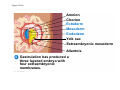
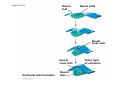
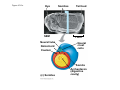
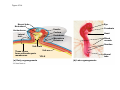
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
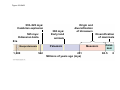
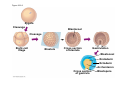
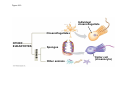
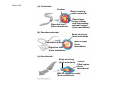
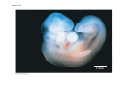
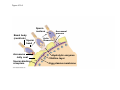
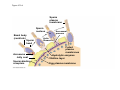
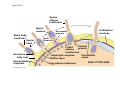
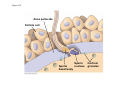
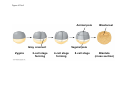
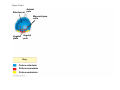
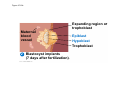
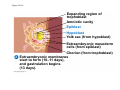
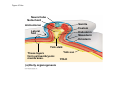
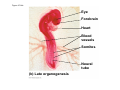
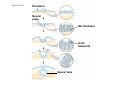
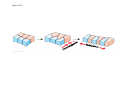
ANIMAL DEVELOPMENT E l ti off Evolution Development (Evo/Devo) ( ) evolutionaryy developmental bi l biology Figure 47.6 50 m (a) Fertilized egg (b) Four-cell stage (c) Early blastula ( ) Later blastula (d) Figure 27.UN01 Eukarya Archaea Bacteria Figure 32.11 Ct Ctenophora h Eumeta azoa Me etazoa ANCESTRAL COLONIAL FLAGELLATE Porifera Cnidaria Acoela Bilateria Chordata Platyhelminthes L Lophotro ochozoa a Ecdysozoa Deute erostomiia Echinodermata Rotifera Ectoprocta Brachiopoda Mollusca Annelida Nematoda Arthropoda ontogeny recapitulates phylogeny phylogeny"" (ORP ORP)) 我們的身體裡有一條魚 我們的身體裡有 條魚 Recapitulation theory The theory of recapitulation, recapitulation, often expressed as "ontogeny recapitulates phylogeny"" (ORP phylogeny ORP))- is a hypothesis that in developing from embryo to adult, animals go through g stages g resembling g or representing p g successive stages in the evolution of their remote ancestors.. Th conceptt originated The i i t d iin th the 1790 1790s among the German Natural philosophers and, as proposed p p by y Étienne Serres in 1824– 1824–26,, became known as the "Meckel"Meckel-Serres Law". In 1866, the German zoologist Ernst Haeckel proposed that the embryonic development of an individual organism (its ontogeny) followed the same path as the evolutionary history of its species (its phylogeny). Ontogeny recapitulates Phylogeny"" (ORP Phylogeny ORP)) Ontogeny recapitulates Ph l Phylogeny can nott explain l i chick development p This is clearly not the case — a fact recognized by many scientists even when the idea of ontogeny recapitulating phylogeny was introduced. If you observe a chick's chick s development, development you will find that the chick embryo may resemble the embryos of reptiles and fish at points in its development, but it doesn't recapitulate the forms of its adult ancestors. Root of EvoEvo-Devo Edward B. Lewis discovered homeotic genes rooting the emerging discipline of genes, evo--devo in molecular genetics. evo genetics. In 2000, a special section of the Proceedings of the National Academy of Sciences (PNAS) was devoted to "evo"evo evo-devo devo",and and an entire 2005 issue of the Journal of Experimental Zoology Part B: Molecular and Developmental Evolution was devoted to the key evoevo-devo topics of evolutionary innovation and morphological novelty novelty. Edward B. Lewis a Nobel Prize winner in 1995 His Nobel Prize winning studies with Drosophila,, (including the discovery of the Drosophila Drosophila Bithorax complex and elucidation of its function) function), founded the field of developmental genetics and laid the groundwork for our current understanding of th universal, the i l evolutionarily l ti il conserved d strategies controlling animal development. He is credited with development of the complementation l t ti test. test t t. His Hi kkey publications bli ti iin the fields of genetics genetics,, developmental biology, biology, radiation and cancer are presented in the book G Genes, Development and Cancer, Cancer C , which was released in 2004. Model Og i Organisms 生物 experimental i t l models d l Escherchia coli 大腸菌 Saccharomyces cerevisiae Dictyostelium discoideum 酵母菌 黏菌 Dictyostelium y discoideum 黏菌 Caenorhabditis elegans 秀麗線蟲 Drosophila melanogaster Xenopus laevis 蛙 果蠅 Danio rerio 斑馬魚 Arabidopsis p thaliana 阿拉伯芥 The mouse as a model for human development 人 小鼠 Ontogeny recapitulates Phylogeny Phylogeny"" (ORP ORP)) This idea is an extreme one. If it were strictly true, it would predict, for example, that in the course of a chick's development, it would go through the following stages: g a single g celled organism, g a multi-celled invertebrate ancestor, a fish, a lizard-like reptile, an ancestral bird,, and then finally, y, a babyy chick. Ancestral characters are often, but not always, preserved in an organism organism'ss development. For example, both chick and human embryos go through a stage where they have slits and arches in their necks like the gill slits and gill arches of fish. These structures are not gills and do not develop into gills in chicks and humans humans, but the fact that they are so similar to gill structures in fish at this p point