Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
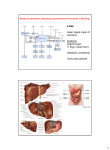
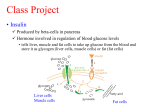
University of Groningen Metabolic adaptations in models of fatty liver disease Hijmans, Brenda IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite from it. Please check the document version below. Document Version Publisher's PDF, also known as Version of record Publication date: 2017 Link to publication in University of Groningen/UMCG research database Citation for published version (APA): Hijmans, B. (2017). Metabolic adaptations in models of fatty liver disease: Of mice and math [Groningen]: Rijksuniversiteit Groningen Copyright Other than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons). Take-down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons the number of authors shown on this cover page is limited to 10 maximum. Download date: 15-06-2017 Chapter 6 Zonation of glucose and fatty acid metabolism: mechanism and metabolic consequence Brenda S. Hijmans, Aldo Grefhorst, Maaike H. Oosterveer and Albert K. Groen Adapted from: Biochimie (2014): 96, 121-129. 103 104 Chapter 6 ABSTRACT The liver is generally considered a relatively homogeneous organ containing four different cell types. It is however well-known that the liver is not homogeneous and consists of clearly demarcated metabolic zones. Hepatocytes from different zones show phenotypical heterogeneity in metabolic features, leading to zonation of metabolic processes across the liver acinus. Zonation of processes involved in glucose and fatty acid metabolism is rather flexible and therefore prone to change under (patho)physiological conditions. Hepatic zonation appears to play an important role in the segregation of different metabolic pathways in the liver. As a consequence, perturbations in metabolic zonation may be a part of metabolic liver disease. The metabolic syndrome is characterized by the inability of insulin to adequately suppress hepatic gluconeogenesis, leading to hyperglycemia, hyperinsulinemia and eventually to type II diabetes. As insulin promotes lipogenesis through the transcription factor sterol regulatory element binding protein (SREBP)-1c, one would expect that lipogenesis should also be impaired in insulin-resistant states. In the metabolic syndrome however, hepatic de novo lipogenesis is increased, leading to hyperlipidemia and hepatosteatosis, primarily in the pericentral zone. These observations suggest the co-existence of insulin resistant glucose metabolism and insulin sensitive lipid metabolism in the metabolic syndrome. Here we provide a theoretical framework to explain this so-called insulin signaling paradox in the context of metabolic zonation of the liver. Hepatic zonation and insulin resistance 105 INTRODUCTION Due to its rather uniform histological appearance the liver is often unjustly regarded as a homogeneous organ. Already in 1856, when describing the anatomy of the liver in great detail, Lionel Beale noticed a heterogeneity of hepatocytes with regard to bile secretion and deposition of oil [237]. After this, it took more than a century until Jungermann and Sasse proposed a functional significance of heterogeneous enzyme distribution in the liver, thereby introducing the concept of metabolic zonation [238]. It was recognized that the liver may be divided into functional units which were designated acini. Inside the acinus blood flow is directional on basis of which the acinus may be subdivided into different zones [239, 240]. The periportal zone receives nutrient rich blood from the portal vein and blood rich in oxygen from the hepatic artery. In the pericentral zone blood is drained from the liver by the central vein. Hepatocytes lining the sinusoids can be classified according to their location on the portocentral axis of the acinus (see Figure 6.1). Three different zones are distinguished: 1 = periportal, 2 = intermediate, 3 = pericentral). The flow of blood through the liver generates gradients of oxygen tension, hormones and nutrients which causes hepatocytes to be exposed to different metabolic conditions depending on their location along the portocentral axis. Gradients in signals arise through interaction of blood borne components with hepatocytes, such that periportal hepatocytes are exposed to higher concentrations of blood borne components than pericentral hepatocytes [241]. Hepatocytes from different zones of the liver show phenotypical heterogeneity in metabolic features, leading to zonation of metabolic processes across the acinus [242]. Regarding glucose and fatty acid metabolism, periportal hepatocytes are more involved in gluconeogenesis and β-oxidation, while pericentral hepatocytes are more engaged in glycolysis and lipogenesis [243, 244]. Other metabolic properties of the liver, such as ammonia metabolism, xenobiotic reactions, cytoprotective functions and protein metabolism also show zonation along the portocentral axis [241]. Zonation of glucose [244] and fatty acid metabolism [245] shows flexibi-lity in different conditions in such a manner that dynamic adaptation of gene and protein expression can be observed in different nutritional states [246]. Interestingly, metabolic pathways performing opposing functions seem to follow inverse gradients and are distributed in a complementary manner. Interdependent metabolic pathways (e.g. lipogenesis and glycolysis) are co-localized 106 Chapter 6 Figure 6.1: Schematic depiction of the hepatic acinus. to allow for synergistic action, whereas opposing pathways are segregated in different zones, likely