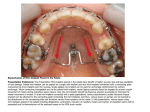
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
41 Development 103 Supplement, 41-60 (1988) Printed in Great Britain © The Company of Biologists Limited 1988 Palate development MARK W. J. FERGUSON Animal & Human Reproduction, Development and Growth Research Group, Department of Cell & Structural Biology, University of Manchester, Coupland 3 Building, Manchester M13 9PL, UK Summary In all vertebrates, the secondary palate arises as bilateral outgrowths from the maxillary processes. In birds and most reptiles, these palatal shelves grow initially horizontally, but do not fuse with each other resulting in physiological cleft palate. In crocodilians, shelf fusion occurs resulting in an intact secondary palate. Mammalian palatal shelves initially grow vertically down the side of the tongue, but elevate at a precise time to a horizontal position above the dorsum of the tongue and fuse with each other to form an intact palate. Palatal shelf-elevation is the result of an intrinsic shelf elevating force, chiefly generated by the progressive accumulation and hydration of hyaluronic acid. In all vertebrates the nasal epithelium differentiates into pseudostratified ciliated columnar cells and the oral epithelia differentiates into stratified squamous cells, but the medial edge epithelial (MEE) phenotype differs in different groups. In mammals, the MEE of opposing shelves adhere to each other to form an epithelial seam which then disrupts by cell death and cell migration into the mesenchyme accompanied by an epitheliomesenchymal transformation. In birds, the MEE keratinize resulting in cleft palate whereas, in alligators, the MEE migrate onto the nasal aspect of the palate. In all vertebrates, this regional, temporal and species-specific epithelial dif- ferentiation is specified by the underlying mesenchyme. Signalling of this interaction is complex but involves both extracellular matrix and soluble factors e.g. minor collagen types, tenascin, EGF, TGFa-, TGF£, PDGF, FGF. These soluble growth factors have a biphasic effect: directly on the epithelia and on the mesenchyme where they stimulate or inhibit cell division and synthesis of specific extracellular matrix molecules. The extracellular matrix molecules (and bound growth factors) synthesized by the mesenchymal cells may then directly affect the epithelium. These signals cause differential gene expression via second messenger systems e.g. cAMP, cGMP, Ca 2 + , pH, pi etc. Molecular markers for nasal, medial and oral epithelial cell differentiation include the types of cytokeratin intermediate filaments and specific cell surface molecules recognized by monoclonal antibodies: the genes for such molecules are probably expressed in response to mesenchymal signals. Using such an approach, it is possible to go from a morphological description of palate development to a cellular analysis of the mechanisms involved and then to identification of candidate genes that may be important for screening and diagnosis of cleft palate. Introduction investigating fundamental mechanisms common to the embryogenesis of many structures as it appears relatively late in embryogenesis, can be easily excised and cultured under chemically defined, serum-free conditions (Ferguson, Honig & Slavkin, 1984) and exhibits morphogenetic movements (Ferguson, 1978), extracellular matrix synthesis (Brinkley & Morris Wiman, 1984, 1987; Pratt & King, 1971; Silver, Foidart & Pratt, 1981), neurotransmitter synthesis (Zimmerman & Wee, 1984), cell adhesion Development of the mammalian secondary palate is a complex and critical event, which, in man, is frequently disturbed resulting in the common and distressing birth defect of cleft palate. Thus palate embryogenesis has been the target of much research with the long-term aim of rational prevention or improved treatment for cleft palate. Moreover, the developing palate is often used as a model system for Key words: palate, extracellular matrix, epitheliomesenchyme interaction, growth factor. 42 M. W. J. Ferguson (Greene & Pratt, 1977), epithelial-mesenchymal interactions and regional patterning (Ferguson & Honig, 1984). Several reviews summarize the literature on palate development (Greene & Pratt, 1976; Ferguson, 1978; Shah, 1979; Pratt & Christiansen, 1980; Melnick, Bixler & Shields, 1980; Zimmerman, 1984; Pratt, 1984; Pisano & Greene, 1986; Ferguson, 1987). Rather than re-review the extensive literature on palate development in this paper, I first provide an overview of secondary palate development in vertebrates and then focus on important developmental questions, particularly in relationship to recent work from our laboratory on the mechanism of regional specification of palatal epithelial differentiation by the underlying mesenchyme. Overview Mesenchymal cells from the neural crest migrate to the primitive oral cavity where in association with craniopharyngeal ectoderm they form the bilateral maxillary processes. Bilateral palatal shelves arise from these maxillary processes at embryonic day 12 in mice, day 6 in chickens, day 17 in alligators and day 45 in man. In mammals (both eutherian and protherian), these bilateral palatal shelves at first grow vertically down the sides of the tongue (Fig. 1), but at a precise developmental stage they rapidly elevate to a horizontal position above the dorsum of the tongue (Fig. 2). The medial edge epithelia of the approximating palatal shelves then fuse with each other to form a midline epithelial seam (Fig. 2), which rapidly degenerates, so establishing mesenchyme continuity across the intact horizontal palate. At approximately the same time as the midline epithelial cells die, the epithelia on the nasal aspect of the palate differentiate into pseudostratified ciliated columnar cells, whilst those on the oral aspect of the palate become stratified squamous, nonkeratinizing cells. Osteogenic blastemata for the palatal processes of the maxillary and palatine bones differentiate in the mesenchyme of the hard palate (and include secondary cartilages in the midline region of the palatal suture) whilst several patterned myogenic blastemata develop in the soft palate. Cleft palate may result from disturbances at any stage of palate development: defective palatal shelf growth, delayed or failed shelf elevation, defective shelf fusion, failure of medial edge cell death, postfusion rupture and failure of mesenchymal consolidation and differentiation (Ferguson, 1987). In birds, the bilateral palatal shelves arise from the maxillary processes, but instead of growing vertically (as in mammals) they initially develop and grow horizontally above the dorsum of the tongue (Shah & Crawford, 1980; Koch & Smiley, 1981; Ferguson etal. 1984; Ferguson & Honig, 1985; Shah, Cheng, Suen & Wong, 1985; Shah, Cheng, MacKay & Wong, 1987). The horizontal avian shelves approximate and contact each other, but their medial edge epithelia neither adhere, fuse nor die, but rather keratinize so that birds have a naturally cleft palate (Koch & Smiley, 1981; Ferguson el al. 1984; Ferguson & Honig, 1984, 1985; Shah etal. 1985, 1987). In amphibians and some reptiles e.g. certain species of turtles and snakes, the roof of the mouth is largely formed by a posterior growth of the primary palate which is of frontonasal process origin (Voeltzkow, 1903; Fuchs, 1907; Fleischmann, 1910). In other reptiles e.g. lizards, secondary palatal shelves arise from the maxillary processes, grow horizontally above the dorsum of the tongue, either never contact each other, resulting in a large choanal groove in the palate, or else contact each other but their medial edge epithelia do not adhere, fuse or die but keratinize resulting in natural cleft palate (Goppert, 1903; Hofmann, 1905; Sippel, 1907; Fuchs, 1908; Fleischmann, 1910). The general developmental scenario is analogous to that in birds. One group of reptiles; the crocodilians, however, have an intact, fused mammal-like secondary palate (Ferguson, \9%\a,b, 1984, 1985). In alligators and crocodiles, bilateral palatal shelves arise from the maxillary processes and in the anterior four fifths of the palate, grow horizontally above the dorsum of the tongue. In the posterior one fifth of the palate, the shelves grow vertically and remodel into a horizontal position later in development, except in their posterior extremities where they fuse with the floor of the mouth to form the basihyal valve (Ferguson, 1981a, 1984, 1985). Upon contact, however, the medial edge epithelia of alligator palatal shelves show a very restricted region (near the oral edge) of adherence, fusion and cell death. Instead the shelves establish mesenchymal continuity largely by a merging process with mesenchymal infilling and migration of the medial edge epithelia onto the nasal aspect of the palate (Ferguson, 1981a, 1984, 1985). A fused epithelial seam with medial edge cell death is therefore not a feature of alligator secondary palate development. Secondary palate development appears to be absent in lower vertebrates including fish. Phylogenetically, therefore, cleft palate is ancestoral to intact palate, initial vertical growth of the palatal shelves is peculiar to mammals, keratinization of the medial edge palatal epithelia (and consequently cleft palate) occurs in birds and some reptiles, limited medial edge cell adhesion, fusion and death, but massive cell migration (and intact palate) occurs in alligators and Palate development crocodiles, whilst medial edge epithelial cell adherence, fusion to form a seam and death (and intact palate) occurs in mammals. These species-specific morphogenetic and differentiative events also occur under identical chemically defined, serum-free culture conditions (Ferguson et al. 1984) and can be experimentally exploited to investigate fundamental mechanisms in palate development (Ferguson & Honig, 1984, 1985). Why do mammalian palatal shelves develop initially in a vertical direction? There has been almost no research on the initial phases of palatal shelf outgrowth