in development p supports the idea that chicks and humans share a common ancestor with fish. Thus, developmental characters, along with other lines of evidence, can be used for constructing phylogenies LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 47 Animal Development p Lectures by Erin Barley Kathleen Fitzpatrick © 2011 Pearson Education, Inc. Figure 32.11 Ct Ctenophora h Eumeta azoa Me etazoa ANCESTRAL COLONIAL FLAGELLATE Porifera Cnidaria Acoela Bilateria Chordata Platyhelminthes L Lophotro ochozoa a Ecdysozoa Deute erostomiia Echinodermata Rotifera Ectoprocta Brachiopoda Mollusca Annelida Nematoda Arthropoda Figure 32.10 Porifera Cnidaria Eum metazoa M Metazoa ANCESTRAL COLONIAL FLAGELLATE Ctenophora Pro otostomiia Billateria Deutero ostomia a Ectoprocta Brachiopoda p Echinodermata Chordata Platyhelminthes R tif Rotifera Mollusca Annelida Arthropoda Nematoda Figure 32.UN02 535–525 mya: Cambrian explosion 565 mya: Ediacaran biota 365 mya: Early land animals Origin and diversification of dinosaurs Diversification of mammals Era Paleozoic Neoproterozoic 1,000 542 251 Millions of years ago (mya) Mesozoic Cenozoic 65.5 0 Concept : Animal are multicellular, heterotrophic eukaryotes with tissues that d l from develop f embryonic b i layers l • Th There are exceptions ti to t nearly l every criterion it i ffor distinguishing animals from other life-forms • Several S l characteristics, h t i ti ttaken k ttogether, th sufficiently ffi i tl define the group • Animals A i l are h heterotrophs t t h th thatt iingestt th their i ffood d © 2011 Pearson Education, Inc. Cell Structure and Specialization • Animals are multicellular eukaryotes • Their cells lack cell walls g by y structural • Their bodies are held together proteins such as collagen • Nervous tissue and muscle tissue are unique, defining characteristics of animals • Tissues are groups of cells that have a common structure, function, or both © 2011 Pearson Education, Inc. Reproduction and Development • Most animals reproduce sexually sexually, with the diploid stage usually dominating the life cycle • After a sperm fertilizes an egg egg, the zygote undergoes rapid cell division called cleavage • Cleavage Cl leads l d to t formation f ti off a multicellular, lti ll l hollow blastula • The blastula undergoes gastrulation, forming a gastrula with different layers of embryonic tissues © 2011 Pearson Education, Inc. Figure 32.2-1 Zygote Z t Cleavage Eight-cell stage g Figure 32.2-2 Zygote Z t Cleavage Blastocoel Cleavage Eight-cell stage g Blastula Cross section of blastula Figure 32.2-3 Zygote Z t Cleavage Blastocoel Cleavage Eight-cell stage g Blastula Cross section of blastula Gastrulation Blastocoel Endoderm Ectoderm Archenteron Cross section of gastrula Bl t Blastopore RESULTS 1 Early stages of d development l 100 m Figure 32.6 2 32-cell stage Site of gastrulation 3 Early gastrula stage g 4 Embryos with blocked -catenin activity Site of gastrulation • Many animals have at least one larval stage • A larva is sexually immature and morphologically distinct from the adult; it eventually undergoes metamorphosis • A juvenile j il resembles bl an adult, d lt but b t iis nott yett sexually mature © 2011 Pearson Education, Inc. • Most animals, animals and only animals animals, have Hox genes that regulate the development of body form • Although the Hox family of genes has been highly conserved, it can produce a wide diversity of animal morphology © 2011 Pearson Education, Inc. Concept : The history of animals spans more than half a billion years • The animal kingdom includes a great diversity of living species and an even greater diversity of extinct ones • The Th common ancestor t off living li i animals i l may h have lived between 675 and 800 million years ago • This ancestor may have resembled modern choanoflagellates, protists that are the closest living relatives of animals © 2011 Pearson Education, Inc. Figure 32.3 Individual choanoflagellate Ch Choanoflagellates fl ll t OTHER EUKARYOTES Sponges An nimals Other animals Collar cell (choanocyte) Neoproterozoic Era (1 Billion Billion–542 542 Million Years Ago) • Early members of the animal fossil record include the Ediacaran biota biota, which dates from 565 to 550 million years ago © 2011 Pearson Education, Inc. Mesozoic Era (251 (251–65 65.55 Million Years Ago) • Coral reefs emerged emerged, becoming important marine ecological niches for other organisms • The ancestors of plesiosaurs were reptiles that returned to the water • During D i th the M Mesozoic i era, di dinosaurs were th the dominant terrestrial vertebrates • The first mammals emerged gp plants and insects diversified • Flowering © 2011 Pearson Education, Inc. Cenozoic Era (65.5 Million Years Ago to the Present) • The beginning of the Cenozoic era followed mass extinctions of both terrestrial and marine animals • These extinctions included the large, nonflying di dinosaurs and d th the marine i reptiles til • Mammals increased in size and exploited vacated ecological niches global climate cooled • The g © 2011 Pearson Education, Inc. Concept : Animals can be characterized by “body plans” • Zoologists sometimes categorize animals according to a body plan, plan a set of morphological and developmental traits • Some S developmental d l t l characteristics h t i ti are conservative For example, the molecular control of gastrulation is conserved among diverse animal groups © 2011 Pearson Education, Inc. Symmetry • Animals can be categorized according to the symmetry of their bodies, or lack of it • Some animals have radial symmetry symmetry, with no front and back, or left and right © 2011 Pearson Education, Inc. Figure 32.7 (a) Radial symmetry (b) Bilateral symmetry • Two Two-sided sided symmetry is called bilateral symmetry • Bilaterally symmetrical animals have – – – – A dorsal (top) side and a ventral (bottom) side A right and left side Anterior (head) and posterior (tail) ends Cephalization, the development of a head © 2011 Pearson Education, Inc. • Radial animals are often sessile or planktonic (drifting or weakly swimming) • Bilateral animals often move actively and have a central nervous system © 2011 Pearson Education, Inc. Tissues • Animal body plans also vary according to the organization of the animal’s tissues • Tissues are collections of specialized cells isolated from other tissues by membranous layers • During D i d development, l t th three germ llayers give i rise i tto the tissues and organs of the animal embryo © 2011 Pearson Education, Inc. • Ectoderm is the germ layer covering the embryo’s embryo s surface • Endoderm is the innermost germ layer and lines the developing digestive tube, called the archenteron © 2011 Pearson Education, Inc. Figure 32.8 (a) Coelomate Coelom Digestive tract (from endoderm) Body covering (f (from ectoderm) t d ) Tissue layer lining coelom and d suspending di internal organs (from mesoderm) (b) Pseudocoelomate Body covering (from ectoderm) Pse docoelom Pseudocoelom Digestive tract (from endoderm) Muscle layer (from mesoderm) (c) Acoelomate Body covering Tissue(f (from ectoderm) d ) Ti filled region (from mesoderm) Wall of digestive cavity (from endoderm) • Coelomates and pseudocoelomates belong to the same grade • A grade is a group whose members share key biological features • A grade d iis nott necessarily il a clade, l d an ancestor t and all of its descendents © 2011 Pearson Education, Inc. Protostome and Deuterostome Development • Based on early development development, many animals can be categorized as having protostome development or deuterostome development © 2011 Pearson Education, Inc. Cleavage • In protostome development development, cleavage is spiral and determinate • In deuterostome development development, cleavage is radial and indeterminate • With iindeterminate d t i t cleavage, l each h cellll iin th the early l stages of cleavage retains the capacity to develop i t a complete into l t embryo b • Indeterminate cleavage makes possible identical twins, and embryonic stem cells © 2011 Pearson Education, Inc. Figure 32.9 Protostome development (examples: molluscs, annelids) ( ) Cleavage (a) Cl Deuterostome development (examples: echinoderms, chordates) Eight-cell stage Eight-cell stage Spiral and determinate Radial and indeterminate (b) Coelom formation Coelom Archenteron Coelom Mesoderm Blastopore Blastopore Solid masses of mesoderm split and form coelom. (c) Fate of the blastopore Mesoderm Folds of archenteron form coelom. Anus Mouth Digestive tube Key Ectoderm Mesoderm Endoderm Mouth Mouth develops from blastopore. Anus Anus develops from blastopore. Figure 32.9a (a) Cleavage Protostome development (examples: molluscs, annelids)) Eight-cell stage Deuterostome development (examples: echinoderms, chordates)) Eight-cell stage Key Ectoderm Mesoderm Endoderm Spiral and determinate Radial and indeterminate Coelom Formation • In protostome development development, the splitting of solid masses of mesoderm forms the coelom • In deuterostome development development, the mesoderm buds from the wall of the archenteron to form the coelom © 2011 Pearson Education, Inc. Figure 32.9b (b) Coelom formation Protostome development (examples: molluscs, annelids) Deuterostome development (examples: echinoderms, chordates) C Coelom Archenteron c e te o Coelom Key ey Ectoderm Mesoderm Endoderm Mesoderm Blastopore Solid masses of mesoderm p and form coelom. split Blastopore Mesoderm Folds of archenteron form coelom. Fate of the Blastopore • The blastopore forms during gastrulation and connects the archenteron to the exterior of the gastrula • In protostome development, the blastopore becomes the mouth • In deuterostome development, the blastopore b becomes th the anus © 2011 Pearson Education, Inc. Figure 32.9c (c) Fate of the blastopore Protostome development (examples: molluscs, annelids) Deuterostome development (examples: echinoderms, chordates) Anus Mouth Digestive tube Key Ectoderm Mesoderm Endoderm Mouth Mouth develops from blastopore. Anus Anus develops from blastopore. Concept : New views of animal phylogeny are emerging from molecular data • Zoologists recognize about three dozen