during the experiment the fish were fed with Tubificidae. The water temperature
was 20°C±1°C. Three animals were killed with ±0.3U/Oo MS222 (Sandoz, Basel, Switzerland) after
3h, 24h, 2days, 5, 10, 15and 21days.Tissues were fixed inBouin's fixative and embedded in paraffin.
Sections of 3um were stained with haemalum and eosin. Radioautograms were made with slides
preparedinKodak NT B2emulsion(Eastman-Kodak, Rochester,NewYork),diluted 1:1with distilled
water. The slides were exposed for three weeks.
Forelectronmicroscopicpurposes,7-9monthsoldgrasscarpwerestarvedfor 15days(2animals) or
40days(4animals).In addition, two specimens wereregularly fed. Three ofthe starvelings and the two
regularly fed fishes were given tubifex approximately 3h before being killed. All these animals were
anaesthetized in0.1°/ 00 MS222,and small fragments ofintestinal tissuewereexcised from the locations
showninFig. 1.For electron microscopicalexamination, tissueswereprefixed for 15mininamixtureof
1% osmium tetroxide and 2 % glutaraldehyde buffered with 0.05M Na-cacodylate (pH 7.2) at 0"C.
Postfixation was made in a similar mixture to which 1 %potassium bichromate was added. The tissues
weresectionedonaLKBIIIultramicrotome.Thesectionswerecontrasted withuranylacetate, followed
by lead citrate according to Reynolds (1963), and photographs were made on a Philips 300 electron
microscope.
Results
1. The Renewal of the Intestinal Epithelium
3h after injection of 3 H-thymidine, labelled epithelial cells can be noticed in the
three segments of the gut. Most of these cells are found in the central segment
(Table1), and they are concentrated in the basal third of the folds (Figs.2,4;
Table1).
Grasscarp Intestinal Epithelium
183
Table1.Percentageoflabelledcellsinagivenareaofthemucosalfoldsintwospecimens(IandII),3h
after 3H-thymidine injection
Area(in % of
total fold length)
0%= baseof
the fold
0-10
10-20
20-30
30-40
40-50
50-60
Percentage oflabelled cells
in agiven area measured in 5-8 folds/segment
proximal segment
middle segment
distal segment
I
II
I
II
I
II
3.3±0.5
1.1±0.4
1.3±0.7
6.5±1.0
2.9±1.6
8.2±1.8
4.6±1.1
2.5±0.8
2.0±1.0
0.9±0.4
0.7±0.4
10.2±2.6
2.3±1.7
3.3±0.9
3.7±1.1
0.7±0.5
1.7±1.3
3.9±1.1
1.5±0.4
0.6±0.3
-
-
-
-
-
Figure4shows that labelled cellsmigrate totheapex ofthemucosal folds.The
result after 10-15days are shown in Figure 3.Theheight inthefolds reached by
labelled cells 48hafter 3 H-thymidineinjection andthelarge standard deviation in
thiscolumn suggest aconsiderable individual variation intheproliferative activity
of the intestinal epithelium.
Three hours after injection labelling in the intestinal epithelium may be quite
different from place to place; this indicated that proliferation varies in the folds
with time. The well-labelled areas have been used for quantification.
2. The Ultrastructure of the Epithelium of theProximal Segment
a) In FedFish.Most ofthecellsofthe(columnar) epithelium consist of absorptive
cells.Goblet cellsarepresenttoo,andalso someendocrinecells(Fig.18;Rombout,
1977) and migrating cells (lymphocytes, granulocytes and macrophages).
The apical part ofthe absorptive cellbears numerous microvilli(Fig.5)with an
asymmetric cell membrane (Fig.8). The part of the cell adjacent to the lumen is
covered with a thin fuzzy coating or glycocalyx. The cytoplasm of the microvilli
containsanumber oflongitudinal fibrillar structures,penetratingtheterminalweb.
The tips ofthe microvilli have a dense cap(Fig.5),named capitulum by Krementz
and Chapman (1974).
The lateral plasma membranes of adjacent cells run closely together from the
microvillous border to thebasement membrane without complex interdigitations.
Desmosomes are frequently encountered, most of them lying in the terminal web
area whereajunctional complex ispresent(Fig.9).Inthebasal halfof thecells,the
lateral plasma membranes form long lamellar infoldings parallel tothelong axisof
the cells. These infoldings can be recognized by the regular distance between the
membranes( ± 250Â) andbythepresence oflumen material with a relatively high
electron density. Themembranes are relatively thick and asymmetrical, just asin
the microvilli (Fig.6). The same type of infoldings originates in the basal plasma
membrane(Fig.7).Similar structures weredescribed byNoaillac-Depeyre andGas
H.W.J. Stroband and F.M.H. Debets
184
anus
«^
.è #•'
... «,
. i
• *
j>
-I
*r-
<.
> $ ,
i551t ..
•v>; f
.•• i
... V _ _
À
' l :'• ;4A>.*V
•t v >- •'t1-."'*•• «
HA» • >/
yt.
t -*
%i;'~ '
\t,-'
• - . 'i
* % % . ' • - • •
2b
2a
,*
? " .'
iV-
*.t
'% **
••'•.•••
. **
^ t,
• •ƒ••
*•<;'?-w
t' %
<•
.
-'.#.
'
• ' ^
>. * *
< •/,;- ' , . « - .
».
-
•
v.
j,
••••*
%'..
. _
• -
#*
**..
..',
$':
>••"
• .
-..''*
1?
'•
\'
A-I *-
m
A l ;
t
••
X. !
Grasscarp Intestinal Epithelium
185
im
10096
"T"l
à
3h
rb
>
< 24h
<
5d
< lOd >
< I5d
Fig.4.Areasofthemucosal foldscontaininglabelledcells.Eachvaluerepresentsthemean of3animals;
5-8 folds have been measured per fish. Ordinate: % of fold height. Abscissa: time after injection
(1973b)incellsofthe"rectum"ofthecarp,anditwassuggestedthattheymayplay
a role in ion transport.
Between the microvilli, pinocytotic vesicles may be present (Fig.9), which do
not contain fat droplets, most membranes of which have a coated appearance.
A terminal webispresent below the microvillous border (Fig.5). In this area
cytoplasmicorganellesarescarce,andribosomesandsomevesiclesmaybepresent
also. Mitochondria, located in the basal parts of the cells(Fig.13),are generally
closely connected with the lamellar structures.
Thenucleusoftheabsorptivecellisovalandlocatedinthebasalpartofthecell
at about one third of the height.A few ribosomes may be attached to the outer
membrane of the nuclear envelope, which isgenerally smooth (Fig. 10).
Rough-(RER) and smooth-(SER) endoplasmic reticulum are overall present,
except in the terminal web area. In the upper part of the cells, the SER is
predominant. Many small fat particles, approximately 700Â in diameter, are
present in the SER, RER, Golgi apparatus (which is located in the perinuclear
cytoplasm) and intercellular spaces (Figs.9,10,11,12). Large droplets of fat are
present too(Fig.10),especially in thecytoplasm above thenucleus,and theseare
surrounded by a membrane.
b) InFastedFish.Themoststrikingchangeintheabsorptivecellsafter fastingisthe
absenceoffat particlesand fat dropletsinthecellsand intheintercellular spaces.
Fig.1. Schemeoftheposition ofthegrasscarpintestine.Theoesophagus(O)opensdirectlyinto the gut.
In the wide first limb, or intestinal bulb (IB), the ductus choledochus (DC) opens a few mm from the
oesophagus (O). From theend of the first limb theintestine proper (IP) narrows and leads to the anus.
Thearrowsindicatethelocationsofthetissuesusedinthisstudy.A andZJrepresenttheboundariesofthe
middle segment. GB gallbladder
Fig.2aandb. Radioautograph of the middle segment of the intestine 3h after injection with 3 Hthymidine.TheDNA precursor ispresentinnucleiincellsofthebasalpart ofthemucosalfolds, x 125.b
As Fig.2a. x250
Fig.3. Radioautograph of the middle segment 10days after injection. Labelled cells have reached the
apex of the mucosal folds, x 125
Figs.5-8.Absorptive cellsin the proximal segment
Grasscarp Intestinal Epithelium
Mitochondriawerenotonlyfoundinlargenumbersinthebasalpartofthecells,
asin fedfish,but they areconcentrated in theupper third of theabsorptivecells
also.Thissuggeststhat severalofthemitochondria havemigrated tomoreapical
parts of the cellsduring the fasting period. This type of apical concentration of
mitochondria isnot noticed three hours after feeding, following a long period of
fasting. The matrix of mitochondria of fasted specimens (Fig.5) contains dense
granules, which disappear after feeding.
c) CellsatDifferentLocationsoftheMucosalFolds.Allcellorganellesappeartobe
welldeveloped,eveninthemostbasalpartofthefolds.Adistinctindicationofcells
at thebaseofthefoldsbeinglessdifferentiated istheirnarrow shapecompared to
theshapeinotherareasofthefold(theaveragenumberofmicrovillipercellis15in
sectionsatthebaseofthefoldsand 30insectionsatthetopofthemucosalfoldsin
theproximal segment) and thegreat number offree polyribosomes(Fig.15).One
typeofcolumnar cellisrestricted to thebaseofthefolds. Thiscellcontainsmany
large mitochondria with electron dense granules intermingled with ER and free
ribosomes (Fig.17). This type may be found in groups of two or three cells.
