Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
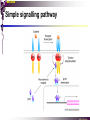
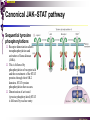
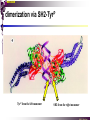
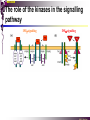
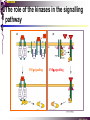
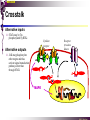
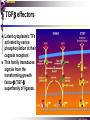
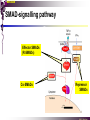
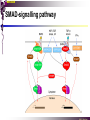
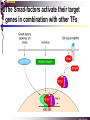
The STAT family MBV4230 Class IIB(3)(b) latent cytoplasmic factors These families not present in fungi or plants, hinting at an important evolutionary divergence leading to animals. MBV4230 STATs - a signal responsive TF family STATs: Signal Transducers and Activators of Transcription 1. Transducers for signals from many cytokines two functions given in the name Broad spectrum of biological effects 2. Transcriptional activators characteristic activation mechanism activation at the cell membrane, response in the nucleus Rapid signal response The activation/deactivation cycle of STAT molecules is quite short, about 15 min for an individual molecule. MBV4230 Simple signalling pathway MBV4230 The JAK-STAT signalling pathway Function: regulation of gene expression in response to cytokines 1. cytokines bind and aggregate the cytokine receptors in the cell membrane 2. associated JAK-type tyrosine kinases are activated by aggregation and tyrosine-phosphorylates neighbouring-JAK (transphosphorylation) as well as the C-terminal tail of the receptor (multiple sites) 3. Tyr-phosphates recruit inactive STAT-factors in the cytoplasm which are bound through their SH2-domains 4. STATs become tyrosine-phosphorylated by JAK 5. phosphorylated STATs dissociate, dimerize (homo-/hetero-) and migrate to the nucleus 6. STAT-dimers bind DNA and activates target genes MBV4230 Canonical JAK–STAT pathway Sequential tyrosine phosphorylations Receptor dimerization allows transphosphorylation and activation of Janus kinases (JAKs). This is followed by phosphorylation of receptor tails and the recruitment of the STAT proteins through their SH-2 domains. STAT tyrosine phosphorylation then occurs. Dimerization of activated (tyrosine phosphorylated) STAT is followed by nuclear entry. MBV4230 IFN-response: two variants signalling pathway first discovered in studies of interferon-response (IFN) IFN/ IFN/ activation of Jak1+Tyk2 DNA-binding complexes (trimer: STAT1+STAT2+p48, together designated ISGF3) activation of target genes with ISRE (IFN-stimulated response element) IFN IFN activation of Jak1+Jak2 DNA-binding complex (dimer: 2x STAT1) activation of target genes having GAS elements (IFN activated sequence) MBV4230 IFN-response: two variants MBV4230 STAT-family members STAT1 - involved in IFN/- and IFN-response STAT2 - involved in IFN/-response Mainly acting as partner for STAT1/p48 STAT3 - involved in response to several cytokines including IL6. It activates several genes involved in acute phase response Important in growth regulation, embryonic development & organogenesis Activation of STAT3 correlated with cell growth, link to cancer, bind c-Jun STAT4 - involved in IL12-response STAT5a & 5b - involved in response to several cytokines including prolactin, IL-2, and regulates expression of milk proteins in breast tissue in response to prolactin STAT6 - involved in IL4-response non-mammalian family members (e.g. Drosophila) MBV4230 STAT-members SH2 Y MBV4230 STAT-STAT interaction occurs through reciprocal phospho-Tyr - SH2 interactions SH2-domain SH2 = “Src-homology domain 2” function: phospho-tyrosine binding Three important functions in STATs: important for recruitment of STAT to receptor important for interaction with the JAK kinase important for dimerization of STATs to an active DNA-binding form Tyr-701 conserved key Tyr residue located just C-terminal to SH2 essensiell for dimerdannelse to an active DNA-binding form function: TyrP bindingssted for SH2 in partner P + Y P Y Y MBV4230 dimerization via SH2-TyrP TyrP from the left monomer SH2 from the right monomer MBV4230 STAT-members SH2 Y MBV4230 STATs - structure and function dimerization Reciprocal SH2- TyrP interaction Homodimers Heterodimers (STAT1)2 STAT1-STAT2 STAT1-STAT3 GAS= TTN5-6AA ISRE= AGTTTN3TTTCC DNA-binding domain DBD located in the middle of the protein Unique motif - se