animal phyla • Phylogenies now combine morphological, molecular, and fossil data • Current debate in animal systematics has led to the development p of multiple p hypotheses yp about the relationships among animal groups © 2011 Pearson Education, Inc. • One hypothesis of animal phylogeny is based mainly on morphological and developmental comparisons © 2011 Pearson Education, Inc. Figure 32.10 Porifera Cnidaria Eum metazoa M Metazoa ANCESTRAL COLONIAL FLAGELLATE Ctenophora Pro otostomiia Billateria Deutero ostomia a Ectoprocta Brachiopoda p Echinodermata Chordata Platyhelminthes R tif Rotifera Mollusca Annelida Arthropoda Nematoda • One hypothesis of animal phylogeny is based mainly on molecular data © 2011 Pearson Education, Inc. Figure 32.11 Ct Ctenophora h Eumeta azoa Me etazoa ANCESTRAL COLONIAL FLAGELLATE Porifera Cnidaria Acoela Bilateria Chordata Platyhelminthes L Lophotro ochozoa a Ecdysozoa Deute erostomiia Echinodermata Rotifera Ectoprocta Brachiopoda Mollusca Annelida Nematoda Arthropoda Points of Agreement 1. All animals share a common ancestor 2. Sponges are basal animals 3 Eumetazoa 3. E t is i a clade l d off animals i l (eumetazoans) with true tissues 4. Most animal phyla belong to the clade Bilateria, and are called bilaterians 5. Chordates and some other phyla belong to the clade Deuterostomia © 2011 Pearson Education, Inc. Progress in Resolving Bilaterian Relationships • The morphology-based tree divides bilaterians into two clades: deuterostomes and protostomes • In contrast, recent molecular studies indicate three bil t i clades: bilaterian l d D Deuterostomia, t t i E Ecdysozoa, d and d Lophotrochozoa • Ecdysozoans shed their exoskeletons through a process called ecdysis © 2011 Pearson Education, Inc. Vertebrate Development: p A Bodyy Building Plan • A human embryo y at about 7 weeks after conception shows development of distinctive features © 2011 Pearson Education, Inc. Figure 47.1 1 mm • Development occurs at many points in the life cycle of an animal • This includes metamorphosis and gamete production, as well as embryonic development © 2011 Pearson Education, Inc. Figure 47.2 EMBRYONIC DEVELOPMENT Sperm Zygote Adult frog Egg Metamorphosis Blastula Larval stages Gastrula Tail-bud embryo • Although animals display different body plans plans, they share many basic mechanisms of development and use a common set of regulatory genes • Biologists use model organisms to study development, chosen for the ease with which they can be studied in the laboratory © 2011 Pearson Education, Inc. Concept 47.1: Fertilization and cleavage initiate embryonic development • Fertilization is the formation of a diploid zygote from a haploid egg and sperm © 2011 Pearson Education, Inc. Fertilization • Molecules and events at the egg surface play a crucial role in each step of fertilization – Sperm penetrate the protective layer around the egg – Receptors on the egg surface bind to molecules on the sperm surface – Changes at the egg surface prevent polyspermy polyspermy, the entry of multiple sperm nuclei into the egg © 2011 Pearson Education, Inc. The Acrosomal Reaction • The acrosomal reaction is triggered when the sperm meets the egg • The acrosome at the tip of the sperm releases hydrolytic enzymes that digest material surrounding the egg © 2011 Pearson Education, Inc. Figure 47.3-1 Basal body (centriole) p Sperm head Acrosome Jelly coat Sperm-binding receptors Vitelline layer Egg plasma membrane Figure 47.3-2 Basal body (centriole) p Sperm head Acrosome Jelly coat Sperm-binding receptors Hydrolytic enzymes Vitelline layer Egg plasma membrane Figure 47.3-3 Sperm nucleus Basal body (centriole) p Sperm head Acrosome Jelly coat Sperm-binding receptors Acrosomal process Actin filament Hydrolytic enzymes Vitelline layer Egg plasma membrane Figure 47.3-4 Sperm plasma membrane Sperm nucleus Basal body (centriole) p Sperm head Acrosome Jelly coat Sperm-binding receptors Acrosomal process Actin filament Fused plasma membranes b Hydrolytic enzymes Vitelline layer Egg plasma membrane Figure 47.3-5 Sperm plasma membrane Sperm nucleus Basal body (centriole) p Sperm head Acrosome Jelly coat Sperm-binding receptors Fertilization envelope l Acrosomal process Actin filament Cortical Fused granule plasma membranes b Hydrolytic enzymes Perivitelline space Vitelline layer Egg plasma membrane EGG CYTOPLASM • Gamete contact and/or fusion depolarizes the egg cell membrane and sets up a fast block to polyspermy © 2011 Pearson Education, Inc. The Cortical Reaction • Fusion of egg and sperm also initiates the cortical reaction • Seconds after the sperm binds to the egg egg, vesicles just beneath the egg plasma membrane release their contents and form a fertilization envelope • The fertilization envelope acts as the slow block t polyspermy to l © 2011 Pearson Education, Inc. • The cortical reaction requires a high concentration of Ca2 ions in the egg • The reaction is triggered by a change in Ca2 concentration • Ca C 2 spread d across th the egg correlates l t with ith th the appearance of the fertilization envelope © 2011 