from the maxillary processes, despite the fact that this is the developmental time at which most drugs are administered to experimentally induce cleft palate (Salomon & Pratt, 1979). Recently it has been shown that there are two peaks of DNA synthesis around the time of initial palate development, one corresponding with initial shelf outgrowth and the other with elongation of the shelf in a vertical direction (Burdett, Waterfield & Shah, 1988). Why mammalian palatal shelves initially grow vertically and subsequently elevate, as opposed to growing horizontally ab initio, as in birds and reptiles, is unresolved. Arguments relating to the volume of potential space in the primitive oronasal cavity with the evolution of the large muscular mammalian tongue and cheeks have been advanced (Shah, 1977; Ferguson, 1981) but remain intellectually unsatisfying. To date, no fossil or extinct animal with vertical palatal shelves in the adult state has been described so that phylogenetic arguments relating to the function of vertical palatal shelves in respiration, feeding or hearing are highly speculative. What is the mechanism of mammalian palatal shelf elevation? Numerous theories have been advanced to account for palatal shelf elevation (see Ferguson, 1978 for a review). In principle, an intrinsic force is progressively generated within the palatal shelves; once this reaches a threshold level which exceeds the force of resistance factors (e.g. frictional tongue resistance) shelf elevation occurs. Shelf elevation is a rapid event, probably occurring over a matter of minutes or hours in vivo (Ferguson, 1978; Brinkley, 1980). It involves a swinging 'flip up' mechanism in the anterior one third of the palate but an oozing remodelling 'flow' mechanism in the posterior two thirds of the palate (Figs 1, 2) (Brinkley & Morris-Wiman, 1984, 1987a). The intrinsic shelf elevating force is 43 multifactorial, but the chief component appears to be a regionally specific accumulation (Figs 3, 4) of glycosaminoglycans, predominantly hyaluronic acid, as development progresses (Pratt, Goggins, Wilk & King, 1973; Ferguson, 1978; Brinkley & MorrisWiman, 1984, 1987a,b). There is more hyaluronic acid in the anterior palate than posteriorly and more in the future oral aspect than the future nasal aspect (Knudsen, Bulleit & Zimmerman, 1985; Brinkley & Morris-Wiman, 1987a). Hyaluronic acid is a highly electrostatically charged, open coil molecule, capable of binding up to ten times its own weight in water. Small changes in the concentration of hyaluronic acid result in large changes in osmotic concentrations. Regional accumulation of hyaluronic acid results in swelling of the extracellular matrix and a corresponding decrease in mesenchyme cell density (Brinkley & Bookstein, 1986). This separation of cells may also be important in preventing contact and cell-cell, cellmatrix interactions at this stage of development: in later palatal development such interactions are critical and their onset corresponds with a decrease in hyaluronic acid content and an increase in cell density. Synthesis of hyaluronic acid by palatal mesenchyme cells is stimulated by epidermal growth factor (EGF) (Turley, Hollenberg & Pratt, 1985; Dixon, Foreman, Schor & Ferguson, 1988) and transforming growth factor beta (TGF/?) (Sharpe, Foreman, Carette, Schor & Ferguson, 1988; Sharpe & Ferguson, 1988). Vertical palatal shelves increase in size as much by extracellular matrix swelling as by mesenchymal cell division (Brinkley & Bookstein, 1986). The latter is greatest at the shelf tip (Jelinek & Dostal, 1974; Nanda & Romeo, 1975; Cleaton-Jones, 1976). The erectile shelf elevating force is partly directed by stout bundles of type I collagen which run down the centre of the vertical shelf from its base (where they are associated with the bony blastemata) to its tip (Fig. 5). Moreover the epithelial covering and associated basement membrane of the palatal shelf exhibit differential traction, which serve to constrain and direct the swelling osmotic force, in much the same way that the skin constrains an inflating balloon (Bulleit & Zimmerman, 1985; Brinkley, 1984). The alignment of mesenchymal cells within the core of the palatal shelf may further serve to direct the elevating force (Zimmerman & Wee, 1984). On the basis of in situ cell morphology, palatal mesenchyme has been divided into three principal regions which undergo different changes in cell shape and orientation during elevation (Babriarz, Wee & Zimmerman, 1979; Zimmerman & Wee, 1984). Palatal mesenchymal cells are themselves contractile (Zimmerman, Clark, Ganguli & Venkatasubramanian, 1983) and secrete various neurotransmitters e.g. serotonin, acetylcholine (Zimmerman & Wee, 1984). These neurotransmitters 44 M. W. J. Ferguson Fig. 1. Histological section through the anterior region of the palate in a day-13 embryonic mouse head. Note the vertical palatal shelves (p) firmly wedged against the tongue (t) in preparation for shelf elevation. x21. Fig. 2. Transverse histological section through the mid-palate region of an early day-14 and late day-14 mouse embryo. The palatal shelves first approximate each other, their medial edge epithelia (arrowed), then contact each other and fuse to form the midline epithelial seam. x21. Fig. 3. Transverse cryosection through the vertical palatal shelf of a day-13 mouse embryo in the anterior region. The section has been immunocytochemically stained for hyaluronic acid using the hyaluronectin/anti-hyaluronectin technique (Girard, Delpech & Delpech, 1986) and immunoperoxidase. Note the intense staining for hyaluronic acid within the palatal shelf mesenchyme. x84. Fig. 4. Transverse cryosection through the anterior palatal region of a day-14 mouse embryo immunocytochemically stained for hyaluronic acid using the hyaluronectin/anti-hyaluronectin technique. Note the concentration of hyaluronic acid along the medial edge mesenchyme adjacent to the midline epithelial seam. x84. affect both mesenchymal cell contractility and glycosaminoglycan degradation, and so may play a modulatory role in palate morphogenesis (Zimmerman & Wee, 1984). Palatal shelf elevation occurs in a conducive orofacial environment. During the period of shelf elevation, there is almost no growth in head width, but constant growth in head height (Diewert, 1978). This means that the position of least resistance for the expanding palatal shelves to occupy is above the dorsum of the tongue. Moreover, the tongue muscles become functional around the time of shelf elevation (Wragg, Smith & Borden, 1972) and fetal reflexes are present (Humphrey, 1968,1969). Recently it has been demonstrated that human embryos start to hiccup around the time of palatal shelf elevation, but that jaw opening, sucking and swallowing behaviour are not observed until after the time of palatal closure (De Vries, Visser & Prechtl, 1982, 1985). It is tempting to speculate that the profound pressure Palate development 45 Fig. 5. Transverse cryosection of a day-13 vertical anterior embryonic mouse palatal shelf immunocytochemically stained with antibodies against collagen type I. Note the stout bundles of collagen type I down the centre of the palatal shelf orientated from the base towards the tip. xl79. Fig. 6. Transverse cryosection through a day-14 fusing mouse palatal shelf. The medial edge epithelia are just contacting each other. The section has been stained immunocytochemically with antibodies against desmoplakin. Note the localization of these molecules on the surfaces of the medial edge epithelia (antibodies kindly supplied by Dr D. Garrod, Southampton). x450. changes produced in the oronasal cavity by hiccuping may be a trigger for rapid palatal shelf elevation. How does the epithelial seam form? After elevation, the palatal shelves approximate and contact each other, first in the region of the second ruga (middle third of the palate) from which point fusion spreads in posterior and anterior directions. The medial edge epithelia of opposing palatal shelves adhere to each other by means of a sticky cell surface glycoprotein coat (Greene & Kochhar, 1974; Pratt & Hassell, 1975; Souchon, 1975; Greene & Pratt, 1977) and desmosomes (De Angelis & Nalbandian, 1968; Morgan & Pratt, 1977) to form an epithelial seam. This is an interesting developmental phenomenon: (1) medial edge epithelial cells, which were originally superficial, develop cell adhesion molecules and desmosomes and become the central cells of the seam and (2) medial edge epithelial cell adherence is specific: the medial edge epithelia will not normally fuse with other epithelia e.g. floor of mouth, tongue, cheek etc. (Ferguson et al. 1984). Recently, we have demonstrated that mouse medial edge epithelial cells rapidly form desmosomes and accumulate desmoplakin - one of the desmosomal plaque proteins - on their superficial cell membranes just prior to shelf contact (Fig. 6). This suggests that desmosomal components are rapidly assembled, specifically in medial edge epithelial cells, just prior to and upon medial edge epithelial cell contact: this may be one of the mechanisms conferring specificity on medial edge epithelial cell adherence. The turnover rates of various desmosomal components prior to, during and after medial edge epithelial cell adherence may be an important controlling mechanism in palatal fusion and require investigation. Moreover, we have also produced monoclonal antibodies that recognize cell surface molecules on palatal epithelial cells (Dixon, White & Ferguson, 1988). These molecules vary in distribution by palatal region, by layers of the epithelium and with developmental time. One molecule is distributed between the layers of epithelial cells but is absent from their superficial surfaces in vertical palatal shelves. It appears on the superficial surface of the medial edge epithelia prior to and during shelf contact. It is tempting to speculate that these are epithelial cell adhesion molecules which might again confer specificity on medial edge epithelial cell adher- 48 