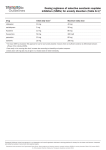
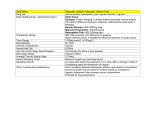
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Pharmacognosy wikipedia , lookup
Pharmaceutical industry wikipedia , lookup
Drug discovery wikipedia , lookup
Prescription drug prices in the United States wikipedia , lookup
Prescription costs wikipedia , lookup
Drug design wikipedia , lookup
Pharmacogenomics wikipedia , lookup
Polysubstance dependence wikipedia , lookup
Plateau principle wikipedia , lookup
Psychopharmacology wikipedia , lookup
Drug interaction wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Neuropharmacology wikipedia , lookup
Theralizumab wikipedia , lookup
DEVELOPMENT OF TOLERANCE AND DEPENDENCE IN BARBITURATE USE: A SYSTEMS MODELING APPROACH by Ali Osman Konuray B.S., Chemical Engineering, Istanbul Technical University, 2005 Submitted to the Institute for Graduate Studies in Science and Engineering in partial fulfillment of the requirements for the degree of Master of Science Graduate Program in Industrial Engineering Bo aziçi University 2008 ii DEVELOPMENT OF TOLERANCE AND DEPENDENCE IN BARBITURATE USE: A SYSTEMS MODELING APPROACH APPROVED BY: Prof. Yaman Barlas ………………… (Thesis Supervisor) Assist. Prof. Aybek Korugan ………………... Assoc. Prof. Cengizhan Öztürk ………………... DATE OF APPROVAL: 22.09.2008 iii ACKNOWLEDGEMENTS I am deeply grateful to Professor Yaman Barlas, my thesis supervisor, for being a great example of an enthusiastic scientist. Without him, I would never indulge the field of System Dynamics which seemed, at first, very distinct from my scientific background. His contribution to my studies in recent years is invaluable. I would like to thank Assist. Prof. Aybek Korugan and Assoc. Prof. Cengizhan Öztürk for taking part in my thesis jury and providing valuable feedback. I would like to thank Ceyhun Eksin and Genco Fas for their company, during and after intense academic moments. I would also like to thank members of SESDYN Research Group for their support and friendship, and all the bright people in the department for contributing to my academic development. I would like to thank Süheyla Ayar for sharing her life with me in the last couple of years. I would like to express my deepest gratitude to my mother Gülsün Konuray for inspiring me with her artistic personality. Her wisdom is my guiding light. Lastly, I would like to thank my late father Dr. M. Mehmet Konuray for installing in me an unfailing respect for science. iv ABSTRACT DEVELOPMENT OF TOLERANCE AND DEPENDENCE IN BARBITURATE USE: A SYSTEMS MODELING APPROACH A system dynamics model is constructed to study the development of tolerance and dependence to phenobarbital in prolonged use. Phenobarbital is a sedative barbiturate drug whose target of action is the brain. Although its use has decreased over the years, phenobarbital is still being prescribed to many patients. As a side effect, phenobarbital enhances the synthesis of its own metabolic enzymes in the liver. This enzyme induction problem causes increased tolerance to phenobarbital over time. Moreover, the brain adapts to the presence of the drug and its sensitivity decreases with time. The resulting decrease in drug effectiveness urges the drug user to increase the dose. A feedback loop results, as the increased dose in turn leads to more metabolic induction and neuroadaptation. Furthermore, the brain’s adaptation to the drug plays a major role in rendering the user dependent on the drug hence complicating withdrawal from the drug. Because adaptive changes persist even after drug intake stops, upon abrupt discontinuation to the drug, the user experiences unwanted rebound effects. The model incorporates phenobarbital absorption, distribution, metabolism, and elimination processes with enzyme induction and neuroadaptation related structures. We start with validating the model by assuming a normal person. We then consider three scenarios: An epilepsy patient, a normal person taking an enzyme inhibitor drug concurrently with phenobarbital, and a normal person adopting different dosing schemes. We finally search for dosing regimens that facilitate gradual withdrawal from the drug so that rebound effects are avoided. Results show that an epilepsy patient is more prone to developing tolerance and dependence. Also, it is shown that concurrent intake of an enzyme inhibitor drug weakens rebound effects after sudden discontinuation since phenobarbital is cleared slower. Experiments with dosing frequencies show that the patient is more prone to tolerance and dependence development if dosing frequency is decreased. Finally, experiments confirm that in order to withdraw from the drug safely, doses should be reduced gradually. v ÖZET BARB TURAT KULLANIMINDA TOLERANS VE BA IMLILIK OLU UMU: B R S STEM MODELLEMES Sürekli fenobarbital kullanımında tolerans ve ba ımlılık olu umunu ara tırmak için bir sistem dinami i modeli kurulmu tur. Fenobarbital, beyni etkileyen sedatif (sakinle tirici) bir ilaçtır. Geçmi yıllara kıyasla kullanımı azalmı olmasına ra men bir çok insan halen fenobarbital kullanmaktadır. Fenobarbital bir yan etki olarak kendini metabolize eden karaci er enzimlerinin sayısını arttırır. Bu enzim artı ı ilaca tolerans olu umuna neden olur. Bunun yanında, zamanla beyin ilaca adapte olur ve dolayısıyla ilaca kar ı hassasiyeti azalır. Bu iki faktör, ilacın etkinli ini azalttı ından kullanıcının aynı etkiyi hissedebilmesi için dozu arttırması gerekir. Artan dozlar metabolizma ve nöroadaptasyon etkilerini güçlendirerek kısır bir geri bildirim döngüsü olu turur. Nöroadaptasyon, kullanıcıyı ilaca ba ımlı kılarak ilacın bırakılmasını zorla tırır. laç alımı kesilmesine ra men adaptif de i imler hemen yokolmaz ve dolayısıyla kullanıcı ilacı bıraktıktan kısa bir süre sonra yoksunluk sendromu ya ar. Kurulan model, fenobarbital ilacının emilimi, da ılımı, metabolizması ve atılımı süreçlerini içermektedir. Enzim artı ı ve nöroadaptasyon mekanizmaları da modele eklenmi tir. Tezde öncelikle normal bir insan ele alınmakta ve model empirik veriler kullanılarak gerçeklenmektedir. Bunun ardından, bir epilepsi hastasının, bir enzim inhibitörüyle birlikte fenobarbital kullanan bir insanın, ve normal bir insanın uyguladı ı farklı doz uygulamalarının modellendi i üç ayrı senaryo incelenmi tir. Son olarak yoksunluk sendromunu engelleyebilecek doz stratejileri ile deneyler yapılmı tır. Sonuçlar epilepsi hastalarının tolerans ve ba ımlılık geli imine daha hassas olduklarını göstermi tir. Di er taraftan, fenobarbital ile beraber enzim inhibitörü bir ilaç alınırsa, fenobarbital vücuttan daha yava atılmakta, dolayısıyla da fenobarbital alımı aniden kesildi inde ortaya çıkan yoksunluk sendromunun iddeti daha az olmaktadır. Farklı doz stratejileriyle yapılan deneylerde, doz alım sıklı ı azaldıkça tolerans ve ba ımlılık geli iminin hızlandı ı görülmü tür. Son olarak, yoksunluk sendromundan kaçınmak için, dozun kademeli bir ekilde azaltılması gerekti i gösterilmi tir. vi TABLE OF CONTENTS ACKNOWLEDGEMENTS.............................................................................................. iii ABSTRACT...................................................................................................................... iv ÖZET ................................................................................................................................. v LIST OF FIGURES ........................................................................................................ viii LIST OF TABLES............................................................................................................ xi LIST OF ABBREVIATIONS.......................................................................................... xii 1. INTRODUCTION ......................................................................................................... 1 1.1. Neurotransmission in the Central Nervous System ................................................ 4 1.2. Definition of Pharmacokinetics .............................................................................. 7 1.3. Pharmacokinetics of Barbiturates ........................................................................... 7 1.4. Action mechanism of barbiturates .......................................................................... 9 1.5. Development of Tolerance and Dependence to Barbiturates ............................... 11 2. RESEARCH OBJECTIVE AND DYNAMIC HYPOTHESIS ................................... 16 3. METHODOLOGY ...................................................................................................... 19 4. MODEL DESCRIPTION ............................................................................................ 20 4.1. Pharmacokinetics Sector....................................................................................... 20 4.1.1. Fundamental Approach and Assumptions .................................................. 20 4.1.2. Description of the Structure........................................................................ 20 4.2. Central Nervous System Sector ............................................................................ 26 4.2.1. Fundamental Approach and Assumptions .................................................. 26 4.2.2. Description of the Structure........................................................................ 28 4.3. Dose Sector ........................................................................................................... 33 4.3.1. Fundamental Approach and Assumptions .................................................. 33 4.3.2. Description of the Structure........................................................................ 33 4.4. Model Parameters ................................................................................................. 35 5. VALIDATION OF THE MODEL .............................................................................. 37 5.1. Simulation Results ................................................................................................ 37 5.1.1. Single Dose ................................................................................................. 37 5.1.2. Continuous Drug Intake with Constant Dose ............................................. 40 vii 5.1.3. Continuous Drug Intake with Dose Increase as a Result of Feedback ....... 42 5.1.3.1. Drug Treatment for Seven Days .................................................... 43 5.1.3.2. Drug Treatment for 20 Days .......................................................... 47 5.1.3.3. Drug Treatment for 60 Days .......................................................... 49 5.2. Model Validity Discussion ................................................................................... 52 6. SCENARIO ANALYSES............................................................................................ 56 6.1. Epilepsy Patient .................................................................................................... 56 6.2. Co-administration of a Drug That Causes Enzyme Inhibition ............................. 60 6.3. Different Dosing Frequencies ............................................................................... 64 7. ANALYSIS OF WITHDRAWAL POLICIES ............................................................ 70 7.1. Withdrawal after 20 days of treatment ................................................................. 70 7.1.1. An unsuccessful regimen ............................................................................ 70 7.1.2. A successful regimen .................................................................................. 72 7.2. Withdrawal after 60 days of treatment ................................................................. 74 7.2.1. An unsuccessful regimen ............................................................................ 74 7.2.2. A Successful Regimen ................................................................................ 76 8. CONCLUSIONS AND FUTURE RESEARCH DIRECTIONS................................. 78 APPENDIX. EQUATIONS OF THE MODEL............................................................... 80 REFERENCES ................................................................................................................ 89 viii LIST OF FIGURES Figure 1.1. Relative safety of barbiturates and benzodiazepines....................................... 1 Figure 1.2. Frequency of barbiturate use among twelfth grade high school students ....... 2 Figure 1.3. Different types of synapses (top). A schematic representation of neurotransmission (bottom) ............................................................................ 5 Figure 1.4. Steps in excitatory and inhibitory neurotransmission ..................................... 6 Figure 1.5. Blood plasma concentration – time data for a single IV dose of thiopental ... 8 Figure 1.6. GABAA receptor-chloride channel complex ................................................... 9 Figure 1.7. Pre- and post-synaptic neuroinhibition by barbiturates................................. 10 Figure 1.8. Proposed mechanism of enzyme induction by phenobarbital ....................... 12 Figure 1.9. Administered doses of PB ............................................................................. 13 Figure 1.10. Change in the intensity of rebound behavior with time .............................. 14 Figure 1.11. The Himmelsbach hypothesis ..................................................................... 15 Figure 2.1. Causal loop diagram for tolerance and dependence development ................ 17 Figure 4.1. Stock-flow structure of the pharmacokinetics sector .................................... 21 Figure 4.2. Saturability of enzyme induction .................................................................. 25 Figure 4.3. Concentration-response data for phenobarbital............................................. 27 Figure 4.4. Stock-flow structure of the CNS sector......................................................... 28 Figure 4.5. Graphical function for IndAdptnRate ............................................................ 29 Figure 4.6. Graphical function for EffSatur..................................................................... 30 Figure 4.7. Graphical function for EffAdptnOnNormClCur ............................................ 31 Figure 4.8. Graphical function for EffPBOnReadptn ...................................................... 32 Figure 4.9. Stock-flow structure for the Dose Sector ...................................................... 34 Figure 5.1. Absorption and distribution of a single dose................................................. 38 Figure 5.2. Increasing chloride current in the brain after a single dose........................... 39 Figure 5.3. Dynamics of enzyme induction and neuroadaptation for a single dose ........ 40 Figure 5.4. Constant doses (a) and drug profiles in the brain (b) in both a seven day and a 20 day treatment. ................................................................................. 41 Figure 5.5. Enzyme induction and neuroadaptation and the resulting chloride current profile when the user takes constant doses (for seven and 20 days)............. 42 ix Figure 5.6. Dose profile (a) and drug amount in the brain (b) in the seven day drug treatment followed by abrupt withdrawal..................................................... 44 Figure 5.7. Enzyme and neuroadaptation dynamics in the seven day drug treatment followed by abrupt withdrawal ..................................................................... 45 Figure 5.8. Behavior of chloride current in the seven day drug treatment ...................... 46 Figure 5.9. Dose profile (a) and drug amount in the brain (b) in the 20 day drug treatment followed by abrupt withdrawal ..................................................... 47 Figure 5.10. Enzyme and neuroadaptation dynamics in the 20 day drug treatment followed by abrupt withdrawal ................................................................... 