Lymphocytes arepresentin thebasal part ofthemucosal folds, generally inlarge
numbers, most of which are located near the basement membrane.
After feeding, thecellsinthebasalpartofthefoldsaresimilartocellsatother
locations. The ER (Fig.16)and Golgi vesicles are filled with lipid particles, and
mostmitochondria arelocatedinthebasalpart ofthecells,butthisislessdistinct
than at other locations of the fold. After fasting neither lipid particles nor lipid
droplets have been observed.
After feeding thecellsat the topofthefolds aredifferent from thoseat other
locations. Next to structures mentioned above, several large lipid droplets(up to
4u) are present, most of them in the supranuclear cytoplasm (Fig.14). The
extracellular spaces are very large in this area and the different location of the
mitochondria in fed and fasted fish ismost conspicuous in theseparts.
Fig.5. Theapicalpartofsomecellsinthemiddleareaofamucosalfoldinafastedfish.Manyvesiclesof
SERwithoutlipidparticlesbelowtheterminalweb(TfV). Notetheabundantpresenceofmitochondria
(M)in thispart of thecells,containing densegranules x16,600
Fig.6.Thelateralplasmamembranes(LM)ofadjacent cellsinthenuclear(NU)area.Themembranes
form infoldings(I)containinganelectrondensematerial.Themembranesareasymmetricand thicker
than the lateral plasma membrane, x81,500
Fig.7.Basalportionofacellwithinfoldingsofthebasalplasmamembrane(arrows),closelyconnected
withmitochondria(M).Betweenthebasalcellmembraneandthebasementmembrane(BM)anarrow
spacecan be noticed, x41,000
Fig.8.Transverse section through microvilli. Many fibrils form the core of the microvilli. The unit
membrane isasymmetric. Themicrovilli are covered with a thin fuzzy coating, x104,000
1
187
« •«•»w n om. .£vi^y >A ^«*<fe^W^iASBrî
200
H.W.J. Stroband and F.M.H. Debets
Noaillac-Depeyre, J., Gas, N.: Absorption of protein macromolecules by the enterocytes of the carp
(Cyprinus carpio L.). Z. Zellforsch. Mikrosk. Anat. 146, 525-541 (1973a)
Noaillac-Depeyre,J.,Gas,N.:Miseenévidenced'une zoneadaptéeautransport desionsdans l'intestine
decarpcommune(Cyprinus carpioL.).C.R.Hebd. Séance.Acad. Sei.(Paris), 276, 773-776(1973b)
Noaillac-Depeyre, J., Gas, N.: Fat absorption by the enterocytes of the carp(Cyprinus carpioL.). Cell
Tiss. Res. 155, 353-365 (1974)
Noaillac-Depeyre,J., Gas,N.: Electronmicroscopicstudyongutepithelium ofthetench(TineatineaL.)
with respect to its absorptive functions. Tissue & Cell 8, 511-530 (1976)
Palay, S.L., Karlin, L.J.: An electron microscopic study of the intestinal villus II. The pathway of fat
absorption. J. biophys. biochem. Cytol. 5, 373-383 (1959)
Reynolds, E.S.: The use of lead at high pH as an electron-opaque stain in electron microscopy. J. Cell
Biol. 17, 208-212 (1963)
Rombout, J.H.W.M.: Enteroendocrine cells in the digestive tract of Barbus conchonius (Teleostei,
Cyprinidae). Cell. Tiss. Res. 185, 435^150 (1977)
Sarbahi,D.S.:Studiesofthedigestivetracts and thedigestiveenzymesof thegoldfish, Carassiusauratus
(L) and the largemouth black bass, Micropterus salmoides (Lacépède) Biol. Bull. mar. biol. lab.
Woods Hole, 100, 244-257 (1951)
Strauss, E.W.: Electron microscopic study of intestinal fat absorption in vitro from mixed micelles
containing linolic acid, monolein and bile salt. J. Lipid Res. 7, 307-323 (1966)
Stroband, H.W.J.: Growth and diet dependant structural adaptations of the digestive tract in juvenile
grasscarp (Ctenopharyngodon idella, Val.). J. Fish. Biol. 11, 167-174 (1977)
Toner, P.G.: Cytology of intestinal epithelial cells. Int. Rev. Cytol. 24, 233-243 (1968)
Trier,J.S.,Rubin, C E .: Electronmicroscopy ofthesmallintestine.Areview.Gastroenterology 49,574603 (1965)
Vickers, T.:A study of the intestinal epithelium of the goldfish Carassiusauratus: its normal structure,
thedynamics ofcellreplacement, and thechanges induced by saltsofcobalt and manganese. Quart.
J. Micr. Sei. 103,93-110(1962)
Yamamoto, T.:An electron microscope study of the columnar epithelial cell in the intestine of fresh
water teleosts:goldfish (Carassius auratus)and rainbow trout(Salmo irideus).Z. Zellforsch. 72,6687 (1966)
Accepted October 28, 1977
"»••' r'-fi-WJfm-'1,'-' "^f>F"ISJJ**"
ï
'
•
ï -< ,
»? ' IA- * \ > «.« »'S
*.~,i
i.*li4f
3 t *• •
(
.;>*
±LL
Hl stoche ml str y 64, 235-249 (1979)
Histochemistry
© by Springer-Verlag 1979
Regional Functional Differentiation in the Gut
of the Grasscarp, Ctenopharyngodonidella (Val.)
H.W.J. Stroband, H. v.d. Meer, and Lucy P.M. Timmermans
Agricultural University, Department Experimental Animal Morphology and Cell Biology,
Zodiac, Marijkeweg 40, Wageningen, The Netherlands
Summary. A regional differentiation - reflecting structural differences - of
the intestine of larval andjuvenile grasscarps can be illustrated by studying
the activity of alkaline phosphatase and the uptake of orally administered
horseradish peroxidase.
Pinocytosis of peroxidase takes place in a welldefined area of about
23% of the length of the gut (segment II). Neither the rostral ±68% (segment I)nor the caudal ±9% (segment III) shows absorption of the enzyme.
Alkaline phosphatase activity, mainly localized at the microvilli of the
enterocytes is high in the first segment of the gut and low in the second
segment. In larvae, the activity decreases sharply at the transition from
segment I to segment II. The activity is weak or absent in the caudal third
segment.Quantitativehistochemical data areconfirmed bybiochemical analyses.
Alkaline phosphatase activity is found all over the mucosal folds of
the first segment, with relatively weak activity at the base and at the tip
of the folds. This may be related to a renewal of the epithelium.
Our results suggest that active absorption of digested food takes place
mainly in the rostral first segment, while the uptake of macromolecules
by pinocytosis is a function of the second segment. Comparison of the
results with information available in literature leads to a rejection of the
hypothesis that the uptake of protein macromolecules in Cyprinids is to
be attributed to the absence of a stomach and therefore to an inefficient
digestion of proteins.
Introduction
An understanding ofabsorption processes and related morphological and physiologicalcharacteristics in larval,juvenile and adult stomachless teleosts isvaluable for fish culture. This is especially true because most teleosts pass through
a larval period (Balon, 1975)in which exogenous food is used when a stomach
0301-5564/79/0064/0235/$03.00
LOCALIZATION OF PROTEIN ABSORPTION DURING
TRANSPORT OF FOOD IN THE INTESTINE OF THE
GRASSCARP, CTENOPHARYNGODON IDELLA (VAL.)
H.W.J.StrobandandF.H.vanderVeen,
AgriculturalUniversity,DepartmentofExperimentalAnimalMorphologyand
CellBiology,Zodiac,Marijkeweg 40,Wageningen,The Netherlands
SUMMARY
Theuptake of proteinand aminoacids in the intestine of young (18months
old) and adult grasscarp has been studied with the inertmarker (Cr„0~)method.
Theabsorption ofprotein takesplace in theanterior 40-50% of thegut.
The second intestinal segment (+25%of gut length)has the ability to
absorbmacromolecules ofprotein.Ferritin (MW300.000)was found inpinocytotic
vesicles inenterocytes of this second segment.On thebasis ofmorphological and
histochemical evidence,ithas previously been suggested that the pinocytotic
uptake ofwholeproteinmoleculesmight be related to the lack of pepticdigestion instomachless fishes.Amajor quantitative function inprotein absorption
however cannotbe attributed to the second gut segment,and proteins appear tobe
digested adequately in these stomachless animals.The function of this partof
thegut and therole of the stomach inprotein digestion inother vertebrates
remain tobe explained.
The absorption of individual aminoacids in the intestine of the grasscarp
suggeststhatessential aminoacids arepreferentially absorbed. The rapid disappearance of lysine and arginine from the chyme points toa tryptic breakdown of
proteins.
I
REFERENCES
Austreng,E.(1978).Digestibilitydeterminationinfishusingchromicoxide
markingandanalysisofcontentsfromdifferentsegmentsofthegastro
intestinaltract.AquacultureJ3^265-272.
Barrington,E.J.W. (1957).Thealimentarycanalanddigestion.In:"ThePhysiologyofFishes"(M.E.Brown,ed.),VolI109-161.AcademicPress,NewYork,
Bell,G.H.,Davidson,J.N.&Emslie4smith,D. (1972)."Textbookofphysiology
andbiochemistry"ChurchillLivingstone,Edinburgh,London.
