Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
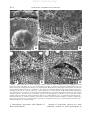
Interxylary phloem: Diversity and functions Sherwin Carlquist Brittonia ISSN 0007-196X Brittonia DOI 10.1007/s12228-012-9298-1 1 23 Your article is protected by copyright and all rights are held exclusively by The New York Botanical Garden. This e-offprint is for personal use only and shall not be selfarchived in electronic repositories. If you wish to self-archive your article, please use the accepted manuscript version for posting on your own website. You may further deposit the accepted manuscript version in any repository, provided it is only made publicly available 12 months after official publication or later and provided acknowledgement is given to the original source of publication and a link is inserted to the published article on Springer's website. The link must be accompanied by the following text: "The final publication is available at link.springer.com”. 1 23 Author's personal copy Interxylary phloem: Diversity and functions SHERWIN CARLQUIST Santa Barbara Botanic Garden, 1212 Mission Canyon Road, Santa Barbara, CA 93105, USA; e-mail: [email protected] Abstract. Interxylary phloem is here defined as strands or bands of phloem embedded within the secondary xylem of a stem or root of a plant that has a single vascular cambium. In this definition, interxylary phloem differs from intraxylary phloem, bicollateral bundles, pith bundles, and successive cambia. The inclusive but variously applied terms included phloem and internal phloem must be rejected. Histological aspects of interxylary phloem are reviewed and original data are presented. Topics covered include duration of interxylary phloem; relationship in abundance between sieve tubes in external phloem and interxylary phloem; distinctions between interxylary and intraxylary phloem; presence of parenchyma, fibers, and crystals in the interxylary phloem strands; development of cambia within interxylary phloem strands; three-dimensionalization and longevity of phloem, systematic distribution of interxylary phloem; physiological significance; and habital correlations. No single physiological phenomenon seems to explain all instances of interxylary phloem occurrence, but rapidity and volume of photosynthate transport seem implicated in most instances. Key Words: Bicollateral bundles, included phloem, intraxylary phloem, photosynthate conduction, successive cambia. Interxylary phloem consists of strands of sieve tubes, companion cells, and adjacent parenchyma or other cells embedded within the secondary xylem of a stem or root that has a single vascular cambium. This definition is presented to distinguish interxylary phloem from a series of other histological phenomena that may have similar functions but are histologically and ontogenetically different. For example, the term successive cambia denotes a series of vascular increments, each with secondary phloem, secondary xylem, and a vascular cambium, each of which ultimately originates from the master cambium at the periphery of a stem or root (Carlquist, 2007). The master cambium produces secondary cortex (0 parenchyma) to the outside, and to the inside, conjunctive tissue and vascular cambia to the inside of an axis. Each vascular cambium then produces secondary phloem to the outside and secondary xylem to the inside. The term included phloem was misapplied to successive cambia in some (but not all) Nyctaginaceae by Chalk and Chattaway (1937), but used also for instances of interxylary phloem. Misapplications of this sort render the term included phloem imprecise, and, in any case, are based on topographic phloem distribution without regard to ontogenetic factors. The ontogeny of phloem and xylem within various cambia variants can be easily determined from the mature histology, and thus can readily be included in definitions of cambial variants. The term "included" suggests that the phloem in instances of successive cambia is embedded within secondary xylem (as it is in the case of interxylary phloem). In fact, the phloem in examples of successive cambia lies between secondary xylem (internal to it) and conjunctive tissue (external to it) in each vascular increment. The term internal phloem has likewise been contaminated by conflicting usages and should be rejected. "Internal phloem" has been used to refer to intraxylary phloem, but has been applied to other histological conditions. Significantly, like "included" phloem, the term Brittonia, DOI 10.1007/s12228-012-9298-1 ISSN: 0007-196X (print) ISSN: 1938-436X (electronic) © 2013, by The New York Botanical Garden Press, Bronx, NY 10458-5126 U.S.A. Author's personal copy [VOL BRITTONIA internal phloem is vague with respect to ontogeny as well as location of phloem. Intraxylary phloem, although readily distinguishable from interxylary phloem, may have a similar physiological significance and is covered in a later section of this paper. The present usages are consistent with those adopted in earlier accounts of cambial variants (Carlquist, 1988, 2001, 2007). For the present, workers would be well advised to define the terms they use for cambial variants. Interpretation of functions of interxylary phloem is further complicated by the fact that in woody angiosperms as a whole, interxylary phloem occurs in only a relatively small number of families and species. Even within a genus such as Combretum or Strychnos, some species have interxylary phloem, others lack it, with no clear differences in habit or size of plant (van Vliet, 1979; Mennega, 1980). However, there is some correlation with systematic units within genera such as these (van Vliet, 1979; Mennega, 1980). There is no unique function for interxylary phloem; other phloem distributions seem to be adequate alternates. That does not mean, however, that interxylary phloem is not a physiologically significant way of meeting a plant's photosynthate conduction requirements. Wood anatomy contains many examples of alternative ways of serving particular functions (e.g., vestured pits, helical sculpture of vessel surfaces, and vasicentric tracheids are probably all methods of minimizing embolism formation —or reversing that). Interxylary phloem, like vestured pits, is a device that is homoplastic in woody angiosperms. In both instances, genetic information for the formation of these structures has not been frequently achieved phylogenetically, perhaps because a complex series of genetic changes is required. The data presented here may offer interesting examples that lend themselves to physiological studies. Plant physiological studies have traditionally been done on economically important plants, and none of the species with interxylary phloem has any major economic value. materials and methods used in those studies are given in the papers listed below. In all cases, however, the photographs and observations are new. Re-studied materials include the following: Figure 1: Turbina stenosiphon (Hallier f.) A. Meeuse (Convolvulaceae): Carlquist and Hanson, 1991. Figure 2A–C: Thunbergia laurifolia Lindl. (Acanthaceae): Carlquist and Zona, 1988 Figure 2D: Stylidium glandulosum Salisb. (Stylidiaceae): Carlquist, 1981 Figure 4: Pseudolopezia longiflora Rose and Oenothera linifolia Nutt. (Onagraceae): Carlquist, 1975. Figure 6: Salvadora persica L.