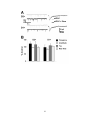
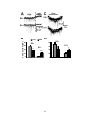
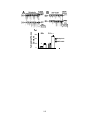
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
REGULATION OF STRIATAL MEDIUM SPINY NEURON GABAA RECEPTOR MEDIATED TONIC CURRENT A Dissertation Submitted to the Faculty of the Graduate School of Arts and Sciences of Georgetown University in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Pharmacology By Megan J Janssen Schroeder, B.S. Washington, DC July 2, 2010 Copyright 2010 by M. Janssen Schroeder All Rights Reserved ii REGULATION OF STRIATAL MEDIUM SPINY NEURON GABAA RECEPTOR MEDIATED TONIC CURRENT Megan J Janssen Schroeder, B.S. Thesis Advisor: Stefano Vicini, Ph.D. ABSTRACT The majority of striatal neurons are GABAergic projecting medium spiny neurons (MSNs), of which there are two types based on their primary axonal projection pathway and dopamine receptor expression. MSNs that express the dopamine D2 receptor (D2+) inhibit the globus pallidus to inhibit movement, while those that express the dopamine D1 receptor (D1+) inhibit the substantia nigra to facilitate movement. Although these neurons are morphologically and electrophysiologically indistinguishable, D2+ MSNs express greater GABAergic inhibitory tonic currents than D1+ MSNs in young mice. In this thesis, I identify putative GABAA receptor subunits that underlie tonic current in striatal MSNs. Using whole-cell recordings in acute mouse brain slices from bacterial artificial chromosome transgenic mice where EGFP is driven by the Drd2 promoter and tdTomato protein is driven by the Drd1a promoter, I show that the GABAA receptor β3 subunit largely regulates tonic current based on phosphorylation state and dopamine receptor activation. Thus, D2+ MSNs express tonic current through a basally phosphorylated β3 subunit, while under proper conditions where the β3 subunit becomes phosphorylated, D1+ MSNs also express tonic current. Indeed, a transgenic mouse with β3 subunits selectively knocked-out of D2+ MSNs failed to demonstrate the typical tonic iii current pattern seen in wild type mice, suggesting that this subunit is particularly important in mediating tonic current. In this thesis, I also compare striatal MSNs to a subtype of striatal GABAergic interneurons that express Neuropeptide Y (NPY +). These neurons have a characteristic cell-attached firing pattern, making them a likely source for the ambient GABA necessary for MSN tonic currents. My data suggest that these interneurons receive fewer inhibitory inputs with different presynaptic origins than MSNs. My thesis work has revealed important players in striatal tonic current and such insight offers innumerable pharmacological targets in the treatment of Parkinson‟s disease, and other striatally relevant diseases, where an imbalance in the D2+ and D1+ MSN projection pathways causes symptomatic akinetic behaviors. iv ACKNOWLEDGMENTS This thesis dissertation has come to fruition due to the support of many individuals. Each person has either helped to shape my research or my personal skills, without which this project would not have been completed. My thesis mentor, Dr. Stefano Vicini, has been extremely instrumental in my development as a scientist. I am grateful for his excitement when experiments were successful and his words of encouragement when experiments did not go quite like we had hoped. I am equally appreciative for the high standards that he expects his students to meet; I was challenged each and every day, and these challenges have made my successes even more significant. I am honored to continue working for him as a postdoctoral fellow, as I continue to progress towards becoming an independent scientist. Dr. John Partridge has also been a tremendous help to me during my time in the lab. Not only has he answered my many questions, but has also offered the guidance and constructive criticism to perfect my understanding of, and love for, striatal physiology. Dr. Kristen Ade introduced me to the wonderful world of GABAergic currents in striatal MSNs. Although the research has changed and evolved over the years, the project was originally hers, and she handed it off to me when she completed her PhD, for which I am grateful. I am greatly appreciative to the members of my thesis committee. Drs. David Lovinger, Gerard Ahern, Molly Huntsman, and Barry Wolfe have spent numerous hours with me, going over data, discussing possible interpretations, and offering constructive criticisms to my research. v I would also like to thank other members, both past and present, of Dr. Vicini‟s lab: Dr. Zhanyan Fu, Mary Adedoyin, Alfredo Gonzalez-Sulser, and Rupa Lalchandani. Their friendship has meant so much to me over the years, and made my time in this lab truly enjoyable and memorable. My family has been instrumental in my successes, both past and present. My parents emphasized the importance of education and pursing dreams. Their encouragement, support, and praise has made me want to work harder and achieve more each and every day. Most importantly, I dedicate this dissertation to my husband, Pete. His unvarying patience, support, love, and encouragement have gotten me through the toughest times of these past four years, and made the pleasant times even more exciting and meaningful. vi TABLE OF CONTENTS PAGE ABSTRACT iii ACKNOWLEDGEMENTS v TABLE OF CONTENTS vii LIST OF FIGURES xii LIST OF TABLES xiv INTRODUCTION 1 I. GABA Receptor Mediated Inhibition 1 a. Phasic and Tonic Inhibitory Currents 2 b. GABAA Receptor Subunits 5 II. The Basal Ganglia 7 III. Striatal Medium Spiny Neurons 11 a. MSN Electrophysiological Characteristics 14 b. GABAA Receptor Subunit Expression in Striatal MSNs 17 Striatal Interneuons 18 a. The Cholinergic Interneuron 18 b. The GABAergic Calretinin-Positive Interneuron 19 c. The GABAergic Fast-Spiking Interneuron 20 d. The GABAergic Low-Threshold Spiking Interneuron 24 V. Feedforward and Feedback Inhibition 29 VI. Modulation of Striatal MSNs 31 a. Dopamine 31 IV. vii VII. b. Adenosine 34 Post-translational Modifications of MSN GABAergic Currents 36 a. Receptor Assembly and Membrane Insertion 36 b. Phosphorylation 38 i. Dopamine‟s Influence on Phosphorylation c. Endocytosis and Recycling 41 45 HYPOTHESIS 50 MATERIALS AND METHODS 51 I. Animals 51 II. Cre-lox System 54 III. Genotyping 55 IV. Acute Slice Preparation 59 V. Cell Identification 59 VI. Whole Cell Recordings 60 VII. Drugs and Peptides 62 VIII. Measurement of GABA Currents 63 a. Phasic GABAergic Measurements 63 b. GABAergic Tonic Current Measurements 64 c. Evoked Current Measurements 65 IX. HEK 293 Cell Culture and Transfection 65 X. Immunofluorescence and Confocal Imaging 67 XI. Statistics 68 RESULTS 69 viii I. GABAergic Tonic Conductances in Striatal MSNs 69 a. Identification of MSNs in Acute Slice Preparation 69 b. Unique GABAA Receptor Subunits Mediate MSN Tonic Current 72 i. Importance of β Subunit Expression in MSNs 80 ii. Etomidate is Selective for β3 Subunits in Recombinant Receptors 81 iii. MSNs have Different Etomidate Responses 87 iv. Etomidate does not Affect mIPSC Properties 87 c. PKA Phosphorylation Affects MSN Tonic Conductance 92 i. Intracellular PKA Application Modulates D1+ MSN BMR-Sensitive Tonic Current ii. Basal Phosphorylation of β3 Subunits in D2+ MSNs 92 96 iii. sIPSC Properties are Largely Unaffected by PKA and PKI 101 d. Dopamine Modulation of MSN Tonic Current 102 i. Little Basal Dopamine Present in Acute Slice Preparation 102 ii. Dopamine Receptor Agonists Alter MSN Tonic Current 103 iii. The Link Between Dopamine Receptor Activation, Phosphorylation, and GABA-Mediated Tonic Current 106 iv. Tonic Current in Adult MSNs 106 v. Dopamine Regulates MSN Excitability 112 vi. Dopamine Receptor Agonists do not Affect sIPSC Properties 112 ix e. The Importance of the GABAA Receptor β3 Subunit in MSN Tonic Tonic Current 115 i. Cre-lox Method Decreases β3 GABAA Receptor Subunit Expression 115 ii. Conditional β3 Subunit KO Mice Show Enhanced Lethality 116 iii. Conditional β3 Subunit KO Mice have Decreased Tonic Current 117 iv. Etomidate Sensitivity in Conditional β3 Subunit KO Mice 121 v. Synaptic Responses in β3 Subunit KO Mice II. Characterizing NPY+ Interneurons 122 125 a. Identification of LTS/NPY+ Interneurons in Acute Slice Preparation 125 b. Intrinsic NPY+ Interneuron Characterization 125 c. GABAergic Signaling in NPY+ Interneurons 129 d. Inhibitory Short Term Plasticity 135 DISCUSSION 144 I. Summary 144 II. Etomidate Sensitivity 145 III. PKA Phosphorylation of GABA Receptor Subunits 147 IV. Dopamine Receptor Regulation of Tonic Current 149 V. Potential Role for GABAA Receptor Endocytosis 154 x VI. Interpretations of Conditional β3 Subunit KO 156 VII. Potential GABAA Receptor Composition in Striatal MSNs 158 VIII. Developmental Regulation of MSN Tonic Current 160 IX. Functional Implications of Striatal MSN GABAergic Tonic Current 161 X. Inputs to NPY+ Interneurons 162 XI. GABAergic Drugs in the Management and Treatment of Movement XII. Disorders 164 Implications to Parkinson‟s Disease Research 165 CONCLUSIONS 167 APPENDIX 169 Copyright Permissions 169 REFERENCES 171 xi LIST OF FIGURES PAGE Figure 1. Phasic and Tonic GABAA Receptors Mediate Inhibition 4 Figure 2. Striatal Circuitry 10 Figure 3. Medium Spiny Neuron Characteristics 13 Figure 4. Fast Spiking Interneuron Characteristics 22 Figure 5. Characteristics of Low Threshold Spiking Interneurons 27 Figure 6. PKA Phosphorylation Cascade is Regulated by Dopamine 43 Figure 7. β3 Subunit-Mediated Endocytosis 47 Figure 8. Generation of Conditional β3 Subunit KO Mice using the Cre-Lox System 53 Figure 9. Striatopallidal and Striatonigral Pathways are Distinct 71 Figure 10. Tonic Current Expression in BAC Drd2 EGFP;BAC Drd1a tdTomato Mice 74 Figure 11. δ Subunits do not Mediate MSN Tonic Current 76 Figure 12. β3 Subunit Expression in Striatal MSNs 79 Figure 13. GABA Dose Response Curves for Recombinant GABAA Receptors 83 Figure 14. Etomidate‟s Direct Effects are Selective for β3 Subunit-Containing Recombinant Receptors 86 Figure 15. Etomidate Selectively Activates Extrasynpatic Receptors in D2+ MSNs 89 Figure 16. Etomidate does not Affect Striatal Synaptic GABAA Receptors 91 Figure 17. Internal PKA Application Regulates GABA-Mediated Tonic Current 94 Figure 18. D2+ MSNs are Basally Phosphorylated by PKA and PKC 98 xii Figure 19. Synaptic Receptors are Unaffected by PKA and PKI 100 Figure 20. Dopamine Agonists Alter MSN Tonic Conductance 105 Figure 21. SKF-81297 Increases Etomidate Response in D1+ MSNs 108 Figure 22. Adult Animals have Different Tonic Current Expression 111 Figure 23. Dopamine Receptor Agonists Modulate MSN Excitability 114 Figure 24. Tonic Current Patterns in β3 Subunit KO Mice 119 Figure 25. Synaptic Responses in Drd2-β3 Subunit KO Mice 124 Figure 26. Intrinsic NPY+ Interneuron Characteristics 127 Figure 27. GABA Mediated Synaptic Transmission in NPY+ Interneurons 131 Figure 28. Evoked GABAergic Currents in NPY+ Interneurons and MSNs 134 Figure 29. Short Term Plasticity in NPY+ Interneurons 138 Figure 30. Alterations to Presynaptic Release Probabilities 142 Figure 31. Tonic Conductance is Mediated through a Phosphorylated β3 Subunit 153 xiii LIST OF TABLES PAGE Table 1. PCR Reaction Protocols 56 xiv INTRODUCTION I. GABA Receptor Mediated Inhibition GABA (γ-aminobutyric acid) is the primary inhibitory neurotransmitter in the adult central nervous system (CNS), and is largely implicated in many developmental stages, as well as disease states and disorders, ranging from epilepsy to sleep disturbances. GABA binds to and activates both fast-acting ionotropic GABAA and GABAC receptors and slower metabotropic G-protein coupled GABAB receptors. GABAA receptors are pentameric ligand-gated ion channels that undergo a conformational change upon binding two agonist molecules to allow Cl- and, to a lesser extent, HCO3- ion flow (Bormann et al, 1987; Kaila, 1994). The inward or outward flow of these ions is based upon the electrochemical gradient. In mature neurons, the postsynaptic membrane potential typically becomes hyperpolarized upon receptor activation due to the inward ionic flow, resulting in reduced neuronal activity. GABAA receptors are responsible for the majority of GABAergic inhibition in the adult CNS, and are the GABA receptors investigated in this thesis. GABAC receptors are ionotropic, chloride-sensitive channels that are related to the GABAA receptor, but are insensitive to the GABAA receptor antagonist, bicuculline (Bormann, 2000). GABAB receptors are chloride-independent receptors that mediate inhibition by heterotrimeric G protein activation. These metabotropic GABAB channels also regulate cell excitability and are located both pre- and post-synaptically. Presynaptic GABAB receptors serve as autoreceptors and mediate a reduction in calcium 1 currents at the nerve terminal, while post-synaptic GABAB channels activate potassium channels that hyperpolarize the neuron (Bormann, 1988; Owens & Kriegstein, 2002). a. Phasic and Tonic Inhibitory Currents GABAA receptors produce two forms of inhibition based on certain biophysical features and cell surface location (Figure 1). Receptors in the post-synaptic active zone experience a rapid increase in GABA concentration due to pre-synaptic vesicular release, and GABA binding causes ion channel openings. This phasic mode of inhibition is defined by the short duration (< 1 ms) in which the receptors are exposed to high GABA concentrations (0.3 - 1 mM) due to rapid reuptake and diffusion away from the synapse (Nusser et al, 2001; Mozrzymas et al, 2003). Activation of phasic receptors induces inhibitory postsynaptic currents (IPSCs), characterized by high amplitudes and fast decay kinetics. Phasic inhibition is essential in setting the temporal window for synaptic integration (Farrant & Nusser, 2005), and may also generate rhythmic network activities (Cobb et al, 1995; Huntsman et al, 1999). GABAA receptors are also located outside the active zone, and play a role in the slower form of inhibition, tonic inhibition. Continuous, or tonic, activation of these receptors likely occurs from ambient neurotransmitter (near µM levels) (Tossman et al, 1986) present in the extracellular space that binds to the high affinity extrasynaptic and perisynaptic GABAA receptors. Although tonic current is small in amplitude, the persistent inhibition is likely central for modulating cellular excitability (Mody, 2001). Tonic currents are revealed by their blockade and application of a GABA antagonist that reduces the amount of current required to keep the membrane potential fixed (holding 2 Figure 1. Phasic and Tonic GABAA Receptors Mediate Inhibition. (A) Single vesicular release from a presynaptic terminal activates GABAA receptors in the postsynaptic active zone (yellow). The blue shading indicates the spread of released GABA. The current trace (below) shows a representative example of a mIPSC, with the charge transfer colored in green. (B) Action-potential mediated release of multiple presynaptic vesicles activates both synaptic (yellow) and extrasynaptic (blue) receptors due to the large spread of GABA, or “spillover.” The current trace (below) shows a representative example of a sIPSC with the light green showing the resultant charge transfer with the mIPSC charge transfer superimposed (green). (C) Low concentrations of ambient GABA fail to activate synaptic receptors, but activate the high-affinity extrasynaptic receptors (blue) to mediate tonic inhibition. The current trace (below) shows that application of the GABA receptor antagonist gabazine reduces the holding current and RMS noise, revealing tonic current. The green shaded area represents the charge transfer via these extrasynaptic receptors. Reprinted with permission from Farrant & Nusser, 2005. See Appendix for permission rights. 3 4 current). This decrease in input conductance is also accompanied by a reduction in current variance due to the reduced number of open GABAA channels (Brickley et al, 1996; Wall & Usowicz, 1997; Glykys & Mody, 2007). By increasing a cell‟s input conductance, tonic inhibition decreases the size and duration of excitatory postsynaptic potentials (EPSPs) while narrowing the temporal and spatial window in which synaptic integration can occur (Semyanov et al, 2004; Farrant & Nusser, 2005). b. GABAA Receptor Subunits There are 19 known subunits that comprise the GABAA channel: α1-6, β1-3, γ1-3, δ, ε, θ, π, and ρ1-3 (Macdonald & Olsen, 1994; Owens & Kriegstein, 2002; Seighart & Sperk, 2002). Each GABAA receptor typically expresses two α subunits, two β subunits, and either a δ or γ subunit to form the pentameric channel (McKernan & Whiting, 1996; Tretter et al, 1997). The subunit composition of a GABAA receptor determines the channel‟s subcellular location and biophysical properties. Therefore, the extent and type of inhibition exhibited by a neuron relies heavily on subunit composition. Although no GABAA subunit appears to have an exclusive synaptic location, α1, α2, α3, β2, β3, and γ2 subunits are enriched at GABAergic synapses in many brain regions (Craig et al, 1994; Somogyi et al, 1996; Brünig et al, 2002). Indeed, it has been shown that the γ2 subunit clusters receptors at the synaptic localization (Essrich et al, 1998; Nusser et al, 1998), although α5βXγ2 receptors are usually located extrasynaptically (Brünig et al, 2002) and mediate tonic current (Caraiscos et al, 2004; Prenosil et al, 2006; Ade et al, 2008). Receptors that incorporate the δ subunit appear to be exclusively present in the extrasynaptic plasma membrane in cerebellar granule cells 5 (Nusser et al, 1998) and hippocampal dentate gyrus cells (Nusser & Mody, 2002; Wei et al, 2003). δ subunits prefer to form receptors with α6, α4, and β2/β3 subunits (Jones et al, 1997; Barnard et al, 1998) to mediate tonic inhibition in many brain regions (Nusser & Mody, 2002; Stell et al, 2003; Cope et al, 2005; Jia et al, 2005; Mangan et al, 2005; Drasbek & Jensen, 2006; Mtchedlishvili & Kapur, 2006; Glykys et al, 2007). Although post-synaptic localization of the β subunit has not been well characterized, immunostaining in cultured hippocampal pyramidal neurons suggests that β1 GABAA receptor subunits are not synaptically located, while β2/β3 subunits are present at both synaptic and extrasynaptic locations (Mangan et al, 2005). An additional study found that α4β2δ GABAA receptors mediate tonic current in dentate gyrus granule cells (Herd et al, 2008). As β subunit expression may play a unique role in mediating both types of inhibitory currents because of its phosphorylation capability and potential changes in biophysical properties (McDonald et al, 1998; see Introduction, Section VIIb), its expression and function is of central importance in this thesis. GABAA receptor subunits also dictate GABA sensitivity and other biophysical properties of the receptor. These properties prime the receptors with characteristics of phasic or tonic currents. In recombinant GABAA receptors, α subunits dictate much of the GABA sensitivity; α6 subunits have the highest affinity for GABA (Ducic et al, 1995), a property necessary for receptors in the extrasynaptic space due to low ambient GABA levels. Additionally, the incorporation of the δ subunit increases GABA affinity in receptors heterologously expressed in fibroblast cells (Saxena & Macdonald, 1994; Fisher and Macdonald, 1997). Receptor deactivation and desensitization rates are also dependent upon subunit composition. α1 subunit-containing receptors have the fastest 6 deactivation rate (Bianchi et al, 2002), whereas γ2 subunit-containing receptor have more rapid kinetics than δ subunit-containing receptors (Haas and Macdonald, 1999), ideal for their role in synaptic inhibition. Compared with γ2-containing receptors, αβδ receptors have slower and less extensive desensitization (Haas & Macdonald, 1999; Bianchi et al, 2001; Bianchi et al, 2002), priming them to generate inhibitory tonic current. II. The Basal Ganglia. The basal ganglia are a group of forebrain structures, associated with motor control and learning, that include the striatum, globus pallidus, substantia nigra, and subthalamic nucleus. In a complex array of organization, these regions integrate excitatory input from the cortex and generate a net inhibitory output to the thalamus that largely dictates motor control and movement (Kreitzer, 2009). Excitatory input from the cortex and thalamus converge in the largest basal ganglia structure, the striatum. These connections are topographically complex, such that an area of the cortex forms more connections in a particular region of the striatum compared to others (Voorn, 2004). This complex patterning of corticostriatal connections conditions areas of the striatum for procedural learning. The dorsomedial striatum receives much of its input from the association cortex, while the dorsolateral striatum receives most input from the sensorimotor cortex (Brown et al, 1998). The ventral striatum, also known as the nucleus accumbens (NAc), receives excitatory input from both the frontal cortex and limbic regions (Voorn, 2004), but can be further divided into shell and core regions, which also differ in their input. Because the ventral striatum is not the subject of this thesis, further discussion of the striatum will be referring to the 7 dorsal striatum, although concepts investigated in the dorsal striatum may also be relevant for ventral striatal pathways. Another important input structure for the striatal function, the substantia nigra pars compacta (SNpc), provides the striatum with dopamine. Due to the large dopaminergic input, the striatum contains the highest density of dopamine receptors in the rodent brain (Richtand et al, 1995). Although dopamine does not fit the classic definition of a neurotransmitter, it modulates striatal output from both the “direct” and “indirect” pathways (Gerfen et al, 1990). Histochemically, the striatum is often divided into two distinct compartments, the patch and matrix. Each region is distinct in neuronal connectivity and biochemical characteristics. The patch, also called striosome, consists of cells surrounded by the larger matrix of grey matter, and while it receives input from the superficial layers of layer V of the cortex, deeper areas of layer V and layer VI constitute the primary input to the matrix (Gerfen, 1992). Patches express high levels of opiate receptors, dopamine receptor 1 (D1R), Substance P, and tyrosine-hydroxylase reactive fibers (Graybiel & Ragsdale, 1978; Gerfen, 1992). Acetylcholinesterase-, calbindin-, and somatostatinpositive neurons largely populate the matrix (Gerfen, 1992). Striatal output is GABAergic and acts to inhibit neurons in other basal ganglia structures through one of two pathways. Striatal pathways that project to the internal segment of the globus pallidus (GPi) and the substantia nigra pars reticulata (SNr) comprise the “direct” pathway, which serves to disinhibit thalamocortical projections and facilitate movement (Kreitzer & Malenka, 2008). The striatal “indirect” pathway sends 8 Figure 2. Striatal Circuitry. Cartoon depicting the striatal circuitry within the basal ganglia. The striatum receives excitatory input from the cerebral cortex and thalamus (green arrows). Striatal neurons are comprised of dopamine D1 receptor (D1+) and Substance P (SubP) expressing neurons that constitute a direct pathway (blue arrows) that inhibits the substantia nigra pars reticulata (SNr) and medial globus pallidus (GPm). Both of these areas are output nuclei of the basal ganglia. The indirect pathway of striatal neurons (red arrows) consists of dopamine D2 receptor (D2+) and Enkephalin (Enk) expressing neurons, whose axons terminate in the globus pallidus (GP). GP neurons project to the subthalamic nucleus (STN), which then projects to the output structures of the GPm and SNr. Modified and reprinted with permission from Gerfen, 2006. See appendix for permission rights. 9 10 inhibitory projections to the external segment of the globus pallidus (GPe) and subthalamic nucleus (STN). The STN is not an output structure, and projects within the basal ganglia to the GPi and SNpr. Therefore, this pathway inhibits thalamocortical projections to inhibit movement (Kreitzer & Malenka, 2008). The balance between these two output pathways ensures proper movement control. Figure 2 attempts to simplify the complex basal ganglia organization and circuitry. III. Striatal Medium Spiny Neurons. Striatal neurons represent a heterogeneous population of neurons that either release acetylcholine (from Cholinergic interneurons, see Introduction, Section IVa) or GABA (from GABAergic interneuron and medium spiny neurons). Medium spiny neurons (MSNs) represent 90-95% of the striatal population (Graveland & DiFiglia, 1985) and nearly equal amounts of MSNs belong to the “direct” and “indirect” pathways (Gerfen, 2004). MSNs of the “direct” pathway primarily express the dopamine D1 receptor (D1+) and substance P (SubP), while MSNs of the “indirect” pathway primarily express the dopamine D2 receptor (D2+) and enkephalin (Enk) (Figure 2). In the larger basal ganglia circuit, D1+ MSN output acts to facilitate movement, while D2+ MSN output inhibits movement control. Although the two subtypes of MSNs have distinct output pathways and deliver opposing messages on movement initiation and control, these neurons are indistinguishable in size and basic physiology. Anatomically, all MSNs are medium sized (soma size ranges between 10-20 µm), and their dense dendritic tree is covered with small bulbular protrusions, called spines (Figure 3A). GABA and dopamine terminals 11 Figure 3. Medium Spiny Neuron Characteristics. (A) Confocal z-stack image of biocytin injected MSN. Calibration bar: 25 µm. Inset is 1.5 times magnification of the boxed red region, showing densely spiny dendrites. (B) Representative trace from a current clamp recording of a striatal MSN with a set holding potential of -70 mV. Current injection steps are 10 pA. Hyperpolarized current injections show significant inward rectification (i). Depolarizing current injections to threshold are characterized by delayed action potential firing (ii). Further depolarizing current injections show repetitive action potential firing (iii). Scale bars: 20 pA and 500 ms. 12 A B iii ii i 13 are both found on the spine neck, in a unique position to filter and regulate excitatory input, which is found on spine heads (Smith et al, 1994). MSN dendritic arborization is spherical, offering dense local innervation to other MSNs. The distal dendritic arborization of MSNs receives dense glutamatergic, excitatory innervation from the cortex and intralaminar thalamus, while modulatory dopaminergic afferents arise from the SNpc. Glutamatergic synapses primarily act through excitatory AMPA and NMDA receptors, while dopamine acts through metabotropic G-protein coupled receptors (GPCRs). In addition to sending inhibitory projections out of the striatum, some MSNs also form axon collaterals with other MSNs at distal portions of the dendritic tree (Tunstall et al, 2002; Czubayko & Plenz, 2002; Taverna et al, 2004; Tepper et al, 2004), forming a feedback inhibitory network (see Introduction, Section V). These connections allow neurons of both pathways to influence and be influenced by neurons of the same and opposite output pathway, adding to their complex circuitry. MSNs also receive inhibitory input from neighboring GABAergic interneurons that only make connections within the striatum, constituting an inhibitory feedforward circuitry (see Introduction, Section V). The inhibitory connections between presynaptic MSNs and interneurons and among interneurons remain subject for debate (Partridge et al, 2009; Gittis et al, 2010; Planert et al, 2010), although this issue will be revisited. a. MSN Electrophysiological Characteristics Morphologically indistinguishable, striatal D1+ and D2+ MSNs are characterized by several defining properties including hyperpolarized resting membrane potential 14 (RMP), low input resistance, delayed onset firing patterns, as well as both “up” and “down” states. Unique potassium channels shape many of these characteristics. At rest, a striatal MSN‟s membrane potential resides around -80 mV (Yasumoto et al, 2002; Shen et al, 2005; Ade et al, 2008). Because the MSN RMP resides far from the reversal potential for Cl- (-60 mV) (Mercuri et al, 1991), much of the GABAergic input results in membrane depolarization (Kita, 1996; Tunstall et al, 2002; Ade et al, 2008). This hyperpolarized membrane potential is principally determined by an inwardly rectifying potassium channel (Kir) (Bargas et al, 1988) where the current magnitude depends on the voltage difference between the RMP and reversal potential for potassium (Jiang & North, 1991, Nisenbaum & Wilson, 1995). At membrane potentials more negative than the chloride reversal potential, these channels are open, priming the MSN with unique inward rectification evoked by hyperpolarized current and low input resistance (Nisenbaum & Wilson, 1995) (Figure 3Bi). Therefore, Kir channels are thought to shunt the amplitude of excitatory synaptic input and reduce the efficacy for temporal summation of excitatory currents (Nisenbaum & Wilson, 1995). As the MSN membrane potential becomes more depolarized due to a barrage of excitatory input, Kir channels close and polyamines block the pore (Ficker et al., 1994). At potentials near -60 mV, transient-outwardly rectifying potassium channels (IAs) open, and function to slow the depolarization (Bargas et al, 1988; Nisenbaum & Wilson, 1995; Nisenbaum et al, 1996; Shen et al, 2004). Such Kv1.2 channels (Shen et al, 2004) have been shown to delay first spike latency in response to depolarized currents and slow the repetitive spike discharge of these cells (Nisenbaum et al, 1994), which are two additional MSN characteristics (Figure 3Bii, iii). 15 Lastly, while membrane “up” and “down” states are also regulated by these potassium channels, they are under direct influence of cortical and thalamic excitatory input. At rest, the mature MSN displays a hyperpolarized membrane potential (-90 to -80 mV) due to the Kir channels and the lack of synchronous excitatory input. Upon a barrage of synchronous excitatory input in vivo, Kir channels close, and the membrane potential reaches its “up” state, near -55 mV, due to outwardly rectifying potassium channels. In the “up” state, MSNs are more likely to fire action potentials, and modest excitatory input has greater impact on MSN firing. Therefore, closure of Kir channels increases MSN excitability as it allows for easier transitions into the more excitable “up” state. The advent of bacterial artificial chromosome (BAC) technology combined with fluorescent protein labeling has allowed identification of D1+ and D2+ MSNs in acute striatal slices (Gong et al, 2003; Shuen et al 2008), and has exponentially increased both investigation into and reliability of MSN subtype studies. These studies showed that in addition to their different projection patterns, striatonigral and striatopallidal neurons have different electrophysiological characteristics; D2+ MSNs are significantly more excitable than D1+ MSNs (Ade et al, 2008; Day et al, 2008; Gertler et al, 2008). In addition, D1+ and D2+ MSNs express different amounts of GABAergic tonic current. While young D1+ MSNs express small amounts of tonic current, young D2+ MSNs express a significant amount of tonic current (Ade et al, 2008; Santhakumar et al, 2010); this difference in tonic current is likely due to differences in GABAA receptor subunit expression. 16 b. GABAA Receptor Subunit Expression in Striatal MSNs. The GABAA receptor subunits expressed in the striatum have been thoroughly researched for many years with the aid of antibody staining (Liste et al, 1997; Waldvogel et al, 1997; Pirker et al, 2000; Rodriguez-Pallares et al, 2000; Schwarzer et al, 2001; Ade et al, 2008), single cell PCR (Flores-Hernandez et al, 2000), and pharmacological sensitivity (Ade et al, 2008; Santhakumar et al, 2010). Although all studies do not agree on subunit expression, the striatum stained positive for α1-α5 subunits, all β subunits, as well as γ2 and δ subunits in a comprehensive GABAA receptor antibody staining study performed by Pirker et al. (2000). While this study did not address which types of striatal neurons expressed which subunits, it has been shown that α3 and α6 subunits are not expressed in MSNs (Liste et al, 1997; Rodriguez-Pallares et al, 2000; Schwarzer et al, 2001), while they do express α2 and β2/β3 subunits (Liste et al, 1997). Although the β subunit antibody used in that study does not discriminate between β2 and β3 subunits, single-cell PCR studies suggest that β1 and β3 subunits are expressed by MSNs (FloresHernandez et al, 2000), while β2 subunits appear to be solely expressed on striatal interneurons (Schwarzer et al, 2001). The GABAA receptor subunits that mediate tonic current in striatal MSNs have been an interest in our lab for many years. Ade et al. (2008) showed that receptors containing α5 and γ2 subunits contribute to tonic current in young mice (p 16-21), while both α1 and δ subunits do not contribute to MSN tonic current. The β subunit responsible for MSN tonic current is a central component to this thesis, and I demonstrate that the β3 subunit largely mediates MSN GABAA tonic current in young animals (Janssen et al, 2009). 