Pearson Education, Inc. Figure 47.4 EXPERIMENT 10 sec after fertilization 25 sec 35 sec 1 min 10 sec after f tili ti fertilization 20 sec 30 sec 500 m RESULTS 1 sec before f tili ti fertilization CONCLUSION Point of sperm nucleus entry Spreading wave of Ca2 Fertilization envelope 500 m m Figure 47.4a EXPERIMENT 10 sec after fertilization 25 sec 35 sec 1 min 10 sec after fertilization 20 sec 30 sec 500 m RESULTS 1 sec before fertilization 500 m Figure 47.4b CONCLUSION Point P i t off sperm nucleus entry Spreading wave of Ca2 Fertilization envelope Egg Activation • The rise in Ca2+ in the cytosol increases the rates of cellular respiration and protein synthesis by the egg cell • With these rapid changes in metabolism, the egg is said to be activated • The proteins and mRNAs needed for activation are already l d presentt iin th the egg • The sperm nucleus merges with the egg nucleus and cell division begins © 2011 Pearson Education, Inc. Fertilization in Mammals • Fertilization in mammals and other terrestrial animals is internal • Secretions in the mammalian female reproductive tract alter sperm motility and structure • This Thi is i called ll d capacitation, it ti and d mustt occur b before f sperm are able to fertilize an egg © 2011 Pearson Education, Inc. • Sperm travel through an outer layer of cells to reach the zona pellucida, the extracellular matrix of the egg • When the sperm binds a receptor in the zona pellucida it triggers a slow block to polyspermy pellucida, • No fast block to polyspermy has been identified in mammals l © 2011 Pearson Education, Inc. Figure 47.5 Zona pellucida Follicle cell Sperm basal body Sperm nucleus Cortical granules • In mammals the first cell division occurs 1236 12 36 hours after sperm binding • The diploid nucleus forms after this first division of the zygote © 2011 Pearson Education, Inc. Cleavage • Fertilization is followed by cleavage, cleavage a period of rapid cell division without growth • Cleavage partitions the cytoplasm of one large cell into many smaller cells called blastomeres • The Th blastula bl t l is i ab ballll off cells ll with ith a flfluid-filled id fill d cavity called a blastocoel © 2011 Pearson Education, Inc. Figure 47.6 50 m (a) Fertilized egg (b) Four-cell stage (c) Early blastula ( ) Later blastula (d) Cleavage Patterns • In frogs and many other animals animals, the distribution of yolk (stored nutrients) is a key factor influencing the pattern of cleavage • The vegetal pole has more yolk; the animal pole has less yolk • The difference in yolk distribution results in animal and d vegetal t lh hemispheres i h th thatt diff differ iin appearance © 2011 Pearson Education, Inc. • The first two cleavage furrows in the frog form four equally sized blastomeres • The third cleavage is asymmetric asymmetric, forming unequally sized blastomeres © 2011 Pearson Education, Inc. • Holoblastic cleavage, cleavage complete division of the egg, occurs in species whose eggs have little or moderate amounts of yolk yolk, such as sea urchins and frogs • Meroblastic cleavage, cleavage incomplete division of the egg, occurs in species with yolk-rich eggs, such as reptiles and birds © 2011 Pearson Education, Inc. Figure 47.7 Zygote 2-cell 2 ll stage forming Gray crescent 0.25 mm 8-cell stage (viewed f from the h animal i l pole) l ) 4-cell stage forming 8 cell 8-cell stage Animal pole 0.25 mm Blastula (at least 128 cells) V Vegetal l pole l Blastula (cross section) Blastocoel Figure 47.7a-1 Zygote Figure 47.7a-2 Gray crescent Zygote 2-cell stage forming Figure 47.7a-3 Gray crescent Zygote 2-cell stage forming 4-cell stage forming Figure 47.7a-4 Animal pole Gray crescent Zygote 2-cell stage forming Vegetal pole 4-cell stage forming 8-cell stage Figure 47.7a-5 Animal pole Gray crescent Zygote 2-cell stage forming Blastocoel Vegetal pole 4-cell stage forming 8-cell stage Blastula (cross section) Figure 47.7b 0 25 mm 0.25 Animal pole 8-cell stage (viewed from the animal pole) Figure 47.7c 0 25 mm 0.25 Blastocoel Blastula (at least 128 cells) Regulation of Cleavage • Animal embryos complete cleavage when the ratio of material in the nucleus relative to the cytoplasm is sufficiently large © 2011 Pearson Education, Inc. Concept 47.2: Morphogenesis in animals involves specific changes in cell shape, position, ii and d survival i l • After f cleavage, the rate off cell division slows and the normal cell cycle is restored • Morphogenesis, the process by which cells occupy their appropriate locations, involves – Gastrulation, the movement of cells from the blastula surface to the interior of the embryo – Organogenesis, the formation of organs © 2011 Pearson Education, Inc. Gastrulation • Gastrulation rearranges the cells of a blastula into a three-layered embryo, called a gastrula © 2011 Pearson Education, Inc. • The three layers produced by gastrulation are called embryonic germ layers – The ectoderm forms the outer layer – The endoderm lines the digestive tract – The mesoderm partl partly fills the space bet between een the endoderm and ectoderm • E Each h germ llayer contributes