M. W. J. Ferguson Fig. 7. Transmission electron micrograph through the midline epithelial seam (e) of a pair of fusing mouse palatal shelves. The specimen was prepared in tannic acid containing fixative. Note the intact basement membrane on both sides of the epithelial seam and the accumulation of numerous lysosomes within the medial edge epithelial cells. X39OO. Fig. 8. Transmission electron micrograph through a disrupted midline epithelial seam. Note the mesenchymal penetration and the resynthesis of the basement membrane along the epithelial seam cells (e). X3900. ence. None of the known cell adhesion molecules (Obrink, 1986) exhibit a similar staining pattern when localized immunocytochemically. Superficial medial edge epithelial cells also stain markedly with certain lectins e.g. ConA and Ulex europaeus agglutinin suggesting the presence of surface molecules rich in carbohydrate. Chick medial edge epithelia show none of these specializations of surface coat, do not form desmosomes and normally do not adhere. How does the epithelial seam disrupt? Almost as soon as the epithelial seam has formed (Fig. 7), it starts to thin ultimately becoming two or three cells thick. Thinning is achieved by expansion in palatal height (oronasally) and epithelial cell migration onto the oral and nasal aspects of the palate. The seam cells also rapidly accumulate lysosomal enzymes (Mato, Aikawa & Katahiva, 1966) and undergo apototic cell death (Figs 7,8). Palatal mesenchyme becomes continuous in areas where the seam has disrupted (Fig. 8). The basement membrane on each side of the epithelial seam remains intact even when it has completely thinned (Fig. 9) and a basal lamina is rapidly reconstituted around the ends of the seam or isolated epithelial islands, in areas of mesenchymal penetration (Fig. 8) (K. Hall & M. W. J. Ferguson, unpublished data). Medial edge epithelial cells cease DNA synthesis 24-36h prior to shelf contact (Hudson & Shapiro, 1973) and this has been referred to as programmed cell death. Cyclic AMP increases transiently just prior to shelf fusion, and much has been made of this observation in relation to competitive inhibitor studies of the prostaglandin, cyclic AMP, metabolic pathways (for reviews see Greene & Garbarino, 1984; Goldman, 1984; Pratt, 1984; Pisano & Greene, 1986; Ferguson, 1987). Exogenous cyclic AMP causes precocious medial edge epithelial cell death in mammals (Pratt & Martin, 1975) but does not induce it in birds (Tyler, 1986). Epidermal growth factor inhibits medial edge epithelial cell death (Hassell, 1975; Pratt, Kim & Groove, 1984; Pratt, 1984) but only in the presence of palatal mesenchyme (Tyler & Pratt, 1980): exogenous cyclic AMP blocks this EGFinduced inhibition (Hassell & Pratt, 1977). Despite a wealth of data on the biosynthesis and effects of cyclic AMP on palatal mesenchymal and epithelial cells (Greene & Garbarino, 1984; Goldman, 1984; Pratt, 1984; Pisano & Greene, 1986), it must be remembered that cyclic AMP, like intracellular calcium, pH and phosphatidylinositol lipids, is physiologically an intracellular second messenger. Its peak prior to shelf fusion suggests that it may be mediating differential gene expression, as a result of events at the cell surface e.g. binding of various molecules. Recently, we have demonstrated that epithelial cell death is not the only method of seam disruption. A large number (perhaps up to 50 %) of epithelial seam cells migrates into the palatal mesenchyme initially carrying with them fragments of their disrupted basement membranes (Fig. 10) (Fyfe & Ferguson, 1988; K. Hall & M. W. J. Ferguson, unpublished Palate development 47 Fig. 9. Cryosection through a pair of fusing palatal shelves in an embryonic day-14 mouse immunocytochemically stained with antibodies against type IV collagen. Note that the basement membrane is intact even though the seam is only one to two cells thick. Note also the staining of blood vessel basement membranes. X112. Fig. 10. Cryosection through an embryonic day-14 mouse head illustrating recently fused palatal shelves immunocytochemically stained with antibodies against type IV collagen. Note the disruption of the midline epithelial seam and the dispersion of clumps of basement membrane material with the epithelial cells which migrate into the mesenchyme. X179. Fig. 11. Cryosection through a horizontal day-14 embryonic mouse palatal shelf immunocytochemically stained with antibodies against tenascin. Note the fibrils of tenascin emerging from the basement membrane region and running perpendicularly into the mesenchyme particularly around the medial edge of the palatal shelf. X179. Fig. 12. Cryosection through an embryonic day-13 mouse palatal shelf immunocytochemically stained with a monoclonal antibody recognizing heparan sulphate proteoglycan. This specimen has been prepared using the immunogold silver enhancement technique. Note the distribution of heparan sulphate proteoglycan in the basement membrane on the stout collagen I bundles (Fig. 5), and on the surfaces of the palate mesenchymal cells. X179. 48 M. W. J. Ferguson data). These fragments disappear after a few hours, the cells lose their staining for cytokeratins and express vimentin intermediate filament staining and so quickly become indistinguishable from other palatal mesenchyme cells. For this reason, the ultimate developmental fate of these cells is unknown. It is unclear whether these migratory cells constitute a specific epithelial subpopulation e.g. basal stem cells which do not die. During seam disruption, the seam epithelial cells are less adherent to their basement membranes and display blebbing of their basal surfaces. They also develop well-organized bundles of microfilaments and microtubules in their cytoplasms. Interestingly in other cell systems increased cyclic AMP levels cause marked decreases in the numbers of cell surface fibronectin receptors (Allen-Hoffmann & Mosher, 1987), and it would be interesting to determine if this is true for palatal medial edge epithelial cells. Moreover, the basement membranes of the epithelial seam show a progressive loss of laminin, whilst fibrils of tenascin and type III collagen accumulate in the sub-basement membrane zone at right angles to the basement membrane (Fig. 11) (Fyfe & Ferguson, 1988; Fyfe, Ferguson & ChiquetEhrisman, 1988). The medial edge epithelial cells appear to migrate into the underlying mesenchyme along these fibrils. Chick medial edge epithelia do not cease DNA synthesis, show no peak of cyclic AMP (Greene et al. 1983) and no vertical fibrils of tenascin or type III collagen (Fyfe & Ferguson, 1988; Fyfe et al. 1988). Recently a mesenchymally produced soluble factor (scatter factor) has been isolated which causes the rapid disruption of epithelial junctions (Stoker, Gherardi, Perryman & Gray, 1987). Such a factor could be important in epithelial seam disruption. We therefore investigated the production of scatter factor by regionally derived clones of mouse embryonic palate mesenchyme cells from the oral, nasal and medial surfaces grown in vitro. The prediction was that medial edge mesenchymal cells would synthesize more scatter factor. Table 1 shows the results of scatter factor activity in the conditioned media of such cells. Although palate mesenchymal cells synthesize scatter factor, regional differences in production are insignificant, so that the role of scatter factor in palatal epithelial seam disruption is currently unclear and awaits experimental and immunological investigations. Does medial edge epithelial cell differentiation (death) depend upon shelf contact? Experiments in which mouse, alligator or chick palatal shelves were organ cultured either in isolation or Table 1. The scatter factor activity (units ml ') of palate mesenchymal cells derived from different regions of the palate (nasal, medial or oral) when cultured in vitro for various passages Palate mesenchyme Cell type Passage number 3 4 5 6 Nasal 24 48 24 24 Medial 24 48 12 24 Oral 48 48 24 48 These values are not statistically significant. Analysis by Sir Michael Stoker, FRS, IJniversity of Cambridge. in homologous pairs, revealed that medial edge epithelial cell differentiation (death in mice, migration in alligators, keratinization in chicks) occurred in single palatal shelves and so was independent of shelf contact (Ferguson et al. 1984). Regional differentiation of the nasal epithelia into pseudostratified ciliated columnar cells and the oral epithelia into stratified squamous cells also occurred in single palatal shelves of all three species. Each palatal shelf is therefore a developmental field with three defined regions of epithelial differentiation (nasal, medial, oral), one of which (medial) varies between vertebrate species. Is mouse palatal medial edge epithelial cell death a suicide or a murder? An extensive series of epithelial-mesenchymal recombination experiments has been performed including homologous, heterologous, heterochronic and isochronic combinations of mandibular, limb and palatal tissues both within and between mouse, chick and alligator embryos (Ferguson & Honig, 1984). In alligator and chick, the situation is simple. The mesenchyme signals nasal, medial and oral epithelial differentiation in a species-specific fashion, even to heterologous epithelia. The palatal epithelium appears to play a passive role, receiving its instructions from the underlying mesenchyme (Ferguson & Honig, 1984). The results in mice are similar, except that mouse palatal epithelia are biased to differentiate into nasal, medial and oral phenotypes from the onset of palatogenesis and will do so if placed on a 'neutral' e.g. mandibular, mesenchyme. Mouse palatal mesenchyme, however, signals regionally specific epithelial differentiation to heterologous epithelia or to palatal epithelia from other species e.g. chick, alligator at all stages of palatogenesis (Ferguson & Honig, 1984). If mouse palatal epithelial sheets are Palate development recombined with mouse mesenchyme with the original medial edge of the epithelial