to avoid interference and thereby waste of energy. Altogether, the heterogeneity of the liver enables the simultaneous performance of different and even opposing metabolic pathways while allowing for flexible adaptation to differing circumstances [241]. This chapter will first briefly discuss the concept and mechanisms of hepatic metabolic zonation, with a special focus on zonation of glucose and fatty acid metabolism. Second, the physiological consequences of segregation of metabolic pathways in the liver will be discussed via examples of disturbed zonation in metabolic diseases of the liver. Finally we will provide a theoretical framework that may explain the relevance of hepatic zonation to aberrant metabolic states, such as insulin resistance and non-alcoholic fatty liver disease (NAFLD). Hepatic zonation and insulin resistance 107 METABOLIC ZONATION Signals underlying hepatic metabolic zonation Many different signals are involved in the establishment of zonal heterogenic properties of hepatocytes. First of all, liver innervation seems to play a substantial role (e.g. [247]). Sympathetic and parasympathetic nerve fibers enter the liver near the hepatic artery and the portal vein. The extent of further innervation of the acinus differs between species. In rats and mice innervation is limited to the periportal zone, while in humans also areas surrounding the central vein are innervated [246]. Furthermore, the directional blood flow inside the acini establishes oxygen, nutrient, nutrient intermediate and hormonal gradients along the portocentral axis [241, 246]. In the periportal zone oxygen tension and concentrations of hormones and nutrients are higher. Hepatocytes in the pericentral zone are exposed to blood enriched in CO2 and other products of metabolism. While as of now it is not entirely clear how gradients of enzymes along the portocentral axis arise, blood borne humoral factors (e.g. oxygen and hormones) are thought to be mainly involved in the dynamic adaptation of their heterogeneous expression [248] as will be discussed below. This dynamic adaptation is mostly seen in enzymes that show a gradient-like distribution, such as those involved in glucose [244] and fatty acid [245] metabolism. Other enzymes, such as carbamoyl phosphate synthetase I (CPS I) [249] and glutamine synthetase (GS) [250] the key enzymes in ammonia metabolism, show a more compartmentalized distribution. Recent studies have implicated different signaling pathways and molecules in the establishment of hepatic zonation. The following sections will provide a brief description of several of these pathways. Wnt/β-catenin signaling The involvement of β-catenin was suggested by the finding that liver tumours that contain mutations in activation of β-catenin express a pericentral-like transcription pattern [251, 252]. Immunohistochemical studies in mice have shown that adenomatous polyposis coli (APC), which is part of the β-catenin degradation complex, shows a heterogeneous distribution along the portocentral axis with a higher expression in periportal hepatocytes. The active unphosphorylated form of β-catenin was increased in hepatocytes surrounding the central vein [253, 254]. Thus, there seems to be a mutually exclusive localization of active β-catenin and one of its negative regulators across the acinus. By modulating the Wnt signaling pathway it was shown that Wnt/β-catenin 108 Chapter 6 promotes the expression of pericentral genes while downregulating periportal gene expression. When APC expression is downregulated, β-catenin is activated and an induction of normally exclusive pericentral localized genes can be observed in periportal hepatocytes. On the other hand, specific blockade of Wnt signaling induces periportal gene expression in pericentral hepatocytes [253]. Finally, pharmacological inhibition of glycogen synthase kinase 3β, a component of the β-catenin degradation complex, leads to suppression of periportal gene expression and activation of pericentral gene expression in a resident liver stem cell line [255]. Ha-RAS/MAPK signaling While β-catenin signaling imposes a pericentral pattern of gene expression, the periportal pattern is likely regulated by the Ha-RAS pathway. The first implications for involvement of Ha-RAS signaling in metabolic zonation were derived from the observation that mutated Ha-RAS liver tumours display a periportal pattern of gene expression [256]. Also, a portal to central gradient was found for the phosphorylated form of extracellular signal-regulated kinase (ERK), a downstream target of RAS. A discrepancy in this finding is however described: in female rats phosphorylated ERK (p-ERK) levels are higher in pericentral areas, especially during the pro-oestrous phase [257]. It may therefore be possible that the difference in zonation of pERK between females and males is dependent on female sex-hormone levels. Activation of the RAS/MAPK pathway induces ERK expression in cells that do not express GS or other pericentral genes [252, 256]. Furthermore, induction of the RAS/MAPK pathway by serum components also suppressed GS expression [258]. Combined, these studies indicate that Ha-RAS favours a periportal pattern of gene expression, while it abolishes a pericentral pattern. HNF4α Hepatic nuclear factor 4α (HNF4α) is a transcription factor that is highly expressed in liver, kidney, intestine and pancreas. HNF4α can bind to the promoter region of 12% of the genes expressed in the liver [259]. A rather homogeneous expression of this transcription factor was found throughout the hepatic lobule, with