Ben-Ghedalia,D.,Tagari,H.,Bondi,H.&Tadmor,A. (1974).Proteindigestion
intheintestineofsheep.Br.J.Nutr.3J_>125-142.
Bergot,P. (1976).Demonstrationwithrutheniumredofdeepinvaginationsinthe
apicalplasmamembraneofenterocytesinrainbowtroutintestine.
Ann.Biol.Anim.Biochem.Biophys.16,37-43.
Cowey,C.B.&Sargent,J.R. (1979).Nutrition.In:"FishPhysiology",HoarW.S.,
Randall,D.J.&Brett,J.R. (ed|s).Vol.VIII,1-69.Acad.Press,NewYork,
London.
Crampton,R.F. (1972).Nutritionalandmetabolicaspectsofpeptidetransport.
In:"Peptidetransportinbacteriaandmammaliangut",CibaFoundation
symposium 11nov.1971.Ass.Sei.publ.Amsterdam.
Creach,P.V. (1963).Lesenzymesprotéolytiquesdespoissons.Ann.Nutr.Alim.
J2.375-471.
Davenport,H.W. (1977)."Physiologyofthedigestivetract".Yearbookmedical
publishers,Inc.,Chicago.
Dijke,J.M.van&Sutton,D.L. (1977).Digestionofduckweed (Lemmaspp)bythe
grasscarp (Ctenopharyngodon idella) J.Fish.Biol.JJ_,273-278.
Farmanfarmaian,A.,Ross,A.&Mazal,D. (1972).Invivointestinalabsorption
ofsugarinthetoadfish (Marineteleost, Opsanus tau). Biol.bull.142,
427-455.
Furakawa,A.&Tsukahara,H.(1966).Ontheaciddigestionmethodforthedeterminationofchromicoxideasanindexsubstanceinthestudyofdigestibilityoffishfeed.Bull.Jap.Soc.Sei.fish.32,502-506.
Gauthier,G.F.&Landis,S.C. (1972).Therelationshipofultrastructuraland
cytochemicalfeaturestoabsorptiveactivityinthegoldfishintestine.
Anat.Rec.172,675-701.
Hickling,C F . (1966).OnthefeedingprocessinthewhiteAmur, Ctenopharyngodon idella. J.Zool.140,408-419.
Hsu,Y.L.&Wu,J.L. (1979).Therelationshipbetweenfeedinghabitsanddigestiveproteasesofsomefreshwaterfishes.Bull.Inst.Zool.Acad.Sin.18,
45-53.
Iwai,T.(1968).Thecomparativestudyofthedigestivetractofteleostlarvae
Vfatabsorptioninthegutepitheliumofgoldfishlarvae.Bull.Jap.Soc.
Sei.Fish34,973-978.
Iwai,T.(1969).Finestructureofgutepithelialcellsoflarvalandjuvenile
carpduringabsorptionoffatandprotein.Arch,histol.jap.30_,183-189.
Jacobshagen,E.(1937).DarmsystemIVMittelundEnddarm.Rumpfdarm.In:
"HandbuchderVergleichendenAnatomiederWirbeltiere"(L.Bolk,E.Göppert
E.KalliusandW.Lubosch,eds).Vol.Illpp.563-724.
UrbanandSchwärzenberg,BerlinundWien.
Jany,K.D.(1976).Studiesonthedigestiveenzymesofthestomachlessbonefish
Carassius auratus Gibelio (Bloch):Endoptidases.Comp.Biochem.Physiol.
53B,31-38.
Kawai,S.&Ikeda,S.(1973).Studiesondigestiveenzymesoffishes-IVdevelopmentofthedigestiveenzymesofcarpandblackseabreamafterhatching.
Bull.Jap.Soc.Sei.Fish.39,877-881.
14
Krause,W.J.,Cutts,J.H.&Leeson,C R . (1977).Thepostnataldevelopmentof
thealimentarycanalintheopossum:IIIsmallintestineandcolon.
J.Anat.123,21-46.
Krementz,A.B.&Chapman,G.B. (1974).Ultrastructureoftheposteriorhalfof
theintestineofthechannalcatfish, Iotalurus
punotatus.
J.Morph._U5,441-482.
tfepham,T.B.&Smith,M.W. (1966).Aminoacid transportinthegoldfishintestine.
J.Physiol.184,673-684.
Noaillac-Depeyre,J.&Gas,N.(1973)• Absorptionofproteinmacromoleculesby
theenterocytesofthecarp {Cyprinus aarpio L.).Z.Zellforsch.146,
525-541.
Noaillac-Depeyre,J.&Gas,N.(1974).Fatabsorptionbytheenterocytesofthe
carp {Cyprinus aarpio L.)CellTiss.Res.155,353-365.
foaillac-Depeyre,J.&Gas,N. (1976).Electronmicroscopicstudyongutepitheliumofthetench {Tinaa tinea L)withrespecttoitsabsorptivefunctions.
Tissue&Cell8,511-530.
Joaillac-Depeyre,J.&Gas,N. (1979).Structureandfunctionoftheintestinal
epithelialcellsintheperch {Peraa fluviatilis
L).Anat.Rec.195,
621-640.
)nishi,T.,Murayama,S.&Takeuchi,M. (1976).Changesindigestiveenzyme
levelsincarpafterfeeding:IIIresponseofproteaseandamylasetotwice
adayfeeding.Bull.Jap.Soc.Sei.Fish42,921-929.
)rten,A.U.(1963).Intestinalphaseofaminoacidnutrition.Fed.Proc.22,
1103-1109.
ihcherbina,M.A. (1973).Astudyofdigestiveprocessesinthecarp {Cyprinus
aarpio) CommunicationIabsorptionofcrudefatintheintestinefrom
syntheticdiets.Vopr.Ikhtiol.J_3,119-127.
ihcherbina,M.&Sorvatchev,K. (1969).Afewdataconcerningtheabsorptionof
aminoacidsinthedigestivetractoftwoyearsoldcommoncarp.
Vopr.Prud.Rubov. \6, 315-322.
Ihcherbina,M.A.etal. (1976).Trofimova,L.N.&Kazlauskene,O.P.Theactivity
ofproteaseandtheresorptionintensityofproteinwith theintroduction
ofdifferentquantitiesoffatintothefoodofthecarp Cyprinus aarpio
J.Ichthyol.J6^632-636.
ihcherbina,M.A.,Shcherbina,T.V.&Kazlauskene,O.P. (1977).Amylaseactivity
andrateofcarbohydrateresorptionwiththeintroductionofvariousamounts
offatintothedietofthecarp, Cyprinus aarpio. J.Ichthyol.17,
327-331.
mith,M.W. (1966).Sodium-glucoseinteractioninthegoldfishintestine
J.Physiol.182,559-573.
taley,T.E.,Corley,L.D.,Bush,L.J.&Jones,E.W. (1972).Theultrastructure
ofneonatalcalfintestineandabsorptionofheterologousprotein.
Anat.Rec.172,559-579.
troband,H.W.J. (1977).Growthanddietdependantstructuraladaptationsof
thedigestivetractinjuvenilegrasscarp {Ctenopharyngodon idella Val.)
J.Fish.Biol. H.»167-174.
troband,H.W.J.&Debets,F.M.H. (1978).Theultrastructureandrenewalofthe
intestinalepitheliumofthejuvenilegrasscarp Ctenopharyngodon
idella
(Val.).Cell.tiss.Res.187,181-200.
troband,H.W.J.,Meer,H.v.d.&Timmermans,L.P.M.(1979).Regionalfunction
differentationinthegutofthegrasscarp Ctenopharyngodon idella (Val.).
Histochemistry64,235-249.
troband,H.W.J.&Dabrowski,K.R.Morphologicalandphysiologicalaspectsofthe
digestivesystemandfeedinginfreshwaterfishlarvae.In:"LaNutrition
desPoissons"CNERNA(Paris),inpress.
15
Stroband,H.W.J.&AnnetG.Kroon.Thedevelopmentofthestomachin Clarias
lazera andtheintestinalabsorptionofproteinmacromolecules.
SubmittedtoCell.tiss.Res.June1980.
Tanaka,M. (1969).Studiesonthestructureandfunctionofthedigestivesyste
inteleostlarvaeII.Characteristicsofthedigestivesysteminlarvaeat
thestageoffirstfeeding.Jpn.J.Ichthyol.J_6,141-149.
Tanaka,M. (1970).Studiesonthestructureandfunctionofthedigestivesyste
inteleostlarvaeIII.Developmentofthedigestivesystemduringpostlarv
stage.Jap.J.ofIchthyologyJ_8,164-174.
Walter,W.A.&Isselbacker,K.J. (1974).Uptakeandtransportofmacromolecules
bytheintestine.Possibleroleinclinicaldisorders.GastroEnterol.67,
531-550.
Yamamoto,T. (1966).Anelectronmicroscopestudyofthecolumnarepithelialce
intheintestineoffreshwaterteleosts:goldfish (Cavassius auratus) and
rainbowtrout (Salmo irideus). Z.Zeilforsch.72,66-78.
16
THE DEVELOPMENT OF THE STOMACH IN CLARIAS LAZERA
AND THE INTESTINAL ABSORPTION OF PROTEIN
MACROMOLECULES
H.W.J.Stroband andAnnetG. Kroon
Agriculture University, Department of Experimental Animal Morphology and Cell
Biology,Wageningen, The Netherlands.