sheet at right angles to the medial edge of the mesenchyme (crossed recombination), then there is complete respecification of palatal epithelial differentiation into the regions defined by the orientation of the new mesenchyme. If, however, mouse palatal epithelia are recombined with alligator mesenchyme in a crossed recombination, then the alligator mesenchyme completely respecifies the nasal and oral fields and a new medial edge with a cobblestoned migratory phenotype typical of the alligator; but a line of cell death dividing the new oral and nasal fields still persists along the original medial edge of the mouse epithelial sheet (Ferguson & Honig, 1984). This suggests that there is some kind of signalling hierarchy between vertebrate mesenchymes and epithelia. In summary, epithelial-mesenchymal recombination experiments have demonstrated that nasal, medial and oral palatal epithelial differentiation is specified by the mesenchyme, and that signalling of medial edge epithelial differentiation goes across different vertebrates in a species-specific fashion (Ferguson & Honig, 1984). Medial edge epithelial cell death in mammals is therefore a murder by the underlying mesenchyme rather than an intrinsic epithelial suicide. What are the physlcochemlcal properties of the mesenchymal signal? In theory, there are four principle ways in which mesenchyme could signal epithelial differentiation; (1) extracellular matrix molecules, (2) soluble factors, (3) direct cell-cell contact, (4) combinations of 1-3. Extensive TEM surveys of the epithelial-mesenchymal interface at all stages of mouse palate development reveal that direct mesenchymal-epithelial cell contacts are very rare, even in the degenerating epithelial seam! (K. Hall & M. W. J. Ferguson, unpublished data). Direct cell-cell contact is therefore an unlikely signalling mechanism. There are, however, differences in the number and density of mesenchymal pseudopodial processes adjacent to the basement membrane at different developmental times and in different regions of the palate: these may be important in extracellular matrix synthesis/remodelling and soluble factor secretion. We have conducted extensive immunocytochemical surveys of all stages of the developing mouse and chick palate using affinity-purified polyclonal or monoclonal antibodies against: collagen types I-X, laminin, fibronectin, tenascin, heparan sulphate proteoglycan, chondroitin 0,4,6, sulphates; (Fyfe & Ferguson, 1988; Fyfe et al. 1988). Collagen types II, VIII and X are absent from the developing mouse palate. 49 Collagen types I, III, chondroitin sulphates and fibronectin are fairly ubiquitously distributed in the palate at all stages of development (Fig. 5) (Silver et al. 1981). These molecules are rapidly synthesized in the fusion zone following epithelial seam degeneration. Collagen type IV and laminin are present in the basement membranes of the palatal epithelia and blood vessels at all stages of development (Figs 9,10). Their distribution during seam disruption was described earlier. Collagen types V and VI and heparan sulphate proteoglycan (Fig. 12) are present in basement membranes and in certain regions of the mesenchyme e.g. type VI collagen is present around the nasal angle and in the midoral regions of an individual palatal shelf. Types V and VII collagen are found around epithelial cells particularly of the midline epithelial seam. Tenascin accumulates preferentially beneath the medial edge basement membranes of horizontal mouse palatal shelves (Fig. 11). Heparan sulphate proteoglycan is present in the basement membranes and on the mesenchyme cell surfaces, particularly in vertical tracts of collagen down the centre of the shelf (Figs 5 and 12). Type IX collagen is the most interesting extracellular matrix molecule in terms of signalling epithelialmesenchymal interactions in the mouse palate (Fyfe & Ferguson, 1988). Type IX collagen is absent from the basement membranes, mesenchymal and epithelial cell surfaces of the medial edge epithelia of day-12 to -13 embryonic mouse palates (Fig. 13). It is present on the cell surfaces of floor of the mouth epithelia at these stages (Fig. 13). However, just prior to shelf elevation type IX collagen appears on the cell surfaces of palatal medial edge epithelial cells (Fig. 14). This corresponds precisely with the time of signalling of medial edge epithelial differentiation as determined by recombination experiments (Ferguson & Honig, 1984), so the molecule appears in the right place at the right time. It is absent from developing chick palates. Cryoimmunoelectron microscopy reveals that type IX collagen is present at intersections of other collagen molecules (e.g. types III and I) with either themselves or the basal lamina (Fig. 15). It is also present on the medial edge epithelial cell surfaces (Fig. 16). We believe that type IX collagen acts as a linker molecule, physically and functionally connecting components of the extracellular matrix both with themselves and with the epithelial cell surface (Fyfe & Ferguson, 1988). If this is true, then its appearance at the time of signalling of epithelialmesenchymal interactions could represent a significant developmental control mechanism. Within such a hypothesis all of the elements of the signalling mechanism (different collagen types, basal lamina, cell surface) could be developed and in place, awaiting only the synthesis of a linker molecule (type IX 50 M. W. J. Ferguson collagen) to activate the mechanism. Type IX collagen has been described as linking together type II collagen fibrils at their intersections in cartilage (Muller-Glauser et al. 1986). Recently, a new collagen molecule, type XII, has been described, and it is suggested that this molecule acts as a minor collagen linking fibrils of types I and III collagen (Olsen, personal communication). As the types IX and XII molecules allegedly exhibit homology in at least one of the regions where our antibodies bind, it is possible that what we have described as type IX collagen in the palate, is in fact type XII. This possibility will be tested once details of type XII collagen are elucidated and published, and typespecific antibodies generated against both types IX and XII collagens. It may be that minor collagen types (including species yet to be described) play a crucial role in embryogenesis and that such a role need not be structural but, rather, predominantly one of directing morphogenesis and differentiation. Indeed, it could be intuitively argued that if the chief role of collagen was structural then it is unlikely that there would be so many different types, particularly of a minor variety. Knowledge that type IX collagen appears in the right place at the right time to be a candidate signalling molecule for mouse palatal medial edge epithelial cell differentiation, merely begs the question as to what controls the regional synthesis of type IX collagen. We have investigated this coincident with investigating the role of certain soluble factors as mesenchymal-signalling agents of epithelial differentiation. It is known that palate mesenchymal cells have large numbers of receptors for epidermal growth factor (EGF) (Yoneda & Pratt, 1981a,/>; Pratt et al. 1984) and that EGF (1) inhibits palatal medial edge epithelial cell death, but only in the presence of mesenchyme (Tyler & Pratt, 1980), and (2) stimulates oral epithelial cell division, hypertrophy and keratinization (Grove & Pratt, 1984) and mesenchymal synthesis of type V collagen and hyaluronic acid (Silver, Murray & Pratt, 1984; Turley et al. 1985). Transforming growth factor alpha (TCFcr) is likely the embryonic homologue of EGF. Transforming growth factor beta (TGF/3) is known to kill cells, particularly of epithelial lineage (Sporn, Roberts, Wakefield, de Crombrugghe, 1987; Cheifetz et al. 1987; Massague, 1987) and to stimulate fibronectin synthesis (Ignotz & Massague, 1986). It has been localized in the developing mouse palate (Heine et al. Fig. 13. Cryosection through the medial edge (m) and floor of the mouth epithelia (/) of an embryonic day-13 mouse head immunocytochemically stained with antibodies against type IX collagen. Note the absence of type IX collagen staining on the medial edge epithelia of the tip of the palatal shelf. Such staining is present in the floor of the mouth epithelia. X280. Fig. 14 Cryosection through an early day-14 embryonic mouse head illustrating the approximating medial edge epithelia (m) of the two palatal shelves immunocytochemically stained with an antibody against type IX collagen. Note the localization of type IX collagen around the surfaces of these medial edge epithelial cells. x280. Palate development 51 Fig. 15. Cryoelectron microscopy of the medial edge epithelia (w), basement membrane (b) and underlying mesenchyme of an embryonic day-14 mouse palatal shelf. The specimen has been prepared with 15 nm gold probes linked to a secondary antibody detecting the primary antibody against laminin which distributes in the basement membrane. The specimen is double labelled with 5nm immunogold probes identifying a type IX collagen antibody. Note the type IX collagen appears to be linking the basement membrane with an underlying type I collagen fibril (arrow), x 16500. Fig. 16. Cryotransmission electron micrograph of a medial edge epithelial cell (m) on a day-14 embryonic mouse palatal shelf. The specimen has been prepared with 5 nm immunogold probes detecting a type IX collagen antibody. Note that the type IX collagen is localizing on the surface of the epithelial cells. X30250. Fig. 17. Scanning electron micrograph of regionally derived mouse embryonic palatal mesenchymal cells (m) cultured to confluence on the surface of a type I collagen gel (c). X137. Fig. 18. Transmission electron micrograph illustrating an embryonic mouse palatal mesenchymal cell (m) embedded within a type I collagen gel. Note the collagen fibrils, the loose nature of the collagen gel and the reorganization of extracellular matrix molecules around the mesenchymal cell, x 11000. 