next slide All DBDs bind similar motifs in DNA symmetric inverted half sites Only difference to STATs: preference for central nucleotide MBV4230 STAT-DBD structure Known structures [STAT1]2-DNA and [STAT3]2-DNA, as well as an N-terminal of STAT4 Characteristic feature of DBD Symmetry-axis through DNA, each monomer contacts a separate half site structure resembles NFkB and p53 (immunoglobuline fold). The dimer forms a Cshaped ”clamp” around DNA. The dimer is kept together by reciprocal SH2- TyrP interactions between the SH2 domain in one monomer and the phosphorylated Tyr in the other. The SH2 domain in each monomer is closely linked to the core DBD and is itself close to DNA, and is assumed also to contribute to DNA-binding. N-terminal coiled-coil region not close to DNA, probably involved in prot-prot interaction with flexible position MBV4230 3D STAT domain structure and protein binding sites. MBV4230 Promoter recognition and selectivity Mechanisms to achieve specific trx responses. Inverted repeat TTN5–6AA motif common. Binding specificity to individual elements based on evolved preferences for specific positions. In the ISGF3 heterotrimeric complex, STAT1–STAT2 heterodimers bind to a third protein, p48/ISGF3, a TF that recognizes the ISRE sequence. STAT N-domains mediate dimer–dimer interactions allowing high-avidity binding to tandemly arranged lowaffinity GAS elements. Adjacent response elements bind to other TFs. Cooperativity and synergy. STAT directly recruit co-activators that alter chromatin dynamics. MBV4230 TAD transactivation domain C-terminal part of the protein, less conserved variants generated by alternative splicing + proteolysis STAT1 lacking the last 38aa has all functions retained except transactivation Regulation through TAD-modification Activity of TAD is regulated through Ser phosphorylation (LPMSP-motif) Ser727 in STAT1 Kinase not identified - candidates: p38, ERK, JNK A role in recruitment of GTF/coactivator Proteins identified that bind TAD in a Ser-dependent manner MCM5 BRCA1 TAD in STAT2 binds C/H-rich region of CBP STAT2 carries the principal TAD of the ISGF3-complex MBV4230 Other functional domains The N-domain is important for stabilizing interactions between STAT dimers, bound to tandemly arranged response elements Tyr kinases MBV4230 The JAK-family of tyrosine kinases Family members JAK1 (135 kDa) JAK2 (130 kDa) JAK3 (120 kDa) Tyk2 (140 kDa) Common feature C-terminal kinase + pseudokinase ≠ RTK by lacking transmembrane domains and SH2, SH3, PTB, PH several regions homologous between JAK-members Associated with cytokine receptors (type in and II) Function Associated with cytokine receptors in non-stimulated cells in an inactive form MBV4230 The role of the kinases in the signalling pathway INF-signalling INF-signalling MBV4230 The cytokine-receptor superfamily A receptor-family that mediates response to more than 30 different cytokines Common feature: conserved extracellular ligand-binding domain Are associated with tyrosine-kinases in the JAK-family Ligand-binding Receptor dimerization or oligomerization leads to JAK apposition associated JAK Tyr kinases are activated transphosphorylation of neighbour-JAKs tyrosine-phosphorylation of C-terminal tail of receptors on multiple sites several cellular substrate-proteins associate (including STATs) multiple signalling pathways are activated MBV4230 The role of the kinases in the signalling pathway INF-signalling INF-signalling MBV4230 Specificity in response Specific cytokines activate distinct STATs and lead to a specific response - what mediate specificity? each cytokine activates a subgroup STAT some cytokines activate only one specific STAT One contribution: the SH2 - receptor interaction specific for certain combinations swaps-experiments of SH2 between STATs change specificity affinity of the SH2-receptor interaction is affected by the sequence context of the Tyr Another contribution: different STAT-dimers bind different response elements in the genome and turn on different genes STAT1 knock-out mice illustrate biological specificity STAT1-/- phenotype: total lack of IFN-response highly sensitive to virus-infection MBV4230 Several signalling pathways linked STATs may also be Tyr-phosphorylated and hence activated by other receptor families receptor tyrosine kinases (RTKs) such as