SUMMARY
The development of the stomach of the teleost Clarias
lazera is described
for theearly posthatching period.A comparison ismade to the stomach of adult
Clarices.
The stomach develops in two distinct parts:the corpus,which is theearliestdifferentiated part,and the pylorus.The corpus contains amucous surface
epithelium, arranged infolds,and atubular gland system containing only one
type of gland cell, towhich thesecretion of pepsinogen and HCl is attributed.
Thepyloric region does not contain tubular glands.
From theultrastructure of thegland cells,the H thymidine labeling index,
and theonset of acid production (asdetermined with pH indicators) it isconcluded that afunctional stomach ispresent in juvenileswith astandard length
of +_ 11mm. (approximately 12days after fertilization at 23-24C).
Inaddition to the stomach, theultrastructure of the intestinal epithelium
hasbeen studied. The gut shows three segments,similar asdescribed for stomachless teleosts and anumber of fish larvae. In larvae aswell as injuveniles,the
enterocytes of the second segment showpinocytosis ofhorse radish peroxidase,
although in thejuveniles thestomachhas already developed. This second segment
has the same relative length inall studied larvae and juveniles and isalso present inadult
Clarias•
Therefore itmust be concluded that the ability of absorbing proteinmacromolecules isnot definitely related to the absence of a functional stomach in this
teleost species.
Key words: Stomach - Intestine -Epithelium -Teleost -larvae.
INTRODUCTION
Instomachless teleosts the intestine shows aregional differentiation. The
anterior segment has themorphological characteristics of lipid absorption, while
the shorter second segment has the ability toabsorb proteinmacromoleculesby
pinocytosis (Yamamoto,1966;Iwai,1969;Gauthier&Landis,1972;Noaillac-Depeyre&Gas,1973a,1976;Stroband,1977;Stroband&Debets,1978;Strobandet
al.,1979).Thethirdsegmentisprobablyinvolvedinionandwatertransport
(Noaillac-Depeyre&Gas,1973b;Stroband&Debets,1978).
Itwaspostulatedbytheabovementionedauthorsthattheabsorptionof
macromoleculesmightberelatedtothelackofastomachandtoaninadequate
proteindigestion.Thisisalsoknowninsucklingmammals,inwhichpinocytosis
ofproteinmacromoleculestakesplaceinthesmallintestinebeforethestomach
isfullyfunctional(Cornell&Padykula,1965;Kraehenbühl&Campiche,1969;
Krauseetal.,1977).Inourviewtherearetwowaysfortestingthementioned
hypothesis.Ifnoappreciablequantitiesofproteinareabsorbedbythesecond
gutsegmentinstomachlessfish,thehypothesiswouldbewithoutanyfoundation.
Experimentsareinprogressandthefirstresultswillbepublishedelsewhere
(Stroband&VanderVeen,inpreparation).Thesecondwayistostudythepost
hatchingdevelopmentofthedigestivetractofateleostwithastomach.Usuall)
teleostlarvaelackafunctionalstomachuntilseveralweeksafterthestartof
exogenousfeeding (Tanaka,1972).Duringthisperiodtheintestineshowssimilai
morphologicalcharacteristicsasfoundinthestomachlessfishes.Consequently,
thesecondsegmentoftheintestineofteleostlarvaeislikelytoabsorbprote:
macromoleculesalso.
Theprincipalobjectiveofthepresentstudyistoascertainwhetherchange
occurintheformandfunctionoftheintestine,especiallyinthesecondgut
segment,whenthestomachbecomesfunctional.Therefore,thedevelopmentofthe
digestivetracthasbeenstudiedwithlight-andelectronmicroscopicalmethods.
3
Theperiodofdevelopmentofthestomachwasalsodeterminedwith Hthymidineasamarkerofcellproliferation.Moreoveritwasexpectedthatthelabelingindexdecreaseswhencellsbecomefunctionallyactive.
pHIndicatorshavebeenusedtofindoutwhenacidityreachespH5-4,as
cathepsinaswellaspepsinhasanoptimumpHbelow5.Theuptakeofhorseradi:
peroxidaseservedasamarkerformacromoleculeabsorption.
MATERIALANDMETHODS
Twobatchesofnew-hatched Clarias
lazera (Cuv.andVal.)larvaeand6adu:
specimenwerekindlyprovidedbytheDepartmentofFishCultureandInland
FisheriesoftheAgriculturalUniversity.Althoughthelarvaewererearedin
exactlythesameway,thetwobatchesshowedsomewhatdifferentgrowthrates
(fig. 1).
g.1.Growthratesofthetwobatchesof Clavias larvae,(a)and(b).
E
z 10.
Thelarvaewererearedat23-24C.At5,6,8,9,12and15daysafterfer.lizationanumberoffishesofbath (a)wereinjectedwith+30-40nl.ofa
Ithymidinesolution (1Ci/L,spec.act.24Ci/mmol,RadiochemicalCentre,
ïersham,England).After3-4hincubationsomespecimenswerefixedinBouin's
jcativeandsubsequentlyembeddedinparaffin.4pSectionswerestainedwith
emalumandeosinorwithAlcianBlueandSchiffsreagens (Rcmeis,1968).
dioautographsweremadewithKodakNTB-2liquidemulsion(Eastman-Kodak,
ehester,NewYork),diluted1 : 1 . Theslideswereexposedfortwoweeks.For
eexperimentsdescribedbelow,thesecondbatch (b)ofClariaslarvaewasused,
ecimensof4,7,9,10,11,12and15daysoldwerepreparedforelectron
croscopy:theywerefixedinamixturecontaining 2%osmiumtetroxide, 2%gluraldehydeand M potassiumbichromate,bufferedwith0,05MNa-cacodylate
H7.2)at0C.1 \iSectionswerepreparedforradioautography.UltrathinseconsweremadeonaReichertUltratomeIVmicrotome,contrastedwithuranyl
etateandleadcitrateaccordingtoReynolds (1963),andphotographedwitha
ilips300electronmicroscope.
Furthermore,specimensof4to21dayswerefedwith Artemia satina ina
horseradishperoxidasesolution(Sigma,St.Louis;typeII,mol.Wt.+
.000).Theyswallowedtheenzymetogetherwiththefood.After2hthefishes
refixedfor2hin3%glutaraldehydebufferedwith0,05MNa-cacodylate(pH
2),rinsedwithwaterfor3h,frozeninliquidnitrogenandfreezedriedfor
h.Afterembeddinginparaffin,5ysectionswerepreparedandincubatedat
Dmtemperaturefor10-30minutesinamediumaftervanDuyn (Pearse,1972).
1 2
3 4 5
Fig. 2.a. schematic drawings of thedigestive tract of Clarias (batch a)at t
stages ofdevelopment.Dorsalview: o= oesophagus; s= stomach; i= intestin«
a= anus.A and Brepresent theboundaries of the second gut segment
=wall of thecorpus of the stomach.
= wall of thepyloric part of the stomach.
b. ordinate:Mean percentages of H thymidine labeled epithelial cells
three fishes at the same 6stages of development as fig.2a. abscissa: 1,2,
4 and 5represent the caudal oesophagus,themost rostral,mid and caudal
(pyloric)part of the stomach,and the intestine respectively. For thestomacl
2 slideshavebeen counted per location ineach fish.In the intestine thepel
centage of labelled cellswas counted at 7locations over theentire length.
Duringtheexperiments,everydaysomespecimensofbath (b)wereanaestheLzedwith0,1 /ooMS222immediatelyafterfeeding.Asmalldoseofmethyl-red
rcongo-redwasinjectedintothestomachthroughaglasscapillary.
Partsofthedigestivetractsofadult Clarias
lazera werefixedandpre-
iredforlightandelectronmicroscopyasdescribedabove.
5SULTS
, The morphology and regional differentation
of the digestive
tract.
Fig.2ashowsthemorphologyofthedigestivetractasreconstructedfrom
Lghtmicroscopicpreparations.Beforeday5,an"Anlage"ofthefuturestomach
)uldnotbedetected.Theintestineshowedthethreesegmentsasdescribedfor
:omachlessfish.Theesophagusisdirectlyfollowedbythefirstsegment,and
iebileductentersthisfirstsegmentveryclosetotheesophagus.Asfromday
,themostcaudalpartofthe"esophagus"graduallydevelopsintothestomach,
liehiseasilyrecognizedhistologicallybytheglandulartissueinthethickened
.11(figs.3,4,5).Betweenday9andday15thepyloricpartofthestomachis
veloped.
Afterthisstagenoimportantchangesseemtotakeplaceinthemorphology
thedigestivetract,apartfromaproportionalgrowthofthevariousparts,
erelativelengthofthegut(ascomparedtobodylength)andtherelative
ngthsoftheintestinalsegmentsremainthesame(table 1). Inadultsthethird
gmentisrelativelyshort,butthesecondsegmenthasthesamelengthasin
rvae.