48 Figure 5.11. Behavior of chloride current in the 20 day drug treatment ......................... 49 Figure 5.12. Dose profile (a) and drug amount in the brain (b) in the 60 day drug treatment followed by abrupt withdrawal ................................................... 50 Figure 5.13. Enzyme and neuroadaptation dynamics in the 60 day drug treatment followed by abrupt withdrawal ................................................................... 51 Figure 5.14. Behavior of chloride current in the 60 day drug treatment ......................... 52 Figure 5.15. Progression of enzyme induction in 20 days of continuous PB use............ 53 Figure 5.16. Comparison of tolerance dynamics generated by the model (a) against real data (b) from Gay et al (1983). ............................................................ 53 Figure 5.17. Tolerance and dependence indicators for 60 days of continuous PB intake, (a) Model output, (b) Real data. ...................................................... 54 Figure 5.18. Differences in withdrawal dynamics between a partially dependent (20 day user) and a completely dependent (60 day user). ........................... 55 Figure 6.1. Dose profiles (a) and drug profiles in brain tissue (b) of both a healthy and an epileptic individual in 20 days of continuous PB use .............................. 58 Figure 6.2. Enzyme and neuroadaptation dynamics in both a healthy and an epileptic individual taking PB for the last 60 days ...................................................... 59 Figure 6.3. Chloride current in a healthy and an epileptic individual) ............................ 59 Figure 6.4. Flurbiprofen average clearance as influenced by fluconazole pre-treatment ................................................................................................. 61 Figure 6.5. Dose profiles (a) and drug amounts in the brain (b) with and without fluconazole pre-treatment ............................................................................. Figure 6.6. Enzyme and neuroadaptation dynamics with and without fluconazole pre- 62 x treatment ....................................................................................................... 63 Figure 6.7. Enzyme and neuroadaptation dynamics in different dosing schemes (No feedback to increase the doses)..................................................................... 65 Figure 6.8. Comparative behavior of chloride current (No feedback to increase the doses) ............................................................................................................ 66 Figure 6.9. Difference in the extent of tolerance development w.r.t dosing schemes (Feedback allowed to increase doses)........................................................... 67 Figure 6.10. Neuroadaptation dynamics for different dosing schemes (Feedback allowed to increase doses) ........................................................................... 67 Figure 6.11. Dependence dynamics for different dosing schemes (Feedback allowed to increase doses) ........................................................................................ 68 Figure 6.12. Behavior of chloride current in different dosing schemes (Feedback allowed to increase doses) ........................................................................... 69 Figure 7.1. Dynamics of an unsuccessful withdrawal regimen after partial dependence.................................................................................................... 71 Figure 7.2. Severity of withdrawal signs after an unsuccessful dosing strategy in partial dependence ......................................................................................... 72 Figure 7.3. Dynamics in a successful withdrawal regimen after partial dependence ...... 73 Figure 7.4. Severity of withdrawal signs after a successful dosing strategy in partial dependence..................................................................................................... 73 Figure 7.5. Results for an unsuccessful withdrawal regimen after complete dependence..................................................................................................... 75 Figure 7.6. Severity of withdrawal signs after an unsuccessful dosing strategy in complete dependence..................................................................................... 75 Figure 7.7. Results for a gradual withdrawal regimen of 30 days following a 60 day drug treatment ................................................................................................ 76 Figure 7.8. Severity of withdrawal signs after a successful dosing strategy in complete dependence..................................................................................................... 77 xi LIST OF TABLES Table 1.1. Classification and properties of barbiturates .................................................... 3 Table 4.1. Main pharmacokinetic parameters used in the model .................................... 36 Table 4.2. Other pharmacokinetic parameters ................................................................. 36 Table 5.1. Initial values for the stocks ............................................................................. 37 xii LIST OF ABBREVIATIONS Adptn Adaptation Arterial Arterial Blood Braincapil Brain Capillary Braintis Brain Tissue C Concentration (of phenobarbital in) ClCur Chloride Current ClCurWOP B Chloride Current Without Phenobarbital CNS Central Nervous System CYP Cytochrome P Eff Effect GABA Gamma Amino Butyric Acid GI Gastrointestinal Ind Indicated Induc Induction M Amount (of phenobarbital in) Norm Normal P Tissue-Blood Partitition Coefficient (of phenobarbital in) PB Phenobarbital Q Blood Volumetric Flow Rate (through) Readptn Re-adaptation Real Realized ThresholdSedat Sedation Threshold V Volume (of) Venous Venous Blood 1 1. INTRODUCTION Barbiturates are classified as central nervous system (CNS) depressants. They act generally on the CNS. In low doses, they cause sedation and as the dose is increased, the user experiences hypnosis (i.e. sleep). Further increase in the dose results in anesthesia and finally coma. Overdose of barbiturates causes severe respiratory depression and may lead to death. For instance, Jimi Hendrix, the famous rock artist, died of barbiturate overdose in the year 1970. Because of having high abuse potential, they are being replaced by the safer benzodiazepines. Figure 1.1 gives an idea about the relative safety of barbiturates and benzodiazepines. Figure 1.1. Relative safety of barbiturates and benzodiazepines (Katzung 2004) The dose-effect relationship of barbiturates is rather linear and lethal overdoses are more likely. On the other hand, this relationship is saturable for benzodiazepines. At high doses, as the dose is further increased, CNS depression stays almost constant. This enables a wider margin of safety. 2 Despite their high abuse potential, barbiturates are still being used as anticonvulsants (i.e. anti-epileptic drugs), intravenous anesthetics, and death inducing agents (Hardman and Limbird, 2001). Furthermore, a lot of people still use barbiturates for sedation or to fall asleep. Alarmingly, a statistical study revealed that the frequency of barbiturate use among twelfth grade high school students in the U.S. has increased slightly over the last few years (See Figure 1.2 below). Figure 1.2. Frequency of barbiturate use among twelfth grade high school students (From http://www.monitoringthefuture.org) Barbiturates are classified with respect to their onset and duration of action. However, the action mechanism is the same for all barbiturates. Different barbiturate classes are tabulated in Table 1.1. 3 Table 1.1. Classification and properties of barbiturates (Hardman and Limbird, 2001) CLASS COMPOUND (TRADE NAMES) ROUTES OF ADMINISTRATION Ultra-shortacting Methohexital (BREVITAL) I.V.† HALFLIFE, HOURS 3-5* Thiopental (PENTHOTAL) I.V., rectal 8-10* Pentobarbital (NEMBUTAL) Oral, I.M. †, I.V., rectal 15-50 Secobarbital (SECONAL) Oral, I.M., I.V., rectal 15-40 Amobarbital (AMYTAL) Oral, I.M., I.V. 10-40 Aprobarbital (ALURATE) Butabarbital (BUTISOL, others) Oral 14-34 Oral 35-50 Butalbital Oral 35-88 Mephobarbital (MEBARAL) Phenobarbital (LUMINAL, others) Oral 10-70 Oral, I.M., I.V. 80-120 Short-acting Intermediateacting Long-acting † THERAPEUTIC USES Induction and/or maintenance of anesthesia Induction and/or maintenance of anesthesia, preoperative sedation, emergency management of seizures Insomnia, preoperative sedation, emergency management of seizures Insomnia, preoperative sedation, emergency management of seizures Insomnia, preoperative sedation, emergency management of seizures Insomnia Insomnia, preoperative sedation Marketed in combination with analgesic agents Seizure disorders, daytime sedation Seizure disorders, status epilepticus, daytime sedation I.M.: intramuscular injection, I.V.: intravenous administration * Value represents terminal half-life due to metabolism by liver; redistribution following intravenous administration produces effects lasting only a few minutes 4 Other than the therapeutic uses mentioned in Table 1.1, some barbiturates have had different uses. For example, other than its common use as an inducer of anesthesia, the ultra-short acting thiopental is used in large doses in the United States to execute prisoners on death row. In lower doses, it is sometimes used as a truth serum. The drug does not itself force people to tell the truth, but is thought to make subjects more likely to be caught off guard when questioned (Stevens and Bannon, 2007). Barbiturate use can cause dependence. This dependence may be psychological in the initial stages of barbiturate treatment. However, as treatment continues, tolerance and then physical dependence develops. As people develop tolerance for barbiturates, they may need more of the drug to get the desired effect. This can lead to an overdose. As Weil and Rosen (2004) point out in From Chocolate to Morphine, “People who get in the habit of taking sleeping pills every night to fall asleep might start out with one a night, progress to two, and then graduate to four to get the same effect. One night the dose they need to fall asleep might also be the dose that stops their breathing." Overdoses occur because tolerance to the lethal effects of the drug is less than tolerance to its therapeutic effects (e.g. sedation). In physical dependence, the user experiences difficulties in stopping drug treatment. Upon discontinuation of the drug, the user experiences a withdrawal syndrome in which he/she goes through a state of rebound hyperexcitability manifested as excessive nightmarish dreaming, restlessness, irritability, and convulsions (Liska, 2001). Although their use is decreasing, mechanism of action of barbiturates is just recently being clarified. Before reviewing the mechanism, it would be useful to briefly overview first the subject of neurotransmission and then pharmacokinetics. 1.1. Neurotransmission in the Central Nervous System Neurotransmission means the communication of nerve cells (i.e. neurons). This is accomplished by billions of interconnected neurons. The point where two neurons meet is called a synapse. Different types of synapses exist and these are shown in Figure 1.3. 5 Figure 1.3. Different types of synapses (top). A schematic representation of neurotransmission (bottom) (From http://www.answers.com/topic/synapse?cat=health) The message between two neurons is conveyed through synapses via substances called neurotransmitters. Neurotransmitters are stored in specialized sacs (i.e. vesicles) inside the presynaptic nerve endings (i.e. nerve terminals). When a reversal of electrical charge is experienced in the nerve terminal, the vesicles translocate and bind to the neuronal membrane. This process is called docking. The reversal of charge is called the action potential. It is accomplished through an influx of sodium ions and efflux of potassium ions through specialized ion channels located on the axon of the presynaptic neuron. This depolarization is conveyed to the nerve ending and causes ion channels to open and allow an influx of calcium. The influx of calcium ions induces the release of the neurotransmitter to the synaptic cleft by exocytosis of the docked vesicles. The neurotransmitter then travels to the postsynaptic neuron and binds to specific receptor proteins on its membrane and changes the membrane electrical potential. If the neurotransmitter is excitatory, an influx of sodium ions to the postsynaptic neuron causes depolarization and this initiates an action potential in the neuron. However, if the 6 neurotransmitter is inhibitory, an influx of chloride and potassium ions occurs which hyperpolarizes the membrane and thus an action potential is inhibited (Hardman and Limbird, 2001). In figure 1.4, inhibitory and excitatory neurotransmission are summarized. Figure 1.4. Steps in excitatory and inhibitory neurotransmission (Hardman and Limbird, 2001) The most widespread excitatory and inhibitory transmitters in the CNS are glutamate and gamma-aminobutyric acid (GABA), respectively (Powis and Bunn, 1995). As mentioned previously, there exist receptors on neuronal membranes that are specialized to bind neurotransmitters. Each receptor is specialized to bind a specific type of neurotransmitter. Furthermore, there are many sub-types of a receptor for a specific 7 neurotransmitter and functions of each of these subunits are modulated by different mechanisms (Hardman and Limbird, 2001). 1.2. Definition of Pharmacokinetics There are several phases before an administered drug causes a response. After administration, the drug goes through many phases during which it may lose effectiveness. After oral administration, the drug must dissolve in stomach fluids, and it must be absorbed from the gastrointestinal tract. Once absorbed, it is directly transported to the liver via the hepatic portal vein. The metabolism in liver at this stage is referred to as first-pass metabolism. In drug development, it is aimed to design drugs that have little first-pass metabolism since it has a negative impact on drug efficacy. Furthermore, a drug may also undergo elimination in different regions such as the gastrointestinal wall which too is an undesired property. After first-pass metabolism, the remaining drug enters blood circulation and reaches the target organ. There, it binds its receptor to exert its effect. While in blood circulation, the drug is transported to the liver once more and it undergoes further elimination. Also while in circulation, it may bind to blood plasma proteins or tissues of different organs. Once bound, a drug molecule is ineffective. This process of drug delivery in the body is referred to as pharmacokinetics. 1.3. Pharmacokinetics of Barbiturates Most barbiturates are rapidly absorbed into the blood following oral intake. The most important factor that plays a role in the entrance of a barbiturate into the brain is its lipid solubility. To exemplify the differences in pharmacokinetic profiles of barbiturates, we consider two barbiturates: ultra-short acting thiopental and long acting phenobarbital (See Table 1.1). Due to its high lipid solubility, the ultra-short acting thiopental has a very rapid onset of effects in the CNS. In comparison, the long acting phenobarbital has low lipid solubility and thus penetrates into the brain slower. 