17 IV. Striatal Interneurons As indicated by their name, striatal interneurons‟ axons are restricted to the striatum and form connections with MSNs to modulate network activity and regulate inhibitory output. Although they only represent a small portion (about 5%) of striatal neurons, aspiny interneurons can be categorized into cholinergic and GABAergic cells (Kawaguchi et al, 1995). a. The Cholinergic Interneuron Cholinergic interneurons are giant (20-50 µm diameter soma) aspiny interneurons, representing approximately 2% of all striatal neurons. They express choline acetyltransferase (ChAT) (Kawaguchi et al, 1995) and release acetylcholine (Ach) in the striatum. Electrophysiologically, ChAT+ cells are characterized by their depolarized RMP and high input resistance (Kawaguchi, 1993), as well as a membrane potential sag in response to hyperpolarizing current injections (Bennett & Wilson, 1998). Limited cortical and thalamic excitatory input (Meredith & Wouterlood, 1990; Lapper & Bolam, 1992) at proximal locations along the neuron (Wilson et al, 1990) is able to produce enough depolarization (1-5 mV) for ChAT+ interneurons to tonically fire action potentials, giving them the name “tonically active neurons” (TANs) (Zhou et al, 2002). These interneurons also receive GABAergic inputs originating from MSN axon collaterals (Bolam et al, 1986; Martone et al, 1992; DeBoer & Westerink, 1994; Bennett & Wilson, 1998) that may influence cell excitability. Dopamine is also thought to modulate ChAT+ neuronal output due to their expression of both dopamine D1- and D2- like receptors. Activation of D1-like receptors 18 increases Ach release through depolarization (Aosaki et al, 1998; Damsma et al, 1990; Consolo et al, 1992), while D2 activation inhibits Ach release (Lehmann & Langer, 1983; Stoof et al, 1992; Maurice et al, 2004). Their dendritic arborization offers dense, local innervation, primarily onto MSN distal dendrites (Bolam et al, 1984; Phelps et al, 1985; Zhou et al, 2002), overlapping with dopamine terminal fields (Bolam et al, 2000). In addition to synaptic transmission, these interneurons are also thought to release Ach via volume transmission, which has been shown to depolarize GABAergic fast-spiking interneurons (Chang & Kita 1992; Koós & Tepper, 2002; Zhou et al, 2002). ACh, the output transmitter of ChAT+ interneurons, is thought to alter MSN activity primarily by acting on M1 and M4 muscarinic receptors located on both types of MSNs (Hersch et al, 1994, Yan et al, 2001). Activation of these receptors will modulate voltage-gated ion channels and alter MSN excitability. ACh may reduce striatal glutamatergic transmission at corticostriatal synapses through presynaptic muscarinic receptors (Hernández-Echeagaray et al, 1998; Malenka & Kocsis, 1988; Sugita et al, 1991; Calabresi et al, 2000; Pakhotin & Bracci, 2007) and attenuate GABAergic inhibition of MSNs through postsynaptic nicotinic receptors on GABAergic interneurons (Koos & Tepper, 2002; Perez-Rosello et al, 2005). b. The GABAergic Calretinin-Positive Interneuron The least well described type of striatal GABAergic interneuron expresses calretinin (CR), a calcium binding protein, and GAD67, an enzyme used in GABA synthesis (Kubota et al, 1993). However, no other biochemical marker is expressed to 19 allow differentiation of these neurons, and they have not been described electrophysiologically. Immunostaining has revealed that CR+ interneurons are medium sized, and contain few aspiny, infrequently branched, dendrites (Bennett & Bolam, 1993). In the rodent, these cells account for less than 1% of striatal neurons (Tepper & Bolam, 2004), but are the most abundantly expressed interneuron in the human striatum (Cicchetti et al, 1998). Human striatal immunochemistry shows that CR is also expressed in a subset of large, cholinergic interneurons (Cicchetti et al, 1998) further compounding the role of CR+ interneurons in striatal physiology. c. The GABAergic Fast-Spiking Interneuron. A second type of aspiny striatal GABAergic interneuron, which expresses the calcium binding protein parvalbumin (PV) (Gerfen et al, 1985), has been characterized biochemically and electrophysiologically. In addition to expressing PV, these neurons also express GAD67 and the Kv3.1 potassium channel (Chesselet & Robbins, 1989; Lenz et al, 1994). These neurons are also called fast-spiking (FS) interneurons, based on their firing patterns evoked by depolarization. Just above threshold, FS interneurons fire several spikes in bursts, followed by a quiescent period (Kawaguchi, 1993; Koós & Tepper, 1999; Kubota & Kawaguchi, 2000), a pattern also known as “stuttering” (Figure 4A). With stronger depolarization, these interneurons fire repetitively with frequencies around 200 Hz (Koós & Tepper, 1999; Plotkin et al, 2005; Taverna et al, 2007) with largeamplitude afterhyperpolarizations and abrupt peaking (Koós & Tepper, 1999). These neurons are also characterized by their low input resistance and RMPs that are more 20 Figure 4. Fast Spiking Interneuron Characteristics. (A) Representative trace from a current clamp recording of a striatal FS/PV+ interneuron with a set holding potential of 60 mV. Current injection steps are 10 pA. Upon depolarizing current injections at threshold, FSI fire action potentials in bursts followed by a short quiescent period. Scale bars are 20 pA and 500 ms. (B) Biocytin injected FS/PV+ interneuron from rat. Calibration bar: 25 µm. Panel B reprinted with permission from Planert et al, 2010. See appendix for permission rights. 21 A B 22 negative than other striatal interneurons (approximately -75 mV) (Kawaguchi, 1993; Kubota & Kawaguchi, 2000; Taverna et al, 2007). Because these interneurons are identified by both biochemical and physiological characteristics, they will be called FS/PV+ interneurons for the entirety of this thesis. Morphologically, these cells‟ somata are slightly larger than MSNs, averaging 17 μm in diameter (Kita et al, 1990; Kawaguchi et al, 1995). Their aspiny dendrites form a restricted, yet compact and elaborate, arborization (Cowan et al, 1990; Kawaguchi, 1993) (Figure 4B). Electron microscopy studies have shown that FS/PV+ cells form synaptic connections on the soma and proximal dendrites of MSNs (Kita et al, 1990; Kubota & Kawaguchi, 2000) and other FS/PV+ interneurons (Kita et al, 1990; Chang & Kita, 1992). These findings have been confirmed electrophysiologically, indicating that FS/PV+ interneurons synaptically innervate MSNs (Taverna et al, 2007; Gittis et al, 2010) and other FS/PV+ interneurons (Gittis et al, 2010). In addition to the classical chemical synapse, FS/PV+ interneurons have also been shown to form gap junctions, or electrical synapses, with other FS/PV+ interneurons (Kawaguchi et al, 1995; Gibson et al, 1999; Koos & Tepper 1999; Hjorth et al, 2009; Galarreta & Hestrin 2001), suggesting that they integrate excitatory input on a much larger scale than the single cell. FS/PV+ cells receive cortical excitatory input onto their soma and proximal dendrites (Plotkin et al, 2005), while cortical-MSN asymmetric synapses are formed on distal dendritic spines (Bennett & Bolam, 1994). In addition, these interneurons receive multiple contacts from a single corticostriatal axon within a short distance (Ramanathan et al, 2002; Plotkin et al, 2005), priming the FS/PV+ interneuron with more powerful excitatory input than the MSN. Paired recordings with a presynaptic MSN and a 23 postsynaptic FS/PV+ cell have not found MSN-FS/PV+ connections (Koós & Tepper 1999; Taverna et al, 2007), suggesting that inhibitory input on the FS/PV+ cells originate from other striatal interneurons. Inhibitory pallidal axons may provide inhibitory input to striatal interneurons (Bevan et al, 1998; Bolam et al, 2000). A subpopulation of neurons originating in the globus pallidus project to the striatum and were anatomically shown to form synaptic contacts with PV+ interneurons (Bevan et al, 1998), although these results have not been electrophysiologically verified. Dopamine D5 (D1-like) and D2 receptors are also expressed on FS/PV+ interneurons. Dopamine mediates the depolarization of FS/PV+ cells via postsynaptic D5 receptors, while presynaptic D2 receptors are known to decrease inhibitory synaptic input onto these cells (Bracci et al, 2002). As described previously, FS/PV+ interneurons also receive cholinergic input that depolarizes the postsynaptic cell through nondesensitizing nicotinic receptors (Chang and Kita, 1992; Koós & Tepper, 2002). d. The GABAergic Low-Threshold Spiking Interneuron. The third type of striatal GABAergic interneuron co-expresses neuropeptide Y (NPY), somatostatin (SS), nitric oxide synthase (NOS), and NADPH diaphorase (Figueredo-Cardenas et al, 1996), and represents approximately 1-2% of striatal neurons (Kawaguchi et al, 1995). These aspiny interneurons were first electrophysiologically characterized by Dr. Kawaguchi, and based on their firing pattern are called persistent low thresholding spiking (PLTS) neurons due to their persistent depolarizing spikes in response to depolarizing current injection (Kawaguchi, 1993). Further studies revealed another type of striatal interneuron, similar to the PLTS, but that lacks the persistent 24 depolarization (Koós & Tepper, 1999). These cells were named low-threshold spiking (LTS) cells. Electrophysiological characteristics between the PLTS and LTS cells are similar, with a relatively depolarized RMP compared to FS/PV+ interneurons and MSNs, large input resistance, and action potential firing to small depolarizing current injections (Kawaguchi, 1993; Koós & Tepper, 1999; Kubota & Kawaguchi, 2000). These cells are also prone to firing “rebound spikes” after cessation of hyperpolarizing current pulses (Kawaguchi, 1993) and a time dependent Ih voltage sag during hyperpolarization (Kawaguchi, 1993; Koós & Tepper, 1999; Tepper et al, 2008) (Figure 5A). Because these interneurons are identified by both biochemical and physiological characteristics, they will be called LTS/NPY+ interneurons for the entirety of this thesis, although these cells co-express other peptides and have other physiological characteristics. Morphologically, LTS/NPY+ interneurons‟ soma is somewhat larger than that of a MSN (approximately 18 µM in diameter) (Kawaguchi, 1993; Kubota & Kawaguchi, 2000). Their aspiny dendrites radiate longitudinally with infrequent branching (Kawaguchi, 1993) (Figure 5B). These cells target the dendrites of spiny neurons, with smaller synaptic junctional areas (a morphological measure of postsynaptic receptor number) than FS/PV+ cells (Kubota & Kawaguchi, 2000). These LTS/NPY+ striatal interneurons receive their excitatory drive from the cortex (Vuillet et al, 1989), but the origin of their inhibitory drive remains poorly understood. Like FS/PV+ interneurons, LTS/NPY+ cells may also receive inhibitory inputs from the globus pallidus (Bevan et al, 1998), although it has not yet been shown electrophysiologically. In addition, LTS/NPY+ cells may be electrically coupled like striatal FS/PV+ interneurons (Kawaguchi, 1995; Koós & Tepper, 1999; Galarreta & 25 Figure 5. Characteristics of Low Threshold Spiking Interneurons. (A) Representative trace from a current clamp recording of a striatal LTS/NPY+ cell with a set holding potential of -60 mV. Current injection steps are 10 pA. Small depolarizing current injections are sufficient for LTS/NPY+ interneuron action potential firing (i). At cessation of hyperpolarizing current injection steps, LTS/NPY+ cells often have rebound firing (ii). Upon hyperpolarizing current injections, LTS/NPY+ cells display a voltage sag (iii). Scale bars are 40 pA and 1 s. (B) Z-stack image of biocytin injected NPYexpressing, LTS interneuron with long apical dendrites. Calibration bar: 50 µm. 26 A B i ii iii 27 Hestrin, 2001) and neocortical LTS neurons (Gibson et al, 1999), providing an added layer of complexity to their physiology. Paired recordings have shown that these striatal interneurons may participate in feedforward inhibition with MSNs (Koós & Tepper, 1999), although other studies have been unable to find traditional synaptic connections between these two types of cells, or any other striatal neuron (Gittis et al, 2010). These conflicting results will need to be fleshed out to more fully understand the role of LTS/NPY+ cells in striatal physiology. Dopamine D1Rs are also expressed on LTS/NPY+ interneurons (Rivera et al, 2002), and activation of these receptors has been shown to increase action potential firing through a membrane depolarization (Centonze et al, 2002). Upon stimulation, these cells are thought to also co-release nitric oxide (NO), SS, and NPY, and therefore can regulate brain physiology aside from inhibitory neurotransmission (Kawaguchi et al, 1995; Fu & van den Pol, 2007). NO is known to promote vasodilation, and therefore, LTS/NPY+ cells are likely to control striatal blood flow (Kelly et al, 1994). In addition, NO may act on ion channels and modulate the release of striatal neurotransmitters (Manzoni et al, 1992; Kawaguchi et al, 1995). A recent study showed that neocortical interneurons with similar biochemical and electrophysiological characteristics are inhibited by both NPY and SS application, suggesting an intrinsic control in LTS/NPY+ excitability (Fu & van den Pol, 2007). 28 V. Feedforward and Feedback Inhibition. Both GABAergic interneurons and MSNs provide striatal MSNs with local synaptic GABAergic input. The feedforward circuit of striatal inhibition consists of GABAergic interneurons which synaptically inhibit both D1+ and D2+ MSNs, while the feedback circuit with MSN lateral connections provides substantially less GABAergic input to MSNs (Planert et al, 2010). Anatomical studies show that both FS/PV+ and LTS/NPY+ interneurons form symmetric synapses with spiny cells (Koós & Tepper, 1999). The synaptic connections between FS/PV+ cells and MSNs are more widely studied than those connections between LTS/NPY+ cells and MSNs, and therefore much of the current knowledge regarding feedforward inhibition relies on the FS/PV+ type of interneuron. Extensive research with immunostaining, single cell filling, and paired recordings has shown that the predominant target of FS/PV+ cells is the soma and proximal dendrites of MSNs (Kita et al, 1990; Bennett & Bolam, 1994; Koós & Tepper, 1999; Kubota & Kawaguchi, 2000; Taverna et al, 2007). Paired recordings have shown high connection rates between these cell types and that single evoked spikes in FS/PV+ cells can evoke large IPSCs in connected MSNs, decreasing the transitory MSN firing rate (Koós & Tepper, 1999; Koós et al, 2004; Gustafson, et al 2006; Gittis et al, 2010). Two recent studies independently investigated FS/PV+ interneuron input onto both types of MSNs, and found that these interneuron target both D1+ and D2+ MSNs (Gittis et al, 2010; Planert et al, 2010). Because spiny cells receive inhibitory inputs from as many as 27 FS/PV+ interneurons and one FS/PV+ cell may be connected to 130 MSNs (Koós & Tepper, 1999), 29 feedforward inhibition is both powerful and widespread, regulating striatal output of both pathways. When LTS/NPY+ cells‟ connections onto MSNs were studied, presynaptic spikes evoked IPSCs with a high failure rate in the postsynaptic MSN, suggesting few connections between these cell types (Koos & Tepper, 1999; Gittis et al, 2010). Further insight between LTS/NPY+ and MSN connections will be offered below (see Discussion, Section X). GABAergic inhibition onto MSNs through MSN axon collaterals, also known as feedback inhibition, was once widely thought to provide the majority of inhibition onto MSNs because the major contact of MSN collaterals is other MSNs (Wilson & Groves, 1980; Somogyi et al, 1982; Bolam et al, 1983). These axons form synapses with MSN distal dendrites (Wilson & Groves, 1980), a poor location for extensive control, and it is currently thought that feedback inhibition does not provide the majority of GABAergic inhibition in spiny cells. In recent years, paired recordings between MSNs show synaptic connections, albeit small and with a high failure rate (Czubayko & Plenz, 2002; Tunstall et al, 2002; Koós et al, 2004; Taverna et al, 2004; Venance et al, 2004; Gustafson et al, 2006; Planert et al, 2010). These studies also found that MSN-MSN synaptic connections were unidirectional (Tepper et al, 2008). A brief hyperpolarization in a presynaptic MSN has been shown to delay action potential firing in a postsynaptic MSN (Koós & Tepper, 1999), and therefore feedback inhibition may function to help synchronize neuronal population activity (Plenz, 2003) or provide strong effects on local dendritic processing. 30 Immunostaining with SubP and Enk initially showed that D1+ and D2+ MSNs formed connections with each other (Bolam et al, 1983; Yung et al, 1996), but more comprehensive studies have shown that D1+ MSNs primarily form synapses with other D1+ MSNs, whereas D2+ MSNs form unidirectional connections with both types of MSNs (Taverna et al, 2008; Planert et al, 2010). Taverna et al. (2008) also showed that synapses with presynaptic D2+ MSNs are stronger due to the number of postsynaptic receptors that these synapses target, suggesting that the connectivity between MSNs is highly ordered and systematic in controlling output. The presence of gap junctions among MSNs is controversial as some groups have seen evidence for such coupling (Czubayko & Plenz 2002; Venance, 2004), while others have not (Taverna et al, 2008; Koós et al, 2004; Taverna et al, 2004; Tecuapetla et al, 2007). Although the age of mice may influence the presence or absence of electronic coupling, the role of gap junctions among striatal MSNs remains unknown. Therefore, although the majority of GABAergic contacts onto MSNs are made from axon collaterals, the primary source of GABA input comes from local GABAergic interneurons. VI. Modulation of Striatal MSNs a. Dopamine Dopamine‟s importance in proper striatal function is sufficiently illustrated by the detrimental effects of Parkinson‟s disease (PD), where the degeneration of dopaminergic cells in the SNpc results in dopamine loss in the striatum. Because dopamine receptors 31 are GPCRs, receptor activation leads to numerous downstream effects through various second messenger systems (see Introduction, Section VIIbi). Early studies investigated dopamine‟s regulation of MSN excitability, showing that D1 receptor (D1R) activation decreases spike activity in response to current injections (Uchimura et al, 1986; Akaike et al, 1987; Calabresi et al, 1987), while D2 receptor (D2R) activation enhances (Uchimura et al, 1986; Akaike et al, 1987) or suppresses MSN firing (Hooper et al, 1997). Although dopamine receptor agonists may not significantly alter D1+ or D2+ striatal MSN RMP, (cf. Uchimura et al, 1986 and Calabresi et al, 1987; Onn et al, 2003), many studies have investigated dopamine‟s influence on individual ionic currents, which may ultimately affect cell excitability (reviewed in Nicola et al, 2000). D1R agonists have been shown to decrease peak Na+ currents (Zhang et al, 1998; Surmeier et al, 1992; Schiffmann et al, 1995) and increase rectifying Kir2 currents (Pacheco-Cano et al, 1996), which set the RMP of striatal MSNs (Mermelstein et al, 1998). D1R activation has also been shown to increase L-type Ca2+ currents (Surmeier et al, 1995; Song & Surmeier, 1996; Hernández-López et al, 1997; Nicola et al, 2000), while decreasing P/Q/N- type Ca2+ currents (Surmeier et al, 1995; Salgado et al, 2005). These modulations underlie D1R activation‟s ability to promote the “down” state of MSN RMP, reducing cell excitability. However, at subthreshold membrane potentials, the increased L-type Ca2+ currents may promote D1+ MSN excitability. D2R mediated changes to MSN ionic conductances are less clear, as the predicted effects (opposite to those of D1Rs) are not often seen. For example, Na+ channels can be enhanced or suppressed by D2R agonists (Surmeier et al, 1992; Perez et al, 2006). It has also been shown that D2R agonists suppress L-type Ca2+ channel currents (Olson et al, 32 2005; Salgado et al, 2005), decreasing D2+ MSN excitability (Hernández-López et al, 2000), further compounding original theories of D2R function in striatal MSN physiology. A recent computational study included these alterations to ion channels, and the resultant model suggested that dopamine decreases the excitability of D2+ MSNs, while increasing D1+ MSN excitability (Moyer et al, 2007). Dopamine‟s effect on inhibitory neurotransmission is not completely understood. In general, D1R agonists increase GABAergic transmission (Harsing & Zigmond 1997; Radniknow & Misgeld, 1998; Arias-Montaño et al, 2007; Tecuapetla et al, 2007), while D2R agonists decrease inhibitory current (Delgado et al, 2000; Cooper & Stanford, 2001; Momiyama & Koga, 2001; Centonze et al, 2002; Guzmán et al, 2003; Tecuapetla et al, 2007) onto both D1+ and D2+ MSNs via a presynaptic mechanism (Tecuapetla et al, 2009). However, one study found that dopamine does not alter inhibitory currents in the dorsal striatum (Nicola & Malenka, 1998), suggesting that general statements about dopamine‟s influence over GABA currents are too simplistic. Indeed, dopamine may modulate both pre- and post-synaptic receptors, adding to the confusion over their effects on GABAergic currents. Further evidence of dopamine‟s role in the striatum comes from animal models of dopamine depletion. MSN excitability is enhanced when animals are subjected to dopamine depletion (Fino et al, 2007; Azdad et al, 2009); this modulation was attributed to a decrease in inactivating potassium currents, which play an important role in MSN repetitive spiking characteristics (Azdad et al, 2009). However, these studies did not differentiate between D1+ and D2+ neurons, which are likely to respond differently to dopamine depletion. Other studies have shown specifically that D2+ MSN excitability is 33 increased by dopamine depletion (Day et al, 2006; Day et al, 2008; Mallet et al, 2006; Shen et al, 2007). In addition, dopamine depletion has been shown to markedly increase glutamatergic input onto MSNs (Calabresi et al, 1993), while the inhibitory connections between MSNs decreases (Taverna et al, 2008), suggesting further implications in dopamine modulation in striatal MSN connectivity. b. Adenosine Adenosine is produced locally in the striatum, and therefore is an intrinsic signal/modulator for striatal function. With increased workload and cellular firing, intracellular ATP becomes dephosphorylated, causing adenosine levels to rise. Membrane transporters move the intracellular adenosine into the extracellular space where adenosine is able to modulate striatal function (Mori & Shindou, 2003; Schiffmann et al, 2007) by activating A1 and A2A receptors. Additionally, neuronal firing activates vesicular release of ATP (Cunha, 2001), and offers another mechanism to increase adenosine levels at the synapse. A1 receptors are found throughout the brain, while A2A receptors are found primarily in the striatum, nucleus accumbens, and the olfactory tubercle (Jarvis et al, 1989). Striatal A1 receptors are known to colocalize with D1Rs in striatonigral MSNs, while A2A receptors colocalize with D2Rs in striatopallidal MSNs (Fink et al, 1992; Schiffmann & Vanderhaeghen, 1993; Augood et al, 1994; Svenningsson et al, 1997). In these unique locations, adenosine and dopamine receptors have antagonistic functions because A1 receptors are inhibitory (D1Rs are faciliatory) and decrease presynaptic release, while A2A receptors have faciliatory actions (D2Rs are inhibitory) (Ferré et al, 1991; Ferré et al, 1997). 34 In addition, both A1 and D1 receptors and A2A and D2 receptors are thought to form heteromeric complexes (Ginés et al, 2000; Hillion et al, 2002; Canals et al, 2003; Ciruela et al, 2004), and intermembrane interactions may further antagonize receptor signaling. Studies have shown that stimulation of A1 receptors decreases dopamine‟s binding affinity to D1Rs (Ferré et al, 1998; Ginés et al, 2000). Similarly, A2A stimulation decreases dopamine‟s binding to the D2R (Ferré et al, 1991). Although the majority of A2A receptors are located post-synaptically (Rosin et al, 2003), adenosine modulates both excitatory and inhibitory transmission in the striatum. Intrastriatal stimulation in the presence of A2A agonists decreases evoked IPSCs in D2+ neurons (Mori et al, 1996), whereas agonists enhance GABA release in the GP through pre-synaptic mechanisms in striatopallidal terminals (Floran et al, 2005). Stimulation of presynaptic A2A receptors at glutamate terminals has been shown to increase glutamate release (Popoli et al, 1995; Rosin et al, 2003), whereas postsynaptic receptors reduce the amplitude of excitatory currents in D2+ MSNs (Nörenberg et al, 1997). In periods of high activity, adenosine provides additional fine-tuning in striatal function by increasing the efficiency of excitatory transmission in striatopallidal neurons. However, when considering striatal modulating transmitters, it is crucial to not only evaluate dopamine and adenosine, but the effects of their antagonistic, and complex, signaling. For the studies presented in this thesis, adenosine receptor activation is not considered although previous work in our lab has shown that A2A receptors are active in our striatal slice preparation (Ade, unpublished observations), and other studies have shown effects with A1 receptor antagonists (Choi & Lovinger, 1995). 35 VII. Post-translational Modifications of MSN GABAergic Currents. Post-translational modifications can alter receptor number and spatial orientation, as well as receptor biophysical properties. These modifications are essential to inhibitory network efficiency, and are a critical component in understanding the basic physiology of GABAA receptors. Although many of the experiments referenced here were not conducted in the striatum, these concepts may be generalized to the striatum, where similar events are likely to occur. a. Receptor Assembly and Membrane Insertion Receptor assembly begins in the endoplasmic reticulum where individual subunits interact to form heteromeric channels that typically contain 2α-, 2β-, and 1γ-subunits (Moss & Smart, 2001; Lüscher & Keller, 2004). Subsequent expression in the plasma membrane relies on the interplay of several key anchoring and receptor targeting proteins. The protein Plic-1 has been shown to enhance membrane insertion by increasing the half-life of intracellular pools of GABAA receptors and inhibiting receptor degradation (Bedford et al, 2001; Saliba et al, 2008). Blocking the interactions between Plic-1 and GABAA receptors has been shown to decrease GABAA receptor mediated currents (Bedford et al, 2001), presumably through a decrease in receptor insertion into the plasma membrane. Surface transport and synapse targeting is another area in which the GABAA receptor is under tight regulation by proteins such as the GABAA receptor-associated protein (GABARAP). This protein binds specifically in the intracellular domain of γ GABAA receptor subunits (Nymann-Anderson et al, 2002) and has been shown to play an 36 important role in membrane trafficking as well as anchoring a receptor to the cytoskeleton (Kittler & Moss, 2003). Once receptors arrive at the synapse, additional proteins provide receptor stability within the plasma membrane. The majority of inhibitory synaptic targeting is thought to depend on potential interactions between the γ2 receptor subunit and the gephyrin protein (Kneussel & Betz, 2000; Jacob et al, 2005), as deletion of subunit or protein disrupts the number of receptors that reach the cell surface (Essrich et al, 1998; Kneussel et al, 1999). Therefore, intracellular and cell-membrane mediated GABAA receptor stability is dependent upon various proteins that help to maintain the efficiency of fast synaptic inhibition. It has been suggested that receptor insertion primarily occurs at extrasynaptic sites (Lüscher & Keller, 2004; Bogdanov et al, 2006), and GABAA receptors may utilize lateral mobility to increase synaptic receptor numbers in the post-synaptic zone (Bogdanov et al, 2006; Bannai et al, 2009). Indeed, unique receptor binding studies showed that when synaptic receptors were blocked, extrasynaptic receptors diffused through the plasma membrane to become functional „synaptic‟ receptors (Thomas et al, 2005). The investigators also found that this lateral mobility only takes approximately two minutes for extrasynaptic receptors to move into synaptic locations (Thomas et al, 2005), suggesting that receptor diffusion may provide a means for receptor number consistency and maintaining homeostasis. 37 b. Phosphorylation Kinases are a group of proteins that catalyze the phosphorylation of many proteins. This constitutively active intracellular process involves the transfer of ATP‟s highly charged phosphate group to the hydroxyl group on a serine or threonine residue. Hydrolysis of the phosphate group is aided by a phosphatase. The protein‟s charge is altered after phosphorylation, often resulting in a conformational change and changes in protein activity. Biochemical studies have shown that various protein kinases target ion channels, and therefore a great number of studies have been devoted to determining the effects and implications to GABAA receptor phosphorylation. Biochemical studies have shown that GABAA receptor subunits can be phosphorylated by 3'-5'-cyclic adenosine monophosphate (cAMP) dependent kinase (PKA) (Heuschneider & Schwartz, 1989), Ca2+/phospholipid-dependent protein kinase (PKC) (Sigel & Baur, 1988), type II Ca2+/calmodulin-dependent protein kinase (Machu et al, 1993), and protein tyrosine kinase (Moss et al, 1995). This thesis will only focus on PKA and PKC actions because most of the phosphorylation studies on striatal GABAA receptors have shown effects with these two kinases. PKA becomes activated when cAMP binds to the inactive form of the kinase which frees the catalytic subunit from the regulatory domains. PKC, on the other hand, is activated when Ca2+ binds to the kinase‟s regulatory domain, separating it from the catalytic domain. Protein phosphatase 1 (PP-1) and calcineurin are phosphatases that have been shown to inhibit both PKA and PKC activity in vitro (Surmeier et al, 1995; Yan & Surmeier, 1997; Flores-Hernandez et al, 2000; Foster et al, 2003; Hourez et al, 2005), providing additional regulation to the phosphorylation cascade. PP-1 also controls 38 the phosphorylation state of many other phosphoproteins including receptors, ion channels, and transcription factors. PKA phosphorylates serine residues (S) in β1 (S409) and β3 (S408/9) GABAA receptor subunits, while the β2 subunit is not phosphorylated by this kinase (McDonald et al, 1998). PKC targets all GABAA receptor β subunits on a conserved serine residue (S409 in β1 and β3 subunits and S410 in β2 subunits) in the intracellular domain (McDonald et al, 1998). This latter kinase has also been shown to phosphorylate γ2 GABAA receptor subunits on S327 (Moss et al, 1992a). The mechanisms behind PKA phosphorylation appear to rely on key signaling scaffold molecules because PKA itself is unable to directly bind to the GABAA receptor subunits (Brandon et al, 2003). AKAP79/150, a neuronal A-kinase anchoring protein, has been shown to bind to PKA as well as both β1 and β3 subunits, effectively targeting PKA to these GABAA receptor subunits (Brandon et al, 2003). Similar studies have shown that PKC is able to bind directly and, with the help of the receptor for activated C kinase (RACK-1), to target β residues (Brandon et al, 1999). Because RACK-1 is not necessary for PKC to bind to these subunits, it may increase affinity between kinase and subunit (Brandon et al, 1999). GABAA receptor function is altered by PKA phosphorylation, although its effects on GABAA receptors appears diverse, since some studies show an enhancement in GABAergic whole-cell currents (Kano & Konnerth 1992; Feigenspan & Bormann, 1994; Veruki & Yeh, 1994; Kapur & Macdonald, 1996; Nusser et al, 1999), while others show inhibition (Porter et al, 1990; Moss et al, 1992b; Robello et al, 1993; Brünig et al, 1999). These discrepancies are likely due to the varying brain regions investigated since GABAA 39 receptor β subunit expression differs throughout the brain. Indeed, PKA‟s effects on β1 and β3 GABAA receptor subunits are quite different. PKA increases whole-cell GABAA currents in cells expressing the β3 subunit (McDonald et al, 1998; Nusser et al, 1999), while phosphorylation of the β1 subunit decreases whole-cell GABAA currents (Moss et al, 1992b; Flores-Hernandez et al, 2000). Mutation of S408 to an uncharged alanine residue on the β3 subunit showed effects of phosphorylation similar to those seen with the β1 subunit, suggesting that distinct phosphorylated serine residues underlie the functional differences in β subunit phosphorylation (McDonald et al, 1998). It has also been found that β3 subunits exhibit basal phosphorylation by PKA (McDonald et al, 1998; Poisbeau et al, 1999; Brandon et al 2003), which may also be due to the additional phosphorylation site on the β3 subunit. PKC‟s effects on GABAA receptors‟ functional activity are also confounded as PKC has been shown to enhance (Poisbeau et al, 1999; Choi et al, 2008) and inhibit GABAA receptors (Brandon et al, 2000; Brussaard et al, 2000; Filappova et al, 2000; Herring et al, 2005). Although GABAA receptor phosphorylation may affect cell surface expression, and changes in receptor levels may modulate inhibitory conductance (see Introduction, Section VIIc), short-term effects of phosphorylation have also been noted. PKA phosphorylation of GABAergic currents has been shown to decrease mIPSC amplitude in cortical granule cells (Poisbeau et al, 1999) and cerebellar Purkinje cells (Kano & Konnerth, 1992), but increase it in olfactory granule neurons (Nusser et al, 1999); these differences are likely due to β subunit expression. On the other hand, many studies have shown that PKC phosphorylation increases mIPSC amplitude (Poisbeau et al, 1999; 40 Jovanovic et al, 2004; Terunuma et al, 2008) and tonic current (Choi et al, 2008) in cortex and hippocampus. Many studies suggest that phosphorylation may alter GABAergic currents through receptor desensitization, changes in single-channel open probability, and ligandreceptor interactions. Experiments in rat hippocampal neurons showed that phosphorylation induces a significant conformational change that directly affects and destabilizes ligand affinity for the receptor (Jones &Westbrook, 1997). Further, a study investigating both β1- and β3- containing receptors expressed in HEK 293 cells showed that PKA phosphorylation regulates desensitization and deactivation parameters by increasing the fast phases of desensitization and prolonging deactivation (Hinkle & Macdonald, 2003). A recent study found that PKA phosphorylation increased current through α4β3δ receptors by increasing their single channel mean open frequency (Tang et al, 2010), although a previous study by Angelotti et al. (1993) failed to discern changes in open and close times, bursting parameters, and conductance due to PKA phosphorylation. i. Dopamine’s influence on phosphorylation. Dopamine receptors are GPCRs whose second messenger system provides another layer of regulation to PKA phosphorylation. D1Rs are coupled to the Gs protein, which promotes adenylyl cyclase (AC) and further activates cAMP and PKA phosphorylation (Stoof & Kebabian, 1981; Walaas & Greengard, 1991; Stoof & Kebabian, 1984). On the other hand, D2Rs are inhibitory as they activate the Gi/o protein that blocks cAMP accumulation, preventing protein phosphorylation (Stoof & Kebabian, 1981). Additionally, all striatal MSNs express the dopamine- and cAMP- regulated- 41 Figure 6. PKA Phosphorylation Cascade is Regulated by Dopamine. D1R stimulation (left) promotes AC activation and allows cAMP to accumulate and activate PKA. PKA phosphorylates target proteins such as β subunits of the GABAA receptor that affect the amount of current passing through receptors. PKA also phosphorylates DARPP-32 at T34, which blocks PP-1 activity. PP-1 dephosphorylates target proteins. PKA also activates PP-2A which dephosphorylates T75 on DARPP-32. The phosphorylated T75 protein further blocks PKA activity. Cdk-5 is the kinase responsible for phosphorylating T75 of DARPP-32. D2R activation (right) inhibits AC activity and further blocks PKA phosphorylation by activating PP-2B, a phosphatase that dephosphorylates DARPP-32 at T34. Abbreviations: AC, adenylyl cyclase; cAMP, 3'-5'-cyclic adenosine monophosphate; PKA, protein kinase A; DARPP-32, dopamine- and cAMP- regulated-phosphoprotein of 32 kD; PP-2A, protein phosphatase 2A ; T, threonine residue; PP-1, protein phosphatase 1; Cdk-5, cyclin-dependent kinase 5; PP-2B, protein phosphatase 2B. 42 D1R D2R Gi/o Gs AC cAMP PKA PP-2A D32 T75 D32 T34 D32 PT75 Cdk-5 D32 PT34 PP-2B Target protein Target protein P PP-1 β3 β1 β3P GABA currents β1P GABA currents 43 phosphoprotein of 32 kD (DARPP-32) (Ouimet et al, 1984; Walaas & Greengard, 1984; Ouimet & Greengard, 1990; Matamales et al, 2009; Rajput et al, 2009). This protein is phosphorylated at threonine residue (T) T34 by PKA (Hemmings et al, 1984), and in this phosphorylated state, DARPP-32 acts as a key inhibitor of PP-1 (Hemmings et al, 1984; reviewed in Greengard et al, 1999) to further regulate the phosphorylation cascade. Under low cAMP and PKA levels, DARPP-32 is phosphorylated on T75 by cyclindependent kinase 5 (Cdk5); in this state, DARPP-32 inhibits PKA activity (Bibb et al, 1999). When PKA levels rise, protein phosphatase 2A (PP-2A) catalyzes the dephosphorylation of T75, providing a positive feedback loop in PKA phosphorylation (Nishi et al, 2000). D1R agonists have been shown to increase DARPP-32 phosphorylation (Nishi et al, 1997; Svenningsson et al, 1998; Kuroiwa et al, 2008), while D2 agonists decrease this phosphorylation (Nishi et al, 1997; Lindskog et al, 1999) due to the receptors‟ influence over cAMP. Thus, D1R activation will activate PKA phosphorylation by both cAMP accumulation and phosphorylated T34-DARPP-32 activity; D2R stimulation will block PKA phosphorylation by blocking AC activity and dephosphorylating T75-DARPP-32, rending the protein unable to block PP-1 activity (Figure 6). Although dopamine is not thought to directly activate PKC in young mice (Rashid et al, 2007), dopamine may regulate downstream players involved in the inactivation of PKC. Dopamine receptors are linked to the DARPP-32/PP-1 pathway that primarily regulates PKA activity and function, yet PP-1 has also been shown to dephosphorylate PKC-phosphorylated residues in the nucleus accumbens (Snyder et al, 1998), suggesting that dopamine may regulate PKC activity though indirect mechanisms. 