Table1.Ratioofthelengthoftheintestinetothestandardbodylength,andthe
relativelengthofthethreegutsegmentsasdetermined for3specimensofeach
age(batcha).
age standardlength
(days)
(mm)
lengthintestine
bodylength
relativelengthofgutsegments (%)
I
II
III
5
8,0*
0,36
68,3+1,6 22,0+1,6
6
8,4 + 0,5
0,44
73,0+4,0 16,7+_3,8 10,3+1,5
9,8+2,4
8
10,4*
0,34
67,7+3,5 20,0+2,6 12,3+1,5
9
11,9 + 0,5
0,29
69,7+2,1 22,0+1,0
12
14,3 + 0,6
0,35
64,3+5,5 20,7+4,0 12,0+1,0
8,3+1,5
15
15,2 + 0,8
0,37
66,0+1,0 23,7+0,6 10,1+1,5
adult
257,8 + 15,7
0,45
70,8+3,1 23,9+2,4
5,2+0,5
* :extrapolationsfromfig.1.
2. The differentiation
of the digestive
3
H-thymidine labeling index.
tract epithelium as determined by its
3
Thepercentageof H-thymidine labeledcellsinseveralpartsofthedigestivetractandatdifferentstagesofdevelopmentarepresentedinfig.2b.In
larvaeof5days,21to231-oftheepithelialcellswerelabeledovertheentire
lengthoftheintestine,butlargevariationswerenoticedineachspecimen.
Labeledcellswerefoundallovertheepithelium,butmostarepresentinthe
lowerpartsofthemucosalfolds (fig.6).Duringdevelopmentthepercentageof
labeledcellsinthegutgraduallydimishedto+ 31onday15,whenlabeledcells
3
Fig.3-6:Incorporationof Hthymidineinthegutof Clarias
lazera.
Fig.3.Larva,4daysold.Lightmicroscopicradioautographofa1yeponsection
stainedwithtoluidineblue.Notethehighlabelingindexofthetissues.
S=surfaceepithelium;Gl=undifferentiatedglandularcells,(x500)
Fig.4.1ueponsectionthroughthecorpus (C)pylorus (P)androstralintestine
(Int)ofa7daysold Clarias. Notetherelativelyhighlabelingindexinthe
pyloricareaandtheverylowpercentagesoflabeledcellsinthegut (x250)
Fig.5.Developingstomachofa12daysoldlarva.Afewlabeledcellsinthe
anteriorcorpusofthestomach (C)andahighlabelingindexintheposterior
pyloricarea(P). x250
Fig.6.Mucosalfoldofthesecondintestinalsegmentofa4daysoldlarva.
Manysupranuclearvacuolesintheenterocytesandmostlabeledcellsatthebase
ofthefolds,x500
irerestrictedtothebasalpartofthemucosal folds.Intheareaofthedevelo)ingstomach,thelabeling indexdiminishes later,firstintheanteriorpartand
atermorecaudally (figs.2-5).Asfromday9thepyloricareacanberecognized.
tslabelingindexof+30°sdecreasesbetweenday12andday15tosimilarvalues
.sfoundinotherareasofthedigestive tract.Asintheintestinal epithelium,
iroliferativesurfacemucus cellsincorpusandpylorusaremerelyfoundinthe
asalpartofthefoldsafterthedropofthelabeling index.Glandularcells
emain labeledatrandominallstages.
Fortheproliferationoftheepitheliumofthesuccessivepartsofthediestive tract,thefollowingisofinterest.Acomparisonofradioautographsof
psectionswithultrathin sectionsdidnotshowanydistinctdifferencesbetween
3
hecytologyof H-thymidinelabeledcellsandothercells,neitherinthesurface
pithelium (fig.21)andglandularepitheliumofthestomachnorintheepithelium
ftheintestineofthestudied fishes (upto10days old). FunctionalcellsappaentlyhavethefacilitytosynthesizeDNA;consequently theyareabletodivide.
. Ultrastructure
and regional differentiation
of the
intestine.
Inlarvaeof4daystheesophagus,mainly linedwithmucous cells,enters
irectlyintothefirstsegment.Thisfirstsegmentisinvolvedinlipidabsorpion.Absorptive enterocytes,recognizedbytheapicalborderofmicrovilli,conainchylomicrons intheirendoplasmaticreticulum (ER)andGolgi apparatus,
irgelipiddropletsarealsopresent (fig.7).Chylomicronsarealsofoundin
ieintercellular spacesbetweentheenterocytes.Pinocytotic vesiclesarescarce
rabsent.MitochondriaandstrandsofERarepresent throughoutthecytoplasm
Lgs. 7-9: Electronmicrographs of gut absorptive cells in Clarias
lazera.
Lg.7.Apical part of an absorptive cell in the caudal part of the first
itestinal segment of a4days old larva.MV =microvilli;TW= terminalweb;
L=mitochondria; L = lipid droplet;Ch = chylomicrons;G =Golgi apparatus
Lthchylomicrons,x 17.000
Lg. 8. Some absorptive cells in the second intestinal segment.Many pinocytotic
:sicles (arrows)are present beneath themicrovillous border (MV),Supranuclear
icuoles (SV),probably originating from fusing pinocytotic vesicles and lyso>mes.4Days old larva,x 26.000
.g.9.Enterocytes in the third segment of thegut of a7days old larva.Many
imellar infoldings of theplasmamembrane in stacks (arrows)and afew small
.crovilli (MV).x 13.000
10
exceptinthatoftheterminalweb.Lamellarinfoldingsofthebasalandlateral
plasmamembranesoccur,justasinenterocytesofotherteleosts.
Absorptiveenterocytesofthesecondsegmentshowheavypinocytosisbeneath
theirmicrovillousborder (fig.8).Moreovermanypinocytoticvesiclesandone01
morelargesupranuclearvacuolesmaybepresent (figs.6, 8).Chylomicronshave
notbeenfoundintheER,intheGolgiapparatus,andintheintercellularspaces
butsomefatdropletsmaybepresentinthecytoplasm.
Themostcaudalsegmentdoesnothavethecharacteristicsofthefirstand
secondsegments;itlackslipiddroplets,chylomicrons,pinocytoticvesicles,and
supranuclearvacuolesintheenterocytes.Themicrovilliarerelativelyshort
andfewinnumber.Asinotherteleosts,foodabsorptiondoesnotseemtobean
importantfunctionofthesecells.Lamellarinfoldings,however,arefoundin
largenumbers,generallyconcentratedinstacks (fig. 9).
Duringdevelopmentnoimportantchangescanbenoticedinthemorphologyof
theabsorptiveenterocytes.Itisnoteworthythatthesecondsegmentremainsof
thesamelengthinallstages.Inadult Clarias thissegmenthasthesamecharacteristicsasinlarvae (table 1).
4. Ultrastrueture
of the developing stomach.
Thefirst"Anlage"ofthestomachcanberecognizedonday4withtheelectronmicroscopebythepresenceofafewlobulesoffutureglandularcells,forme
byinvaginationsofthesurfaceepitheliumofwhatseemstobethemostcaudal
partoftheesophagus.
Epithelialcells (fig.10)areelongate,andthenucleus (withlargenucleolus)issituatedinthebasalhalfofthecells.Neighbouringcellshavecomplex
interdigitationsofthelateralcellmembranes.AdistinctGolgiapparatusis
presentandmanystrandsofRERarefoundthroughoutthecells.Mucoidgranules
maybefoundinthecytoplasm,butareaccumulatedintheapexofthecell.The
roundtoovoidgranulesaremembranebound (fig.10).TheyprovedtobeP.A.S.
positiveafterAlcian-blue-P.A.S.reactiononparaffinmaterial;thereforethese
granulesprobablycontainneutralmucopolysaccharides.Thecytologyoftheepitheliumliningofthelumendoesnotchangemuchinlaterstagesofdevelopment,but
Fig. 10.Epithelial cells from thestomach of an 11days old larva.
NU =nucleus;Int.= complex interdigitations of the lateral plasmamembranes;
G =Golgi apparatus;M =Mucoid granules;Mi =mitochondria; RER= rough endoplasmatic reticulum,x 28.000
11
moregranulesandstrandsofendoplasmicreticulumandfewerfreeribosomesare
foundinolderspecimens.Afewveryshortmicrovillimaybepresentonthesurfaceoftheepitheliumlining.Inadultspecimensthesecretiongranulesare
flatterandmorenumerous (fig.19);theyaremoreelectrondensethaninlarvae.
Thetransitionfromsurfaceepitheliallayertotubularglandepitheliumis
veryabrupt,withoutatransitionor"neck"zone.
Theglandularcellscanbeeasilydistinguishedfromtheepitheliumlining
cellsbythelackofmucousgranulesandthepresenceofanumberofsmooth
vesiclesintheapicalarea(figs.11-16).Thecellsarepyramidalandshowcomplexinterdigitationsofthelateralcellmembranes.Thenucleicontainprominent
nucleoli.Manymitochondriaarepresentinthecytoplasm.AwelldevelopedGolgi
apparatusisfoundnearthenucleus.At4daysonlyafewstrandsofRERhave
beennoticed.Atthatagethecytoplasmisfilledwithfreeribosomes (fig. 11).
Apartfromtheliningepitheliumonlyonetypeofstomachglandularcell
wasfoundinthevariousstages.Theapicaltubularsystem,inearlystagesconsistingofanumberofvesicles,developsrapidly (fig.13-14)andfillsupthe
greaterpartofthecellsinspecimensof11days.Sincethemembranesofthe
vesiclesstronglyresembletheapicalplasmamembrane (fig.14),thesemembranes
mayhavethesameorigin.Importantchangeshavebeennoticedinthemorphology
ofthetubularsystemduringlaterontogeny.Inlarvaefrom11daysoldthe
vesiclesweremoreelongated,possiblybecauseofmutualfusion.Inadultspecimensthemoreelongatetubulesaremerelyextendinginlongitudinaldirection
(fig. 16).