(Salvadoraceae): Carlquist, 2002. Sources for material not previously studied are as follows: Figure 3: Orphium frutescens E. Mey. (Gentianaceae): Carlquist 8212, June 28, 2011 (SBBG). Figure 5: Craterosiphon scandens Eng. & Gilg (Thymeleaceae): Breteler 1227 (WAG). Figure 7A–B. Strychnos madagascariensis Poir. (Loganiaceae): David Lorence 10285 (PTBG), National Tropical Botanical Garden living collections accession number 801348. Figure 7C–D. Combretum erythrophyllum Sond. (Combretaceae): cultivated in the Vavra Garden (formerly owned by University of California, Los Angeles). The sections in Figs. 3 and 7C–D were derived from living material that was preserved in 50 % aqueous ethanol, sectioned on a sliding microtome, and stained with a Safranin-Fast Green combination. The section in Fig. 7A–B was derived from living material that was preserved in aqueous 50 % ethanol. The sections were prepared by driving a single-edged razor blade into a stem with the aid of a hammer. The sections derived were subjected to changes of distilled water, and dried between glass slides under pressure (to prevent curling), then sputter-coated with gold and examined with a Hitachi S2600N scanning electron microscope (SEM). Aspects of Interxylary Phloem Materials and methods Some of the examples cited here are derived from earlier wood anatomical surveys. The 1. Ontogenetic and Histological Criteria; Allied Phenomena. Because secondary xylem consists mostly of cells with rigid walls, it is a clear and Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM FIG. 1. Transections of stem of Turbina stenosiphon (Convolvulaceae), to show diverse types of vascular histology. A–B. Successive cambia. A. Three vascular increments, each with secondary xylem (sx) and secondary phloem (sp); the middle vascular increment has an inverted orientation (ivi), atypical for successive cambia. B. Higher power, area corresponding to center of A. The inverted increment (ivi) above has produced secondary phloem adaxially and secondary xylem abaxially; the crushed secondary phloem (csp) was produced by the inverted increment. The normal increment below has produced secondary phloem abaxially and secondary xylem adaxially. C. Strand of interxylary phloem, surrounded by fibrous secondary xylem. D. Intraxylary phloem strand (itp) adaxial to protoxylem (upper left). A cambium (arrows) has developed in the strand and has produced secondary phloem (sp) adaxially. Pith parenchyma, some of which has been converted to pith sclereids (ps) below. easily read record of the action of the vascular cambium. The products of cambial action are unambiguous, so that there is no need to develop definitions that exclude ontogenetic aspects and are merely topographic in their frame of reference (e.g., included phloem). To be sure, xylarium specimens have poor preservation of meristematic cells and of phloem, but the location and sequences of cell types with secondary walls (which are readily seen in sections of xylarium specimens) are quite sufficient to permit any appearance to be referred to one of the categories accepted here. The difference between interxylary phloem and successive cambia can be seen in Turbina stenosiphon (Fig. 1A–C) of the Convolvula- Author's personal copy BRITTONIA [VOL FIG. 2. Sections of rayless stems containing interxylary phloem. A–C. Thunbergia laurifolia (Acanthaceae). A–B. Transections. A. Low power to show secondary phloem (sp) above), and secondary xylem below, with the vascular cambium (vc) between them. The secondary xylem is composed of an axial parenchyma (pax: gray) background in which bands or strands of fibrous vessel-bearing xylem (fx: darker) are embedded. B. Higher power photograph to show the strands of sieve tubes (st) in the axial parenchyma backgrounds. Fibrous xylem may deiverge from each other (di) by means of parenchyma or may actually break apart (br) due to tensions during growth. C. Radial section to show sieve plates (sp) in sieve tubes (darker gray), sheathed by parenchyma of the interxylary strand (pix, lighter gray); the fibrous xylem cells (fx) are narrower and stain more darkly. Stylidium glandulosum (Stylidiaceae), transection of stem portion with secondary growth. The vascular cambium (vc) is essentially unifacial, producing secondary xylem externally but nothing more than perhaps a single layer of secondary cortex externally; a phellogen (pg) has developed in the innermost cortical cells. Seven arrows indicate strands of interxylary phloem, which are very narrow and consist of only two to four cells each. ceae. This species is the first, to my knowledge, in which both successive cambia (Fig. 1A–B) and interxylary phloem (Fig. 1C) have been shown to coexist in a single stem. Interestingly, a third phenomenon, intraxylary phloem, is also represented in Turbina stenosiphon (Fig. 1D). Successive cambia are increments of secondary xylem and phloem, each produced by a vascular cambium. The cambia, as well as conjunctive tissue and secondary cortex (0 parenchyma) are produced by a master cambium (Carlquist, 2007). Conjunctive tissue is not Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM FIG. 3. Transections of stem of Orphium frutescens (Gentianaceae). A. Low power photograph to illustrate that secondary phloem (sp) contains only a few strands of sieve tubes (st); the remainder is parenchyma. Vascular cambium (vc) at the juncture with secondary xylem. The secondary xylem consists of fibrous xylem (fx) in which strands or bands of interxylary phloem (ip) are located. B. Intermediate power photograph to illustrate that the interxylary phloem may take the form of strands (ip) or bands (ipb) located in a background of vessel-bearing fibrous xylem (fx). C–E. Higher power photographs to illustrate intersections between rays (r) and interxylary phloem (ip) in relation to the fibrous xylem (fx) background. C. Two strands of interxylary phloem are separated by a ray. D–E. Instances in which sieve-tube elements and companion cells are derived from ray initials, and in which phloem ray tissue therefore contains phloem. a form of axial parenchyma; axial parenchyma is produced only by a vascular cambium. The example illustrated here, Turbina stenosiphon, is unusual in having one vascular increment inverted in orientation, with that vascular cambium producing phloem internally and xylem externally. This is the only instance I know in which a vascular increment has this inverted orientation. This occurrence does underline the fact that occasionally one does see a cambial variant with atypical ontogeny. At the same time, the rarity of such occurrences reinforces the regularity of phenomena described by the definitions given: cambial variants are orderly, and not "anomalous." Author's personal copy BRITTONIA [VOL FIG. 4. Transections of stems of Onagraceae. A–B. Pseudolopezia longiflora. A. Tangentially wide interxylary phloem band (ipb) embedded in a background of starch-rich fibers (srf). B. Section showing secondary phloem (sp) in upper half, divided from the secondary xylem, below, by the vascular cambium (vc). Strands of sieve tubes (st) are scarce in the secondary phloem. Fibers of the secondary xylem are rich in starch (srf). C–D. Stem transections of Oenothera linifolia. C. First-year xylem: strands of interxylary phloem (ip) are relatively numerous, but short-lived; gaps appear in them due to collapse of sieve tubes; phloem parenchyma cells surround the gaps. D. Second-year xylem: strands of interxylary phloem (ip) are few but functional. The interxylary phloem strand illustrated in Turbina stenosiphon (Fig. 1C) exemplifies all of the features claimed by the definition. It is a strand of sieve tube elements and companion cells surrounded by a sheath of parenchyma. Because the parenchyma is formed by a vascular cambium, it can be termed axial parenchyma. Intraxylary phloem is also present in Turbina stenosiphon (Fig. 1D). Intraxylary phloem occurs at the adaxial tips of vascular bundles of a number of woody angiosperms (for