44 A relationship between dopamine, phosphorylation, and GABA receptor function was established in hippocampal slices (Terunuma et al, 2004): a D1R agonist was shown to increase GABAA currents in D1+ neurons via a PKA phosphorylated β3 subunit. A study by Jim Surmeier‟s group has also made this link in striatal MSN GABAergic currents. They showed a reduction in MSN GABA currents in response to D1R activation, through the PKA/DARPP-32/PP-1 signaling cascade‟s phosphorylation of the GABAA receptor β1 subunit (Flores-Hernandez et al, 2000). This thesis serves to provide the essential link among dopamine, PKA phosphorylation, and MSN tonic currents. c. Endocytosis and Recycling Receptor recycling is a common feature of GABAA receptor populations that affects receptor numbers in the cell membrane and, therefore, the extent of GABAergic inhibition. GABAA receptor endocytosis is predominately mediated through a clathrindependent internalization process (Kittler et al, 2000). Under basal conditions, approximately 25% of β3-containing receptors can be internalized within 30 minutes (Kittler et al, 2004), suggesting that receptor numbers are dynamic. A clathrin-adaptor protein 2 (AP2) complex is critical in recruiting cell surface proteins into clathrin-coated pits. Direct binding sites between the GABAA receptor β1-3 and γ2 subunits and the μ2 subunit of AP2 link the two proteins together (Kittler et al, 2000; Kittler et al, 2005). The β2 GABAA receptor subunit contains a di-leucine motif that is important for clathrin-dependent endocytosis (Herring et al, 2003), while β1 and β3 subunits bind to AP2 in an atypical basic binding pocket (401KTHLRRRSSQLK412 in 45 Figure 7. β3 Subunit-Mediated Endocytosis. Endocytosis is blocked when the β3 GABAA receptor subunit is phosphorylated (left). Phosphorylation of this subunit also increases current through the receptor. Upon dephosphorylation, AP2 binds to the β3 subunit and mediates endocytosis, with the help of clathrin and dynamin (right). Dephosphorylation of the subunit also results in decreased current through the receptor. Abbreviations: P, phosphorylated; AKAP, neuronal A-kinase anchoring protein; PKA, protein kinase A; PP-1, protein phosphatase 1; AP2, adaptor protein 2. 46 β3 subunit β3 subunit P S408/409 PP1 A K A P Cl-PKA AP2 Cl- Clathrin Dynamin AP2 Endocytosis 47 the β3 subunit), which also contains the major PKA and PKC phosphorylation sites (S408/409 in the β3 subunit) (Kittler et al, 2005). Phosphorylation of these serine residues reduces subunit association with AP2 (Kittler et al, 2005), increases receptor surface expression (Jacob et al, 2009), and increases GABAergic inhibition (Kittler et al, 2005). Likewise, blockade of the phosphorylation site (with peptide blockade or mutations of serine residues to alanine residues) increases mIPSC amplitudes (Kittler et al, 2005; Chen et al, 2006; Jacob et al, 2009) (Figure 7). Further evidence for an interaction between endocytosis and PKA/PKC phosphorylation on GABAA receptor β3 subunits comes from a study that pharmacologically enhanced β3 subunit PKC phosphorylation and reported increased cell-surface levels of β3-containing GABAA receptors and decreased AP2 binding (Brandon et al, 2002). The γ2 subunit has two distinct AP2 binding sites which can regulate receptor endocytosis. AP2 can bind at tyrosine residue (Y) binding site 365YGYECL370 with high affinity (Kittler et al, 2008). Both tyrosine residues are principal phosphorylation sites for Src tyrosine kinases and phosphorylation of these residues decreases AP2 binding to the γ2 subunit, resulting in increased cell surface receptor expression and mIPSC amplitudes (Kittler et al, 2008; Smith et al, 2008). Another AP2 binding site on the γ2 subunit is similar to the binding pocket for β subunits as it is highly basic (324RKPSKDKDKKK335) (Smith et al, 2008). Phosphorylation of these serine residues in the γ2 subunit blocks AP2 binding, resulting in increased IPSC amplitude (Smith et al, 2008). Because dopamine is known to alter phosphorylation rates in the brain, this neuromodulator may alter the integrity of inhibitory transmission through subunit 48 phosphorylation state and receptor numbers at the cell surface. Indeed, activation of a dopamine D3 receptor (of the D2R family) was shown to suppress the efficacy of GABAergic transmission by increasing phospho-dependent GABAA receptor endocytosis (Chen et al, 2006). It was further shown that decreased β3 phosphorylation due to status epilepticus induced GABAA receptor internalization, contributing to compromised GABAergic signaling (Terunuma et al, 2008). When AP2 binds to the membrane-associated receptor, other key elements of the endocytotic mechanism, clatherin and dynamin, are recruited to form the clathrin coat (Marsh &McMahon, 1999; Takei et al, 1999). Associations with dynamin have been shown to support endocytosis since blocking this protein, and therefore inhibiting endocytosis, has been shown to increase receptor numbers at the cell surface (Kittler et al, 2000) and increase mIPSC amplitudes (Kittler et al, 2000; Herring et al 2003; van Rijnsoever et al 2005; Chen et al, 2006). Upon internalization, clatherin coats are removed and some GABAA receptors are degraded by lysosomes (Jacob et al, 2009), while the majority are recycled to the plasma membrane (Kittler et al, 2004; Thomas et al, 2005). 49 HYPOTHESIS: GABAergic medium spiny neuron tonic current and excitability is largely regulated by dopamine receptor activation and PKA phosphorylation of the GABAA receptor β3 subunit. 50 MATERIALS AND METHODS I. Animals Bacterial artificial chromosome (BAC) Drd2 EGFP (Gong et al, 2003; courtesy of Dr. Lovinger, National Institutes of Health), BAC Drd1a tdTomato (Shuen et al, 2008; courtesy of Dr. Calakos, Duke University) mice, and their progeny (BAC Drd2 EGFP;BAC Drd1a tdTomato) were used for the reliable detection of D2+ and D1+ MSNs in the same animal. GABAA receptor δ subunit knock-out (KO) (Mihalek et al, 1999; Dr. Huntsman, Children‟s Hospital) mice were also used in a subset of experiments. In addition, GABAA receptor subunit β3 conditional KO mice were generated from floxβ3 mice (Jackson Lab, B6;129-Gabrb3tm2.1Geh/J; Ferguson et al, 2007) and either Drd2-Cre (Gong et al, 2007) or Dlx5/6-Cre (Stenman et al, 2003) (both courtesy of Dr. Xu, Georgetown University) mice. In some experiments D2+ MSNs were visually identified by a cross with the Rosa-EGFP reporter mouse where cells that express Cre recombinase also express EGFP (Jackson Lab, B6;129-Gt(Rosa)26Sor<tm2Sho>; Mao et al, 2001). To accurately identify NPY+ interneurons, BAC NPY mice (Jackson Lab, B6.FVB-Tg(Npy-hrGFP)1Lowl/J; Fu & van den Pol, 2007; courtesy of Dr. Zukowska, Georgetown University) were used and crossed with BAC Drd1a tdTomato mice for simultaneous visualization of NPY+ interneurons and D1+ MSNs. In general, mice of postnatal day (p) 15-21 were used for the experiments, but in a subset of studies, adult mice (p33-38) were used as well. The sex of the animal was noted, but as there were no detectable electrophysiological differences between males and females, these data were pooled. 51 Figure 8. Generation of Conditional β3 Subunit KO Mice using the Cre-Lox System. (A) Mice that express the floxed β3 allele were crossed with mice that express Cre recombinase under the expression of the D2R promoter. Their progeny had D2R expressing neurons that lacked the β3 subunit, while non-D2R expressing neurons still contained the β3 subunit. (B) Mice that express EGFP under the ROSA promoter, with a STOP codon between, were crossed with Drd2-Cre recombinase expressing mice, so progeny would have EGFP+, D2+ neurons that expressed Cre recombinase. Black arrow is loxP site. 52 A X Floxed β3 subunit Cre under D2 promoter Excised β3 subunit B Gt(ROSA)26Sor STOP EGFP X Drd-2 Drd-2 EGFP+ Cells expressing Cre 53 CRE II. Cre-lox System The Cre-lox recombination system uses the enzyme Cre recombinase to remove a targeted sequence of DNA. In short, the target gene is flanked by two 34- base pair loxP sequences situated in the same direction, and Cre recombinase (with telomerase- and restriction enzyme-like activities) excises portions of each loxP site. The strands of DNA are reannealed with DNA ligase, and in this new strand of DNA, the gene of interest is deleted. For a conditional KO animal, Cre recombinase is expressed under a promoter that is specific for a certain cell type or tissue. Therefore, a conditional KO animal lacks the gene of interest only in Cre-expressing tissue. Floxβ3 mice express two loxP sites that flank exon 3 of the GABAA receptor β3 subunit gene (flox = flanked by loxP). Excision of the 68 basepair exon 3 will leave a mutant β3 subunit product with a premature stop codon, rendering the subunit nonfunctional, as exon 3 contains the genetic code for all transmembrane domains of the subunit (Ferguson et al, 2007). Drd2-Cre mice express Cre recombinase under the dopamine D2 receptor promoter, and therefore, Cre recombinase is solely expressed in D2+ striatopallidal neurons (Figure 8A). Dlx5/6-Cre mice express Cre recombinase under the Dlx5/6 promoter, and therefore Cre recombinase is expressed in GABAergic forebrain neurons (Stenman et al, 2003). Rosa-EGFP mice express EGFP under the ubiquitous Rosa promoter, with a floxed STOP codon between them. When crossed with mice expressing Cre recombinase, progeny will express EGFP in neurons where Cre recombinase is present (Figure 8B). 54 Dlx5/6-Cre+/-;floxTrkB+/- mice were originally obtained (Dr. Xu, Georgetown University), and crossed with wild-type C57BL/6J mice to yield pure Dlx5/6-Cre+/mice. These mice were then crossed with floxβ3+/+ animals to yield Dlx5/6-Cre+/-;floxβ3+/- mice, which were crossed with floxβ3+/+ mice to yield conditional β3 subunit KO (Dlx5/6-KO) and heterozygous animals (Dlx5/6-Cre;Hetβ3) that were used for experiments. Drd2-Cre+/- mice were bred in a similar fashion (described thereafter as Drd2-KO, Drd2-Cre;Hetβ3) (Figure 8A). Rosa-EGFP+/+ reporter mice were crossed with Drd2-Cre+/- mice and floxβ3+/+ mice, and their progeny were crossed (Drd2-Cre+/-;Rosa-EGFP+/- X Rosa-EGFP+/-;floxβ3+/-). Progeny (Drd2-Cre+/-;Rosa-EGFP+/+;floxβ3+/-) were mated with Rosa-EGFP+/-;floxβ3+/- mice to generate the Drd2-Cre+/-;Rosa-EGFP+/+;floxβ3+/+ mouse. Therefore, these mice have the β3 subunit knocked out in EGFP+, D2+ neurons. III. Genotyping BAC Drd1a tdTomato, BAC Drd2 EGFP;BAC Drd1a tdTomato, and BAC NPY mice were genotyped as 1-2 day old pups using a fluorescence microscope. Heads were placed under the lens, and fluorescent light allowed visualization of fluorescent cortical areas. This method proved to be nearly 100% accurate in determining if an animal expressed a fluorescent protein. However, this method of genotyping is not useful for BAC Drd2 EGFP mice because levels of D2R expressing neurons are low in cortical areas. δ subunit KO mice were bred as heterozygotes and genotyped for the specific δ subunit deletion by Peijun Li with Southern blot analysis. 55 Table 1. PCR Reaction Protocols. Details for Cre recombinase and Rosa-EGFP PCR thermocycler protocols. 56 Cre Recombinase (400 bp) Rosa-EGFP (Mutant; 173 bp) Rosa-EGFP (Wild Type; 410 bp) Holding 92°C; 2 min 95°C; 15 min 95°C; 15 min Melting 92°C; 30 sec 94°C; 30 sec 94°C; 30 sec Annealing 61°C; 45 sec 60°C; 1 min 66.8°C; 1 min Holding 72°C; 1 min 72°C; 1 min 72°C; 1 min Repeat steps 24 30 cycles 35 cycles 35 cycles Capping 72°C; 7 min 72°C; 2 min 72°C; 2 min Maintain 4°C 10°C 10°C 57 Conditional β3 KO mice were genotyped using the polymerase chain reaction (PCR) method for the floxβ3 allele, Cre recombinase expression, and (when appropriate) Rosa-EGFP. Snips of mouse tails were cut between p7-10, and total genomic DNA was isolated using the DNeasy Tissue Kit (Qiagen, Valencia, CA) according to the manufacturer‟s instructions. Genomic DNA was amplified using primers (all from Invitrogen) for the loxP site in exon 3 of the β3 subunit (FATTCGCCTGAGACCCGACT, R- GTTCATCCCCACGCAGAC), Cre recombinase (FGGATGAGGTTCGCAAGAACC, R-CCATGAGTGAACGAACCTGG), Rosa-EGFP (Mutant F- AAFTTCATCTGCACCACCG, R- TCCTTGAAGAAGATGGTGCG; Wild Type F- CGTGATCTGCAACTCGACTC, R- GGAGCGGGAGAAATGGATATG). The floxβ3 PCR product was quite difficult to detect with standard PCR, so samples were sent to Transnetyx, Inc., where the presence of this allele was determined using real time PCR. Forward and reverse primers (1 µL of each), 2 µL of isolated DNA, and distilled water (to a final volume of 25 µL) were added to Ready-to-Go Beads (Amersham Pharmacia Biotech, Piscataway, NJ). The PCR assay for Rosa-EGFP required the use of Thermo-Start (Thermo Scientific), and therefore 12.5 µL of 2X Thermo-Start PCR Master Mix, 1 µL of each primer, 2 µL of DNA, and distilled water (to a final volume of 25 µL) were added to PCR reaction tubes. DNA was amplified in a PTC-100 thermocycler (MJ-Research, Inc.) with varying protocols, based on the gene of interest (Table 1). Amplified DNA (10 µL) and 6x Loading dye (2 µL) were mixed together and added to a 2% agarose gel. DNA samples were separated by electrophoresis at 90 V for 58 approximately 40 minutes. Gels were stained with ethidium bromide (Gibco BRL) and bands were visualized with a Kodak EDAS 290 camera (Kodak, Rochester, NY) and Foto/UV 21 lamps (Fotodyne, Harland, WI). IV. Acute Slice Preparation Mice were sacrificed by decapitation in agreement with the guidelines of the AMVA Panel on Euthanasia and the Georgetown University ACUC. The whole brain was removed and placed in an ice-cold slicing solution containing (in mM): NaCl (85), KCl (2.5), CaCl2 (1), MgCl2 (4), NaH2PO4 (1), NaHCO3 (25), glucose (25), sucrose (75) (all from Sigma). Corticostriatal coronal or sagittal slices (250 – 300 µm) were prepared using a Vibratome 3000 Plus Sectioning System (Vibratome) and were incubated in slicing solution at 32°C for 30 minutes. Slices recovered for 30 minutes at 32°C in artificial cerebrospinal fluid (aCSF) containing (in mM): NaCl (124), KCl (4.5), Na2HPO4 (1.2), CaCl2 (2), MgCl2 (1), and dextrose (10) of 305 mOsm and were maintained at room temperature. During experiments, slices were submerged in the recording chamber and continuously perfused (2-3 mL/min) with aCSF at room temperature, 22-24°C, or physiological temperature, 32°C, where noted. All solutions were maintained at pH 7.4 by continuous bubbling with 95% O2/5% CO2. V. Cell Identification. Although BAC Drd2 EGFP;BAC Drd1a tdTomato mice allow for simultaneous visualization of D2+ and D1+ MSNs (Shuen et al., 2008), these mice were not available for the entirety of this thesis research. When BAC Drd2 EGFP mice were used, “green” 59 neurons were identified as D2+ neurons, and “non-green” neurons as D1+ neurons. When BAC Drd1a tdTomato mice were used, “red” neurons were identified as D1+ MSNs and “non-red” neurons as D2+ MSNs. No remarkable differences were observed between D1+ and D2+ cells from either mouse strain, and data was pooled as D2+ and D1+ MSNs, regardless of the transgenic mouse from which they originated. NPY cells in the BAC NPY strain were identified as “green” neurons, and “nongreen” cells with MSN-like firing properties were deemed to be MSNs. When BAC NPY; BAC Drd1a tdTomato mice were used to aid in the detection of MSNs, NPY+ interneurons were “green”, while “red” neurons were identified as D1+ MSNs. MSN properties observed in both BAC NPY strains were confirmed with MSNs from other strains where MSNs were fluorescently labeled. In experiments where the β3 subunit was deleted, MSNs were not tagged with fluorescent proteins, and MSN identification was largely determined by soma size. In a few experiments, Rosa-EGFP expression allowed visualization of Cre-expressing D2+ MSNs, and “green” neurons were assumed to be D2+ neurons, while the “non-green” cells were assumed to be D1+ MSNs. VI. Whole Cell Recordings Slices were visualized under an upright microscope (E600FN; Nikon) equipped with Nomarski optics and an electrically insulated 60X water-immersion objective with a long working distance (2 mm) and a high numerical aperture (1.0). Recording pipettes were pulled on a vertical pipette puller from borosilicate glass capillaries (Wiretrol II; 60 Drummond) and filled with CsCl-, KCl- or KGluconate-based internal solutions. The CsCl-based internal solution (CsCl internal) contained (in mM): CsCl (145), HEPES (10), ATP.Mg (5), GTP.Na (0.2), EGTA (10), adjusted to pH 7.2 with CsOH. In KClbased internal solution (KCl internal), CsCl was replaced with equimolar KCl and pH was adjusted with KOH. For Kgluconate-based internal solutions (Kgluconate internal), CsCl was replaced with equimolar Kgluconate and pH was adjusted with KOH. Single and dual voltage-clamp recordings were performed using the whole-cell configuration of the patch-clamp technique at a pipette voltage of -60 mV using the Axopatch 200B and 1D amplifiers (Molecular Devices). Access resistance was monitored during the recordings, and experiments with > 20% change were discarded. When potassium based solutions were used, the baseline membrane potential for currentclamp recordings was set at -70 mV or -60 mV, as noted, before each series of current step injection protocols. Rheobase current was defined as the first current step, within a series of 10 or 20 pA steps, from the set membrane potential that elicited an action potential. For evoked current recordings, a single bipolar stimulating electrode was placed within the striatal tissue (in coronal slices) or the globus pallidus (in sagittal slices). Stimulation parameters were altered to evoke inhibitory currents of 100 – 400 pA in amplitude. For anatomical reconstructions, biocytin (AnaSpec, Fremont, CA) was added at a final concentration of 1% to the intracellular solution. Upon achieving whole-cell recordings, biocytin was allowed to diffuse through the cell for approximately 5 minutes. After the whole cell recording, the pipette was gently removed to form an outside-out 61 patch, and the slices were fixed in 4% paraformaldehyde (PFA) for 30 minutes. After fixation, slices were washed in phosphate buffer saline (PBS) and stained with Texas redconjugated avidin-D dye (Invitrogen, Carlsbad, CA) at 2.5 µL/mL for 2 hours. Slices were mounted on microscope slides and processed for confocal imaging as described in Methods, Section X. VII. Drugs and Peptides Stock solutions of bicuculline methbromide (BMR), 6-chloro-1-phenyl-2,3,4,5tetrahydro-1H-3-benzazepine-7,8-diol (SKF-81297), quinpirole, sulpiride, 7-chloro-3methyl-1-phenyl-1,2,4,5-tetrahydro-3-benzazepin-8-ol (SCH 23390), GABA, 1,2,3,4Tetrahydro-6-nitro-2,3-dioxo-benzo[f]quinoxaline-7-sulfonamide disodium salt hydrate (NBQX), and (±)-3-(2-Carboxypiperazin-4-yl)propyl-1-phosphonic acid (CPP) (all from Sigma) were prepared in water. Tetrodotoxin (TTX), from Alamone Labs, and Neuropeptide Y, a kind gift from Dr. Kitlinska (Georgetown University), were also dissolved in water. Etomidate and taurine (Sigma) were dissolved in dimethylsulfoxide (DMSO; <0.0001% final concentration). Drugs were diluted into aCSF prior to experiments, and applied to the slice through the Y-tube method (Murase et al., 1989), modified for optimal solution exchange in brain slices (Hevers and Luddens, 2002). This method allows rapid solution exchange in the immediate area of the target cell, without affecting the entire slice, as does bath application of drug. Additionally, phosphorylation peptides were supplemented into the internal solution in a subset of experiments: catalytic subunit of PKA (PKA), PKA Inhibitor Fragment 6-22 amide (PKI), and Protein Kinase C Fragment 19-36 (PKC-I) (all from 62 Sigma). For evoked IPSC experiments, lidocaine N-ethyl bromide (QX-314; Sigma) was added to the internal solution. VIII. Measurement of GABA currents Internal solutions with isotonic chloride were used to detect GABAergic currents, setting the reversal potential for Cl- to 0 mV and allowing increased detection of inhibitory currents with a holding potential of -60 mV. Currents were filtered at 2 kHz with a low-pass Bessel filter and digitized at 5-10 kHz using a personal computer equipped with Digidata 1322A data acquisition board and both pCLAMP9 and pCLAMP10 software (all from Molecular Devices). Off-line data analysis, curve fitting, and figure preparation were performed with Clampfit 9 and Clampfit 10 software (Molecular Devices). a. Phasic GABAergic Measurements Spontaneous and miniature inhibitory postsynaptic synaptic currents (sIPSCs and mIPSCs) were identified using a semiautomated threshold based minidetection software (MiniAnalysis; Synaptosoft) and were visually confirmed. Event detection threshold was set at 5 times the root mean square (RMS) level of baseline noise. NBQX was not included in IPSC measurements so as not to perturb network activity. AMPA-mediated sEPSCs could easily be identified by the rapid decay kinetics (<2 ms) (Ortinski et al., 2004), and were excluded from the analysis. IPSC averages were based on more than 50 events, and decay kinetics were determined using double exponential curve fittings and reported as weighted time constants (Tw): 63 Tw = η1*A1/(A1 + A2) + η2*A2/(A1 + A2) where ηx is the decay time constant for a particular component of the curve and Ax is the peak amplitude of the corresponding component. mIPSCs were isolated with Y-tube application of TTX (0.5 µM). All detected events were used for event frequency, but superimposing events were eliminated for amplitude, rise time, and decay kinetic analyses. b. GABAergic Tonic Current Measurements Although much of the tonic current data presented in this thesis was analyzed with an all-points histogram that measured the mean holding current 10 seconds before and during BMR application (Ade et al, 2008), recent studies have shown that this method may not detect small changes in tonic current (Glykys & Mody, 2007). Therefore, when indicated, changes in tonic current were also measured by changes in RMS noise before and during BMR application. These analyses were based on an allpoints histogram that was fitted to the Gaussian function: (x) = A·exp[–(x – µ)2/2 2] where ζ represents the RMS noise during the 10 second period before and during drug application and µ represents the mean holding current. Tonic noise was then measured as a difference in RMS noise between two conditions. When peptides were supplemented in the internal solution, phasic and tonic currents were analyzed at least 4 minutes after break-in to allow proper peptide function and equilibration with the cellular internal components. Likewise, effects of 64 dopaminergic agents were not analyzed until 5 minutes of application to allow proper downstream functions. c. Evoked Current Measurements Evoked IPSCs (eIPSCs) were isolated with NBQX (5 µM) and CPP (10 µM), and weighted decay kinetics were analyzed as described in Introduction, Section VIIIa with averages of at least 10 eIPSC events. For short term plasticity measurements, postsynaptic plasticity responses to 16 and 33 Hz stimulations were measured with two distinct methods: Each of the responses were measured from the original baseline (baseline plasticity) and from the individual response‟s baseline (global plasticity). Short term plasticity ratios were calculated: Plasticity ratio = [amplitude of IPSC7/amplitude of IPSC1] to demonstrate the degree of depression or facilitation. A ratio less than 1.0 represents depression, while a ratio greater than 1.0 represents facilitation. BMR was applied at the end of each experiment to verify that all measured currents were mediated through GABAA receptors. IX. HEK 293 Cell Culture and Transfection Human embryonic kidney 293F (HEK 293) cells (American Type Culture Collection; CLF1573) were grown in minimal essential medium, supplemented with 10% fetal bovine serum, 100 units/mL penicillin, and 100 units/mL streptomycin (all from Invitrogen) in a 5% CO2 incubator at 36°C. The colony was maintained by replacing the 65 medium twice a week and spitting the cells once a week. For transfection, growing cells were dispersed with trypsin and seeded at approximately 2 x 105 cells/35 mm dish in 2 mL of culture medium on 12 mm glass coverslips coated with poly-D-lysine. After 24 hours from plating, the cells were transfected with rat GABAA receptor subunit cDNAs, because of their high homology to mouse receptors, and EGFP using calcium phosphate precipitation. The following GABAA receptor subunit plasmid combinations were used: α2β1γ2, α2β3γ2, α5β1γ2, and α5β3γ2 at a ratio of 1:1:4. Mixed plasmids (5 µg total) were added to the dish containing 2 mL culture medium for 8–12 hours at which point the media was refreshed. The cells were used for electrophysiological recording 2-3 days after transfection. EGFP expression in these HEK 293 cells was used as a marker for successful transfection, although EGFP uptake does not guarantee receptor subunit expression. This method was often successful for about 70% transfection of total HEK 293 cells plated. GABAA receptor dose-response curves were measured with four GABA subsaturating concentrations which were normalized to a saturating GABA concentration (3 mM) in each cell. Dose-response curves were fitted using GraphPad Prism (La Jolla, CA) software with the logistic function: Ipeak = Imin + (Imax – Imin/1 + ([Drug]slope/EC50)) where Imin and Imax are minimal and maximal evoked currents, [Drug] is the GABA concentration, and slope is slope of the curve. 66 X. Immunofluorescence and Confocal Imaging Brain sections were prepared from BAC Drd2 EGFP and BAC Drd2 EGFP;BAC Drd1a tdTomato mice aged ~3 weeks postnatally. Whole brains were dissected and stored in 4% PFA overnight at 4°C. 100 μm coronal or sagittal sections were cut with a Lancer Vibrotome (Series 1000, Sherwood Medical, St. Louis, MO). Fixed, free floating tissue sections were blocked with 4% normal donkey serum in PBS for 1 hour at room temperature, then washed 3 x 10 minutes in PBS/0.1% Triton-X100 (Tx). The rabbit αGABAA receptor β3 1° antibody (Millipore, Temecula, CA; AB5563; 1:1000) was diluted in PBS/Tx/1% bovine serum albumin (BSA). The 1° antibody incubation step was 12-18 hours in duration at 4°C, and slices were then washed 3 x 10 minutes in PBS/Tx. Indocarbocyanine (Cy3)-conjugated 2° antibodies (Jackson ImmunoResearch, West Grove, PA) were diluted 1:1000 in PBS/Tx/BSA and exposed to tissue for 2-4 hours at room temperature. The 2° antibody was washed 3 x 10 minutes with PBS/Tx and sections were placed on microscope slides and sealed under glass cover slips with VectaShield H1000 mounting media (Vector Labs Inc., Burlingame, CA). Images were captured using an Olympus Fluoview-FV300 laser scanning confocal microscope (Olympus Optical, Tokyo, Japan) or the Nikon Eclipse E600 microscope (Nikon Instruments, Melville, NY), exciting endogenous EGFP and Alexa555 fluorophores. For confocal imaging, Plan-Acromat objectives with sequential acquisition of separate color channels were used. Stacks of consecutive sections (Z-stack images; 1 μm thick) were processed with MetaMorph software (Universal Imaging, Downingtown, PA). When noted, confocal images represent a projection image of 14-16 z-stacks. 67 XI. Statistics. Box and whisker plots were generated for more complete representation of certain data. The whiskers include the minimum and maximum values, while the box outlines the 25th and 75th percentile of data points. The mean value is represented by a bar inside the box. Scatterplots of data points are also included in these plots to further show data spread. Statistical significance was determined using the paired two-tailed Student's t test to compare predrug conditions with recordings made under drug conditions of the same cell population. Unpaired two-tailed Student‟s t test was used for comparisons across cell groups. Significance criteria was set at p < 0.05, and all values in text and figures are expressed as mean ± SEM. n represents the number of cells/animals. In text and figures, *p < 0.05, **p < 0.005, and ***p < 0.0005. 