slightly more transcripts in pericentral hepatocytes [260]. Liver-specific ablation of HNF4α induces GS expression in periportal hepatocytes while weakening the expression of periportal PEPCK [261]. This indicates that HNF4α stimulates the expression of periportal enzymes while inhibiting expression of pericentral enzymes. Interestingly, recent findings support a con- Hepatic zonation and insulin resistance 109 vergence of the Wnt/β-catenin signaling and HNF4α in control of metabolic zonation. Using co-immunoprecipitation HNF4α was shown to interact with lymphoid enhancer binding factor-1 (LEF1), a Wnt target [255]. MicroRNAs MicroRNAs (miRNAs) are a group of small noncoding RNAs of approximately 18-25 nucleotides long. In animals, miRNAs repress translation of their target messenger RNAs (mRNAs) through partial complementary binding to the coding region or the 3'untranslated region (3'UTR) [262]. In addition to this, miRNAs can direct cleavage of mRNAs, thereby reducing their amount [263]. Thus, miRNAs regulate gene expression at the posttranscriptional level, inhibiting mRNA translation into proteins. It is becoming increasingly clear that these molecules play a prominent role in regulation of gene expression. One study has examined a general role of miRNAs in metabolic zonation by knocking out Dicer1. This gene encodes for the protein Dicer, which cleaves pre-miRNAs into mature, functioning miRNAs. Loss of Dicer and thus a loss of miRNAs resulted in impaired localization of the examined periportal proteins. Zonal expression of pericentral proteins was less affected, with only expression of some proteins extending beyond the normal boundaries while remaining mainly pericentrally located [264]. Interestingly, ablation of either β-catenin or Dicer induced a similar phenotype. Both models show a loss of heterogeneous localization of periportally expressed proteins, and a rather diffuse expression pattern throughout the liver acinus [253, 255, 264]. This suggests that both β-catenin and Dicer are required for normal localization of periportal enzyme expression. Conserved expression of pericentral gene expression indicates that Wnt/β-catenin signaling is intact in Dicer-deficient livers. Furthermore, it was found that Dicer expression is not affected by β-catenin deficiency [265]. Microarray analysis did not reveal individual miRNAs that are directly activated by β-catenin signaling but identified four miRNAs that are modestly upregulated [264]. Therefore it seems that β-catenin in concert with yet to be specified microRNAs impact another factor which in turn inhibits periportal gene expression. Although loss of Dicer expression indicates a role for miRNAs in zonation, it is far from clear which specific miRNAs are involved. Presently, over 1000 miRNA have been found in humans, which are predicted to regulate the activity of more than 60% of all protein-coding genes. This suggests that miRNAs play a role in most, if not all, (patho)physiological processes. However, bioinformatic predictions and experimental approaches indicate that a single miRNA 110 Chapter 6 may target more than a hundred mRNAs [265]. Given the current popularity of this area of research our insight into the mechanisms of miRNA action is expected to significantly increase within the coming years. In summary, many signals have been implicated in zonation of protein expression alongside the portocentral axis. In this respect, the Wnt/β-catenin signaling and its antagonist, the Ha-RAS signaling pathway appear to have a major contribution. It is however not clear which factors induce the heterogeneity in these signaling pathways and how the different signals interact. MiRNAs also play a role, but the mechanistic basis of their actions remains to be established. Although our insights into the mechanisms of metabolic zonation are still limited, it is important to evaluate the zonation of metabolic processes under different conditions. Before discussing this issue, further details about zonation of glucose and fatty acid metabolism will be outlined. Zonation of glucose metabolism The liver plays a central role in the maintenance of glucose homeostasis. During the fed state it takes up excess amounts of glucose and converts it to glycogen and triglycerides (TG) while oxidizing it to CO2 . To maintain normoglycemia during fasting states, the liver releases glucose derived from glycogenolysis and gluconeogenesis into the bloodstream [266]. The bulk of research on acinar zonation of enzymes involved in glucose and fatty acid metabolism dates back to the eighties and nineties of the last century. Partitioning of different processes involved in glucose metabolism along the liver acinus constituted the basis for the concept of metabolic zonation [241, 246]. According to the concept of metabolic zonation, gluconeogenesis mainly takes place in the periportal zone, while glycolysis is performed in pericentral hepatocytes. Many studies have reported increased gluconeogenic gene and protein expression in periportal hepatocytes, whereas glycolytic enzymes show higher expression in pericentral hepatocytes (for reviews, see: [267, 268, 269]). More importantly the rate of glucose formation from gluconeogenic precursors was found to be higher in cultured periportal hepatocytes isolated from fasted animals after collagenase-digitonin perfusion [270, 271]. Glycogen synthesis via the indirect pathway was also found to be higher in periportal cells [270], showing that gluconeogenic flux is increased in periportal zones. While glucokinase activity is higher in