8 In order to be cleared from the body, barbiturates must be transformed into more water-soluble forms so that they can be filtered in the kidneys. Only insignificant quantities (less than 1per cent) of thiopental are excreted unchanged in the urine. Unlike thiopental, 20 to 30 percent of the administered dose of phenobarbital is excreted unchanged. The elimination-half life of phenobarbital is 4 to 5 days. For thiopental, the situation is much more complex. Upon intravenous administration, thiopental rapidly penetrates into the brain due to its very high lipid solubility and if the dose is sufficient, produces loss of consciousness in one circulation time. The blood plasma-brain equilibrium is reached in less than a minute. After that, thiopental diffuses out of the brain and out of other tissues that receive high blood supply and is redistributed to all the remaining less perfused tissues such as muscle and fat. It is because of this rapid redistribution that a single dose of thiopental is very short acting (Katzung, 2004). The redistribution phenomenon causes the half-life of thiopental to be time dependent. Initially, the fall in plasma concentration is very rapid corresponding to a half-life of less than ten minutes. It is denoted as t1/2α in Figure 1.5 below. After redistribution to less perfused areas, the fall of concentration slows down. The half-life increases to more than ten hours. This half-life is denoted as t1/2β in the figure. Figure 1.5. Blood plasma concentration – time data for a single IV dose of thiopental (From http://www.accessmedicine.com/popup.aspx?aID=414128&print=yes) 9 1.4. Action mechanism of barbiturates It was shown that barbiturates exert their CNS-depressant effects by both potentiating the inhibitory effects of GABA and suppressing excitatory effects of glutamate. However, suppression of excitatory neurotransmission does not contribute to their sedative/hypnotic effects (Powis and Bunn, 1995; Joo et al., 1999). At low to moderate concentrations, barbiturates bind to the GABAA receptor. The GABAA receptor is a sub-type of GABA receptors which is classified as a ligand-gated ion channel meaning that the binding of a ligand (a molecule) to the receptor causes the ion channel to open. The GABAA receptor is composed of different sub-units. The distribution of these sub-units in the CNS is widespread and heterogeneous and this heterogeneity has yet to be fully defined (Hardman and Limbird, 2001). Schematically, the GABAA receptorion channel complex is as in Figure 1.6. Figure 1.6. GABAA receptor-chloride channel complex. There are five binding sites (subunits) on the complex (From http://www.ifcc.org) By binding to its specific site, barbiturates enhance the inhibitory chloride ion currents mediated by GABA. Essentially, barbiturates increase the time for which GABAactivated channels are open. At higher concentrations, they activate the chloride channels even in the absence of GABA. This action is regarded as postsynaptic inhibition. In 10 addition to postsynaptic effects, barbiturates induce GABA-mediated presynaptic inhibition as well. This takes place in axo-axonic synapses (See Figure 1.3). GABA released from the ending of the inhibitory neuron binds to GABA receptors on the terminal of the excitatory neuron and causes a modest depolarization which decreases excitatory neurotransmitter release. It was also shown that especially at higher concentrations, barbiturates directly suppress excitatory transmission mediated by glutamate. The postand pre-synaptic inhibition effects of barbiturates are shown in Figure 1.7. Figure 1.7. Pre- and post-synaptic neuroinhibition by barbiturates (Powis and Bunn, 1995) Also, at anesthetic concentrations, barbiturates inhibit calcium influx to the presynaptic nerve ending and thus reduce transmitter release. In addition to these, barbiturates reduce axonal conduction through ion channels and thus increase the threshold for electrical excitability and decrease the rate of rise of the action potential. However, these 11 effects are realized at very high concentrations which are practically irrelevant (Powis and Bunn, 1995). 1.5. Development of Tolerance and Dependence to Barbiturates Barbiturates have been shown to cause the phenomenon of enzyme induction. In the liver, there exists a system of enzymes that are responsible for converting many endogenous and exogenous substances into active and/or inactive forms. The so-called cytochrome P450 family of enzymes constitutes the majority of the enzyme population in the liver (Hardman and Limbird, 2001). By convention, cytochrome enzymes have the prefix CYP. The CYP enzymes catalyze various destructive reactions such as oxidation. The inducing effect of barbiturates causes more enzymes to be synthesized and thus a faster metabolism of the substrates of these enzymes. When the set of substrates include the drug itself, this is called autoinduction. In time, a tolerance to the barbiturate occurs and higher doses are required to induce the same drug effect. Among barbiturates, phenobarbital (will be denoted by PB hereafter) is the most potent inducer of CYP2C subfamily of enzymes. Since PB itself is mostly metabolized by this subfamily of enzymes (Tanaka, 1999), it has autoinduction properties. This was also reported by Magnusson (2007). Induction of enzymes by PB in rats is studied by Magnusson et al. (2006). Their purpose is to characterize the magnitude, time course, and specificity of PB mediated enzyme induction, and to develop an integrated pharmacokinetic model that represents the change in the activities of different CYP enzymes. In another study, Raucy et al. (2002) work with human liver cells in vitro to investigate the extent of induction of CYP2C enzymes by several inducers including PB. The mechanism of induction is not fully understood. Nevertheless, there is progress. A variety of drugs and xenobiotics cause enzyme induction and each is believed to have its own mechanism. It is believed that inside liver cells, there exist several receptors that respond to different types of chemicals. These receptors are called nuclear receptors. An excellent review on the topic is provided by Handschin and Meyer (2003). It is believed 12 that upon exposure to the chemical to which it is sensitive, these nuclear receptors translocate to the nucleus of the cell and bind to specific regions on the DNA molecule and modulate protein synthesis. PB is believed to activate the CAR (Constitutively Active Receptor) type of nuclear receptors. The proposed mechanism of enzyme induction by PB in a liver cell is shown in Figure 1.8. Figure 1.8. Proposed mechanism of enzyme induction by phenobarbital (Simplified from Zelko and Negishi, 2000). PB: Phenobarbital, HSPs: Heat Shock Proteins, CAR: Constitutively Active Receptor, RXR: Retinoid X Receptor Upon exposure to PB, the heat shock proteins dissociate from the CAR receptor by a dephosphorylation reaction. The true PB target in this event is not known. Upon liberation, the CAR receptor enters the nucleus and it is activated by a phosphorylation reaction. The CAR receptor then heterodimerizes (i.e. combines with another molecule of different 13 structure) with the RXR type of receptor and finally binds to a specific area on the DNA. The binding eventually leads to an increased rate of enzyme synthesis. The increased rate of enzyme synthesis in turn leads to faster metabolism of the drug and thus a tolerance develops to the effects of the drug. Development of tolerance to PB during chronic treatment is studied on rats by Gay et al. (1983). Their aim is to quantify the development of tolerance. They give rats two daily doses of PB so as to achieve the same level of CNS depression with each dose. They show that the doses show an increasing trend (Figure 1.9). This is also referred to as pharmacokinetic tolerance. Figure 1.9. Administered doses of PB. Half-filled circles are morning doses, filled circles are total daily doses (sum of morning and evening doses). All doses result in the same level of CNS depression (Gay et al., 1983) Physical dependence to barbiturates develops over a time period of weeks to months as opposed to pharmacokinetic tolerance which peaks in a few days to a week (Hardman and Limbird, 2001). The major cause of physical dependence is brain’s adaptation to the drug. This adaptation is called neuroadaptation. Physical dependence renders withdrawal from barbiturates difficult. Upon withdrawal, a dependent barbiturate user experiences rebound effects such as seizures and is urged to continue the drug. 14 In a research, although PB enhances inhibitory neurotransmission by increasing the rate of GABA binding, it is shown that after rats were treated with PB for a long time, they show decreased GABA binding. It is believed that this is due to an adaptive response by the rats which results in desensitized or down-regulated GABAA receptors (Ito et al., 1996). This down-regulation decreases chloride flow through the channel and thus inhibitory neurotransmission weakens. In the same study by Gay et al., it is shown that upon abrupt cessation of PB treatment, rats experience rebound effects such as ear twitches, tremor, and tail erection. However, these withdrawal syndromes weaken with time as shown in Figure 1.10. Figure 1.10. Change in the intensity of rebound behavior with time. Rats are observed twice daily for withdrawal signs following abrupt termination of 35 days of PB treatment (Gay et al., 1983) It is believed that all chemicals promoting inhibitory neurotransmission trigger similar mechanisms of neuroadaptation. The mechanisms of neuroadaptation induced by chronic ethanol use that lead to tolerance and dependence are studied by many researchers (Brailowsky and Garcia, 1999; Finn and Crabbe, 1997; Kokka et al., 1993; Littleton, 1998). Similar to barbiturates, ethanol acutely promotes the inhibitory effects of the 15 neurotransmitter GABA by increasing chloride ion flow through the GABAA channel. It is being speculated that, as an adaptation, the receptor-channel complex counteracts this effect by changing the composition of its subunits, and thus reducing the chloride flow. Unintended effects such as hallucinations or seizures occur upon withdrawal from chronic ethanol exposure. This is called alcohol withdrawal syndrome. It is believed that the adaptive changes on GABAA receptors and calcium ion channels persist during alcohol withdrawal and contribute to the withdrawal syndrome. This suggestion is in agreement with the Himmelsbach hypothesis which illustrates the development of tolerance and dependence. Schematically, the hypothesis is as in Figure 1.11. Figure 1.11. The Himmelsbach hypothesis (Littleton, 1998) The hypothesis can be applied as well to the barbiturate case since it involves similar neuroadaptative changes and it is shown that a disrupt discontinuation of barbiturate use results in rebound hyperexcitability, characterized by excessive nightmarish dreaming, restlessness, irritability and convulsions. It is generally suggested that barbiturate dosage must be reduced gradually to avoid these unwanted effects (Liska, 2001). 16 2. RESEARCH OBJECTIVE AND DYNAMIC HYPOTHESIS This thesis focuses on phenobarbital (PB) use. Other than its use as a sedative drug, it is also an anti-epileptic drug of choice. Continuous use of PB unfolds interesting dynamics that are likely to be counter-intuitive and thus require careful research. Prolonged use of PB enhances liver enzymes in a few days so that the rate of metabolism approximately doubles (enzyme induction). As the drug is continued, the body tries to counteract the increase in inhibitory neurotransmission by down-regulating the GABAA receptors. This neuroadaptation is much slower than enzyme induction. Peak of neuroadaptation is reached after several weeks and the number of down-regulated receptors comes to stagnation. Down-regulated receptors reduce the efficiency of inhibitory transmission and together with enzyme induction, they decrease the efficacy of the drug. The decreased efficacy urges the drug user to increase the doses. Upon abrupt withdrawal, the drug is cleared much rapidly but the reduced efficiency in inhibitory neurotransmission persists. This disrupts the normal activity of the CNS since excitatory neurotransmission is not balanced by inhibitory neurotransmission, which is manifested by a chloride current lower than normal. The result is a withdrawal syndrome. Nevertheless, as re-adaptation commences with decreasing levels of the drug, the physiology gradually returns to normal and withdrawal syndrome ceases. The causal loop structure is given in Figure 2.1. There are three negative feedback loops in the system. The first one is related to the development of pharmacokinetic tolerance as a result of enzyme induction. The loop is 12-3-1. Sustained levels of PB in the body lead to a higher rate of enzyme synthesis. This leads to a faster PB metabolism and thus the amount of drug in the body decreases. 17 Re-adaptation rate 8 + Adaptation rat e 6 + - + Number of down-regulated recep tors 7 Chloride current + 5 - Enzy me synthesis rate + 2 + Extent of CNS depression 9 Functionality of GABA neurotransmission 4 + 3 M etabolism rat e + + Intensity of withdrawal syndrome 11 - - + Amount of phenobarbital in the body Rate of phenobarbital intake 10 1 Figure 2.1. Causal loop diagram for tolerance and dependence development The second negative feedback loop is related to neuroadaptive changes in the brain. The loop is 4-5-6-7-4. The primary effect of PB is to increase chloride current which leads to the depression of the CNS. If treatment is continued, continuous potentiation of chloride current is counteracted by desensitization of GABAA receptors. This weakens the inhibitory neurotransmission system. There is a third negative feedback loop which is a consequence of the two aforementioned loops. The decrease in inhibitory neurotransmission as a result of increased metabolism and desensitized receptors leads to less CNS depression. This urges the drug user to increase the administered dose, which leads to stronger inhibitory neurotransmission. The loop is 4-5-9-10-1-4. This loop is operational only at later phases when the functionality of inhibitory neurotransmission is weakened. When all three loops are operational, they result in positive feedbacks that lead to continuously increasing doses. The most potent positive feedback is through 18 neuroadaptation rather than enzyme induction and it is 4-5-9-10-1-4-5-6-7-4. Verbally, as functionality of inhibitory neurotransmission weakens as a result of neuroadaptation, the user compensates by increasing the dose which leads to further neuroadaptation and thus inhibitory neurotransmission is further weakened. The aim of this research is to build a simulation model that represents a regular PB user taking into account the two related aspects: Enzyme induction and neuroadaptation. Tolerance development will be traced by monitoring the dose increase decisions of the user. To provide insight on dependence development, the situation after withdrawal will also be studied. In addition to a hypothetical healthy person who takes PB for sedation, we will study three other cases: An epilepsy patient, a person taking another drug that interacts with PB, and a normal person employing dfferent dosing frequencies. These different scenario analyses will improve our insights on prolonged PB use. Finally for the hypothetical healthy person, a feasible dosing scheme during withdrawal will be investigated, so that the unwanted rebound effects are avoided. 19 3. METHODOLOGY In Section 2, we have defined a medical problem that involves several interdependent variables and feedback relationships. Indeed, this is a rather complex system: Human body exposed to an exogenous chemical. To capture the long-term dynamics, one has to study the system as a whole rather than focusing one at a time on individual elements of the system. By creating a mathematical model of the system and defining accurately the relationships, one can unfold the behavior of the system in the long term. System Dynamics (SD) methodology is most suitable for this task. In general, SD is a simulation modeling methodology for studying and managing complex feedback systems. SD models contain sets of differential/difference equations which when solved simultaneously, produce certain dynamics of behavior. The focus is on pattern prediction rather than point prediction, unlike the “black box” statistical models. As a result, SD models are descriptive in the sense that they explain the direct causalities (rather than drawing correlations) that give birth to the dynamic behavior of interest. SD can be applied to all sorts of systems (e.g. businesses, medical systems, socio-economic systems) that contain complex feedback relationships. The methodology first identifies a problem, then develops a dynamic hypothesis explaining the causes of the problem, builds a computer simulation model of the system with regard to the root of the problem, validates the model against structural and behavioral information seen in the real world, suggests policies to address the problem and implements the solution. The process is not purely sequential since one usually finds him/herself visiting some previous steps and revising decisions (Sterman, 2000; Barlas, 2002; Forrester, 1961). 