Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Natural environment wikipedia , lookup
Introduction to evolution wikipedia , lookup
History of biology wikipedia , lookup
Cambrian explosion wikipedia , lookup
Paleontology wikipedia , lookup
Precambrian body plans wikipedia , lookup
Microbial cooperation wikipedia , lookup
Developmental biology wikipedia , lookup
Evolutionary developmental biology wikipedia , lookup
Palaeos:
CNIDARIA
CNIDARIA
CNIDARIA
Page Back
Unit Back
Metazoa
Metazoa Dendrogram
Metazoa References
Taxon Index
Pieces
Page Next
Unit Next
Unit Home
Unit Dendrogram
Unit References
Glossary
Time
Cnidaria
METAZOA
|--PORIFERA (paraphyletic?)
`--Radiata
|--Ctenophora
`--+--Cnidaria
| |--Anthozoa
| `--+--Hydrozoa
|
`--+--+--Scyphozoa
|
| `--Cubozoa
|
`--Conulata
`--Bilateria
|--DEUTEROSTOMIA
`--PROTOSTOMIA
|--Ecdysozoa
`--Lophotrochozoa
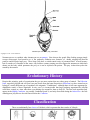
The Cnidaria
Symmetry
Body Form
The deadly Nematocysts
Evolutionary history
Classification
Links
Ptilosarcus gurneyi
A close-up of the Orange Sea Pen
Class Anthozoa - Subclass Octocorallia - Order Pennatulacea - Suborder Subselliflorae - Family Pennatulidae
image copyright © Keith Clements and Jon Gross Marine Life of the Northeast Pacific
Page Back
Page Top
Unit Home
Page Next
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
Text by M. Alan Kazlev (Creative Commons Attribution)
page uploaded 16 June 2002, last modified 2 May 2003
(originally uploaded on Kheper Site 3 June 1999)
checked ATW060205
Palaeos:
CNIDARIA
CNIDARIA
CNIDARIA
Page Back
Unit Back
Metazoa
Metazoa Dendrogram
Metazoa References
Taxon Index
Pieces
Page Next
Unit Next
Unit Home
Unit Dendrogram
Unit References
Glossary
Time
Cnidaria
Jellyfishes, Corals, Etc. -- Cambrian to Recent
METAZOA
|--PORIFERA (paraphyletic?)
`--Radiata
|--Ctenophora
`--+--Cnidaria
| |--Anthozoa
| `--+--Hydrozoa
|
`--+--+--Scyphozoa
|
| `--Cubozoa
|
`--Conulata
`--Bilateria
|--DEUTEROSTOMIA
`--PROTOSTOMIA
|--Ecdysozoa
`--Lophotrochozoa
The Cnidaria
Symmetry
Body Form
The deadly Nematocysts
Evolutionary history
Classification
Links
A zoanthid of the genus Acrozoanthus.
Image: Gary Cranitch, © 2008-2010 Australian Institute of Marine Science, Lizard Island 2010, Creative Commons
Attribution license
The Cnidaria
This diverse group of very simple-bodied animals includes corals, sea anemones, hydras, jellyfishes, and their
relatives. About 9,000 living species are known. The Cnidaria are the simplest Metazoa, and do not even possess
organs. All they have is a stomach and a mouth surrounded by tentacles.
Cubozoa
Scythozoa
Hydrozoa
Anthozoa
Symmetry
All cnidarians are radially symmetrical (the body is symmetrical around a central axis). As with the plant, there is no
front or rear, but there is a top and bottom.
Body Form
Polyp
anchored - mouth up
Medusa
free-floating - mouth down
There are two major body forms among the Cnidaria - the polyp and the medusa. Sea anemones and corals are typical
of the polyp form, while jellyfish are typical medusae When you look at it, you can see that these are the same body
form, except that one is upside down, while the other is not.
Lack of Organs
Cnidaria have no organs (i.e. groupings of different tissues to perform complex functions e.g. hearts, lungs, kidneys,
etc). There is a gastrovascular cavity ("stomach" in the broad sense) with a mouth but no anus, a body wall with outer
epidermis and inner gastrodermis, between which is a jelly-like mesogloea.
A mouth but no brain and no head
The nervous system is arranged as a decentralized network (‘nerve net’), with one or two nets present. There is no
brain or ganglia.
Note that although there is no head, there is a mouth, surrounded by a crown of tentacles. The tentacles are armed
with deadly nematocysts (stinging cells) .
A Diffusion based Physiology
All the metabolic functions of the body - respiration, digestion, elimination - are carried out by diffusion. Diffusion is
only an efficient means of exchange of materials only over short distances (e.g. over about 1 mm for oxygen
exchange). This means that all the tissues of a cnidarian which require oxygen (all its living cells) must be within
about 1 mm of a surface which is bathed by oxygenated water. This is why cnidarians are either very tiny (e.g. coral
polyps) or have very thin or flat bodies or long thin tentacles (both of which increase surface area). And also because
of this diffusion based physiology, cnidarians can only live in water (most species are marine, but some Hydrozoa live
in fresh water). There are only two surfaces in a cnidarian through which this diffusion can occur - the epidermis (the
outer layer or skin, which is bathed in water at all times) and the gastrodermis (the inner layer, representing the
stomach lining).
The deadly Nematocysts
Nematocyst with barb (top) extended
copyright © xxxx - source unknown
Nematocysts are to cnidaria what choanocysts are to sponges. But whereas the gentle filter-feeding sponge simply
sweeps microscopic food particles to it, the predatory cnidarian uses batteries of deadly stinging cells that can
paralyze and kill quite large prey. These large cells have a sealed central cavity of poisonous fluid. A small sensory
hair is sensitive to vibrations in the water. Any luckless creature passing too close triggers the nematocyst which then
shoots out the barb, which penetrates the prey so it can be injected with poison. The prey is then conveyed to the
mouth by the tentacle.
Evolutionary History
Despite this primitive grade of organization they are no more ancient than any other group of animal. The Edicarian
biota, supposed jellyfish and soft coral (sea pen) fossils from the latest Proterozoic (Ediacaran) era have turned out to
belong to a totally different type of organisms, the enigmatic "Vendobionta". although there is still some argument for
diploblastic natuer of these organisms. In any case it is inconceivable that large predatory organisms like jellyfish
could have existed at a time when there was nothing else around for them to feed on! The first coral organisms only
appear in the Middle Cambrian, (or possibly Cambrian Epoch 2), and only diversified during the Ordovician. the first
unambiguous jellyfish are also not known before the Cambrian.
Classification
There are traditionally four classes of Cnidaria, which represent the three modes of lifestyle:
Cubozoa
Scythozoa
Hydrozoa
Anthozoa
(medusa)
(medusa)
(polyp and medusa)
(polyps)
It is generally agreed that the Anthozoa are different from the others, but there has been disagreement for over a
century on whether they branch first or last. The majority of recent work finds that they branched early, and we
follow that trend.
References and Links
Cnidaria (Coelenterata)
Cnidaria
Cnidaria
The Cnidaria Home Page - links to material on the Cnidaria, arranged accordng to topic
Phylum Cnidaria - short intro and general information
Cnidaria - a good non-technical intro. Part of Keith Davey's Life on Australian Seashores website
Systema Naturae 2000 / Classification - Phylum Cnidaria
Cnidaria - basic outline
Page Back
Page Top
Unit Home
Page Next
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
Text by M. Alan Kazlev (Creative Commons Attribution)
page uploaded 16 June 2002, last modified 2 May 2003
(originally uploaded on Kheper Site 3 June 1999)
checked ATW060205
Palaeos:
SCYPHOZOA
CNIDARIA
SCYPHOZOA
Page Back
Unit Home
Page Next
Page Down
Cladogram
Class Scyphozoa
(Cambrian - Recent)
image © xxxx
Members of this class are large free swimming marine organisms. The medusa is the dominant phase in the life cycle
and the polyp stage is either absent or it is small and gives rise to medusae by asexual budding. There is a strong
four-fold symmetry.
<==o SCYPHOZOA
|--o STAUROMEDUSAE
| |-- Cleistocarpidae
| `-- Eleutherocarpidae
|--o CORONATAE
| |-- Atollidae
| |-- Atorellidae
| |-- Linuchidae
| |-- Nausithoidae
| |-- Paraphyllinidae
| |-- Periphyllidae
| `-- Tetraplatidae
|--o SEMAEOSTOMEAE
| |-- Cyaneidae
| |-- Pelagiidae
| `-- Ulmariidae
`--o RHIZOSTOMEAE
|-- Cassiopeidae
|-- Catostylidae
|-- Cepheidae
|-- Lobonematidae
|-- Lychnorhizidae
|-- Mastigiidae
|-- Rhizostomatidae
|-- Stomolophidae
|-- Thysanostomatidae
`-- Versurigidae
Cladogram Reference(s):
Parker, S. P. (ed.), 1982: Synopsis and classification of living organisms. Vols. 1 & 2 --McGrew-Hill Book Company
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
page uploaded 16 June 2002
checked ATW030411
(originally uploaded on Kheper Site 4 June 1999)
dendrogram by Mikko Haaramo
Palaeos:
CUBOZOA
CNIDARIA
CUBOZOA
Page Back
Page Up
Unit Home
Page Next
Page Down
Cladogram
Class Cubozoa
image © xxxx
Originally included as an order of the class Scyphozoa (Jellyfish) the Cubozoa are now considered to warrant
separate class status. As with the Scyphozoa the medusa is the dominant phase in the life cycle. They differ from
Scyphozoa in that they have a velum like structure, the velarium, the bell has four flattened sides and a simple margin,
and each polyp produces a single medusa by complete metamorphosis. The name refers to the cubic form of the
organism. They occur in tropical and subtropical oceans. There is only one order, the Cubomedusae
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
Text by M. Alan Kazlev (Creative Commons Attribution) 1999-2002
page uploaded 16 June 2002
(originally uploaded on Kheper Site 4 June 1999)
checked ATW050516
Parent: Cnidaria
Palaeos:
HYDROZOA
CNIDARIA
HYDROZOA
Sister: Anthozoa
Child: Siphonophora
Child: Hydroidea
Other Children:
See below
Dendrogram
Hydrozoa:
Hydroids and Hydromedusae.
From the Ordovician
Garveia annulata - Golden Hydroid
Class Hydrozoa - Order Hydroida - Family Bougainvilliidae
image copyright © Jon Gross Marine Life of the Northeast Pacific
This class includes hydroids and hydromedusae. $ Medusae small. $ Cnidocysts epidermal.
This very diverse class of cnidarians many of which have a true alternation of generation in its reproductive cycle.
That is, they passes through both the medusa and the polyp phase. Many groups secrete a chitinous skeleton in the
polyp stage. In some groups the polyp stage is colonial and secretes a calcareous skeleton, like a coral. There are
seven orders altogether, including:
Order
Order
Order
Order
Order
Order
Order
Order
Trachylina
Hydroida
Milliporina (Hydrocorallina)
Stylasterina
Siphonophora
Spongiomorphida (extinct)
Chondrophora
Actinulida
Trachylina - medusa stage only. These are perhaps the most primitive Hydrozoa. : Gonionemus
Hydra - a small simple freshwater Hydroid,
named after a mythical Greek monster
image copyright xxxx
Hydroida - Ordovician? to Recent - colonial attached polyps.
The free-floating medusa stage may be
present or absent. Includes most types of Hydrozoans. Freshwater or marine, solitary or colonial, softbodied or with skeleton. There are two suborders: Anthomedusae (Hydra, etc) and Leptomedusae. In the
latter the polyps always colonial. e.g. Obelia (left), Sertularia
Siphonophora - Large and complex swimming or floating colonies, with division of labour. Jellyfish-like,
but with a float or sail-like structure. Mainly tropical. Includes the unusual Portuguese Man-of-War (genus
Physalia).