pericentral hepatocytes, no zonal differences in glycolytic flux were found [270, 271]. These in vitro studies were however not performed under physiological oxygen tensions and did not account for the dif- Hepatic zonation and insulin resistance 111 ferent oxygen tensions and hormonal gradients along the portocentral axis. It has been shown that glucose metabolism is significantly modified upon oxygen supply, and that glucose conversion to lactate is higher under pericentral-like oxygen tension and hormonal concentrations [272]. Furthermore, it was shown that perfusion of liver with glucose promotes glycogen deposition in pericentral zones while perfusion with lactate induces glycogen formation in periportal hepatocytes [273]. This suggests that glucokinase flux is higher in pericentral hepatocytes. It has recently been shown that glucose flux is required for nuclear localization of β-catenin in enteroendocrine cells [274]. A higher glucose flux through glucokinase in pericentral hepatocytes may therefore provoke a portocentral gradient of active β-catenin in liver. Glycogen synthesis and utilization are also distributed in a zonated manner across the acinus. Glycogen degradation is initiated in the periportal zone and ends in the pericentral zone while glycogen stores are replenished in the opposite direction [275]. Zonation of fatty acid metabolism Evidence is abundant that fatty acid oxidation takes place at higher rates in periportal hepatocytes, while pericentral hepatocytes are more engaged in lipogenesis. It was shown that the rate of fatty acid synthesis is higher in pericentral hepatocytes, while the rate of β-oxidation is higher in periportal cells [276, 277]. In microdissected rat liver tissue, Katz and colleagues reported higher activities and higher presence of the lipogenic enzymes acetyl-CoA carboxylase (ACC) and ATP citrate lyase (ACL) in pericentral areas. Furthermore, the activity of carnitine palmitoyltransferase-1 (CPT1), a key enzyme in β-oxidation, was found to be higher in periportal hepatocytes [278]. CPT-1 also shows a lower sensitivity to inhibition by the lipogenic intermediate malonyl-coA in periportal as compared to pericentral areas [243, 279]. Esterification of fatty acids and synthesis of very low density lipoprotein (VLDL) is slightly higher in hepatocytes derived from pericentral areas. This uneven distribution has been suggested to avoid TG accumulation due to increased lipogenesis in this part of the liver. However, the same study found no differences in TG excretion rate for hepatocyte fractions derived from either periportal or pericentral zones of the liver [243]. There are indications for higher fatty acid uptake in periportal hepatocytes. Immunohistochemical staining shows that liver fatty acid binding protein (LFABP) is present in a portocentral gradient, declining in its expression from periportal to pericentral in rodents [280] and in humans [281]. L-FABP is a 112 Chapter 6 cytoplasmic lipid binding protein involved in intracellular lipid transport. Its heterogeneous distribution indicates a higher uptake and utilization of fatty acids in periportal liver cells. However, digitonin/collagenase isolated periportal and pericentral cells show no differences in radiolabeled oleic acid incorporation into TG and phospholipids and no changes in lipogenic rates and water-soluble oxidation products [282]. This does however not exclude the possibility that in an intact organism, where a complex interplay of gradients of hormones and substrates exists, a differential uptake of fatty acids in periportal versus pericentral hepatocytes may occur. Indeed, upon perfusion with a fluorescent-labeled stearic acid a portal to central decline in fluorescence could be observed in isolated liver, indicating higher uptake of fatty acids in periportal as compared to pericentral hepatocytes. Interestingly, this pattern was reversed upon retrograde perfusion, indicating the presence of a 'first pass'-effect [282, 283]. Flexibility of metabolic zonation It is intriguing to realize that the localization of enzymes involved in fatty acid and glucose metabolism shows flexibility. Some enzymatic distribution gradients partly overlap and may switch according to physiological needs [279]. The zonal distribution of enzymes involved in glucose and fatty acid metabolism may vary under different (patho)physiological circumstances. For instance, in alloxan-induced diabetes in rats, zonal heterogeneity of gluconeogenic and glycolytic enzymes is perturbed. This diabetic state is characterized by a zonation pattern resembling that of fasted rats [284]. Similarly, streptozotocin-induced diabetic ketoacidosis was shown to increase hepatic glucose output and ketone production in perfused rat liver. The same study also applied different degrees of digitonin-induced pericentral hepatocyte damage to demonstrate that diabetic ketoacidosis led to an increase in gluconeogenesis in pericentral hepatocytes specifically [285]. Disturbances in zonation of hepatic fatty acid metabolism were also observed in a rat model for alcoholic fatty liver disease (AFLD) [286]. Ethanol feeding caused a selective deposition of TG in pericentral hepatocytes, a pattern also seen in humans. In control animals, lipogenic rate and ACC activity were found to be higher in pericentral areas, whereas this heterogeneity was blunted after ethanol feeding. The ethanol group exhibited a higher rate of β-oxidation in pericentral hepatocytes, while this process was more abundant in periportal areas of control animals. Furthermore, in this