1987; Sharpe & Ferguson, 1988) and is therefore a potentially interesting molecule. Acidic and basic fibroblast growth factors (FGF) or related molecules are known to be important in signalling epithelial mesenchymal interactions during amphibian gastrulation (Slack, Darlington, Heath & Godsave, 1987) and so are of potential interest in the palate. Plateletderived growth factor (PDGF) is important in wound healing (Lynch, Nixon, Colvin & Antoniades, 1987) and so of potential interest in terms of epithelial seam formation, degeneration and mesenchymal consolidation. We have therefore investigated the individual ef- fects of EGF, TGFar, TGF/3, PDGF, FGF acidic and basic (FGFa/FGFb), on mouse palatal development in organ culture, mesenchyme cell culture and epithelial cell culture (Sharpe et al. 1988; Sharpe & Ferguson, 1988; Dixon, Foreman, Schor & Ferguson, 1988). For mesenchyme cell culture the general strategy has been to determine the dose-response curves for the growth factor and then to study its effects on cell division and extracellular matrix biosynthesis when mouse palatal mesenchyme cells are cultured on plastic, on collagen films and within three-dimensional hydrated collagen gels (Figs 17, 18) i.e. physiologically relevant substrata. Sum- 52 M. W. J. Ferguson Fig. 19. Cryosection of a type I collagen gel with mouse embryonic palatal mesenchymal cells cultured on its surface (see Fig. 17). The section has been stained with antibodies against type IX collagen and virtually none is present. The cultures were performed in the presence of 2-5 % donor calf serum in the DMEM/F12 media, x 132. Fig. 20. Cryosection of a type I collagen gel with embryonic mouse palatal mesenchymal cells cultured on its surface. These cells were cultured in the presence of DMEM/F12 media supplemented with 2-5 % donor calf serum and lOngml"' EGF. The section is immunocytochemically stained for type IX collagen. Note the appearance of type IX collagen in a basement-membrane-like structure on the surface of the collagen gel. Type IX collagen synthesis has been stimulated by epidermal growth factor (compare with Fig. 19). X132. Fig. 21. Immunocytochemical staining of the matrix deposited by mouse embryonic palatal mesenchymal cells on a plastic glass slide immunocytochemically stained with antibodies against fibronectin. The embryonic palatal mesenchymal cells were cultured in the presence of transforming growth factor /3 which stimulated the synthesis of fibronectin. xl68. Fig. 22. Scanning electron micrograph of a mouse palatal epithelial sheet cultured on the surface of a type I collagen gel which had previously been conditioned (see Figs 19-21) by the prior culture of mouse embryonic palatal mesenchymal cells on the surface of the gel followed by lysis of these cells using ammonia. The epithelial sheet was derived from a late day-13 mouse embryo and was therefore already determined. Note differentiation of the palatal epithelia into nasal pseudostratified ciliated columnar cells (n), medial edge cell death (m) and oral stratified squamous cells (O). X180. marized, these so called growth factors have major specific effects on extracellular matrix biosynthesis which are much more important than their mitogenic effects when cells are cultured in a physiologically relevant environment. TGF/3 increases markedly the synthesis of fibronectin and also of collagen types III, IV and V and possibly IX (Fig. 21). It inhibits the synthesis of type I collagen and only increases the synthesis of hyaluronic acid in sparse cultures but not in dense ones (Sharpe etal. 1988; Sharpe & Ferguson, 1988). TGF/3 inhibits palatal mesenchymal cell growth and kills some cells. EGF (or TGFa) stimu- ates the synthesis of all collagen types and increases the synthesis of glycosaminoglycans and proteoglycans (Figs 19, 20). Interestingly EGF stimulates the synthesis of collagen which is underhydroxylated (Dixon et al. 1988). EGF (or TGFo-) increases the rate of palatal mesenchymal cell division, but less so when cells are cultured in or on collagen gels. PDGF stimulates cell division in a similar way (but to a lesser extent) to EGF, but has no effect on extracellular matrix biosynthesis. FGFa or FGFb inhibits extracellular matrix biosynthesis, which is interesting in view of its previously documented effect of Palate development stimulating cell migration during gastrulation (Slack et al. 1987). Therefore, the synthesis of extracellular matrix molecules including type IX collagen is stimulated by certain soluble factors e.g. TG¥a, TGF0 known to be present in the palate and inhibited by others e.g. FGF. We have investigated the effects of these various combinations of extracellular matrix molecules on palatal epithelial differentiation by lysing the palatal mesenchymal cells off the conditioned gel surface using ammonia and plating on sheets of isolated palatal epithelia (Fig. 22). To date, we have been able to maintain the differentiation of already determined palatal epithelial sheets in these mesenchyme-free culture conditions (Fig. 22): experiments are in progress to investigate the effects of these conditioned matrices in directing epithelial differentiation. It should also be noted that many soluble factors bind to extracellular matrix molecules such as heparan or fibronectin (Smith, Singh, Lillguist, Goon & Stiles, 1982; Fava & McClure, 1987) and this may be important both in the embryo and in interpreting these conditioned gel experiments. Thus one population of mesenchymal cells could secrete extracellular matrix molecules and bound growth factors. These may remain either inactive or locally active (Smith et al. 1982; Fava & McClure, 1987) until at a subsequent developmental time the ECM is remodelled by another population of cells, the bound growth factors released and exert their effects on this new population of cells. Cause and effect may therefore be separated by developmental time. Mouse palatal shelves have been organ cultured in the presence of these various growth factors (M. J. Dixon, D. M. Fyfe & M. W. J. Ferguson, unpublished data). No effects were noted in cultures at the air-gas interface, presumably due to the failure of the growth factor to penetrate the tissue despite the omission of Millipore filters as a culture substratum. By contrast in submerged organ cultures effects were noted. In EGF-supplemented cultures, the medial edge of the palatal shelf develops a nipple-like bulge; interestingly, this is the area in vivo where cells have the highest mitotic indices (Jelnick & Dostal, 1974; Nanda & Romeo, 1975; Cleaton-Jones, 1976). The mesenchyme had increased quantities of extracellular matrix molecules, medial edge epithelial cell death was absent and the medial edge plus oral epithelia were heavily keratinized. In TGF/3-supplemented cultures, the epithelia were very thin and poorly differentiated, MEE cell death was marked, mesenchyme cells were clumped and the fibronectin content of the mesenchyme was increased. These results indicate that exogenous growth factors exert effects on organ-cultured mouse palatal shelves in a fashion 53 similar to their effects in cell culture and that controlled physiological levels of such factors may be important in mouse palate development. How Is mesenchymal signalling of epithelial differentiation regionally specified? Our model of mesenchymal signalling of terminal palatal epithelial differentiation is complex and involves a dynamic interaction between soluble growth factors and the extracellular matrix. The model supposes that the mesenchyme produces a growth factor, or factors, which may (1) act directly on the epithelia to induce a phenotype, (2) act by autocrine stimulation of extracellular matrix production by mesenchyme cells and these extracellular matrix molecules may then directly influence epithelial differentiation. Additionally, the extracellular matrix may modulate the response of epithelia to growth factors or vice versa. The system may selfregulate via a biphasic response of the epithelia and mesenchyme to the same growth factor i.e. the factor may directly affect the epithelia and the mesenchyme, the extracellular matrix molecules produced by the mesenchyme may then modulate (e.g. turn off) the direct effects of the factor on the epithelia. Regional specification of epithelial differentiation may depend upon the clonal heterogeneity of palatal epithelial and mesenchymal cells. Not all the palatal mesenchymal cells produce all the extracellular matrix molecules known to be present (Sharpe et al. 1988; Dixon et al. 1988). Heterogeneity in extracellular matrix production therefore exists. Not all the palatal mesenchymal cells respond to soluble growth factors (Sharpe et al. 1988; Dixon et al. 1988) either in relationship to mitosis or extracellular matrix production. This may reflect the presence or absence of receptors for these various growth factors on the mesenchyme cells. Preliminary immunocytochemical studies of the regional localization of the EGF receptor during different stages of mouse palate development suggest that there is regional heterogeneity (M. W. J. Ferguson, unpublished data). When the palatal shelves are vertical, the mesenchyme cells beneath the medial edge appear to lack EGF receptors, but the latter appear when the shelves are horizontal and the epithelial seam is degenerating (Fig. 23). Growth factors and extracellular matrix molecules may cause the clonal expansion of subpopulations of mesenchyme cells which may be important in the regional signalling of epithelial mesenchymal interactions (Schor & Schor, 1987). Palatal mesenchyme cells are heterogeneous not only in terms of production of extracellular matrix molecules and response to soluble factors but also in 5* M. W. J. Ferguson Fig. 23. (A) Histological section through a day-13 vertical mouse palatal shelf which had been stained en bloc with antibodies against the epidermal growth factor receptor. These antibodies were then detected via secondary antibodies linked to colloidal gold particles