a list, see Metcalfe and Chalk, 1983, appendix). Cambial activity is found in a minority of instances of intraxylary phloem, and there is no list of genera in which it is Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM FIG. 5. Stem sections of Craterisiphon scandens (Thymeleaceae). A. Low power photograph of wood transection to illustrate that earlier-formed secondary xylem is devoid of interxylary phloem, whereas more recently-formed wood contains bands of interxylary phloem (ipb). B. Intermediate power photograph of transection to illustrate the extent of two interxylary phloem bands (ixb). C. High-power wood transection photograph of an interxylary phloem strand and a ray (r); ap 0 axial parenchyma (related to vessel); cs 0 crystal sand; f 0 intrusive fiber in interxylary phloem; sp 0 sieve plate as seen in transection. D. Longisection of interxylary phloem strand: f 0 intrusive fibers; pix 0 parenchyma of interxylary phloem; sp 0 sieve plates (seen obliquely). present or absent. It is present in the strand illustrated in Fig. 1D. Where present, cambium in intraxylary phloem acts unidirectionally, producing secondary phloem adaxially rather than (as with an ordinary vascular cambium) abaxially. Secondary xylem is produced by this cambium only rarely, but has been figured in Operculina (Carlquist & Hanson 1991; Carlquist, 2012,). Bicollateral bundle is a term that denotes phloem at both the abaxial and adaxial surfaces of a bundle (as opposed to a collateral bundle, which has phloem only abaxially). Although the term does not spe- Author's personal copy BRITTONIA [VOL FIG. 6. Transections of stem of Salvadora persica (Salvadoraceae). A. Low power photograph to illustrate secondary phloem (top 2/5 of picture) and secondary xylem (sx), bottom 3/5 of photograph, indicating various quantities of parenchyma (pa) within the secondary xylem. B. High power photograph of a portion of the transection shown in A; secondary phloem (sp, above) contains a single strand of sieve tube elements (to right of the letters sp); below the vascular cambium (arrows) is a strand of fibrous xylem containing two vessels; the remainder of the secondary xylem consists of parenchyma (pa). C–E. High power photographs to show interxylary phloem. C. Juncture between secondary phloem (sp) and secondary xylem, with vascular cambium (arrow) between them. In the secondary phloem, a strand of sieve tubes is seen (left); in the secondary xylem, a ray (r) and a young strand of interxylary phloem (left of the letters ip) are embedded in axial secondary xylem that consists of parenchyma (pa), with no fibers present. D. A strand of interxylary phloem of intermediate age, farther from the vascular cambium; some of the phloem is crushed (cip); below that, phloem is function, and the formation of a cambium in the interxylary phloem strand is denoted by arrows. E. An older strand of interxylary phloem; all of the phloem is crushed (cip). cifically exclude the presence of secondary xylem in such a bundle, it is more commonly applied when there is little or no secondary growth in the bundle, as in Cucurbita or Solanum (Lycopersicon). Thus, there is an overlap with the term "intraxylary phloem." Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM FIG. 7. Stem transections to show interxylary phloem. A–B. Strychnos madagascariensis (Loganiaceae). Stem transections seen with SEM. A. Low power micrograph, to show a typical large interxylary phloem strand (ip) in a fibrous xylem (fx) background; bark (b) at top of photograph. B. High power photograph corresponding to lower left portion of the strand shown in A; a cambium (c, plus arrows) has formed within the interxylary phloem strand; cpa 0 crushed parenchyma, fx 0 fibrous xylem; np 0 newer phloem; op 0 older phloem. C–D. Light photomicrographs of stem transections of Combretum erythrophyllum (Combretaceae). C. An interxylary phloem strand near the vascular cambium (which is not shown, but is just above the top of the photograph); a cambium (c, plus arrow) has recently formed within the interxylary phloem strand, but there is no crushed phloem; cpa 0 crystal-bearing parenchyma; fp 0 functional phloem; fx 0 fibrous secondary xylem. D. An older strand of interxylary phloem, in which a cambium (c, plus arrow) has been active; crushed phloem (cp) forms a conspicuous band; cpa 0 crystal-bearing parenchyma; fp 0 functional phloem; fx 0 fibrous xylem. 2. Parenchyma Associated with Strands of Interxylary Phloem. Strands of interxylary phloem are most commonly strands of axial parenchyma in Author's personal copy BRITTONIA which there is a central core of sieve tube elements and companion cells. This is most easily seen in Thunbergia (Fig. 2A–C). Thunbergia is a vine and the diameter of the sieve tubes is larger than in the average nonvining eudicot. In longisections, one can see sieve plates in the strands of interxylary phloem (Fig. 2C). The fibrous xylem consists of nonseptate libriform fibers (Fig. 2C, fx). Thunbergia is rayless, so all of the thin-walled parenchyma seen in Fig. 2A–C is axial parenchyma. Parenchyma associated with interxylary phloem is transversely subdivided into strands, much like axial parenchyma in typically woody eudicots. Axial parenchyma in Thunbergia separates bands and strands of vessel-containing fibers (Fig. 2A, B), which occasionally break (Fig. 2C, br) in response to stem growth and torsion. Axial parenchyma in Thunbergia thus serves a mechanical function that can often be served by wide rays in scandent woody plants. Stylidium glandulosum (Fig. 2D), a subshrub, also has rayless wood. The strands of interxylary phloem (arrows) consist of little more than a sieve tube element plus a companion cell each, and are quite inconspicuous. An occasional axial parenchyma cell is present in these strands. Stylidium and Thunbergia form extremes with respect to quantity of parenchyma associated with phloem. Intermediate quantities of parenchyma characterize the strands of most species that have interxylary phloem (Figs. 3A–E, 4A and 6C–E). The varied quantities of parenchyma observed in strands of interxylary phloem may be keyed to diverse physiological functions, but there has been no experimental work on this topic. 3. Rays and Interxylary Phloem. By implication, interxylary phloem occurs as vertical strands in axial xylem. This is clearly demonstrated by most species (e.g., Thunbergia, Fig. 2A–C). In most species that have interxylary phloem, rays either do not cross strands of interxylary phloem (Fig. 6A– E) or if they do (Fig. 5A–B), The ray cells retain their typical histological characteristics. Orphium (Fig. 3) provides some examples of this latter condition, but it also, in a few places, forms sieve tube elements and companion cells in rays. In Fig. 3C, instances of typical interxylary phloem strands that do not intersect rays are shown. [VOL In Fig. 3D and E, instances in which portions of rays have been converted to sieve tube elements and companion cells are shown (r 0 rays). One should note that ray cells in Orphium are predominantly upright, as one would expect from a secondarily woody plant (Carlquist, 1962, 2009), so conversion of ray cells to sieve tube elements and companion cells is really not contrary to the expected direction of conduction. If one scans larger areas of Orphium wood transections, one sees that interxylary phloem occurs as either strands or bands (Fig. 3A, B). The ratio of bands that contain sieve tube elements and companion cells in ray areas to those that do not is perhaps only one band out of twenty. 