model VLDL secretion and TG incorporation into VLDL were inhibited in pericentral hepatocytes. Combined, these data show that AFLD is characterized by disturbed zonal heterogene- Hepatic zonation and insulin resistance 113 ity of hepatic lipid metabolism. The outcome of this study also suggests that a reduction in VLDL-TG excretion may be the primary cause of pericentral steatosis in AFLD. The collected evidence for altered heterogeneity in metabolic enzyme expression under pathophysiological conditions leads to the question whether dysregulation of metabolic zonation is involved in or may even induce liver pathology. AFLD and NAFLD are both characterized by disturbances in fatty acid metabolism and show similar histological liver pathology. It is therefore tempting to speculate that, as is the case for AFLD, also in NAFLD metabolic zonation will be disturbed. NON-ALCOHOLIC FATTY LIVER DISEASE In NAFLD TG accumulate in hepatocytes due to imbalances in hepatic fatty acid metabolism [11]. The precise mechanisms that lead to steatosis in NAFLD have not been elucidated. The deposition of TG in the liver most likely has different underlying origins that may vary between individuals. Hepatic TG accumulation may for instance either result from an increased fatty acid influx, a decreased fatty acid efflux, or a combination of the two. There are different ways in which TG may reach the liver. Dietary TG that are transported via chylomicrons from the intestine may be delivered directly to the liver, or may first reach adipose tissue and reach the liver in the form of FFA after lipolysis. Fatty acids for esterification inside the liver are provided through either the plasma FFA pool or via de novo lipogenesis (DNL). Decreased efflux of hepatic TG may be due to a decrease in β-oxidation or a decrease in transport of lipids out of the liver in VLDL particles [11]. A dynamic balance of the above mentioned pathways is needed to maintain homeostasis of hepatic TG content. To adequately evaluate the basis of steatosis in NAFLD, all pathways involved in influx and efflux of hepatic TG should be studied in parallel. Donnelly et al. quantified fatty acid metabolism and the origin of hepatic TG in NAFLD patients using 13 C-labeled isotope enrichment of free fatty acids (FFA), TG and a fatty acid precursor in the hepatic TG pool. They found that in NAFLD patients, postprandially, hepatic and VLDL TG are for the largest part (59%) derived from circulating FFA. The contributions of DNL and dietary fatty acids were found to be smaller, 26% and 15%, respectively. During fasting about 80% of hepatic and VLDL TG were derived from the plasma compartment, suggesting that they are mainly derived from adipose 114 Chapter 6 tissue lipolysis [119]. While healthy subjects show an increase in fractional DNL from 5% in the fasted to 23% in the postprandial state [287], NAFLD patients [119] and hyperinsulinemic obese subjects [288] did not shown an elevation of DNL in response to feeding. Instead, in NAFLD and hyperinsulinemic obese subjects, DNL was already elevated in the fasted state and contributed for up to 25% to hepatic and VLDL TG in both fasted and postprandial conditions [119, 288]. Regarding the efflux of hepatic TG, in NAFLD patients generally an induction of genes involved in β-oxidation is reported [289, 290]. In support of this, higher levels of circulating ketone bodies in these subjects are indicative of increased hepatic β-oxidation [291]. The excretion of TG-rich VLDL particles was furthermore elevated [292]. Although VLDL-TG secretion increased with higher intrahepatic lipid content, it reached a plateau at higher TG concentrations, suggesting that TG secretion in VLDL somehow fails to increase further above a certain threshold [292]. Therefore it seems that plasma derived FFA are the main contributor to hepatic TG in NAFLD, suggesting that the main contributor of hepatic steatosis in the metabolic syndrome is increased adipose tissue lipolysis. Zonation of steatosis Deposition of TG in NAFLD is often found to be distributed in a zonated manner. In humans, steatosis associated with AFLD, and many forms of NAFLD is described to initiate in pericentral areas, with advancement to intermediate and periportal areas upon disease progression [293, 294]. As stated earlier, under normal physiological circumstances periportal zones show higher rates of fatty acid oxidation while lipogenesis is more pronounced in pericentral zones. In addition, changes in this segregation of fatty acid metabolism have been observed under aberrant metabolic conditions. Unfortunately, not many studies have evaluated metabolic zonation in NAFLD. Some studies have found increased lipid peroxidation and oxidative damage in pericentral areas in liver biopsies from human non-alcoholic steatohepatitis (NASH) patients [295, 296]. This indicates a zonation of mitochondrial dysfunction to pericentral areas in steatohepatitis. Enhanced lipid peroxidation and oxidative stress are known to occur at lower oxygen tensions [297], which may account for predominant pericentral hepatocyte damage due to obesity or ethanol use. While in adults steatosis is most often localized in pericentral zones, this is not the case for paediatric steatosis. Some studies found paediatric steatosis to be typically azonal or to have a homogeneous distribution [298, 299], while others report a predominant periportal pattern [298]. In malnutrition disorders, Hepatic zonation and insulin resistance 115 infections and toxic insults, steatosis seems to originate in the periportal zone [300]. These