Asfromtheageof7dayssmallsecretiongranulesappearinthecytoplasm
oftheglandularcells (fig.12).Inthefollowingdayslargezymogengranules
areformed (figs.4,13)andonday11thefirstindicationsofexocytosiscan
beobserved (fig. 15).
InadultspecimenstheRERfillsupthebasalpartoftheglandularcells
(fig.17).Adifferenceinthemorphologyofglandularcellsofyounganimals
andadultsisthepresenceofmanymicrofilamentsinadults,runningfromthe
junctionalcomplexintothecytoplasm (fig.18).Glandularcellsareabsent
beneaththeepitheliumlininginthepyloricareaofthestomach,whichisalso
Fig. 11. Corpus gland cells of a4days old larva.T=apical tubular system;
Nu =nucleus;Mi =mitochondria; G =Golgi apparatus x 22.000
Fig. 12.Corpus gland cells ina 7days old larva. Some small secretion granules
(arrows).Legends as fig. 11.x 22.000
13
14
'ecognizedbyarelativelywelldevelopedmuscularwall.Thecellsoftheepihelial liningaresimilartothose inthecorpusregion.
. pHindicator
test.
ThepHindicatormethyl-red (colourchange fromyellowtoredbetweenpH
nd4,4)onlyshowsaredcolourwhen injected intothestomachof13daysan<
lderlarvae.However,afterinjectingalargedoseofindicator this colour
hangewasnotnoticedat13days.Inlarvaeof14daysandolderthecolour
hangewasdistinctlyvisible,evenwith largerquantitiesofindicator.Conge
ed (pH5,2to3,3)changestoblueinlarvaeof15days.Duringthenextfew
aysthereactionwasevenmoredistinct.This shows thattheacidityoftheÏ
achcontents afterfeedingishigherthanpH5inyoung larvaeofupto11,5
tandardlength.DuringthenextfewdaysthepHdropstovaluesof5,2-3,3.
Anumberofolder larvae survivedthetreatmentwith indicator.Theday£
heexperimentthedyeswere foundinthegutlumen.Theircolourswereyellov
ndred(methyl-red,resp.Congo-red),which indicates thatthepHinthegut
jmenishigherthan5.
. Uptake of horse radish peroxidase by
enterocytes.
Aftertheingestionofhorseradishperoxidase,enzymeactivityisnotice
ienterocytesofthesecondsegmentonly (fig.20).Apositive reactionwas i
ialllarvaeandjuvenileswestudied (4to15days).Thelocalizationofthe
îroxidaseactivitysuggests thattheenzymeispresentinthesupranuclearpa
Etheabsorptive cells,evidentlywithinthepinocytoticvesiclesandthelar
ipranuclearvacuoles.Thelengthofthesegmentwithperoxidaseabsorptionan
s locationcorrespondtothesecondsegmentasdetermined histologically.
g. 13.Corpus glandular cells ina12daysoldlarva.Many large zymogengra
îles(Zy)especially inthebasalpartofthecells.Theapical partisfille
thtubules.Legendsasfig.11.x 15.000
g. 14.Theapical portionofacorpus gland cellofa12daysoldlarva.The
ructureoftheapicalplasmamembraneissimilar tothatofthemembranesof
emany apical tubules,sincebothareasymmetrical,x52.000
g. 15.Apical partofacorpus gland cellofa12daysoldlarva,showing
ocytosisofasecretion granule,x26.000
15
16
DISCUSSION
1. Regional differentiation
of the
Intheintestineoflarval Clarias
intestine.
lazera aregionaldifferentiationin
threesegmentscanbemade,similartotheoneinstomachlessfish.In Clarias
larvaetheenterocytesinthedifferentsegmentshavethesamehistological
characteristicsas
instomachlessfish(Yamamoto,1966;Iwai,1969;Gauthier&
Landis,1972;NoaillacDepeyre&Gas,1974,1976;Stroband&Debets,1978).
Similarfeatures,includingthepresenceof
asecondsegmentwithpinocytosisof
macromolecules,havebeendescribedfortheintestineofotherinitiallystomachlesslarvaeoffishspecieswhichpossessastomachinthejuvenileandadult
stages(Iwai&Tanaka,1968;Tanaka,1972).
2. Generalmorphology of the epitheliwn of the stomach.
Forthemorphologyofthestomachepithelium,theresultsaresimilar
to
thoseobtainedbyothersinultrastructuralstudies (Ling&Tan,197SinChelmon
rostratus;
Huebner&Chee,1978in Hoplosternum sp.;jäs&Noaillac-Depeyre,1978
in Ameiurus sp.andNoaillac-Depeyre&Gas,1978,inperch).Alsoinmanylightmicroscopicalstudiesonfishes,onlyonetypeofglandularcellwasfoundinthe
corpus,apartfromthesurfacemucouscells (AlHussaini,1946;Weinreb&Bilstad,
1955;Verma&Tyagi,1974;Moitra&Ray,1977;Reifel&Travill,1978;Weisel,
1973).Thedifferentiationoftheglandularcellsintotwotypes,oneofwhich
producespepsinogenandtheotherhydrochloricacid,isonlyknownforthemammalianstomach (Smit,1968).Specialneckcells,asfoundin Mulloides sp.(AlHussaini,1946),stickleback (Hale,1965)andperch (Gas&Noaillac-Depeyre,1978),
havenotbeenfoundinClarias
lazera, justas
inthesearobin(Blake,1936)in
Colisa sp.(Moitra&Ray,1977),andin
anumberofotherteleostspecies(Reifel
&Travill,1978).
Figs. 16-18.Stomachglandcellsofadult Clarias
lazera.
Fig.16.Apicaltubularsystem.Mi
=mitochondrium;Zy=zymogengranule,x23.000
Fig. 17.RERinthebasalpartofacell,x40.000
Fig.18.Desmosomesandmanymicrofibrillsintheapicalpartofaglandular
cell,x40.000
Fig.19.Adult Cl. lazera',secretion granuleslocatedintheapexofasurface
mucouscell,x18.000
17
Fig. 20.Lightmicrograph ofpart of the gut after horse
radish peroxidase ingestion.Black deposits of reaction product indicate the
presence of enzyme activity. 1= segment 1;3= segment 3;A = anus.
Fig. 21.Functional cell of the epithelial lining of the stomach,x 10.000
The inset shows a lightmicroscopical radioautogram. The arrowpoints to the
H-thymidine labeled nucleus of the cell in the electronmicrograph,x 400
18
Thecellsoftheepitheliallininghavethesameappearanceinallstudied
shspecies.UsuallythesecellsshowPASpositivityintheirapicesafter
cianBlue-PASstaining,andthissuggeststhattheycontainneutralmucopolyccharides.Inpike,however,Reifel&Travill (1978)foundacidcarbohydrates
thesecells.
Development of stomach
functions.
Itisdifficulttodeterminewhenthestomachbecomesactually"functional",
evolumeincreasesrapidlyasfromday8,andoneoftheimportantstomach
nctions,thestorageoffoodmaybeginasofthatday.Thedigestivefunction
thestomachclearlydevelopslater,betweentheformationofsecretiongranules
thecorpusglandcellsandtheproductionofasmuchhydrochloricacidasrearedforreachinglowpHvaluesinthestomachlumen.
Fromthemorphologicalevidenceitcanbeconcludedthatin Clarias
lazera
ecorpusofthestomachisformedbetweenday4whenthefirstglandularcells
efound,andday12whenexocytosisofsecretiongranulesisfirstobserved.
Anacidenvironmentinthestomachasfromday13allowscathepticorpeptic
gestion.Fromday12theapicalvesicularsystemshowsatendencytodevelop
bules,whicharedistinctinadults.Thesevesiclesandtubulespossessmemaneswithacoatingattheinnerside.Thiscoating,alsopresentonthememanesofthemicrovilli,consistofglycoproteinsin Ameiurus nebulosus (Gas&
aillac-Depeyre,1978)andin Perca fluviatilis
(Noaillac-Depeyre&Gas,1978).
similartubulo-vesicularsystemisalsopresentinstomachglandcellsof
liernon-mammalianvertebratesandapparentlyplaysacertainroleinhydrochloc-acidproduction (Sedar,1961).Inmammalsthevolumeofthetubulo-vesicular
stem,asonlyfoundinparietalcells,isininverseratiotothevolumeof
3socalledintracellularcanaliculi,whicharemaximallydevelopedwhenmuch
irochloricacidisproduced (Padykula,1977).Intracellularcanaliculi,however,
reneitherbeenfoundin Clarias
i Chelmon rostratus
lazera, norin A. nebulosus,
P.
fluviatilis,
(Ling&Tan,1975).
Afterday13theacidityincreases.Thepyloricsphincterispresentas
3mday12.Theseresultsareingeneralaccordancewiththeradioautographic
suits;thelabelingindexdecreasedsharplyinthestomachcorpusglandsbetween
f9andday12andinthepyloricareabetweenday12and15.Differentiation
theepitheliummusttakeplacefirstintheintestine,thenintherostral
rtofthestomach,andfinallyinthepylorus.