EGF-receptor may phosphorylate STATs EGF stimulation activation of STAT1, STAT3 non-receptor tyrosine kinases such as Src and Abl may also phosphorylate STATs (?) G-protein coupled 7TMS receptors such as angiotensine receptor (?) STAT may also be modified by Ser-phosphorylation DNA-binding reduced (STAT3) Transactivationdomain Ser-phosphorylated (important for transactivation in STAT1 and STAT3) Responsible kinases not identified - MAPkinases candidates, probably also others JAKs may activate other signalling pathways than STATs TyrP will recruit several protein-substrates and lead to phosphorylation and activation of other signalling pathways e.g. JAK activation activation of MAP-kinases e.g. substrates: IRS-1, SHC, Grb2, HCP, Syp, Vav MBV4230 Crosstalk Alternative inputs Alternative outputs JAK may phosphorylate other targets and thus activate signal transduction pathways other than through STATs Receptor tyrosinee kinase Cytokine receptor P P P JAK P P P P P P Y P MAPK Y STATs may be Tyrphosphorylated by RTKs Y P Y SH2 MBV4230 Variations in mechanisms of STAT activation SMAD family MBV4230 SMAD-family - a logic resembling the STAT-family The Smad-factors mediate response to TGF-related growth- and differentiation factors STAT-related logic Membrane-bound receptors (such as the TGFß-receptor) are activated by binding of ligand (TGF). The receptors here are transmembrane serine/threonine-kinases Activated kinases phosphorylate specific Smad-factors phosphorylated Smad-factors associate with a common Smad-factor (Smad4) The generated heteromeric complexes migrate to the nucleus as transcription factors MBV4230 TGF effectors Latent cytoplasmic TFs activated by serine phosphorylation at their cognate receptors This family transduces signals from the transforming growth factor- (TGF-) superfamily of ligands. MBV4230 Classification Smad-factors - design and classification Nine different Smad-factors identified in vertebrates common conserved domains: N-terminalt MH1-domain (DBD) + C-terminalt MH2domain Can be divided into three groups 1. Receptor-activated Smad-factors - become phosphorylated by activated receptors in their C-terminal (SSXS) 2. common Smad-factors associated with activated Smad-factors and participate in several signalling pathways 3. Inhibitoriske Smad-factors MBV4230 SMAD-signalling pathway Effector SMADs (R-SMADs) Co-SMADs Repressor SMADs MBV4230 Three groups of SMADs First group: The effector SMADs (also called the R-SMADs) become serine-phosphorylated in the C-terminal domain by the activated receptor. Smad1, Smad5, Smad8, and Smad9 become phosphorylated in response to bone morphogenetic morphogenetic protein (BMP) and growth and differentiation factor (GDF), and Smad2 and Smad3 become phosphorylated in response to the activin/nodal branch of the TGF- pathway. Second group: regulatory or co-SMADs (common SMADs). There are two regulatory SMADs: Smad4 and Smad4 (also called Smad10). Smad4 binds to, and is essential for, the function of Smad1 and Smad2. The regulatory Smad4 binds to all effector SMADs in the formation of transcriptional complexes, but it does not appear to be required for nuclear translocation of the effector molecules. Third group: two inhibitory SMADs, Smad6 and Smad7. provide negative regulation of the pathway by blocking Smad4 binding. MBV4230 SMAD-signalling pathway MBV4230 Final steps - target gene activation Once an activated, serine-phosphorylated effector SMAD binds Smad4 and escapes the negative influences of Smad6 and Smad7, nuclear accumulation and regu-lation of specific target genes can occur. In most cases, SMADs require partner transcription factors with strong DNA binding capacity that determine the gene to be activated. The DNA binding is then strengthened by association with SMADs that on their own bind weakly to adjacent DNA sites. The SMADs furnish transcriptional activation capacity. The specificity of response among different ligands can be partially explained by the choice of DNA binding partner proteins. For example, activin activation of SMADs results in combinations with FAST1 and a particular set of genes is activated. Signaling by BMP ligands results in association of activated SMADs with a DNA binding protein called OAZ. MBV4230 The Smad-factors activate their target genes in combination with other TFs