68 RESULTS I. GABAergic Tonic Conductances in Striatal MSNs a. Identification of MSNs in Acute Slice Preparation Transgenic mice where EGFP was driven under the promoter for the D2 receptor and tdTomato fluorescent protein was driven under the D1 receptor promoter (BAC Drd2 EGFP;BAC Drd1a tdTomato mice) were used when available to accurately and efficiently compare tonic currents between D2+ and D1+ MSNs. In these acute slices, there was very little overlap of fluorescent proteins (< 1%), suggesting the two primary striatally expressed dopamine receptors are largely segregated onto different striatal neurons. Additionally, images taken from sagittal sections of this mouse brain showed distinct, non-overlapping pathways to the GP (D2+ neurons) and SNr (D1+ neurons), confirming distinct striatopallidal and striatopallidal pathways (Figure 9A). While these results agree with those found recently in the same strain of mice (Shuen et al, 2008), previous reports found that D1Rs and D2Rs colocalize within the same neuron (Surmeier et al, 1993; Surmeier et al, 1996; Aizman et al, 2000). Therefore, under acute slice preparations, D1+ and D2+ MSNs were identified by their unique red and green fluorescence, respectively. No recordings were made from the few cells that expressed both fluorescent proteins because their dopamine receptor identity remained ambiguous. Because the majority of experiments were conducted using CsCl internal solutions, neuronal firing patterns could not be used to confirm MSN identity. Therefore, in many cases, fluorescent proteins were the only identifying marker used to identify MSNs in the slice preparation. In a subset of experiments, to be described later, 69 Figure 9. Striatopallidal and Striatonigral Pathways are Distinct. (A) Sagittal slice taken from a BAC Drd2 EGFP;BAC Drd1a tdTomato mouse, showing distinct, nonoverlapping striatonigral (red) and striatopallidal pathways (green). (B) Confocal image of BAC Drd2 EGFP;BAC Drd1a tdTomato striatal slice (75 µm) with DIC and EGFP (i) and Tomato red (ii) fluorescence. Merged image (iii) of EGFP and Tomato red shows little overlap. Dotted box denotes magnified section shown in (iv). The yellow cell is the only doubly labeled cell in view. Calibration bar: 150 µm. 70 A B A i B ii C iii D iv 71 Kgluconate internal solutions were used to determine MSN firing patterns. In these experiments, MSN identity was confirmed by both fluorescence and firing pattern. Additionally, slices from β3 subunit conditional KO mice lacked fluorescently labeled cells. CsCl internal solutions further complicated accurate MSN identification since firing patterns could not be recorded. In these cases, MSNs were visually identified by soma size, and therefore, these data sets may contain a few GABAergic interneurons. However, the large sample population size coupled with the high percentage of MSNs in striatal slices is likely to offset potential inaccuracies. b. Unique GABAA Receptor Subunits Mediate MSN Tonic Current. A previous study in our lab (Ade et al, 2008), showed that D2+ MSNs have significantly larger tonic current than D1+ MSNs in young mice (p16-25). I have confirmed these results in BAC Drd2 EGFP;BAC Drd1a tdTomato mice and see distinct differences in D2+ and D1+ tonic conductances (Figure 10) in the absence of glutamate blockers (which may affect neuronal excitability and alter tonic current) and any uptake blockers that are often used to increase tonic current visualization (Santhakumar et al, 2010). Ade et al. (2008) hypothesized that differences in tonic current were due to different GABAA receptor composition between the two types of MSNs. Indeed, pharmacological studies showed that the α5 GABAA receptor subunit mediated some tonic current in D2+ MSNs, but none in D1+ MSNs (Ade et al, 2008). A δ subunit KO mouse was used to determine the role of this subunit in young MSN tonic current, as it is implemented in tonic current elsewhere in the rodent brain (Nusser and Mody 2002; Stell et al, 2003; Cope et al, 2005; Drasbek & Jensen 2005; Jia 72 Figure 10. Tonic Current Expression in BAC Drd2 EGFP;BAC Drd1a tdTomato Mice. (A) Representative current traces displaying tonic current in striatal D2+ and D1+ MSNs from BAC Drd2 EGFP;BAC Drd1a tdTomato mice. Right, all-points histogram and Gaussian fit from each segment. (B) Summary graph for the tonic current expressed in D2+ (n = 10/5) and D1+ (n = 15/5) MSNs from BAC Drd2 EGFP;BAC Drd1a tdTomato mice. 73 A B BMR Tonic Current (pA) 30 D2+ BMR D1+ D2+ 20 10 *** 0 30 pA 5s 74 D1+ Figure 11. δ Subunits do not Mediate MSN Tonic Current. (A) Representative current traces from two MSNs in δ subunit KO mice. (B) Summary graph of the tonic current expression pattern from δ subunit KO mice (n = 8/1), which resembles the expression pattern seen in BAC D2 EGFP mice (D2+: n = 8/2; D1+: 8/2). 75 BMR B Tonic Current (pA) A 20 pA 15 s BMR 76 30 D1+ D2+ 20 10 0 δ -/ -/- BAC Drd2 EGFP et al, 2005; Mangan et al, 2005; Mtchedlishvili & Kapur 2006; Glykys et al, 2007). These animals did not express fluorescent proteins, and D1+ and D2+ MSNs could not be identified, but the pattern of tonic current in the δ subunit KO mouse resembled the pattern in BAC Drd2 EGFP mice, suggesting that the δ subunit does not contribute to tonic current in young striatal MSNs (Figure 11). In addition to the α5 subunit, α4 GABAA receptor subunits are also known to mediate tonic inhibition (Jia et al, 2005; Mangan et al, 2005), and are expressed in the striatum (Pirker et al, 2000). Therefore, pharmacological sensitivity to the α4 GABAA receptor subunit agonist taurine (Jia et al, 2008) was investigated for striatal MSNs, and compared between the two MSN subtypes to determine if the α4 subunit contributes to the differential tonic current pattern expressed by D1+ and D2+ MSNs. Although this drug selectively activates glycine receptors at low concentrations, taurine activates α4containing GABAA receptors at much higher concentrations (Jia et al, 2008). Therefore, to investigate taurine‟s effects on MSNs, a high concentration was used (50 µM), which was also likely to activate glycine receptors. Yet, in 3 D2+ MSNs, taurine (50 μM) produced a substantial amount of inward tonic current (24.5 ± 13.4, n = 3/1). The effect of this α4 subunit selective agonist was smaller in D1+ MSNs (7.2 ± 2.5 pA, n = 3/1, p = 0.2), although this difference was not significant, likely due to the small number of cells in each group. Follow up experiments are planned to further investigate the potential differences in α4 subunit expression in MSNs and the subunit‟s importance in mediating tonic inhibition. 77 Figure 12. β3 Subunit Expression in Striatal MSNs. (A) Fluorescent image of endogenous EGFP in striatal BAC Drd2 EGFP slice. (B) Fluorescent image of β3-subunit antibody staining. (C) Merged image of panels A, B. (D) Magnification of boxed area in (C), showing β3-antibody staining on both green (D2+ MSN) and non-green (D1+ MSN) cells. Images shown in A,B,C show the same field of view. Calibration bar: 20 µm for A,B,C. 78 A B C D 79 i. Importance of β Subunit Expression in MSNs. The next step in identifying the receptor composition required for MSN tonic current was to investigate the β subunit composition in these cells. Immunostaining in adult striatum suggested that all β subunits are expressed (Pirker et al, 2000), however a single-cell PCR study showed that D1+ MSNs have β1 and β3 mRNA, while β2 mRNA was absent from MSNs (Flores-Hernandez et al, 2000). However, this study did not investigate mRNA levels for D2+ MSNs, but it remains likely that D2+ MSNs also lack the β2 subunit as the subunit is proposed to be expressed solely in striatal interneurons (Schwarzer et al, 2001). To confirm the presence of β3 subunit expression in MSNs, I performed immunohistochemistry on BAC Drd2 EGFP slices and stained for the β3 subunit. As shown in Figure 12, both D1+ and D2+ MSNs express this subunit, and although fluorescence intensity analysis has not been conducted, the subunit appears to be more highly expressed on D2+ MSNs. Although β subunit expression is not often investigated in studies of tonic current, one study from Dr. Lambert‟s group found that a component of dentate gyrus tonic current is mediated through the β2 subunit, while β1 and β3 GABAA receptor subunits are located in synaptic areas (Herd et al, 2008). Therefore, the contributions of β1 and β3 subunits to striatal MSN tonic current were investigated subsequently. To identify differences in β1 and β3 function, I used etomidate, a general anesthetic with selective agonist properties at β2/β3 subunits of the GABAA receptor (Belelli et al, 1996; Herd et al, 2008). Since MSNs are presumed to lack the β2 subunit, this drug should be specific for β3 subunits in striatal MSNs. 80 Another anesthetic, loreclezole, has also been shown to be highly selective for β2/β3 over β1 GABAA receptor subunits (Wafford et al, 1994). However, etomidate offers a potential experimental benefit over loreclezole. Etomidate‟s ability to potentiate GABAA mediated currents is greater compared to the actions of loreclezole (Wafford et al, 1994), offering greater disparity between β3 and β1 subunits in my experimental setup. However, loreclezole has been shown to inhibit α1β1γ2 single channel units when applied with high doses of GABA (Fisher et al, 2000), offering a potential means to pharmacologically identify the presence of the β1 subunit. ii. Etomidate is Selective for β3 Subunits in Recombinant Receptors It remains necessary, however, to investigate the selectivity of this compound in recombinant receptors as interactions with other subunits may alter etomidate‟s selectivity. The general anesthetic‟s selective properties to β2/β3 GABAA receptor subunits were maintained in recombinant receptors that contained the α1-, α2-, α3-, or α6 subunits, with γ2 (Slany et al, 1995; Hill-Venning et al, 1997; Sanna et al, 1997). Because previous studies showed the importance of α5 subunits in MSN tonic currents (Ade et al, 2008), etomidate‟s pharmacological selectivity needed to be tested in recombinant receptors expressing α5 and γ2 subunits. For these experiments, HEK 293 cells were transfected with striatally relevant α2β1γ2, α2β3γ2, α5β1γ2, and α5β3γ2 receptors. Etomidate has both direct effects and potentiating effects (when combined with GABA) on GABAA receptors, and therefore, both effects were investigated in cells expressing these recombinant GABAA receptors. Each transfected cell was subjected to a 81 Figure 13. GABA Dose Response Curves for Recombinant GABAA Receptors. Cells were subjected to 4 submaximal doses of GABA, and responses were compared to 3 mM GABA. α5β1γ2 receptors (n = 12, black solid line) have increased GABA sensitivity compared to α2β1γ2 receptors (n = 5, grey solid line). The α2β3γ2 receptors (n = 5, grey dashed line) showed enhanced GABA sensitivity to the α2β1γ2 receptors (n = 5, grey solid line) at 3 µM and 10 µM GABA. 82 % Current 100 80 * 60 α5β3γ2 α5β1γ2 α2β3γ2 α2β1γ2 * 40 20 0 -7 -6 -5 -4 log [GABA] 83 -3 -2 series of GABA concentrations (250 nM, 1 µM, 3 µM, 10 µM, and 3 mM saturating dose) to effectively determine the EC10 with which to compare etomidate‟s direct effects with those of GABA potentiation. α5β1γ2 receptors had increased GABA sensitivity compared to α2β1γ2 receptors, suggestive of their role in tonic current (p < 0.05). Interestingly, in α2-containing receptors, the β3 subunit dictated enhanced GABA sensitivity over β1-containing receptors, shifting the dose-response curve to the left (p < 0.05) (Figure 13). A similar shift in GABA sensitivity was not observed in α5-containing receptors. Etomidate (3 µM) selectively activated α2β3- and α5β3-containing receptors (p < 0.05), confirming etomidate‟s selective direct effects on β3-containing receptors. When etomidate and each cell‟s EC10 for GABA were applied together, etomidate selectively potentiated α2β3γ2 currents (α2β1γ2: p = 0.6; α2β3γ2: p < 0.05), while this distinction was not apparent between α5β1γ2 and α5β3γ2 transfected cells (α5β1γ2: p < 0.0005; α5β3γ2: p < 0.05) (Figure 14) and etomidate potentiated GABA currents in both types of α5-containing receptors. It should also be noted that the degree of potentiation did not differ between α5β3- and α5β1-containing receptors (p = 0.3). These data confirm previous reports of etomidate‟s selectivity for direct activation of β3-containing receptors (Hill-Venning et al, 1997), although the drug‟s GABA potentiating role is not selective in α5-containing receptors. As MSNs are known to express the α5 subunit, care will be taken to interpret data involving etomidate‟s effects on MSNs. 84 Figure 14. Etomidate’s Direct Effects are Selective for β3 Subunit-Containing Recombinant Receptors. (A) Representative current traces from HEK 293 cells transfected with α2β1γ2 or α2β3γ2 receptors with the EC10 GABA concentration (3 µM for α2β1γ2 and 1 µM for α2β3γ2) and etomidate (3 µM) application, showing β3-subunit selectivity for direct and potentiating effects. Calibration bars: 200 pA, 10 s. (B) Representative current traces from HEK 293 cells transfected with α5β1γ2 or α5β3γ2 receptors with the EC10 GABA concentration (250 nM) and etomidate (3 µM) application, showing β3-subunit selectivity for direct effects only. Calibration bars: 50 pA, 10 s. (C) Summary of etomidate‟s direct effects on striatally relevant GABAA receptors (α2β1γ2, n = 5; α2β3γ2, n = 10; α5β1γ2, n = 5; α4β3γ2, n = 7). (D) Summary of etomidate‟s potentiating effects of GABA currents evoked by EC10 concentrations of GABA on each receptor type (α2β1γ2, n = 5; α2β3γ2, n = 4; α5β1γ2, n = 6; α6β3γ2, n = 13). 85 β1 A GABA β3 GABA Etom GABA + Etom Etom GABA + Etom α2 B GABA Etom GABA + Etom GABA Etom GABA + Etom α5 β1 β3 * 50 * D 800 % GABA Response 100 Etom Response (pA) C 0 α2 400 0 α5 86 ** α2 α5 iii. MSNs have Different Etomidate Responses Taking these potential restrictions on etomidate selectivity into account, etomidate was applied in the acute striatal slice preparation to determine β3 subunit activity in D1+ and D2+ MSNs. Etomidate (3 µM) was applied to both D1+ and D2+ MSNs, where it produced substantial tonic current in D2+, but not D1+, MSNs (D2+: 42 ± 13.2 pA, n = 12/3; D1+: 9.0 ± 3.0 pA, n = 14/3; p < 0.05). Because local interneurons may express β2/β3 GABAA receptor subunits, etomidate‟s activation of these receptors may increase GABA levels in the acute slice (Ade et al, 2008), these experiments were repeated in TTX (500 nM). D2+ MSN etomidate responses remained significantly greater than those from D1+ cells (p < 0.005, Figure 15), suggesting that D2+ MSNs express more β3 GABAA receptor subunits than D1+ MSNs. Etomidate‟s potentiating role was also assessed as it may give a more accurate account of MSN physiology, because low levels of ambient GABA are always present in the slice and contribute to tonic GABAA currents. Therefore, etomidate‟s potentiating roles were determined by comparing exogenously applied GABA (1 µM) to coapplication of etomidate and GABA, in the presence of TTX. Etomidate potentiated GABA currents 188 ± 68.6% (n = 9/4, p < 0.005) in D2+ MSNs, but only 56.6 ± 29.6% (n = 3/3, p = 0.09) in D1+ MSNs, also suggesting that D2+ MSNs have a greater number of β3-containing GABAA receptors that may mediate tonic current. iv. Etomidate does not Affect mIPSC Properties Because the composition of synaptic GABAA receptors also remains unknown, I investigated etomidate‟s effects on mIPSCs in both types of MSNs. Although etomidate 87 Figure 15. Etomidate Selectively Activates Extrasynaptic Receptors in D2+ MSNs. (A) Representative current traces of a D2+ and a D1+ MSN in 0.5 µM TTX showing greater etomidate-mediated currents in the D2+ MSN. (B) Representative traces of two individual MSNs, showing the direct effect of etomidate (3 µM), response to 1 µM GABA, and etomidate‟s potentiating role with 1 µM GABA, all in 0.5 µM TTX. (C) Summary data of etomidate‟s direct (D2+: n = 21/9; D1+: n = 17/8) and potentiating (D2+: n = 10/4; D1+: n = 6/4) effects on D2+ and D1+ MSN tonic current. 88 A D2+ D1+ Etom BMR Etom BMR 30 pA 40 s B Etom GABA GABA + Etom D2+ 100 pA 20 s D1+ C GABA D2+ ** GABA + Etom Response (pA) 150 100 D1+ D2+ 50 D1+ ** 0 Direct Potentiation 89 Figure 16. Etomidate does not Affect Striatal Synaptic GABAA Receptors. (A) Examples of mIPSCs in D2+ and D1+ neurons before (grey) and after (black) etomidate application. Averaged mIPSC traces are normalized and overlaid (right) to demonstrate that etomidate had little effect on current decay. (B) Summary graph of mIPSC data demonstrating that etomidate had little effect on frequency (D2+: n = 13/4; D1+: n = 11/4), amplitude (D2+: n = 13/4; D1+: n = 11/4), weighted tau (D2+: n = 12/4; D1+: n = 8/4), or rise time (D2+: n = 3/2; D1+: n = 3/2) in both D2+ and D2+ MSNs. 90 mIPSC mIPSC + Etom 91 did not significantly alter mIPSC frequency, amplitude, weighted tau, or rise time (Figure 16), the presence of β3 subunits in synaptic receptors cannot be ruled out. Heterogeneity in β subunit expression at this site may mask any effects of etomidate on β3 subunitcontaining GABAA receptors. Additionally, β1 and β3 subunit expression within the same synaptically-located GABAA receptor is a unique possibility which may underlie these results. c. PKA Phosphorylation Affects MSN Tonic Conductance i. Intracellular PKA Application Modulates D1+ MSN BMRSensitive Tonic Current The large etomidate response in D2+ MSNs suggests that β3 subunits are present in D2+ cells and that this subunit contributes to MSN tonic current. Because both β1 and β3 GABAA receptor subunits are targets of PKA phosphorylation (McDonald et al, 1998), it was crucial to determine the role PKA phosphorylation plays in mediating and regulating MSN tonic current. In order to alter the post-synaptic cell‟s phosphorylation environment, many studies apply cell-permeable drugs to the extracellular space that will activate the PKA phosphorylation cascade in neurons. For example, cell-permeable cAMP or forskolin is often applied exogenously to activate PKA phosphorylation in target neurons (McDonald et al, 1998; Flores-Hernandez et al, 2000). However, these drugs are permeable to every neuron, and therefore affect not only the target postsynaptic neuron, but also pre-synaptic terminals and release probability as well. Therefore, these drugs add another layer of complexity in understanding a post-synaptic neuron‟s response to changes in PKA phosphorylation. To investigate an individual 92 Figure 17. Internal PKA Application Regulates GABA-Mediated Tonic Current. (A) Representative traces of two individual MSNs where PKA was supplemented in the internal solution, showing BMR-sensitive tonic current in both cells. (B) Summary graph for tonic current in D2+ and D1+ MSNs in control conditions (D2+: n = 16/10; D1+: n = 12/9) and with internal PKA application (D2+: n = 10/3; D1+: n = 15/7). (C) Representative traces from simultaneous dual recording of D2+ and D1+ MSNs, showing etomidate responses with internal PKA application. (D) Summary graph for current induced by etomidate with internal PKA application. D2+ MSN responses decreased slightly (CsCl: n = 21/9, CsCl + PKA: n = 5/2), while etomidate responses in D1+ MSNs were significantly greater with internal PKA application (CsCl: n = 17/8, CsCl + PKA: n = 9/4). 93 C Tonic Current (pA) B CsCl 40 D 40 D2+ 30 20 PKA * 30 D1+ 20 ** 10 10 0 0 94 D2+ D1+ * neuron‟s response to phosphorylation, kinases and phosphatases were included in the internal recording solution in this study. The catalytic subunit of PKA (50 – 75 μg/mL; used for only 3 days following dilution) was supplemented into the CsCl internal solution for ideal phosphorylation of PKA substrates. The first step in identifying the effects of phosphorylation on striatal MSNs was to monitor tonic current in both D1+ and D2+ MSNs upon intracellular application of PKA. In these conditions, tonic current in D1+ MSNs increased (p < 0.005), while D2+ MSN tonic current decreased (p < 0.05) (Figure 17). Since β3 subunit phosphorylation results in larger currents (McDonald et al, 1998; Nusser et al, 1999), the increase in D1+ MSN tonic current could reasonably be attributed to a phosphorylated β3 GABAA receptor subunit. Etomidate‟s effects were also measured in the presence of TTX and intracellular application of PKA. In these conditions, the D1+ MSN etomidate response increased (p < 0.05), while the D2+ MSN response was slightly decreased (p = 0.2) (Figure 17C&D). Interestingly, etomidate responses between D1+ and D2+ MSNs did not differ with intracellular PKA application (p = 0.5), indicating that the two MSN subtypes contain similar amounts of extrasynaptic β3 subunit-containing GABAA receptors which may mediate tonic current when phosphorylated. These data suggest that D1+ MSN tonic current is mediated through a phosphorylated β3 subunit. 95 ii. Basal Phosphorylation of β3 Subunits in D2+ MSNs Because intracellular PKA application did not increase D2+ MSN tonic current, the next obvious question to investigate was if D2+ MSN β3 subunits were basally phosphorylated to mediate tonic current. Indeed, many studies have also suggested that β3 GABAA receptor subunits are basally phosphorylated by PKA (McDonald et al, 1998; Poisbeau et al, 1999; Brandon et al 2003). If D2+ MSNs mediate their tonic current through basally phosphorylated β3 subunits, PKA blockade should eliminate tonic current. Therefore, I investigated the PKA phosphorylation blockade next by including the PKA inhibitor, PKI (20 μM), in the internal recording solution and measuring MSN tonic current. In these conditions, D2+ MSN tonic current decreased (p < 0.0005), while D1+ tonic current remained unaffected (p = 0.3), suggesting that D2+ MSNs are basally phosphorylated by PKA (Figure 18B). Because D1+ MSN tonic current was unaltered with internal PKI application, but increased with internal PKA application, β3 subunits in these cells are unlikely to be basally phosphorylated. Basal phosphorylation has also been attributed to PKC activity in cortical neurons (Brandon et al, 2000), so I also supplemented the specific PKC inhibitor, PKC fragment 19-36 (PKC-I, 10 µM), into the internal solution to determine its effects on D2+ MSN tonic current. In these conditions, D2+ MSNs expressed just 8.0 ± 1.13 pA (p < 0.0005) of tonic current, while D1+ MSN tonic current was largely unaffected (p = 0.09) (Figure 18B). Therefore, it appears that both PKA and PKC may be responsible for basal phosphorylation of D2+ MSNs, although it is likely that the pharmacological inhibitors are not selective enough for assigning the role of basal phosphorylation to PKA or PKC exclusively. 96 Figure 18. D2+ MSNs are Basally Phosphorylated by PKA and PKC. (A) Representative traces of normal D2+ tonic current (left) and D2+ tonic current with intracellular PKI application (right). (B) Summary graph for D2+ and D1+ MSN tonic current in normal conditions (D2+: n = 16/10; D1+: n = 15/7), with intracellular application of PKI (D2+: n = 15/8; D1+: n = 4/3), and intracellular application of PKC-I (D2+: n = 12/2; D1+: n = 4/2). 97 A D2+ MSN CsCl BMR D2+ MSN CsCl+PKI BMR 30 pA 5s Tonic Current (pA) B 30 CsCl CsCl+PKI CsCl+PKC-I 20 *** 10 *** 0 D2+ D1+ 98 Figure 19. Synaptic Receptors are Unaffected by PKA and PKI. (A) Summary graph showing no changes in sIPSC frequency in control conditions (D2+: n = 31/14; D1+: n = 26/11), internal PKA application (D2+: n = 16/5; D1+: n = 21/5), and internal PKI application (D2+: n = 16/5; D1+: n = 7/5). (B) Summary graph showing no changes in sIPSC amplitude in control conditions (D2+: n = 31/14; D1+: n = 26/11), internal PKA application (D2+: n = 16/5; D1+: n = 20/5), and internal PKI application (D2+: n = 16/5; D1+: n = 7/5). (C) Summary graph of changes in weighted tau among control conditions (D2+: n = 31/14; D1+: n = 26/11), internal PKA application (D2+: n = 14/5; D1+: n = 21/5), and internal PKI application (D2+: n = 15/5; D1+: n = 7/5). Internal application of PKI significantly decreased D2+ MSN decay time from control. 99 Frequency (Hz) A 3 2 1 Amplitude (pA) 0 B CsCl CsCl+PKA CsCl+PKI 40 D2+ D1+ D2+ D1+ 30 20 10 0 30 Tau (ms) C 20 ** 10 0 D2+ D1+ 100 However, due to striatal dopamine‟s importance in regulating the PKA second messenger cascade (Stoof & Kebabian, 1981), I have largely focused on PKA‟s influence on MSN tonic current. Future studies are warranted to further investigate the potential role for PKC in this system. iii. sIPSC Properties are Largely Unaffected by PKA and PKI To better understand the role of PKA phosphorylation in striatal MSNs, sIPSC properties were also measured with intracellular PKA and PKI application and compared to those properties measured in control conditions. Intracellular application of PKA did not affect sIPSC frequency, peak amplitude, or decay time in MSNs as shown in Figure 19. Because PKA phosphorylation modulates β1 and β3 subunits differently, these results are somewhat expected since synaptic receptor β subunit composition is likely heterogeneous, and presence of both subunits will likely mask the effects of the other. Additionally, intracellular application of PKI did not affect sIPSC properties in MSNs, although it decreased D2+ MSN decay time (p < 0.005, Figure 19). The decrease in decay kinetics may also suggest that β3 subunits in D2+ MSN synaptic receptors are basally phosphorylated, as dephosphorylation will decrease the amount of current through the channel. 101 d. Dopamine’s Regulation of MSN Tonic Current. i. Little Basal Dopamine Present in Acute Slice Preparation. Given dopamine‟s extensive control over PKA phosphorylation and PKA phosphorylation‟s role in MSN tonic current, I next sought to reconcile these two observations and determine how dopamine may modulate striatal MSN tonic current. Striatal dopamine will promote PKA activity in D1+ MSNs, while inhibiting it in D2+ MSNs, and ambient dopamine affects the system in drastically different ways. Although basal dopamine levels are not usually measured with real-time voltammetry studies, one study investigated dopamine release in the nucleus accumbens of rat acute slices, and found that the neuromodulator is only released when electrical or pharmacological stimulus is provided (Lee et al, 2001). Although conducted in the nucleus accumbens where dopamine concentrations are lower than the dorsal striatum, these studies suggest that dopamine is not present in the acute slice preparation, and dopamine receptors are unaffected. Therefore, to investigate the basal levels of dopamine in the dorsal striatum of the acute slice preparation, I applied D1R and D2R selective antagonists to MSNs and measured tonic currents. In 4 D2+ MSN whole-cell recordings, the D2R antagonist sulpiride (2 µM) did not affect tonic currents (19.5 ± 2.1 pA, n = 4/3, p = 0.2). Likewise, SCH23390 (10 µM), the D1R antagonist, had no effect on D1+ MSN tonic current (1.8 ± 1.0 pA, n = 4/2, p = 0.2), suggesting that basal levels of dopamine do not affect D1 and D2 receptors in young animals. 102 ii. Dopamine Receptor Agonists Alter MSN Tonic Current. In order to determine if dopamine affects the PKA phosphorylation cascade in striatal MSNs, dopamine‟s effects on MSN tonic current were assessed by applying D1Rand D2R-like agonists to the slice, and measuring tonic current in D1+ and D2+ MSNs, respectively. The endogenous compound dopamine was not applied because it will bind and activate both D1R and D2Rs, and may alter presynaptic functions, muddling the effects of dopamine on postsynaptic MSNs. Quinpirole (10 µM), the D2R-like agonist, was applied to D2+ MSNs, and BMR (25 µM) was applied following at least 5 minutes of whole-cell recording (to allow full GPCR effects). In these conditions, quinpirole decreased the D2+ MSN tonic current (p < 0.05, Figure 20), while the D1R-like agonist (SKF-81297, 10 µM), increased D1+ MSN tonic current (p < 0.05). To be confident that these effects were due to activation of their dopamine receptors, I applied SKF-81297 and quinpirole to D2+ and D1+ MSNs, respectively. In these conditions, the D1R-like agonist did not alter the D2+ MSN tonic current (p = 0.3) nor did the D2R-like agonist alter the D1+ MSN tonic current (p = 0.9). Therefore, D2R activation decreases tonic current, which is in line with a GPCR-mediated inhibition of PKA phosphorylation which I found to decrease D2+ MSN tonic current. On the other hand, D1R activation increases tonic current; this effect is likely mediated through PKA activation, which I found to increase D1+ MSN tonic current. These same experiments were repeated at more physiological temperatures (32 ºC) since phosphorylation rates are likely altered under such conditions. In these conditions, D2+ MSN tonic currents remained greater than those expressed by D1+ MSNs (D2+: 21.7 ± 11.4 pA, n = 3/2; D1+: 7.0 ± 2.5 pA, n = 3/2, p = 0.2). At 103 Figure 20. Dopamine Agonists Alter MSN Tonic Conductance. (A) Representative current traces from individual D2+ and D1+ MSNs illustrating that the D2R agonist, quinpirole, reduces tonic current in the D2+ MSN, whereas it does not affect tonic current in the D1+ MSN. (B) Representative traces of a simultaneous dual recording between a D2+ and D1+ MSN illustrating that the D1R agonist, SKF-81297, induces a tonic current in the D1+ MSN, while not affecting tonic current in the D2+ MSN. (C) Summary graph showing effects of tonic current with quinpirole and SKF-81297 application on D2+ (n = 5/3, 7/3) and D1+ neurons (n = 5/3, 3/3). 104 % Control Tonic Current C 300 D2+ D1+ * Quinpirole 200 SKF- 81297 100 * 0 105 physiological temperatures, quinpirole continued to decrease the D2+ MSN tonic current to 38.9 ± 18.3 % of control (n = 3/2; p = 0.1), while SKF-81297 continued to increase the D1+ tonic current to 183.6 ± 33.9 % of control (n = 3/2, p = 0.08). Although these findings were not significant, likely due to the small numbers of cells where control tonic current could be compared to the effects of agonists within the same cell, the effects seen at room temperature are consistent with the trends seen at physiological temperatures. These experiments suggest that the dopamine-PKA phosphorylation-tonic current relationships remain intact at more physiological temperatures. iii. The Link Between Dopamine Receptor Activation, Phosphorylation, and GABA-Mediated Tonic Current To confirm that the increased D1+ MSN tonic current with SKF-81297 application was due to a phosphorylated β3 subunit, etomidate was applied alone and after the cell was bathed in SKF-81297 for over 5 minutes. When co-applied with the D1R-like agonist, etomidate produced a significantly greater response than when it was applied alone (etomidate, 8.3 ± 2.2 pA; SKF-81297 + etomidate, 16.0 ± 4.2 pA, n = 4/1, p < 0.05; Figure 21). These data support the hypothesis that D1+ MSNs express tonic current through phosphorylated β3 subunit-containing GABAA receptors upon dopamine receptor activation. iv. Tonic Current in Adult MSNs. While such experiments in young mice (p 15-21) are necessary and important to the understanding of neuronal physiology, experiments in adult animals offer additional 106 Figure 21. SKF-81297 Increases Etomidate Response in D1+ MSNs. Representative current trace from a D1+ MSN where etomidate was given before and during SKF-81297 application. SKF-81297 was given for over 5 minutes before co-application with etomidate. In a total of 4 cells, SKF-81297 + etomidate currents were significantly larger than etomidate currents alone from the same cells (p < 0.05). 107 Etom SKF - 81297 20 pA 20 ms 108 Etom insight into potential disruptions that occur in primarily adult-onset diseases, like PD. For this reason, I recorded MSN tonic current from adult mice (p 33-37) and investigated how tonic current in these cells was modulated by dopamine receptor agonists and regulators of PKA phosphorylation. Strikingly, older D1+ MSNs expressed significantly more tonic current than young neurons (18.3 ± 1.19 pA, n = 6/2, p < 0.0005), while tonic current was decreased in D2+ MSNs (8.25 ± 3.1 pA, n = 4/2, p < 0.05) (Figure 22). These findings were confirmed by a recent study in adult mice by Santhakumar et al. (2010) who found that adult (p > 30) D1+ MSNs express more tonic current than D2+ MSNs, the opposite pattern from young (p 16-25) MSNs. Basal dopamine levels are unable to activate receptors in young mice, but increased dopamine release in the adult striatum may provide a reasonable mechanism for the developmental changes in MSN tonic current expression. Application of the D1Rlike antagonist decreased the adult D1+ MSN tonic current to 35.1 ± 14.9 % of control (n = 4/1, p = 0.05), suggesting that basal levels of dopamine may be altered in the adult animal, and underlie the observed changes in MSN tonic current. To investigate the role of PKA phosphorylation in these older animals, PKI was supplemented in the internal solution and tonic currents from D1+ and D2+ MSNs were measured. In older animals with intracellular PKI application, tonic current in D2+ MSNs did not change (p = 0.9), while D1+ MSN tonic current was significantly reduced (p < 0.0005) (Figure 22). Although the dopamine receptor profile for tonic current is reversed in adult mice (likely due to altered dopamine release), the relationship between dopamine, PKA phosphorylation, and tonic current remains intact throughout development. 109 Figure 22. Adult Animals have Different Tonic Current Expression. Summary graph showing the amount of tonic current expressed in young MSNs (p 15-21) (D2+: n = 16/10; D1+: n = 12/9), old MSNs (p 33-37) (D2+: n = 4/2; D1+: n = 6/2), and old MSNs with intracellular PKI application (D2+: n = 4/2; D1+: n = 4/2). * denotes significance between young and old mice. # denotes significance between control and intracellular PKI application in old mice. 110 Tonic Current (pA) 40 30 D2+ D1+ *** 20 * ## 10 0 111 v. Dopamine Regulates MSN Excitability Because dopamine modulates MSN tonic currents and GABAergic tonic current regulates cell excitability (Ade et al, 2008), dopamine‟s effects on MSN rheobase and firing frequency were also investigated. For these experiments, the Kgluconate internal solution was used with the current-clamp technique. Cells were held at a pre-determined membrane potential (-70 mV), and subjected to 1 s hyperpolarizing and depolarizing current injections before and during dopamine receptor agonist application. As previously reported, D2+ MSNs exhibited a lower rheobase compared to D1+ MSNs (Figure 23C), suggesting that D2+ MSNs are more excitable (Ade et al, 2008; Gertler et al, 2008). Quinpirole (10 µM) significantly increased the rheobase and significantly decreased the firing frequency in D2+ MSNs (Figure 23). However, no changes to rheobase or firing frequency were observed with the D1R agonist, SKF-81297 (10 µM). These data suggest that activation of D2Rs reduces D2+ MSN excitability, feasibly through dopamine regulation of the D2+ MSN tonic current. Further investigation is necessary to make a definitive statement on how D1Rs regulate D1+ MSN excitability. vi. Dopamine Receptor Agonists do not Affect sIPSC Properties In order to determine the effects of dopamine receptor agonists on D2+ and D1+ MSN synaptic receptors, I measured sIPSC properties in these neurons after they were exposed to quinpirole or SKF-81297. No changes to decay kinetics were observed in these conditions (D2+: 124.6 ± 17.7 % control, n = 4/3, p = 0.09; D1+: 99.5 ± 11.3 % control, n = 10/7, p = 0.8), in agreement with a study from Robert Malenka‟s lab (Nicola & Malenka, 1998) where dopamine failed to affect IPSCs in the dorsal striatum. 