Milleporina (should be Milleporida?) (Tertiary - Recent) - Reef builders, massive calcium carbonate skeleton. Marine.
Also called Stinging Coral and Fire Coral. The Milleporina and Stylasterina have in the past been included together
under the order Hydrocorallida
Stylasterina (should be Stylasterida?) (Tertiary - Recent) Reef builders, very like the Milleporina, but with a thick layer
of tissue overlying the skeleton
Stylaster venustus - Pink (purple) Hydrocoral
Class Hydrozoa - Order Stylasterina - Family Stylasteridae
image copyright © Jon Gross Marine Life of the Northeast Pacific
Spongiomorphida (Triassic - Jurassic). Massive colonies with radial pillars united by horizontal bars. Resemble
Stromatoporoids.
Chondrophora: Colonial or specialized polypoid individuals, e.g. Velella
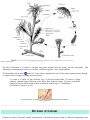
Alternation of Generations - Ontogenic Metamorphosis
image copyright xxxx
The above illustration is of Obelia, a Hydroid that passes through both the medusa and the polyp phase. This
organism goes through stages where it is in effect a different organism, a true metamorphosis .
The philosopher and inventor
Arthur M. Young observes that the life cycle of this simple organism passes through
a complete series of stages according to process theory:
"(1) starts as a single cell [the fertilized egg]; (2) becomes multicellular [3] acquires a shape
(identity) [planular stage]; (4)fastens to the ocean floor [hydroid colony]; (5) grows in plant-like
fashion; (6) flowers break off into mobile jellyfish (the animal stage); (7) fertilization."
[The Reflexive Universe, p.122]
an Obelia colony, uncannily resembling a branching plant with flowers
image copyright xxxx
Division of Labour
Hydrozoan colonies frequently exhibit polymorphism, with different polyps taking on different morphologies and
performing different functions (reproduction, feeding, etc.). Each colony is a super-organism, like an ant-hive.
Part of a colony of the marine hydrocorallid Millepora, showing division of labour.
image copyright xxxx
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
Text by M. Alan Kazlev (Creative Commons Attribution)
page uploaded 16 June 2002, last modified 2 May 2003
(originally uploaded on Kheper Site 4 June 1999)
checked ATW050611
Palaeos:
CNIDARIA
CNIDARIA
ANTHOZOA
Page Back
Unit Back
Metazoa
Metazoa Dendrogram
Metazoa References
Taxon Index
Pieces
Page Next
Unit Next
Unit Home
Unit Dendrogram
Unit References
Glossary
Time
Anthozoa
Corals, Sea anemones, Sea Pens, Etc.
CNIDARIA
|--Anthozoa
| |--Octocorallia (Alcyonaria)
| `--Hexacorallia (Zoantharia)
|
|--Actiniaria
|
`--+--Tabulata
|
`--+--Rugosa
|
`--Scleractinia
`--+--Hydrozoa
`--+--+--Scyphozoa
| `--Cubozoa
`--Conulata
The Cnidaria
Symmetry
Body Form
The deadly Nematocysts
Evolutionary history
Classification
Links
Cambrian to Recent
Epizoanthus scotinus
Class Anthozoa - Subclass Ceriantipatharia - Order Ceriantharia - Family Zoanthidae
image copyright © Keith Clements and Jon Gross Marine Life of the Northeast Pacific
Anthozoans include corals, sea anemones, sea pens, and related organisms. These animals are either solitary or
colonial polyps that live attached to a substrate (surface). This is the largest and ecologically the most important group
of cnidarians. Anthozoans are unique in their absence of a medusiod phase and polyps much larger than are found in
the other two classes. The polyp shows biradial symmetry, with the body cavity divided by septa There are 6,000
known recent species.
Phylogeny
<==o ANTHOZOA
|-- OCTOCORALLIA
`--o ZOANTHARIA [Hexacorallia]
|--+-- †KILBUCHOPHYLLIDA
| `--+?-o CERIANTHARIA
|
`--+--o CORALLIMORPHARIA
|
|--+?- Ptychodactiidae* [Ptychodactiatria]
|
| `-- ACTINIARIA (merivuokot)
|
`--+-- †NUMIDIAPHYLLIDA
|
`-- SCLERACTINIA [Madreporaria]
`--+-- †TABULOCONIDA
`--+-- †COTHONIIDA
`--+-- †TABULATA
`--+-- †HELIOLITIDA
`--+--+?- †HETEROCORALLIA
| `-- †RUGOSA
`--+?- ANTIPATHARIA
`-- ZOANTHIDEA
Cladogram Reference(s):
Conway Morris, S., 1993: The fossil record and early evolution of the Metazoa. --Nature, vol. 361, 21 January,pp.
219-225
--iNet: The Tree of Life
Fautin, D. G., Romano, S. L. & Oliver, W. A. Jr., 1999: Zoantharia - Sea Anemones and corals. --iNet: The Tree of
Life
Parker, S. P. (ed.), 1982: Synopsis and classification of living organisms. Vols. 1 & 2 --McGrew-Hill Book Company
Links
Introduction to the Anthozoa
Tree of Life - Anthozoa
Page Back
Page Top
Unit Home
Page Next
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
page uploaded 16 June 2002
checked ATW050516
(originally uploaded on Kheper Site 3 June 1999)
page by M. Alan Kazlev (Creative Commons License)
dendrogram by Mikko Haaramo
Palaeos:
ANTHOZOA
CNIDARIA
CERANTIPATHARIA
Page Back
Page Up
Unit Home
Page Next
Page Down
Cladogram
Subclass Cerantipatharia
Horny Corals
Pachycerianthus fimbriatus
Tube Anemone
Class Anthozoa - Subclass Cerantipatharia - Order Ceriantharia - Family Cerianthariidae
image copyright © Keith Clements and Jon Gross Marine Life of the Northeast Pacific
Included in this subclass are two superficially rather distinct forms. Alternatively they may be included with Rugose
and Hexacorals in the subclass Zoantharia. It is possible that the Ceriantipatharia is a polyphyletic taxon
Subclass Ceriantipatharia
Order Antipatharia: black corals
Order Ceriantharia: tube anemones
Order Cerantharia - anemone-like forms with elongate bodies adapted for burrowing - the so-called "burrowing
sea anemones". These are large solitary Anthozoa adapted for life in soft bottoms. The body is lodged with a secreted
mucous tube buried in sand or mud e.g.. Cerianthus (above)
Order Antipatharia: Black or Horny Corals. These are gorgonian-like upright plant-like forms. Polyps are
arranged around an axial skeleton of black horny material bearing thorns. Most forms are deep water and live in the
tropics.
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
Text by M. Alan Kazlev (Creative Commons Attribution) 1999-2002
page uploaded 16 June 2002
(originally uploaded on Kheper Site 4 June 1999)
checked ATW050516
Palaeos:
ANTHOZOA
CNIDARIA
HEXACORALLIA
Parent: Anthozoa
Sister: Octocorallia
Child: Actiniaria
Child: Tabulata
Hexacorallia or Zoantharia
This very important subclass includes six
mesenteries or multiples thereof. Included here
are sea anemones, true corals, and two similar
groups. Sometimes the subclass Zoantharia is
used instead, in which case hexacorals are
included with Rugose corals (Rugosa) and
Horny Corals (Cerantipitharia).
The following four orders have generally been
included:
Actinaria: Sea anemones
Scleractinia: True or stony corals
Zoanthidea: Colonial sea-anemone-like
animals
Corallimorpharia: Coral-like anemones
As of this revision (040930), the most recent
phylogeny is that of Daly et al. (2003).
Unfortunately, these workers were restricted to
living taxa, since they used a combined
molecular and morphological approach. Their phylogeny of the hexacorals may represented as follows:
Root
|--Ceriantharia
`--Hexacorallia
|--Actiniaria
`--+--Zoanthidea
`--+--Antipatharia
`--+--Corallimorpharia
`--Scleractinia
The Tabulata and Rugosa are usually assigned positions just basal to the Zoanthidea and Antipatharia. For lack of anything
more sensible to do, we insert them so as to preserve that relationship.
Root
|--Ceriantharia
`--Hexacorallia
|--Actiniaria
`--+--Tabulata
`--+--Rugosa
`--+--Zoanthidea
`--+--Antipatharia
`--+--Corallimorpharia
`--Scleractinia
The basic anatomy of a coral is shown in the image. Scleractinian larvae (i.e. the planula stage of polyp development) forms
a basal plate on a suitable substrate. It then secretes vertical partitions (septa) which are joined by the outer body wall
(theca). The septa may meet at the axis during at least part of their vertical extent. If so, they may form a vertical columella
in the center. This is common in scleractinian corals, but rare in rugose corals and absent in the Tabulata. In addition, the
polyp forms soft tissue partitions or mesenteries. Ultimately, these control the central pharynx which opens on the dorsal
surface. The opening of the pharynx is surrounded by a row of tentacles.
Image: from Coral Reefs Biology 200 Lecture Notes and Study Guide by Prof. David A. Krupp of Windward Community
College.
ATW040930. Text public domain. No rights reserved.
Actiniaria:
sea anemones
From the
Cambrian
Urticina piscivora: Fish-Eating
Urticina. Class Anthozoa Subclass Hexacorallia - Order
Actiniaria - Family Actiniidae.
Image copyright � Keith
Clements and Jon Gross
Marine Life of the Northeast
Pacific.
The soft-bodied sea anemones are large soft-bodied corals. The polyp is usually brightly colored, always solitary, and found
in coastal waters world-wide, but are particularly common in the tropics. In size they are usually from 1.5 to 5 cm in
diameter, although giant forms like Tealia columbiana (North Pacific USA) and Stoichactus sp. (Great Barrier Reef,
Australia) may reach over a meter at the oral (mouth) end.
The outer tentacles are much divided, giving a frilly appearance.
The mouth occurs in the middle of a flat area called the oral disc; the mouth and pharynx below are oval in shape. There are
usually two ciliated grooves called siphonoglyphs, found the length of the pharynx, that create a current of water into the
"stomach" (gastrovascular cavity), which maintains a hydrostatic skeleton, against which the muscles can act.
Zoanthidea
From the Triassic
These are small, solitary or colonial, anemone-like hexacorals. Most forms are tropical, some are common reef inhabitants,
and some live attached to other invertebrates. The body may be columnar, but is more often short and button-like. A short
fringe of tentacles surrounds a the broad oval disc.
Corallimorpharia:
False Corals,
Mushrooms, etc.
No fossil record -presumably evolved
before the Triassic
Amplexidiscus fenestrafer. From
CORALLIMORPHARIA (Ordre des Corallimorphaires)
These anthozoans resemble stony corals (Scleractinia) but lack skeletons. In fact, they are consistently classified as the sister
group of the Scleractinia by a wide variety of taxonomic methods. However, they secrete no skeleton of any kind. The
Corallimorpharia have a flattened adherent base, similar to Scleractinia, but without a base plate or basilar muscles. The theca
around the column is smooth, sometimes with weak longitudinal muscles. The sphincter is weak or absent. The tentacles
around the mouth are disposed in concentric circles, usually forming a series of radial lines rather than being alternately
arranged. Both the muscles of the oral disk and the longitudinal muscles of the tentacles are weak. However, the disk is
well-supplied with cilia which create a current carrying suspended materials into the pharynx. The polyps can also form the
dorsal surface into a sac, lined with nematocysts to trap and subdue larger prey. The mesenteries are often irregular.
Corallimorpharia are often solitary, or connected to other polyps by coenenchyme.
Corallimorpharia are well known to aquarists because of their striking forms and colors, the latter often due to symbiotic
zooxanthellae within the tissues of the polyp. In spite of this trait, Corallimorpharia generally prefer low light levels or
indirect light. They feed on a wide variety of food materials. All feed on dissolved and particulate matter, some feed on
zooplankton, and some are even known to trap and digest fish. Corallimorpharians usually reproduce vegetatively (e.g., by
budding or fission), and often form colonies of cloned individuals. Sexual reproduction is assumed to occur, but has not been
well documented.