20 4. MODEL DESCRIPTION 4.1. Pharmacokinetics Sector 4.1.1. Fundamental Approach and Assumptions This sector models the absorption, distribution, metabolism, and excretion phases. We model each organ separately and calculate the amount of drug in each organ at a given instance. We regard only the organs and tissues that are large in volume and those that receive high blood supply. These are brain, lungs, heart, muscle tissue, fat tissue, kidney, gastrointestinal tissue, and liver. We model the organs as stock variables, each stock representing the amount of drug accumulated in that organ. Blood is divided into two parts: Arterial blood and venous blood. Arterial blood flows into organs whereas venous blood flows out of organs. Preliminary simulation runs revealed that lungs and heart do not decouple arterial and venous blood phenobarbital (PB) content. That is, for both lungs and heart, the amount of PB entering the organ is practically equal to the amount leaving the organ at a given time. There is no significant drug uptake into or drug elimination in these tissues. Therefore, we do not model lungs and heart as stock variables. We only include the flow of drug from venous blood to arterial blood through the lungs and the flow of drug from arterial blood to venous blood through the heart. Another assumption is that within an organ, the drug is distributed uniformly, so that the concentration of drug inside the organ is equal to the concentration of drug in the blood that flows out of the organ. Finally, in all our simulation experiments, we assume oral administration of PB in the form of tablets. 4.1.2. Description of the Structure The stock-flow structure of the pharmacokinetics sector is given in Figure 4.1 21 VBraintis CBraintis VBraincapil MBraintis BraintisTo Braincapil FR DR CBraincapil Bplasma Braincapil ToBraintis MBraincapil QBrain ArterialToBrain BrainToVenous PFat VFat QFat CFat MFat ArterialToFat FatToVenous QHeart ArterialToVenous MArterial QMuscle CArterial VMuscle MMuscle ArterialTo Muscle VArterial MKidney ArterialTo Kidney VenousToArterial CMuscle QTotal MuscleTo Venous PMuscle QKidney Excretion KidneyToVenous DaysTreatment CKidney VKidney Kexcr MGItissue CGItissue QGItissue Intake2 Absorption VGItissue GItissue ToLiver ArterialToLiver IndInducByPB Intake 5 Intake 4 Intake3 Synthesis CLiver MLiver NormKmet Metabolism LiverTo Venous PLiver QLiver Kmet Figure 4.1. Stock-flow structure of the pharmacokinetics sector kout Rin VLiver Kabs PGItissue <Dose> HalflifeEnzyme ReaIInducbyPB PKidney Intake1 MGIlumen ArterialToGItis MVenous CVenous VVenous EnzymeFactor Degradation 22 As mentioned before, the stocks represent the amounts of drug in different organs. The flows represent the amounts flowing in blood. In modeling absorption and distribution, the assumptions used by El-Masri and Portier (1998) were utilized. Absorption is assumed to follow a first-order rate equation. Its equation is given below. Absorption = Kabs * MGIlumen (4.1) where Kabs (min-1) is the absorption constant, and MGIlumen (mg) is the amount of drug present in the gastrointestinal lumen. To calculate concentrations, we divide the amounts to volumes. For example, the concentration of PB in brain tissue is given by Equation 4.2 below. CBraintis = MBraintis / VBraintis (4.2) where Mbraintis (mg) is the amount of drug in brain tissue, and VBraintis (L) is the volume of brain tissue. The amounts flowing via arterial blood into all organs are assumed to be flow-limited. To exemplify, the rate of PB transfer from arterial blood to brain is given in Equation 4.3 below. ArterialtToBrain = CArterial * QBrain (4.3) where CArterial (mg/L) is the concentration of the drug in arterial blood and QBrain (L/min) is the rate of blood flow through the brain. The outflows of all organs except liver and brain are formulated considering that only unbound drug can flow out of the organ into venous blood. For example, the rate of PB flow from kidney to venous blood is given by Equation 4.4. KidneyToVenous = CKidney * QKidney / PKidney (4.4) 23 where QKidney is the rate of blood flow through the kidney; CKidney is the concentration and PKidney is the tissue-blood partition coefficient in the kidney. The tissue-blood partition coefficient in an organ is simply the equilibrium ratio of the concentration of drug in the blood (mobile) to the concentration of drug bound to tissue (immobile) in that organ. The liver is perfused by both the arterial blood and also by the blood coming from GI tissue via the hepatic portal vein. Therefore, its outflow towards venous blood is LiverToVenous = CLiver * (QLiver + QGItissue) / PLiver (4.5) where CLiver (mg/L) is the concentration of drug in the liver, QLiver (L/min) is the blood flow rate through the liver, PLiver is the tissue-blood partition coefficient in the liver, and Qgi (L/min) is the blood flow rate through the GI tissue. The brain is divided into two parts: Blood (in capillaries) and tissue. Blood in the brain is denoted by the stock “Brain capillary”. The amount of drug flowing from the brain into the venous blood is simply QBrain*CBraincapil where QBraincapil (L/min) is the blood flow rate through the brain and CBraincapil (mg/L) is the concentration of the drug in brain capillaries. The equations for the flow of drug between brain capillary and brain tissue are derived by El-Masri and Portier (1998). We copy these equations in our formulations for BraincapilToBraintis and BraintisToBraincapil as follows. BraincapilToBraintis =VBraintis * DR * Cbraincapil / (1+Bplasma) (4.6) BraintisToBraincapil =VBraintis * DR * CBraintis * FR 4.7) BraincapilToBraintis (mg/min) is the amount of drug diffusing from brain capillaries into brain tissue, BraintisToBraincapil (mg/min) is the amount of drug diffusing out of brain tissue to the capillaries, VBraintis is the volume of brain tissue (ml), DR is the diffusion rate constant (min-1), CBraincapil (mg/L) is the concentration of drug in brain capillary, CBraintis is the concentration of the drug in brain tissue, Bplasma is the bound fraction of drug in red blood cells, and FR is the ratio of free to tissue concentrations of the 24 drug. The values for blood flow rates, organ volumes used in calculating concentrations, partition coefficients, bound fractions and rate parameters are taken from the paper by ElMasri and Portier (1998) and are given in Tables 4.1 and 4.2 (and in the Appendix together with all the equations of the model). Urinary excretion was assumed to be a first-order rate process. It is given in Equation 4.8. Excretion = Kexcr * MKidney (4.8) In modeling metabolism rate (mg/min), we use the following equation. Metabolism = CLiver* Kmet (4.9) As a matter of fact, Kmet is a function of CLiver. This functional relationship underlies the process of enzyme induction. To clarify, we start with the equation for Kmet given below. Kmet = NormKmet * EnzymeFactor (4.10) NormKmet (L/min) is a constant and EnzymeFactor is modeled as a stock variable (See Figure 4.1). Initially, it equals 1, and its inflow and outflow are equal to each other. Its differential equation is given below. d(EnzymeFactor) / dt = Synthesis – Degradation (4.11) As drug concentration in the liver increases, the inflow Synthesis also increases. The following equation holds for Synthesis. Synthesis = Rin *(1+RealInducByPB) (4.12) Rin is the synthesis rate of the enzyme when no drug is present. RealInducByPB is a smoothed version of IndInducByPb, the latter being a saturable function defined by 25 Equation 4.13. We assume a smoothing time of 2 days. The reason for the delay is that enzyme induction is a process of protein synthesis involving several genetic processes such as transcription of genes, mRNA synthesis, etc. which take time. IndInducByPb = E max * CLiver EC 50 + CLiver (4.13) Emax is the maximal induction effect and EC50 (mg/L) is the concentration of the drug that causes half the maximal effect. This function is linear in CLiver for small values of CLiver since when CLiver << EC50, IndInducByPB ≅ Emax * CLiver. On the other hand, when CLiver is large so that CLiver >> EC50, IndInducByPb ≅ Emax, thus the function becomes constant (i.e. the function saturates). We plot the function in Figure 4.2 to clarify further. Emax and EC50 are as given in Table 4.2. 1,2 IndInducByPB 1 0,8 0,6 0,4 0,2 0 0 5 10 15 20 25 CLiver (mg/L) Figure 4.2. Saturability of enzyme induction The outflow Degradation is given by the following equation. Degradation = kout * EnzymeFactor (4.14) 26 where kout= ln(2) which has units of min-1. HalflifeEnzyme To establish a baseline situation, initially (i.e. when no drug is present) we set EnzymeFactor = 1, and we also set Rin = kout. We assume an enzyme half-life of 2 days regarding the information in the literature that half-lives of CYP enzymes range between 1 to 6 days (Michalets, 1998). Other mathematical aspects of the model will be explained where relevant. For numerical values of model parameters, refer to Appendix. 4.2. Central Nervous System Sector 4.2.1. Fundamental Approach and Assumptions As mentioned in Section 1.4, for sedation, the major effect of barbiturates is their potentiation of inhibitory neurotransmission mediated by GABA. The effect on excitatory neurotransmission is seen in relatively high concentrations of the barbiturate. Therefore, the model does not take into account the effects on excitatory neurotransmission. Any adaptation that may be experienced in glutamate (excitatory neurotransmitter) receptors is also not taken into account. In the literature, there is no data regarding the quantitative relationship between PB concentration and GABA mediated inhibitory neurotransmission in humans. However, there are animal data. In their research, Ffrench-Mullen et al. (1993) use in vitro assays from pig brains and derive concentration-response functions for several drugs including PB by measuring peak chloride currents with special equipment. Their results are given in Figure 4.3. To use this data, we assume that pigs and humans respond equally to PB treatment. 27 PB Figure 4.3. Concentration-response data for phenobarbital (Ffrench-Mullen et al., 1993). The concentration-response relationship of phenobarbital given in the figure can be represented by the following equation. Response = Pmax * C PB GABAEC50 + C PB (4.15) where Pmax is the maximum chloride current increase percentage and GABAEC50 is the concentration of PB that causes half the maximal response. The numerical values of these parameters are 600 per cent and 2.79 mg/L, respectively. Neuroadaptation rate is dictated by the extent of inhibitory neurotransmission. However, as the number of desensitized receptors grows very large, neuroadaptation saturates. This is presumably achieved by a decreased adaptation rate rather than by an opposing re-adaptation process. Nevertheless, re-adaptive mechanisms are operational once the drug concentration drops below a certain threshold. In the human CNS, there exist around 100 billion neurons and GABA receptors can be found amongst 60-80 per cent of all neurons. Furthermore, the GABAA subtype of 28 GABA receptors is claimed to be present in ubiquitous amounts (Birnir, 2008). These facts led to the assumption that there are approximately 60 billion GABAA receptors. 4.2.2. Description of the Structure The stock-flow diagram of the CNS sector is given in Figure 4.4. EffPBOnClCur EffPBOn Readptn WithdSignIntensity <CBraintis> EffAdptn OnNorm ClCur ClCurWOPB IndAdptnRate ReadptnFrac EffSatur NormClCur ClCur TotalNoRecep Adaptation NoDownreg Recep Readaptation RealAdptnRate Figure 4.4. Stock-flow structure of the CNS sector EffPBOnClCur is simply the concentration-response function. It is a proxy for how PB presence affects the central nervous system. PB presence not only affects chloride currents, it also influences the rate of re-adaptation. Unless PB concentration in the CNS is below a certain level, re-adaptation is inhibited. The concentration-response function for PB was given in Equation 4.15. We use this function for EffPBOnClCur. EffPBOnClCur is used in the following equation. ClCur = NormClCur*(1+EffPBOnClCur/100) (4.16) It was not possible to find numerical data on chloride currents in the human brain. Thus, we model the chloride current variables as multiplicative factors. We assume that when no drug is present in the body, 29 ClCur= NormClCur = ClCurWOPB = 1 (4.17) We define the number of down-regulated GABAA receptors as a stock variable called NoDownregRecep having units of billions. It is an indicator of the extent of brain’s adaptation to the drug. Its differential equation is given below. d (NoDownregRecep) / dt = Adaptation – Readaptation (4.18) Adaptation = RealAdptnRate * EffSatur (4.19) where Adaptation involves several steps at the cellular level which delay the desensitization of GABAA receptors. Therefore, we model RealAdptnRate as a third order smoothing of IndAdptnRate. The smoothing time is assumed to be 15 days. The rate of adaptation is assumed to be proportional to the discrepancy between a base chloride current (without PB) and the actual chloride current. IndAdptnRate is therefore defined as a function of ClCur / ClCurWOPB and is given in Figure 4.5. EffSatur, as the name implies, slows down neuroadaptation as the number of desensitized receptors approach the total number of receptors. It is therefore defined as a function of NoDownregRecep / TotalNoRecep and is given in Figure 4.6. Figure 4.5. Graphical function for IndAdptnRate. Abscissa is ClCur / ClCurWOPB 30 Figure 4.6. Graphical function for EffSatur. Abscissa is NoDownregRecep / TotalNoRecep As can be seen, the saturation effect is operational after 80 per cent of the receptor population is down-regulated. Since adaptation modifies brain physiology, normal chloride current is affected. NormClCur = ClCurWOPB* EffAdptnOnNormClCur (4.20) EffAdptnOnNormClCur = F (NoDownregRecep/TotalNoRecep) (4.21) where and F is assumed to be a decreasing function given in Figure 4.7. 31 Figure 4.7. Graphical function for EffAdptnOnNormClCur. Abscissa is NoDownregRecep / TotalNoRecep As can be seen, when all receptors are down-regulated, physiology becomes such that chloride current is 30 per cent less than that in a healthy person. In modeling the re-adaptation process, we use the following equation. Readaptation = EffPBOnReadptn* ReadptnFrac * NoDownregRecep (4.22) We assume that there is a critical concentration of the drug above which no readaptation can occur. This is captured by EffPBOnReadptn which is given in Figure 4.8. 32 Figure 4.8. Graphical function for EffPBOnReadptn. Abscissa is EffPBOnClCur Figure 4.8 implies that when PB concentration in the brain is such that when the concentration-response function (i.e. EffPBOnClCur) indicates less than a 70 per cent potentiation of the chloride current, re-adaptation can commence. Finally, to see the intensity of withdrawal signs, we define a variable called WithdSignIntensity which is merely a shifted and inverted version of ClCur and is given below. WithdSignIntensity = - (ClCur – 1) (4.23) The variable is only meaningful after drug treatment stops. We assume that when ClCur drops below its base value of 1, WithdSignIntensity becomes greater than 0 implying that inhibitory neurotransmission is compromised. Given that WithdSignIntensity is greater than 0, the larger it is, the less the inhibitory neurotransmission and the more likely the outburst of a withdrawal syndrome. To interpret this variable, we will first establish reference values that imply insignificant and significant withdrawal signs. This will be clarified in Section 5. 