4. Diversity in Patterns: Strands, Bands, Relative Abundance. Interxylary phloem is usually seen as cylindrical strands (Figs. 1C and 2B). These strands can range from inconspicuous and few celled, as in Stylidium, to large and obvious, even perceptible without microscopy, as in Strychnos (Fig. 7A) and Combretum (Fig. 7C, D). Interxylary phloem is often present in the form of tangential bands (Figs. 4A and 5A. Bands and strands may occur together. Salvadora (Fig. 6) appears to have bands rather than cylindrical strands of interxylary phloem (e.g., Fig. 6C) because the parenchyma surrounding the strands occurs as tangential bands (Fig. 6A, B). Salvadora exemplifies the point that parenchyma may be much more abundant in transectional area than the strands of phloem in embedded in the parenchyma. 5. Cell Contents: Crystals, Starch, etc. Crystals occur in parenchyma that sheathes phloem in a large number of the species that have interxylary phloem. Exceptions can be cited in the case of Stylidium (Fig. 2D) Orphium (Fig. 3) and Salvadora (Fig. 6). Although not shown here, prismatic crystals occur in the axial parenchyma bands that contain phloem in stems of Thunbergia alata Bojer ex Sims (Carlquist and Zona, 1988; Carlquist, 2001). Raphides are present in such parenchyma in Onagraceae (Carlquist, 1975). Crystal sand is present in axial parenchyma of interxylary phloem strands in Craterosiphon (Fig. 5C; SEM photos in Carlquist, 2001). Druses occur in the phloem-ensheathing paren- Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM chyma of Combretum (Fig. 7C–D, cpa). The positioning of druses in interxylary phloemadjacent parenchyma in Combretum as well as in cases of crystal occurrence in other genera listed in the present study suggests that crystal-bearing sheaths may deter predation of interxylary phloem by chewing beetles. Such positioning of crystals is often seen in relation to phloem in bark of many woody species. In woods of Onagraceae, starch is common in libriform fibers adjacent to strands of interxylary phloem (Fig. 4A–B, srf; Carlquist, 1975). However, starch is notably absent in the parenchyma sheathing the phloem strands in that family (Fig. 4A–B). This circumstance suggests that starch storage and active transport of soluble photosynthates are distinct functions performed by these two respective tissues. 6. Libriform Cells in Interxylary Phloem. Because the root word "liber" refers to phloem, sieve tube elements (and their associated companion cells) could be included as "libriform elements" but that is not usually done. "Libriform" implies an elongated form, and usually refers to fibers. Certainly sievetube elements in Craterisiphon are elongate, their length easily determined from presence of sieve plates (Fig. 5D). Curiously, however, extraxylary fibers mature in the interxylary phloem bands of Craterisiphon (Fig. 5B, C). These fibers are gelatinous, and in permanent slides, the secondary walls shrink away from the primary walls (Fig. 5C). The fibers are instrusive, and their tips (Fig. 5D, f) do not align with the sieve plates of the sieve tube elements, which are shorter than the fibers. 7. Timing of Interxylary Phloem Onset. Presence of interxylary phloem may change in abundance with age of stem. This is evident in Fig. 5A for Craterisiphon. In this stem, interxylary phloem is absent in earlier-formed secondary xylem. This has been reported in other species, such as Azima tetracantha Lam. of the Salvadoraceae. Den Outer and van Veenendaal (1995) describe interxylary phloem strands throughout the stem of this species. Their stem was larger in diameter than the one I studied (Carlquist, 2002); at the periphery of the stem I studied, interxylary phloem production had just begun. In other Salvadoraceae, such as Dobera (Carlquist, 2002) and Salvadora (Fig. 6), interxylary phloem production begins early and remains constant in abundance. Mennega (1980) in her study of wood of Strychnos and other Loganiaceae mentions that interxylary phloem has not been reported in some species (cf. Pfeiffer, 1926) in which only small diameter stems were available, whereas larger-diameter stems prove to have interxylary phloem. An apparent exception to this trend occurs in Oenothera linifolia, in which interxylary phloem strands are fewer and smaller in second-year wood of stems (Fig. 4D) as compared to first-year wood (Fig. 4C). The occurrence of this trend is somewhat masked by the fact that in firstyear wood, parenchyma cells of the strands enlarge and develop secondary walls after the collapse of sieve tube elements and companion cells. Pfeiffer (1926), following Schenck (1895) reports sieve tube elements in secondary xylem of Mucuna altissima DC. as a late or subsequent (nachträglich) development compared with maturation of other cell types nearby. 8. Comparison between Interxylary Phloem and Secondary Phloem in a Single Stem. Comparisons of this sort are lacking, presumably because dried material rather than liquid-preserved material has been studied, and because soft tissues do not survive sliding microtome sectioning as well as harder (fibrous) tissues. The question that arises in connection with comparison of these two phloem regions within a given stem is whether in stems that have interxylary phloem, sieve tube elements are less abundant in secondary phloem (outside the vascular cambium) than in stems that lack interxylary phloem. The answer to this question, based on a small sample, is yes, although variability is evident. Whether bark thickness relates to distribution of sieve tube elements in interxylary phloem vs. secondary (0 external, or bark) secondary phloem is not known, and needs investigation. The most notable example is found in Stylidium species that have secondary growth (most species of the genus lack secondary growth). Stylidium glandulosum (Fig. 2D) Author's personal copy BRITTONIA produces at most one layer of cells external to the vascular cambium, and the cells of that layer are best described as parenchyma. These cells are not radially aligned with the innermost cortical parenchyma layer, in which phellogen develops (Fig. 2D, pg). Thus, there is no true secondary phloem, and no sieve tubes external to the cambium. This was reported, although not thoroughly illustrated, earlier (Carlquist, 1981). Orphium (Fig. 3A) and Pseudolopezia (Fig. 4B) show a relative paucity of sieve tubes and companion cells in secondary phloem. Relatively extensive areas of secondary phloem consist exclusively of phloem parenchyma, with only a few, isolated strands of sieve tube elements and companion cells present. Some degree of variability occurs in Salvadora (Fig. 6). In some areas of secondary phloem, sieve tube elements and companion cells are relatively sparse (Fig. 6A–B), whereas in others, they are rather more common (Fig. 6C). As a generalization, however, Salvadora—as well as the other interxylary-phloembearing eudicots for which data are reviewed here—have fewer sieve tube elements and companion cells in secondary phloem (bark) than is typically observed in eudicots that lack interxylary phloem. 