112 Figure 23. Dopamine Receptor Agonists Modulate MSN Excitability. (A) Representative current-clamp recording from a D2+ MSN illustrating the responses to a series of depolarizing current injections (20 pA steps) from -70 mV, recorded with Kgluconate internal in the absence and presence of quinpirole and BMR. (B) Representative example of a D1+ MSN in the same conditions as (A), but with the D1Rlike selective agonist, SKF-81297. (C) Summary plot showing the averaged rheobase current in D2+ (n = 5/2) and D1+ (n = 8/3) MSNs with dopamine receptor agonist and BMR application. Rheobase measured from -70 mV. (D) Summary of action potential firing frequency in response to increasing depolarizing current injections in D2+ MSNs (black square) in the absence and presence of quinpirole (white square) and BMR (black circle) (n = 5/2). (E) Summary of action potential firing frequency in D1+ MSNs (black square) in the absence and presence of SKF-81297 (white square) and BMR (black circle) (n = 7/3). * denotes significance to D2+ control cells. # denotes significance between D2+ and D1+ MSNs. Calibration bar: 20 µM and 500 ms. 113 D2+ A D1+ B Control Control 40 pA Quinpirole 60 pA SKF-81297 80 pA 60 pA BMR BMR 60 pA 80 pA Rheobase (pA) C80 ***** 0 Quinpirole 0 Control Quinpirole BMR 100 Frequency (Hz) E ** ** * 0 # BMR 20 10 Control SKF D2+ 30 D1+ 40 D Frequency (Hz) D2+ D1+ 30 20 Control SKF-81297 BMR 10 0 200 0 I (pA) 100 I (pA) 114 200 e. The Importance of the GABAA Receptor β3 Subunit in MSN Tonic Current Because global β3 subunit KO mice are prone to developmental problems resulting in neonatal death (Homanics et al, 1997; Ferguson et al, 2007), conditional β3 subunit KO mice were generated to identify the importance of this subunit in mediating striatal MSN tonic conductance. Two strains of conditional KO animals were generated using the Cre-lox system, which as previously described, excises a gene of interest from a particular neuronal tissue or cell type. Drd2-β3 subunit KO mice have Cre-mediated excision of the β3 GABAA receptor subunit only in cells that express the D2 receptor; cells that lack the D2 receptor still contain the GABAA receptor subunit. Dlx5/6-β3 subunit KO animals have Cre-mediated excision of the β3 subunit in cells that express Dlx5/6, which has been characterized in neurons that originate from the lateral ganglionic eminence (Stenman et al, 2003). Thus, the β3 subunit is excised in all GABAergic forebrain neurons, including both types of striatal MSNs and interneurons, in Dlx5/6-β3 subunit KO mice. i. Cre-lox Method Decreases β3 GABAA Receptor Subunit Expression Western blot analysis on adult pan-neuronal β3 subunit KO animals showed a significant Cre recombinase-mediated reduction in the β3 GABAA receptor subunit expression in all areas studied (Ferguson et al, 2007). Although the subunit was not completely deleted, these animals offer unprecedented insight into the importance and function of the β3 subunit. Western blot analysis on the conditional KO mice discussed 115 in this thesis was not conducted due to the complexity of the conditional KO tissue. For example, D2R expressing neurons need to be effectively sorted from non-D2R expressing neurons for a meaningful Western blot analysis. It can be assumed, however, that Cre recombinase expression in a conditional KO excises a similar amount of β3 GABAA receptor subunits as reported in the pan-neuronal KO because these two studies share the floxed β3 mouse strain. ii. Conditional β3 Subunit KO Mice Show Enhanced Lethality While a global deletion of the β3 GABAA receptor subunit resulted in 90% lethality within 24 hours, possibly due to issues involving cleft palate (Homanics et al, 1997), pan-neuronal deletion of the subunit was found to cause neonatal lethality unrelated to cleft palate (Ferguson et al, 2007). A similar finding was observed with the Dlx5/6-β3 subunit KO mice when the number of Cre expressing animals that were heterozygous for the floxβ3 allele (Hetβ3) surviving to p7 was compared to the number of Cre expressing animals with the homozygous floxβ3 allele (KO). The number of Cre recombinase expressing animals that were Hetβ3 was 75%, while Cre recombinase expressing animals with both floxβ3 alleles represented just 25% of animals (n = 8). This ratio of 3:1 differs from the predicted 1:1 ratio with my breeding scheme and Mendelian genetics, and suggests that the conditional Dlx5/6-β3 subunit KO mice may be prone to neonatal lethality before p7. Neonatal lethality was not observed in Drd2-β3 subunit KO animals (n = 10). Despite these neonatal deaths, both Dlx5/6- and Drd2-β3 subunit KOs lacked any abnormal phenotypes like cleft palate and hyperactivity, which are observed in global β3 116 subunit KOs (Homanics et al, 1997), but not pan-neuronal KOs (Ferguson et al, 2007). Under general observation, conditional β3 subunit KOs were not behaviorally different from heterozygous littermates. iii. Conditional β3 Subunit KO Mice have Decreased Tonic Current Although dopamine receptor identity was not able to be determined in these mice, a large number of cells were sampled, and therefore the sample population likely contains both D1+ and D2+ MSNs. Because cell identity was not known, box and whisker plots were generated to observe the spread and pattern of tonic current. Presenting only averages and SEM would be misleading as these data contain two types of cells with differences in the measured characteristics. However, averages and SEM were used to determine significance in these cells for lack of a better way to present statistical data. Tonic inhibition was measured and compared between Drd2-β3 subunit KO and Drd2-Cre;Hetβ3 mice to verify that any differences observed in tonic current were due to the lack of the β3 subunit. As seen in Figure 24B, the presence of the wide box representing the 25th and 75th percentile demonstrates the high degree of variability in tonic current amplitudes (0 to 25.1 pA, n = 14/4) from MSNs in Drd2-Cre;Hetβ3 mice. This wide range is consistent with the inclusion of D1+ and D2+ MSN tonic currents among the recorded neurons, and suggests that any potential changes to tonic current in Drd2-β3 subunit KO mice is due to the lack of β3 subunit expression. Indeed, Drd2-β3 subunit KO mice had tonic current amplitudes that were confined to a much more narrow 117 Figure 24. Tonic Current Patterns in β3 Subunit KO Mice. (A) Individual representative current traces displaying tonic current in Drd2-Cre;Hetβ3 mice (left) and Drd2-β3 subunit KO mice (right). (B) Summary box and whisker plots displaying the tonic current patterns in Drd2-Cre;Hetβ3 (n = 14/4), Drd2-β3 subunit KO (n = 26/4), Dlx5/6-Cre;Hetβ3 (n = 13/3), and Dlx5/6-β3 subunit KO (n = 8/2) mice. Small squares represent each cell‟s tonic current. (C) Summary box and whisker plots displaying the tonic current patterns in Floxβ3 (n = 13/2) and wild type BAC Drd2 EGFP;BAC Drd1a tdTomato mice from D1+ (n = 15/5) and D2+ (n = 10/5) MSNs. Small squares represent each cell‟s tonic current. 118 Cre;Hetβ3 A KO BMR BMR BMR 10 s 60 pA C Tonic Current (pA) 40 Drd2 Dlx5/6 20 40 Tonic Current (pA) B 20 0 0 Het KO Het KO 119 Floxβ3 range (0 to 6.4 pA, n = 26/4), suggesting that the tonic current expression in Drd2-β3 subunit KO mice does not differ between D1+ and D2+ MSNs. In addition, the average tonic current between Drd2-β3 subunit KO and Drd2-Cre;Hetβ3 was significantly different; amplitudes were smaller in Drd2-β3 subunit KO animals (Drd2-Cre;Hetβ3: 11.0 ± 2.3 pA, n = 14/4; Drd2-β3 subunit KO: 2.8 ± 0.4 pA, n = 26/4, p < 0.0005). These results suggest that β3 subunit deletion in D2+ MSNs decreases the D2+ MSN tonic current to levels characteristic of D1+ MSNs. Tonic current was also measured by RMS noise analysis in these cells. Although the difference was not significant, the average RMS noise difference tended to be lower in Drd2-β3 subunit KO animals (1.0 ± 0.15, n = 26/4) than Drd2-Cre;Hetβ3 animals (1.5 ± 0.24, n = 14/4, p = 0.07). Again, this trend suggests that Drd2-β3 subunit KO animals express a lower average tonic current amplitude than the Drd2-Cre;Hetβ3 mice. Experiments conducted in Dlx5/6-β3 subunit KO and Dlx5/6-Cre;Hetβ3 mice gave similar results (Figure 24B). While Dlx5/6-Cre;Hetβ3 mice showed a tonic current pattern suggestive of D1+ and D2+ MSNs (range of 0 to 22.3 pA), the spread of tonic current was more narrow in Dlx5/6-β3 subunit KO animals (0.4 to 12.7 pA). The average tonic current between Dlx5/6-β3 subunit KO and Dlx5/6-Cre;Hetβ3 mice was not significantly different (Dlx5/6-Cre;Hetβ3: 10.9 ± 2.8 pA, n = 13/3; Dlx5/6-β3 subunit KO: 5.8 ± 1.6 pA, n = 8/2, p = 0.2), although may become significant with increased numbers of experimental Dlx5/6-KO animals. These results suggest that β3 subunit deletion in striatal MSNs and interneurons decreases MSN tonic current. It is also possible that the floxβ3 allele alters β3 subunit and functionality, and therefore tonic currents were measured from mice that lacked Cre recombinase 120 expression, but were homozygous for the floxβ3 allele (Floxβ3). Suggesting inclusion of both D1+ and D2+ MSNs in the sample population, Floxβ3 mice showed quite varied tonic current amplitudes (0 to 34.3 pA) (Figure 24C), agreeing with the Western blot analysis (Ferguson et al, 2007) that suggested that the subunit was not affected by the inclusion of loxP sites. iv. Etomidate Sensitivity in Conditional β3 Subunit KO Mice. Based on experiments already presented, etomidate (3 µM) is a selective pharmacological agent that identifies β3 subunit-containing GABAA receptors. Therefore, the effects of etomidate in TTX were tested on Drd2-β3 subunit KO and Drd2Cre;Hetβ3 mice. Responses to etomidate (3 µM) averaged 43.6 ± 9.3 pA (n = 9/2) in Drd2-Cre;Hetβ3 mice, with a wide range of amplitudes (4.3 to 95.4 pA), suggesting that this population of MSNs contains both etomidate-sensitive D2+ MSNs and relatively etomidate-insensitive D1+ MSNs. Interestingly, a similar wide range of etomidate responses was found in the Drd2-β3 subunit KO animals (0.4 to 34.6 pA), while the average was significantly smaller (14.9 ± 2.7 pA, n = 15/4, p < 0.005). These same experiments were repeated in Dlx5/6-KO mice with similar results. Although Dlx5/6-Cre;Hetβ3 mice had etomidate currents averaging 44.3 ± 13.6 pA (n = 6/2) with a wide range of amplitude (6.5 to 95.2 pA), Dlx5/6-β3 subunit KO mice expressed somewhat lower average etomidate responses (33.4 ± 9.2 pA, n = 7/2, p = 0.5), although the range of currents was still quite wide (4.8 to 65.9 pA). Data from both types of conditional β3 KO mice suggest that etomidate responses in β3 KOs are smaller compared to heterozygous littermates. 121 As previously reported, responses to 1 µM GABA in TTX are significantly smaller in D1+ MSNs compared to D2+ neurons (Ade et al, 2008). Therefore GABA sensitivity was also tested in β3 subunit Drd2-β3 subunit KO mice in the presence of TTX. The average response to GABA (1 µM) in the Drd2-Cre;Hetβ3 mice was 61.5 ± 13.1 pA (n = 7/4) with a range of 15.3 to 100.5 pA. In contrast, Drd2-β3 subunit KO mice expressed an average GABA current of 14.0 ± 2.7 pA (n = 6/2, p < 0.05), with a more narrow range of amplitudes (9.1 to 24.3 pA). These data suggest that the β3 subunit may also affect GABA sensitivity in some receptors. v. Synaptic Responses in β3 Subunit KO Mice Although D1+ and D2+ MSNs do not differ in their mIPSC profile (Ade et al, 2008), I sought to more fully understand the composition of phasic GABAA receptors, and mIPSC characteristics were compared between β3 subunit KO and Cre;Hetβ3 mice. As shown in Figure 25C, averaged mIPSC decay times were significantly shorter in Drd2-β3 subunit KO mice compared to Drd2-Cre;Hetβ3 mice. On the other hand, both mIPSC frequency and peak amplitudes did not differ between the two transgenic animal strains. Although there was no significant difference between mIPSC decay times in the Dlx5/6-β3 subunit KO and Dlx5/6;Hetβ3 mice (p = 0.4), β3 subunit KO animals tended to have more rapid decay kinetics. As seen in the Drd2-β3 subunit KO mice, mIPSC frequency and peak amplitudes were no different from their Cre;Hetβ3 littermates (p = 0.9, p = 1.0). 122 Figure 25. Synaptic Responses in Drd2-β3 Subunit KO Mice. (A) Summary graph for mIPSC frequencies in Drd2-β3 subunit KO (n = 17/4) and Drd2-Cre;Hetβ3 (n = 9/4) mice. (B) Summary graph for mIPSC peak amplitudes in Drd2-β3 subunit KO (n = 17/4) and Drd2-Cre;Hetβ3 (n = 9/4) mice. (C) Summary graph for mIPSC decay time in Drd2β3 subunit KO (n = 14/4) and Drd2-Cre;Hetβ3 (n = 9/4) mice. 123 Frequency (Hz) A Drd2-Cre;Hetβ3 Drd2-β3 subunit KO 1 0.5 B Amplitude (pA) 0 30 20 10 0 C Tw (ms) 40 ** 20 0 124 II. Characterizing NPY+ Interneurons. a. Identification of LTS/NPY+ Interneurons in Acute Slice Preparation. To accurately identify LTS/NPY+ interneurons and compare their electrophysiological characteristics with those of MSNs, BAC NPY mice were used that express EGFP under the NPY promoter. Therefore, in acute slice preparations made from these mice, “green” neurons were assumed to be NPY+ interneurons. Because firing pattern was not tested in all “green” NPY+ interneurons, these cells will be labeled as NPY+ interneurons for the purposes of this results section. In these acute slices, NPY+ interneurons were sparse in the striatum compared with cortical areas (Partridge et al, 2009), consistent with the observation that striatal NPY+ interneurons represent just 1-2% of the neuronal population (Kawaguchi et al, 1995). Because these experiments were conducted with a KCl-based internal solution, “green” cells were also confirmed with previously established characteristic firing patterns of LTS and PLTS-type interneurons (Kawaguchi, 1993; Koós & Tepper, 1999). In these slices, MSNs were identified as “non-green” cells with MSN characteristic firing patterns (Ade et al, 2008). To confirm the MSN properties found in these slices, these experimental conditions were repeated in fluorescently labeled animals, where MSN identity was known. b. Intrinsic NPY+ Interneuron Characterization. Striatal NPY+ interneuron electrophysiology had remained largely unexplored since initial work by Kawagushi in 1993, until Dr. John Partridge and I began a study that utilized the BAC NPY strain of mice to identify these cells in the acute slice preparation 125 Figure 26. Intrinsic NPY+ Interneuron Characteristics. (A) Representative cellattach recording of spontaneous firing in an NPY+ interneuron. (B) Superimposed representative current-clamp traces to hyperpolarizing and depolarizing current injections in distinct examples from NPY+ cells (left, center) and an MSN (right). (C) Spikefrequency plots of NPY+ interneurons (n = 8/4) and MSNs (n = 8/3) as a function of depolarizing injection current amplitude. All cells held at -70 mV. (D) Box and whisker plot shows the range of input resistance values in NPY+ interneurons (n = 8/3) and MSNs (n = 6/3). 126 A 60 pA 2s B NPY+ MSN- NPY+ 20 mV 35 30 25 20 15 10 5 0 NPY+ MSN ** 0 50 D ** 100 ** 150 R input (MΩ) C Frequency (Hz) 500 ms 200 1000 800 600 400 * 200 0 Injected Current (pA) NPY+ 127 MSN (Partridge et al, 2009). These cells were previously identified only by firing pattern characteristics and post-hoc immunohistochemistry, and were difficult to study due to their low abundance in rodent striatum. One of the first characteristics we observed was that about 85% of “green” NPY+ cells displayed spontaneous firing (16 of 19/7, Figure 26A), while “non-green” MSNs did not when a Kgluconate internal solution was used. These findings suggest that in the acute slice preparation, NPY+ interneurons are continually releasing GABA, and may be providing the ambient GABA needed to activate tonic current in striatal MSNs. Based on this hypothesis, further characterization was necessary to determine basic properties of these striatal interneurons and their potential role in providing ambient GABA in the acute slice preparation. To identify firing pattern characteristics of NPY+ interneurons and MSNs from BAC NPY mice, I performed current clamp recordings with KCl internal solutions in both types of cells. Each cell was subjected to 1 s hyperpolarizing and depolarizing current injection steps of 10 pA from a set membrane potential of -60 mV for NPY+ interneurons and -70 mV for MSNs based on their reported RMPs. Of 20 NPY+ interneuron recordings, most cells exhibited rebound action potentials in response to hyperpolarizing steps and repetitive firing in response to depolarizing current injections (n = 15/9; Figure 26B, left). The remaining cells also exhibited the rebound action potentials, but featured persistent depolarization in response to depolarizing current injections (n = 5/4; Figure 26B, center). These defining characteristics have been previously noted for LTS and PLTS interneurons (Kawaguchi, 1993; Tepper & Bolam, 128 2005), and the striatal NPY+ interneuron population highlighted in the BAC NPY mice is likely to contain both types of these cells. Further analysis of these firing patterns showed that MSNs had a greater rheobase (55.0 ± 5.3 pA, n = 8/3) than NPY+ interneurons (27.5 ± 8 pA, n = 8/4, p < 0.005) when measured from -70 mV. The relationship between frequency of action potentials and amount of injected current was also measured, and the frequencies were greater in LTS/NPY+ interneurons compared to MSNs from the same animals (Figure 26C). These data suggest that LTS/NPY+ cells fire action potentials with greater frequency within a physiological membrane potential range than MSNs due to their Kv1.2 channel expression (Shen et al, 2007), and the interneurons may also provide a source of ambient GABA to the striatal acute slice preparation. In addition, NPY+ interneurons had a significantly greater input resistance than MSNs (Figure 26D). These findings agree with previously reported characteristics of LTS/PLTS interneurons (Kawaguchi, 1993), and verify the use of BAC NPY mice in the study of striatal LTS/NPY+ interneurons. c. GABAergic signaling in NPY+ interneurons. Although much is known about pre-synaptic inputs onto MSNs, much less is known about pre-synaptic input to NPY+ interneurons. GABAergic IPSC data has not been described for striatal NPY+ interneurons, and therefore, I also investigated the differences in GABAergic signaling between striatal NPY+ interneurons and MSNs. GABAergic synaptic events were recorded with a KCl internal solution, and the cells were held at -60 mV in the voltage-clamp configuration. There were no major differences in whole-cell synaptic activity parameters between NPY+ interneurons and 129 Figure 27. GABA Mediated Synaptic Transmission in NPY+ Interneurons. (A) Representative traces of GABAergic sIPSCs in an NPY+ interneuron (top) and MSN (bottom) with KCl-based internal solution. (B) Summary graph showing no differences between NPY+ interneurons and MSNs in sIPSC (NPY+: n = 9/3; MSN: 7/3) and mIPSC (NPY+: 9/3; MSN: 9/3) frequency. (C) Summary graph showing no differences between NPY+ interneurons and MSNs in sIPSC and mIPSC amplitude. (D) Summary graph showing no differences between NPY+ interneurons and MSNs in sIPSC and mIPSC weighted decay kinetics. 130 NPY+ A MSN 20 pA D Amplitude (pA) C Tw (ms) B Frequency (Hz) 500 ms 2.5 2 1.5 1 0.5 0 NPY+ MSN sIPSC mIPSC sIPSC mIPSC sIPSC mIPSC 30 20 10 0 40 30 20 10 0 131 MSNs (Figure 27), suggesting that these two neuronal populations contain GABAA receptors with similar subunit composition. However, evoked GABAergic current responses (eIPSCs) to intrastriatal stimulation were significantly different between these types of neurons. It must be noted, however, that intrastriatal stimulation does not stimulate a particular type of pre-synaptic cell (as dual recordings are able to control), and currents from intrastriatal stimulation are evoked from a population of striatal neurons. eIPSCs were pharmacologically isolated with application of NBQX (5 µM) and CPP (10 µM), glutamatergic receptor antagonists. At the end of each experiment, BMR (25 μM) was applied to further ensure pure GABAergic eIPSCs. In whole-cell recordings with KCl internal solutions supplemented with QX-314 (4 mM), the decay time of eIPSCs in NPY+ interneurons and MSNs were significantly longer than sIPSCs from the same cells (NPY+: 213.3 ± 21.5 % sIPSC decay, n = 19/9, p < 0.0005; MSNs: 459.4 ± 74.0 % sIPSC decay, n = 9/4, p < 0.005) (Figure 28B). In addition, MSN eIPSCs were significantly longer than NPY+ interneuron eIPSCs (MSN: 175.6 ± 28.0 ms, n = 9/4; NPY+: 69.5 ± 6.6.ms, n = 19/9; p < 0.0005), suggesting that MSNs receive more inhibitory input than NPY+ interneurons. MSN data from BAC NPY strains was confirmed in BAC Drd2 EGFP;BAC Drd1a tdTomato mice, and the eIPSC properties were found to be similar in confirmed MSNs (322.6 ± 41.0 % of sIPSC decay, n = 11/2, p = 0.1). Because there were no differences between MSNs from BAC NPY mice and those from BAC Drd2 EGFP;BAC Drd1a tdTomato mice, the majority of the MSN data was obtained from the latter mouse strain so distinctions could be made between the two MSN subtypes. 132 Figure 28. Evoked GABAergic Currents in NPY+ Interneurons and MSNs. (A) Representative current traces of eIPSCs in the presence of NBQX and CPP in response to intrastriatal stimulation (black lines). BMR was applied to confirm a purely GABAergic event (grey line). (B) Summary graph comparing decay kinetics between sIPSCs and eIPSCs in the same NPY+ (n = 18/10) interneuron or MSN (n = 9/5). * denotes significance between sIPSCs and eIPSCs of the same cell type. # denotes significance between eIPSCs in NPY+ interneurons and MSNs. 133 A NPY+ MSN 50 pA 20 pA 200 ms 200 ms ### B Tw (ms) 250 ** 200 150 *** 100 50 0 sIPSC eIPSC NPY+ 134 sIPSC eIPSC MSN Application of the modulating neuropeptide NPY was shown to decrease the spontaneous firing of action potentials in cortical NPY+ interneurons (Fu & van den Pol,2007), and the effects of the NPY peptide (1 µM) were investigated on striatal eIPSC measurements with CsCl-based internal solutions supplemented with QX-314. While these experiments were only conducted in 3 NPY+ interneurons from one mouse, the NPY peptide tended to reversibly decrease the eIPSC amplitude (control: 128.8 ± 67.3 pA; NPY peptide: 97.7 ± 44.5 pA, n = 3/1, p = 0.3), suggesting that striatal NPY+ interneurons may also be inhibited by NPY peptide application. d. Inhibitory Short Term Plasticity Differences in short term plasticity were characterized next to discern differences in pre-synaptic influences due to the apparent differences in GABAergic input between striatal NPY+ interneurons and MSNs. In this set of experiments, intrastriatal stimulation was set at 33 Hz with 7 pulses spaced 30 s apart, and cells were recorded with CsClbased internal solutions supplemented with QX-314 (4 mM). Figure 29A shows examples of these stimulus train eIPSC traces for both striatal cell types. In these conditions, NPY+ interneurons displayed short term depression (STD) with an STD ratio ([amplitude of IPSC7/amplitude of IPSC1]) of 0.18 ± 0.02 (n = 42/14), with both D1+ and D2+ MSNs from BAC Drd2 EGFP;BAC Drd1a tdTomato mice also expressing STD with a ratio of 0.32 ± 0.03 (n = 73/22). However, the STD ratio was significantly greater in NPY+ cells than MSNs (p < 0.0005). When this type of measurement of short term plasticity is used in this thesis, it will be referred to as baseline plasticity (Figure 29B). 135 Antidromic pallidal stimulation has also produced STD in striatal MSNs that was enhanced by D1R agonist application in the striatum (Tecuapetla et al, 2007). In my hands, pallidal stimulation produced STD in MSNs (0.47 ± 0.15, n = 4/3) from sagittal slices in BAC Drd2 EGFP;BAC Drd1a tdTomato mice. To determine if dopamine affects baseline plasticity differently between NPY+ interneurons and MSNs with intrastriatal stimulation, modulation by dopamine receptor agonists and antagonists was investigated. In these experiments, the D1R agonist was applied to NPY+ interneurons for over 5 minutes because they are known to express D1-like receptors (Rivera et al, 2002). In 4 NPY+ interneurons, SKF-81297 (10 µM) failed to alter STD (NPY+: 0.15 ± 0.04, SKF-81297: 0.16 ± 0.08, n = 4/2, p = 0.8). In these experiments, D1R and D2R agonists also failed to significantly alter STD in D1+ and D2+ MSNs (D1+: 0.36 ± 0.05, SKF-81297: 0.41 ± 0.06, n = 14/7, p = 0.9; D2+: 0.31 ± 0.04, Quinpirole: 0.31 ± 0.05, n = 20/10, p = 0.9). In order to determine whether striatal stimulation was evoking dopamine release in the striatum and preventing dopamine receptor agonists from having any significant effects, these same experiments were conducted in the presence of dopamine receptor antagonists. However, SCH 23390 (10 µM) failed to have an effect on either NPY+ interneurons (NPY+: 0.08 ± 0.03, SCH 23390: 0.12 ± 0.02, n = 3/1, p = 0.4) or D1+ MSNs (D1+: 0.29 ± 0.04, SCH23390: 0.36 ± 0.08, n = 14/8, p = 0.6), while Sulpiride (2 µM) slightly decreased the D2+ MSN STD (D2+: 0.26 ± 0.03, Sulpiride: 0.38 ± 0.05, n = 16/8, p < 0.05). Effects of D2R antagonists may be reflective of the increased dopamine sensitivity at D2Rs (Missale et al, 1998). 136 Figure 29. Short Term Plasticity in NPY+ Interneurons. (A) Representative traces of postsynaptic responses in NPY+ interneurons (left) and MSNs (right) to intrastriatal stimulation of 7 pulses (arrows) at 33 Hz. (B) Summary graph showing baseline STD in NPY+ interneurons (n = 42/14) and MSNs (n = 73/22). (C) Summary graph showing global depression in some NPY+ interneurons (n = 15/8), while others showed facilitation (n = 15/7). MSNs invariably expressed global facilitation (n = 78/22). 137 A NPY+ D2+ MSN NPY+ D1+ MSN 40 pA B Baseline Plasticity Ratio 1.0 NPY+ MSN .75 .50 .25 0 0 1 2 3 4 5 6 Stimulus # 7 138 Global Plasticity Ratio 100 ms C NPY+ (facil) NPY+ (depr) MSN 3.0 2.0 1.0 0 0 1 2 3 4 5 Stimulus # 6 7 Upon closer examination of individual short term plasticity traces, two types of NPY+ interneurons could be identified if the baseline was set at the cell‟s holding current. If short term plasticity was measured this way, a population of NPY+ interneurons expressed short term facilitation (STF) (1.64 ± 0.07, n = 15/7), while others expressed STD (0.67 ± 0.02, n = 15/8), or no change at all (1.0 ± 0.03, n = 11/5). When recorded from BAC Drd2 EGFP;BAC Drd1a tdTomato mice, both D1+ and D2+ MSNs showed STF (2.28 ± 0.17, n = 78/22) with this analysis method. Although this short term plasticity measurement is not used in the literature, it may suggest that there are two populations of NPY+ interneurons that receive inhibitory connections with differing release probabilities. When this type of measurement of short term plasticity is used in this thesis, it will be referred to as global plasticity (Figure 29C). Due to the differences in global plasticity among NPY+ interneurons, their plasticity was also measured with a stimulus frequency of 16 Hz to potentially identify the origin of differences in these interneurons. In a population of 10 NPY+ interneurons, 5 expressed global facilitation with an STF ratio of 1.33 ± 0.02 (n = 5/2), while the other 5 expressed global depression (0.72 ± 0.03, n = 5/3) when stimulated with 33 Hz. In response to 16 Hz stimulation, the 5 cells that expressed global facilitation with 33 Hz stimulation showed neither facilitation nor depression (0.92 ± 0.06, n = 5/2, p < 0.005). The 5 cells that showed depression with 33 Hz stimulation continued to show global depression with 16 Hz stimulation (0.68 ± 0.05, n = 5/3, p = 0.6). These data do not clearly indicate a presynaptic or postsynaptic origin for differences between these two types of NPY+ interneurons as these data can be explained by a frequency dependent presynaptic release or differences in postsynaptic receptor desensitization characteristics. 139 To determine if there are differences in presynaptic release probabilities onto these two types of striatal neurons, NPY+ interneurons and MSNs were subjected to two differing concentrations of Ca2+ and Mg2+, to alter presynaptic release conditions. The extracellular solution contained 2 mM Ca2+, 1 mM Mg2+ (2 Ca / 1 Mg), and was altered to contain 4 mM Ca2+, 1 mM Mg2+ (4 Ca /1 Mg) or 1 mM Ca2+, 2 mM Mg2+ (1 Ca / 2 Mg). The 4 Ca / 1 Mg solution was hypothesized to increase the presynaptic release of neurotransmitter, making the postsynaptic response as much as four times larger. On the other hand, the 1 Ca / 2 Mg solution was hypothesized to decrease neurotransmitter release as the greater Mg2+ concentration would block many presynaptic Ca2+ binding sites. As shown in Figure 30B, enhancing presynaptic release significantly increased the first evoked current amplitude in MSNs, but did not alter any other characteristics in MSNs or NPY+ interneurons. Decreasing presynaptic release probability, however, significantly decreased the amplitude of the first evoked current and the charge transfer in both NPY+ interneurons and MSNs. Additionally, 1 Ca / 2 Mg tended to increase the baseline short term plasticity in both striatal cell types, further enhancing their existing STD, and suggesting that these two types of striatal interneurons receive presynaptic inputs with similar release probabilities. If short term plasticity was measured as global plasticity, 4 Ca / 1 Mg and 1 Ca / 2 Mg largely had no impact on NPY+ interneurons, while they decreased and increased global facilitation in MSNs, respectively (Figure 30C). Thus, these experiments did not reveal any striking differences in presynaptic release probabilities between NPY+ interneurons and MSNs suggesting that differences between NPY+ interneuron and MSN global plasticity may be due to differences in 140 Figure 30. Alterations to Presynaptic Release Probabilities. (A) Representative current traces from a MSN in response to different concentrations of Ca2+ and Mg2+. Calibration bars: 100 pA and 50 ms. (B) Summary graph showing % control changes with 4 Ca / 1 Mg and 1 Ca / 2 Mg solutions on 1st amplitude, baseline plasticity, and charge transfer in NPY+ interneurons (n = 10/4) and MSNs (n = 12/3). (C) Summary graph showing % control changes with 4 Ca / 1 Mg and 1 Ca / 2 Mg solutions on global plasticity in NPY+ interneurons (facil: n = 3/2; depr: n = 7/3) and MSNs (n = 12/3). 141 A 2 Ca / 1 Mg B 2 Ca / 1 Mg 200 120 * * 150 80 * * 50 4 Ca / 1 Mg 1 Ca / 2 Mg 0 % Control Global Plasticity 150 100 * * 50 0 4 Ca / 1 Mg * 1st Ampl. Baseline Plasticity Charge 4 Ca / 1 Mg 1 Ca / 2 Mg 200 C BMR * MSN 100 40 0 1 Ca / 2 Mg 250 NPY+ 160 % Control 4 Ca / 1 Mg 1 Ca / 2 Mg 142 NPY + (facil) NPY + (depr) MSNs postsynaptic receptors. Although not investigated in this thesis, it is possible that the two groups of NPY+ interneurons differ in the sheer number of active synapses or postsynaptic receptor desensitization properties. For example, if MSNs have greater GABAergic synaptic inputs than NPY+ interneurons, as suggested with the eIPSC decay shown in Figure 28B, GABAergic synapses on NPY+ interneurons are likely to become desensitized faster than GABAergic synaptses on MSNs. Although both globally facilitating and depressing cells were seen within the same slice, the discrepancy between these two could be an issue of maturation. For example, globally depressing cells may be the result of less pre-synaptic contacts due to their relative immaturity compared to globally facilitating cells within the same slice. Because NPY+ interneurons showed two different firing patterns and two different types of global plasticity, experiments in KCl-based solutions were used to determine if these two characteristics were related. However, NPY+ interneurons with the PLTS-like firing patterns expressed both types of global plasticity and cells with LTS-like firing patterns also expressed both types of global plasticity, suggesting that these two characteristics are not related. Although these distinct groups of neurons may be regulated by development, the distinctive firing properties of both neuron types, as well as distinct plasticity characteristics were observed within the same slice at various ages tested. These data argue against these characteristics‟ developmental regulation. 143 DISCUSSION I. Summary The major finding of these studies indicates that tonic GABAergic conductance expression patterns differ between striatonigral and striatopallidal MSNs based on their expression of phosphorylated GABAA receptor β3 subunits. D2+ MSNs express tonic current in the acute slice preparation without pharmacological manipulation due to basally phosphorylated β3 receptor subunits. Intracellular application of the PKA inhibitor, PKI, significantly decreases D2+ MSN GABA-mediated tonic currents. Quinpirole, the D2R-like agonist also decreases D2+ MSN tonic current through G protein-coupled inhibition of the PKA phosphorylation cascade. Further, intracellular PKA phosphorylation was shown to induce significant β3 GABAA receptor subunitmediated tonic current in D1+ MSNs. Application of SKF-81297, the D1R-like agonist, also induces D1+ MSN tonic current by promoting the PKA phosphorylation cascade. In addition, studies using the β2/β3 selective agonist etomidate and conditional Drd2-β3 subunit KO animals show that this subunit is essential to the GABA-mediated tonic current characteristic of these striatal neurons. Prior to my thesis work, both the players and mechanism behind striatal GABAergic tonic current was unknown. My findings provide novel insight into the modulation and regulation of striatal MSN output, as well as the pathogenesis of PD. My work also characterizes the striatal LTS/NPY+ interneuron, which may provide the striatal slice with the ambient GABA necessary for tonic current. The importance of these neurons in striatal physiology is only beginning to be recognized and understood. 