differences indicate that under different pathophysiological circumstances zonation of steatosis may vary. More insight is needed into specific zonal alterations that occur in the liver acinus under these different conditions. Whether zone-dependent initiation and progression of steatosis and development into NASH exists is also not known. Further insight into disease progression is needed to reveal by what mechanisms TG accumulation progresses, and how steatosis may develop into NASH. Zonation of steatosis in animal models of NAFLD Genetic or diet-induced animal models are commonly used to examine pathophysiology of hepatosteatosis. Unfortunately not many studies report on zonal distribution of TG deposition. Those models in which a zonal distribution of steatosis has been described will be discussed here. Genetic models A distinction can be made between spontaneous and induced genetic models of NAFLD. The two most often used models are spontaneous genetic models: ob/ob and db/db mice. Ob/ob mice exhibit a naturally occurring mutation that results in the absence of the satiety hormone leptin that inhibits feeding behaviour and stimulates energy expenditure [301]. These animals display hyperphagia and obesity. Db/db mice possess a genetic defect that leads to a similar phenotype. These animals do not have functioning receptors for leptin, resulting in a loss of leptin signaling (e.g. [302]). Both genetic models develop severe obesity, accompanied by a reduction in insulin sensitivity, abnormalities in (hepatic) fatty acid metabolism and hepatic steatosis [303]. Hepatosteatosis is located in pericentral hepatocytes in ob/ob mice [304]. To our knowledge, it has not been examined whether lipogenesis or other aspects of fatty acid metabolism are deregulated in a zonal manner in these animals. To date, the localization of TG accumulation in db/db mice has not been reported. However, histological pictures of the liver reveal a clearly zonated fat accumulation [305]. Diet-induced models C57BL/6 mice fed a high fat diet (HFD) are often used as a model to study the metabolic syndrome. On a normal rodent diet, this mouse strain spontaneously develops obesity, hyperinsulinemia and glucose intolerance with age, which can be induced earlier in life by feeding a HFD [306]. In addition, C57BL/6 mice 116 Chapter 6 develop hepatosteatosis on a HFD. Not many studies report on zonation of diet-induced steatosis. Some recent articles report predominant fat accumulation in pericentral hepatocytes in response to a HFD in mice [307, 308, 309]. However, in another study high fat feeding was shown to predominantly lead to deposition of TG in hepatocytes in areas surrounding the portal vein [310]. Overfeeding mice a high fat liquefied diet was shown to cause steatosis throughout the whole liver acinus. Interestingly, this led to upregulation of lipogenic genes in pericentral hepatocytes [310]. It should be noted that the diets used in these studies differ in fat and carbohydrate content. Because glucose and fructose are metabolized in a different manner, the source of carbohydrates may also be of importance [311]. This content difference may account for the difference in histological findings, particularly because the diet used in the study of Gaemers and colleagues had a relatively high carbohydrate content (47%). It has been shown that different dietary fat types may have different metabolic consequences, leading to differences in for instance liver TG content [312]. Diets high in carbohydrates seem to induce periportal steatosis. In Sprague-Dawley rats high dietary concentrations of sucrose (40-50%) lead to periportal fat accumulation [313]. Mice fed a high fat/high fructose diet initially also show TG accumulation in periportal hepatocytes. In this model, small fat droplet accumulation in pericentral zones was noticed after eight weeks of feeding [314]. There is no published research on the basis of the difference in zonation of steatosis that occurs when feeding different diets. A sucrose or fructose-rich diet would lead to an increased flux of fructose to the liver via the portal vein. The loss of fructose in the urine in persons that have an inherited deficiency of fructokinase [315] shows that fructose is not well metabolized in extrahepatic tissues. Furthermore, the isoform fructokinase-C, which is considered the most active in metabolizing fructose [316] is only expressed in liver, intestine and kidney [317]. A high availability of fructose will lead to rapidly decreasing hepatic intracellular ATP levels and a decline in free phosphate due to the low Km of fructokinase for fructose and the absence of negative feedback by energy status on its metabolism [318]. Therefore, triose-phosphate derivatives of fructose will be metabolized rapidly by glycolysis or will enter the gluconeogenic pathway. It has been shown in humans that prolonged fructose, but not glucose consumption leads to increased postprandial hepatic DNL and elevated postprandial plasma TG [319]. It could be argued that a fructose rich diet will primarily result in periportal zonation of steatosis, since in rats fructose is taken up mainly by periportal hepatocytes [320]. On the other hand, as discussed in more detail in a subsequent section, Hepatic zonation and insulin resistance 117 a high fat diet may lead to a portocentral gradient in insulin sensitivity, which may account for pericentral accumulation of TG. INSULIN RESISTANCE AND STEATOSIS NAFLD is regarded as the hepatic component of the metabolic syndrome [2], showing clear correlations with