33 4.3. Dose Sector 4.3.1. Fundamental Approach and Assumptions PB depresses the activity of the CNS by enhancing chloride currents. We assume that the PB user is content as long as his/her level of CNS depression corresponds to a chloride current that is 250 per cent higher than normal. This value was found by regarding the expected concentration of PB in the venous blood. The therapeutic range of PB plasma levels (sampled from venous blood) is 10-40 mg/L. Since sedation is the primary effect and the user is assumed to take the drug for this purpose, the venous blood concentration is expected to be around 10 mg/L. Hence the threshold was calibrated to yield our expectation. We assume the user is urged to take a larger dose at the next dosing time if the chloride current drops below the sedation threshold. That is, if he/she had taken the single daily dose, say, 2 hours before he/she has realized the reduced effectiveness, he/she waits until the next day to increase the dose. Therefore, the frequency of dosing does not change. We also assume constant dose increments. 4.3.2. Description of the Structure We model the amount of administered dose as a stock which only has a single inflow. The stock-flow structure is given in Figure 4.9. The single differential equation in this sector is given below. d(Dose)/dt = DoseIncr (4.24) where DoseIncr=11*12*13*14*IncrRate (4.25) 34 ThresholdSedat <ClCur> IncrRate I1 <DaysTreatment> Dose DoseIncr I2 <Time> I3 LoadDose I4 Figure 4.9. Stock-flow structure for the Dose Sector IncrRate is equal to 10 mg/min. The variables I1, I2, I3 and I4 are 0,1 binary indicator variables. We want the dose dynamics to be operational only after the initial dose is effective. The variable I3 serves this purpose and is given below. I3 = IF THEN ELSE(Time>1440, 1, 0 ) (4.26) That is, if Time is later than 1440 minutes, I3 = 1, and I3 = 0 otherwise. The purpose of the variable I2 is to stop dose increase decisions after the end of PB treatment. It is given in Equation 4.27. I2 = IF THEN ELSE (Time<DaysTreatment, 1, 0) (4.27) During drug treatment, I1 helps start the inflow when chloride current is below the sedation threshold and stop it when the threshold is exceeded. I1 = IF THEN ELSE (ClCur<ThresholdSedat, 1, 0) (4.28) where Sedation threshold is 2.5 as explained and dose incr rate is calibrated to give 1/3 mg/min. 35 To model constant dose increments, we use I4. For example, in one-a-day dosing and for a constant increment of 30 mg, I4 is as in Equation 4.29. I4 = IF THEN ELSE (MODULO(Time,1440)>=1437, 1, 0) (4.29) Since the inflow DoseIncr, when it is open, equals 10 mg/min, after 3 minutes of inflow, 30 mg accumulate in the stock Dose. Additionally, since the inflow opens before a day is over, the dose increase decision can be implemented at the beginning of the next day. 4.4. Model Parameters The model represents the actual physical structure of a human being. For the pharmacokinetic sector, we use data from a pharmacokinetic modeling study by El-Masri and Portier (1998). Rate parameters are assumed regarding the variations between the three human subjects who had participated in their work. Note that rate parameters naturally vary among humans. The parameters we use in the pharmacokinetic sector are given in Tables 4.1 and 4.2. 36 Table 4.1. Main pharmacokinetic parameters used in the model ORGAN VOLUME, L BLOOD PARTITION FLOW, L/min COEFFICIENT, Dimensionless GI tissue 1.19 0.9 1 † Liver 1.925 0.235 2.25 Kidney 0.308 0.875 2.05 Fat 16.394 0.26 1 Muscle 28 1.67 1.12 Brain capillary 0.0447 0.57 Brain tissue 1.3553 Arterial blood 1.556* Venous blood 3.811* Heart 0.2 Lungs 4.475†† * : Blood volume : Contribution of the hepatic portal vein is not included †† : Sum of all flows † Table 4.2. Other pharmacokinetic parameters PARAMETER NUMERICAL VALUE Kabs 0.02 min-1 NormKmet 0.00314 L/min DR 0.02 FR 1.75 Bplasma 0.438 Emax 1.15 EC50 1 mg/L In the CNS sector, we mostly use graphical functions which were explained in detail in Section 4.2.2. 37 5. VALIDATION OF THE MODEL In this section, we present evidences of the model’s validity with respect to the real system. We first present the simulation results under basic assumptions. We then introduce new assumptions and shed light on dynamics generated by different structures in the model. Having presented all relevant results, we then draw comparisons between model outputs and real data and present discussions. 5.1. Simulation Results In these structural validation runs, we assume that the user employs one-a-day dosing. He/she is assumed to continue with 30 mg tablets when allowed after an initial loading dose of 180 mg (i.e. first dose). We use the initial conditions given in Table 5.1 for the stocks. Table 5.1. Initial values for the stocks STOCK INITIAL VALUE All (except MGIlumen 0 and Dose) MGIlumen 180 Dose 30 EnzymeFactor 1 NoDownregRecep 0 5.1.1. Single Dose To observe the initial pharmacokinetic processes such as absorption from the gastrointestinal lumen, distribution to organs and tissues, and elimination, we give the results for the first 300 minutes (5 hours) after the loading dose of 180 mg only. We display only the most informative stocks for this run. 38 MGITissue 6 150 4.5 100 3 mg mg MGILumen 200 1.5 50 0 0 0 30 60 90 120 150 180 Time (Minute) 210 240 270 0 300 30 60 90 120 150 180 Time (Minute) a 270 300 210 240 270 300 210 240 270 300 b MVenous MLiver 20 20 15 15 10 10 mg mg 240 MGITissue : singletablet MGILumen : singletablet 5 5 0 0 0 30 60 90 120 150 180 Time (Minute) 210 240 270 0 300 30 60 90 120 150 180 Time (Minute) MVenous : singletablet MLiver : singletablet c d MFat MMuscle 100 60 75 45 50 30 25 15 0 0 0 30 60 90 120 150 180 Time (Minute) 210 240 270 0 300 30 60 90 120 150 180 Time (Minute) MFat : singletablet MMuscle : singletablet e f MBraintis 2 1.5 mg mg 210 1 0.5 0 0 30 60 90 120 150 180 Time (Minute) 210 240 270 300 MBraintis : singletablet g Figure 5.1. Absorption and distribution of a single dose 39 After diffusing from the gastrointestinal lumen into the gastrointestinal tissue, the drug does not stay here and it is immediately distributed to various organs, its first destination being the liver. The sharp increase in liver PB content during the first 15 minutes confirms this (Figure 5.1c). From Figure 5.1e, we note that about half the amount administered is distributed to muscle tissue. This is expected since muscle tissue constitutes 40 per cent of total body volume and receives approximately 35 per cent of total blood supply. The amount of PB accumulated in fat is also large (Figure 5.1f). Similar to muscle tissue, fat constitutes a large percentage of total body volume. As can be seen from Figure 5.1g, the amount of drug in the target site (i.e. brain tissue) reaches a plateau in 3 hours. Although, it is only a small fraction of the amount administered, its effect is not insignificant. This can be seen in the following figure. ClCur 4 Dmnl 3 2 1 0 0 30 60 90 120 150 180 Time (Minute) 210 240 270 300 ClCur : singletablet Figure 5.2. Increasing chloride current in the brain after a single dose It can be seen that chloride current (i.e. inhibitory neurotransmission) has more than doubled. As expected, no enzyme induction or neuroadaptation took place in such a short time. Enzyme amount stays at the undrugged level (Figure 5.3a). The number of downregulated receptors is an insignificant fraction of the total receptor population of 60 billion (Figure 5.3b). 40 NoDownregRecep 4e-007 1.75 3e-007 Billions Dmnl EnzymeFactor 2 1.5 2e-007 1e-007 1.25 0 1 0 30 60 90 120 150 180 Time (Minute) 210 240 270 300 0 30 60 90 120 150 180 Time (Minute) 210 240 270 300 NoDownregRecep : singletablet EnzymeFactor : singletablet a b Figure 5.3. Dynamics of enzyme induction and neuroadaptation for a single dose 5.1.2. Continuous Drug Intake with Constant Dose In this section, we give the results of simulation experiments in which we assume a regular user of PB. We use the terms “drug use”, “drug intake”, and “drug treatment” interchangeably in the following sections. Recall that our dynamic hypothesis defends that the user would be urged to increase the doses as tolerance develops to the effects of the drug. In this section, however, we assume that the user is not urged and takes constant doses of 30 mg after the loading dose. We therefore show the failure of constant doses to maintain a constant level of sedation. We comparatively study two scenarios to show different levels of tolerance and dependence development: Intake for seven days and intake for 20 days. The results are as follows. 41 Dose 40 mg 35 30 25 20 0 7200 14400 Time (Minute) Dose : d7nofeedback 21600 28800 Dose : d20nofeedback a MBraintis 2 mg 1.5 1 0.5 0 0 18000 36000 Time (Minute) 54000 72000 MBraintis : d7nofeedback MBraintis : d20nofeedback b Figure 5.4. Constant doses (a) and drug profiles in the brain (b) in both a seven day and a 20 day treatment. 42 NoDownregRecep 6 1.7 4.5 Billions Dmnl EnzymeFactor 2 1.4 1.1 3 1.5 0.8 0 0 18000 36000 Time (Minute) 54000 72000 0 EnzymeFactor : d7nofeedback EnzymeFactor : d20nofeedback 18000 36000 Time (Minute) 54000 72000 NoDownregRecep : d7nofeedback NoDownregRecep : d20nofeedback a b ClCur 4 Dmnl 3 2 1 0 0 18000 36000 Time (Minute) 54000 72000 ClCur : d7nofeedback ClCur : d20nofeedback c Figure 5.5. Enzyme induction and neuroadaptation and the resulting chloride current profile when the user takes constant doses (for seven and 20 days). The constant 30 mg doses can be seen in Figure 5.4a. Although the extent of enzyme induction is the same in seven days and 20 days (Figure 5.5a), neuroadaptation progresses much slower (Figure 5.5b). As a result of enhanced metabolism, the amount of PB in the brain decreases constantly (Figure 5.4b). Although in the 20 day treatment the amount in the brain approaches a steady state, chloride current continues to fall as can be seen in Figure 5.5c. These results demonstrate that to maintain sedation, the doses must be increased. Starting from the following section, we incorporate this feedback loop into our analyses. 5.1.3. Continuous Drug Intake with Dose Increase as a Result of Feedback We consider the following drug treatment durations all of which end with abrupt withdrawal: 7 days, 20 days, and 60 days. In all drug treatments, the user is assumed to 43 start with 30 mg tablets after the loading dose of 180 mg. As time elapses, the user would increase the dose in constant increments to compensate the reduced effectiveness. To model the daily drug administration process, we use a pulse function. The inflow named Intake1 of the stock MGIlumen is given in Equation 5.1. Intake1 = (Dose/TIME STEP)*PULSE TRAIN( 1440, TIME STEP, 1440, DaysTreatment*1440+TIME STEP) (5.1) Dose is a stock variable previously explained in detail in Section 4.3, and DaysTreatment is simply the number of days of PB administration. The first term in parentheses is the pulse amplitude. The function PULSE TRAIN is a built-in function in Vensim whose arguments are start time of pulse, pulse duration, pulse repeat time, and final time of pulse, respectively. 5.1.3.1. Drug Treatment for Seven Days To see the situation after withdrawal as well, we set the final time to 27 days (38,880 minutes), and DaysTreatment to 6. Recall that at time zero, the stock MGIlumen contains the loading dose. The tablets are administered starting from the second day (i.e. Time=1440) and for four days. The sum is seven days of drug treatment. We obtain the following dynamics for the key variables. We first present the dose profile (excluding the loading dose) and the profile for the amount of drug in the brain. 44 Dose 80 mg 65 50 35 20 0 2520 5040 Time (Minute) 7560 10080 29160 38880 Dose : d7 a MBraintis 2 mg 1.5 1 0.5 0 0 9720 19440 Time (Minute) MBraintis : d7 b Figure 5.6. Dose profile (a) and drug amount in the brain (b) in the seven day drug treatment followed by abrupt withdrawal 45 NoDownregRecep 4 1.7 3 Billions Dmnl EnzymeFactor 2 1.4 1.1 2 1 0.8 0 0 9720 19440 Time (Minute) 29160 38880 0 EnzymeFactor : d7 9720 19440 Time (Minute) 29160 38880 NoDownregRecep : d7 a b Intensity of withdrawal signs 0.1 Dmnl 0.075 0.05 0.025 0 8640 16200 23760 Time (Minute) 31320 38880 WithdSignIntensity : d7 c Figure 5.7. Enzyme and neuroadaptation dynamics in the seven day drug treatment followed by abrupt withdrawal Looking at figure 5.6a we see that the user increases the third dose. This is because chloride current drops below the threshold as can be seen in Figure 5.8. By doubling the dose, the user avoids a decrease below the threshold. 46 ClCur 4 Dmnl 3 2 1 0 0 7220 14440 Time (Minute) ClCur : d7 ClCurWOPB : d7 21660 28880 ThresholdSedat : d7 Figure 5.8. Behavior of chloride current in the seven day drug treatment In Figure 5.7a, it is interesting to note that although drug treatment stops on the seventh day (8640 minutes), enzyme induction continues its progress until around the ninth day (12,000 minutes). Furthermore, there is an onset of enzyme induction. This inertia is due to genetic processes related to enhanced synthesis of enzymes such as gene transcription, mRNA synthesis, etc. which take time. Nevertheless during drug treatment, EnzymeFactor approaches 2, implying that induction is almost complete (Recall that at maximal induction, rate of metabolism doubles). The inertia in neuroadaptation is more significant. Observe from Figure 5.7b that although drug intake stops, similar to enzyme induction, down-regulation continues its progress six more days (i.e. the curve peaks around the 13th day). However, only a very small fraction of total receptor population is down-regulated implying that dependence has not yet developed. We therefore assume that the peak intensity in Figure 5.7c is insignificant and thus establish a reference. Hereafter, we regard any peak intensity below 0.025 as insignificant. The reports in literature stating that dependence to barbiturates develops in several weeks also support the validity of our assumption. 47 5.1.3.2. Drug Treatment for 20 Days Following are the results for 20 days of 30 mg one-a-day doses ending with abrupt withdrawal. Dose 200 mg 150 100 50 0 0 7200 14400 Time (Minute) 21600 28800 54000 72000 Dose : d20 a MBraintis 4 mg 3 2 1 0 0 18000 36000 Time (Minute) MBraintis : d20 b Figure 5.9. Dose profile (a) and drug amount in the brain (b) in the 20 day drug treatment followed by abrupt withdrawal 48 NoDownregRecep 20 3 15 Billions Dmnl EnzymeFactor 4 2 1 10 5 0 0 0 18000 36000 Time (Minute) 54000 72000 0 EnzymeFactor : d20 18000 36000 Time (Minute) 54000 72000 NoDownregRecep : d20 a b Intensity of withdrawal signs 0.3 Dmnl 0.225 0.15 0.075 0 27360 38520 49680 Time (Minute) 60840 72000 WithdSignIntensity : d20 c Figure 5.10. Enzyme and neuroadaptation dynamics in the 20 day drug treatment followed by abrupt withdrawal The inertia in enzyme and neuroadaptation dynamics is again evident. The onset of enzyme induction is shorter than that of receptor down-regulation. Figure 5.10a shows that in a few days, enzyme induction peaks and although intake stops on the 20th day, fast metabolism persists six more days (until the 36,000th minute). As can be seen in Figure 5.10b, more than a quarter of the receptor population is down-regulated. This weakens inhibitory neurotransmission by decreasing normal chloride current (See the variable named NormClCure in Section 4.2.2). Together with fast metabolism this reduces the effectiveness of the drug, urging the user to increase the dose several times (Figure 5.9a). The decrease in drug effectiveness is so severe that the final dose is five times the initial dose. Looking at Figure 5.11 below, we conclude that the dose increase decisions are justified since chloride current is maintained above the threshold with a few insignificant undershoots throughout 20 days. 49 Chloride current 4 Dmnl 3 2 1 0 0 18000 ClCur : d20 ThresholdSedat : d20 36000 Time (Minute) 54000 72000 ClCurWOPB : d20 Figure 5.11. Behavior of chloride current in the 20 day drug treatment The peak intensity of withdrawal signs is around 0.1 as can be seen in Figure 5.10c. As mentioned in Section 1.5, clinical research suggests that dependence to barbiturates develops in a few weeks. This suggestion and our outputs showing that more than a quarter of total receptors are desensitized in 20 days lead to the conclusion that the user has become dependent-at least partially-to PB and thus upon abrupt discontinuation, he/she would experience a significant withdrawal syndrome. In Figure 5.10c, the peak intensity of withdrawal signs is around 0.1. Accordingly, hereafter we shall regard any withdrawal sign intensity above 0.1 as severe. We now have two reference points to help us assess the significance of withdrawal signs in further simulation experiments. Finally, the delay in the outburst of the withdrawal syndrome is due to the long half-life (despite enhanced metabolism) of PB. 5.1.3.3. Drug Treatment for 60 Days We set the final time to 90 days (129,600 minutes) and DaysTreatment to 59 days. We obtain the following results. 