Currently, four families of Corallimorpharia are recognized:
Corallimorphidae
Discosomatidae
Ricordeidae
Sideractiidae
Corynactis: source unknown
Amplexidiscus: from
Corallimorphs
Ricordea: from Sanjay's
Reef Aquariums
Sideractis: from
Hexacorallians of the World
There does not appear to be any good indication of the relative phylogenetic position of these families.
Links:
classification_path_2.cfm-taxonname=Corallimorpharia,
http://web.nhm.ku.edu/inverts/carlgren_1949/corallimorpharia.html, Order Corallimorpharia (ERMS taxonomic hierarchy),
Aquarium Invertebrates, Corallimorpharia (looks like great site, but my Icelandic is a bit rusty ...).
ATW040930. Text public domain. No rights reserved.
Scleractinia
From the Triassic
All Mesozoic and Cenozoic reef-building corals are members of this order (sometimes, subclass).
Reef-building corals are also known as "hard corals" or "stony corals." They are similar to and
closely related to sea anemones but unlike those soft-bodied forms they secrete an aragonitic
(calcium carbonate) skeleton. It is this skeleton that forms coral reefs.
Some forms, like the Indo-Pacific Fungia, are solitary, with single polyps reaching 25 cm in
diameter. The majority however are colonial, with very small polyps (about 1 to 3 mm),
although the colony as a whole can grow very large.
As with all the Hexacorallia, septal insertion occurs in multiples of six, and many scleractinial
corals have 6-fold symmetry.
Over 60 genera of Scleractinia have a symbiotic relationship with a type of microscopic algae,
called zooxanthellae, living within the coral polyp's tissue (within the gastrodermal cells).
Although deep water and some cold-water corals lack zooxanthellae, virtually all reef-building
possess them. These corals are generally found in clear water at depths of less than 50 meters,
the zone where sunlight penetrates. The algae not only provide food but help polyp calcification, and may account for up to
50% of the protein nitrogen of the coral, as well as giving then a yellow-brown or dark brown colour. These so-called
hermatypic corals can lay down massive amounts of limestone in the photic zone of shallow tropic seas.
Image: Cross section through a coral organism
Unless otherwise attributed, text on this page may be used under the terms of a
Creative Commons License.
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
Text by M. Alan Kazlev (Creative Commons Attribution) 1999-2002
page uploaded 16 June 2002
(originally uploaded on Kheper Site 4 June 1999)
checked ATW050611
Palaeos:
ANTHOZOA
CNIDARIA
OCTOCORALLIA
Page Back
Page Up
Unit Home
Page Next
Page Down
Cladogram
Octocorallia
Calcigorgia spiculifera
Class Anthozoa - Subclass Octocorallia - Order Gorgonacea - Suborder Holaxonia - Family Acanthogorgiidae
image copyright © Keith Clements and Jon Gross Marine Life of the Northeast Pacific
In this group of mostly soft corals, the polyps are almost always formed into colonies, each polyp having eight
pinnate (side- branching) tentacles. Octocorallians include a number of diverse forms
Subclass Octocorallia: Octocorals. Eight mesenteries.
Order Gorgonacea: Sea fans, sea whips
Order Telestacea
Order Pennatulacea: Sea pens, sea pansies
Order Alcyonaea: Soft corals
Order Helioporacea
Order Stolonifera: Organ pipe corals (among others)
Alcyonacea - sometimes known as 'soft corals'. Often inhabit low tidal zone of rocky shores
Gorgonacea - members secrete a horny material called gorgonin to act as skeletal support. Abundant on coral reefs,
especially in the Caribbean.
<==o ALCYONARIA
|--o PROTOALCYONARIA
| |-- Haimeidae
| `-- Taiaroidae
|--o STOLONIFERA
| |-- Clavulariidae
| |-- Cornulariidae
| `-- Tubiporidae
|--o TELESTACEA
| |-- Coelogorgiidae
| |-- Pseudocladochonidae
| `-- Telestidae
|--o GASTRAXONACEA
| `-- Pseudogorgiidae*
|--o GORGONACEA
| |--o HOLAXONIA
| | |-- Acanthogorgiidae
| | |-- Ainigmaptilidae
| | |-- Chrysogorgiidae
| | |-- Ellisellidae
| | |-- Gorgoniidae
| | |-- Ifalukellidae
| | |-- Isididae
| | |-- Keroeididae
| | |-- Paramuriceidae
| | |-- Plexauridae
| | `-- Primnoidae
| `--o SCLERAXONIA
|
|-- Anthothelidae
|
|-- Briareidae
|
|-- Coralliidae
|
|-- Melithaeidae
|
|-- Paragorgiidae
|
|-- Parisididae
|
`-- Subergorgiidae
|--o ALCYONACEA
| |-- Alcyoniidae
| |-- Astrospiculariidae
| |-- Maasellidae
| |-- Nephtheidae
| |-- Siphonogorgiidae
| `-- Xeniidae
|?- PENNATULACEA (merisulat)
`--o HELIOPORACEA
|-- Helioporidae
`-- Lithotelestidae
Pennatulacea - the sea pens, have a single long main polyp forms a stalk which anchors the colony in its mud
substrate. Branches from this bear the feeding and ciliated polyps and generate an inward current of water that keeps
the whole colony inflated. It used to be thought that some Edicarian forms were sea-pens, this does not seem to be
the case.
See also Race Rocks - Ptilosarcus gurneyi - for more on the Sea Pens.
<==o PENNATULACEA
|--o SESSILIFLORAE
| |-- Anthoptilidae
| |-- Chunellidae
| |-- Echinoptilidae
| |-- Funiculinidae
| |-- Kophobelemnidae
| |-- Protoptilidae
| |-- Renillidae
| |-- Scleroptilidae
| |-- Stachyptilidae
| |-- Umbellulidae
| `-- Veretillidae
`--o SUBSELLIFLORAE
|-- Pennatulidae
|-- Pteroeididae
`-- Virgulariidae
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
page uploaded 16 June 2002, last modified 2 May 2003
(originally uploaded on Kheper Site 3 June 1999)
checked ATW050611
page by M. Alan Kazlev (Creative Commons License)
cladograms © Mikko Haaramo
this material may be freely used for non-commercial purposes
Palaeos:
ANTHOZOA
CNIDARIA
RUGOSA
Parent: Anthozoa
Rugosa
Horn Corals
Middle Ordovician to Late Permian
Heliophyllum halli
Devonian period
image © xxxx
The Rugosa or "rugose corals" (referring to their wrinkled appearance), also known as "horn corals" were an
important group of Paleozoic organisms. Both solitary and colonial forms are known, but the former are more
common. Solitary rugosans usually have a horn shaped (hence the alternative term, "horn corals"), while the colonial
types commonly have hexagonal corallites. The skeleton is made of calcite and is generally quite massive. Solitary
rugose corals range in size from a few millimeters in diameter and in length to 14 centimeters in diameter and a height
of close to one meter. Some colonies may be 4 meters in diameter. Like modern corals (e.g. sea anemones, which
can be observed in intertidal rock pools), the coral animal (or polyp) fed by using tentacles to capture and sweep
organisms into their mouths
As a very general rule, rugose coral have stronger radial septa than they do transverse platforms. The septa radiate out
from the center. Rugose corals have both major and minor septa. Rugose corals differ from other corals by the
pattern by which they add septa through their ontogeny (development and growth). Both solitary and colonial
rugosans have this distinctive septal insertion pattern, which gives most rugose corals bilateral symmetry. The six
prosepta are added first, including the cardinal septum and counter septum, which are at 180 degrees from one
another. . After this the major septa (metasepta) are inserted serially in four positions; minor septa short and inserted
between major septa, probably serially also. It is this four-fold developmental pattern that gives rugose corals their
alternative name of tetracorals (tetra meaning four).
New corallites may bud asexually, although they also reproduced sexually. The buds have four septa. As the corallite
grows, the septa begin to spread further apart, and new septa are added, generally, four septa at a time are added to
maintain a rigid structure.
The growth lines on the coral span its length from the calice (top) to the base. Rugose corals generally added a new
layer of growth each day (a new wrinkle), and the days in the Paleozoic year have been determined through counting
growth rings on rugose corals. It is now known for example that the Devonian period had a year of 400 days (in the
past the Earth rotated more quickly around it's axis; this rotation is being gradually but continually slowed by the tidal
"brake" exerted by the moon's gravity).
It is not known whether rugose corals had symbiotic photosymbiotic zooxanthellae (algae) as modern corals do.
Some have suggested not, but personally I see no reason why they would not have.
They were rarely reef-builders as modern corals are. The reason being they were not able to attach themselves to the
bottom the way modern scleractinian corals can.
Classification
Classification of the Rugose corals is provisional and will no doubt be revised as more detailed investigation of the
microstructure of the different groups is made. The present arrangement (from the Treatise of Invertebrate
Paleontology) has three suborders:
suborder Streptelasmatina (Ordovician to Late Permian) - includes both solitary and colonial types. The tabulae are
domed (convex upward). The periphery of the corallum has either a stereozone or a dissepimentarium. Examples
include Streptelasma, Heliophyllum.
suborder Columnariina (Ordovician—Permian). Usually colonial, rarely solitary forms. Examples: include
Spongophyllum and Lonsdaleia. The septa are thin, the tabulae flat, depressed or downwardly convex
suborder Cystiphyllina (Ordovician—Devonian). Solitary or colonial forms. The septa are large with complex
microstructure. A wide dissepimentarium or a stereozone is present. Examples include Cystiphyllum, Calceola, and
Goniophyllum.
An alternative classification has instead two orders, and elevates the Rugosa from ordinal to subclass ranking:
Subclass Rugosa
Order Stauriida (Mid Ordovician - Late Permian)
Order Cystiphyllida (Mid Ordovician - Mid Devonian)
Evolution
The general trend among rugose corals was to evolve a strong skeleton. Several different lineages show convergent
trends toward similar morphologies. For example, several lineages developed carinae and columella to strengthen the
septa and the central axis of the coral.
The simplest and earliest grades of corallite organization appeared during the
Middle Ordovician (left). These had only simple walls, septa and tabulae. The
polyp lived on top of a tabula in a depression in the top of the coral called the
Lambeophyllum - Sandbian
(Ordovician)
calyx. A little later some types developed a layer (the marginal stereozone) of
thickened calcite around the periphery of the corallite. This would have doubtless
served a strengthening function.
For this whole period, and until the early Silurian, rugose corals remained small and solitary.
Streptelasmatina and Columnariina were dominant.
The suborders
The period from the early Silurian onwards saw the emergence of colonial forms, and an adaptive radiation and
exploitation of the reef habitat by both solitary and colonial types. During this time small horizontal internal blisterlike plates called dissepiments appeared in several lineages. These have a strengthening function and also make
incremental growth of the corallite possible, as there is no need to lift the whole polyp and create entire new tabula at
once. Nevertheless the rugose corals remained subsidiary components of reefs relative totabulate corals and
stromatoporoids. The suborder Columnariina dominated.
Few Silurian forms survived into the Devonian. A new adaptive radiation of solitary and colonial rugosans occurred in
the Middle Devonian. These were all relatively large forms with wide dissepimentaria.
above: A Middle Devonian (Eifelian) reef featuring the crinoid Dolatocrinus, a tabulate coral
(Favosites) and a trilobite (lower right corner), and a number of species of Rugose corals. The
giant one in the center is probably Siphonophrentis gigantea. The squat white ones at the middle
right would be Heterophrentis prolifica. The cluster of yellow ones in the middle foreground and
right middle background are a colonial corals of the genus Eridophyllum.