diabetes [321] and obesity [5]. Although a strong correlation exists between insulin resistance and NAFLD, the causality behind this association is uncertain. For instance, when hepatic steatosis is induced by pharmacological inhibition of β-oxidation this does not lead to hepatic insulin resistance [322]. In genetic diseases that promote hepatic FFA influx, such as glycogen storage disease type 1, steatosis is also found without the presence of insulin resistance [146]. Moreover, in liver insulin receptor knockout (LIRKO) mice, which display complete hepatic insulin resistance, no hepatic steatosis is found [323]. In liver X receptor (LXR) agonist treated mice severe hepatic steatosis is induced without hepatic insulin resistance [324]. Finally, in mice with a liver-specific deletion of histone deacetylase 3 (HDAC3), a fatty liver is even accompanied by an increase in insulin sensitivity [309]. These findings show that hepatic fat deposition per se does not lead to insulin resistance and insulin resistance in itself does not produce steatosis. For further reading on this subject the reader is referred to a recent review [325]. Since insulin resistance and fatty liver are both aspects of the metabolic syndrome, it is important to elucidate the mechanisms by which these symptoms are associated and may influence each other. According to the concept of metabolic zonation, one can even wonder whether it is relevant to correlate TG accumulation with the failure of insulin to inhibit gluconeogenesis because these processes are normally localized in different hepatic zones. In NAFLD, pericentral hepatocytes more often show TG accumulation [293, 294], while gluconeogenic activity is mainly located in periportal hepatocytes [326]. The insulin signaling paradox It has been suggested previously that in the metabolic syndrome the liver shows a mixed pattern of insulin resistance and sensitivity [327, 328]. In persons suffering from the metabolic syndrome gluconeogenesis is not (sufficiently) suppressed by insulin, leading to hyperinsulinemia, and eventually to hyperglycaemia and type II diabetes. As insulin stimulates lipogenesis through SREBP-1c (reviewed in [329]), one would expect that in the presence of hepatic insulin resistance, 118 Chapter 6 lipogenesis should also be impaired. However, a paradox exists with respect to lipid metabolism: in type II diabetes DNL is increased, leading to hyperlipidemia and hepatosteatosis [288]. These findings suggest the presence of selective insulin signaling in the metabolic syndrome, with insulin resistance of gluconeogenesis, while lipid metabolism appears to remain insulin sensitive. It has been suggested that mixed insulin sensitivity may arise from different signaling pathways for insulin to lipid and glucose metabolism. Insulin receptor substrate (IRS) 1 was proposed to signal to lipid metabolism, while IRS 2 would signal to glucose metabolism [330]. Recent studies have however pointed out that IRS1 and IRS2 together signal to both hepatic glucose and lipid metabolism under different dietary circumstances [331, 332]. Other manners of branching of the insulin signaling pathway downstream of the insulin receptor have also been suggested [333, 334, 335, 336], but do not (fully) explain mixed insulin sensitivity and zonation of steatosis. More recently, Sun and colleagues [309, 325] have proposed an interesting theory which may explain the insulin signaling paradox from a biochemical viewpoint. It is hypothesized that under pathological conditions of overnutrition the oversupply of metabolic intermediates is the driving force behind the co-existence of gluconeogenesis and lipogenesis. In these conditions the flux of FFA to the liver is thought to be too high for lipogenesis to keep up, thereby leading to accumulation of lipid intermediates which may cause hepatic insulin resistance [309]. As discussed before, gluconeogenesis mainly takes place in periportal hepatocytes, while pericentral hepatocytes are more engaged in lipogenesis. Here, we propose an alternative theory which explains how hepatic metabolic zonation may lead to selective insulin signaling. The insulin signaling pathway that inhibits gluconeogenesis is well studied. Insulin binds to the insulin receptor present on cell membranes, leading to a cascade of phosphorylation of downstream enzymes: IRS, phosphoinositide 3-kinase (PI3K) and Akt. Eventually phosphorylation of Forkhead Box 1 (Fox01) results in its nuclear exclusion, thereby reducing gene expression of gluconeogenic enzymes (reviewed in [336]). The signaling pathway that leads to increased SREBP-1c expression is however less well studied. Liver-specific deletion of the insulin receptor in mice (LIRKO mice) leads to hyperinsulinemia and hyperglycaemia with normal plasma and hepatic TG levels. These mice show an increase in Srebp-1c expression [323], accompanied by an increase in Pepck expression [337]. Thus, total hepatic insulin resistance leads to a phenotype that does not match the metabolic syndrome. When downstream targets in the IRS/PI3K/Akt Hepatic zonation and insulin resistance 119 pathway are inhibited, a similar phenotype is found: hyperglycaemia, with normal to low plasma lipid levels. For instance, mice deficient for the two major insulin receptor substrates (IRS1 and IRS2) in the liver show hyperglycaemia, but have normal plasma lipid values [331]. Moving even further downstream, it has been shown that inhibition of PI3K or Akt leads to blocking of insulininduced increases in Srebp-1c and decreases in Pepck expression [333]. It is also known