9. Organographic Distribution. Work in wood anatomy remains biased toward stems, for understandable reasons. In fact, so few xylarium specimens are of root material that no indication of site of origin on the plant is given on most specimens; the default assumption is that stem material is involved. (Whether the material comes from main stem or branches is likewise never indicated on xylarium labels). Some workers have mentioned interxylary phloem in roots or rhizomes. For example, Pfeiffer (1926) reports interxylary phloem "islands" (interxylären Inseln) in transections of roots and rhizomes of Cochlearia armoracia L., Brassica napus L., B. rapa L., and Raphanus sativus L. Metcalfe and Chalk (1950), citing Pfeiffer's (1926) reports, mentioned "secondary interxylary bundles" in the unlignified xylem of the rhizomes of Armoracia lapathifolia Gileb. and in the root of Brassica napus, B. rapa, and Raphanus sativus. The overlapping nature of these reports [VOL and the impreciseness of the term "secondary interxylary bundles" underlines the need for more extensive studies not merely to confirm the histological nature of these instances, but to determine the ontogenetic origin of these "bundles" or "islands." The lack of reports of such appearances in the stems of Brassicaceae is, however, notable in this regard. Pfeiffer (1926) assigned histological appearances in the roots of Scolopia atropoides Bercht. & Presl (Solanaceae) to the concept of interxylary phloem. He likewise referred similar vascular tissue in roots of Browallia viscosa HBK. (Solanaceae) to this category. Pfeiffer (1926) also reproduces a believable drawing of interxylary phloem in thin-walled root secondary xylem of Atropa belladonna L. (Solanaceae) by Leisering (1899), so there is reason to credit his concept of interxylary phloem in that species as the same as mine. Stem interxylary phloem has not been reported in Atropa. Pfeiffer (1926) reports interxylary phloem for roots of Ipomoea versicolor Meissn. (Convolvulaceae), but does not note it in stems of this species. There are obviously many residual opportunities for confirmation or reassignment into other catergories of interxylary phloem reports. New discoveries remain to be made, as in Turbina stenosiphon (Fig. 1C). The material available to earlier workers was limited, and often was biased in favor of plants which were naturally-occurring or grown in Europe, and in favor of stems rather than roots of those. 10. Cambial Activity within Interxylary Phloem. Scott and Brebner (1889) described development of cambial activity in the interxylary phloem strands of Strychnos, based upon living material cultivated in greenhouses. Scott and Brebner figured large strands of interxylary phloem much like the one figured here (Fig. 7A, B). They reported crushed phloem on the abaxial side of the strands with cambial activity, so that the cambia in these strands produces secondary phloem externally, thereby in the same direction as the vascular cambium. This activity agrees with the findings reported here (Fig. 7A, B). Despite the disadvantage of working with herbarium material, Mennega (1980) reported the above facts accurately in a survey of Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM woods of Strychnos (and other Loganiaceae). As Mennega (1980) stated, not all species of Strychnos have interxylary phloem in the stems, even when large-diameter stems are examined. A study devoted to one species, S. millepunctata Leeuwenberg (van Veenendaal & den Outer, 1993) includes some excellent SEM images that reinforce the findings of Scott and Brebner (1889). Note should be taken that cambial activity in the interxylary phloem strand begins soon after a strand is produced by the vascular cambium. Cambial action is evident from the radial seriation of the cells produced by the interxylary phloem cambium (Fig. 7B, np). The earlier-formed interxylary phloem cells may not show radial seriation (Fig. 7B, op). The amount of secondary phloem produced by cambial activity within a strand is evident from the quantity of radially seriate phloem cells in the phloem as well as the amount of crushed phloem on the abaxial side of the strand. Bark of Strychnos (Fig. 7A, top) is relatively poor in sieve tube element production. Large interxylary phloem strands occur in the African species of Combretum of the Combretaceae (van Vliet, 1979), but not all of them. Histological and ontogenetic details given to date are relatively few because most specimens studied are from herbarium material or xylarium blocks. Somewhat thick sliding microtome sections of liquid-preserved material presented here (Fig. 7C, D) illustrate that Combretum interxylary phloem strands are histologically similar to those in Strychnos and like them in the timing of cambial initiation within the strand. Strands close to the vascular cambium (Fig. 7C) already show the beginning of cambial activity on the adaxial side of the strand. Older strands (Fig. 7D) show crushed phloem (cp) conspicuously, and a continuation of cambial activity within the strands. Interxylary phloem strands in Combretum are composed of fibriform (Fig. 7C, fp) cells (when seen in longisection) and crystalliferous parenchyma cells (cpa) that contain druses. Cambial activity is reported here in the interxylary phloem strands of one other family, Salvadoraceae, although attention has not hitherto been called to this phenomenon (Carlquist, 2002), presumably because it is so inconspicuous. In young strands of interxylary phloem of Salvadora persica (Fig. 6C, bottom, ip) there are only sieve tube elements and companion cells that have been derived from the vascular cambium. Crushed phloem and cambial activity within younger strands is not observed, contrary to the conditions in Combretum and Strychnos. In moderately old interxylary phloem strands of Salvadora, no radial seriation of cells is evident (Fig. 6D). Cambial activity (arrows) is minimal. Some collapsed phloem (Fig. 6D, cip) is evident, but the accumulation is not prominent. Earlier formed (older) interxylary phloem strands show a greater amount of crushed phloem (Fig. 6E). There is a correlation between size of interxylary phloem strands and presence of cambial action within these strands, when one compares the interxylary phloem of Combretaceae, Loganiaceae, and Salvadoraceae to that of other families. These families are woody, ranging from shrubs to trees, and preservation of phloic pathways by means of active replacement of sieve tube elements and companion cells by cambial activity within the strands seems a strategy that is correlated with habit. 11. Relationship between Intraxylary Phloem and Interxylary Phloem. The present study endorses the term intraxylary phloem to refer to phloem strands that occur adjacent to protoxylem, at margins of the pith. This term does not equate entirely to the term "bicollateral bundle" (see “Aspects of Interxylary Phloem”, section 1), in which minimal accumulation of secondary xylem is implied. Approximately equal amounts of phloem are seen external and internal to the xylem in species with bicollateral bundles, whereas in instances referred to the concept of intraxylary phloem, the amount of phloem formed externally from the vascular cambium can be relatively large, whereas the intraxylary phloem strands are relatively finite in size. Because of the uneasy coexistence of these terms, listings of families that exemplify one or the other concept have not always been assembled based on critical review of material. In addition, misapplication of these terms creates problems. Pfeiffer (1926) cited instances of phloic strands in the pith, for example. One can, however, cite particular families and genera in which intraxylary phloem is characteristically present. Metcalfe and Chalk Author's personal copy BRITTONIA (appendices, 1950, 1983) presented such lists. In their listings, Metcalfe and Chalk distinguished between families that characteristically have intraxylary phloem (bold face) and those in which intraxylary phloem is occasionally reported (ordinary font) or infrequent or perhaps dubious (italics). The families of Myrtales figure prominently in the list (Combretaceae, Crypteroniaceae, Lythraceae, Melastomataceae, Myrtaceae, Oliniaceae, Onagraceae, Penaeaceae, and Punicaceae). Interestingly, interxylary phloem also occurs