Coral faunas were seriously affected by the late Devonian mass-extinction events. Then during theearly
Carboniferous, a new adaptive radiation occurred in both solitary and colonial forms. At this time a number of types
developed a column through the middle of the corallite (through a thickening of the end of the counter septum) called
the columella. It is assumed this would also aid in strengthening and growth of the structure. In addition the
microstructure of the skeletons became quite complex. The rugose corals of this period were the most advanced types
that evolved. The organizational grades would seem to represent a tendency towards greater efficiency, like the three
grades of sponge organization. Yet paradoxically the more primitive types continue to co-exist alongside the more
advanced forms.
The final radiation of rugose corals occurred during the Permian period. Varieties with prominent prosepta appear.
These flourished until the end of the period, when the entire group was exterminated by the huge Permo-Triassic
extinction event.
References & Links
Rugose Corals (in notational form, with a few photos) and Rugose Corals (notational form)
Introduction to the Rugosa: only a brief mention
Unless otherwise attributed, text on this page may be used under the terms of the
Creative Commons License.
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
Text by M. Alan Kazlev
page uploaded 16 June 2002, last modified 2 May2003
(originally uploaded on Kheper Site 3 June 1999)
checked ATW040930
Palaeos:
ANTHOZOA
CNIDARIA
TABULATA
Page Back
Page Up
Unit Home
Page Next
Page Down
Cladogram
Subclass Tabulata
Furongian? / Early Ordovician to Late Permian
Tabulates are a Paleozoic group of corals that produce calcite skeletons of varying shapes. Unlike the contemporary
Rugose corals, they are always colonial, and never found as solitary forms, and the individual corallites are small.
The group takes its name from the organization of the colony. It is built around prominant horizontal "floors" or
tabulae. Other skeletal elements, such as septa, are reduced or absent. Because of this it has even been suggested that
these are not corals at all, but a type of extinct algae.
The larger tabulates were important reef-builders, being found in association with Stromatoporoids. Some species,
like the well known Favosites, form mound-like colonies, but there are also sheet-like, branching, and chain-like
forms.
The usual classification is as follows:
Subclass Tabulata
Order
Order
Order
Order
Order
Order
Chaetetida
Tetradiida
Sarcinulida
Heliolitida
Halysitida
Auloporida
References and Links
Introduction to the Tabulata
Corals - Kansas Fossils
Tabulata
E.N.K. Clarkson, Invertebrate Paleontology and Evolution
Unless otherwise attributed, text on this page may be used under the terms of the
Creative Commons License.
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
page by M. Alan Kazlev
page last modified 2 May 2003
ATW020819
Palaeos:
CNIDARIA
CNIDARIA
REFERENCES
Unit Home
Invertebrates
Cnidaria: References
Daly, M, DG Fautin & VA Cappola (2003), Systematics of the Hexacorallia (Cnidaria: Anthozoa). Zool. J. Linn. Soc.
139: 419�437.
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact webmaster
page uploaded 7 May 2002
checked ATW030701
page by M. Alan Kazlev (Creative Commons License)
Unless otherwise noted,
the material on this page may be used under the terms of a
Creative Commons License.
Palaeos
BILATERIA
METAZOA
OVERVIEW
Page Back
Bilateria Home
Glossary
Pieces
Page Next
Unit Back
Metazoa Home
Dendrogram
Taxon Index
Unit Next
Bilateria
Animals with Bilateral Symmetry
Metazoa
|--Porifera (paraphyletic?)
`--+--Radiata
`--Bilateria
|--Deuterostomia
`--Protostomia
|--Ecdysozoa
`--Spiralia
Introduction
Building Bilateria
The Problem of Middle Earth
Trees in the Entwood
"Where Many Paths and Errands Meet"
References
This is a holding page, pending material being added.
Page Back
Page Top
Unit Home
Page Next
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
page uploaded MAK020407, last modified ATW050724
checked ATW050724
this material may be freely used as long as attribution is given
Palaeos
BILATERIA
METAZOA
BILATERIA -1
Page Back
Bilateria Home
Glossary
Pieces
Page Next
Unit Back
Metazoa Home
Dendrogram
Taxon Index
Unit Next
Bilateria
Animals with Bilateral Symmetry
Metazoa
|--Porifera (paraphyletic?)
`--+--Radiata
`--Bilateria
|--Deuterostomia
`--Protostomia
|--Ecdysozoa
`--Spiralia
Introduction
Building Bilateria
The Problem of Middle Earth
Trees in the Entwood
"Where Many Paths and Errands Meet"
References
Introduction
This category includes all Metazoa with bilateral symmetry
(having a definite front and rear, and left and right body surfaces),
either in their adult stage or (in the case of types with secondary
radial symmetry, such as echinoderms) in the larval stage. Many
bilaterian animals exhibit cephalization, which is the evolutionary
trend toward concentration of sensory structures, mouth, and nerve
ganglia, at the anterior end of the body. This body type is related
to the further elaboration of hox genes
All bilaterians are triploblastic, which means they develop three
germ layers during embryonic development. The three germ
layers are
1. Ectoderm - Covers the surface of the embryo and
forms the outer covering of the animal and the
central nervous system in some phyla.
2. Endoderm - the innermost germ layer which lines the
archenteron (primitive gut). It forms lining of the
digestive tract and out-pocketing give rise to the
liver and lungs of vertebrates.
3. Mesoderm - located between the ectoderm and endoderm. Forms the muscles and most organs located
between the digestive tract and outer covering of the animal. The circulatory and (in vertebrates) the
skeletal system stems from this
Note that only Bilateria have a mesoderm.
Bilateria is defined by reference to a hypothetical organism,Urbilateria, who was the last common ancestor of
Deuterostomia (echinoderms and the various chordate phyla) and Protostomia (all other "higher" animals).
Classically, these two groups were said to differ in a number of respects. Some of these characteristics have been
challenged recently, but they remain generally true:
Update (April 2007): A number of
papers published in the last few years
have made it clear that the whole
traditional concept of Bilateria is probably
mistaken. Bilateria supposedly have three
unique features: (1) bilateral symmetry,
(2) hox gene patterning of the
anteroposterior axis, and (3) mesoderm.
It turns out that developing cnidarians
clearly have (1), probably have (2), and
may even have (3). It seems even more
likely now that the Cnidaria are
paraphyletic. That is, all of us bilaterian
animals are just jellyfish who happened to
use a particular type bilateral symmetry
and used the hom genes in a particular
way.
This may have important phylogenetic
implications. We cannot assume that any
bilateral organism with a couple of the
usual hom genes is inside the crown
group of Bilateria. Consequently, the
predictions we made last year (see end of
this essay) about Metazoan phylogeny are
looking good.
Most of this is covered in Brooke &
Holland (2003), which we cited -- but
obviously didn't pay enough attention to.
To see how far things have progressed at
this point, see Ryan et al. (2007).
ATW070404
One important difference involves the cleavage patterns; the
division of cells in the early embryo. Protostomes are
characterized by a spiral (the planes of cell division are diagonal
to the vertical axis of the embryo) and determinant (the goal of
each embryonic cell is established very early) cleavage.
Deuterostomes undergo radial (parallel or perpendicular to the
vertical axis of the embryo) and indeterminate cleavage (each
early embryonic cell retains the capacity to develop into a
complete embryo if isolated from other cells). But see, Halanych
(2004); Hejnol & Schnabel (2004).
There is a strange symmetry in the fate of the blastopore (the first
opening of the archenteron which forms during gastrulation.). In
Protostomes ("mouth first") the blastopore forms the mouth; in
Deuterostomes ("mouth second") - blastopore forms the anus. (see
following diagram) .
Another difference involves how the mesoderm surrounds the
body cavity. It can either pouch off or split entirely. In
Schizocoela or Protostomia, it splits. In the Enterocoela or
Deuterostomia, the mesoderm (or archenteron, the original gut)
pouches off
This diagram is showing the difference of the two major types of coelomates:
the protostomes (molluscs, annelids, arthropods, ...) and deuterostomes
(echinoderms, vertebrates, ...). These groups differ in several characteristics of
early development; In deuterostomes blastula devisions is called "radial
cleavage" because it occurs parallel or perpendicular to the major polar axis. In
protostomes the cleavage is called "spirale" because division planes are oriented
obliquely to the polar major axis. During gastrulation, protostomes embryos'
mouth was given first by the blastopore while the anus was formed later and vis
versa for the deuterostomes
Diagram and caption by YassineMrabet via Wikipedia. Creative Commons Attribution-Share Alike,
GNU Free Documentation License
Descriptions
Bilateria Hatschek, 1888 : fish > jellyfish
Range: Fr Ediacaran
Phylogeny: Eumetazoa : Cnidara + * : Protostomia + Deuterostomia
Characters: Primary bilateral symmetry, occaisonally secondarily modified to pentameral or radial, organ-system
grade of organization, most triploblastic with well-developed mesoderm of endodermal origin, most with body cavity
other than the digestive cavity, anus typically present.
Comments: For simplicity's sake, minor and problematic phyla have been excluded from the Phylogeny line; these
will be added at some later point. The Bilateria evolved from a paraphyletic Radiata; early bilateral phylogeny
remains obscure, MAK120423
Page Back
Page Top
Unit Home
Page Next
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
this page MAK120423 (from bilateria.htm)
Palaeos
BILATERIA
METAZOA
INTRODUCTION
Page Back
Bilateria Home
Glossary
Pieces
Page Next
Unit Back
Metazoa Home
Dendrogram
Taxon Index
Unit Next
Bilateria-1
Animals with Bilateral Symmetry
Metazoa
|--Porifera (paraphyletic?)
`--+--Radiata
`--Bilateria
|--Deuterostomia
`--Protostomia
|--Ecdysozoa
`--Spiralia
Introduction
Building Bilateria
The Problem of Middle Earth
Trees in the Entwood
"Where Many Paths and Errands Meet"
References
Building Bilateria
In general terms, it isn't hard to map out the place of the Bilateria
in the scheme of things. The first "animals" were probably
uninteresting hollow balls of cells closely related to the living
choanoflagellate colonies and -- somewhat more distantly -- to the
Fungi. These aggregates were clonal. That is, they all derived
from a single parent cell, and might be thought of as a single
organism. Since maintaining a hollow ball geometry is trickier
than it sounds, the first animals became adept at manipulating cellcell interactions, which naturally led to the ability to specialize
different parts of the "body." After a bit, some deviant globe of
cells developed the ability to create new globes and attach them to
the original ball.
After a few more million years of
experimentation, the result was the Porifera, the sponges, which
often have no particular shape, but have a respectable degree of
specialization within small subunits.
The really difficult part came next -- body-level organization.
How exactly this was accomplished we won't even guess, at least
not right now, since it would be off-topic. The bottom line is that
the ball of cells continued to grow, rather than budding new cell
aggregates. Since a soft, hollow sphere is not an easy shape to
maintain without collapsing, it promptly collapsed. However, it did
so in a very interesting and specific way. The maneuver involved creating an inner pocket, as shown in the image.
By geometrical necessity, this creates a population of "inside" cells in addition to the original "outside" cells, and a
more or less circular boundary region. This is pretty much all we need to create the hydra body plan of a cnidarian a
(sea anemone or jellyfish). This is also exactly what happens in the early development of all animal embryos, during
the gastrula phase, which is why we have used embryological terminology to label the parts in the diagram from
Technau & Scholz (2003).
Ernst Haeckel (1834-1919), whose many accomplishments included looking just exactly like Gandalf the Grey, was
the first person to recognize this striking parallel between early animal development and the probable course of early
animal evolution. He called his idea the Gastraea Hypothesis, and it is the real source of Haeckel's "Law" that
"ontogeny recapitulates phylogeny." This was, and remains, a powerful insight -- true in many cases. It is not a
"law" in the same sense as the physical laws of Kepler or Newton. Like many biological "laws," it is, instead, a
recurring pattern. It may not apply in any given case, but it happens often enough that it ought to be the default
hypothesis until something better comes along. However, in the case of early metazoan evolution, nothing better has
come along even now, more than 100 years after Haeckel's time. So, the next time you hear someone belittle Haeckel
(as it has become fashionable to do), remind them of this remarkable insight. Ask them if they have a better idea.