144 II. Etomidate Sensitivity As a general anesthetic, etomidate is an agonist at GABAA receptors containing either the β2 or the β3 subunit (Hill-Venning et al, 1997). Although this drug directly affects GABA receptors, and therefore has GABA-mimetic activity, etomidate also modulates or potentiates GABA currents. Because striatal MSNs are reported to lack the β2 subunit (Flores-Hernandez et al, 2000), etomidate proved to be an important pharmacological tool to investigate the role of the β subunit in mediating GABAA receptor tonic current. Previous studies have tested etomidate sensitivity in α1-, α2-, α3-, and α6containing recombinant GABAA receptors (Hill-Venning et al, 1997), and investigation into α5-containing recombinant receptors was needed to understand etomidate‟s effect in striatal MSNs since this subunit mediates tonic current in the striatum (Ade et al, 2008). My results show that etomidate lacks GABA potentiating specificity to β3 subunits since it potentiated GABA currents in α5β1γ2 and α5β3γ2 receptors to the same degree (see Figure 14). Etomidate‟s GABA-mimetic actions on these recombinant receptors were specific for the β3 subunit. β2 subunit specificity was not investigated since this subunit is not expressed in striatal MSNs (Flores-Hernandez et al, 2000). Because this is the first study to suggest that etomidate is not selective at β2/β3 subunits when combined with the α5 subunit, one can only speculate on potential causes for its lack of specificity. It is possible that etomidate‟s selectivity is determined by the β subunit, while maximal effects may be determined by interactions with other subunits. It was shown that etomidate‟s maximal potentiation was greater for β2- than β1- containing receptors in α1, α2, and α6-containing receptors, the maximal enhancement of the inward 145 currents produced by etomidate was no different between α3β2- and α3β1-containing receptors (Hill-Venning et al, 1997). Results from these recombinant receptor studies suggest that etomidate‟s ability to potentiate GABA responses is likely to depend on the identity of both α and β isoforms present in the receptor. The presence of the δ subunit may also increase etomidate‟s efficacy in recombinant receptors. One study showed that etomidate-potentiated GABA currents were approximately ten times greater in α4β3δ receptors than α4β3γ2 receptors, whereas the responses to other anesthetics did not differ between the δ- and γ-containing recombinant receptors (Brown et al, 2002). This study did not, however, determine the selectivity of etomidate in α4β1δ receptors. These three studies suggest that etomidate‟s ability to potentiate GABA responses in β2/β3 subunit-containing receptors are both influenced by and dependent upon the α subunit isoform and the presence of δ or γ subunits. Because etomidate‟s GABA-mimetic effects are likely dependent upon other subunits present in the GABAA receptors, pharmacological effects in the slice preparation are further complicated as the exact receptor composition of GABAA receptors in striatal MSNs remains unknown. It cannot be simply concluded that D2+ MSNs express substantially more β3 subunits than D1+ MSNs due to larger etomidate responses (see Figure 15) because other subunits may be influencing etomidate‟s effects in both types of striatal output neurons. However, it is likely that in TTX, and without presynaptic GABA release, etomidate‟s effects in the slice may more closely resemble the direct effects seen in the recombinant study, where etomidate showed selectivity for the β3containing receptors over those expressing the β1 subunit, independent of the α subunit. 146 Therefore, the data presented in this thesis suggests that because striatopallidal MSNs had greater etomidate responses, D2+ MSNs have more functional β3 subunits than D1+ MSNs. Intracellular PKA application and the D1-like receptor agonist, SKF-81297, increases the etomidate response in striatonigral MSNs and provides two important details: 1) Etomidate‟s direct effects are modulated by β subunit phosphorylation, and 2) D1+ MSNs also express functional β3-containing receptors when they are phosphorylated. This first finding is to be expected as β3 subunit phosphorylation has been reported to enhance GABA whole cell currents (McDonald et al, 1998), but implies that basal phosphorylation of GABAA receptor subunits may also modulate etomidate‟s activity and selectivity of β2 and β3 subunit-containing receptors. The second finding from these studies regarding a larger etomidate response in D1+ MSNs under certain intracellular conditions suggests that the difference between striatonigral and striatopallidal MSNs does not lie in their β3 subunit expression, but rather the extent of phosphorylation on their β3 subunits. III. PKA Phosphorylation of GABAA Receptor Subunits Due to the importance of β3 subunits in mediating tonic current and their distinct regulation by PKA phosphorylation (McDonald et al, 1998), the role of PKA phosphorylation in modulating GABA-mediated MSN tonic currents was subsequently investigated. To study the purely postsynaptic effects of PKA phosphorylation on MSN tonic current, kinases and phosphatases were supplemented into the internal solution to avoid unwanted effects on presynaptic terminal release of neurotransmitter (Asaumi et al, 147 2006). Due to these experimental parameters, many of the changes seen with PKA phosphorylation modulators are likely due to postsynaptic regulation. These experiments do not, however, rule out potential retrograde influences of the postsynaptic cell on presynaptic terminals (Sivakimaran et al, 2009), but they certainly decrease potential presynaptic effects. Although PKA phosphorylation was shown to decrease whole cell currents from acutely-dissociated striatal MSNs (Flores-Hernandez et al, 2000), the results presented in this thesis suggest, for the first time, that PKA phosphorylation regulates GABAmediated tonic current and that PKA phosphorylated β3 subunits regulate striatal MSN tonic conductance. Blockade of the PKA phosphorylation cascade and dephosphorylation of β3 subunits dramatically reduces tonic current in striatopallidal MSNs (see Figure 18). On the other hand, promoting intracellular PKA phosphorylation with PKA‟s catalytic subunit increases striatonigral MSN tonic current through a phosphorylated β3 subunit (see Figure 17). As tonic current regulates cell excitability in striatal output neurons (Ade et al, 2008), these studies suggest that a neuron‟s phosphorylation state may regulate tonic current as well as striatal excitability and output. However, a particularly noticeable caveat to this experimental method is that intracellular application of kinases and phosphatases also affects other target proteins within the postsynaptic neuron. PKA phosphorylation has been shown to modulate intracellular processes that lead to cell survival and cell division (Cross et al, 2000), as well as the function of many neurotransmitter channels (Moss et al, 1992b; Carvalho et al, 2000; Chen & Roche, 2007) and ion channels (Schiffmann et al, 1995; Chahine et al, 2005) which may affect cell excitability themselves. Therefore, intracellular application 148 of these peptides does not ensure that the changes observed in MSN tonic current are specifically due to the phosphorylation state of the β3 GABAA receptor subunit as PKA phosphorylation of other channels may also affect RMP and cell excitability themselves. Recently, a new transgenic mouse was generated by Dr. Moss‟s laboratory at Tufts University, where the PKA targeted serine residues of the β3 receptor subunit were mutated to alanine residues (β3(S408/9A)). Therefore, in these mice the GABAA receptor β3 subunit is unable to be phosphorylated. I expect that MSN tonic currents in this transgenic mouse will represent a narrow range of amplitudes, suggestive of a decrease in D2+ MSN tonic current, confirming the present studies that suggest the phosphorylation state of the β3 GABAA receptor subunit is responsible, at least in part, for mediating MSN tonic current. In a recent collaborative effort between Drs. Moss and Vicini, I have obtained these mice and plan to conduct these experiments to further verify the phosphorylated β3 subunit‟s role in striatal GABA-mediated tonic current and cell excitability. IV. Dopamine Receptor Regulation of BMR-Sensitive Tonic Current Although dopamine has been shown to affect whole cell currents in D1+ MSNs through interactions with the PKA phosphorylation cascade, the results presented here suggest that dopamine affects not only MSN tonic current, but also cell excitability as well. D2+ MSNs express GABA-mediated tonic current in the absence of dopaminergic tone in young striatal slices, while D1+ MSNs lack tonic current due to inactivated D1Rs. In fact, while pharmacologically stimulating the D2R with quinpirole significantly decreases D2+ MSN tonic current, stimulating the D1R with SKF-81297 significantly 149 increases the D1+ MSN tonic current due to its interactions with the PKA phosphorylation cascade (see Figure 20). Therefore, the dopaminergic tone of the striatal slice largely regulates the tonic current expression in striatal output neurons. It has been shown that MSN tonic current is excitatory and facilitates MSN cell excitability (Ade et al, 2008; Janssen et al, 2009). Indeed, application of quinpirole significantly decreases the D2+ MSN cell excitability by decreasing tonic current. Although the lack of tonic current in D1+ cells makes them less excitable than D2+ MSNs (Ade et al, 2008; Day et al, 2008; Gertler et al, 2008; Janssen et al, 2009), increasing tonic current with SKF-81297 application failed to affect young MSN excitability. The reasons why the D1R-like agonist failed to affect cell excitability while increasing tonic current remains unknown. It is possible that presynaptic activation of D1Rs on striatal interneurons will affect D1+ MSN excitability. For example, D1R agonists have been shown to depolarize LTS/NPY+ and FS/PV+ GABAergic interneurons (Chang & Kita, 1992), and their increased activity may enhance D1+ MSN tonic current due buildup of extracellular, ambient GABA. Additionally, D1R activation will also activate cholinergic interneurons, and increase their release of ACh (Aosaki et al, 1998; Damsma et al, 1990; Consolo et al, 1992). Downstream effects of ACh are complex, however, and vary with the type of postsynaptic receptor they activate. It is also likely that activation of postsynaptic dopamine receptors will alter the function of other proteins, including other ion channels. Dopamine-mediated modifications to these channels may also affect cell excitability, and counter any changes in cell excitability due to increased GABA-mediated tonic current. Although no mechanism has been 150 established to explain the lack of increased cell excitability on D1+ MSNs with D1R activation, it is likely to depend on the striatal physiology and abundance of striatal D1Rs. Several years ago, it was thought that the striatal output pathways were not distinct due to co-expression of D1R and D2R mRNA in single MSNs (Surmeier et al, 1996). However, two pieces of data presented in this thesis argue against this theory. First, receptor co-localization, as visualized with BAC Drd2 EGFP;BAC Drd1a tdTomato mice, is largely absent in young striatal brain slices (see Figure 9), as suggested by Shuen et al. (2008) with this same transgenic mouse. In addition, pharmacological studies presented here also support the theory of separate striatal output pathways with distinct, non-overlapping dopamine receptor expression. Application of D1R agonists had no effect on D2+ MSN synaptic or extrasynaptic receptors, and D2R agonists had no effect on D1+ MSNs. Taken together, these data strongly suggest that D1+ MSNs primarily express D1Rs, while D2+ MSNs primarily express D2Rs with little overlap in receptor expression. These data also indicate that dopamine regulates specific striatal output nuclei, offering more specific avenues for PD therapeutics. Based on my studies with etomidate, PKA phosphorylation, and dopamine receptor agonists, I propose the following mechanisms regulate MSN GABAergic tonic conductance (Figure 31): First, in basal conditions where little dopamine is released in the striatal slice preparation, D2+ MSN tonic current is mediated through a phosphorylated β3 subunit since the Gi/o protein remains inactive. The phosphorylated β3 subunit passes more current than the dephosphorylated subunit and D2+ MSNs express tonic current. Under the same conditions, D1+ MSNs lack tonic current because their β3 151 Figure 31. Tonic Conductance is Mediated through a Phosphorylated β3 Subunit. Under basal conditions (little to no dopamine), D2Rs do not activate the Gi/o protein to inhibit PKA phosphorylation, and β3 subunits are basally phosphorylated (green β3). Because the phosphorylated β3 subunit yields increased currents and may be more plentiful than extrasynaptic β1 subunits, D2+ MSNs exhibit GABA-mediated tonic current. Without dopamine, D1Rs do not activate the Gs protein to promote PKA phosphorylation, and the dephosphorylated β3 (red β3) subunits yield smaller tonic current in the D1+ than the D2+ MSNs. When stimulated, the D2R activates the Gi/o protein to inhibit PKA activity, dephosphorylating the β3 subunits. A dephosphorylated β3 subunit results in smaller tonic currents compared with basal conditions, reflective in the amount of tonic current observed. During D1R stimulation, the Gs protein activates cAMP and PKA phosphorylates the β3 subunit and increases D1+ MSN tonic currents. 152 Basal Conditions D1R D2R β3 β3 β3 Overall: β3 PKA β1 β3 β1 β3 β3 Tonic current β1 PKA β1 β3 Tonic current Quinpirole SKF-81297 D2R D1R β3 β3 β3 β1 cAMP, PKA β1 cAMP, PKA β1 β3 β3 β3 Overall: β3 Tonic current β1 β3 Tonic current 153 subunits remain dephosphorylated since the PKA phosphorylation cascade remains inactive due to the unstimulated D1R-coupled Gs protein. Next, when the D2R-like agonist quinpirole is applied, the D2R is stimulated and cAMP levels drop. Thus, the β3 subunits become dephosphorylated and tonic current is significantly reduced. When the D1R-like agonist SKF-81297 is applied, β3 subunits become phosphorylated due to Gs stimulation and increased levels of cAMP. β3 subunit-containing receptor currents increase and D1+ MSN tonic current is observed. V. Potential Role for GABAA Receptor Endocytosis The data presented here suggest that dopamine and intracellular phosphorylation regulate MSN tonic current. However, enhancement or suppression of GABAA receptor endocytosis may also explain the results presented in this thesis. As explained in the Introduction, Section VIIc, GABAA receptor endocytosis is regulated by PKA phosphorylation of the β3 subunit, and possibly the γ2 subunit. Blockade of phosphorylation allows the proper endocytotic machinery to bind to the β3 subunit. The receptors undergo endocytosis and the number of receptors at the cell surface decreases, leading to reduced current (Terunuma et al, 2008). In this dissertation I show that intracellular PKI application decreases D2+ MSN tonic current. While it is presumed that this decrease in current is due to decreased current through β3 subunits, it is possible that decreased cell surface β3-containing receptors may also be contributing to this observation. On the other hand, phosphorylation prevents the endocytotic machinery from binding to the β3 subunit, and effectively increases the numbers of GABAA receptors at 154 the cell surface (Kittler et al, 2000). Indeed, intracellular PKA application increases D1+ MSN tonic current. Again, while it is presumed that this increase in current is due to increased current through phosphorylated β3 subunits, it is possible that increased cell surface β3 subunit-containing receptors may also be contributing to this observation. Therefore, the results presented here may be due to complementing effects between altered current and altered receptor numbers at the cell surface although significant receptor endocytosis may take longer to occur than the 5 minute observation periods tested in this thesis (Kittler et al, 2004). Further experiments to address the endocytosis mechanism in these experimental conditions are planned. For such experiments, the dynamin inhibitory p4 peptide (QVPSRPNRAP, 50 µM) and PKA modulators will be added to the internal pipette solution as in Chen et al. (2006), and MSN tonic currents will be measured. This peptide blocks interactions between dynamin and amphiphysin, preventing endocytosis. Because the δ subunit was shown to not mediate tonic inhibition in young mice (Ade et al, 2008), the γ2 subunit remains important to striatal tonic current. The role that PKA phosphorylation has over this subunit and endocytosis was not investigated in this thesis, and therefore could also be playing a role in observed changes to tonic current. While altered rates of endocytosis may play a complementing role in the tonic current modulation presented in this thesis, I believe that the majority of the effects are indeed mediated through changes in individual receptor conductances. 155 VI. Interpretations of Conditional β3 Subunit KO Because the pharmacological sensitivity of etomidate does not give a definitive picture of β3 subunit expression and GABA-mediated tonic current in MSNs, I complemented these pharmacological studies with genetic studies where the β3 subunit was deleted. Conditional β3 subunit KO mice were generated in which the subunit was deleted from Drd2-expressing and Dlx5/6-expressing cells since pan-neuronal β3 subunit KO animals experience a high degree of neonatal mortality (Homanics et al, 1997). As with any KO animal, it is important to remain cognizant that since the β3 subunit is absent since birth, adaptive changes through development may affect normal physiology, making it difficult to make definitive statements about the protein of interest from genetic manipulation alone. The most significant, yet hypothesized, finding with conditional β3 subunit KO mice was the dramatic reduction in tonic current that was shown to be specifically due to the lack of the GABAA receptor β3 subunit (see Figure 24). Tonic current remained unchanged with Cre recombinase expression or homozygous floxed β3 alleles. These data strongly indicate the importance of this subunit in mediating D2+ MSN tonic current. These experiments do not address, however, the role of subunit phosphorylation in mediating tonic current. As previously mentioned, future experiments will further address this issue using a transgenic mouse where the specific β3 subunit PKA phosphorylation sites are mutated. It was originally hypothesized that β3 subunit KO mice should not respond to the β2/β3 subunit agonist etomidate due to its relatively specific actions at β3 subunitcontaining GABAA receptors in striatal MSNs. Surprisingly, mice from both strains of 156 conditional β3 subunit KOs expressed etomidate-mediated inward currents, although the average etomidate-mediated current was smaller in β3 subunit KO mice. Several explanations can be offered as to why etomidate responses are still intact, albeit reduced, in β3 subunit KO mice. First, Western blot data conducted in pan-neuronal β3 subunit KO mice suggest that the subunit is still present in small amounts (Ferguson et al, 2007) and, therefore, remaining β3 subunits may be responsible for the reduced etomidate response. Second, deletion of the β3 subunit may result in upregulation of β2 subunit expression to maintain homeostasis, and observed etomidate responses could be due to etomidate‟s actions on β2-containing GABAA receptors in MSNs. As presented in the Results, Section Ibii, etomidate‟s potentiating effects did not differ between α5β1- and α5β3-containing GABAA recombinant receptors. Therefore, another possible explanation for the etomidate responses seen in β3 subunit KO animals is due to its effects at α5β1containing receptors. Following this latter scenario, etomidate currents in wild-type mice would be mediated by a combination of α5β3- and α5β1-containing receptors. Thus, smaller etomidate currents would be expected in the β3 subunit KO mice as etomidate would be only affecting α5β1-containing receptors. Support for the theory that a homeostatic mechanism dictates the observed etomidate response is reflected in synaptic receptor properties in the β3 subunit KO mice. Since a reduction in postsynaptic receptors is electrophysiologically reflected by decreased IPSC amplitude, and the mIPSC peak amplitudes did not differ between Cre;Hetβ3 and KO mice, it is likely that another type(s) of GABAA receptor is upregulated to take the place of the non-functional β3-subunit containing receptors. The results presented here suggest that the α2 subunit primarily clusters with the β3 subunit, 157 as previously shown by Ramadan et al. (2003), since deletion of the β3 subunit reveals mIPSCs with significantly more rapid mIPSC decay kinetics. Deletion of the β3 subunit and removal of its associated receptors from the plasma membrane may leave more rapidly decaying α1 subunit-containing GABAA receptors. Additionally, it is possible that α1β2- or α1β1-containing receptors are upregulated upon β3 subunit deletion to further speed mIPSC decay. Because GABA-mediated tonic current has been shown to affect striatal MSN excitability and the β3 subunit affects tonic current, further experiments are planned to link the β3 GABAA receptor subunit with changes in MSN excitability. Furthermore, global β3 subunit KO mice were observed to have epilepsy (Homanics et al, 1997), suggestive of the subunits‟ essential role in neuronal excitability. Many studies have noted the crucial importance of the GABAA receptor β3 subunit; this subunit has been linked to many autism spectrum disorders (Buxbaum et al, 2002; Handforth et al, 2005) and childhood absence epilepsy (Feucht et al, 1999; Urak et al, 2006). Although the mechanism that links the β3 subunit with these developmental disorders is unknown, data presented in this thesis suggests that the β3 subunit is important in cell excitability and tonic current. It is plausible, therefore, that improper excitation and GABAergic tonic currents may underlie many of the physiological alterations characteristic of these disorders. VII. Potential GABAA Receptor Composition in Striatal MSNs One of the original goals for my thesis research was to determine the composition of GABAA receptors in striatonigral and striatopallidal MSNs. Although much of the 158 work presented in this thesis investigates receptor composition one subunit at a time, together these studies offer unprecedented pharmacological insight into MSN GABAA receptor composition. Indeed, the studies presented here investigated the function of α4, β2/β3, and δ subunits using a combination of pharmacological and genetic techniques. It is possible that D1+ and D2+ MSNs have a similar synaptic receptor composition since these two types of neurons have similar decay kinetics and respond similarly to the α1/2 subunit receptor agonist, zolpidem (Ade et al, 2008; personal observations). Etomidate‟s effects on mIPSCs did not differ between MSN subtypes, also suggesting that MSNs express similar synaptic receptors. Additionally, etomidate failed to significantly prolong mIPSC decay kinetics suggesting that synaptic MSN GABAA receptors contain both β1 and β3 subunits. Therefore, it is likely that both D1+ and D2+ MSN synaptic GABAA receptors are composed of α1β1γ2, α1β3γ2, α2β1γ2, α2β3γ2 receptors. Extrasynaptic or perisynaptic GABAA receptors that mediate tonic current have been shown to differ between D1+ and D2+ MSNs. Pharmacological manipulation of the α5 subunit revealed that this subunit was primarily expressed on D2+ MSNs (Ade et al, 2008), while pharmacology and genetic deletion of the δ subunit determined that this subunit was not mediating young MSN tonic current (Janssen et al, 2009), although it was expressed in MSNs (Ade et al, 2008). In addition, studies presented in this thesis suggest that the α4 subunit is expressed in both MSN subtypes, with greater pharmacological effects in D2+ MSNs (see Results, Section Ib). As the β2 subunit was not found to be expressed in striatal MSNs (Flores-Hernandez et al, 2000), etomidate sensitivity and subunit immunohistochemistry has shown that both D1+ and D2+ MSN 159 express β3 subunits (see Figure 12). Since these studies did not directly investigate the presence of the β1 subunit, definitive statements about their expression in MSNs cannot be made. Assuming that extrasynaptic and perisynaptic GABAA receptors regulate tonic current, D2+ MSNs are likely to express α4β1γ2, α4β3γ2, α5β1γ2, and α5β3γ2 receptors, while D1+ MSNs are likely to express α4β1γ2 and α4β3γ2 extrasynaptic receptors, as well as receptors with an unknown α subunit; extrasynaptic α2 subunit expression is possible. Whether these combinations of subunits form fully functioning GABAA receptors has not been explored. VIII. Developmental Regulation of MSN Tonic Current Because this thesis research focuses on tonic current in young mice, it is important to note the developmental changes in GABAA receptor composition (Laurie et al, 1992) and their potential impact on striatal MSN tonic current. The developmental regulation of tonic current is clearly demonstrated by the shifts observed in MSN tonic current patterns (Janssen et al, 2009; Santhakumar et al, 2010). While the α5 subunit was reported to mediate tonic current in young D2+ MSNs (Ade et al, 2008), this subunit failed to alter either D1+ or D2+ MSN tonic current in adult mice (Santhakumar et al, 2010). On the other hand, the δ subunit was not found to mediate tonic current in young mice (Janssen et al, 2009), whereas this subunit was essential to MSN tonic current in adult animals (Santhakumar et al, 2010). mRNA expression studies have shown that young striatal tissue expresses primarily α2, α5, β3, and γX GABAA receptor subunits, while adult tissue abundantly expresses α2, α4, β3, and δ subunits (Laurie et al, 1992). Importantly, striatal β subunit expression remains intact throughout development, 160 although it is more developmentally regulated in other brain regions (Laurie et al, 1992). Based on the available data, young MSN tonic current is likely mediated through α5β3γ2 receptors, while tonic current in older MSNs is likely mediated through αXβ3δ receptors. IX. Functional Implications of Striatal MSN GABAergic Tonic Current My thesis work has shown that under certain conditions, both striatonigral and striatopallidal MSNs express tonic GABA currents, regardless of age. These data suggest that these tonic currents are essential to the maintenance and proper function of striatal MSNs. In agreement with Ade et al. (2008), I found that blockade of D2+ MSN tonic current shifts the current versus frequency plots to the right (see Figure 23), suggesting that GABAergic tonic current facilitates cell excitability. However, a recent study showed that adult MSN tonic current protects neurons from excitotoxic cell death (Santhakumar et al, 2010), suggesting that tonic current reduces neuronal excitability that can lead to cell death. Therefore, my results suggest that GABAA receptor mediated tonic current enhances MSN excitability, while the study by Santhakumar et al. (2010) implies that tonic current reduces excitability. These studies differ not only in the age of animals tested, but also in their ways of observing and interpreting neuronal excitability. My study focused on young mice, and used whole-cell electrophysiology in current-clamp mode to determine changes in cell excitability. The study from Dr. Mody‟s lab investigated tonic current from adult MSNs and determined excitotoxicity with application of NMDA and measurements of MSN swelling (Santhakumar et al, 2010). Due to these different experimental parameters, it is possible that GABA-mediated tonic current exhibits a bimodal effect on neuronal excitability 161 based on the extracellular environment. For example, under basal conditions where the striatal slice is relatively quiet and MSNs have a hyperpolarized membrane potential, GABAergic tonic current may be depolarizing and promote neuronal excitation. Under a barrage of glutamatergic input (mimicked by the application of NMDA), the MSN membrane potential may sufficiently depolarize so that GABA-mediated tonic current is inhibitory, to prevent MSNs from potential toxicity and cell death. X. Inputs to NPY+ Interneurons The results presented here and in Partridge et al. (2009) on striatal NPY+ interneurons indicate that these interneurons receive inhibitory input. Although several analyses of sIPSC and mIPSC characteristics did not differ between striatal NPY+ interneurons and MSNs, data from striatally-evoked IPSCs strongly suggest that NPY+ interneurons receive far fewer inhibitory contacts than MSNs. Extensive experiments investigating short term plasticity and presynaptic release probabilities (see Figures 29, 30) failed to identify a clear explanation for this difference. Most likely, input onto NPY+ interneurons is sparser than the inhibitory input to MSNs. Or perhaps the postsynaptic receptors on NPY+ interneurons desensitize more rapidly than those on MSNs. While the experiments presented here only suggest that these two populations of striatal neurons receive different inhibitory input, this research does not investigate or identify the possible pre-synaptic terminals on NPY+ interneurons. In fact, intrastriatal stimulation in glutamatergic blockers will stimulate MSN axon collaterals, GABAergic interneuron axons, cholinergic interneuron axons, and inhibitory axons that originate in 162 the GP. Axons arising from the GP originate from only about a quarter of all pallidal neurons (Kita & Kitai, 1994), and selectively innervate striatal interneurons (Bevan et al, 1998). While most pallidostriatal terminals make contact with FS/PV+ interneurons, about 3-32% of terminals make contact with LTS/NPY+ cells on the proximal dendritic arborization (Bolam et al, 2000), although the extent of their inhibitory input has not been determined. Few contacts traveling over a great distance are likely to dampen the postsynaptic response seen in the striatal interneuron, and may actually make the response negligible. Although eIPSCs from NPY+ interneurons were smaller in amplitude and exhibited faster decay kinetics than MSNs, it is likely that NPY+ cells also receive intrastriatal inhibitory inputs to complement their nearly negligible pallidostriatal inputs. Although no studies have identified additional pre-synaptic contacts onto LTS/NPY+ interneurons, the possibility remains that, in addition to pallidal inputs, these neurons receive inhibitory contacts from within the striatum. The role that LTS/NPY+ interneurons play in striatal physiology remains unclear as studies have failed to find synaptic connections between LTS/NPY+ interneurons and striatal MSNs (Gittis et al, 2010). However, due to their spontaneous firing of action potentials, these interneurons remain prime candidates for providing the striatal slice with ambient GABA. If classical synaptic transmission does not occur with these pre-synaptic neurons, these cells may provide the slice with ambient GABA through volume transmission (for review, see Agnati et al, 2010). Although this type of communication has not been shown in the striatum, volume transmission originating from NPYexpressing neurogliaform cells has been shown to provide enough GABA for tonic 163 activation of extrasynaptic receptors in the cortex (Olah et al, 2009). It remains to be determined if these striatal LTS/NPY+ interneurons communicate with MSNs through volume transmission; such studies would further establish the crucial role of NPY+ interneurons in the striatal network and pathological diseases. XI. GABAergic Drugs in the Management and Treatment of Movement Disorders Although the causative physiology for many movement disorders has not been established, it is likely that striatal MSN output may play a role in the processing of motor control in many movement disorders. My thesis research suggests that MSN excitability and output is dependent upon GABA-mediated tonic current, and therefore it is likely that modulating the GABAergic network may ameliorate many symptoms that are characteristic of movement disorders such as PD, Tourette‟s syndrome, Huntington‟s disease, tardive dyskinesia, and restless leg syndrome. While GABAergic drugs are not first-line therapy for any of these movement disorders, there is sufficient clinical and basic science data to suggest that these agents may play a role in future therapeutics for movement disorders. My thesis works implies that MSN excitability and output is largely controlled by GABA-mediated tonic current (Janssen et al, 2009), and therefore, GABAergic agents may play a pivotal role in future therapeutics for PD. In the treatment of Tourette‟s syndrome, clinical trials have shown that baclofen (a GABAB receptor agonist) is effective in controlling tics in children (Singer et al, 2001), and basic science research has shown that there are altered levels of PV+ interneurons in the basal ganglia (Kalanithi et al, 2005), suggesting that control of 164 GABA release in basal ganglia structures may alleviate motor tics. While various treatments with GABAergic agents are being tested in Huntington‟s disease, tiagabine (a GABA reuptake inhibitor) has shown promise to reverse several behavioral and biochemical markers characteristic of the disease in experimental animal models (Dhir et al, 2008). For treatment of tardive dyskinesia, one study has shown preliminary evidence suggesting that benzodiazepines may have some effects in treatment of this movement disorder (Bhoopathi et al, 2006). Although dopaminergic agents are currently the firstline therapy for treatment of restless leg syndrome, gabapentin (a GABA analogue) has shown to be beneficial in some clinical trials (Merlino et al, 2009). It is likely that GABAergic drugs will be utilized in the management and treatment of many movement disorders in the near future. XII. Implications to Parkinson’s Disease Research The data presented here offers a possible mechanism by which striatal MSN excitability becomes imbalanced during PD. Specifically, PD is characterized by increased excitability and output of striatopallidal D2+ MSNs, while the striatonigral pathway exhibits decreased activity (Raz et al, 2000; Galeffi et al, 2003; Starr et al, 2005; Mallet et al, 2006). These imbalances result in the inability to move effectively. While previous work has shown that GABAergic tonic conductance modulates MSN excitability (Ade et al, 2008), this thesis work suggests three additionally important concepts in regards to the generation and treatment of this disease. First, dopamine alters both inhibitory tonic conductance and MSN excitability. My data show that application of a D2R-like agonist decreases both the D2+ MSN tonic current and D2+ MSN 165 excitability. It is likely that the converse is also true: dopamine depletion increases D2+ MSN excitability (Day et al, 2006; Mallet et al, 2006; Shen et al, 2007; Day et al, 2008). Secondly, dopamine modulation of tonic inhibition is mediated through its interactions with the PKA phosphorylation cascade. Lastly, GABAA receptor β3 subunits are largely responsible for tonic current in MSNs. These data suggest additional means of regulating MSN output, and present a possible therapeutic avenue for alleviating PD symptoms that arise due to imbalances in the striatal output pathways. 