in an appreciable number of species in three of these families (Combretaceae, Melastomataceae, and Onagraceae). A similar link between intraxylary phloem presence and interxylary phloem occurrence can be cited for other families in the Metcalfe and Chalk (1950) list (Gentianaceae, Loganiaceae, Stylidiaceae, and Thymeleaceae) as well as Leptadenia, an asclepioid genus of Apocynaceae (Singh, 1943; Patil & Rajput, 2008). Thus, intraxylary phloem may be a kind of "precursor" for interxylary phloem formation in a given species. In developmental terms, the genetic information for the formation of strands of phloem within the xylem (interxylary) as well as internal to (adaxial to) the xylem (intraxylary) may be similar. Exceptions to this concept can certainly be listed (e.g., Salvadoraceae lack intraxylary phloem), and this theory may apply only in particular clades. The physiological implications of this connection between the two sites of phloem formation are, however, even more interesting. In, say, Myrtales, why do only some of the species that have intraxylary phloem go on to produce interxylary phloem? One of the most interesting aspects of interxylary phloem is the development of a cambium in intraxylary phloem in some instances. This is illustrated for Turbina stenosiphon (Fig. 1D), but occurs in other eudicots, such as Cucurbitaceae (Carlquist, 1992; Patil et al., 2011). When cambium develops within a strand of intraxylary phloem, the secondary phloem it yields is always produced toward the center of the stem, rather than toward the outside (the latter, of course, is what happens in the formation of bark by the vascular cambium). The inverted nature of the secondary phloem production by cambia at intraxylary phloem [VOL sites is also indicated by the fact that in a few species, the intraxylary phloem cambium also produces some secondary xylem (in an external, or abaxial, direction). This has been illustrated for Operculina palmeri (Wats.) Howe of the Convolvulaceae (Carlquist & Hanson 1991; Carlquist 2012,). 12. Systematic Occurrence of Interxylary Phloem. As noted above, understanding the systematic occurrence of interxylary phloem is a work in progress. Several reports must be regarded as tentative, while others are likely incorrect. The latter are difficult to prove definitively, because interxylary phloem may occur infrequently in a few species. The late onset of interxylary phloem production, mentioned for Azima and Strychnos, is another reason to be cautious where lists are concerned. The multiplicity of individuals who report instances of interxylary phloem results in variable criteria and thus lack of precision in application of the concept. The following list contains instances that appear well substantiated on the basis of supporting drawings or photographs. Earlier workers occasionally conflated interxylary phloem (formed from a single vascular cambium) with instances of successive cambia under the inclusive rubric "included phloem." That vague umbrella usage was followed by IAWA Committee (1989). Instances of successive cambia are not included in this list. The listing is similar to that presented earlier (Carlquist, 2001), but with some emendations. Following this list, a compilation of dubious, incorrect, or unusual instances that do not conform to the working definition of interxylary phloem. Apocynaceae (including Aslepiadaceae): Asclepias, Ceropegia, Leptadenia (Singh, 1943; Patil & Rajput, 2008). Brassicaceae: roots and rhizomes of Brassica spp., Cochlearia, and Raphanus (Pfeiffer, 1926). Combretaceae: Calycopteris, Combretum, Guiera, Thiloa (van Vliet, 1979; den Outer & van Veenendaal, 1995; Rajput et al., 2009). Convolvulaceae: Ipomoea versicolor Meissn. roots and hypocotyl (Scott, 1891); Turbina stenosiphon (infrequent; new report, above). Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM Fabaceae: Mucuna altissima DC. (Schenck 1893). Gentianaceae: Crawfordia, Chiroma, etc. (Pfeiffer, 1926); Ixanthus (Carlquist, 1984); roots of some other genera (Pfeiffer, 1926). Icacinaceae: Chlamydocarya, Sarcostigma, etc. (Lens et al., 2008). Lythraceae: roots of Lythrum salicaria L. (Gin, 1909). Malpighiaceae: Dicella, Stigmaphyllon, Tetrapteris (Pfeiffer, 1926). Melastomataceae: six genera (Chalk & Chattaway, 1937; Metcalfe & Chalk, 1950). Onagraceae: at least seven genera (Carlquist, 1975, 1977, 1983, 1987). Salvadoraceae: all genera (den Outer & van Veenendaal, 1981; Carlquist, 2002). Solanaceae: roots and rhizomes of Atropa belladonna L.; roots of Datura stramonium L. and Scolopia sp. (Pfeiffer, 1926). Stylidiaceae: Stylidium (Carlquist, 1981). Thymeleaceae: Aquilaria and eight other genera (Pfeiffer, 1926; Solereder, 1908; Metcalfe & Chalk, 1950); Craterosiphon (above). Special cases: Coccinia (Cucurbitaceae) develops cambia adjacent to rays (Carlquist, 1992) or within axial parenchyma of secondary xylem (Patil et al., 2011). In both of these instances, these unusual cambia produce secondary phloem, but no secondary xylem. Because the secondary phloem in both instances lies within the confines of secondary xylem, the phloem produced by these cambia can be called interxylary phloem. The terminological choice by Patil et al. (2011) is therefore acceptable, but one should note that Coccinia represents an unusual instance. Excluded instances or dubious cases in need of re-examination: Acanthaceae: Barleria (Pfeiffer, 1926) Apocynaceae: Lyonsia, Mandevilla, and Parsonsia were cited by Pfeiffer (1926), but his definition of interxylary phloem was wider than mine and is not followed here. Asteraceae: Stoebe (Adamson, 1934). Bignoniaceae: Distictis, Haplolophium, and Pithococtenium (Pfeiffer, 1926). Convolvulaceae: Cuscuta (Pfeiffer, 1926) Clusiaceae: Endodesmia roots (Pfeiffer, 1926). Euphorbiaceae: Dalechampia (Pfeiffer, 1926) Loranthaceae: Nuytsia (original data; reported as a case of interxylary phloem by Pfeiffer, 1926; Nuytsia has successive cambia). Sapindaceae: Serjania (Pfeiffer, 1926). Urticaceae: Myriocarpa is cited by Chalk and Chattaway (1937) on the basis of large parenchyma strands within the secondary xylem. In fact, these strands, as they conceded, do not contain phloem. Rather, the parenchyma strands exemplify the phenomenon of fiber dimorphism (Carlquist, 1958, 1961). The occurrence of this kind of parenchyma in Urticaceae has been confirmed by Bonsen and ter Welle (1984). Study of liquid-preserved material is needed to resolve cases considered dubious here, because sieve tube elements do not survive drying very well. The erroneous report of Myriocarpa exemplifies this. Likewise, roots provide logistical problems for investigation. The few reports of interxylary phloem in roots (Weiss, 1880; Gin 1909; Solereder, 1908; Pfeiffer, 1926) are tantalizing because they suggest more instances might be found. 