19
Grizzle&Curd (1978)foundalarvalperiodofapproximately35daysin
logperch (Percina eaprodes),
rearedat22C.Thefirstgastricglandswerefoi
onday27,whereasthepyloricsphincterdevelopedsomedayslater.Thisiso:
interest,asTanaka(1971)concludedforalargenumberofteleostspeciesth;
gastricglandsdevelopafterabout3/4ofthetotallarvalstage.Thestomach
developsveryearlyin Clarias
lazera andthelarvalstageisevidentlyvery
shortwhenthefisharerearedat23-24C.Thelatterisbeginningwhen,at+
30hafterfertilization,exogenousfeedingstarts.Thecompletionofthestoi
isusuallyconsideredtocoincidewiththeendofthelarvalstage (Tanaka,1!
4. Physiological
relevance
of the second gut
segment.
Themorphologyoftheintestinedoesnotchangeafterthetimewhenthe:
machbecomesfunctional.Asthesecondsegmentwiththefacilityforabsorbin;
proteinmacromoleculesremainstobepresentbeyondthisperiod,thisfacilit;
cannotberelatedtothelackofastomachin Clarias
lazera. Thisisunderlii
bythefactthatthelengthofthesegmentremainsthesameandalsobythep
senceofpinocytoticenterocytesintheposteriorintestineofadult
Clarias.
lastargumentisinaccordingwiththeresultsofKrementz&Chapman (1974),
Bergot (1976)andNoaillac-Depeyre&Gas(1979),whofoundpinocytosisinthe
posteriorpartofthegutofadultcatfish {Ictalurus
punctatus),
in15cmla
trout,andinadultperch,respectively.However,supranuclearvacuoleshave:
beenfoundinadult Ictalurus
. Thismightbeattributedtothelongtimeinte
betweenfeedingandfixingofthetissues;starvationcausesthedisappearanc
supranuclearvacuolesbutnotofpinocytosis,aswasfoundin Coregonus larva
(Stroband&Dabrowski,inpress).Intheadultperchthesecondsegmentseeme<
relativelyshort(+111ofgutlength,but_+20%in Clarias
lazera larvae,ju
nilesandadults).Tanaka(1972)studied15fishspecies.In5oftheseheno
cedatthetransitionalstagefromlarvatojuvenile,whenthegastricglands
becomefunctional,thegradualdisappearanceofacidophylicgranules (i.e.su
nuclearvacuoles)intheposteriorintestine.Thiswasnotobservedintheot'
10species.Inhisexperimentssomefixationswerepossiblycarriedoutlong
afterfeeding,andultrastructuralstudiesmightshowpinocytosisintheseca
segmentofthegut.
Thefacilityofabsorbingmacromoleculesisprobablyageneralfeatureo
theteleosteanintestine,butitmightbemorecommoninlarvaeandstomachle
speciesthaninspecimenswithafunctionalstomach.Thephysiologicalroleo
20
"secondsegment"infishlarvaemightbedifferentfromthatinadults.In
irvaethefoodreachesthecaudalintestineinaveryshorttime,andabsorption
:macromoleculesmightbeofquantitativeimportance (Iwai,1968).Inadultspe.mens,includingstomachlessfish,manyhoursgobybeforethefoodreachesthe
»steriorgut,anddigestionofproteinthenmaybeinanadvancedstage.That
(sorptionofundigestedfoodinthesecondgutsegmentplaysaroleinfishlarieisalsoindicatedbytheabsenceofsupranuclearvacuolesinstarved Coregonus
irvae(O.C.).TheobservationofIwai(1969)thatafterfeedingthenumberof
'sosomesincarplarvaeincreasestogetherwiththesupranuclearvacuolespoints
iintracellulardigestionofabsorbedmaterial.
Inadults,eveninstomachlessfish,absorptionofproteintakesplacemain'intheanteriorpartoftheintestine (Shcherbina&Sorvatchev,1969;Stroband
jVanderVeen,inpress;studyingcarpandgrasscarprespectively).Thereforeit
emslikelythatadultstomachlessfishareabletodigestproteinsefficiently
thoughapepsin-hydrochloricacidsystemisabsent (Creach,1963;Jany,1976).
fferenceshavenotyetbeenfoundinthepartoffoodproteinabsorbedbyfishes
thorwithoutastomach.Thequestioniswhyotherspeciesdevelopedastomach
dapepsin-HClsystem.Otherstomachfunctions,e.g.storageoffood,partioningoffooditems,ordefenceagainstmicroorganismsmayhavetobeconsidered
thisrespect.
Renewal of the
epithelium.
3
Therandomdistributionof H-thymidine labeledcellsinthedigestivetract
itheliumintheearlystagesofdevelopmentisinaccordancewiththeresults
Romboutetal.(inpress)obtainedin Barbus aonohonius larvae,inwhichthe
Delingindexsharplydeclinedbeforethefirstexogenousfeeding.In Clarias
zera thedecreasewasnoticedtwodayslater.Inbothspecies,thelocationof
jeledcellsduringthisdecreasechangestowardsthebasalpartsoftheinteslalmucosalfolds.Thepresenceofthymidinelabelinnucleioffunctional
Lis(glandularandsurfacestomachcellsaswellasabsorptivecellsin
xrias lazera larvae)wasobservedalsobyRomboutetal.andbyStroband&
jets(1978)intheintestinalabsorptiveenterocytesof Barbus larvaeand
senilegrasscarp,respectively.Theabilitytoproliferatewasalsofoundin
ictionalsmall-intestinalcellsin Xenopus laevislarvaebyMarshall&Dixon
)78).
21
Itisinteresting thatnotwithstanding thedecreasing labeling index,wh<
thetissueshavedifferentiated andbecome functionally active,onlyfunction;
proliferating cellsappeartobepresent.This incontrast tothemammaliansi
intestine,whereonlyundifferentiated cryptcellsare abletoproliferate
(Lipkip,1973).Theresults indicatethat fishenterocytes andstomach cells
loosetheirproliferative activityinarelatively latestageofdifferentiat:
TheauthorswishtothankProf.Dr.LucyP.M. Timmermans andProf.Dr.J.W.M.0:
fortheirconstructive comments,MissJ.J. Thiele fortechnical assistance in
electronmicroscopy,andMessrs.W.ValenenH.G.Eleri (T.F.D.L.)forprepar:
the illustrations.Special thanksaredueDr.L.Boomgaartfor correcting
linguisticerrors.
22
REFERENCES
Al-Hussaini,A.H.The anatomy andhistology of the alimentary tract of the
bottomfeeder Mulloides
auriflamma
(Forsk.). J.Morph. 78.» 121-153 (1946).
Bergot,P. Demonstrationwith ruthenium red of deep invaginations in the apical
plasmamembrane of enterocytes in rainbow trout intestine.
Ann. Biol.Anim. Biochem. Biophys.J_6>37-43 (1976).
Blake,I.H.Studies on the comparative histology of thedigestive tube of certain
teleost fishes.J.Morph. 60,77-102 (1936).
Creach,P.V. Les enzymes protéolytiques des poissons.Ann.Nutr.Alim. 17,
375-471 (1963).
Cornell,R. &Padykula,H.A. A cytochemical study of intestinal absorption in
thepostnatal Rat.Anat.Rec. L5_i> 139 (1965).
Gas,N. &Noaillac-Depeyre,J. Types cellulaires de l'estomac, assurant la productiondu suc gastrique chez Ameiurus nebulosus L. Biologie Cellulaire 31,
181-190 (1978).
Gauthier,G.F. &Landis,S.C. The relationship of ultrastructural and cytochemical
features toabsorptive activity in the Goldfish intestine.Anat.Rec.172,
675-701 (1972).
Grizzle,J.M. &Curd,M.R. Posthatching histological development of the digestive
system and swimbladder of logperch, Peraina oaprodes . Copeia 3^ 448-455
(1978).
Hale,P.A. Themorphology andhistology of thedigestive systems of two fresh-
waterteleosts, Vo&ciVia reticulata
and Gasterosteus
aauleatus.
J. Zool.J_46, 132-149 (1965).
Huebner,E. &Chee,G.Histological and ultrastructural specialization of the
digestive tract of the intestinal airbreather Hoplosternum
thoraoatum
(Teleost). J.Morph. 157,301-328 (1978).
Iwai,T.Fine structure and absorption patterns of intestinal epithelial cells
in rainbow trout alevins.Z.Zellforsch. 9J_,366-379 (1968).
Iwai,T.Fine structure of gut epithelial cells of larval and juvenile carpdurinj
absorption of fat and protein.Arch.Histol.jap. 30,183-189 (1969).
Iwai,T. &Tanaka,M. The comparative study of the digestive tract of teleost
larvae III.Epithelial cells in the posterior gut ofhalfbeak larvae.
Jap. Soc. Sei. Fish. 34,44-48 (1968).
Jany,K.D. Studies on thedigestive enzymes of the stomachlessbonefish
Cavassius
auvatus Gibelio (Bloch): Endopeptidases. Comp.Biochem. Physiol.53B,
31-38 (1976).
Krause,W.J. & Cutts,J.H. The postnatal development of the alimentary canal in
theopossum: III small intestine and colon.J. Anat. 123,21-46 (1977).