which were enhanced using the silver enhancement technique. The specimen is viewed in epipolarizing light and the EGF receptor appears as a white spot. Note the distribution of this receptor in the nasal and oral regions particularly in the core of the palatal shelf, but its absence from the mesenchymal cells at the tip of the palate. This absence is a real result and not due to a penetration-of-antibody technical problem as we have deliberately disrupted the epithelium at the tip of the palatal shelf. (B) A similar preparation of a pair of horizontal fusing palatal shelves in a day-14 mouse embryo. Note the appearance of mesenchymal cells expressing the EGF receptor adjacent to the midline epithelial seam (e). xl69. growth factor production (Heine et al. 1987). Synthesis of growth factors at particular locations within the palate may give rise to diffusion gradients and thus to differing regional responses by both epithelial and mesenchymal cells. In this context, it should be noted that the same growth factor can stimulate cell division at one concentration and inhibit it at another; populations of cells can respond differently to the same concentration of a growth factor depending upon their density and the nature of their substratum (Sporn et al. 1987; Massague, 1987). Gradients of growth factors within the developing palate may also be generated by (1) the binding and sequestration of growth factors in specific locations by binding proteins or extracellular matrix molecules, (2) their presence in amniotic fluid and local diffusion into palatal tissues via the oronasal cavity. Gradients of growth factors may be superimposed upon gradients of receptors on responding cells and the density and substrata of the latter. The situation is therefore potentially highly complex and interactive. Moreover different growth factors interact with each other to produce either synergistic or antagonistic effects, thus layering on another complicating tier (Sharpe et al. 1988; Dixon et al. 1988; Sporn et al. 1987). Palatal epithelial tissues may also be heterogeneous in terms of their response (e.g. possession of receptors) to either growth factors or extracellular matrix molecules. Investigation of such a complex model is not as daunting as it might appear. First, we have recently demonstrated that regionally derived, uncloned populations of mesenchyme cells from the nasal, medial, and oral regions of the palate, exhibit differences in response (extracellular matrix pro- Palate development duction and mitosis) to various growth factors even after several passages in vitro (M. J. M. Carette & M. W. J. Ferguson, unpublished data). It is therefore possible to conduct mesenchyme conditioning/epithelial target experiments on collagen gels (as described in the previous section) using either regionally derived or cloned palatal mesenchyme cells. Second, we have produced monoclonal antibodies against specific cell surface determinants on mouse palatal nasal, medial and oral epithelial cells, before, during and after epithelial differentiation (Fig. 25) (Dixon, White & Ferguson. 1988). These antibodies can be used to regionally sort (using FACS) isolated palatal epithelial populations into regional types which can then be used in targeting experiments using cloned mesenchyme cells. Such a dynamic interactive model of mesenchyme signalling is even more intriguing when it is realized (1) that specific extracellular matrix molecules can bind and sequester growth factors (Smith et al. 1982; Fava & McClure, 1987) and (2) that certain growth factors (e.g. TGF#) can stimulate the expression of cell adhesion protein receptor sites in mouse epithelial cell lines (Ignotz & Massague, 1987). Thus, presence of a certain growth factor could stimulate extracellular matrix production by mesenchyme cells, bind locally to this newly synthesized matrix and simultaneously stimulate the expression of receptors for such matrix and soluble factors in adjacent epithelial cells: epithelia and mesenchyme would then interact. In this context, soluble factors such as EGF, TGFa-, TGF£, PDGF, FGF are somewhat misnamed as 'growth factors'. They act more like 'differentiation factors' during embryonic development and their principal effect is not on mitogenesis when cells are investigated in a physiologically relevant situation (Sharpe etal. 1988; Dixon etal. 1988). Interestingly, it has recently been shown that proteins encoded by pattern-forming genes in Drosophilia are homologous with EGF and TGF/3 (Padgett, Johnston & Gelbart, 1987). How do the responding epithelial cells process the epigenetlc signals? It is hypothesized that palatal epithelial cells possess receptors for extracellular matrix and soluble factors. Apart from some data for the EGF receptor (Yoneda & Pratt, 1981a,b) nothing is known about the nature or affinity of such receptors, or how they vary by region or with developmental time. Changes in the extracellular matrix substrata underlying the epithelia may affect the polarity and cytoskeletal organization of the latter. Such shape changes may directly induce differential gene expression and/or affect the dispo- 55 sition, numbers and affinity of other receptors on the cell surface e.g. for soluble factors. Most events at the cell surface are transduced by second messenger systems, intracellular: cyclic nucleotides, calcium, pH and phosphatidylinositol lipids. There are extensive data on the regulation of palatal cyclic AMP by prostaglandins, some data on cyclic GMP, few data on phosphatidylinositol lipids and almost no data on intracellular pH and calcium (reviewed in Greene & Garbarino, 1984; Goldman, 1984; Pisano & Greene, 1986; Ferguson, 1987). Surprisingly, there have been few investigations of second messenger systems using defined palatal cell populations - even epithelia or mesenchyme! There have been equally few studies of the chain of intracellular events following stimulation of the cell surface with one particular factor either extracellular matrix or soluble. How do these signals result In the terminal differentiation of the epithelial cells? Ultimately, this question asks what genes and their encoded proteins make a nasal cell pseudostratified ciliated columnar, an oral cell stratified squamous and a medial edge cell in the mouse die? We have addressed this issue by trying to develop better molecular markers for nasal, medial and oral epithelial cells. First, we have mapped immunocytochemically the distribution of cytokeratin and vimentin intermediate filaments in epithelial and mesenchymal cells during mouse and and chick palate development (Ferguson, Fyfe & Lane, 1988). In mouse, cytokeratin 18 is present only in nasal epithelia, not in medial edge nor oral epithelia (Fig. 24A). It appears progressively from the palatal shelf nasal base (day 12) to the shelf tip (day 14-5) corresponding with the progressive differentiation of the nasal epithelia into pseudostratified ciliated columnar cells. Cytokeratins 1 and 19 are present in all epithelia at all stages of development. Cytokeratin 8 is present in the future nasal and oral regions of vertical palatal shelves but absent from the medial edge epithelia (Fig. 24B). Cytokeratin 10 appears in the oral epithelia at day 14 when the shelves are fused. Vimentin is present in shelf mesenchyme cells and appears in medial edge epithelia at the time of midline epithelial seam disruption. Second, we have produced monoclonal antibodies that recognize cell surface determinants on mouse nasal, medial and oral epithelia at different stages of development (Dixon, White & Ferguson, 1988). Of the 104 antibody secreting hybridomas produced, four (designated 2H6, 3H2, 6A11, 6D3) are of considerable interest (Fig. 25). 2H6 and 3H2 recog- 56 M. W. J. Ferguson Fig. 24. (A) Histological section through the superior region of a vertical palatal shelf (p) in a day-13 embryonic mouse head. The specimen has been immunocytochemically stained with antibodies against cytokeratin 18. Note the presence of this cytokeratin in the developing nasal epithelia at the superior aspect of the palatal shelf and their absence from the adjacent tongue (r). x375. (B) A similar preparation stained with an antibody against cytokeratin 8. Note this cytokeratin is present in the future nasal and oral epithelia but is absent from the tip of the palatal shelf and the future medial edge epithelia. xl69. Fig. 25. Cryosection through a day-13 embryonic mouse palatal shelf in the anterior region. The specimen has been stained with monoclonal antibody 6D3 which recognizes future nasal and some basal medial edge epithelial cells but not future oral epithelia. Note also the staining in the tongue (r). x210. (B) A similar preparation stained with monoclonal antibody 6A11 which recognizes predominantly cells in the medial edge of the palatal shelf and also on the future nasal aspect as well as the tongue (/). p, palatal shelf. x84. Palate development nize an epitope which is present on palatal epithelia from the onset of palatogenesis but which progressively disappears as regional differentiation occurs. 6D3 recognizes an epitope which is expressed in differentiating epithelia in a specific temporal and spatial pattern. 