t ib t tto specific ifi structures t t in the adult animal © 2011 Pearson Education, Inc. Figure 47.8 ECTODERM (outer layer of embryo) • Epidermis of skin and its derivatives (including sweat glands, hair follicles) • Nervous and sensory systems • Pituitary gland gland, adrenal medulla • Jaws and teeth • Germ cells MESODERM (middle layer of embryo) • Skeletal and muscular systems • Circulatory and lymphatic systems • Excretory and reproductive systems (except germ cells) • Dermis of skin • Adrenal cortex ENDODERM (inner layer of embryo) • Epithelial lining of digestive tract and associated organs (liver, pancreas) • Epithelial lining of respiratory, excretory, and reproductive tracts and d ducts d • Thymus, thyroid, and parathyroid glands Gastrulation in Sea Urchins • Gastrulation begins at the vegetal pole of the blastula • Mesenchyme cells migrate into the blastocoel • The vegetal plate forms from the remaining cells of th vegetal the t l pole l and db buckles kl iinward d th through h invagination © 2011 Pearson Education, Inc. • The newly formed cavity is called the archenteron • This opens through the blastopore, blastopore which will become the anus © 2011 Pearson Education, Inc. Figure 47.9 Animal pole Blastocoel Mesenchyme cells Vegetal plate Vegetal g pole Blastocoel Filopodia Mesenchyme cells Bl t Blastopore Archenteron 50 m Blastocoel Ectoderm Key Future ectoderm Future mesoderm Future endoderm Mouth Mesenchyme (mesoderm forms future skeleton) A h t Archenteron Blastopore Di Digestive i tube b (endoderm) ( d d ) Anus (from blastopore) Figure 47.9a-1 Blastocoel Animal pole Mesenchyme cells Vegetal Vegetal pole plate Key Future ectoderm Future mesoderm Future endoderm Figure 47.9a-2 Blastocoel Animal pole Mesenchyme cells Vegetal Vegetal pole plate Key Future ectoderm Future mesoderm Future endoderm Figure 47.9a-3 Blastocoel Animal pole Mesenchyme cells Filopodia Vegetal Vegetal pole plate Archenteron Key Future ectoderm Future mesoderm Future endoderm Figure 47.9a-4 Blastocoel Animal pole Mesenchyme cells Filopodia Vegetal Vegetal pole plate Archenteron Blastocoel Archenteron Key Future ectoderm Future mesoderm Future endoderm Blastopore Figure 47.9a-5 Blastocoel Animal pole Mesenchyme cells Filopodia Vegetal Vegetal pole plate Archenteron Blastocoel Archenteron Key Future ectoderm Future mesoderm Future endoderm Digestive tube (endoderm) Ectoderm Blastopore Mouth Mesenchyme (mesoderm forms future skeleton) Anus (from blastopore) Figure 47.9b Blastocoel Filopodia Mesenchyme cells Blastopore Archenteron 50 m Gastrulation in Frogs • Frog gastrulation begins when a group of cells on the dorsal side of the blastula begins to invaginate g • This forms a crease along the region where the gray g y crescent formed • The part above the crease is called the dorsal lip of the blastopore p © 2011 Pearson Education, Inc. • Cells continue to move from the embryo surface into the embryo by involution • These cells become the endoderm and mesoderm • Cells C ll on th the embryo b surface f will ill fform th the ectoderm © 2011 Pearson Education, Inc. Figure 47.10 1 CROSS SECTION SURFACE VIEW Animal pole Blastocoel Dorsal lip of blastopore Early Vegetal pole gastrula Blastopore Blastocoel shrinking 2 3 Blastocoel remnant Dorsal lip of blastopore Archenteron Ectoderm Mesoderm Endoderm Key Future ectoderm Future mesoderm Future endoderm Late gastrula Blastopore Blastopore Bl t Yolk plug Archenteron Figure 47.10a 1 CROSS SECTION SURFACE VIEW Animal pole Bl t Blastocoel l Key Future ectoderm Future mesoderm Future endoderm Dorsal lip p of blastopore Early E l Vegetal pole gastrula Blastopore Dorsal lip of blastopore Figure 47.10b 2 Key Future ectoderm Future mesoderm Future endoderm Blastocoel shrinking Archenteron Figure 47.10c 3 Key Future ectoderm Future mesoderm Future endoderm Late gastrula Blastopore Blastocoel remnant Ectoderm Mesoderm Endoderm Blastopore Yolk plug A h Archenteron Gastrulation in Chicks • Prior to gastrulation, gastrulation the embryo is composed of an upper and lower layer, the epiblast and hypoblast respectively hypoblast, • During gastrulation, epiblast cells move toward the midline of the blastoderm and then into the embryo toward the yolk © 2011 Pearson Education, Inc. • The midline thickens and is called the primitive streak • The hypoblast cells contribute to the sac that surrounds the yolk and a connection between the yolk and the embryo embryo, but do not contribute to the embryo itself © 2011 Pearson Education, Inc. Figure 47.11 Fertilized egg Primitive streak Embryo Yolk Primitive streak Epiblast p Future ectoderm Blastocoel Migrating cells (mesoderm) Endoderm Hypoblast YOLK Gastrulation in Humans • Human eggs have very little yolk • A blastocyst is the human equivalent of the blastula • The inner cell mass is a cluster of cells at one end d off the th blastocyst bl t t • The trophoblast is the outer epithelial layer of the blastocyst and does not contribute to the embryo, but instead initiates implantation © 2011 Pearson Education, Inc. • Following implantation, implantation