Actually, there are a number of
topologically reasonable ways to create an
inner compartment, and most can be found
within the embryological diversity of the
Cnidaria and Ctenophora.
All such
methods create endoderm and ectoderm,
but the gastrula maneuver creates a critical
ring-shaped
boundary
area
where
ectoderm and endoderm meet. It isn't clear
yet why this should have such importance,
but it plainly does. In cnidarians, this is
the mouth or hypostome. It is the most
specialized region of the body, and around
it develop the other specialized structures,
particularly the tentacles and a large
proportion of the sensory cells. This area is
clearly a critical link, but extraordinarily hard to understand, as we will discuss later.
The hydra body plan, described above, lacks two important elements which are shared by allBilateria, i.e.,
Protostomia (bugs, worms, clams, etc.) plus Deuterostomia (sea urchins, people, etc.). These were mentioned in the
introduction: (1) mesoderm and (2) bilateral symmetry. Mesoderm is a third embryological "germ layer"
(fundamental cell type) which develops in the blastocoel after gastrulation. There are several different ways in which
mesoderm is produced, which tend to differ between protostomes and deuterostomes. In general, mesoderm
formation appears to be based on a key transcription factor coded by the regulatory gene brachyury. We go into this
in excessive detail below. The evolution of bilateral symmetry is actually the more difficult development to explain,
and we have no easy answers.
The net result of all this reconstruction is a sort of short, worm-like animal with a mouth (but not necessarily an anus),
a primitive gut, a specialized area around the mouth with a tendency to develop sensory structures, mesoderm (and,
so, probably some internal specializations such as body wall muscles), and bilateral symmetry. That, folks, is
Urbilateria.
The Problem of Middle Earth
Urbilateria the ur-bilaterian, the last common ancestor of Attila the Hun and Atta the ant, is a rather critical player in
our story because of the way we have defined our clades. As a reminder, the bare bones of our working phylogeny
looks like this:
Metazoa (animals) = [toads > toadstools]
|--Porifera (sponges)
`--+--Cnidaria (anemones, jellyfish, etc.)
`--BILATERIA = [men + mollusks] = Urbilateria and all of its descendants
|--DEUTEROSTOMIA = [men > mollusks]
`--"Middle Earth " (paraphyletic)
`--PROTOSTOMIA = [bugs + slugs]
|--Ecdysozoa = [bugs > slugs]
`--Lophotrochozoa = [slugs > bugs]
|--Annelida
`--Mollusca
Bilateria is a crown group. It is defined as
the extended family of the last common
ancestor of men and mollusks, or Attila and
Atta if you prefer. In short, it consists of
Urbilateria and all of its descendants. Since
it is a crown group, it has two branches, one
leading to men and the other leading to
mollusks. Deuterostomia is a stem group,
defined to take in everything more closely
related to men than mollusks. That is, it
takes up all of the branch of Bilateria
leading toward humans.
However
Protostomia is not a stem group. It is another crown group, defined by the intersection of bugs (Ecdysozoa) and slugs
(Lophotrochozoa). Therefore, there is a gap of unknown size and composition between Urbilateria and Protostomia
containing animals which are bilaterians, but neither deuterostome nor protostome. We have labeled this part of
phylospace "Middle Earth," since it may be completely mythical, or quite real and filled with all manner of unfamiliar
creatures. In order to get a handle on Bilateria, the challenge is to find out if Middle Earth exists and, if so, who lives
there. That, in turn, may depend on what characteristics Urbilateria might have had.
We will use two distinct approaches to investigate the population of Middle Earth: phylogeny and embryology. The
phylogenetic approach involves the usual business of comparing warring cladograms and methodologies. This
approach focuses on the point where bugs and slugs diverge. Anything bilaterian, but basal to that point, is a hobbit.
The embryological approach, by contrast, focuses on Urbilateria. It tends to be difficult, theoretical and often quite
speculative. However, as we will see, there are excellent reasons for resorting to this sort of thing, as a check on the
usual phylogenetic methods.
Trees in the Entwood
We would love to say that no one could possibly
review all of the recent literature on bilaterian
phylogeny. Unfortunately several inconsiderate
show-offs evidently had nothing better to do, and
have published just such reviews, too recently to
ignore. Zrzav� (2001); Halanych (2004),
Philippe et al. (2005). Confronted with the
undeniable evidence of all this industry, we can
claim only that no one as shiftless as we are could
possibly review all of the recent literature on
bilaterian phylogeny.
Nonetheless, we have
dabbled in the literature in our usual cursory
manner -- at least to the extent of casually
skimming the said reviews of Zrzav�, Halanych,
and Philippe et al.
Fortunately, a few examples will suffice to make
the relevant point. What we will find is that, while
we can obtain some hints, the trees of Middle Earth often turn out to be Ents. That is, they are not fixed in place, but
shift their limbs in unexpected ways, with a tendency to ensnare unwary travelers with their long branches. In short,
these trees are unstable and exhibit long branch attraction.
Consider Steinauer et al. (2005). One of the problems with investigating Middle Earth is that its inhabitants are
poorly known. Most candidate species are rare and/or obscure parasites. The Acanthocephala are a case in point. The
acanthocephalan Leptorhynchoides is a parasite of vertebrate carnivores which looks a little like a nemertine worm
with training wheels. Virtually the only persons with an interest in Leptorhynchoides are a few, notoriously singleminded, public health types whose interest begins and ends with finding new methods for making the Acanthocephela
yet rarer and/or more obscure.
To the extent that anyone has cared at all, the understanding has
been that acanthocephalans are some kind of evil rotifer
cousin (Herlyn et al., 2003) or just some kind of evil rotifer
(Zrzav�, 2001). Undeterred by considerable morphological
evidence, Steinauer et al. sequenced the entire mitochondrial
genome of Leptorhynchoides. They found many useful bits of
information which, as in most mtDNA studies, they proceeded to
ignore completely (see methodological critique at Insectivora).
These workers then applied the usual sequence-based techniques
to construct a phylogenetic tree. Perversely, they failed to include
any sequences from rotifers, gnathostomulids, chaetognaths, or, in
fact, anything that might actually be comparable to
Acanthocephala. They found that (a) nothing lives in Middle
Earth; (b) acanthocephalans are close to Platyhelminthes; (c) both
are mollusks; and (d) that long branch attraction might have
played a part in the results.
We can certainly endorse the last conclusion. If a group of DNA
sequences are similar, one can sometimes create a valid
phylogenetic tree by comparing the changes at homologous sites.
Over time, however, the same site will have changed multiple times. The nucleotide found at the site becomes
randomized and carries no valid phylogenetic signal. See discussion at Insectivora. The relevant time frame varies
with the site, the species, and by blind luck. Consequently, when one is comparing sequences which diverged long
ago and are quite different ("long branches"), the phylogenetic signal is swamped by random noise and by other, nonphylogenetic, effects, giving rise to all kinds of spurious results.
This is called "long branch
attraction." Unfortunately, long branch attraction actually becomes worse as more data are added to the matrix.
Philippe et al. (2005). This effect may be analogous to a subtle problem related to missing data in morphological
work. See discussion at Pythonomorpha. However, contrary to a wrong, but oft-repeated, scientific fable,
morphological parsimony methods are generally much less subject to this source of error. Siddall & Whiting (1998).
For reasons we really ought to discuss one day, small, parasitic organisms tend to mutate a lot faster than others.
Most of the possible inhabitants of Middle Earth are just such organisms and are known to mutate faster than a comic
book superhero. Lavrov & Lang (2005) (nematode mtDNA "statistically indistinguishable from randomly shuffled
genomes with the same gene content") Thus these organisms quickly reach branch lengths at which any phylogenetic
signal is overwhelmed by noise.
Treehugging
In the last few years, a number of groups have
attempted various strategies to confine these
phylogenetic Ents. While they have had somewhat
more success than Sauruman the White, the results
have been unsatisfactory.
Lavrov & Lang (2005) used the promising
technique of comparing mitochondrial gene order
(not sequence); but they were forced to eliminate
taxa with unacceptably high rates of branch
growth. They obtained believable results, but
resolution was poor and all of the likely inhabitants
of Middle Earth had to be pruned to make their
approach work. Much the same might be said of
Philippe et al. (2005), who use 18S rRNA
sequences.
Both recognize the long-branch
problem, attempt to deal with it, and recover
reasonable trees -- but with very little phylogenetic resolution and no information on the phyla of interest to us.
A more productive approach may be that of Ruiz-Trillo et al. (2002), who used one of the myosin genes to map
bilaterian phylogeny. Myosin is one of the critical contractile proteins of all metazoan cells -- and almost all
eukaryotic cells. The reasoning, which seems to have been sound, was that myosin is under more or less uniform
structural constraints and ought to evolve both slowly and regularly. The authors conclude that acoel worms and
nemertodermatids (collectively, the Acoelomorpha) are basal to the rest of the former Platyhelminthes which are
protostomes. The only difficulty is that the Acoelomorpha also end up basal to the crown group Bilateria! As it turns
out, this is entirely possible. See discussion of Cook et al. (2004), toward the end of all this noise. Nevertheless,
Ruiz-Trillo et al. (2002) requires no panic-driven redefinition of the Bilateria. We certainly thought about it, but soon
stopped screaming (when we ran out of breath). Restored to our customary state of cheerful pomposity, we set about
the serious business of making excuses. Fortunately, the branch order of a number of the major clades seems to be
slippery in this study. Accordingly, we can't take it as solid authority that Acoelomorpha has fallen right out of
Bilateria.
Halanych (2004) has attempted to
summarize those results in the
figure reproduced here. However,
it is unclear what definition he is
applying to Bilateria.
In our
scheme, there are only three kinds
of
Bilateria:
deuterostomes,
protostomes and hobbits (stem
protostomes -- the yellow lines in
the figure).
Since Halanych
supplies no phylogenetic definition,
we were forced against our will,
and with much gnashing of teeth,
rending or garments, etc., to consult
the primary sources for the three
groups marked with red lines.
As it turns out, the Orthonectida are
almost
certainly
metazoans,
particularly under our stem group
definition; but it's anyone's guess
whether they, or any other
members of the mysterious
Mesozoa, are also bilaterians.
Hanelt et al. (1996). At any rate
neither Hanelt's study nor any later
work exclude the Mesozoa from
Middle Earth. Zrzav� (2001).
The Myxozoa may well reside in
Middle Earth. Monteiro et al.
(2002). Zrzav� (2001) asserts that
myxozoans are more primitive, but
his review was written before
Monteiro's
report
that
the
morphologically
bilaterian
Buddenbrockia is a myxozoan.
The Acoelomorpha are consist of two different phylum-level groups, the Nemertodermatida and the Acoela. Both of
these groups were parts of the classical Phylum Platyhelminthes. However, molecular studies consistently separate
the acoelomorphs, and the separation is plausible on morphological grounds. We will return to these candidate
hobbits later. For the moment, it is enough to note that both are likely bilaterians, but either or both may be
protostomes.
Finally, Halanych places the Chaetognatha right in the middle of Middle Earth. This is recent work from Halanych's
own lab, and it has not yet attracted much support from elsewhere. Chaetognaths ("arrow worms") were considered
to be deuterostomes until quite recently, when they were relocated to the protostomes. Possibly the thought of
moving this lot yet again has inspired some resistance.
"It is a lovely language, but it takes a very long time to say anything in it."
Phylogenetic methods may ultimately be able to sort out the long branch problem by using carefully chosen, highly
conserved sequences. But phylogenetic methods, particularly molecular methods, can also suffer from an opposite
problem, i.e. not just long branches, but short stems. A branching pattern can best be resolved if the radiation occurs
in a relatively slow and orderly way, so that each critical divergence is clearly separate from the previous one.