6A11 recognizes an epitope which is expressed on the tips of the palatal shelves just prior to fusion i.e. in determined and differentiating medial edge epithelia. It should also be noted that these cells exhibit a specific lectin-staining pattern (see earlier section entitled 'How does the epithelial seam form?'). Thus the molecular message that makes a nasal cell pseudostratified ciliated columnar or an oral cell stratified squamous may involve the synthesis of a specific repertoire of cytokeratins and cell surface (? adhesion) molecules, not to mention receptors for various matrix and soluble factors. Use may be made of these molecular markers of regional palatal epithelial differentiation for experimental analysis of the nature of mesenchyme signalling of epithelial differentiation. What are the Important candidate genes for normal and cleft palate? At a phenomenological level the different developmental mechanisms (e.g. failure of shelf elevation, inhibition of cell division, excessive head width etc.) causing cleft palate have been summarized previously (Ferguson, 1987). It follows from the previous account that the genes coding for the following molecules could be important in normal and cleft palate development: extracellular matrix molecules particularly hyaluronic acid, minor collagen types e.g. type IX, tenascin; soluble factors e.g. EGF, PDGF, TGFo-, TGF/3, FGF, cytokeratins, cell surface molecules. Moreover, as many aspects of the intracellular signalling pathway utilized by growth factors can be subverted by oncogene products, it is likely that certain proto-oncogenes e.g. erb b, c cis etc, may be particularly important in palate development. Adamson (1987) has recently summarized the literature on proto-oncogenes in developmental systems and the differential expression of a panel of proto-oncogenes during mouse palate development is currently being investigated in my laboratory. There are also data implicating the H-2 locus in cleft palate development (Bonner & Slavkin, 1975; Goldman, 1984) whilst a gene involved in X-linked cleft palate has recently been mapped to the Xql3-Xq21 regions of the human X chromosome (Moore et al. 1987). However, in general genes controlling pattern formation in the palate or indeed the head are virtually unknown. To date few homeo- 57 tic genes have been localized in the cranial region. However, there exists the tantalizing possibility that mechanisms of pattern formation in the head (and palate) may be similar to that elsewhere in the body, for instance the limbs. The investigation of such possibilities awaits the isolation of homeotic genes localizing in the head and palate. Work reported in this review has been and is supported, by grants from: The Medical Research Council, Wellcome Trust, Action Research for the Crippled Child and Birthright. Numerous collaborators and students have made important contributions especially Larry Honig, Seth Schor, David Garrod, Shirley Ayad, Anne White, David Fyfe, Mike Dixon, Paul Sharpe, David Foreman, Martin Carette, Katy Hall and Bill Moser. References ADAMSON, E. D. (1987). Oncogenes in development. Development 99, 449-471. ALLEN-HOFFMANN, R. L. & MOSHER, D. F. (1987). Matrix assembly sites for exogenous fibronectin are decreased on human fibroblasts after treatment with agents which increase intracellular cAMP. J. biol. Chem. 262, 14361-14365. BABIARZ, B. S., W E E , E. L. & ZIMMERMAN, E. F. (1979). Palate morphogenesis. III. Changes in cell shape and orientation during shelf elevation. Teratology 20, 249-278. BONNER, J. J. & SLAVKIN, H. C. (1975). Cleft palate susceptibility linked to histocompatibility-2 (H-2) in the mouse. Immunogenetics 2, 213-218. BRINKLEY, L. L. (1980). In vitro studies of palatal shelf elevation. In Current Research Trends in Prenatal Craniofacial Development (ed. R. M. Pratt & R. L. Christiansen), pp. 203-220. New York: Elsevier/North Holland. BRINKLEY, L. L. (1984). Changes in cell distribution during mouse secondary palate closure in vivo and in vitro. I. Epithelial cells. Devi Biol. 102, 216-227. BRINKLEY, L. L. & BOOKSTEIN, F. L. (1986). Cell distribution during mouse secondary palate closure. II. Mesenchymal cells. J. Embryol. exp. Morph. 96, 111-130. BRINKLEY, L. L. & MORRIS WIMAN, J. (1984). Role of extracellular matrices in palatal shelf closure. In Palate Development: Normal and Abnormal Cellular and Molecular Aspects (ed. E. F. Zimmerman), Current Topics in Developmental Biology 19, 17-36. New York: Academic Press. BRINKLEY, L. L. & MORRIS WIMAN, J. (1987a). Computer- assisted analysis of hyaluronate distribution during morphogenesis of the mouse secondary palate. Development 100, 629-636. BRINKLEY, L. L. & MORRIS WIMAN, J. (19876). Effects of chlorcyclizine-induced glycosaminoglycan alterations on patterns of hyaluronate distribution during morphogenesis of the mouse secondary palate. Development 100, 637-640. 58 M. W. J. Ferguson R. F. & ZIMMERMAN, E. F. (1985). The influence of the epithelium on palate shelf reorientation. J. Embryol. exp. Morph. 88, 265-279. /3 system, a complex pattern of cross-reactive ligands and receptors. Cell 48, 409-415. CLEATON JONES, P. (1976). Radioautographic study of mesenchymal cell activity in the secondary palate of the rat. J. dent. Res. 55, 437- 440. DE ANGELIS, V. & NALBANDIAN, J. (1968). Ultra structure of mouse and rat palatal processes prior to and during secondary palate formation. Archs Oral. Biol. 13, 601-608. Billett & P. Maderson), pp. 454-720. New York: J. Wiley & Sons. FERGUSON, M. W. J. (1987). Palate development: mechanisms and malformations (Conway Review Lecture). Irish Journal of Medical Sciences 156, 309-315. FERGUSON, M. W. J., FYFE, D. & LANE, B. (1988). The cytokeratin profile of embryonic mouse palatal epithelia. J. Dent. Res. (in press). FERGUSON, M. W. J. & HONIG, L. S. (1984). Epithelialmesenchymal interactions during vertebrate palatogenesis. In Current Topics in Developmental Biology, vol. 19 Palate Development: Normal and Abnormal, Cellular and Molecular Aspects' (ed. E. F. Zimmerman), pp. 137-164. New York: Academic Press. FERGUSON, M. W. J. & HONIG, L. S. (1985). Experimental fusion of the naturally-cleft embryonic, chick palate. J. of Craniofac. Genet, devl Biol. SI, 323-337. DE VRIES, J. I. P., VISSER, G. H. A. & PRECHTL, H. F. FERGUSON, M. W. J., HONIG, L. S. & SLAVKIN, H. C. BULLEIT, BURDETT, D. N., WATERFIELD, J. D. & SHAH, R. M. (1988). Vertical development of the secondary palate in hamster embryos following exposure to 6mercaptopurine. J. Craniofac. Genet, devl Biol. (in press). CHEIFETZ, S., WEATHERBEE, J. A., TSANG, M. L. S., ANDERSON, J. K., MOLE, J. E., LUCAS, R. & MASSAGUE, J. (1987). The transforming growth factor R. (1982). The emergence of fetal behaviour. I. Qualitative aspects. Early Human Development 7, 301-322. DE VRIES, J. I. P., VISSER, G. H. A. & PRECHTL, H. F. R. (1985). The emergence of fetal behaviour. II. Quantitative Aspects. Early Human Development 12, 99-120. DIEWERT, V. M. (1978). A quantitative coronal plane evaluation of cranio-facial growth and spatial relations during secondary palate development in the rat. Archs Oral Biol. 23, 607-629. DIXON, M. J., FOREMAN, D., SCHOR, S. L. & FERGUSON, M. W. J. (1988). Epidermal growth factor and transforming growth factor alpha regulate extracellular matrix production by embryonic mouse palatal mesenchymal cells cultured on a variety of substrata. Development (in press). DIXON, M. J., WHITE, A. & FERGUSON, M. W. J. (1988). Monoclonal antibodies recognising stage and region specific epitopes on mouse palatal epithelium. Differentiation (in press). FAVA, R. A. & MCCLURE, D. B. (1987). Fibronectinassociated transforming growth factor. /. cell. Physiol. 131, 184-189. FERGUSON, M. W. J. (1978). Palatal shelf elevation in the Wistar rat fetus. J. Anat. 125, 555-577. FERGUSON, M. W. J. (1981a). The structure and development of the palate in Alligator mississippiensis. Archs Oral Biol. 26, 427-443. FERGUSON, M. W. J. (1981£>). The value of the American alligator (Alligator mississippiensis) as a model for research in craniofacial development. J. Craniofac. Genet, devl Biol. 1, 123-144. FERGUSON, M. W. J. (1984). Craniofacial development in Alligator mississippiensis. In The Structure, Development and Evolution of Reptiles (ed. M. W. J. Ferguson), pp. 223-273. London: Academic Press. FERGUSON, M. W. J. (1985). The reproductive biology and embryology of crocodilians. In Biology of the Reptilia, vol. 14, Development (ed. C. Gans, F. S. (1984). Differentiation of cultured palatal shelves from alligator, chick and mouse embryos. Anat. Rec. 209, 231-249. FLEISCHMANN, A. (1910). Uber den Begriff Gaumen. Morph. Jb. 41, 681- 707. FUCHS, H. (1907). Untersuchungen uber Ontogenie und Phylogenie der Gaumenbildungen bei den Wirbeltieren Erste Mitteilung Uber den Gaumen der Schildkroten und seine Entwicklungsgeschichte. Z. Morph. und Anth. 10, 409-463. FUCHS, H. (1908). Untersuchungen uber Ontogenie und Phylogenie der Gaumenbilding bei den Wirbeltieren Zweite Mitteilung. Uber das Munddach der Rhynchocephalen, Saurier, Schlangen, Krokodile und Sauger und den zusammenhong zwrischen Mund und Nasenhohle bei diesen Tieren. Z. Morph. u Anth. 11, 153-248. FYFE, D., FERGUSON, M. W. J. & CHIQUET-EHRISMANN, R. (1988). Tenascin immunolocalisation during palate development in mouse and chicken embryos. Anat. & Embryol. (in press). FYFE, D. M. & FERGUSON, M. W. J. (1988). Immunocytochemical localisation of collagen types I—XII, proteoglycans, laminin and fibronectin during mouse secondary palate development. Devl Biol. (in press). GIRARD, N., DELPECH, A. & DELPECH, B. (1986). Characterization of hyaluronic acid on tissue sections with hyaluronectin. J. Histochem. Cytochem. 34, 539-541. GOLDMAN, A. S. (1984). Biochemical mechanism of glucocorticoid and phenytoin induced cleft palate. Curr. Top. devl Biol. 19, 217-240. GOPPERT, E. (1903). Die Bedeutung der Zunge fur den sekundaren Gaumen und den Ductus nasopharyngeus. Morphol. Jahrb. 31, 311-359. GREENE, R. M. & KOCHHAR, D. M. (1974). Surface coat on the epithelium of developing palatine shelves in the mouse as revealed by electron microscopy. J. Embryol. exp. Morph. 31,683-692. Palate development GREENE, R. M. & GARBARINO, M. P. (1984). Role of cyclic AMP, prostaglandins and catecholamines during normal palate development. Curr. Top. devl Biol. 19, 65-80. GREENE, R. M. & PRATT, R. M. (1976). Developmental aspects of secondary palate formation. J. Embryol. exp. Morph. 36, 225-245. GREENE, R. M. & PRATT, R. M. (1977). Inhibition by diazo-oxo-norlucine (DON) of rat palatal glycoprotein synthesis and epithelial cell adhesion in vitro. Expl Cell. Res. 105, 27-37. 