166 CONCLUSIONS In the mouse, D1+ and D2+ MSNs constitute unique projection pathways to the substantia nigra pars reticulata and globus pallidus, respectively. GABA-mimetic etomidate effects are selective for the β3 subunit in recombinant receptors expressed in HEK cells, although etomidate‟s ability to potentiate GABA currents appears to depend on both α and β subunits. D2+ MSNs express enhanced etomidate current compared to D1+ MSNs, suggesting that these cell types differ in their expression of functional β3 subunits. Intracellular PKA application increases D1+ MSN tonic current. Enhanced etomidate responses suggest that D1+ MSN tonic current is mediated through a phosphorylated β3 subunit. Intracellular PKI application reduces D2+ MSN tonic currents, suggesting the basal D2+ MSN tonic current is mediated via basally phosphorylated β3 subunits. Dopamine release in the striatal slice preparation is low, and dopamine receptors remain unstimulated. D2-like agonists decrease D2+ MSN tonic current, presumably through dephosphorylated β3 subunits, while D1-like agonists increase D1+ MSN tonic current through phosphorylated β3 subunits. Current-clamp experiments show that the D2-like agonist quinpirole reduces D2+ MSN excitability, suggesting that D2+ tonic current is excitatory. Drd2-β3 subunit KO animals express significantly less tonic current than wildtype mice, suggesting that the β3 subunit is imperative to D2+ MSN tonic current. 167 NPY+ interneurons and MSNs express similar synaptic GABAA receptor populations. Intrastriatal stimulation suggests that MSNs receive more inhibitory input than NPY+ interneurons due to prolonged evoked IPSC decay kinetics. 168 APPENDIX Order Details: JOURNAL OF NEUROSCIENCE Order detail ID: 42331428 ISSN: 0270-6474 Publication Year: 2010 Publisher: SOCIETY FOR NEUROSCIENCE Rightsholder: SOCIETY FOR NEUROSCIENCE Author/Editor: Planert et al Permission Status: Granted Permission type: Republish into a book, journal, newsletter… Requested use: Dissertation Republishing organization: GEORGETOWN UNIVERSITY Organization status: Not for profit Republication date: 07/02/2010 Circulation/Distribution: 1 Type of content: Figure, diagram, or table Description of requested content: Figure 1, Panel C Page range(s): 3501 Requested content's publication date: 03/03/2010 Order Details: JOURNAL OF NEUROSCIENCE Order detail ID: 42331427 ISSN: 0270-6474 Publication Year: 2009 Publisher: SOCIETY FOR NEUROSCIENCE Rightsholder: SOCIETY FOR NEUROSCIENCE Author/Editor: Janssen et al Permission Status: Granted Permission type: Republish into a book, journal, newsletter… Requested use: Dissertation Republishing organization: GEORGETOWN UNIVERSITY Organization status: Not for profit Republication date: 07/02/2010 Circulation/Distribution: 1 Type of content: Selected pages Description of requested content: Figures 1-8 Page range(s): 5117-5124 Number of pages: 8 Requested content's publication date: 04/22/2009 169 Order Details: JOURNAL OF NEUROPHYSIOLOGY Order detail ID: 42331426 ISSN: 0022-3077 Publication Year: 2009 Publisher: AMERICAN PHYSIOLOGICAL SOCIETY [ETC.] Rightsholder: AMERICAN PHYSIOLOGICAL SOCIETY Author/Editor: Partridge et al Permission Status: Granted Permission type: Republish into a book, journal, newsletter… Requested use: Dissertation Republishing organization: GEORGETOWN UNIVERSITY Organization status: Not for profit Republication date: 07/02/2010 Circulation/Distribution: 1 Type of content: Selected pages Description of requested content: Figures 3 and 6 Page range(s): 3041, 3043 Number of pages: 2 Requested content's publication date: 09/16/2009 License Number: 2427141041878 License date: May 13, 2010 License content publisher: Nature Publishing Group License content publication: Nature Reviews Neuroscience Licensed content Ttitle: Variations on an inhibitory theme: phasic and tonic activations of GABAA receptors Licensed content author: Mark Farrant, Zoltan Nusser Year of Publication: 2005 Portion used: Figures/tables Number of figures/tables: 1 Requestor Type: Student Type of Use: Thesis/Dissertation License Number: 2427141224801 License date: May 13, 2010 License content publisher: Nature Publishing Group License content publication: Nature Neuroscience Licensed content Ttitle: Indirect-pathway neurons lose their spines in Parkinson disease Licensed content author: Charles R Gerfen Year of Publication: 2006 Portion used: Figures/tables Number of figures/tables: 1 Requestor Type: Student Type of Use: Thesis/Dissertation 170 REFERENCES Ade KK, Janssen MJ, Ortinski PI, Vicini S. (2008) Differential tonic GABA conductances in striatal medium spiny neurons. J Neurosci. 28(5):1185-97. Agnati LF, Guidolin D, Guescini M, Genedani S, Fuxe K. (2010) Understanding wiring and volume transmission. Brain Res Rev. 64(1):137-159. Aizman O, Brismar H, Uhlén P, Zettergren E, Levey AI, Forssberg H, Greengard P, Aperia A. (2000) Anatomical and physiological evidence for D1 and D2 dopamine receptor colocalization in neostriatal neurons. Nat Neurosci. 3(3):226-30. Akaike A, Ohno Y, Sasa M, Takaori S. (1987) Excitatory and inhibitory effects of dopamine on neuronal activity of the caudate nucleus neurons in vitro. Brain Res. 418(2):262-72. Angelotti TP, Uhler MD, Macdonald RL. (1993) Enhancement of recombinant gammaaminobutyric acid type A receptor currents by chronic activation of cAMP-dependent protein kinase. Mol Pharmacol. 44(6):1202-10. Aosaki T, Kiuchi K, Kawaguchi Y. (1998) Dopamine D1-like receptor activation excites rat striatal large aspiny neurons in vitro. J Neurosci. 18(14):5180-90. Arias-Montaño JA, Floran B, Floran L, Aceves J, Young JM. (2007) Dopamine D(1) receptor facilitation of depolarization-induced release of gamma-amino-butyric acid in rat striatum is mediated by the cAMP/PKA pathway and involves P/Q-type calcium channels. Synapse. 61(5):310-9. Asaumi Y, Hasuo H, Akasu T. (2006) J Neurophysiol. 96(2):591-601. Dopamine presynaptically depresses fast inhibitory synaptic transmission via D4 receptor-protein kinase A pathway in the rat dorsolateral septal nucleus. Augood SJ, Emson PC. (1994) Adenosine A2a receptor mRNA is expressed by enkephalin cells but not by somatostatin cells in rat striatum: a co-expression study. Brain Res Mol Brain Res. 22(1-4):204-10. Azdad K, Chàvez M, Don Bischop P, Wetzelaer P, Marescau B, De Deyn PP, Gall D, Schiffmann SN. (2009) Homeostatic plasticity of striatal neurons intrinsic excitability following dopamine depletion. PLoS One. 4(9):6908. Bannai H, Lévi S, Schweizer C, Inoue T, Launey T, Racine V, Sibarita JB, Mikoshiba K, Triller A. (2009) Activity-dependent tuning of inhibitory neurotransmission based on GABAAR diffusion dynamics. Neuron. 62(5):670-82. Bargas J, Galarraga E, Aceves J. (1988) Electrotonic properties of neostriatal neurons are modulated by extracellular potassium. Exp Brain Res. (2):390-8. 171 Barnard EA, Skolnick P, Olsen RW, Mohler H, Sieghart W, Biggio G, Braestrup C, Bateson AN, Langer SZ. (1998) International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classification on the basis of subunit structure and receptor function. Pharmacol Rev. 50(2):291-313. Bedford FK, Kittler JT, Muller E, Thomas P, Uren JM, Merlo D, Wisden W, Triller A, Smart TG, Moss SJ. (2001) GABA(A) receptor cell surface number and subunit stability are regulated by the ubiquitin-like protein Plic-1. Nat Neurosci. 4(9):908-16. Belelli D, Callachan H, Hill-Venning C, Peters JA, Lambert JJ. (1996) Interaction of positive allosteric modulators with human and Drosophila recombinant GABA receptors expressed in Xenopus laevis oocytes. Br J Pharmacol. 118(3):563-76. Bennett BD, Bolam JP. (1993) Characterization of calretinin-immunoreactive structures in the striatum of the rat. Brain Res. 609(1-2):137-48. Bennett BD, Bolam JP. (1994) Synaptic input and output of parvalbuminimmunoreactive neurons in the neostriatum of the rat. Neuroscience. 62(3):707-19. Bennett BD, Wilson CJ. (1998) Synaptic regulation of action potential timing in neostriatal cholinergic interneurons. J Neurosci.18(20):8539-49. Bevan MD, Booth PA, Eaton SA, Bolam JP. (1998) Selective innervation of neostriatal interneurons by a subclass of neuron in the globus pallidus of the rat. J Neurosci. 18(22):9438-52. Bhoopathi PS, Soares-Weiser K. (2006) Benzodiazepines for neuroleptic-induced tardive dyskinesia. Cochrane Database Syst Rev. 19;3:CD000205. Bianchi MT, Haas KF, Macdonald RL. (2001) Structural determinants of fast desensitization and desensitization-deactivation coupling in GABAa receptors. J Neurosci. 21(4):1127-36. Bianchi MT, Haas KF, Macdonald RL. (2002) Alpha1 and alpha6 subunits specify distinct desensitization, deactivation and neurosteroid modulation of GABA(A) receptors containing the delta subunit. Neuropharmacology. 43(4):492-502. Bibb JA, Snyder GL, Nishi A, Yan Z, Meijer L, Fienberg AA, Tsai LH, Kwon YT, Girault JA, Czernik AJ, Huganir RL, Hemmings HC Jr, Nairn AC, Greengard P. (1999) Phosphorylation of DARPP-32 by Cdk5 modulates dopamine signalling in neurons. Nature. 1999 402(6762):669-71. Bogdanov Y, Michels G, Armstrong-Gold C, Haydon PG, Lindstrom J, Pangalos M, Moss SJ. (2006) Synaptic GABAA receptors are directly recruited from their extrasynaptic counterparts. EMBO J. 25(18):4381-9. 172 Bolam JP, Hanley JJ, Booth PA, Bevan MD. (2000) Synaptic organisation of the basal ganglia. J Anat. 196(4):527-42. Bolam JP, Ingham CA, Izzo PN, Levey AI, Rye DB, Smith AD, Wainer BH. (1986) Substance P-containing terminals in synaptic contact with cholinergic neurons in the neostriatum and basal forebrain: a double immunocytochemical study in the rat. Brain Res. 397(2):279-89. Bolam JP, Somogyi P, Takagi H, Fodor I, Smith AD. (1983) Localization of substance Plike immunoreactivity in neurons and nerve terminals in the neostriatum of the rat: a correlated light and electron microscopic study. J Neurocytol. 12(2):325-44. Bolam JP, Wainer BH, Smith AD. (1984) Characterization of cholinergic neurons in the rat neostriatum. A combination of choline acetyltransferase immunocytochemistry, Golgi-impregnation and electron microscopy. Neuroscience. 12(3):711-8. Bormann J, Hamill OP, Sakmann B. (1987) Mechanism of anion permeation through channels gated by glycine and gamma-aminobutyric acid in mouse cultured spinal neurones. J Physiol. 385:243-86. Bormann J. (1988) Electrophysiology of GABAA and GABAB receptor subtypes. Trends Neurosci. 11(3):112-6. Bormann J. (2000) The 'ABC' of GABA receptors. Trends Pharmacol Sci. 21(1):16-9. Bracci E, Centonze D, Bernardi G, Calabresi P. (2007) Dopamine excites fast-spiking interneurons in the striatum. J Neurophysiol. 87(4):2190-4. Brandon NJ, Delmas P, Kittler JT, McDonald BJ, Sieghart W, Brown DA, Smart TG, Moss SJ. (2000) GABAA receptor phosphorylation and functional modulation in cortical neurons by a protein kinase C-dependent pathway. J Biol Chem. 275(49):38856-62. Brandon NJ, Jovanovic JN, Colledge M, Kittler JT, Brandon JM, Scott JD, Moss SJ. (2003) A-kinase anchoring protein 79/150 facilitates the phosphorylation of GABA(A) receptors by cAMP-dependent protein kinase via selective interaction with receptor beta subunits. Mol Cell Neurosci. 22(1):87-97. Brandon NJ, Jovanovic JN, Smart TG, Moss SJ. (2002) Receptor for activated C kinase-1 facilitates protein kinase C-dependent phosphorylation and functional modulation of GABA(A) receptors with the activation of G-protein-coupled receptors. J Neurosci. 22(15):6353-61. Brandon NJ, Uren JM, Kittler JT, Wang H, Olsen R, Parker PJ, Moss SJ. (1999) Subunitspecific association of protein kinase C and the receptor for activated C kinase with GABA type A receptors. J Neurosci. 19(21):9228-34. 173 Brickley SG, Cull-Candy SG, Farrant M. (1996) Development of a tonic form of synaptic inhibition in rat cerebellar granule cells resulting from persistent activation of GABAA receptors. J Physiol. 497(Pt 3):753-9. Brown LL, Smith DM, Goldbloom LM. (1998) Organizing principles of cortical integration in the rat neostriatum: corticostriate map of the body surface is an ordered lattice of curved laminae and radial points. J Comp Neurol. 392(4):468-88. Brown N, Kerby J, Bonnert TP, Whiting PJ, Wafford KA. (2002) Pharmacological characterization of a novel cell line expressing human alpha(4)beta(3)delta GABA(A) receptors. Br J Pharmacol. 136(7):965-74. Brünig I, Scotti E, Sidler C, Fritschy JM. (2002) Intact sorting, targeting, and clustering of gamma-aminobutyric acid A receptor subtypes in hippocampal neurons in vitro. J Comp Neurol. 443(1):43-55. Brünig I, Sommer M, Hatt H, Bormann J. (1999) Dopamine receptor subtypes modulate olfactory bulb gamma-aminobutyric acid type A receptors. Proc Natl Acad Sci U S A. 96(5):2456-60. Brussaard AB, Wossink J, Lodder JC, Kits KS. (2000) Progesterone-metabolite prevents protein kinase C-dependent modulation of gamma-aminobutyric acid type A receptors in oxytocin neurons. Proc Natl Acad Sci U S A. 97(7):3625-30. Buxbaum JD, Silverman JM, Smith CJ, Greenberg DA, Kilifarski M, Reichert J, Cook EH Jr, Fang Y, Song CY, Vitale R. (2002) Association between a GABRB3 polymorphism and autism. Mol Psychiatry. 7(3):311-6. Calabresi P, Centonze D, Gubellini P, Pisani A, Bernardi G. (2000) Acetylcholinemediated modulation of striatal function. Trends Neurosci. 23(3):120-6. Calabresi P, Mercuri N, Stanzione P, Stefani A, Bernardi G. (1987) Intracellular studies on the dopamine-induced firing inhibition of neostriatal neurons in vitro: evidence for D1 receptor involvement. Neuroscience. 20(3):757-71. Canals M, Marcellino D, Fanelli F, Ciruela F, de Benedetti P, Goldberg SR, Neve K, Fuxe K, Agnati LF, Woods AS, Ferré S, Lluis C, Bouvier M, Franco R. (2003) Adenosine A2A-dopamine D2 receptor-receptor heteromerization: qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J Biol Chem. 278(47):46741-9. Caraiscos VB, Elliott EM, You-Ten KE, Cheng VY, Belelli D, Newell JG, Jackson MF, Lambert JJ, Rosahl TW, Wafford KA, MacDonald JF, Orser BA. (2004) Tonic inhibition in mouse hippocampal CA1 pyramidal neurons is mediated by alpha5 subunit-containing gamma-aminobutyric acid type A receptors. Proc Natl Acad Sci U S A 101(10):3662-7. 174 Carvalho AL, Duarte CB, Carvalho AP. (2000) Regulation of AMPA receptors by phosphorylation. Neurochem Res. 25(9-10):1245-55. Centonze D, Bracci E, Pisani A, Gubellini P, Bernardi G, Calabresi P. (2002) Activation of dopamine D1-like receptors excites LTS interneurons of the striatum. Eur J Neurosci. 15(12):2049-52. Centonze D, Usiello A, Gubellini P, Pisani A, Borrelli E, Bernardi G, Calabresi P. (2002) Dopamine D2 receptor-mediated inhibition of dopaminergic neurons in mice lacking D2L receptors. Neuropsychopharmacology. 27(5):723-6. Chahine M, Ziane R, Vijayaragavan K, Okamura Y. (2005) Regulation of Na v channels in sensory neurons. Trends Pharmacol Sci. 26(10):496-502. Chang HT, Kita H. (1992) Interneurons in the rat striatum: relationships between parvalbumin neurons and cholinergic neurons. Brain Res. 574(1-2):307-11. Chen BS, Roche KW. (2007) Regulation of NMDA receptors by phosphorylation. Neuropharmacology. 53(3):362-8. Chen G, Kittler JT, Moss SJ, Yan Z. (2006) Dopamine D3 receptors regulate GABAA receptor function through a phospho-dependent endocytosis mechanism in nucleus accumbens. J Neurosci. 26(9):2513-21. Chesselet MF, Robbins E. (1989) Characterization of striatal neurons expressing high levels of glutamic acid decarboxylase messenger RNA. Brain Res. 492(1-2):237-44. Choi DS, Wei W, Deitchman JK, Kharazia VN, Lesscher HM, McMahon T, Wang D, Qi ZH, Sieghart W, Zhang C, Shokat KM, Mody I, Messing RO. (2008) Protein kinase Cdelta regulates ethanol intoxication and enhancement of GABA-stimulated tonic current. J Neurosci. 28(46):11890-9. Cicchetti F, Beach TG, Parent A. (1998) Chemical phenotype of calretinin interneurons in the human striatum. Synapse. 30(3):284-97. Ciruela F, Burgueño J, Casadó V, Canals M, Marcellino D, Goldberg SR, Bader M, Fuxe K, Agnati LF, Lluis C, Franco R, Ferré S, Woods AS. (2004) Combining mass spectrometry and pull-down techniques for the study of receptor heteromerization. Direct epitope-epitope electrostatic interactions between adenosine A2A and dopamine D2 receptors. Anal Chem. 76(18):5354-63. Cobb SR, Buhl EH, Halasy K, Paulsen O, Somogyi P. (1995) Synchronization of neuronal activity in hippocampus by individual GABAergic interneurons. Nature. 378(6552):75-8. 175 Consolo S, Girotti P, Russi G, Di Chiara G. (1992) Endogenous dopamine facilitates striatal in vivo acetylcholine release by acting on D1 receptors localized in the striatum. J Neurochem. 59(4):1555-7. Cooper AJ, Stanford IM. (2001) Dopamine D2 receptor mediated presynaptic inhibition of striatopallidal GABA(A) IPSCs in vitro. Neuropharmacology. 41(1):62-71. Cope DW, Hughes SW, Crunelli V. (2005) GABAA receptor-mediated tonic inhibition in thalamic neurons. J Neurosci. 25(50):11553-63. Cowan RL, Wilson CJ, Emson PC, Heizmann CW. (1990) Parvalbumin-containing GABAergic interneurons in the rat neostriatum. J Comp Neurol. 302(2):197-205. Craig AM, Blackstone CD, Huganir RL, Banker G. (1994) Selective clustering of glutamate and gamma-aminobutyric acid receptors opposite terminals releasing the corresponding neurotransmitters. Proc Natl Acad Sci U S A. 91(26):12373-7. Cross TG, Scheel-Toellner D, Henriquez NV, Deacon E, Salmon M, Lord JM. (2000) Serine/threonine protein kinases and apoptosis. Exp Cell Res. 256(1):34-41. Cunha RA. (2001) Regulation of the ecto-nucleotidase pathway in rat hippocampal nerve terminals. Neurochem Res. 26(8-9):979-91. Czubayko U, Plenz D. (2002) Fast synaptic transmission between striatal spiny projection neurons. Proc Natl Acad Sci U S A. 99(24):15764-9. Damsma G, Tham CS, Robertson GS, Fibiger HC. (1990) Dopamine D1 receptor stimulation increases striatal acetylcholine release in the rat. Eur J Pharmacol. 186(23):335-8. Day M, Wang Z, Ding J, An X, Ingham CA, Shering AF, Wokosin D, Ilijic E, Sun Z, Sampson AR, Mugnaini E, Deutch AY, Sesack SR, Arbuthnott GW, Surmeier DJ. (2006) Selective elimination of glutamatergic synapses on striatopallidal neurons in Parkinson disease models. Nat Neurosci. 9(2):251-9. Day M, Wokosin D, Plotkin JL, Tian X, Surmeier DJ. (2008) Differential excitability and modulation of striatal medium spiny neuron dendrites. J Neurosci. 28(45):11603-14. DeBoer P, Westerink BH. (1994) GABAergic modulation of striatal cholinergic interneurons: an in vivo microdialysis study. J Neurochem. 62(1):70-5. Delgado A, Sierra A, Querejeta E, Valdiosera RF, Aceves J. (2000) Inhibitory control of the GABAergic transmission in the rat neostriatum by D2 dopamine receptors. Neuroscience. 95(4):1043-8. 176 Dhir A, Akula KK, Kulkarni SK. (2008) Tiagabine, a GABA uptake inhibitor, attenuates 3-nitropropionic acid-induced alterations in various behavioral and biochemical parameters in rats. Prog Neuropsychopharmacol Biol Psychiatry. 32(3):835-43. Drasbek KR, Jensen K. (2006) THIP, a hypnotic and antinociceptive drug, enhances an extrasynaptic GABAA receptor-mediated conductance in mouse neocortex. Cereb Cortex. 16(8):1134-41. Ducić I, Caruncho HJ, Zhu WJ, Vicini S, Costa E. (1995) gamma-Aminobutyric acid gating of Cl- channels in recombinant GABAA receptors. J Pharmacol Exp Ther. 272(1):438-45. Essrich C, Lorez M, Benson JA, Fritschy JM, Lüscher B. (1998) Postsynaptic clustering of major GABAA receptor subtypes requires the gamma 2 subunit and gephyrin. Nat Neurosci. 1(7):563-71. Farrant M, Nusser Z. (2005) Variations on an inhibitory theme: phasic and tonic activation of GABA(A) receptors. Nat Rev Neurosci. 6(3):215-29. Feigenspan A, Bormann J. (1994) Facilitation of GABAergic signaling in the retina by receptors stimulating adenylate cyclase. Proc Natl Acad Sci U S A. 91(23):10893-7. Ferguson C, Hardy SL, Werner DF, Hileman SM, Delorey TM, Homanics GE. (2007) New insight into the role of the beta3 subunit of the GABAA-R in development, behavior, body weight regulation, and anesthesia revealed by conditional gene knockout. BMC Neurosci. 8:85. Ferré S, Fredholm BB, Morelli M, Popoli P, Fuxe K. (1997) Adenosine-dopamine receptor-receptor interactions as an integrative mechanism in the basal ganglia. Trends Neurosci. 20(10):482-7. Ferré S, Torvinen M, Antoniou K, Irenius E, Civelli O, Arenas E, Fredholm BB, Fuxe K. (1998) Adenosine A1 receptor-mediated modulation of dopamine D1 receptors in stably cotransfected fibroblast cells. J Biol Chem. 273(8):4718-24. Ferré S, von Euler G, Johansson B, Fredholm BB, Fuxe K. (1991) Stimulation of highaffinity adenosine A2 receptors decreases the affinity of dopamine D2 receptors in rat striatal membranes. Proc Natl Acad Sci U S A. 88(16):7238-41. Feucht M., Fuchs K., Pichlbauer E., Hornik K., Scharfetter J., Goessler R., Fureder T., Cvetkovic N., Sieghart W., Kasper S., et al. (1999) Possible association between childhood absence epilepsy and the gene encoding GABRB3. Biol. Psychiat. 46:997– 1002. 177 Ficker E, Taglialatela M, Wible BA, Henley CM, Brown AM. (1994) Spermine and spermidine as gating molecules for inward rectifier K+ channels. Science. 1994 266(5187):1068-72. Figueredo-Cardenas G, Morello M, Sancesario G, Bernardi G, Reiner A. (1996) Colocalization of somatostatin, neuropeptide Y, neuronal nitric oxide synthase and NADPH-diaphorase in striatal interneurons in rats. Brain Res. 735(2):317-24. Filippova N, Sedelnikova A, Zong Y, Fortinberry H, Weiss DS. (2000) Regulation of recombinant gamma-aminobutyric acid (GABA)(A) and GABA(C) receptors by protein kinase C. Mol Pharmacol. 57(5):847-56. Fink JS, Weaver DR, Rivkees SA, Peterfreund RA, Pollack AE, Adler EM, Reppert SM. (1992) Molecular cloning of the rat A2 adenosine receptor: selective co-expression with D2 dopamine receptors in rat striatum. Brain Res Mol Brain Res. 14(3):186-95. Fino E, Glowinski J, Venance L. (2007) Effects of acute dopamine depletion on the electrophysiological properties of striatal neurons. Neurosci Res. 58(3):305-16. Fisher JL, Hinkle DJ, Macdonald RL. (2000) Loreclezole inhibition of recombinant alpha1beta1gamma2L GABA(A) receptor single channel currents. Neuropharmacology. 39(2):235-45. Fisher JL, Macdonald RL. (1997) Single channel properties of recombinant GABAA receptors containing gamma 2 or delta subtypes expressed with alpha 1 and beta 3 subtypes in mouse L929 cells. J Physiol. 505 2):283-97. Floran B, Gonzalez B, Florán L, Erlij D, Aceves J. (2005) Interactions between adenosine A(2a) and dopamine D2 receptors in the control of [(3)H]GABA release in the globus pallidus of the rat. Eur J Pharmacol. 520(1-3):43-50. Flores-Hernandez J, Hernandez S, Snyder GL, Yan Z, Fienberg AA, Moss SJ, Greengard P, Surmeier DJ. (2000) D(1) dopamine receptor activation reduces GABA(A) receptor currents in neostriatal neurons through a PKA/DARPP-32/PP1 signaling cascade. J Neurophysiol. 83(5):2996-3004. Foster JD, Pananusorn B, Cervinski MA, Holden HE, Vaughan RA. (2003) Dopamine transporters are dephosphorylated in striatal homogenates and in vitro by protein phosphatase 1. Brain Res Mol Brain Res. 110(1):100-8. Fu LY, van den Pol AN. (2007) GABA excitation in mouse hilar neuropeptide Y neurons. J Physiol. 579(Pt 2):445-64. Galarreta M, Hestrin S. (2001) Electrical synapses between GABA-releasing interneurons. Nat Rev Neurosci. (6):425-33. 178 Galeffi F, Bianchi L, Bolam JP, Della Corte L. (2003) The effect of 6-hydroxydopamine lesions on the release of amino acids in the direct and indirect pathways of the basal ganglia: a dual microdialysis probe analysis. Eur J Neurosci. 18(4):856-68. Gerfen CR, Baimbridge KG, Miller JJ. (1985) The neostriatal mosaic: compartmental distribution of calcium-binding protein and parvalbumin in the basal ganglia of the rat and monkey. Proc Natl Acad Sci U S A. 82(24):8780-4. Gerfen CR, Engber TM, Mahan LC, Susel Z, Chase TN, Monsma FJ Jr, Sibley DR. (1990) D1 and D2 dopamine receptor-regulated gene expression of striatonigral and striatopallidal neurons. Science. 250(4986):1429-32. Gerfen CR. (1992) The neostriatal mosaic: multiple levels of compartmental organization in the basal ganglia. Annu Rev Neurosci. 15:285-320. Gerfen CR. (2004) The basal ganglia. In: The rat nervous system, Ed 3 (Paxinos G, ed), pp 455–508. New York: Academic. Gertler TS, Chan CS, Surmeier DJ. (2008) Dichotomous anatomical properties of adult striatal medium spiny neurons. J Neurosci. 28(43):10814-24. Gibson JR, Beierlein M, Connors BW. (1999) Two networks of electrically coupled inhibitory neurons in neocortex. Nature. 402(6757):75-9. Ginés S, Hillion J, Torvinen M, Le Crom S, Casadó V, Canela EI, Rondin S, Lew JY, Watson S, Zoli M, Agnati LF, Verniera P, Lluis C, Ferré S, Fuxe K, Franco R. (2000) Dopamine D1 and adenosine A1 receptors form functionally interacting heteromeric complexes. Proc Natl Acad Sci U S A. 97(15):8606-11. Gittis AH, Nelson AB, Thwin MT, Palop JJ, Kreitzer AC. (2010) Distinct roles of GABAergic interneurons in the regulation of striatal output pathways. J Neurosci. 30(6):2223-34. Glykys J, Mody I. (2007) Activation of GABAA receptors: views from outside the synaptic cleft. 56(5):763-70. Glykys J, Peng Z, Chandra D, Homanics GE, Houser CR, Mody I. (2007) A new naturally occurring GABA(A) receptor subunit partnership with high sensitivity to ethanol. Nat Neurosci. 10(1):40-8. Gong S, Doughty M, Harbaugh CR, Cummins A, Hatten ME, Heintz N, Gerfen CR. Targeting Cre recombinase to specific neuron populations with bacterial artificial chromosome constructs. (2007) J Neurosci. 27(37):9817-23. 179 Gong S, Zheng C, Doughty ML, Losos K, Didkovsky N, Schambra UB, Nowak NJ, Joyner A, Leblanc G, Hatten ME, Heintz N. (2003) A gene expression atlas of the central nervous system based on bacterial artificial chromosomes. Nature. 425(6961):917-25. Graveland GA, DiFiglia M. (1985) The frequency and distribution of medium-sized neurons with indented nuclei in the primate and rodent neostriatum. Brain Res. 327(12):307-11. Graybiel AM, Ragsdale CW Jr. (1978) Histochemically distinct compartments in the striatum of human, monkeys, and cat demonstrated by acetylthiocholinesterase staining. Proc Natl Acad Sci U S A. 75(11):5723-6. Greengard P, Allen PB, Nairn AC. (1999) Beyond the dopamine receptor: the DARPP32/protein phosphatase-1 cascade. Neuron. 23(3):435-47. Gustafson N, Gireesh-Dharmaraj E, Czubayko U, Blackwell KT, Plenz D. (2006) A comparative voltage and current-clamp analysis of feedback and feedforward synaptic transmission in the striatal microcircuit in vitro. J Neurophysiol. 95(2):737-52. Guzmán JN, Hernández A, Galarraga E, Tapia D, Laville A, Vergara R, Aceves J, Bargas J. (2003) Dopaminergic modulation of axon collaterals interconnecting spiny neurons of the rat striatum. J Neurosci. 23(26):8931-40. Haas KF, Macdonald RL. (1999) GABAA receptor subunit gamma2 and delta subtypes confer unique kinetic properties on recombinant GABAA receptor currents in mouse fibroblasts. J Physiol. 514 (1):27-45. Handforth A, DeLorey TM, Homanics GE, Olsen RW. (2005) Pharmacologic evidence for abnormal thalamocortical functioning in GABAA receptor beta3 subunit-deficient mice, a model of Angelman Syndrome. Epilepsia 46(12)1860-1870. Harsing LG Jr, Zigmond MJ. (1997) Influence of dopamine on GABA release in striatum: evidence for D1-D2 interactions and non-synaptic influences. Neuroscience. 77(2):419-29. Hemmings HC Jr, Greengard P, Tung HY, Cohen P. (1984) DARPP-32, a dopamineregulated neuronal phosphoprotein, is a potent inhibitor of protein phosphatase-1. Nature. 310(5977):503-5. Herd MB, Haythornthwaite AR, Rosahl TW, Wafford KA, Homanics GE, Lambert JJ, Belelli D. (2008) The expression of GABAA beta subunit isoforms in synaptic and extrasynaptic receptor populations of mouse dentate gyrus granule cells. J Physiol. 586(4):989-1004. 180 Hernández-Echeagaray E, Galarraga E, Bargas J. (1998) 3-Alpha-chloro-imperialine, a potent blocker of cholinergic presynaptic modulation of glutamatergic afferents in the rat neostriatum. Neuropharmacology. 37(12):1493-502. Hernández-López S, Bargas J, Surmeier DJ, Reyes A, Galarraga E. (1997) D1 receptor activation enhances evoked discharge in neostriatal medium spiny neurons by modulating an L-type Ca2+ conductance. J Neurosci. 17(9):3334-42. Hernández-López S, Tkatch T, Perez-Garci E, Galarraga E, Bargas J, Hamm H, Surmeier DJ. (2000) D2 dopamine receptors in striatal medium spiny neurons reduce L-type Ca2+ currents and excitability via a novel PLC[beta]1-IP3-calcineurin-signaling cascade. J Neurosci. 20(24):8987-95. Herring D, Huang R, Singh M, Dillon GH, Leidenheimer NJ. (2005) PKC modulation of GABAA receptor endocytosis and function is inhibited by mutation of a dileucine motif within the receptor beta 2 subunit. Neuropharmacology 48(2):181-94. Herring D, Huang R, Singh M, Robinson LC, Dillon GH, Leidenheimer NJ. (2003) Constitutive GABAA receptor endocytosis is dynamin-mediated and dependent on a dileucine AP2 adaptin-binding motif within the beta 2 subunit of the receptor. J Biol Chem. 278(26):24046-52. Hersch SM, Gutekunst CA, Rees HD, Heilman CJ, Levey AI. (2004) Distribution of m1m4 muscarinic receptor proteins in the rat striatum: light and electron microscopic immunocytochemistry using subtype-specific antibodies. J Neurosci. 14(5):3351-63. Heuschneider G, Schwartz RD. (1989) cAMP and forskolin decrease gammaaminobutyric acid-gated chloride flux in rat brain synaptoneurosomes. Proc Natl Acad Sci U S A. 86(8):2938-42. Hevers W, Lüddens H. (2002) Pharmacological heterogeneity of gamma-aminobutyric acid receptors during development suggests distinct classes of rat cerebellar granule cells in situ. Neuropharmacology. 42(1):34-47. Hillion J, Canals M, Torvinen M, Casado V, Scott R, Terasmaa A, Hansson A, Watson S, Olah ME, Mallol J, Canela EI, Zoli M, Agnati LF, Ibanez CF, Lluis C, Franco R, Ferre S, Fuxe K. (2002) Coaggregation, cointernalization, and codesensitization of adenosine A2A receptors and dopamine D2 receptors. J Biol Chem. 277(20):18091-7. Hill-Venning C, Belelli D, Peters JA, Lambert JJ. (1997) Subunit-dependent interaction of the general anaesthetic etomidate with the gamma-aminobutyric acid type A receptor. Br J Pharmacol. 120(5):749-56. Hinkle DJ, Macdonald RL. (2003) Beta subunit phosphorylation selectively increases fast desensitization and prolongs deactivation of alpha1beta1gamma2L and alpha1beta3gamma2L GABA(A) receptor currents. J Neurosci. 23(37):11698-710. 181 Hjorth J, Blackwell KT, Kotaleski JH. (2009) Gap junctions between striatal fast-spiking interneurons regulate spiking activity and synchronization as a function of cortical activity. J Neurosci. 29(16):5276-86. Homanics GE, DeLorey TM, Firestone LL, Quinlan JJ, Handforth A, Harrison NL, Krasowski MD, Rick CE, Korpi ER, Mäkelä R, Brilliant MH, Hagiwara N, Ferguson C, Snyder K, Olsen RW. (1997) Mice devoid of gamma-aminobutyrate type A receptor beta3 subunit have epilepsy, cleft palate, and hypersensitive behavior. Proc Natl Acad Sci U S A. 94(8):4143-8. Hooper KC, Banks DA, Stordahl LJ, White IM, Rebec GV. (1997) Quinpirole inhibits striatal and excites pallidal neurons in freely moving rats. Neurosci Lett. 237(2-3):69-72. Hourez R, Azdad K, Vanwalleghem G, Roussel C, Gall D, Schiffmann SN. (2005) Activation of protein kinase C and inositol 1,4,5-triphosphate receptors antagonistically modulate voltage-gated sodium channels in striatal neurons. Brain Res. 1059(2):189-96. Huntsman MM, Porcello DM, Homanics GE, DeLorey TM, Huguenard JR. (1999) Reciprocal inhibitory connections and network synchrony in the mammalian thalamus. Science. 283(5401):541-3. Jacob TC, Bogdanov YD, Magnus C, Saliba RS, Kittler JT, Haydon PG, Moss SJ. (2005) Gephyrin regulates the cell surface dynamics of synaptic GABAA receptors. J Neurosci. 