the trophoblast continues to expand and a set of extraembryonic membranes is formed • These enclose specialized structures outside of the embryo • Gastrulation involves the inward movement from th epiblast, the ibl t th through h a primitive i iti streak, t k similar i il to the chick embryo © 2011 Pearson Education, Inc. Figure 47.12 1 Blastocyst reaches uterus. Uterus Endometrial epithelium (uterine lining) Inner cell mass Trophoblast Blastocoel 2 Blastocyst implants (7 days after fertilization). Expanding region of trophoblast Maternal blood vessel Epiblast Hypoblast Trophoblast 3 Extraembryonic membranes start to form (10–11 days), and gastrulation begins (13 days). Expanding region of trophoblast Amniotic cavity Epiblast Hypoblast Yolk sac (from hypoblast) Extraembryonic mesoderm cells (from epiblast) (f ibl t) Chorion (from trophoblast) 4 Gastrulation has produced a three-layered three layered embryo with four extraembryonic membranes. Amnion Chorion Ectoderm Mesoderm Endoderm Yolk sac Extraembryonic mesoderm Allantois Figure 47.12a Endometrial epithelium (uterine lining) Uterus Inner cell mass Trophoblast Blastocoel 1 Blastocyst reaches uterus. Figure 47.12b Expanding region of trophoblast Maternal blood vessel Epiblast Hypoblast yp Trophoblast 2 Blastocyst implants (7 days after fertilization). Figure 47.12c Expanding region of trophoblast Amniotic cavity Epiblast Hypoblast Yolk sac (from hypoblast) Extraembryonic mesoderm cells (from epiblast) Chorion Ch i (from (f trophoblast) t h bl t) 3 Extraembryonic membranes start to form (10–11 (10 11 days), and gastrulation begins (13 days). Figure 47.12d Amnion Chorion Ectoderm Mesoderm Endoderm Yolk sac Extraembryonic mesoderm Allantois 4 Gastrulation has produced a three-layered three layered embryo with four extraembryonic membranes. Developmental Adaptations of Amniotes • The colonization of land by vertebrates was made possible only after the evolution of – The shelled egg of birds and other reptiles as well as monotremes (egg-laying mammals) – The uterus of marsupial and eutherian mammals © 2011 Pearson Education, Inc. • In both adaptations adaptations, embryos are surrounded by fluid in a sac called the amnion • This Thi protects t t the th embryo b ffrom desiccation d i ti and d allows reproduction on dry land • Mammals and reptiles including birds are called amniotes for this reason © 2011 Pearson Education, Inc. • The four extraembryonic membranes that form around the embryo – – – – The chorion functions in gas exchange The amnion encloses the amniotic fluid Th yolk The lk sac encloses l th the yolk lk The allantois disposes of waste products and contributes t ib t tto gas exchange h © 2011 Pearson Education, Inc. Organogenesis • During organogenesis, organogenesis various regions of the germ layers develop into rudimentary organs • Early in vertebrate organogenesis organogenesis, the notochord forms from mesoderm, and the neural plate forms from ectoderm © 2011 Pearson Education, Inc. Figure 47.13 Eye Neural folds Neural fold Tail bud Neural plate SEM 1 mm Neural fold Somites Neural tube Neural plate Notochord Neural crest cells 1 mm Neural crest cells Coelom Notochord Somite E t d Ectoderm Mesoderm Endoderm Neural crest cells Outer layer of ectoderm Archenteron (a) Neural plate formation Neural tube (b) Neural tube formation Archenteron (digestive cavity) (c) Somites Figure 47.13a Neural folds 1 mm Neural Neural fold plate Notochord Ectoderm Mesoderm Endoderm Archenteron A h t (a) Neural plate formation • The neural plate soon curves inward inward, forming the neural tube • The neural tube will become the central nervous system (brain and spinal cord) © 2011 Pearson Education, Inc. Figure 47.13b-1 (b) Neural tube formation Neural fold Neural plate Figure 47.13b-2 Neural fold Neural plate Neural crest cells (b) Neural tube formation Figure 47.13b-3 Neural fold Neural plate Neural crest cells Neural crest cells (b) Neural tube formation Neural tube Outer layer y of ectoderm • Neural crest cells develop along the neural tube of vertebrates and form various parts of the embryo (nerves (nerves, parts of teeth teeth, skull bones bones, and so on) • Mesoderm lateral to the notochord forms blocks called somites • Lateral L t l tto the th somites, it th the mesoderm d splits lit tto fform the coelom (body cavity) © 2011 Pearson Education, Inc. Figure 47.13c Eye SEM Neural tube Notochord Coelom Somites Tail bud 1 mm Neurall N crest cells Somite (c) Somites Archenteron (di (digestive ti cavity) Figure 47.13d Neural folds 1 mm Figure 47.13e Eye SEM Somites T il bud Tail b d 1 mm • Organogenesis in the chick is quite similar to that in the frog © 2011 Pearson Education, Inc. Figure 47.14 Neural tube Notochord Eye Forebrain Somite Coelom Endoderm Mesoderm Ectoderm Archenteron Lateral fold Heart Blood vessels Somites Yolk stalk These layers form extraembryonic membranes. ((a)) Early y organogenesis g g Yolk sac Neural tube YOLK ((b)) Late organogenesis g g Figure 47.14a Neural tube Notochord Somite Archenteron Coelom Endoderm Mesoderm Ectoderm Lateral