However, Middle Earth was being populated right in the midst of the Cambrian Explosion -- arguably the most rapid
series of evolutionary changes which the Animal world has ever experienced.
The Molecule Mob have always argued that the Cambrian Explosion did not exist, and that the actual divergences
occurred deep in the Neoproterozoic. However, if that were the case, as some of the Moleculons now admit, these
tree-building methods ought to work much better than they in fact do. Rokas et al. (2003).
Indeed, Rokas et al. (2005) have now turned the argument on its head. They make a fairly convincing case that the
failure of sequence-based methods to reconstruct the tree at this point in time tends to prove the existence of a
Cambrian Explosion. They point out that, if the Explosion did occur, and if it lasted significantly less than 50 My
(which is a reasonable bet on morphological grounds), even the complete sequence of the all living lineages from that
event may be mathematically insufficient to reconstruct the branching pattern. In other words, it may not even
theoretically possible to construct the correct tree using these methods. The fact that we cannot seem to get consistent
results using these methods strongly suggests that the metazoan phyla in fact diverged within a short space of time.
Note, however, that Levinton et al. (2004), using the same theoretical framework, claim that that they have recovered
the correct phylogeny using sequence methods, so that the Cambrian Explosion cannot have existed. However, the
phylogeny they rely on (Wray et al., 1996) actually draws a tree of only 6 metataxa, using individual taxa and
methods which are not fully described. We view this dispute with placid ambivalence, suspecting that the truth lies
between the positions garrisoned by these contending forces.
Page Back
Page Top
Unit Home
Page Next
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
page uploaded MAK020407, last modified ATW050724
checked ATW050724
this material may be freely used as long as attribution is given
Palaeos
BILATERIA
METAZOA
BILATERIA -2
Page Back
Bilateria Home
Glossary
Pieces
Page Next
Unit Back
Metazoa Home
Dendrogram
Taxon Index
Unit Next
Bilateria-2
Metazoa
|--Porifera (paraphyletic?)
`--Radiata
|--Cnidaria
`--Bilateria
|--Deuterostomia
`--Protostomia
|--Ecdysozoa
`--Spiralia
Introduction
Building Bilateria
The Problem of Middle Earth
Trees in the Entwood
"Where Many Paths and Errands Meet"
The Fellowship of the Ring
References
"Where many paths and errands meet"
Given the very serious problems with the usual phylogenetic
methods, we are forced back onto embryology. Unfortunately,
although Haeckel's dictum holds remarkably well in getting us
from choanoflagellates to Cnidaria, it makes no robust predictions
about Urbilateria or Middle Earth. Nevertheless, by triangulating
through the known developmental scripts of existing organisms, it
has been possible to say a few useful things about Urbilateria and
the inhabitants of Middle Earth. In a few cases, we can look to
classical developmental biology, as Haeckel did. However, in
most cases, we must depend on the homology of transcription
factors, the proteins that control developmental programs by
promoting or suppressing the expression of particular genes.
Reading the Runes
Runic scripts are writing schemes which tend to fall in between
pictograms and context-dependent constructs like alphabets.
Erwin & Davidson (2002) remind us that transcription factors are
somewhat the same way. They argue that just because (1) many
protostomes and deuterostomes have eyes and (2) the pax-6 transcription factor is involved in the morphogenesis of
eyes in both cases does not mean that the eyes of protostomes and deuterostomes are homologous, or even that
Urbilateria had eyes. The lowest common denominator is pax-6 regulation of photosensitive pigments. These
pigments may be found in eyes in many organisms, but they may also have been spread throughout the ectoderm of
Urbilateria, for all we know, as they are in some echinoderms. On this basis, Erwin & Davidson conclude that
Urbilateria was probably a very simple organism, possibly similar to the Ediacaran Kimberella (or the extant, and
even simpler, Xenoturbella).
It's a good point. The meaning of these runes depends on context and cannot be equated one-to-one with some
particular morphological expression. But they aren't a completely arbitrary alphabet, either; and, besides, we know
something about their context. Like most questions of homology, it is not entirely a yes-or-no affair. The eyes of
protostomes and deuterostomes are homologous -- to the extent that they both use the pax-6 regulatory system in a
way which seems to have been unchanged since their last common ancestor (by definition, Urbilateria). But, whether
we can make stronger claims to homology depends on what other evidence we have. Consider a few examples.
The Underworld of Middle Earth
Perhaps the most fundamental issue relates to the
formation of mesoderm. The single most basic difference
between bilaterians and all other animals is the presence of
this third germ layer. If mesoderm is not homologous in
all bilaterians, then we may as well pack it in and quit
blathering about Bilateria, because Bilateria won't be a
monophyletic group.
Unfortunately, we aren't going to be able to weasel out of
this discussion so easily. Technau & Scholz (2003) have
recently reviewed this area. Here, the case for homology
is strong, and we may be reasonably certain that all
hobbits have mesoderm. To put the matter briefly,
mesoderm formation is run by some combination of the
following four genes (or gene families): twist, snail,
brachyury, and the GATA family. Despite the names, this
is probably not a crime syndicate. The GATA family, for
example, is supposedly named for part of the sequence to
which the GATA gene products bind. On the other hand,
the GATA family are elusive, hard to pin down, and appear to operate by "facilitating" the business of higher level
inducers, "enforcing" their directives at the level of individual genes. So, it's safer to be ... respectful.
The GATAs are unique to Bilateria and may be a synapomorphy of the clade. The other factors play rather
interesting roles in the Cnidaria. Twist is responsible for muscle, or, rather it "induces" endodermal cells to form
muscle. The endoderm might make muscle anyway, if asked politely, but twist is a very convincing inducer. It hangs
around just in case, to keep everyone ... respectful. Snail is also involved in muscle formation, but its role lies in
making the right contacts -- regulating cell-to-cell adhesion or mobility, making the right arrangements. Brachyury
just sits on the lip of blastopore. It doesn't seem to do much of anything, except to offer little suggestions to cells
migrating into the archenteron. Mostly those cells are, as we said, respectful. But, if not, it turns out that brachyury
happens to be very close friends with twist and snail.
Muscle is one of the basic mesodermal tissues in
bilaterians, and it is easy to see how this very
efficient arrangement could lead to the formation
of an entirely new germ layer. Rieger & Ladurner
(2003). All that is needed is to recruit more
lieutenants to take on more specialized jobs, which
is presumably where the GATA Family came in.
While this is an elegant explanation of mesoderm
evolution, it seems that not everyone agrees. See,
e.g., Lartillot et al. (2002), reporting that Patella
orthologs of twist and snail are not involved in
mesoderm specification. Perhaps so, but we are
unconvinced.
Mind you, this has absolutely
nothing to do with the alleged little gift we
allegedly received from the alleged GATA
brothers. They don't even exist. It's all malicious
gossip. Honest! We just believe in being ...
respectful.
Rings of Power
Another point which now seems secure is that Urbilateria was not segmented. Seaver (2003). Segmentation is best
understood in arthropods and, more generally, in the Ecdysozoa. Almost all ecdysozoans are segmented but, tellingly,
their most basal living group, the nematodes, are not segmented. In insects, segmentation involves two types of
transcription factors: polarity genes (particularly engrailed) and pair-rule genes. While polarity genes are quite
commonly used in the same way in other ecdysozoans, the pair-rule genes are not. They appear to play a secondary
role. Seaver (2003).
Most lophotrochozoans are unsegmented. Among the segmented annelids, the segmentation pattern is established
through stereotyped spiral divisions in early embryogenesis, which is unlike the insect model. The initiating
embryological tissue varies a good bit within the Annelida, and even between adult and larval segments. Like
vertebrates, but unlike insects, segmentation proceeds in an anterior to posterior sequence; and; unlike both, the
number of segments is not even approximately fixed, and segments continue to develop after embryogenesis is
complete. Engrailed is involved in
segmentation, but it seems to be a
secondary messenger. The pattern
of engrailed expression differs
markedly
even
among
the
Polychaeta,
and
normal
development is possible after
ablation of engrailed-expressing
cells. The heavy lifting is done,
instead, by a completely different
set of genes, the "gap" genes,
which have no role in insect
segmentation. Seaver (2003)
There is no evidence that any of the basal deuterostomes were segmented. Segmentation was re-invented among early
chordates. Unlike any protostome, chordates use a rather delicate timing mechanism based on oscillating expression
of the notch gene to produce segments during elongation of the dorsal midline. This creates mesodermal aggregates,
somites, which actually create the segmental structures. Pair rule, polarity and gap genes are present, but seem to
have little or no role in segmentation. Accordingly, we can be fairly confident that Urbilateria was not segmented
and that any hobbits, orcs, elves, etc. are unlikely to exhibit segmentation. Seaver (2003).
The Eyes of Sauron
As discussed, Erwin & Davidson (2002) use pax-6 and eye development as an example of the crawling evils of naive
reliance on crude transcription factor homologies. Actually, a more current version of this story, as reviewed by
Arendt & Wittbrodt (2001), is a rather more urbane and polished tale. Arendt and co-workers have further refined
the work since that time. Fortunately, it is not necessary for us to summarize this work, since Pharyngula has
conveniently predigested the data at Rhabdomeric and ciliary eyes. A lot more than pax6 backs up the claim that eyes
have extensive homologies across Bilateria.
For those too indifferent even to read Pharyngula's abbreviated
account, here's the TV Guide version:
Light comedy. Two photosensitive young systems, the ciliary
and rhabdomeric, meet in the Proterozoic where they both
work as photoperiod detectors. In the deuterostomes, they
move in together, but the ciliary system starts seeing others,
and the rhabdomeric system is relegated to a supporting role
and menial household work. In the protostomes, they break
up, and the rhabdomeric system goes on to have a glamorous
career in pictures while the ciliary cells have a dead-end job in
the brain. Cast of billions. 600 My, mostly B&W or limited
Color, PG-13 (adult and/or larval situations, some
nudibranchs).
"Hobbits ... are inclined to be fat in the stomach"
This section could well be called Haeckel's revenge. A number of
workers have recently obtained spectacular results with oldfashioned descriptive embryology. In part, the quality of these
papers is due to the availability of sophisticated molecular probes
for looking at gene expression. However, some of the improved
quality simply results from doing better embryology. The
tendency in the past has been to compare embryos of vertebrates
and Drosophila. But both have very specialized developmental
patterns and are unlikely to tell us much about Urbilateria.
The tale from the vertebrate-fruit fly comparison is well known. In protostomes, the mouth develops "first." More
accurately, the blastopore elongates along the future ventral midline and the mouth and anus develop at opposite
ends. In deuterostomes, the gut elongates. The blastopore forms only the anus, and the mouth is formed by punching
a new hole in the ectoderm along the future ventral midline. This seems very different only if one bases the
comparison on vertebrates, in which mouth formation is delayed and involves some very peculiar behavior by the
surrounding ectoderm. See, e.g., The Basisphenoid and Teeth: Overview.
Arendt et al. (2001) do the sensible thing and compare the surprisingly similar primary ciliated larvae of polychaete
annelids and hemichordates. Despite the fact that these are protostome and deuterostome, respectively, their primary
larvae are strikingly similar in morphology and, as it turns out, in the way they express the key transcription factors
related to gut formation: otx, brachyury and goosecoid. At this point, we are moved to remind the enthusiastic reader
of the teachings of Erwin & Davidson (2002). These factors may not have the same degree of priority in the
developmental scheme. They may not be doing the same thing -- but it's a very pretty picture all the same.
We lack the space for a full description of what Arendt & Co. did, but the bottom line is a strong case for homology
of a three- (actually four-) part gut with a mouth, stomadeum (pharyngeal region in vertebrates), an undifferentiated
middle section, and an anus.