13. Physiological Significance. The study of interxylary phloem (as well as allied phenomena: intraxylary phloem, bicollateral bundles) is obviously still incomplete with respect to descriptive anatomy. The understanding of the physiological significance of these structural modes of phloem occurrence is a promising topic for exploration. Nevertheless, we can ask questions about function based on our present understanding of anatomy. Physiological studies, like anatomical studies, are most actively pursued in species of economic interest. In fact, none of the species known to have interxylary phloem is of any major economic importance. Living material of many of the species is not easy to access. Thus, progress in investigation of how interxylary phloem works has been slow. The topics listed below may be regarded as points for departure of physiological studies. (a) Conduction rather than storage. Onagraceae show that parenchyma associated with sieve tube elements and companion cells in interxylary phloem strands is deficient in starch, but tissues distal to the strands (mostly libriform fibers) are rich in starch (Carlquist, 1975). This clearly suggests a marked division of labor, in which interxylary phloem strands represent a conductive tissue, whereas the ground tissue of the secondary xylem is converted into a significant starch reservoir. Author's personal copy BRITTONIA Lack of starch in the parenchyma that sheathes sieve tube elements is evident in examples other than Onagraceae reviewed here: Acanthaceae (Fig. 2A–C), Stylidiaceae (Fig. 2D), Gentianaceae (Fig. 3), Thymeleaceae (Fig. 5) Salvadoraceae (Fig. 6), Loganiaceae (Fig. 7A–B) and Combretaceae (Fig. 7C–D) can be cited in this regard. (b) Conduction to large photosynthate sinks. A number of species with interxylary phloem strands have large fruit size (Strychnos: Leeuwenberg, 1980)) simultaneous production of large numbers of flowers and fruits (Oenothera) or other organographic features (sudden flushes of growth, Strychnos) that suggest a relationship between interxylary phloem and intense photosynthate utilization. Large inflorescences in which numerous flowers open at about the same time (Combretum) are pertinent in this regard. Instances of bicollateral bundles can be cited here. Large fruits in Cucurbitaceae and Solanaceae require rapid input of photosynthates that may be related to supernumerary phloem formations. (c) Enhanced rate of photosynthate conduction: the case of Orphium. This topic is allied to the above, but differs in stressing the rapid, simultaneous flowering of an entire plant. Orphium frutescens (Gentianaceae) has interxylary phloem. Orphium is a small shrub or subshrub that flowers during its first year of growth. During some subsequent year, flowering is so extensive and simultaneous that the plant devotes its entire reserves of photosynthates to the flowering/fruiting process and dies. At this point, it is a monocarpic plant, although one would not have designated it as such in prior years. I have cultivated Orphium frutescens in my garden and attempted to prolong the vegetative growth of a plant by removing all flowers during its summer flowering season. In the sixth year of growth, it produced only branches that terminated in flowers, with no side branches with vegetative buds. At that point, there was no longer any possibility of deterring flowering, and the plant flowered, fruited, and rapidly died after fruiting. An event of this sort seems correlated with the presence of interxylary phloem throughout the stem of Orphium. [VOL To be sure, there are many monocarpic plants that lack interxylary phloem. We do not know about their phloem abundance or phloem conductive patterns, because phloem of monocarpic plants has not been the subject of a study. One can, however, cite such plants as species of Oenothera (Onagraceae) that are biennials—in a sense, short-lived monocarpic plants. There is interxylary phloem in these. With relationship to Onagraceae as a whole, I suggested (Carlquist, 1975) that "Production of large flowers or large quantities of flowers during a short period might be related to massive starch reserves and interxylary phloem for rapid transport of sugars." In this regard, we may note that Fuchsia (Onagraceae), which does not have interxylary phloem, produces flowers slowly over a long period of time (sometimes throughout the entire year). (d) Phloem pathway multiplication and longevity. If interxylary phloem is produced continuously over a period of time, the aggregate quantity of sieve tube elements and companion cells in a stem (or root) soon exceeds the potential amount of phloem in bark. This is a feature relevant to conduction only if older interxylary phloem stays active. The occurrence of cambial activity producing new sieve tube elements and companion cells in interxylary phloem strands, as in Combretum, Salvadora, and Strychnos, attests to interxylary phloem longevity. We do not know, however, about the longevity of interxylary phloem strands in other species, such as the eight genera of Thymeleaceae (some trees: Aquilaria) that have interxylary phloem. We do know that secondary phloem is active in earlier increments of species with successive cambia (Carlquist, 2007). Anatomical studies show that each vascular cambium continues indefinitely to produce secondary phloem—eventually ceasing activity in older parts of larger stems. Reports of sustained longevity of secondary xylem, related to capability to reverse embolisms (e.g., Sperry, 1985), is indirect evidence of prolonged phloem function. Functioning of phloem without simultaneous functioning of adjacent vessels (or tracheids) is unlikely: The two are probably correlated (although studies of this are lacking). (e) Phloem pathway three-dimensionalization. Strands of interxylary phloem are an Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM ideal way of dispersing phloem throughout a stem or root as a means of aiding storage and retrieval of photosynthates. Validation of this speculation can be found in the examples, cited above for Brassicaceae and Solanaceae, in which interxylary phloem occurs in roots but is apparently absent in stems of particular species. Similar hypotheses were entertained with respect to successive cambia, which are also an ideal mechanism for distributing xylem and phloem throughout a stem or root, as in the beet, Beta (Carlquist, 2007). (f) Phloem pathway protection. Interxylary phloem strands are often ideally protected by their location within a fibrous background. That such fibrous backgrounds function in maintaining or prolonging safety of the strands has not been tested, although simple experiments incising bark to see whether interxylary phloem suffices for conductive needs would be easy to do. The evidence of placement and multiplicity suggests possible isolation from phytophagous insects and possibly other influences. The abundance of crystals in parenchyma sheaths of interxylary phloem strands in Acanthaceae (Thunbergia), Combretaceae (Combretum), Onagraceae (all genera with interxylary phloem), and Thymeleaceae (Craterosiphon) seems like an indirect evidence of predation deterrence. (g) Lianoid correlations and other habit considerations. The proportion of genera and species with interxylary phloem that has a lianoid habit is much higher than one would expect on the basis of the frequency of lianas and vines in eudicots as a whole. Families (and pertinent genera) in this regard include Acanthaceae (Thunbergia), asclepioid Apocynaceae (Asclepias, Ceropegia, Leptadenia), Combretaceae (several genera), Convolvulaceae (Ipomoea, Turbina), Cucurbitaceae (Cucurbita, Lagenaria), Icacinaceae (several genera), Malpighiaceae (several genera), and Thymeleaceae (Craterosiphon). Histologically similar phenomena (e.g., successive cambia) are also represented in a larger than expected number of lianoid genera (Carlquist, 1988, 2001, 2007). These constructions include intraxylary phloem, bicollateral bundles, successive cambia, and secondary xylem dispersed by parenchyma proliferation (e.g., Bauhinia, Mendoncia), as shown by the listings in appendices of Metcalfe and Chalk (1950, 1983). The parenchyma sheathing of phloem strands is conspicuous in many instances of interxylary phloem, such as Thunbergia (Acanthaceae) and Craterosiphon (Thymeleaceae) in the present study. These suggest enhanced flexibility, a feature ascribed to parenchyma of lianas by Schenck (1895). In these examples, interxylary phloem and parenchyma