fold Yolk stalk These layers form extraembryonic membranes. (a) Early organogenesis Yolk sac YOLK Figure 47.14b Eye ye Forebrain Heart Blood vessels Somites Neural tube (b) Late organogenesis • The mechanisms of organogenesis in invertebrates are similar, but the body plan is very different • For example, the neural tube develops along the ventral side of the embryo in invertebrats invertebrats, rather than dorsally as occurs in vertebrates © 2011 Pearson Education, Inc. Mechanisms of Morphogenesis • Morphogenesis in animals but not plants involves movement of cells © 2011 Pearson Education, Inc. The Cytoskeleton in Morphogenesis • Reorganization of the cytoskeleton is a major force in changing cell shape during development • For example, example in neurulation, neurulation microtubules oriented from dorsal to ventral in a sheet of ectodermal cells help lengthen the cells along that axis © 2011 Pearson Education, Inc. Figure 47.15-1 Ectoderm Figure 47.15-2 Ectoderm Neural plate Microtubules Figure 47.15-3 Ectoderm Neural plate Microtubules Actin filaments Figure 47.15-4 Ectoderm Neural plate Microtubules Actin filaments Figure 47.15-5 Ectoderm Neural plate Microtubules Actin filaments Neural tube • The cytoskeleton promotes elongation of the archenteron in the sea urchin embryo • Thi This iis convergentt extension, t i th the rearrangement of cells of a tissue that cause it to become narrower (converge) and longer (extend) • Convergent extension occurs in other d developmental l t l processes • The cytoskeleton also directs cell migration © 2011 Pearson Education, Inc. Figure 47.16 Programmed Cell Death • Programmed cell death is also called apoptosis • At various times during development, individual cells sets of cells cells, cells, or whole tissues stop developing and are engulfed by neighboring cells • For example, example many more neurons are produced in developing embryos than will be needed • Extra neurons are removed by apoptosis © 2011 Pearson Education, Inc. • Later studies of C. elegans used the ablation (destruction) of single cells to determine the structures that normally arise from each cell • The researchers were able to determine the lineage of each of the 959 somatic cells in the worm © 2011 Pearson Education, Inc. Time affter fertilization (h hours) Figure 47.18 Zygote 0 First cell division Nervous system, outer skin,, musculature 10 Musculature, gonads Outer skin, nervous system Germ line (future gametes)) g Musculature Hatching g Intestine Intestine Anus Mouth Eggs Vulva POSTERIOR ANTERIOR 1.2 mm Figure 47.18a • Germ cells are the specialized cells that give rise to sperm or eggs • Complexes of RNA and protein are involved in the specification of germ cell fate • In I C. C elegans, l such h complexes l are called ll d P granules, persist throughout development, and can be b d detected t t d iin germ cells ll off th the adult d lt worm © 2011 Pearson Education, Inc. Figure 47.19 100 m • P granules are distributed throughout the newly fertilized egg and move to the posterior end before the first cleavage division • With each subsequent cleavage, the P granules are partitioned into the posterior-most cells • P granules act as cytoplasmic determinants, fixing germ cellll ffate t att the th earliest li t stage t off development d l t © 2011 Pearson Education, Inc. Figure 47.20 20 m 1 Newly fertilized egg 2 Zygote prior to first division 3 Two-cell embryo 4 Four-cell embryo Axis Formation • A body plan with bilateral symmetry is found across a range of animals • This body plan exhibits asymmetry across the dorsal-ventral and anterior-posterior axes • The Th right-left i ht l ft axis i iis llargely l symmetrical ti l © 2011 Pearson Education, Inc. • The anterior anterior-posterior posterior axis of the frog embryo is determined during oogenesis • The animal-vegetal animal vegetal asymmetry indicates where the anterior-posterior axis forms • The Th d dorsal-ventral l t l axis i iis nott d determined t i d until til fertilization © 2011 Pearson Education, Inc. • Upon fusion of the egg and sperm sperm, the egg surface rotates with respect to the inner cytoplasm • This cortical rotation brings molecules from one area of the inner cytoplasm of the animal hemisphere to interact with molecules in the vegetal cortex • This Thi lleads d tto expression i off d dorsall and d ventralt l specific gene expression © 2011 Pearson Education, Inc. Figure 47.21 Dorsal Right Anterior Posterior Left Ventral (a) The three axes of the fully developed embryo Animal pole Animal hemisphere Vegetal hemisphere Vegetal pole (b) Establishing the axes Point of sperm nucleus entry Gray crescent Pigmented cortex Future dorsal side First cleavage • In chicks, chicks gravity is involved in establishing the anterior-posterior axis • Later, Later pH differences between the two sides of the blastoderm establish the dorsal-ventral axis • In I mammals, l experiments i t suggestt that th t orientation i t ti of the egg and sperm nuclei before fusion may h l establish help t bli h embryonic b i axes © 2011 Pearson Education, Inc.