50 Dose 200 mg 150 100 50 0 0 21600 43200 Time (Minute) 64800 86400 97200 129600 Dose : d60 a MBraintis 6 mg 4.5 3 1.5 0 0 32400 64800 Time (Minute) MBraintis : d60 b Figure 5.12. Dose profile (a) and drug amount in the brain (b) in the 60 day drug treatment followed by abrupt withdrawal 51 NoDownregRecep 60 3 45 Billions Dmnl EnzymeFactor 4 2 1 30 15 0 0 0 32400 64800 Time (Minute) 97200 129600 0 EnzymeFactor : d60 32400 64800 Time (Minute) 97200 129600 NoDownregRecep : d60 a b Intensity of withdrawal signs 0.3 Dmnl 0.225 0.15 0.075 0 84960 96120 107280 Time (Minute) 118440 129600 WithdSignIntensity : d60 c Figure 5.13. Enzyme and neuroadaptation dynamics in the 60 day drug treatment followed by abrupt withdrawal Elevated enzyme levels persist even after drug administration stops as was the case in the shorter treatment durations studied previously. We see from Figure 5.13b that in 60 days, practically all receptor population is desensitized implying that the user has been rendered completely dependent. Around the 45th day, desensitization saturates. Tolerance, on the other hand, is almost complete after the user increases the dose to 150 mg at the 19th dose. Further dose increase is a month later (45th day). Abrupt withdrawal causes a severe withdrawal syndrome as can be verified from Figure 5.13c. The peak intensity is nearly twice our reference of significance. In Figure 5.14 below, we present the chloride current profile. The elevated dosages are efficient in maintaining the desired sedation level. 52 Chloride current 4 Dmnl 3 2 1 0 0 32400 ClCur : d60 ClCurWOPB : d60 64800 Time (Minute) 97200 129600 ThresholdSedat : d60 Figure 5.14. Behavior of chloride current in the 60 day drug treatment 5.2. Model Validity Discussion A point-by-point match with real data is not a major goal, and it is not realistic in SD models (Forrester, 1961; Barlas, 1996; Sterman 2000). The crucial idea is to capture the behavior patterns. We thus draw our comparisons according to this approach. The validity of the pharmacokinetic sector is established since the same structures were used in a previous study and a good fit with real data has been shown (El-Masri and Portier, 1998). Urinary excretion was also included in this model and it is assumed to follow first-order kinetics. In the literature, it is reported that 24 per cent of administered PB is excreted unchanged (Engasser et al., 1981). It was a straightforward issue to calibrate the rate constant using this information. Regarding enzyme induction, it is reported in the literature that the rate of metabolism doubles at maximal induction and this peak occurs in days to weeks. Parameter calibrations were done using this information. We repeat the enzyme induction related model outputs for the 20 day treatment case in Figure 5.15. As can be seen, metabolism 53 doubles in approximately 2 weeks (18,000 minutes). This is in good agreement with literature (Hardman and Limbird, 2001). EnzymeFactor 4 Dmnl 3 2 1 0 0 18000 36000 Time (Minute) 54000 72000 EnzymeFactor : d20 Figure 5.15. Progression of enzyme induction in 20 days of continuous PB use Since no quantitative human data regarding tolerance and dependence development are available in the literature, we use data from studies on animal models such as the one by Gay et al. (1983). Our assumptions are fairly similar to theirs. Similar to our model, they target a constant sedation level in rats while adjusting doses. They administer PB orally to rats for 35 days and observe that tolerance development is complete after the first ten days. To compare, in Figure 5.16 we display their daily dosing history together with our model outputs for the 60 day drug treatment case. Dose 200 mg 150 100 50 0 0 21600 43200 Time (Minute) 64800 86400 Dose : d60 a b Figure 5.16. Comparison of tolerance dynamics generated by the model (a) against real data (b) from Gay et al (1983). 54 Similar to the findings by Gay et al, our drug user increases the doses most aggressively in the first few weeks in order to reach a constant (desired) sedation level. Afterwards, relatively constant doses are enough to sustain the constant sedation level. To confirm this constant level of sedation, refer to Figure 5.14 (See the first 60 days). A good pattern match is thus observed between the model’s dose output and experimental dose data as seen in Figure 5.16a and b, so as to sustain a desired sedation level. Gay et al. also monitor rats for withdrawal signs after abrupt discontinuation to the drug. They quantify the intensity of withdrawal signs which occur a few days after discontinuation. They also observe that the signs attenuate as time elapses. Although their proxy for the intensity of withdrawal signs is different from ours (i.e. their proxy is behavioral outcomes like ear twitches; ours is chloride currents), they indicate the same: the more intense the behavioral sign (the lower the chloride current), the more severe is the withdrawal syndrome. We compare our results in Figure 5.17 below. As can be seen, a sharp-boom-then-decay behavior is well captured by our model. Intensity of withdrawal signs 0.3 Dmnl 0.225 0.15 0.075 0 84960 96120 107280 Time (Minute) 118440 129600 WithdSignIntensity : d60 a b Figure 5.17. Tolerance and dependence indicators for 60 days of continuous PB intake, (a) Model output, (b) Real data. Gay et al. also observe that although tolerance development is almost complete in the ten day group, the rats withdrawn from PB after 35 days of continuous administration experience more intense withdrawal signs. Our 20 day case is analogous to their ten day group. In Figure 5.18 below, we show that the difference in withdrawal signs of the 20 day drug user and the 60 day user is captured by our model. 55 Intensity of withdrawal signs 0.3 0.225 0.225 Dmnl Dmnl Intensity of withdrawal signs 0.3 0.15 0.075 0 27360 0.15 0.075 38520 49680 Time (Minute) WithdSignIntensity : d20 60840 72000 0 84960 96120 107280 Time (Minute) 118440 129600 WithdSignIntensity : d60 a b Figure 5.18. Differences in withdrawal dynamics between a partially dependent (20 day user) and a completely dependent (60 day user). 56 6. SCENARIO ANALYSES 6.1. Epilepsy Patient In epilepsy, the essential balance between excitatory and inhibitory neurotransmission is disrupted. There are various types of epilepsy. The major classification regards the extent to which the brain is affected. According to this classification, a seizure may either be general or partial, the former meaning that all parts of the brain are affected while the latter means that the disease starts in one lobe of the brain only. The causes of epilepsy are difficult to dissect. It may be due to anything that damages the brain such as an infection involving the brain or a head injury. It may even be due to incomplete maturation of the brain (Chappell and Crawford, 2001). As mentioned previously, this thesis focuses on inhibitory neurotransmission mediated by the neurotransmitter GABA. Therefore, in this scenario we consider a form of epilepsy called juvenile myoclonic epilepsy that is caused by decreased inhibitory neurotransmission as a result of mutations in GABAA receptors (Kapur, 2003). As the name implies, this type of seizure is experienced by young people (in juvenile time of their life) and it is characterized by sudden, jerky or shock-like contractions usually in arms and legs (Chappell and Crawford, 2001). All epileptic seizures are tried to be controlled by anti-epileptic drugs. These drugs do not cure epilepsy (From http://www.epilepsy.org). There are numerous antiepileptic drugs and several different prescriptions depending on the type of seizure. Sometimes, it is not possible to control a form of seizure with a single drug in which case multi-drug treatment is employed (Chappell and Crawford, 2001). We assume a form of juvenile myoclonic epilepsy that can be controlled with phenobarbital (PB). Recall that our proxy for inhibitory neurotransmission was the chloride current in the brain and the undrugged state was defined by a constant named ClCurWOPB and we assumed that its value was 1 in a healthy individual (See Section 4.2.2). Since an epilepsy 57 patient is suffering from decreased inhibitory neurotransmission, we have to set this to a lower value. We may assume that a myoclonic seizure is comparable to a state of rebound hyperexcitability. Therefore in assigning the value of ClCurWOPB for an epilepsy patient, we regard our previous assumption that in severe rebound hyperexcitability, value of the model variable named WithdSignIntensity was at least 0.1. Since WithdSignIntensity = -(ClCur-1) (6.1) ClCur = 1 – WithdSignIntensity (6.2) it follows that Therefore, in a state of rebound excitability Cl current < 1 – 0.1 = 0.9 (6.3) As a result, in our epilepsy patient, we assume that Cl current without PB is equal to 0.9. This is an average value. In reality, we expect that between seizures, this value is close to 1. However, being more realistic and assuming a time dependent profile to this variable would not contibute to the quality of our analysis. In order to make sound comparisons, we assume that the epilepsy patient not only wants to control seizures, but also desires sedation similar to the healthy person we have studied in Section 5. That is, ThresholdSedat is assumed to be the same for the epilepsy patient. We assume the same daily doses of 30 mg. Epilepsy patients sometimes continue drug treatment for a life time. Usually, withdrawal is not an issue for epilepsy patients. Therefore, we do not study postwithdrawal dynamics in this scenario. Instead, we comparatively present the dynamics in the first 60 days of PB use by a healthy person and an epilepsy patient to provide insights on the differences in development of tolerance and dependence in a disease state. 58 Dose 400 mg 300 200 100 0 0 21600 43200 Time (Minute) Dose : d60 64800 86400 Dose : d60epileptic a MBraintis 6 mg 4.5 3 1.5 0 0 21240 42480 Time (Minute) 63720 84960 MBraintis : d60 MBraintis : d60epileptic b Figure 6.1. Dose profiles (a) and drug profiles in brain tissue (b) of both a healthy and an epileptic individual in 20 days of continuous PB use. 59 NoDownregRecep 60 3 45 Billions Dmnl EnzymeFactor 4 2 1 30 15 0 0 0 21240 42480 Time (Minute) EnzymeFactor : d60 EnzymeFactor : d60epileptic 63720 84960 0 21240 42480 Time (Minute) 63720 84960 NoDownregRecep : d60 NoDownregRecep : d60epileptic Figure 6.2. Enzyme and neuroadaptation dynamics in both a healthy and an epileptic individual taking PB for the last 60 days. Chloride current 4 Dmnl 3 2 1 0 0 21240 ClCur : d60 ClCur : d60epileptic ClCurWOPB : d60 42480 Time (Minute) 63720 84960 ClCurWOPB : d60epileptic ThresholdSedat : d60 Figure 6.3. Chloride current in a healthy and an epileptic individual (Undrugged levels of chloride current are also given) Since the undrugged level of chloride current in an epilepsy patient is lower as can be seen in Figure 6.3 (gray colored line), the discrepancy between the desired level of chloride current and normal chloride current is larger and thus more drug is necessary to sustain sedation. This can be verified by looking at Figure 6.1. The amount of drug administered and thus the amount of drug in the brain is higher in the epileptic individual. 60 Enzyme induction profile is not different in an epilepsy patient as can be seen in Figure 6.2a. This is because the PB concentration in the liver is at saturation in both cases and thus enzyme synthesis accelerates at maximum rate. In the CNS, the rate of neuroadaptation is alarmingly higher in the epileptic individual due to elevated doses. In about 35 days, the number of downregulated receptors approaches the total receptor population of 60 billion (Figure 6.2b). This shows that an epilepsy patient becomes dependent earlier than a healthy person. When compared to the epilepsy patient, in 35 days, slightly more than half the receptor population is downregulated in a healthy person. As mentioned previously, dependence is not a major concern in an epilepsy since the patient is not expected to discontinue the drug. However, this scenario shows the increased susceptibility of an epilepsy patient to rebound effects had he/she discontinued the drug contrary to a doctor’s advice. 6.2. Co-administration of a Drug That Causes Enzyme Inhibition In this scenario, we study a possible drug-drug interaction. Most drug-drug interactions are due to the effects of drugs on liver enzymes (i.e. CYP enzymes). The CYP enzymes are either inhibited or induced by drugs leading to altered metabolism of the substrates of (chemicals that are metabolised by) these enzymes. Usually, the drugs themselves are also substrates of these enzymes and thus pharmacokinetics of a drug may vary considerably if administered together with another drug. To illustrate, suppose that drug A is taken together with drug B which is an inhibitor of a CYP enzyme. Suppose also that drug A is a substrate of this CYP enzyme. This would lead to a slower metabolism of drug A and a normal dose of drug A might actually be fatal. Therefore, in multi-drug treatment, levels of drugs must be carefully monitored to avoid unwanted results. There may be infinitely many forms of drug-drug interactions. In this scenario, we assume that our hypothetical person has been taking fluconazole, an anti-fungal drug, 61 before starting PB treatment. Fluconazole has been shown to be an inhibitor of PB metabolizing enzymes (Venkatakrishnan, 2000). To investigate the extent of enzyme inhibition by fluconazole, Kumar et al. (2008) use flurbiprofen as a substrate of the inhibited enzyme. They study three groups of subjects. To the first group, they administer flurbiprofen only. To the second group, they administer flurbiprofen after pre-treatment with 200 mg fluconazole for 7 days. Finally to the third group, they administer flurbiprofen after pre-treatment with 400 mg of fluconazole for 7 days. They monitor flurbiprofen clearance in all groups. Their averages are plotted in Figure 6.4. Figure 6.4. Flurbiprofen average clearance as influenced by fluconazole pre-treatment. Values are given as median + 25th percentile (Kumar et al., 2008). Observe from the figure that a seven day pre-treatment with 200 mg fluconazole halves the rate of metabolism of flurbiprofen. Clearing rate drops from 1.6 L/hr to 0.8 L/hr. Although there is no comprehensive clinical study on PB-fluconazole interaction, it is reported in the literature that when co-administered with PB, fluconazole leads to increased PB levels via inhibition of enzymes similar to the flurbiprofen case. Since both 62 PB and flurbiprofen are substrates of the same enyzme, we may argue that extent of inhibition