59 ANTONIADES, H. N. (1987). Role of platelet-derived growth factor in wound healing: Synergistic effects with other growth factors. Proc. natn. Acad. Sci. U.S.A. 84, 7696-7700. MASSAGUE, J. (1987). The TGF/3 family of growth and differentiation factors. Cell 49, 437-438. MATO, M., AIKAWA, E. & KATAHIRA, M. (1966). Appearance of various types of lysosomes in the epithelium covering lateral palatine shelves during a secondary palate formation. Gunma J. med. Sci. 15, 46-56. GREENE, R. M., SHAH, R. M., LLOYD, M. R., CRAWFORD, MELNICK, M., BIXLER, D. & SHIELDS, E. D. (1980). The B. J., SUEN, R., SHANFELD, J. L. & DAVIDOVITCH, Z. Aetiology of Cleft Lip and Cleft Palate. New York: Alan Liss. (1983). Differentiation of the Avian Secondary Palate. J. exp. Zool. 225, 43-52. GROVE, R. I. & PRATT, R. M. (1984). Influence of epidermal growth factor and cyclic AMP on growth and differentiation of palatal epithelial cells in culture. Devi Biol. 106, 427-437. HASSELL, J. R. (1975). The Development of rat palatal shelves in vitro. An ultrastructural analysis of the inhibition of epithelial cell death and palate fusion by the epidermal growth factor. Devi Biol. 45, 90-102. HASSELL, J. R. & PRATT, R. M. (1977). Elevated levels of cAMP alters the effect of epidermal growth factor in vitro on programmed cell death in the secondary palatal epithelium. Expl Cell. Res. 106, 55-62. HEINE, V. I., MUNOZ, E. F., FLANDERS, K. C , ELLINGSWORTH, L. R., LAM, H. Y. P., THOMPSON, N. L., ROBERTS, A. B. & SPORN, M. B. (1987). The Role of transforming growth factor-beta in the development of the mouse embryo. J. Cell Biol. 105, 2861-2867. HOFMANN, O. (1905). Das Munddach der Saurier. Morphol. Jahrb. 33, 3-38. HUDSON, C. D. & SHAPIRO, B. L. (1973). An autoradiographic study of deoxyribonucleic acid synthesis in embryonic rat palatal shelf epithelium with reference to the concept of programmed cell death. Archs Oral Biol. 18, 77-84. HUMPHREY, T. (1968). The development of mouth opening and related reflexes involving the oral area of human fetuses. Alabama J. med. Sciences 5, 126—157. HUMPHREY, T. (1969). The relation between human fetal mouth opening reflexes and closure of the palate. Am. J. Anal. 125, 317-344. IGNOTZ, R. A. & MASSAGUE, J. (1987). Cell adhesion protein receptors as targets for transforming growth factor j8 action. Cell 51, 189-197. JELINEK, R. & DOSTAL, M. (1974). Morphogenesis of cleft palate induced by exogenous factors VII. Mitotic activity during formation of the mouse secondary palate. Folia Morphologica (Praha) 22, 94—101. KNUDSEN, T. B., BULLEIT, R. F. & ZIMMERMAN, E. F. (1985). Histochemical localization of glycosaminoglycans during morphogenesis of the secondary palate in mice. Anal. Embryol. 173, 137-142. KOCH, W. E. & SMILEY, G. R. (1981). In vivo and in vitro studies of the development of the avian secondary palate. Archs Oral Biol. 26, 181-187. LYNCH, S. E., NIXON, J. C , COLVIN, R. B. & MOORE, G. E., IVENS, A., CHAMBERS, J., FARRALL, M., WILLIAMSON, R., PAGE, D. C , BJORNSSON, A., ARNASON, A., JENSSON, O. (1987). Linkage of an X chromosome cleft palate gene. Nature, Lond. 326, 91-92. MORGAN, P. R. & PRATT, R. M. (1977). Ultrastructure of the expected fusion zone in rat fetuses with diazo-oxonorleucine (D.O.N.) induced cleft palate. Teratology 15, 281-290. MULLER-GLAUSER, W., HUMBEL, B., GLATT, M., STRAULI, P., WINTERHALTER, K. H. & BRUCKNER, P. (1986). On the role of type IX collagen in the extracellular matrix of cartilage: type IX collagen is localised to intersections of collagen fibrils. J. Cell Biol. 102, 1931-1939. NANDA, R. & ROMEO, D. (1975). Differential cell proliferation of embryonic rat palatal processes as determined by incorporation of tritiated thymidine. Cleft Palate J. 12,436-443. OBRINK, B. (1986). Epithelial cell adhesion molecules. Expl Cell Res. 163, 1-21. PADGETT, R. W., ST JOHNSTON, R. D. & GELBART, W. M. (1987). A transcript from a Drosophilia pattern gene predicts a protein homologous to the transforming growth factor-/? family. Nature, Lond. 325, 81-84. PISANO, M. M. & GREENE, R. M. (1986). Hormone and growth factor involvement in craniofacial development. IRCS Medical Science 14, 635-640. PRATT, R. M. (1984). Hormones, Growth Factors and their receptors in normal and abnormal prenatal development. In Issues and Reviews in Teratology vol. 2 (ed. H. Kalter), pp. 189-217. New York: Plenum Press. PRATT, R. M. & CHRISTIANSEN, R. L. (1980). Current Research Trends in Prenatal Craniofacial Amsterdam: Elsevier/North-Holland. Development. PRATT, R. M. & HASSELL, J. R. (1975). Appearance and distribution of carbohydrate rich macromolecules on the epithelial surface of the rat palatal shelf. Devi Biol. 45, 192-198. PRATT, R. M., KIM, C. S. & GROOVE, R. I. (1984). Role of glucocorticoids and epidermal growth factor in normal and abnormal palatal development. Curr. Top. devl Biol. 19, 81-101. PRATT, R. M., GOGGINS, J. F., WILK, A. L. & KING, C. T. G. (1973). Acid mucopolysaccharide synthesis in the secondary palate of the developing rat at the time of 60 M. W. J. Ferguson rotation and fusion. Devl Biol. 32, 230-237. PRATT, R. M. & KING, C. T. G. (1971). Collagen synthesis in the secondary palate of the developing rat. Archs Oral Biol. 16, 1181-1185. PRATT, R. M. & MARTIN, G. R. (1975). Epithelial cell death and cyclic AMP increase during palatal development. Proc. natn Acad. Sci. U.S.A. 72, 874-877. SALOMON, D. S. & PRATT, R. M. (1979). Involvement of glucocorticoids in the development of the secondary palate. Differentiation 13, 141-154. SCHOR, S. L. & SCHOR, A. M. (1987). Clonal Heterogenity in Fibroblast Phenotype: Implications for the Control of Epithelial-Mesenchymal Interactions. Bio Essays 7, 200-204. SHAH, R. M. (1977). Palatomandibular and maxillo mandibular fusion, partial aglossia and cleft palate in a human embryo. Report of a case. Teratology 15, 261-272. SHAH, R. M. (1979). Current concepts on the mechanisms of normal and abnormal secondary palate formation. In Advances in the Study of Birth Defects, vol. 1, Teratogenic Mechanisms (ed. T. V. N. Persaud), pp. 69-84. Lancaster: M.T.P. Press Ltd. SHAH, R. M., CHENG, K. M., MCKAY, R. A. & WONG, A. (1987). Secondary palate development in the domestic duck (Khaki Campbell). An electron microscopic, histochemical, autoradiographic and biochemical study. J. Anat. 154, 245-258. SHAH, R. M., CHENG, K. M., SUEN, R. & WONG, A. (1985). An ultrastructural and histochemical study of the development of the secondary palate in Japanese quail. Coturnix coturnix japonica. J. Craniofac. Genet, devl Biol. 5, 41-57. SHAH, R. M. & CRAWFORD, B. J. (1980). Development of the secondary palate in chick embryo: a light and electron microscopic and histochemical study. Invest, cell. Path. 3, 319-328. SHARPE, P. M., FOREMAN, D. N., CARETTE, M. J. M., SCHOR, S. L. & FERGUSON, M. W. J. (1988). The effects of transforming growth factor beta (TGF/3) on the proliferation and biosynthetic activity of mouse embryonic palatal mesenchymal cells in vitro. Development (in press). SHARPE, P. M. & FERGUSON, M. W. J. (1988). Mesenchymal influences on epithelial differentiation in developing systems. J. Cell Sci. Suppl. (in press). SILVER, M. H., FOIDART, J.-M. & PRATT, R. M. (1981). Distribution of fibronectin and collagen during mouse limb and palate development. Differentiation 18, 141-149. SILVER, M. H., MURRAY, J. C. & PRATT, R. M. (1984). Epidermal growth factor stimulates type-V collagen synthesis in cultured murine palatal shelves. Differentiation 27, 205-208. SIPPEL, W. (1907). Das munddach der Vogel und Saurier. Morph. Jb. 47, 490-524. SLACK, J. M. W., DARLINGTON, B. G., HEATH, J. K. & GODSAVE, S. F. (1987). Mesoderm Induction in early Xenopus embryos by heparin-binding growth factors. Nature, Lond. 326, 197-200. SMITH, J. C , SINGH, J. P., LILLGUIST, J. S., GOON, D. S. & STILES, G. D. (1982). Growth factors adherent to cell substrata are mitogenically active in situ. Nature, Lond. 296, 154-156. SOUCHON, R. (1975). Surface coat of the palatal shelf epithelium during palatogenesis in mouse embryos. Anat. Embryol. 147, 133-142. SPORN, M. B., ROBERTS, A. B., WAKEFIELD, L. M. & DE CROMBRUGGHE, B. (1987). Some recent advances in the chemistry and biology of transforming growth factor beta. /. Cell Biol. 105, 1039-1045. STOKER, M., GHERARDI, E., PERRYMAN, M. & GRAY, J. (1987). Scatter factor is a fibroblast-derived modulator of epithelial cell mobility. Nature, Lond. 327, 239-242. TURLEY, E. A., HOLLENBERG, M. D. & PRATT, R. M. (1985). Effect of epidermal growth factor/urogastrone on glycosaminoglycan synthesis and accumulation in vitro in the developing mouse palate. Differentiation 28, 279-285. TYLER, M. S. (1986). Effects of dibutyryl cyclic AMP and theophylline on in vitro development of the secondary palate in the embryonic chick. Archs Oral Biol. 31, 261-265. TYLER, M. S. & PRATT, R. M. (1980). Effect of epidermal growth factor on secondary palatal epithelium in vitro: tissue isolation and recombination studies. J. Embryol. exp. Morph. 58, 93-106. VOELTZKOW, A. (1903). Gesichtsbildung und Entwicklung der ausserven Korperform bei Chelone imibricata Schweigg. Abh. Senck. Naturf. Gesell. 27, 181-190. WRAGG, L. E., SMITH, J. A. & BORDEN, C. S. (1972). Myoneural maturation and function of the foetal rat tongue at the time of secondary palate closure. Archs Oral Biol. 17, 673-682. YONEDA, T. & PRATT, R. M. (1981a). Mesenchyme cells from the human embryonic palate are highly responsive to epidermal growth factor. Science 213, 563-565. YONEDA, T. & PRATT, R. M. (19816). Interaction between glucocorticoids and EGF in vitro in the growth of palatal mesenchyme cells from the human embryo. Differentiation 19, 194-198. ZIMMERMAN, E. F. (ed.) (1984). Palate development: Normal and Abnormal Cellular and Molecular Aspects. Curr. Topics devl Biol. vol. 19. New York: Academic Press. ZIMMERMAN, E. F., CLARK, R. L., GANGULI, S. & VENKATASUBRAMANIAN, K. (1983). Serotonin regulation of palatal cell motility and metabolism. J. Craniofac. Genet, devl Biol. 3, 371-385. ZIMMERMAN, E. F. & WEE, E. L. (1984). Role of neurotransmitters in palate development. In Palate Development: Normal and Abnormal Cellular and Molecular Aspects (ed. E. F. Zimmerman), Curr. Topics devl Biol. 19, 37-63.