Kraehenbuhl,J.P.&Campiche,M.A. Early stages of intestinal absorption of
specific antibodies in thenewborn. J. Cell Biol. 42_,345-365 (1969).
Krementz,A.B. &Chapman,G.B.Ultrastructure of the posterior half of theintestineof thechannal catfish, Iatalurus
punatatus.
J.Morph. 145, 441-482
(1974).
Lin,E.A. &Tan,C.K. Fine structure of the gastric epithelium of theCoral fish,
Chelmon rostratus
Cuvier.Okajmas Fol.Anat. Jap.5J_,285-310 (1975).
Lipkin,M. Proliferation and differentiation of gastro intestinal cells.
Physiol. Rev.53,891-915 (1973).
Marshall,J.A.&Dixon,K.E.Cell proliferation in the intestinal epithelium of
Xenopus laevis tadpoles.J. Exp. Zool. 203,31-40 (1978).
Mepham,T.B. &Smith,11.W.Aminoacid transport in the golsfish intestine.
J. Physiol.J_84,673-684 (1966).
23
Moitra,S.K.&Ray,A.K.Morphohistologyofthealimentarycanalofanindian
freshwaterperch Cotisa faoiata (Bloch)inrelationtofoodandfeeding
habits.Anat.Anz.141,37-58(1977).
Noaillac-Depeyre,J.&Gas,N.Absorptionofproteinmacromoleculesbytheenterocytesofthecarp {Cyprinus oarpio L). Z.Zeilforsch,146,525-541
(1973a).
Noaillac-Depeyre,J.&Gas,N.Miseenévidenced'unezoneadaptéeautransport
desionsdansl'intestinedecarpecommune {Cyprinus oarpio L).
C R .Acad.Sei.(Paris)276,773-776(1973b).
Noaillac-Depeyre,J.&Gas,N.Fatabsorptionbytheenterocytesofthecarp
{Cyprinus oarpio L).Cell.Tiss.Res.J_55,353-365 (1974).
Noaillac-Depeyre,J.&Gas,N.Electronmicroscopicstudyongutepitheliumof
thetench {Tinoa tinea L)withrespecttoitsabsorptivefunctions.Tissue
&Cell,8,511-530 (1976).
Noaillac-Depeyre,J.&Gas,N.Ultrastructuralandcytochemicalstudyofthegastricepitheliuminafreshwaterteleosteanfish {Peroa
fluviatilis).
Tissue&Cell K), 23-37 (1978).
Noaillac-Depeyre,J.&Gas,N.Structureandfunctionoftheintestinalepitheli.
cellsintheperch {Peroa fluviatilis
L).Anat.Rec.195,621-640 (1979).
Padykula,H.A.Thedigestivetract.In:"Histology"(L.Weiss&R.O.Greep,eds.]
NewYork:McGraw-Hill (1977).
Pearse,A.G.E."Histochemistry,theoreticalandapplied".London:
J.&A.ChurchillLtd.(1972).
Rombout,J.H.W.M.,Stroband,H.W.J.&Verstijnen,C.P.H.J.Proliferationand
differentiationofintestinalepithelialcellsduringdevelopmentof Barbus
oonohonius {Teleostei,
Cyprinidae). Submitted toJ.Exp.Zool.(June,1980)
Romeis,B."Mikroskopischetechnik"München:OldenbourgVerlag (1968).
Reifel,C.W.&Travill,A.A.Structureandcarbohydratehistochemistryofthe
stomachineightspeciesofteleosts.J.Morph.158,155-168(1978).
Sedar,A.W.Electronmicroscopyoftheoxynticcellinthegastricglandsofthe
bullfrog, Rana eatesbiana. j. Biophys.Biochem.Cytol.JJ),47-57(1961).
Shcherbina,M.&Sorvatchev,K.Afewdataconcerningtheabsorptionofamino
acidsinthedigestivetractoftwoyearsoldcommoncarp.Vopr.Prud.Rubo1
j_6,315-322 (1969).
Smit,H.Gastricsecretioninlowervertebratesandbirds.In:"Handbookof
Physiology"sect.6."AlimentaryCanal"(CF.Code,ed)Vol.V2791-2805.
Washington,D.C.:Am.Physiol.Soc. (1968).
Stroband,H.W.J.Growthanddietdependantstructuraladaptationsofthedigestivetractinjuvenilegrasscarp {Ctenopharyngodon idella Val.).
J.Fish.Biol._1_U167-174(1977).
Stroband,H.W.J.&Debets,F.M.H.Theultrastructureandrenewaloftheintestin;
epitheliumofthejuvenilegrasscarp, Ctenopharyngodon idella (Val.)Cell.
Tiss.Res.187,181-200(1978).
Stroband,H.J.W.,v.d.Meer,H.&Timmermans,L.P.M.Regionalfunctionaldifferei
tiationinthegutofthegrasscarp, Ctenopharyngodon idella (val.).
Histochemistry64_,235-249 (1979).
Stroband,H.W.J.&Dabrowski,K.R.Morphologicalandphysiologicalaspectsofth<
digestivesystemandfeedinginfreshwater fishlarvae.In:"LaNutrition
desPoissons"CNERNA (Paris)inpress.
Stroband,H.W.J.&v.d.Veen,F.H.Thelocalizationofproteinabsorptionduring
thetransportoffoodalongtheintestineofthegrasscarp, Ctenopharyngodon
idella (Val.),submitted toJ.Fish.Biol.July1980.
Tanaka,M.Studiesonthestructureandfunctionofthedigestivesystemin
teleostlarvaeIII.Developmentofthedigestivesystemduringpostlarval
stage.Jap.J.ofIchthyol.J_8,164-174 (1971).
24
'anaka,M.Studiesonthestructureandfunctionofthedigestivesystemin
teleostlarvaeV.Epithelialchangesintheposteriorgutandprotein
ingestion.Jap.J.Ichthyol. \9_, 172-180 (1972).
'erma,S.R.&Tyagi,M.P.Acomparativehistologicalstudyofthestomachofa
fewteleostfishes.Morph.Jahrb.J_20,244-253 (1974).
(einreb,E.L.&Bilstad,N.M.Histologyofthedigestivetractandadjacent
structuresoftherainbowtrout, Salmo gairdneri irideus. Copeia3^
194-204 (1955).
'eisel,G.F.Anatomyandhistologyofthedigestivesystemofthepaddlefish
(Polyodon spathula). J.Morph.140,243-256 (1973).
amamoto,T.Anelectronmicroscopestudyofthecolumnarepithelialcellinthe
intestineoffreshwaterteleosts:Goldfish (Carassius auratus) andrainbow
trout {Salmo irideus). Z.Zeilforsch.72,66-87 (1966).
25
CURRICULUM VITAE
Het levenslichtwerd doormijaanschouwd in Zeistop 31 december 1946.
Hoewel naar ikmeen nooit doormijn ouders opgejaagd,voelde ik,als ieder
kind,al snel dat ik geborenwas ineen samenlevingwaarin "het presteren"
vangrotebetekeniswordt geacht.Grootwas dan ook mijn frustratie toenik
naeenvliegende start (ikwerd pas eind februari 1947verwacht)al op de
openbare kleuterschool "Griffensteijn" in Zeist mijn eerste doubleren te
verwerken kreeg.Achteraf heeft 3 jaar fröbelen stellig inpositieve zin aan
mijnvormingbijdragen.
Later kon ik aan demaatschappelijke verwachtingen voldoen doorhet
slagenvoorhet toelatingsexamen,na daarop uitstekend voorbereid te zijn op
dejongensschool derEvangelische Broedergemeente.
Hetvoorbereidend wetenschappelijk onderwijswerd aangevangen in 1959
aanhet IeChristelijk Lyceum te Zeist.Eenpoging tothetvolgen vanhet
Gymnasiumwas totmislukken gedoemd,maar in 1965werd het einddiploma H.B.S.-B
verworven.
Na eenkorte periode vanarbeidbijPhilips telecommunicatie industrie
teHilversum volgdeeenoproep voormilitaire dienst. Dezewerd vervuld van
januari 1966 tot juli 1967,grotendeels bij een infanterie bataljon teErmelo.
Hoewel strepenvoormijnietwarenweggelegd,behaalde ikhet diploma "militaire lichamelijke vaardigheid" in 1967,ietswaarop ikvanzelfsprekend nóg
trotsben.
In september 1967ben ik,naenig "voorwerk" in diensttijd (aulapockets e.d.) gestart met de studieBiologie aan de Rijksuniversiteit te
Utrecht.Hetkandidaatsexamen werd, totniet alleen mijn genoegen,behaald in
mei 1970en indecember 1972studeerde ik "met lof"afmet als hoofdvak
Vergelijkende Endocrinologie (prof.v.Oordt)en alsbijvakken Plantenfysiologie (Prof.v. Die) enDidaktiek der Biologie (Drs. Saaltink).
Vanaf januari 1973ben ikwerkzaambij de afdeling Dierkunde,later
vakgroep Experimentele Diermorfologie en Celbiologie aan de Landbouwhogeschool
teWageningen. Naast vele andere aktiviteiten zoals onderwijs aanpre-en
postkandidatenvanverschillende studierichtingen enhetbehalen vanhet
diploma "C-deskundige"aanhet ITAL teWageningen in 1978,bleef er tijd
beschikbaar voorhet doenvanhet onderzoek waarvan deresultaten indit
proefschrift zijn neergelegd.