that Akt drives lipogenesis in multiple ways. Activation of mTORC1 increases lipogenesis through induction of SREBP-1c [338]. Also, Akt mediated inhibition of Insig2, which activity retains SREBP-1c in the endoplasmic reticulum, increases the active form of SREBP-1c [338]. Finally, transgenic mice that express a constitutively active form of Fox01 in liver show hyperglycaemia, but reduced plasma lipid levels [339]. Therefore it seems that insulin-induced upregulation of hepatic lipogenesis and downregulation of hepatic gluconeogenesis may be regulated via a common signaling pathway. From these studies it may be proposed that excessive insulin signaling rather than insulin resistance is responsible for hepatosteatosis and dyslipidemia in type II diabetes. Selective insulin sensitivity across the portocentral axis How can glucose metabolism be insulin resistant and lipid metabolism insulin sensitive if these metabolic pathways are regulated through a common arm of the insulin signaling pathway? Here, we propose a model of zonation of insulin sensitivity along the portocentral axis of the liver. Under normal circumstances high circulating insulin levels suppress adipose tissue lipolysis in the postprandial state. In obese individuals, insulin resistance causes insufficient suppression of hormone sensitive lipase (HSL), leading to excessive lipolysis in adipose tissue. It has indeed been reported that insulin does not suppress adipose tissue lipolysis in NAFLD patients to the same extent as it does in healthy individuals [291]. Metabolites of FFA, such as ceramides, may inhibit hepatocyte insulin signaling in two ways. First, ceramides interfere with the insulin signaling pathway by activating protein kinase C (PKC) ζ [340] and stabilizing interaction between PKCζ and Akt, which blocks Akt translocation to the plasma membrane [341, 342]. Second, ceramides stimulate activity of protein phosphatase 2A (PP2A), which dephosphorylates and thereby deactivates Akt [343, 344]. Circulating FFA may be taken up predominantly by periportal hepatocytes, perhaps due to a first-pass effect [282, 283]. The periportal accumulation of FFA metabolites may cause insulin resistance in these hepatocytes, while pericentral hepatocytes may take up less FFA and may therefore remain relatively 120 Chapter 6 Figure 6.2: Proposed model for zonation of insulin signaling in liver. PP = periportal, PC = pericentral. See text for details. insulin sensitive. This will lead to a zonated maintenance of SREBP-1c mediated lipogenesis and may explain selective accumulation of TG in pericentral hepatocytes. We thus propose a model in which increased circulating FFA concentrations in obese subjects will predominantly impair insulin sensitivity in periportal hepatocytes. This will result in impaired insulin-mediated inhibition of gluconeogenesis and glycogen synthesis and hence enhanced hepatic glucose production by periportal hepatocytes. The increased blood glucose concentrations will promote pancreatic insulin release, hence inducing hyperinsulinemia. The increased insulin levels will enhance pericentral insulin signaling resulting in increased lipid synthesis in the pericentral area. This will lead to accumulation of TG in pericentral zones of the liver, accompanied by an increase in the secretion of large, TG-rich VLDL particles, resulting in hyperlipidemia (see Figure 6.2). Hepatic zonation and insulin resistance 121 FINAL REMARKS In human adults steatosis is in most cases localized in pericentral hepatocytes, while in paediatric cases steatosis is more present in periportal zones of the liver. Research in animal models of NAFLD has shown that under differing circumstances steatosis may occur in different zones of the liver. Although it is crucial for an animal model to closely reflect the human situation, one should also acknowledge that there is a variety in the mechanisms that lead to steatosis in humans. By using these different NAFLD models, future work should provide more detailed insights into associations between the location of hepatic TG accumulation and possible changes in metabolic zonation. Many zonated enzymes in the liver show flexibility of expression patterns under changing conditions. This shows the ability of hepatocytes to change their repertoire to differing physiological conditions and metabolic demands. The variety in zonation of fat deposition between steatosis models illustrates the complexity and flexibility of metabolic zonation. Hepatocytes across the portocentral axis are not homogeneous, nor is TG accumulation. Therefore, zonespecific investigation of changes in fatty acid metabolism and insulin signaling should generate more precise answers to the mechanisms underlying NAFLD. New insights into regulation of metabolic zonation in the metabolic syndrome may also lead to development of therapies that target zone-specific hepatocytes. In this respect, periportal-specific upregulation of insulin signaling may be an interesting approach. It has been reported that adenovirus associated vector-mediated modulation of gene expression is more effective in periportal hepatocytes in non-human primates. This will however be a challenging approach, because an inverse pattern of induction is seen in mice [345]. In conclusion, elucidation of the mechanisms responsible for induction and maintenance of hepatocyte heterogeneity remains a challenge in experimental hepatology. It is however important to consider this heterogeneity of the liver, because of possible influences of (aberrant) metabolic zonation on pathological conditions.