presence is less in earlier-formed wood, then increases with age, suggesting parenchyma becomes more important as self-support decreases and response to torsion and displacement of stems increases. A relatively small number of tree species have interxylary phloem, but there are some notable instances of interxylary phloem occurrence in tree Loganiaceae (Mennega, 1980) and Thymeleaceae (Pfeiffer, 1926). Examples should be examined on the basis of individual species, rather than strictly grouped according to habit. (h) Longevity and other physiological phenomena. Many fascinating questions regarding interxylary phloem occurrence remain to be asked and answered. Among these is the longevity of interxylary phloem. Circumstantial evidence may be obtainable from whether or not secondary xylem of a range of ages in a given stem is functional or not, but phloem itself tends to be good evidence, because it collapses so readily if it no longer functions (e.g., Salvadora, Fig. 6). Correlations between longevity of functioning in phloem and that in xylem vessels are to be expected. Phloem longevity is generally thought to be only a year or two, but greater longevity has been demonstrated in some angiosperms (Parthasarathy 1980). Acknowledgments For providing material, thanks are due Dr. David Lorence for stems of Strychnos madagascariensis (National Tropical Botanical Garden), Dr. Peter H. Raven of the Missouri Botanic Garden for stems of various Onagraceae, and the University of California of California at Santa Barbara living greenhouse collections for material of Salvadora. John Bleck provided seeds of Orphium frutescens from which my specimens were cultivated. Mark Olson and Edward L. Schneider provided Author's personal copy BRITTONIA helpful suggestions. Use of laboratory facilities, including the SEM, at Santa Barbara Botanic Garden, is gratefully acknowledged. Literature Cited Adamson, R. S. 1934. Anomalous secondary thickening in Compositae. Annals of Botany 48: 505–514. Bonsen, K. J. & B. J. H. ter Welle. 1984. Systematic wood anatomy of the Urticaceae. Botanisches Jarhbuch Systematik 105: 49–71. Carlquist, S. 1958. Wood anatomy of Heliantheae (Compositae). Tropical Woods 108: 1–30. ———. 1961. Comparative plant anatomy. Holt, Rinehart & Winston, New York. ———. 1962. A theory of paedomorphosis in dicotyledonous woods. Phytomorphology 12: 30–45. ———. 1975. Wood anatomy of Onagraceae, with notes on alternative modes of photosynthate movement in dicotyledon woods. Annals of the Missouri Botanical Garden 62: 386–424. ———. 1977. Wood anatomy of Onagraceae: additional species and concepts. Annals of the Missouri Botanical Garden 64: 627–637. ———. 1981. Types of cambial activity and wood anatomy in Stylidium (Stylidiaceae). American Journal of Botany 68: 778–785. ———. 1983. Wood anatomy of Onagraceae; Further species; root anatomy; significance of vestured pits and other structures in the dicotyledons. Annals of the Missouri Botanical Garden 69: 755–769. ———. 1984. Wood anatomy of some Gentianaceae: systematic and ecological conclusions. Aliso 120: 573–582. ———. 1987. Wood anatomy of noteworthy species of Ludwigia (Onagraceae) with relation to ecology and systematics. Annals of the Missouri Botanical Garden75: 889–896. ———. 1988. Comparative wood anatomy, ed. 1. Springer Verlag, Berlin & Heidelberg. ———. 1992. Wood anatomy of selected Cucurbitaceae and its relationship to habit and systematics. Nordic Journal of Botany 12: 347–355. ———. 2001. Comparative wood antomy, ed. 2. Springer Verlag, Berlin & Heidelberg. ———. 2002. Wood and bark anatomy of Salvadoraceae: ecology, relationships, histology of interxylary phloem. Journal of the Torrey Botanical Society 129: 10–20. ———. 2007. Successive cambia revisited: ontogeny, histology, diversity, and functional significance. Journal of the Torrey Botanical Society 134: 301–332. ———. 2009. Xylem heterochrony: an unappreciated key to angiosperm origins and diversification. Botanical Journal of the Linnean Society 161: 26–65. ———. 2012. How wood evolves: a new synthesis. Botany 90: 901–140. ——— & M. A. Hanson. 1991. Wood anatomy of Convolvulaceae: a survey. Aliso 13: 51–94. ——— & S. Zona. 1988 Wood anatomy of Acanthaceae: a survey. Aliso 12: 201–227. [VOL Chalk, L. & M. M. Chattaway. 1937. Identification of woods with included phloem. Tropical Woods 50: 1–31. den Outer, R. W. & W. L. M. van Veenendaal. 1995. Development of included phloem in the stem of Combretum nigricans (Combretaceae). IAWA Journal 16: 151–158. Gin, A. 1909. Réchérches sur les Lythracées, Travaux del la Laboratoire des Materiels Medicaux, Paris 6:1–166. IAWA Committee. 1989. List of microscopic features for hardwood identification. IAWA Bulletin, new series, 10: 219–882. Leeuwenberg, A. J. M., ed. 1980. Angiospermae: Ordnung Gentianales Fam. Loganiaceae. Die natürlichen Pflanzefamilien Band 28BI: 1–155. Ducker & Humblot, Berlin. Leisering, B. 1899. Über die Entwicklungeschichte des interxylären Leptoms bei den Dicotyledonen. Thesis, Berlin. Lens, F, J. Kårehed, P. Baas, S. Jansen, D. Rabaey, S. Huysmans, T. Hamann & E. Smets. 2008. The wood anatomy of the polyphyletic Icacinaceae s. l., and their relationships within asterids. Taxon 57: 525–552. Mennega, A. 1980. Anatomy of the secondary xylem. In A. J. M. Leeuwenberg, ed. Angiospermae: Ordnung Gentianales Fam. Loganiaceae. Die natürlichen Pflanzenfamilien Band 28BI. Duncker & Humblot, Berlin.(pp. 112—161. Metcalfe, C. R., & L. Chalk. 1983. Anatomy of the dicotyledons. Clarendon Press, Oxford. ——— & ———. 1983. Anatomy of the dicotyledons, ed. 2. Vol. 2. Wood structure and conclusion of the general introduction. Clarendon Press, Oxford. Parthasarathy, M. V. 1980. Mature phloem of perennial monocotyledons. Berichte der deutschen botanischen Gesellschaft 93: 57–70. Patil, V. S. & K. S. Rajput. 2008. Structure and development of inter- and intraxylary phloem in Leptadenia reticulata (Asclepiadaceae). Polish Botanical Journal 53: 5–13. Patil, V. S., C. R. Marcati & K. S. Rajput. 2011, Development of intra- and interxylary secondary phloem in Coccinia indica (Cucurbitaceae). IAWA Journal 32: 475–491. Pfeiffer, H. 1926. Das abnorme Dickenwachstum. Handbuch der Pflanzenanatomie 2 Abteilung 2 Teil, 9: 1–243. Gebrüder Borntraeger, Berlin. Rajput, K. S., V. S. Patil & K. S. Rao. 2009. Development of included phloem of Calycopteris floribunda Lamk. (Combretaceae). Journal of the Torrey Botanical Society 136: 302–312. Schenck, H. 1893. Beiträge zur Biologie und Anatomie der Lianen in Besonderen der in Brasilien einheimsche Arten. 2. Beiträge zur Anatomie der Lianen. In A. F. W. Schimper, ed. Botanische Mittheilungen der Tropens (pp. 1–271). G. Fischer, Jena. Scott, D. H. 1891. On some points in the anatomy of Ipomoea versicolor Meissn. Annals of Botany 5: 173–180. Scott, D. H. & G. Brebner. 1889. On the anatomy and histology of Strychnos. Annals of Botany 3: 275– 302. Singh, B. 1943. The origin and distribution of inter- and intraxylary phloem in Leptadenia. Proceedings: Plant Sciences 18: 14–19. Author's personal copy 2013] CARLQUIST: INTERXYLARY PHLOEM Solereder, H. 1908. Systematic anatomy of the dicotyledons (trans. Boodle & Fritsch). Clarendon Press, Oxford. Sperry, J. S. 1985. Xylem embolism in the palm Rhapis excelsa. IAWA Bulletin, new series 6: 283–292. Van Veenendaal, W. L. H., & R. W. den Outer. 1993. Development of included phloem and organiza- tion of the phloem network in the stem of Strychnos millepunctata (Loganiaceae). IAWA Journal 14: 253– 265. Van Vliet, G. J. C. M. 1979. Wood anatomy of the Combretaceae. Blumea 25: 141–223. Weiss, J. E. 1880. Anatomie und Physiologie der fleischig verdickter Wurzeln. Flora 70: 81–119.