will be similar for both drugs. Here we assume that before starting PB treatment, the user has been taking 200 mg doses of fluconazole for the past seven days. Therefore, by the start of treatment, metabolism rate of PB is assumed to be half the normal rate (i.e. initially the model variable EnzymeFactor is equal to 0.5). However, enzyme induction still occurs and in a few days the metabolism rate is doubled (i.e. EnzymeFactor becomes approximately 1). We assume that fluconazole has no effect on any other part of the system. Assuming that PB treatment duration is 20 days, we get the following results. Dose 200 mg 150 100 50 0 0 7200 14400 Time (Minute) Dose : d20fluc 21600 28800 54000 72000 Dose : d20 a MBraintis 4 mg 3 2 1 0 0 18000 36000 Time (Minute) MBraintis : d20fluc MBraintis : d20 b Figure 6.5. Dose profiles (a) and drug amounts in the brain (b) with and without fluconazole pre-treatment. PB is taken for 20 days and is discontinued abruptly 63 NoDownregRecep 40 3 30 Billions Dmnl EnzymeFactor 4 2 1 20 10 0 0 0 18000 36000 Time (Minute) 54000 72000 0 EnzymeFactor : d20fluc EnzymeFactor : d20 18000 36000 Time (Minute) 54000 72000 NoDownregRecep : d20fluc NoDownregRecep : d20 a b Intensity of withdrawal signs 0.3 Dmnl 0.225 0.15 0.075 0 27360 38520 49680 Time (Minute) 60840 72000 WithdSignIntensity : d20fluc WithdSignIntensity : d20 c Figure 6.6. Enzyme and neuroadaptation dynamics with and without fluconazole pretreatment. PB treatment duration is 60 days ending with abrupt discontinuation. Figure 6.5a shows that inhibition of metabolism has slowed down the progression of tolerance. Since PB is cleared much slower, a milder increase in dose is enough to yield the same level of sedation. Compared to a total PB dose of 2040 mg, pre-treatment with fluconazole necessitates only 1320 mg. In Figure 6.5b, we see that the PB amount in the brain is not increased significantly and thus there is no toxicity concern. Neuroadaptation and withdrawal dynamics are rather interesting. Observing Figure 6.6b, we see that neuroadaptation has progressed more severely after fluconazole pretreatment. Interestingly, upon withdrawal, the intensity of rebound effects is much lower. This can be explained as follows: Although the number of down-regulated receptors is larger when fluconazole is administered prior to PB, phenobarbital is cleared slower and thus there is more time for re-adaptive mechanisms to restore brain physiology. This can 64 be verified by observing Figure 6.6c and noting that the withdrawal syndrome is not only lighter, but also the outburst of the syndrome is later in the fluconazole pre-treatment case. 6.3. Different Dosing Frequencies In all preceding simulation runs, we assume the drug user employs one-a-day dosing. One may reasonably suspect that in terms of tolerance and dependence, employing different dosing schemes could yield different results. Therefore in this section we compare four different dosing schemes in which we vary the initial doses and dosing frequencies. We experiment with four dosing schemes: One tablet every two days, one tablet every day (our basic assumption), two tablets every day, and finally three tablets every day. The initial dose and dose increment are the same for each case and these are 60, 30, 15, and 10 mg for tablets taken one-every-two, one-a-day, two-a-day, and three-a-day, respectively. Had the doses not increased, the average daily doses would be the same in all dosing schemes. We comparatively show tolerance and dependence dynamics together with the behavior of chloride current. Treatment duration is assumed to be 20 days followed by abrupt discontinuation. As in Section 5, we first disengage feedback and assume constant doses. As shown in Figure 6.7a, without the feedback, the behavior of EnzymeFactor does not change significantly as a function of dosing scheme. On the other hand, the peak number of downregulated receptors is the lowest in one-every-two-days scheme (Figure 6.7b). 65 EnzymeFactor 2 Dmnl 1.7 1.4 1.1 0.8 0 18000 36000 Time (Minute) 54000 72000 54000 72000 EnzymeFactor : d20oneeverytwo-nofeedb EnzymeFactor : d20oneaday-nofeedb EnzymeFactor : d20twoaday-nofeedb EnzymeFactor : d20threeaday-nofeedb a NoDownregRecep 6 Billions 4.5 3 1.5 0 0 18000 36000 Time (Minute) NoDownregRecep : d20oneeverytwo-nofeedb NoDownregRecep : d20oneaday-nofeedb NoDownregRecep : d20twoaday-nofeedb NoDownregRecep : d20threeaday-nofeedb b Figure 6.7. Enzyme and neuroadaptation dynamics in different dosing schemes (No feedback to increase the doses) Observing the comparative behavior of chloride current given in Figure 6.8, we see that in one-every-two-days scheme, the average chloride current in the first few days of 66 treatment (when the chloride current is relatively high) is lower in comparison to the other schemes. The rate of neuroadaptation is more sensitive to chloride current at these levels as can be verified from our effect formulation given in Figure 4.5. Therefore, the overall rate of neuroadaptation in this scheme is lower in comparison to the other three schemes. This results in a lower peak in the number of down-regulated receptors. ClCur 4 Dmnl 3 2 1 0 0 18000 ClCur : d20oneeverytwo-nofeedb ClCur : d20oneaday-nofeedb 36000 Time (Minute) 54000 72000 ClCur : d20twoaday-nofeedb ClCur : d20threeaday-nofeedb Figure 6.8. Comparative behavior of chloride current (No feedback to increase the doses) When the feedback loop is operational, the picture changes drastically. Similar to our base case, we assume that the feedback process is operational once the first dose is taken (the dose after the loading dose). In Figure 6.9, we give the dynamics of tolerance development in all dosing schemes. 67 Dose 400 mg 300 200 100 0 0 7200 14400 Time (Minute) 21600 28800 Dose : d20oneeverytwo Dose : d20oneaday Dose : d20twoaday Dose : d20threeaday Figure 6.9. Difference in the extent of tolerance development w.r.t dosing schemes (Feedback allowed to increase doses) NoDownregRecep 40 Billions 30 20 10 0 0 18000 36000 Time (Minute) 54000 72000 NoDownregRecep : d20oneeverytwo NoDownregRecep : d20oneaday NoDownregRecep : d20twoaday NoDownregRecep : d20threeaday Figure 6.10. Neuroadaptation dynamics for different dosing schemes (Feedback allowed to increase doses) 68 Intensity of withdrawal signs 0.3 Dmnl 0.225 0.15 0.075 0 0 18000 36000 Time (Minute) 54000 72000 WithdSignIntensity : d20oneeverytwo WithdSignIntensity : d20oneaday WithdSignIntensity : d20twoaday WithdSignIntensity : d20threeaday Figure 6.11. Dependence dynamics for different dosing schemes (Feedback allowed to increase doses) It turns out that both tolerance and dependence development is less when frequency of doses is increased. In three-a-day dosing, the total amount of drug administered in 20 days is 1690 mg. As the frequency is decreased, this total amount increases. In the extreme case where the user takes one tablet every two days, the total amount administered is 2460 mg which is approximately 50 per cent more than the three-a-day case. Additionally, in one-every-two-days dosing, the peak number of down-regulated receptors is 23 billion whereas in three-a-day dosing, the peak is 13 billion (Figure 6.10). As anticipated from this, the severity of rebound effects is most potent in one-every-two-days dosing (Figure 6.11). Finally, the amplitude of oscillations in chloride current is less in frequent dosing. This outcome is in favor of homeostasis: The body prefers stability. The comparative behavior of chloride current is given in Figure 6.12 below. In one-every-two-days scheme, the enormous overshoots of chloride current increase the rate of neuroadaptation: Chloride current sometimes exceeds 3.5 (Recall that the threshold is 2.5). The extent of neuroadaptation is proxied by the area between the chloride current and the sedation threshold. 69 Chloride current 4 Dmnl 3 2 1 0 0 18000 ClCur : d20oneeverytwo ClCur : d20oneaday ClCur : d20twoaday 36000 Time (Minute) 54000 72000 ClCur : d20threeaday ClCurWOPB : d20oneaday ThresholdSedat : d20oneaday Figure 6.12. Behavior of chloride current in different dosing schemes (Feedback allowed to increase doses) We may confidently argue that the most appropriate dosing scheme is three-a-day dosing. Although further increases in dosing frequency could prove better, such high frequencies would not be practical since the user would have to remember too often taking a tablet. 70 7. ANALYSIS OF WITHDRAWAL POLICIES It is shown in the preceding sections that abrupt withdrawal results in an unwanted withdrawal syndrome. This suggests that the dose should be reduced gradually. During the withdrawal period, the decision variables are dosing times, dosing amounts and duration of the withdrawal regimen. The best policy would be the one that causes very few or no withdrawal signs with a minimum total amount of administered PB. In this section, we demonstrate both unsuccessful and successful withdrawal dosing regimens after one-a-day dosing for both 20 and 60 days. We assume a healthy user taking PB one-a-day for sedation as in Section 5. It is anticipated that withdrawal would be easier after the 20 day treatment since the drug user would not be totally dependent on the drug as was shown in section 5.1.2.2. On the other hand, we have shown in section 5.1.2.3 that the user reaches a maximum tolerance and dependence level in the 60 day treatment and this would complicate withdrawal. In hypothesizing effective withdrawal regimens, we use intuition and therefore start with relatively good regimens. Simulation experiments were conducted as follows: We start with an initial guess and we check, at the end of the regimen, whether the user experiences rebound effects. If this is the case, we prolong the regimen and/or modify the doses until we observe no withdrawal syndrome. In summary, by improving upon our previous postulations, we try to come up with regimens that help avoid a withdrawal syndrome. For each case of drug treatment duration, we present first an unsuccessful postulation. Then we discuss necessary modifications that lead to a successful regimen. 7.1. Withdrawal after 20 days of treatment 7.1.1. An unsuccessful regimen Since the half-life of PB is long, when drug intake is stopped on the 20th day, the drug stays in the body and is still effective. Trials show that the chloride current stays above the base value (i.e. 1) for at least seven days after the last dose. Thus, we wait for 71 seven days before starting the withdrawal regimen. This regimen lasts for ten days: After taking no tablet in the first seven days, the user is supposed to take one-tenth of the last dose (dose on the 20th day) for the following three days and then discontinue. The dynamics that result are given in Figures 7.1 and 7.2. EnzymeFactor 4 150 3 Dmnl mg MGIlumen 200 100 50 2 1 0 0 0 18000 36000 Time (Minute) 54000 72000 0 MGIlumen : d20w10-2 18000 54000 72000 54000 72000 EnzymeFactor : d20w10-2 a b NoDownregRecep ClCur 20 4 15 3 Dmnl Billions 36000 Time (Minute) 10 5 2 1 0 0 0 18000 36000 Time (Minute) NoDownregRecep : d20w10-2 54000 72000 0 18000 36000 Time (Minute) ClCur : d20w10-2 c d Figure 7.1. Dynamics of an unsuccessful withdrawal regimen after partial dependence 72 Intensity of withdrawal signs 0.3 Dmnl 0.225 0.15 0.075 0 41760 49320 56880 Time (Minute) 64440 72000 WithdSignIntensity : d20w10-2 Figure 7.2. Severity of withdrawal signs after an unsuccessful dosing strategy in partial dependence Looking at Figure 7.1b, we see that although enzyme levels are being restored during withdrawal doses, the duration of the withdrawal regimen falls short complete this restoration. EnzymeFactor is more than 1.5 at the time of complete withdrawal. Figure 7.1c shows that down-regulated receptors merely stop increasing and re-adaptive mechanisms are not operational at all. The result is a severe withdrawal syndrome as can be seen in Figure 7.1d (chloride current undershoots 1) and more clearly in Figure 7.2. 7.1.2. A successful regimen The failure of the ten day withdrawal period suggests a longer withdrawal period with decreased dosages. After trial-and-error, we come up with the following 15 day regimen: We administer one-fiftheenth of the final dose between days 27 and 31; and we administer one-twentieth of the final dose between days 32 and 35. The following dynamics result. 73 EnzymeFactor 4 150 3 Dmnl mg MGIlumen 200 100 50 2 1 0 0 0 21600 43200 Time (Minute) 64800 86400 0 MGIlumen : d20w15-4 21600 43200 Time (Minute) 86400 64800 86400 EnzymeFactor : d20w15-4 a b NoDownregRecep ClCur 4 15 3 Dmnl 20 10 5 2 1 0 0 0 21600 43200 Time (Minute) 64800 86400 NoDownregRecep : d20w15-4 0 21600 43200 Time (Minute) ClCur : d20w15-4 c d Figure 7.3. Dynamics in a successful withdrawal regimen after partial dependence WithdSignIntensity 0.1 0.075 Dmnl Billions 64800 0.05 0.025 0 28800 43200 57600 Time (Minute) 72000 86400 WithdSignIntensity : d20w15-4 Figure 7.4. Severity of withdrawal signs after a successful dosing strategy in partial dependence 74 Our anticipation turned out correct. The duration of withdrawal is now long enough so as to facilitate complete recovery of down-regulated receptors (Figure 7.3c). Although the metabolism is still 50 per cent higher than normal (Figure 7.3b, around the 35th day), complete withdrawal does not lead to a significant withdrawal syndrome as can be seen from Figure 7.4. Observe that the peak intensity of withdrawal signs is well below the 0.025 reference. This suggests that the contribution of enzyme induction to development of dependence is minor. In fact, this is reported in the literature as well. This result is thus an additional clue of our model’s validity. 7.2. Withdrawal after 60 days of treatment 7.2.1. An unsuccessful regimen We now experiment with withdrawal regimens after 60 days of continuous PB use after which the user becomes completely dependent on the drug. As a first trial, we propose a 20 day regimen as follows: We wait seven days before administering reduced doses and after that, between days 67 and 80, we administer one-fifteenth of the final dose. The following dynamics result. 75 EnzymeFactor 4 150 3 Dmnl mg MGIlumen 200 100 50 2 1 0 0 0 36000 72000 Time (Minute) 108000 144000 0 MGIlumen : d60w20-1 36000 72000 Time (Minute) 144000 108000 144000 EnzymeFactor : d60w20-1 a b NoDownregRecep ClCur 60 4 45 3 Dmnl Billions 108000 30 15 2 1 0 0 0 36000 72000 Time (Minute) 108000 144000 NoDownregRecep : d60w20-1 0 36000 72000 Time (Minute) ClCur : d60w20-1 c d Figure 7.5. Results for an unsuccessful withdrawal regimen after complete dependence WithdSignIntensity 0.2 Dmnl 0.15 0.1 0.05 0 86400 100800 115200 Time (Minute) 129600 144000 WithdSignIntensity : d60w20-1 Figure 7.6. Severity of withdrawal signs after an unsuccessful dosing strategy in complete dependence Although chloride current is maintained in an appropriate range during the regimen so that both the down-regulated receptors and elevated enzyme levels are decreased 76 (Figures 7.5b and 7.5c), re-adaptation is partial because the duration of withdrawal falls short. The result is a severe withdrawal syndrome as can be seen in Figure 7.6. 7.2.2. A Successful Regimen We prolong the duration of withdrawal to 30 days. Since the dosage in the previous regimen was shown to be appropriate, the regimen in the first 20 days is exactly the same as in 7.2.1. We then assume that the user is supposed to take one-twentieth of the final dose for the following ten days (i.e. between days 81 and 90). The following dynamics are observed. EnzymeFactor 4 150 3 Dmnl mg MGIlumen 200 100 50 2 1 0 0 0 39600 79200 Time (Minute) 118800 158400 0 MGIlumen : d60w30-1 39600 118800 158400 118800 158400 EnzymeFactor : d60w30-1 a b NoDownregRecep ClCur 60 4 45 3 Dmnl Billions 79200 Time (Minute) 30 15 2 1 0 0 0 39600 79200 Time (Minute) NoDownregRecep : d60w30-1 118800 158400 0 39600 79200 Time (Minute) ClCur : d60w30-1 c d Figure 7.7. Results for a gradual withdrawal regimen of 30 days following a 60 day drug treatment 77 Intensity of withdrawal signs 0.2 Dmnl 0.15 0.1 0.05 0 84960 99720 114480 Time (Minute) 129240 144000 WithdSignIntensity : d60w30-1 Figure 7.8. Severity of withdrawal signs after a successful dosing strategy in complete dependence As anticipated, prolonging the last phase of the regimen cured the failure. The drug user experiences no rebound effects (Figure 7.8). The duration of drug intake is long enough so that almost all down-regulated receptors are restored by the end of the 90th day (Figure 7.7c). 