The Wisdom of the Elves
Last, but far from least, we consider the
information on the nervous system; and, indeed,
the subject makes us nervous. We have readily
confessed that the scope of this essay has forced us
to rely on secondary sources. Here, we have been
schooled by Alain Ghysen (2003) who produces
utterly intimidating work at the intersection of
molecular, developmental, evolutionary and
neurobiology at the Universit� Montpellier II,
when he is not raising orchids or studying poetry
in an interesting array of archaic languages.
Whether Prof. Ghysen is actually an elf, we cannot
say. Before rejecting this hypothesis, consider also
that he is a dedicated archer. We can say that he, like an elf, is sometimes opinionated and, when asked to give his
opinion, he is rarely shy about expressing his views. In Ghysen (2003), he gives fair warning that he will "venture
into somewhat speculative grounds."
He then proceeds to erect a strongly fortified position around the proposition that Bilateria not only had a reasonably
well-organized nervous system, but also the framework of a multi-part brain, brainstem, longitudinal nerve cord, and
several types of specialized sensory cells, particularly mechanosesnsory cells. The bit about the proto-brain sounds
like a stretch, but others, working from wholly different premises, have come to the same conclusion. Meinhardt
(2002) (mainly body patterning in early development). Ghysen, as one might expect, relies on a battery of molecular,
developmental, evolutionary, etc. evidence. We won't attempt to summarize it here, since we have gone on too long
as it is. At least that's our story, and we're sticking to it. In fact, it is already time to move on to the next section:
Page Back
Page Top
Unit Home
Page Next
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
page uploaded MAK020407, last modified ATW050724
checked ATW050724
this material may be freely used as long as attribution is given
Palaeos
BILATERIA
METAZOA
BILATERIA -3
Page Back
Bilateria Home
Glossary
Pieces
Page Next
Unit Back
Metazoa Home
Dendrogram
Taxon Index
Unit Next
Bilateria-3
Metazoa
|--Porifera (paraphyletic?)
`--Radiata
|--Cnidaria
`--Bilateria
|--Deuterostomia
`--Protostomia
|--Ecdysozoa
`--Spiralia
Introduction
Building Bilateria
The Problem of Middle Earth
Trees in the Entwood
"Where Many Paths and Errands Meet"
The Fellowship of the Ring
References
The Fellowship of the Ring
In this final episode, we will attempt to do two
things: (1) sum up the characteristics of Bilateria
and (2) briefly review the candidate Middle
Earthlings in view of those findings. Detailed
comparison must be left for more detailed
consideration of each particular kind of hobbit,
orc, elf, etc.
The One Ring
One of the advantages of really profound
ignorance is that it makes neutrality much easier.
We began looking into this subject with very few
preconceptions about Urbilateria. Having finished
for now with this suppositional ancestor of all
Bilateria, we find that he is a peculiar mixture of
highly derived, and exceedingly primitive.
Urbilateria undoubtedly was a triploblast. That is,
he had a fairly well-established system of mesoderm induction. Technau & Scholz (2003). Like cnidarians, he had
muscles. Since he had a differentiated ectoderm separate from the gut, and also had a longitudinal layer of muscle
and bilateral symmetry, he probably had some degree of motility. Indeed, he would almost have to have been mobile,
unless he were fixed to the substrate. But a sessile lifestyle seems unlikely. Sessile organisms leave trace fossils in
the form of holdfasts, variously reinforced holes in the substrate, and, frequently, mineral remains. Nothing like this is
associated with bilaterians until the Small Shelly Fauna of the Terreneuvian. So, Urbilateria was either motile or he
arrived extraordinarily late, in addition to being oddly designed for a benthic couch potato.
There are several other reasons for
believing Urbilateria was not sessile.
One is trivial. Sessile aquatic animals
are rarely very small. It is simply too
easy for them to be buried in sediment.
Yet we can be relatively confident that
Urbilateria was indeed small, since
there are no indications of anything
large and bilaterian until the middle of
the Early Cambrian. Of course, fossils
from any earlier date are extremely
rare, so some behemoth proto-planarian
might yet show up. However, while the record of Neoproterozoic body fossils is poor, we have a large sample of
worm-like trace fossils to examine, beginning in the Ediacaran. These are all quite small.
The next proof is important: the complex gut. Arendt et al. (2001). Suspension feeding doesn't require a specialized
mouth or stomadeum. In fact, that would be counterproductive. A suspension feeder wants a big surface area to catch
potentially edible particles. Filter feeding is a possibility, but this requires rather specialized equipment, none of
which seems to have survived as homologous structures in the main Bilaterian clades. The path of high probability is
that Urbilateria moved -- if not much -- in search of food.
That leaves two possible lifestyles: on the substrate or in the water column. Kimberella seems to have been a preCambrian bilaterian bottom-feeder. If so, at least some bilaterians had this niche. It has recently been shown that
some Late Proterozoic bottoms may have experienced extensive, millimeter-scale horizontal bioturbation (Dornbos et
al., 2005) -- presumably caused by bilaterians not yet able to penetrate the substrate. In any case, it is nearly
impossible to imagine something this small and ungainly as a swimmer. Doubtless there were planktonic forms, but
Urbilateria seems rather complex for life as a floater, and there would be little point in its various physical
specializations. So, we conclude that Urbilateria was part of the benthic epifauna -- a bottom crawler.
We have two sorts of physical models for Urbilateria. The trace fossils and Acoela suggest something long and
generally worm-like. Kimberella and Xenoturbella look more like very primitive mollusks. Fortuitously, the living
Nemertodermatida are, morphologically, more or less in the middle, which is probably our best bet. Finally, The
evidence indicates that Urbilateria had a nervous system with one or (more likely) two main longitudinal nerve cords
and an anteriorly-placed nerve plexus, sensory cells for the perception of light, and some kind of mechanosensory
system.
The Fellowship of the Ring
All this is excellent news, since it
constitutes a reasonably good fit
with some of the candidates for
Middle
Earth
citizenship,
particularly the Acoelomorpha.
This is the name given to a
supposed clade made up of
Nemertodermatida and Acoela.
Four other groups are possibilities.
Two seem too primitive, and may well turn out to be weird Radiata: the Myxozoa and Mesozoa. Two others are
probably too specialized and may turn out to be protostomes (and ecdysozoans in particular): the Chaetognatha and
Gnathostomulida. Nevertheless, we will include them all in this section for now, although we'll probably change our
minds before it's finished.
Acoela: The Acoela were classically shelved with the Platyhelminthes, those exceedingly dull worms which you
quickly forgot after the first quiz in Introductory Zoology. You heard on good authority that the instructor never asks
about them on the Final. This turned out to be true. In fact, it is always true. No instructor ever asks about the
Platyhelminthes on the final. It is simply one of those unspoken laws of academic nature. So, how could we possibly
prevail on you to stay awake long enough to absorb serious data on the Acoela -- a group of worms so cloddish and
uncouth that they have been demoted from the platyhelminths to an even lower plane of phylospace? We will
perform this impossible task by applying a time-honored academic maxim. "The difficult, we do immediately. The
impossible, by pawning it off on someone else". See the excellent essay on this topic by Prof. Seth Tyler (U. Maine),
or pages 9-11 of Tekle (2006).
Tentatively, we disagree with almost all of Prof. Tyler's
phylogenetic conclusions.
The Acoela are quite
specialized and weird, but this is only to be expected of a
surviving twig at the base of an enormous radiation 600
My ago, or more.
Further, the closely related
Nemertodermatida provide a connecting link to more
conventional bilaterian designs. The acoel gut is not a
conventional 3-part bilaterian gut, but the longitudinal
body wall muscle net, nerves, brain, mode of life, and
body form are correct. For that matter, even the syncytial
"gut" departs further from the cnidarian or poriferan
system than from the bilaterian archenteron. Development
proceeds in a recognizably bilaterian fashion, with a spiral
cleavage pattern (albeit a spiral pattern different from that
of annelids). Morphologically, the Acoela are simply
well-behaved hobbits with full stomachs.
The one real hitch comes from Cook et al. (2004), who
looked at acoel hox genes. To make a long and somewhat uncertain story shorter and more certain than it really is, at
least one acoel has only three hox genes, rather than the usual bilaterian minimum of 7-8. Then again, as these
authors note, nematodes also have a reduced hox set. In that case, it is generally thought that the number is due to
gene loss. So, the number in Nemertodermatida may be due to a similar effect of small body size.
Nemertodermatida: The Nemertodermatida are a small group of acoels in rehab. They're trying to conform. They
have guts. Sure, they've gained a little weight, but their nerves are better. Certainly, their sex lives are simpler and
more conventional. Their developmental pattern is similar to that of acoels. Larsson (2003). They're taking it one
day at a time.
Myxozoa: The Myxozoa are odd parasites, originally thought
to be protists (Brooke & Holland, 2003), with almost
nothing to suggest that they are bilaterians -- usually. The
Myxozoa are generally found as intracellular parasites with
very complicated lives, including two hosts; for example,
annelid and fish. Recently, Monteiro et al. (2002) realized
that the mystery worm and putative bilaterian Buddenbrockia
was actually an unrecognized form of the myxozoan,
Tetracapsula. In this avatar, the myxozoan has body wall
muscles in four regular muscle blocks and vaguely bilateral
symmetry. Id.
Initial molecular and morphological studies placed the myxozoans as close cousins of the parasitic cnidarian
Polypodium. Later molecular work put them in the Bilateria. Halanych (2004). Nevertheless, outside the noisy clan
of molecular enthusiasts, those who have actually studied Myxozoa seem to be convinced that myxozoans are, in fact,
cnidarians. Some are polite enough not to say so directly. Kent et al. (2001). Others are less polite, possibly with
good reason. Siddall & Whiting (1998). As Siddall & Whiting point out, it is hard to accept a bilaterian with
nematocysts, and the polar capsules of Myxozoa are undoubtedly nematocysts modified as crowbars to break into host
cells. Zrzav� (2001) (citing much other evidence).
One obvious problem is, as we mentioned at the beginning, that few
participants in these debates ever define their terms. It is not clear that any of
these folks would assert that Myxozoa are bilaterians if they actually bothered
to give Bilateria a phylogenetic definition. It is entirely possible that this
entire dispute is, quite literally, about nothing. Nevertheless, we will treat the
Myxozoa as presumptive Bilateria for now, simply for lack of a better place
to deal with them.
Mesozoa: The Mesozoa are another group of intracellular parasites -- unlikely
candidates, you'd think, for bilaterian status. They are now believed to be
made up of two, possibly unrelated, groups: the Dicyemida (Rhombozoa) and
Orthonectida. They are known to have hox-like genes, and one of the
dicyemid genes has a lophotrochozoan signature.
Some, generally
conservative, workers have found that the Mesozoa are in fact monophyletic
and that they branch near the Acoelomorpha. Siddall & Whiting (1998);
Zrzav� (2001). This creates difficulties, since the Mesozoa have very little
other than molecular traits to tie them to the Bilateria. However, so far as we
are aware, the members of this taxon are all obligate intracellular parasites.
So, extreme simplification is only to be expected.
Chaetognatha: As mentioned, the chaetognaths ("arrow worms") are clearly
orthodox bilaterians. The question to be answered is, "why aren't they
protostomes?" Classically, they were regarded as deuterostomes, but that
now seems extremely unlikely. The chaetognaths were subsequently moved
to the ecdysozoan region, but this became less likely based on subsequent
detailed study of chaetognath development and neuroanatomy. The pattern of
early development looks "spiralian," i.e. spiral cleavage with each of the early
cells having a fixed role in the organism. (This is seems to be the primitive
condition. Extavour & Akam, 2003.) A full complement of hox genes are
present, but their expression is restricted to the central nervous system, as in
certain (other?) lophotrochozoans. Papillon et al. (2005). Thus, the current
best bet is that the chaetognaths are either lophotrochozoans or basal to
Protostomia. Chances are that they are lophotrochozoans, but mitochondrial gene order phylogenies place them in a
more basal position.