78 8. CONCLUSIONS AND FUTURE RESEARCH DIRECTIONS Although being replaced by safer drugs, a lot of people still use phenobarbital (PB) regularly for sedation or against sleep disorders. As a side effect, phenobarbital enhances the synthesis of its own metabolic enzymes in the liver. This enzyme induction problem causes increased tolerance to phenobarbital over time. Moreover, the brain adapts to the presence of the drug and its sensitivity decreases with time (neuroadaptation). The system dynamics model constructed in this thesis is a representation of prolonged barbiturate use, including phenobarbital absorption, distribution, metabolism, and elimination processes with enzyme induction and neuroadaptation related structures. The validity of the model is first demonstrated using available experimental data and other qualitative information. The model is used as an experimental platform to study various scenarios, including an epileptic patient, potential drug interactions and alternative dosing schemes to minimize withdrawal syndromes. Adaptive changes in the body as a response to drug use result in drug tolerance, dependence and eventually withdrawal syndromes. The situation is further complicated in an epilepsy patient. We show that an epilepsy patient is more prone to development of barbiturate tolerance and dependence. Possible drug-drug interactions should also be taken into account if PB is being taken together with other chemicals. In this thesis, we study a drug-drug interaction involving only the liver. However, in epilepsy, several drugs may be prescribed and concurrent intake of these drugs would involve more complex dynamics especially in the central nervous system (CNS). Additionally, the consumption of alcohol while taking barbiturates has well-known synergistic and thus lethal effects. As such, these could be subjects of future study. The model provides an experimental platform to test different dosing schemes and dose adjustment policies in prolonged use. We experiment with different dosing frequencies and show that the more frequent the doses, the better it is in terms of tolerance and dependence development. However, we neglect the possible impracticality of frequent doses. Since we explicitly model drug content in arterial blood, the simulation model could 79 also be used to simulate clinical settings such as constant intravenous infusion where the infusion of drug is more continuous contrary to cases studied here. Parallel to literature reports, we show that when a dependent user abruptly discontinues PB use, harmful rebound effects are experienced. To avoid, the doses should be reduced gradually. We have proposed relatively efficient withdrawal regimens for both partial and complete dependence cases. As anticipated, a longer period of withdrawal is necessary in complete dependence cases. It is shown that he duration of the withdrawal period should be at least half the actual treatment duration in all cases. The method of search for feasible withdrawal regimens was intuitive. A more systematic approach could prove useful in the future. It may be interesting to define the problem as an optimization problem where it is tried to minimize both the duration of the withdrawal period and the amount of doses while keeping the resulting intensity of withdrawal signs at minimum. The model does not take into account neuroadaptation dynamics in the excitatory neurotransmission system. It is likely that when inhibitory neurotransmission is potentiated by PB, besides desensitizing inhibitory receptors to counteract potentiation, neuroadaptation could up-regulate excitatory neurotransmission as well. Including this mechanism in the future versions of the model could enhance its realism. Finally, the model is built using rather generic structures and generic assumptions. This is especially true for the pharmacokinetic sector. The parameters can be modified so that a different CNS-active drug can be modeled as well. Receptor down-regulation is also a rather common mechanism of neuroadaptation. Therefore, the parameters in the CNS sector of the model can be modified to capture the dynamics of a different drug that causes receptor down-regulation. 80 APPENDIX. EQUATIONS OF THE MODEL The complete stock-flow structure of the model is given in Figure A.1. 81 CBraintis QBrain VBraincapil Eff PBReadptn EffPB WithdSignIntensity CBraincapil MBraintis FR BraincapilToBraintis Bplasma ClCur BrainToVenous CFat QFat QTotal VArterial ArterialToVenous ArterialTo Muscle Kexcr ArterialTo Kidney VGItissue ArterialToGItis CMuscle I3 <Time> Intake 5 Intake2 Intake Intake3 4 MLiver CLiver QLiver Metabolism FINAL TIME DaysTreatment VLiver NormKmet kout Rin Intake1 Absorption GItissue ToLiver <Time> KidneyToVenous VKidney MGIlumen MGItissue ArterialToLiver LoadDose HalflifeEnzyme Kabs QGItissue Dose DoseIncr I2 PKidney MKidney CKidney <Time> I4 I1 <Time> MuscleTo Venous VMuscle QKidney PGItissue RealAdptnRate ThresholdSedat MVenous VenousToArterial Excretion CGItissue IndAdptnRate FatToVenous VVenous CVenous QMuscle PMuscle MMuscle CArterial NoDownreg Recep PFat QHeart MArterial Adaptation ReadptnFrac VFat MFat ArterialToFat EffSatur ClCurWOPB MBraincapil ArterialToBrain TotalNoRecep NormClCur VBraintis BraintisToBraincapil DR EffAdptn OnNorm ClCur <TIME STEP> Synthesis LiverTo Venous Enzyme Factor PLiver ReaIInducbyPB IndInducByPB Kmet Figure A.1. Complete stock-flow diagram of the model Degradation Readaptation 82 Equations of the model are given below for one-a-day treatment lasting for 20 days and ending with abrupt withdrawal. Dose=INTEG (DoseIncr,30) I4=IF THEN ELSE(MODULO(Time,1440)>=1437,1,0) LoadDose=180 MGIlumen=INTEG (Intake2+Intake3+Intake4+Intake5+Intake1-Absorption, LoadDose) DoseIncr=I1*I2*I3*I4*10 I3=IF THEN ELSE(Time>1440, 1 ,0 ) I1=IF THEN ELSE(ClCur<ThresholdSedat, 1 , 0 ) Adaptation=EffSatur*RealAdptnRate RealAdptnRate=SMOOTH3(IndAdptnRate , 15*1440 )/15 Readaptation=EffPBOnReadptn*ReadptnFrac*NoDownregRecep ReadptnFrac=0.000325 VenousToArterial=QTotal*CVenous MVenous=INTEG (ArterialToVenous+BrainToVenous+FatToVenous+KidneyToVenous+ LiverToVenous+MuscleToVenous-VenousToArterial,0) Excretion=MKidney*Kexcr 83 MArterial= INTEG (VenousToArterial-ArterialToMuscle-ArterialToLiverArterialToKidney-ArterialToGItis-ArterialToFat-ArterialToBrain-ArterialToVenous, 0) ArterialToVenous=CArterial*QHeart MFat= INTEG (ArterialToFat-FatToVenous,0) FatToVenous=QFat*CFat/PFat ArterialToFat=QFat*CArterial VFat=16.394 PFat=1 CFat=MFat/VFat QFat=0.26 Intake3=0 Intake4=0 Intake5=0 MKidney= INTEG (ArterialToKidney-KidneyToVenous-Excretion,0) TotalNoRecep=60 84 EffAdptnOnNormClCur= WITH LOOKUP (NoDownregRecep/TotalNoRecep, ([(0,0.6)-(1,1)],(0,1),(0.0825688,0.829825),(0.100917,0.807018), (0.131498,0.784211),(0.153333,0.77193),(0.186544,0.764912), (0.266055,0.750877),(0.33945,0.742105),(1,0.7) )) EffSatur= WITH LOOKUP (NoDownregRecep/TotalNoRecep, ([(0.8,0)-(1,1)],(0,1),(0.8,1),(0.8263,0.969298),(0.849541,0.907895), (0.877676,0.758772),(0.899083,0.605263),(0.933945,0.179825), (0.944342,0.100877),(0.95841,0.0351),(0.975535,0),(1,0) )) ThresholdSedat=2.5 Intake2=0 kout=LN(2)/HalflifeEnzyme HalflifeEnzyme=2880 Rin=LN(2)/(HalflifeEnzyme) Intake1=(Dose/TIME STEP)*PULSE TRAIN( 1440, TIME STEP, 1440,DaysTreatment*1440+TIME STEP) DaysTreatment=19 I2=IF THEN ELSE(Time<DaysTreatment*1440,1,0 ) CLiver=MLiver/VLiver CMuscle=MMuscle/VMuscle CVenous=MVenous/VVenous 85 CArterial=MArterial/VArterial CBraincapil=MBraincapil/VBraincapil CBraintis=MBraintis/VBraintis CGItissue=MGItissue/VGItissue CKidney=MKidney/VKidney Kexcr=0.0035 VKidney=0.308 VArterial=1.556 VBraincapil=0.0447 VMuscle=28 VGItissue=1.19 VLiver=1.925 VVenous=3.811 MBraincapil= INTEG (ArterialToBrain+BraintisToBraincapil-BrainToVenousBraincapilToBraintis,0) EffPBOnClCur=CBraintis*600/(2.79+CBraintis) WithdSignIntensity=-(ClCur-1) 86 NormClCur=ClCurWOPB*EffAdptnOnNormClCur ReaIInducbyPB=SMOOTH3(IndInducByPB, 2*1440) Synthesis=Rin*(1+ReaIInducbyPB) EffPBOnReadptn= WITH LOOKUP ( EffPBOnClCur, ([(0,0)-(800,1)],(0,1),(10.7034,0.938596),(18.9602,0.850877),(25.9939,0.714912), (42.5076,0.232456),(46.1774,0.144737),(52.9052,0.0614035),(60.367,0.0175),(70,0) ,(600,0) )) IndAdptnRate= WITH LOOKUP (ClCur/ClCurWOPB, ([(0,0)-(6,0.04)],(0,0),(1.5,0),(1.88991,0.0008635),(2.04587,0.00185025), (2.20183,0.003455),(2.37156,0.0061675),(2.52294,0.009375),(2.66972,0.0133225), (2.78899,0.0173925),(3,0.028125),(3.04465,0.03),(3.13211,0.0317544), (3.23976,0.0331579),(3.40061,0.0342105),(3.61468,0.035),(4,0.035614),(6,0.03561) )) ClCur=NormClCur*(1+EffPBOnClCur/100) ClCurWOPB=1 NoDownregRecep= INTEG (Adaptation-Readaptation,0) Kmet=NormKmet*EnzymeFactor Metabolism=CLiver*Kmet MBraintis= INTEG (BraincapilToBraintis-BraintisToBraincapil,0) BraincapilToBraintis=VBraintis*DR*CBraincapil/(1+Bplasma) BraintisToBraincapil=VBraintis*DR*CBraintis*FR 87 Absorption=MGIlumen*Kabs ArterialToBrain=QBrain*CArterial ArterialToGItis=CArterial*QGItissue ArterialToKidney=CArterial*QKidney ArterialToLiver=CArterial*QLiver ArterialToMuscle=QMuscle*CArterial Bplasma=0.438 BrainToVenous=QBrain*CBraincapil Degradation=(EnzymeFactor)*kout DR=0.02 EnzymeFactor= INTEG (Synthesis-Degradation,1) FR=1.75 MGItissue= INTEG (Absorption+ArterialToGItis-GItissueToLiver, GItissueToLiver=QGItissue*CGItissue/PGItissue Kabs=0.02 KidneyToVenous=QKidney*CKidney/PKidney NormKmet=3.14/1000 88 MLiver= INTEG (ArterialToLiver+GItissueToLiver-LiverToVenous-Metabolism, 0) LiverToVenous=(QLiver+QGItissue)*CLiver/PLiver MuscleToVenous=QMuscle*CMuscle/PMuscle MMuscle= INTEG (ArterialToMuscle-MuscleToVenous,0) PGItissue=1 PKidney=2.05 PLiver=2.25 PMuscle=1.12 QBrain=0.57 QGItissue=0.9 QHeart=0.2 QKidney=0.875 QLiver=0.235 QMuscle=1.67 QTotal=4.475 IndInducByPB=1.15*CLiver/(1+CLiver) VBraintis=1.3553 89 REFERENCES Barlas, Yaman. 1996. Formal aspects of model validity and validation in system dynamics. System Dynamics Review 12 (3): 183-210 Barlas, Yaman. 2002. System dynamics: Systemic feedback modeling for policy analysis. Knowledge for Sustainable Development - An Insight into the Encyclopedia of Life Support Systems, UNESCO Publishing-Eolss Publishers, Paris, France; Oxford, UK. Birnir, Bryndis. 2008. GABAA receptors, the brakes in the brain, Research project, Medical Faculty, Lund University, Sweden Brailowsky, S., and O. Garcia. 1999. Ethanol, GABA and epilepsy. Archives of Medical Research 30: 3-9 Chappell, Brian, and Pamela Crawford. 2001. Epilepsy: The “At Your Fingertips” Guide. London: Class Publishing Ltd. DailyMed. Current Medication Information. http://dailymed.nlm.nih.gov/dailymed /drugInfo.cfm?id=4889 (Accessed: May 14, 2008) El-Masri, H. A., and C. J. Portier. 1998. Physiologically based pharmacokinetics model of primidone and its metabolites phenobarbital and phenylethylmalonamide in humans, rats, and mice. Drug Metabolism and Disposition 26 (6): 585-594. Engasser, J. M., F. Sarhan, C. Falcoz, M. Minier, P. Letourneur, and G. Siest. 1981. Distribution, metabolism, and elimination of phenobarbital in rats: Physiologically based pharmacokinetic model. Journal of Pharmaceutical Sciences 70 (11): 12331238 90 Ffrench-Mullen, Jarlath M. H., Jeffery L. Barker, and Michael A. Rogawski. 1993. Calcium current block by (-)-pentobarbital, phenobarbital, and CHEB but not (+)pentobarbital in acutely isolated hippocampal CA1 neurons: Comparison with effects on GABA-activated Cl- current. The Journal of Neuroscience 13 (8): 3211-3221 Finn, D. A., and J.C. Crabbe. 1997. Exploring alcohol withdrawal syndrome. Alcohol Health & Research World 21 (2): 149-156 Forrester, Jay Wright. 1961. Industrial Dynamics. Portland, Oregon: Productivity Press Gay, M. H., G. P. Ryan, N. R. Boisse, and J. J. Guarino. 1983. Phenobarbital tolerance and physical dependence: Chronically equivalent dosing model. European Journal of Pharmacology 95 (1-2): 21-29 Handschin, C., and U. A. Meyer. 2003. Induction of drug metabolism: The role of nuclear receptors. Pharm. Rev. 55(4): 649-673 Hardman, J. G., and L. E. Limbird, eds. Goodman & Gilman’s The Pharmacological Basis of Therapeutics, 10th ed. (New York: Mc-Graw Hill Co., 2001) International Federation of Clinical Chemistry and Laboratory Medicine, http://www.ifcc.org (accessed November 21, 2007) Ito, T., T. Suzuki, S. E. Wellman, and I. K. Ho. 1996. Pharmacology of barbiturate tolerance/dependence: GABAA receptors and molecular aspects. Life Sciences 59 (3): 169-195 Joo, D.T, Z. Xiong, J. F. MacDonald, Z. Jia, J. Roder, J. Sonner, and B.A. Orser. 1999. Blockade of glutamate receptors and barbiturate anesthesia: Increased sensitivity to pentobarbital-induced anesthesia despite reduced inhibition of AMPA receptors in GluR2 null mutant mice. Anesthesiology 91: 1329-1341. 91 Kapur, Jaideep. 2003. Role of GABAA receptor-mediated inhibition in the pathogenesis of generalized seizures. Experimental Neurology 184: 1-2 Katzung, Bertram G. 2004. Basic & Clinical Pharmacology, 9th ed. New York: Lange Medical Books / McGraw Hill Kokka, N., D. W. Sapp, A. M. Taylor, and R. W. Olsen. 1993. The kindling model of alcohol dependence. Alcoholism: Clinical and Experimental Research 17 (3): 525531 Kumar, Vikas, Richard C. Brundage, William S. Oetting, Ilo E. Leppik, and Timothy S. Tracy. 2008. Differential genotype dependent inhibition of CYP2C9 in humans. Drug Metabolism and Disposition 36 (7): 1242-1248. Liska, Ken. 2000. Drugs and the human body: With implications for society. New Jersey: Prentice Hall Littleton, John. 1998. Neurochemical mechanisms underlying alcohol withdrawal, Alcohol Health & Research World 22(1): 13-24 Magnusson, Mats O., Mats O. Karlsson, and Rikard Sandstrom. 2006. A mechanism-based integrated pharmacokinetic enzyme model describing the time course and magnitude of phenobarbital-mediated enzyme induction in the rat. Pharmaceutical Research 23(3): 521-532 Magnusson, Mats O. 2007. Pharmacodynamics of Enzyme Induction and its Consequences for Substrate Elimination, Acta Universitatis Upsaliensis. Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Pharmacy 52, Uppsala University, Sweden MedlinePlus Webpage, http://www.nlm.nih.gov/medlineplus/druginfo/medmaster /a682007.html (accessed March 17, 2008) 92 Michalets, E. L. 1998. Update: clinically significant cytochrome P-450 drug interactions. Pharmacotherapy 1998(18): 84-112 Powis, David A., and Stephen J. Bunn, eds. Neurotransmitter Release and Its Modulation (New York: Cambridge University Press, 1995), 315-318 Raucy, Judy L., Lisa Mueller, Kui Duan, Scott W. Allen, Stephen Strom, and Jerome M. Lasker. 2002 Expression and induction of CYP2C P450 enzymes in primary cultures of human hepatocytes. Journal of Pharmacology and Experimental Therapeutics 302(2): 475-482 Sterman, J. D. 2000. Business Dynamics: Systems Thinking and Modeling for a Complex World, McGraw-Hill Co. Stevens, Serita, and Anne Louise Bannon, Book of Poisons (Writer' s Digest Books, 2007) Tanaka, E. 1999. Clinically significant pharmacokinetic drug interactions between antiepileptic drugs, Journal of Clinical Pharmacy and Therapeutics 24: 87-92 Venkatakrishnan K., L. L. von Moltke, and D. J. Greenblatt. 2000. Effects of the antifungal agents on oxidative drug metabolism: clinical relevance. Clinical Pharmacokinetics 38:111-180. Weil, Andrew and Winifred Rosen. 2004. From chocolate to morphine: Everything you need to Know about Mind-altering drugs. Boston/New York: Houghton Mifflin Company Zelko, I., and M. Negishi. 2000. Phenobarbital-elicited activation of nuclear receptor CAR in induction of cytochrome P450 genes. Biochemical and Biophysical Research Communications 277: 1-6