Gnathostomulida: This is, as you may have guessed, one more gutless group of worms. They differ from the others
in having jaws of a sort. As far as we can make out, no one has seriously proposed that they are basal to Protostomia.
Like the Acanthocephala, they are probably evil rotifers. However, no one seems particularly comfortable with them
either. Consequently, we will add them here as candidate orcs. And, while we're at it, perhaps we'll do the
Acanthocephala, too.
Perhaps we ought to be a bit more candid. Our reasons are actually not quite so frivolous -- almost as frivolous, but
not quite. At least since the publication of an influential paper by Giribet et al. (2000), there has been a vague
suspicion that the Ecdysozoa-Lophotrochozoa dichotomy masks a third, possibly rather large, clade. This clade -admittedly of variable composition -- tends to show up in a polytomy with the other two. It might be basal to
Protostomia (bugs + slugs). It might not. Consequently, we have picked two likely members of the group, just to
keep an eye on this possibility.
So. There we have it. Middle Earth, an imaginary land, has been populated with a hypothetical population of odd
creatures. Just for fun, we now add our completely baseless and intuitive guess about how things will turn out
in the next five to ten years.
Radiata = Cnidaria (i.e. Bilaterians are Cnidarians!)
|--Myxozoa
`--+--Acoelomorpha
| |--Nemertodermatida
| `--Acoela
`--Bilateria
|--Deuterostomia (by definition)
`--+--Gnathostomulida
`--Protostomia (bugs + slugs)
|--Ecdysozoa (bugs > slugs)
| |--Acanthocephala?
| `--bugs
`--Lophotrochozoa (slugs > bugs)
|--Chaetognatha
`--+--Mesozoa
`--slugs
ATW060218
References
Arendt, D, U Technau & J Wittbrodt (2001), Evolution of the bilaterian larval foregut. Nature 409: 81-85.
Arendt, D & J Wittbrodt (2001), Reconstructing the eyes of Urbilateria. Phil. Trans. Roy. Soc. B 356: 1545-1563.
Brooke, NM & PWH Holland (2003), The evolution of multicellularity and early animal genomes. Curr. Op. Genet.
Devel. 13: 599-603.
Cook, CE, E Jim�nez, M Akam & Emili Sal� (2004), The Hox gene complement of acoel flatworms, a basal
bilaterian clade. Evol. Devel. 6: 154-163.
Dornbos, SQ, DJ Bottjer & J-Y Chen (2005), Paleoecology of benthic metazoans in the Early Cambrian Maotianshan
Shale biota and the Middle Cambrian Burgess Shale biota: evidence for the Cambrian substrate revolution.
Palaeogeog. Palaeoclimat. Palaeoecol. 220: 47� 67.
Erwin, DH & EH Davidson (2002), The last common bilaterian ancestor. Development 129: 3021-3032.
Extavour, CG & M Akam (2003), Mechanisms of germ cell specification across the metazoans: epigenesis and
preformation. Development 130: 5869-5884.
Ghysen, A (2003), The origin and evolution of the nervous system. Int. J. Dev. Biol. 47: 555-562.
Giribet, G, DL Distel, M Polz, W Sterrer & WC Wheeler (2000), Triploblastic relationships with emphasis on the
acoelomates and the position of Gnathostomulida, Cycliophora, Plathelminthes, and Chaetognatha: A combined
approach of 18S rDNA sequences and morphology. Syst. Biol. 49: 539-562.
Halanych, KM (2004), The new view of animal phylogeny. An Rev. Ecol. Evol. Syst. 35: 229-256.
Hanelt, B, D Van Schyndel, CM Adema, LA Lewis & ES Loker (1996), The phylogenetic position of Rhopalura
ophiocomae (Orthonectida) based on 18S ribosomal DNA sequence analysis. Mol. Biol. Evol. 13: 1187�1191.
Hejnol, A & R Schnabel (2004), The eutardigrade Thulinia stephaniae has an indeterminate development and the
potential to regulate early blastomere ablations. Development 132: 1349-1361.
Herlyn, H, O Piskurek, J Schmitz, U Ehlers, & H Zischler (2003), The syndermatan phylogeny and the evolution of
acanthocephalan endoparasitism as inferred from 18S rDNA sequences. Mol. Phylogen. & Evol. 26: 155-164.
Hooge, MD & N Eppinger (2005), New species of Acoela (Acoelomorpha) from the Gulf of California. Zootaxa
1009, 14pp.
Kent, ML, KB Andree, JL Bartholomew, M El-Matbouli, SS Desser, RH Devlin, SW Feist, RP Hedrick, RW
Hoffmann, J Khattra, SL Hallett, RJG Lester, M Longshaw, O Palenzeula, ME Siddalli & C-X Xiao (2001), Recent
advances in our knowledge of the Myxozoa. J. Eukaryot. Microbiol. 48: 395-413.
Larsson, K (2003), Early Development of Nemertoderma westbladi. Unpub. Degree Project, Uppsala Univ., 13 pp.
WWW.
Lartillot, N, M Le Gouar & A Adoutte (2002), Expression patterns of fork head and goosecoid homologues in the
mollusc Patella vulgata supports the ancestry of the anterior mesendoderm across Bilateria. Dev. Genes Evol. 212:
551-561.
Lavrov, DV & BF Lang (2005), Poriferan mtDNA and animal phylogeny based on mitochondrial gene arrangements.
Syst. Biol. 54: 651-659.
Levinton, J, L Dubb & GA Wray (2004), Simulations of evolutionary radiations and their application to
understanding the probability of a Cambrian Explosion. J. Paleontol. 78: 31-38.
Meinhardt, H (2002), The radial-symmetric hydra and the evolution of the bilateral body plan: an old body became a
young brain. BioEssays 24: 185-191.
Monteiro, AS, B Okamura & PWH Holland (2002), Orphan worm finds a home: Buddenbrockia is a myxozoan. Mol.
Biol. Evol. 19: 968�971.
Papillon, D, Y Perez, L Fasano, Y Le Parco & X Caubit (2005), Restricted expression of a median Hox gene in the
central nervous system of chaetognaths. Dev. Genes Evol. 215: 369�373.
Peterson, KJ, B Waggoner & JW Haggadorn (2003), A fungal analog for Newfoundland Ediacaran fossils? Integr.
Comp. Biol. 43: 127-136.
Philippe, H, N Lartillot & H Brinkmann (2005), Multigene analyses of bilaterian animals corroborate the monophyly
of Ecdysozoa, Lophotrochozoa, and Protostomia. Mol. Biol. Evol. 22: 1246-1253.
Rieger, RM & P Ladurner (2003), The significance of muscle cells for the origin of mesoderm in Bilateria. Integr.
Comp. Biol. 43: 47-54.
Rokas, A, N King, J Finnerty & SB Carroll (2003), Conflicting phylogenetic signals at the base of the metazoan tree.
Evol. Devel. 5: 345-359.
Rokas, A, D Kr�ger & SB Carroll (2005), Animal evolution and the molecular signature of radiations compressed in
time. Science 310: 1933-1938.
Ruiz-Trillo, I, J Paps, M Loukota, C Ribera, U Jondelius, J. Bagu��, & M Riutort (2002), A phylogenetic analysis
of myosin heavy chain type II sequences corroborates that Acoela and Nemertodermatida are basal bilaterians. Proc.
Nat. Acad. Sci. (USA) 99: 11246-11251.
Seaver, EC (2003), Segmentation: mono- or polyphyletic? Int. J. Dev. Biol. 47: 583-595.
Siddall, ME & MF Whiting (1998), Long-branch abstractions. Cladistics 15:9-24.
Steinauer, ML, BB Nickol, R Broughton & G Ort� (2005), First sequenced mitochondrial genome from the Phylum
Acanthocephala (Leptorhynchoides thecatus) and its phylogenetic position within Metazoa. J. Mol. Evol. 60: 706715.
Technau, U & CB Scholz (2003), Origin and evolution of endoderm and mesoderm. Int. J. Dev. Biol. 47: 531-539.
Tekle, YI (2006), Phylogeny and Taxonomy of Childia (Acoela). Unpub. doctoral thesis., Uppsala Univ. WWW.
Wray, GA, JS Levinton & LH Shapiro (1996), Molecular evidence for deep precambrian divergences among
metazoan phyla. Science 274: 568-573.
Zrzav�, J (2001), The interrelationships of metazoan parasites: a review of phylum- and higher-level hypotheses
from recent morphological and molecular phylogenetic analyses. Folia Parasitol. 48: 81-103.
Page Back
Page Top
Unit Home
Page Next
images not loading? | error messages? | broken links? | suggestions? | criticism?
contact us
page uploaded MAK020407, last modified ATW050724
checked ATW050724
this material may be freely used as long as attribution is given
Palaeos
BILATERIA
METAZOA
PROTOSTOMIA
Page Back
Bilateria Home
Glossary
Pieces
Page Next
Unit Back
Metazoa Home
Dendrogram
Taxon Index
Unit Next
Protostomia
Metazoa
|--Porifera (paraphyletic?)
`--+--Radiata
`--Bilateria
|--Deuterostomia
`--Protostomia
|--Ecdysozoa
| |--Tardigrada
| `--Arthropoda
|
|--Trilobita
|
`--Pancrustacea
|
|--Crustacea
|
`--Insecta
`--Spiralia (Lophotrochozoa)
|--Bryozoa
|--Mollusca
`--+--Annelida
`--Brachiopoda
The Bilateral (and Coelomate) phyla can be divided into two groups, variously called the Schizocoela and
Enterocoela, or the Protostomia and Deuterostomia. These are distinguished by embryonic cleavage patterns, the fate
of the blastopore and coelom formation. The Deuterostomes are a relatively small (in terms of species and abundance)
group that includes vertebrates, echinoderms, and a few assorted minor phyla. We probably consider them (and
specifically the vertebrates) important mainly for anthropocentric reasons (we are deuterostomes). The Protostomes in
contrast include the great majority of animal species, such as arthropods, molluscs, annelids, and many less wellknown taxa.
The protostome condition is defined by a spiral and determinant cleavage in the early stages of embryo development,
schizocoelous coelom formation (as the archenteron (embryonic gut) forms the coelom begins as splits within the
solid mesodermal mass, and the formation of the blastopore (the original opening) into the mouth
Beginning with Aguinaldo et. al. 1997, molecular studies in metazoan phylogeny have divided the Protostomia can
into two further groups, one called Ecdysozoa or moulting animals, the other the Lophotrochozoa or Spiralia (the first
name is more widely used, but the second is preferable, both being the senior synonym and because it and describes
the spiral larval development of these invertebrates). Although there was originally some molecular phylogeneticists
and morphologists did not accept this approach, it has bene increasingly confirmed by later studies and is now the
mainstream position. Indeed, over the last decade or so the emphasis has shifted from the distinction between
protostomes and deuterostomes and determining which phyla go where, to that of the phylogenetic relationships of teh
various phyla that make up the Lophotrochozoa/Spiralia and Ecdysozoa (along with a few problematica which may
belong to either group), with the now well resolved Deuterostomes the third party, on the sidelines.
Descriptions
Protostomia Grobben, 1908 : slugs > bugs
Range: Fr Ediacaran
Phylogeny: Bilateria : Deuterostomia + * : Ecdysozoa or moulting animals, the other the Spiralia
Characters: Blastopore becomes mouth, typically schizocoelous with spiral cleavage.
Comments: Various microscopic animals and others with simple body plans like the flatworms (platyhelminthes) are
sometimes included in a third clade, Platyzoa, but this seems to be an artificial assemblage of mostly miniaturised
spiralians. MAK120423
Page Back
page uploaded 7 April 2002, last modified ATW050724
text content © Mikko Haaramo
checked ATW080112
this material may be freely used as long as attribution is given
Page Top
Unit Home
Page Next