25(45):10469-78. Jacob TC, Wan Q, Vithlani M, Saliba RS, Succol F, Pangalos MN, Moss SJ. (2009) GABA(A) receptor membrane trafficking regulates spine maturity. Proc Natl Acad Sci U S A. 106(30):12500-5. Janssen MJ, Ade KK, Fu Z, Vicini S. (2009) Dopamine modulation of GABA tonic conductance in striatal output neurons. J Neurosci. 29(16):5116-26. Jarvis MF, Williams M. (1989) Direct autoradiographic localization of adenosine A2 receptors in the rat brain using the A2-selective agonist, [3H]CGS 21680. Eur J Pharmacol. 168(2):243-6. Jia F, Pignataro L, Schofield CM, Yue M, Harrison NL, Goldstein PA. (2005) An extrasynaptic GABAA receptor mediates tonic inhibition in thalamic VB neurons. J Neurophysiol. 94(6):4491-501. Jia F, Yue M, Chandra D, Keramidas A, Goldstein PA, Homanics GE, Harrison NL. (2008) Taurine is a potent activator of extrasynaptic GABA(A) receptors in the thalamus. J Neurosci. 28(1):106-15. Jiang ZG, North RA. (1991) Membrane properties and synaptic responses of rat striatal neurones in vitro. J Physiol. 443:533-53. 182 Jones A, Korpi ER, McKernan RM, Pelz R, Nusser Z, Mäkelä R, Mellor JR, Pollard S, Bahn S, Stephenson FA, Randall AD, Sieghart W, Somogyi P, Smith AJ, Wisden W. (1997) Ligand-gated ion channel subunit partnerships: GABAA receptor alpha6 subunit gene inactivation inhibits delta subunit expression. J Neurosci. 17(4):1350-62. Jones MV, Westbrook GL. (1997) Shaping of IPSCs by endogenous calcineurin activity. J Neurosci. 17(20):7626-33. Jovanovic JN, Thomas P, Kittler JT, Smart TG, Moss SJ. (2004) Brain-derived neurotrophic factor modulates fast synaptic inhibition by regulating GABA(A) receptor phosphorylation, activity, and cell-surface stability. J Neurosci. 24(2):522-30. Kaila K. (1994) Ionic basis of GABAA receptor channel function in the nervous system. Prog Neurobiol. 42(4):489-537. Kalanithi PS, Zheng W, Kataoka Y, DiFiglia M, Grantz H, Saper CB, Schwartz ML, Leckman JF, Vaccarino FM. (2005) Altered parvalbumin-positive neuron distribution in basal ganglia of individuals with Tourette syndrome. Proc Natl Acad Sci U S A. 102(37):13307-12. Kano M, Konnerth A. (1992) Potentiation of GABA-mediated currents by cAMPdependent protein kinase. Neuroreport. 3(7):563-6. Kapur J, Macdonald RL. (1996) Cyclic AMP-dependent protein kinase enhances hippocampal dentate granule cell GABAA receptor currents. J Neurophysiol. 76(4):262634. Kawaguchi Y, Wilson CJ, Augood SJ, Emson PC. (1995) Striatal interneurones: chemical, physiological and morphological characterization. Trends Neurosci. 18(12):527-35. Kawaguchi Y. (1993) Physiological, morphological, and histochemical characterization of three classes of interneurons in rat neostriatum. J Neurosci. 13(11):4908-23. Kelly PA, Thomas CL, Ritchie IM, Arbuthnott GW. (1994) Cerebrovascular autoregulation in response to hypertension induced by NG-nitro-L-arginine methyl ester. Neuroscience. 59(1):13-20. Kita H, Kosaka T, Heizmann CW. (1990) Parvalbumin-immunoreactive neurons in the rat neostriatum: a light and electron microscopic study. Brain Res. 536(1-2):1-15. Kita H. (1996) Glutamatergic and GABAergic postsynaptic responses of striatal spiny neurons to intrastriatal and cortical stimulation recorded in slice preparations. Neuroscience. 70(4):925-40. 183 Kita H, Kitai ST. (1994) The morphology of globus pallidus projection neurons in the rat: an intracellular staining study. Brain Res. 636(2):308-19. Kittler JT, Chen G, Honing S, Bogdanov Y, McAinsh K, Arancibia-Carcamo IL, Jovanovic JN, Pangalos MN, Haucke V, Yan Z, Moss SJ. (2005) Phospho-dependent binding of the clathrin AP2 adaptor complex to GABAA receptors regulates the efficacy of inhibitory synaptic transmission. Proc Natl Acad Sci U S A. 102(41):14871-6. Kittler JT, Chen G, Kukhtina V, Vahedi-Faridi A, Gu Z, Tretter V, Smith KR, McAinsh K, Arancibia-Carcamo IL, Saenger W, Haucke V, Yan Z, Moss SJ. (2008) Regulation of synaptic inhibition by phospho-dependent binding of the AP2 complex to a YECL motif in the GABAA receptor gamma2 subunit. Proc Natl Acad Sci U S A. 105(9):3616-21. Kittler JT, Delmas P, Jovanovic JN, Brown DA, Smart TG, Moss SJ. (2000) Constitutive endocytosis of GABAA receptors by an association with the adaptin AP2 complex modulates inhibitory synaptic currents in hippocampal neurons. J Neurosci. 20(21):79727. Kittler JT, Moss SJ. (2003) Modulation of GABAA receptor activity by phosphorylation and receptor trafficking: implications for the efficacy of synaptic inhibition. Curr Opin Neurobiol. 13(3):341-7. Kittler JT, Thomas P, Tretter V, Bogdanov YD, Haucke V, Smart TG, Moss SJ. (2004) Huntingtin-associated protein 1 regulates inhibitory synaptic transmission by modulating gamma-aminobutyric acid type A receptor membrane trafficking. Proc Natl Acad Sci U S A. 101(34):12736-41. Kneussel M, Betz H. (2000) Receptors, gephyrin and gephyrin-associated proteins: novel insights into the assembly of inhibitory postsynaptic membrane specializations. J Physiol. 525 Pt 1:1-9. Kneussel M, Brandstätter JH, Laube B, Stahl S, Müller U, Betz H. (1999) Loss of postsynaptic GABA(A) receptor clustering in gephyrin-deficient mice. J Neurosci. 19(21):9289-97. Koós T, Tepper JM, Wilson CJ. (2004) Comparison of IPSCs evoked by spiny and fastspiking neurons in the neostriatum. J Neurosci. 24(36):7916-22. Koós T, Tepper JM. (1999) Inhibitory control of neostriatal projection neurons by GABAergic interneurons. Nat Neurosci. (5):467-72. Koós T, Tepper JM. (2002) Dual cholinergic control of fast-spiking interneurons in the neostriatum. J Neurosci. 22(2):529-35. Kreitzer AC, Malenka RC. (2008) Striatal plasticity and basal ganglia circuit function. Neuron. 60(4):543-54. 184 Kreitzer AC. (2009) Physiology and pharmacology of striatal neurons. Annu Rev Neurosci. 32:127-47. Kubota Y, Kawaguchi Y. (2000) Dependence of GABAergic synaptic areas on the interneuron type and target size. J Neurosci. 20(1):375-86. Kubota Y, Mikawa S, Kawaguchi Y. (1993) Neostriatal GABAergic interneurones contain NOS, calretinin or parvalbumin. Neuroreport. 5(3):205-8. Kuroiwa M, Bateup HS, Shuto T, Higashi H, Tanaka M, Nishi A. (2008) Regulation of DARPP-32 phosphorylation by three distinct dopamine D1-like receptor signaling pathways in the neostriatum. J Neurochem. 107(4):1014-26. Lapper SR, Bolam JP. (1992) Input from the frontal cortex and the parafascicular nucleus to cholinergic interneurons in the dorsal striatum of the rat. Neuroscience. 51(3):533-45. Laurie DJ, Wisden W, Seeburg PH. (1992) The distribution of thirteen GABAA receptor subunit mRNAs in the rat brain. III. Embryonic and postnatal development. J Neurosci. 12(11):4151-72. Lee TH, Balu R, Davidson C, Ellinwood EH. (2001) Differential time-course profiles of dopamine release and uptake changes induced by three dopamine uptake inhibitors. Synapse. 41(4):301-10. Lehmann J, Langer SZ. (1983) The striatal cholinergic interneuron: synaptic target of dopaminergic terminals? Neuroscience. 10(4):1105-20. Lenz S, Perney TM, Qin Y, Robbins E, Chesselet MF. (1994) GABA-ergic interneurons of the striatum express the Shaw-like potassium channel Kv3.1. Synapse. 18(1):55-66. Lindskog M, Svenningsson P, Fredholm BB, Greengard P, Fisone G. (1999) Activation of dopamine D2 receptors decreases DARPP-32 phosphorylation in striatonigral and striatopallidal projection neurons via different mechanisms. Neuroscience. 88(4):1005-8. Liste I, Caruncho HJ, Guerra MJ, Labandeira-Garcia JL. (1997) GABA(A) receptor subunit expression in intrastriatal striatal grafts comparison between normal developing striatum and developing striatal grafts. Brain Res Dev Brain Res. 103(2):185-94. Lovinger DM, Choi S. (1995) Activation of adenosine A1 receptors initiates short-term synaptic depression in rat striatum. Neurosci Lett. 199(1):9-12. Lüscher B, Keller CA. (2004) Regulation of GABAA receptor trafficking, channel activity, and functional plasticity of inhibitory synapses. Pharmacol Ther. 102(3):195221. 185 Macdonald RL, Olsen RW. (1994) GABAA receptor channels. Annu Rev Neurosci. 17:569-602. Machu TK, Firestone JA, Browning MD. (1993) Ca2+/calmodulin-dependent protein kinase II and protein kinase C phosphorylate a synthetic peptide corresponding to a sequence that is specific for the gamma 2L subunit of the GABAA receptor. J Neurochem. 61(1):375-7. Malenka RC, Kocsis JD. (1988) Presynaptic actions of carbachol and adenosine on corticostriatal synaptic transmission studied in vitro. J Neurosci. 8(10):3750-6. Mallet N, Ballion B, Le Moine C, Gonon F. (2006) Cortical inputs and GABA interneurons imbalance projection neurons in the striatum of parkinsonian rats. J Neurosci. 26(14):3875-84. Mangan PS, Sun C, Carpenter M, Goodkin HP, Sieghart W, Kapur J. (2005) Cultured Hippocampal Pyramidal Neurons Express Two Kinds of GABAA Receptors. Mol Pharmacol. 67(3):775-88. Manzoni O, Prezeau L, Marin P, Deshager S, Bockaert J, Fagni L. (1992) Nitric oxideinduced blockade of NMDA receptors. Neuron. 8(4):653-62. Mao X; Fujiwara Y; Chapdelaine A; Yang H; Orkin SH. (2001) Activation of EGFP expression by cre-mediated excision in a new ROSA26 reporter mouse strain. Blood. 97(1):324-6. Marsh M, McMahon HT. (1999) The structural era of endocytosis. Science. 285(5425):215-20. Martone ME, Armstrong DM, Young SJ, Groves PM. (1992) Ultrastructural examination of enkephalin and substance P input to cholinergic neurons within the rat neostriatum. Brain Res. 594(2):253-62. Matamales M, Bertran-Gonzalez J, Salomon L, Degos B, Deniau JM, Valjent E, Hervé D, Girault JA. (2009) Striatal medium-sized spiny neurons: identification by nuclear staining and study of neuronal subpopulations in BAC transgenic mice. PLoS One. 4(3):e4770. Maurice N, Mercer J, Chan CS, Hernandez-Lopez S, Held J, Tkatch T, Surmeier DJ. (2004) D2 dopamine receptor-mediated modulation of voltage-dependent Na+ channels reduces autonomous activity in striatal cholinergic interneurons. J Neurosci. 24(46):10289-301. McDonald BJ, Amato A, Connolly CN, Benke D, Moss SJ, Smart TG. (1998) Adjacent phosphorylation sites on GABAA receptor beta subunits determine regulation by cAMPdependent protein kinase. Nat Neurosci. 1(1):23-8. 186 McKernan RM, Whiting PJ. (1996) Which GABAA-receptor subtypes really occur in the brain? Trends Neurosci. 19(4):139-43. Mercuri NB, Calabresi P, Stefani A, Stratta F, Bernardi G. (1991) GABA depolarizes neurons in the rat striatum: an in vivo study. Synapse. 8(1):38-40. Meredith GE, Wouterlood FG. (1990) Hippocampal and midline thalamic fibers and terminals in relation to the choline acetyltransferase-immunoreactive neurons in nucleus accumbens of the rat: a light and electron microscopic study. J Comp Neurol. 296(2):20421. Merlino G, Serafini A, Young JJ, Robiony F, Gigli GL, Valente M. (2009) Gabapentin enacarbil, a gabapentin prodrug for the treatment of the neurological symptoms associated with disorders such as restless legs syndrome. Curr Opin Investig Drugs. 10(1):91-102. Mermelstein PG, Song WJ, Tkatch T, Yan Z, Surmeier DJ. (1998) Inwardly rectifying potassium (IRK) currents are correlated with IRK subunit expression in rat nucleus accumbens medium spiny neurons. J Neurosci. 18(17):6650-61. Mihalek RM, Banerjee PK, Korpi ER, Quinlan JJ, Firestone LL, Mi ZP, Lagenaur C, Tretter V, Sieghart W, Anagnostaras SG, Sage JR, Fanselow MS, Guidotti A, Spigelman I, Li Z, DeLorey TM, Olsen RW, Homanics GE. Attenuated sensitivity to neuroactive steroids in gamma-aminobutyrate type A receptor delta subunit knockout mice. (1999) Proc Natl Acad Sci U S A. 96(22):12905-10. Missale C, Nash SR, Robinson SW, Jaber M, Caron MG. (1998) Dopamine receptors: from structure to function. Physiol Rev. 78(1):189-225. Mody I. (2001) Distinguishing between GABA(A) receptors responsible for tonic and phasic conductances. Neurochem Res. 26(8-9):907-13. Momiyama T, Koga E. (2001) Dopamine D(2)-like receptors selectively block N-type Ca(2+) channels to reduce GABA release onto rat striatal cholinergic interneurones. J Physiol. 533(Pt 2):479-92. Mori A, Shindou T, Ichimura M, Nonaka H, Kase H. (1996) The role of adenosine A2a receptors in regulating GABAergic synaptic transmission in striatal medium spiny neurons. J Neurosci. 16(2):605-11. Mori A, Shindou T. (2003) Modulation of GABAergic transmission in the striatopallidal system by adenosine A2A receptors: a potential mechanism for the antiparkinsonian effects of A2A antagonists. Neurology. 61(11):S44-8. Moss SJ, Gorrie GH, Amato A, Smart TG. (1995) Modulation of GABAA receptors by tyrosine phosphorylation. Nature. 377(6547):344-8. 187 Moss SJ, Doherty CA, Huganir RL. (1992a) Identification of the cAMP-dependent protein kinase and protein kinase C phosphorylation sites within the major intracellular domains of the beta 1, gamma 2S, and gamma 2L subunits of the gamma-aminobutyric acid type A receptor. J Biol Chem. 267(20):14470-6. Moss SJ, Smart TG, Blackstone CD, Huganir RL. (1992b) Functional modulation of GABAA receptors by cAMP-dependent protein phosphorylation. Science. 257(5070):661-5. Moss SJ, Smart TG. (2001) Constructing inhibitory synapses. Nat Rev Neurosci. 2(4):240-50. Moyer JT, Wolf JA, Finkel LH. (2007) Effects of dopaminergic modulation on the integrative properties of the ventral striatal medium spiny neuron. J Neurophysiol. 98(6):3731-48. Mozrzymas JW, Zarnowska ED, Pytel M, Mercik K. (2003) Modulation of GABA(A) receptors by hydrogen ions reveals synaptic GABA transient and a crucial role of the desensitization process. J Neurosci. 23(22):7981-92. Mtchedlishvili Z, Kapur J. (2006) High-affinity, slowly desensitizing GABAA receptors mediate tonic inhibition in hippocampal dentate granule cells. Mol Pharmacol. 69(2):56475. Murase K, Ryu PD, Randic M (1989) Excitatory and inhibitory amino acids and peptideinduced responses in acutely isolated rat spinal dorsal horn neurons. Neurosci Lett 103:56–63. Nicola SM, Malenka RC. (1998) Modulation of synaptic transmission by dopamine and norepinephrine in ventral but not dorsal striatum. J Neurophysiol. 79(4):1768-76. Nicola SM, Surmeier J, Malenka RC. (2000) Dopaminergic modulation of neuronal excitability in the striatum and nucleus accumbens. Annu Rev Neurosci. 23:185-215. Nisenbaum ES, Wilson CJ, Foehring RC, Surmeier DJ. (1996) Isolation and characterization of a persistent potassium current in neostriatal neurons. J Neurophysiol. 76(2):1180-94. Nisenbaum ES, Wilson CJ. (1995) Potassium currents responsible for inward and outward rectification in rat neostriatal spiny projection neurons. J Neurosci. (6):4449-63. Nisenbaum ES, Xu ZC, Wilson CJ. (1994) Contribution of a slowly inactivating potassium current to the transition to firing of neostriatal spiny projection neurons. J Neurophysiol. 71(3):1174-89. 188 Nishi A, Bibb JA, Snyder GL, Higashi H, Nairn AC, Greengard P. (2000) Amplification of dopaminergic signaling by a positive feedback loop. Proc Natl Acad Sci U S A. 97(23):12840-5. Nishi A, Snyder GL, Greengard P. (1997) Bidirectional regulation of DARPP-32 phosphorylation by dopamine. J Neurosci. 17(21):8147-55. Nörenberg W, Wirkner K, Illes P. (1997) Effect of adenosine and some of its structural analogues on the conductance of NMDA receptor channels in a subset of rat neostriatal neurones. Br J Pharmacol. 122(1):71-80. Nusser Z, Kay LM, Laurent G, Homanics GE, Mody I. (2001) Disruption of GABA(A) receptors on GABAergic interneurons leads to increased oscillatory power in the olfactory bulb network. J Neurophysiol. 86(6):2823-33. Nusser Z, Mody I. (2002) Selective modulation of tonic and phasic inhibitions in dentate gyrus granule cells. J Neurophysiol. 87(5):2624-8. Nusser Z, Sieghart W, Mody I (1999) Differential regulation of synaptic GABAA receptors by cAMP-dependent protein kinase in mouse cerebellar and olfactory bulb neurons. J Physiol 521:421–435. Nusser Z, Sieghart W, Somogyi P. (1998) Segregation of different GABAA receptors to synaptic and extrasynaptic membranes of cerebellar granule cells. J Neurosci. 18(5):1693-703. Nymann-Andersen J, Wang H, Chen L, Kittler JT, Moss SJ, Olsen RW. (2002) Subunit specificity and interaction domain between GABA(A) receptor-associated protein (GABARAP) and GABA(A) receptors. J Neurochem. 80(5):815-23. Oláh S, Füle M, Komlósi G, Varga C, Báldi R, Barzó P, Tamás G. (2009) Regulation of cortical microcircuits by unitary GABA-mediated volume transmission. Nature. 461(7268):1278-81. Olson PA, Tkatch T, Hernandez-Lopez S, Ulrich S, Ilijic E, Mugnaini E, Zhang H, Bezprozvanny I, Surmeier DJ. (2005) G-protein-coupled receptor modulation of striatal CaV1.3 L-type Ca2+ channels is dependent on a Shank-binding domain. J Neurosci. 25(5):1050-62. Onn SP, Fienberg AA, Grace AA. (2003) Dopamine modulation of membrane excitability in striatal spiny neurons is altered in DARPP-32 knockout mice. J Pharmacol Exp Ther. 306(3):870-9. Ortinski PI, Lu C, Takagaki K, Fu Z, Vicini S. (2004) Expression of distinct alpha subunits of GABAA receptor regulates inhibitory synaptic strength. J Neurophysiol. 92(3):1718-27. 189 Ouimet CC, Greengard P. (1990) Distribution of DARPP-32 in the basal ganglia: an electron microscopic study. J Neurocytol. 19(1):39-52. Ouimet CC, Miller PE, Hemmings HC Jr, Walaas SI, Greengard P. (1984) DARPP-32, a dopamine- and adenosine 3':5'-monophosphate-regulated phosphoprotein enriched in dopamine-innervated brain regions. III. Immunocytochemical localization. J Neurosci. 4(1):111-24. Owens DF, Kriegstein AR. (2002) Is there more to GABA than synaptic inhibition? Nat Rev Neurosci. 3(9):715-27. Pacheco-Cano MT, Bargas J, Hernández-López S, Tapia D, Galarraga E. (1996) Inhibitory action of dopamine involves a subthreshold Cs(+)-sensitive conductance in neostriatal neurons. Exp Brain Res. 110(2):205-11. Pakhotin P, Bracci E. (2007) Cholinergic interneurons control the excitatory input to the striatum. J Neurosci. 27(2):391-400. Partridge JG, Janssen MJ, Chou DY, Abe K, Zukowska Z, Vicini S. (2009) Excitatory and inhibitory synapses in neuropeptide Y-expressing striatal interneurons. J Neurophysiol. 102(5):3038-45. Perez MF, White FJ, Hu XT. (2006) Dopamine D(2) receptor modulation of K(+) channel activity regulates excitability of nucleus accumbens neurons at different membrane potentials. J Neurophysiol. 96(5):2217-28. Perez-Rosello T, Figueroa A, Salgado H, Vilchis C, Tecuapetla F, Guzman JN, Galarraga E, Bargas J. (2005) Cholinergic control of firing pattern and neurotransmission in rat neostriatal projection neurons: role of CaV2.1 and CaV2.2 Ca2+ channels. J Neurophysiol. 93(5):2507-19. Phelps PE, Houser CR, Vaughn JE. (1985) Immunocytochemical localization of choline acetyltransferase within the rat neostriatum: a correlated light and electron microscopic study of cholinergic neurons and synapses. J Comp Neurol. 238(3):286-307. Pirker S, Schwarzer C, Wieselthaler A, Sieghart W, Sperk G. (2000) GABA(A) receptors: immunocytochemical distribution of 13 subunits in the adult rat brain. Neuroscience. 101(4):815-50. Planert H, Szydlowski SN, Hjorth JJ, Grillner S, Silberberg G. (2010) Dynamics of synaptic transmission between fast-spiking interneurons and striatal projection neurons of the direct and indirect pathways. J Neurosci. 30(9):3499-507. Plenz D. (2003) When inhibition goes incognito: feedback interaction between spiny projection neurons in striatal function. Trends Neurosci. 26(8):436-43. 190 Plotkin JL, Wu N, Chesselet MF, Levine MS. (2005) Functional and molecular development of striatal fast-spiking GABAergic interneurons and their cortical inputs. Eur J Neurosci. 22(5):1097-108. Poisbeau P, Cheney MC, Browning MD, Mody I. (1999) Modulation of synaptic GABAA receptor function by PKA and PKC in adult hippocampal neurons. J Neurosci. 19(2):674-83. Popoli P, Betto P, Reggio R, Ricciarello G. (1995) Adenosine A2A receptor stimulation enhances striatal extracellular glutamate levels in rats. Eur J Pharmacol. 287:215–217. Porter NM, Twyman RE, Uhler MD, Macdonald RL. (1990) Cyclic AMP-dependent protein kinase decreases GABAA receptor current in mouse spinal neurons. Neuron. 5(6):789-96. Prenosil GA, Schneider Gasser EM, Rudolph U, Keist R, Fritschy JM, Vogt KE. (2006) Specific subtypes of GABAA receptors mediate phasic and tonic forms of inhibition in hippocampal pyramidal neurons. J Neurophysiol. 96(2):846-57. Radnikow G, Misgeld U. (1998) Dopamine D1 receptors facilitate GABAA synaptic currents in the rat substantia nigra pars reticulata. J Neurosci. 18(6):2009-16. Rajput PS, Kharmate G, Somvanshi RK, Kumar U. (2009) Colocalization of dopamine receptor subtypes with dopamine and cAMP-regulated phosphoprotein (DARPP-32) in rat brain. Neurosci Res. 65(1):53-63. Ramadan E, Fu Z, Losi G, Homanics GE, Neale JH, Vicini S. (2003) GABA(A) receptor beta3 subunit deletion decreases alpha2/3 subunits and IPSC duration. J Neurophysiol. 89(1):128-34. Ramanathan S, Hanley JJ, Deniau JM, Bolam JP. (2002) Synaptic convergence of motor and somatosensory cortical afferents onto GABAergic interneurons in the rat striatum. J Neurosci. 22(18):8158-69. Rashid AJ, So CH, Kong MM, Furtak T, El-Ghundi M, Cheng R, O'Dowd BF, George SR. (2007) D1-D2 dopamine receptor heterooligomers with unique pharmacology are coupled to rapid activation of Gq/11 in the striatum. Proc Natl Acad Sci U S A. 104(2):654-9. Raz A, Feingold A, Zelanskaya V, Vaadia E, Bergman H. (1996) Neuronal synchronization of tonically active neurons in the striatum of normal and parkinsonian primates. J Neurophysiol. 76(3):2083-8. Richtand NM, Kelsoe JR, Segal DS, Kuczenski R. (1995) Regional quantification of D1, D2, and D3 dopamine receptor mRNA in rat brain using a ribonuclease protection assay. Brain Res Mol Brain Res. 33(1):97-103. 191 Rivera A, Alberti I, Martín AB, Narváez JA, de la Calle A, Moratalla R. (2002) Molecular phenotype of rat striatal neurons expressing the dopamine D5 receptor subtype. Eur J Neurosci. 16(11):2049-58. Robello M, Amico C, Cupello A. (1993) Regulation of GABAA receptor in cerebellar granule cells in culture: differential involvement of kinase activities. Neuroscience. 53(1):131-8. Rodríguez-Pallares J, Labandeira-García JL, Muñoz A, Caruncho HJ. (2000) Morphology and neurochemistry of two striatal neuronal subtypes expressing the GABA(A) receptor alpha3-subunit in the rat. Brain Res. 876(1-2):124-30. Rosin DL, Hettinger BD, Lee A, Linden J. (2003) Anatomy of adenosine A2A receptors in brain: morphological substrates for integration of striatal function. Neurology. 61(11):S12-8. Salgado H, Tecuapetla F, Perez-Rosello T, Perez-Burgos A, Perez-Garci E, Galarraga E, Bargas J. (2005) A reconfiguration of CaV2 Ca2+ channel current and its dopaminergic D2 modulation in developing neostriatal neurons. J Neurophysiol. 94(6):3771-87. Saliba RS, Pangalos M, Moss SJ. (2008) The ubiquitin-like protein Plic-1 enhances the membrane insertion of GABAA receptors by increasing their stability within the endoplasmic reticulum. J Biol Chem. 283(27):18538-44. Sanna E, Murgia A, Casula A, Biggio G (1997) Differential subunit dependence of the actions of the general anesthetics alphaxalone and etomidate at gamma-aminobutyric acid type A receptors expressed in Xenopus laevis oocytes. Mol Pharmacol 51:484–490. Santhakumar V, Jones RT, Mody I. (2010) Developmental regulation and neuroprotective effects of striatal tonic GABA(A) currents. Neuroscience. 167(3):644-55 Saxena NC, Macdonald RL. (1994) Assembly of GABAA receptor subunits: role of the delta subunit. J Neurosci. 14(11):7077-86. Schiffmann SN, Fisone G, Moresco R, Cunha RA, Ferré S. (2007) Adenosine A2A receptors and basal ganglia physiology. Prog Neurobiol. 83(5):277-92. Schiffmann SN, Lledo PM, Vincent JD. (1995) Dopamine D1 receptor modulates the voltage-gated sodium current in rat striatal neurones through a protein kinase A. J Physiol. 483(Pt 1):95-107. Schiffmann SN, Vanderhaeghen JJ. (1993) Adenosine A2 receptors regulate the gene expression of striatopallidal and striatonigral neurons. Neurosci. 13(3):1080-7. Schwarzer C, Berresheim U, Pirker S, Wieselthaler A, Fuchs K, Sieghart W, Sperk G. (2001) Distribution of the major gamma-aminobutyric acid(A) receptor subunits in the 192 basal ganglia and associated limbic brain areas of the adult rat. J Comp Neurol. 433(4):526-49. Semyanov A, Walker MC, Kullmann DM, Silver RA. (2004) Tonically active GABA A receptors: modulating gain and maintaining the tone. Trends Neurosci. 27(5):262-9. Shen W, Hamilton SE, Nathanson NM, Surmeier DJ (2005) Cholinergic suppression of KCNQ channel currents enhances excitability of striatal medium spiny neurons. J Neurosci 25:7449–7458. Shen W, Hernandez-Lopez S, Tkatch T, Held JE, Surmeier DJ. (2004) Kv1.2-containing K+ channels regulate subthreshold excitability of striatal medium spiny neurons. J Neurophysiol. 91(3):1337-49. Shen W, Tian X, Day M, Ulrich S, Tkatch T, Nathanson NM, Surmeier DJ. (2007) Cholinergic modulation of Kir2 channels selectively elevates dendritic excitability in striatopallidal neurons. Nat Neurosci. 10(11):1458-66. Shuen JA, Chen M, Gloss B, Calakos N. (2008) Drd1a-tdTomato BAC transgenic mice for simultaneous visualization of medium spiny neurons in the direct and indirect pathways of the basal ganglia. J Neurosci.28(11):2681-5. Sieghart W, Sperk G. (2002) Subunit composition, distribution and function of GABA(A) receptor subtypes. Curr Top Med Chem. 2(8):795-816. Sigel E, Baur R. (1988) Activation of protein kinase C differentially modulates neuronal Na+, Ca2+, and gamma-aminobutyrate type A channels. Proc Natl Acad Sci U S A. 85(16):6192-6. Singer HS, Wendlandt J, Krieger M, Giuliano J. (2001) Baclofen treatment in Tourette syndrome: a double-blind, placebo-controlled, crossover trial. Neurology. 56(5):599-604. Sivakumaran S, Mohajerani MH, Cherubini E. (2009) At immature mossy-fiber-CA3 synapses, correlated presynaptic and postsynaptic activity persistently enhances GABA release and network excitability via BDNF and cAMP-dependent PKA. J Neurosci. 29(8):2637-47. Slany A, Zezula J, Fuchs K, Sieghart W (1995) Allosteric modulation of [3H]flunitrazepam binding to recombinant GABAA receptors. Eur J Pharmacol 291:99– 105. Smith KR, McAinsh K, Chen G, Arancibia-Carcamo IL, Haucke V, Yan Z, Moss SJ, Kittler JT. (2008) Regulation of inhibitory synaptic transmission by a conserved atypical interaction of GABA(A) receptor beta- and gamma-subunits with the clathrin AP2 adaptor. Neuropharmacology. 55(5):844-50. 193 Smith Y, Bennett BD, Bolam JP, Parent A, Sadikot AF. (1994) Synaptic relationships between dopaminergic afferents and cortical or thalamic input in the sensorimotor territory of the striatum in monkey. J Comp Neurol 344:1–19. Song WJ, Surmeier DJ. (1996) Voltage-dependent facilitation of calcium channels in rat neostriatal neurons. J Neurophysiol. 76(4):2290-306. Somogyi P, Fritschy JM, Benke D, Roberts JD, Sieghart W. (1996) The gamma 2 subunit of the GABAA receptor is concentrated in synaptic junctions containing the alpha 1 and beta 2/3 subunits in hippocampus, cerebellum and globus pallidus. Neuropharmacology. 35(9-10):1425-44. Somogyi P, Priestley JV, Cuello AC, Smith AD, Takagi H. (1982) Synaptic connections of enkephalin-immunoreactive nerve terminals in the neostriatum: a correlated light and electron microscopic study. J Neurocytol. 11(5):779-807. Starr PA, Rau GM, Davis V, Marks WJ Jr, Ostrem JL, Simmons D, Lindsey N, Turner RS. (2005) Spontaneous pallidal neuronal activity in human dystonia: comparison with Parkinson's disease and normal macaque. J Neurophysiol. 93(6):3165-76. Stell BM, Brickley SG, Tang CY, Farrant M, Mody I. (2003) Neuroactive steroids reduce neuronal excitability by selectively enhancing tonic inhibition mediated by delta subunitcontaining GABAA receptors. Proc Natl Acad Sci U S A. 100(24):14439-44. Stenman J, Toresson H, Campbell K. (2003) Identification of two distinct progenitor populations in the lateral ganglionic eminence: implications for striatal and olfactory bulb neurogenesis. J Neurosci. 23(1):167-74. Stoof JC, Drukarch B, de Boer P, Westerink BH. (1992) In vitro and in vivo acetylcholine release from rat striatum as a functional paradigm of signal transduction via a D-2 dopamine receptor. Neurochem Int. 20:201-205. Stoof JC, Kebabian JW. (1981) Opposing roles for D-1 and D-2 dopamine receptors in efflux of cyclic AMP from rat neostriatum. Nature. 294(5839):366-8. Stoof JC, Kebabian JW. (1984) Two dopamine receptors: biochemistry, physiology and pharmacology. Life Sci. 35(23):2281-96. Sugita S, Uchimura N, Jiang ZG, North RA. (1991) Distinct muscarinic receptors inhibit release of gamma-aminobutyric acid and excitatory amino acids in mammalian brain. Proc Natl Acad Sci U S A. 88(6):2608-11. Surmeier DJ, Bargas J, Hemmings HC Jr, Nairn AC, Greengard P. (1995) Modulation of calcium currents by a D1 dopaminergic protein kinase/phosphatase cascade in rat neostriatal neurons. Neuron. 14(2):385-97. 194 Surmeier DJ, Eberwine J, Wilson CJ, Cao Y, Stefani A, Kitai ST. (1992) Dopamine receptor subtypes colocalize in rat striatonigral neurons. Proc Natl Acad Sci U S A. 89(21):10178-82. Surmeier DJ, Song WJ, Yan Z. (1996) Coordinated expression of dopamine receptors in neostriatal medium spiny neurons. J Neurosci. 16(20):6579-91. Surmeier, D. J., Reiner, A., Levine, M. S. & Ariano, M. A. (1993) Are neostriatal dopamine receptors co-localized? Trends Neurosci. 16:299–305. Svenningsson P, Le Moine C, Kull B, Sunahara R, Bloch B, Fredholm BB. (1997) Cellular expression of adenosine A2A receptor messenger RNA in the rat central nervous system with special reference to dopamine innervated areas. Neuroscience. 80(4):117185. Svenningsson P, Lindskog M, Rognoni F, Fredholm BB, Greengard P, Fisone G. (1998) Activation of adenosine A2A and dopamine D1 receptors stimulates cyclic AMPdependent phosphorylation of DARPP-32 in distinct populations of striatal projection neurons. Neuroscience. 84(1):223-8. Takei K, Slepnev VI, Haucke V, De Camilli P. (1999) Functional partnership between amphiphysin and dynamin in clathrin-mediated endocytosis. Nat Cell Biol. 1(1):33-9. Tang X, Hernandez CC, Macdonald RL. (2010) Modulation of spontaneous and GABAevoked tonic alpha4beta3delta and alpha4beta3gamma2L GABAA receptor currents by protein kinase A. J Neurophysiol. 103(2):1007-19. Taverna S, Canciani B, Pennartz CM. (2007) Membrane properties and synaptic connectivity of fast-spiking interneurons in rat ventral striatum. Brain Res. 1152:49-56. Taverna S, Ilijic E, Surmeier DJ. (2008) Recurrent collateral connections of striatal medium spiny neurons are disrupted in models of Parkinson's disease. J Neurosci. 28(21):5504-12. Taverna S, van Dongen YC, Groenewegen HJ, Pennartz CM. (2004) Direct physiological evidence for synaptic connectivity between medium-sized spiny neurons in rat nucleus accumbens in situ. J Neurophysiol. 91(3):1111-21. Tecuapetla F, Carrillo-Reid L, Bargas J, Galarraga E. (2007) Dopaminergic modulation of short-term synaptic plasticity at striatal inhibitory synapses. Proc Natl Acad Sci U S A. 104(24):10258-63. Tecuapetla F, Koós T, Tepper JM, Kabbani N, Yeckel MF. (2009) Differential dopaminergic modulation of neostriatal synaptic connections of striatopallidal axon collaterals. J Neurosci. 29(28):8977-90. 195 Tepper JM, Bolam JP. (2004) Functional diversity and specificity of neostriatal interneurons. Curr Opin Neurobiol. 14(6):685-92. Tepper JM, Koós T, Wilson CJ. (2004) GABAergic microcircuits in the neostriatum. Trends Neurosci. 27(11):662-9. Tepper JM, Wilson CJ, Koós T. (2008) Feedforward and feedback inhibition in neostriatal GABAergic spiny neurons. Brain Res Rev. 58(2):272-81. Terunuma M, Jang IS, Ha SH, Kittler JT, Kanematsu T, Jovanovic JN, Nakayama KI, Akaike N, Ryu SH, Moss SJ, Hirata M. (2004) GABAA receptor phospho-dependent modulation is regulated by phospholipase C-related inactive protein type 1, a novel protein phosphatase 1 anchoring protein. J Neurosci. 24(32):7074-84. Terunuma M, Xu J, Vithlani M, Sieghart W, Kittler J, Pangalos M, Haydon PG, Coulter DA, Moss SJ. (2008) Deficits in phosphorylation of GABA(A) receptors by intimately associated protein kinase C activity underlie compromised synaptic inhibition during status epilepticus. Neurosci. 28(2):376-84. Thomas P, Mortensen M, Hosie AM, Smart TG. (2005) Dynamic mobility of functional GABAA receptors at inhibitory synapses. Nat Neurosci. 8(7):889-97. Tossman U, Segovia J, Ungerstedt U. (1986) Extracellular levels of amino acids in striatum and globus pallidus of 6-hydroxydopamine-lesioned rats measured with microdialysis. Acta Physiol Scand. 127(4):547-51. Tretter V, Ehya N, Fuchs K, Sieghart W. (1997) Stoichiometry and assembly of a recombinant GABAA receptor subtype. J Neurosci. 17(8):2728-37. Tunstall MJ, Oorschot DE, Kean A, Wickens JR. (2002) Inhibitory interactions between spiny projection neurons in the rat striatum. J Neurophysiol. 88(3):1263-9. Uchimura N, Higashi H, Nishi S. (1986) Hyperpolarizing and depolarizing actions of dopamine via D-1 and D-2 receptors on nucleus accumbens neurons. Brain Res. 375(2):368-72. Urak L, Feucht M, Fathi N, Hornik K, Fuchs K. (2006) A GABRB3 promoter haplotype associated with childhood absence epilepsy impairs transcriptional activity. Hum Mol Genet. 15(16):2533-41. van Rijnsoever C, Sidler C, Fritschy JM. (2005) Internalized GABA-receptor subunits are transferred to an intracellular pool associated with the postsynaptic density. Eur J Neurosci. 21(2):327-38. Venance L, Glowinski J, Giaume C. (2004) Electrical and chemical transmission between striatal GABAergic output neurones in rat brain slices. J Physiol. 559(1):215-30. 196 Veruki ML, Yeh HH. (1994) Vasoactive intestinal polypeptide modulates GABAA receptor function through activation of cyclic AMP. Vis Neurosci. 11(5):899-908. Voorn P, Vanderschuren LJ, Groenewegen HJ, Robbins TW, Pennartz CM. (2004) Putting a spin on the dorsal-ventral divide of the striatum. Trends Neurosci. 27(8):46874. Vuillet J, Kerkerian L, Salin P, Nieoullon A. (1989) Ultrastructural features of NPYcontaining neurons in the rat striatum. Brain Res. 477(1-2):241-51. Wafford KA, Bain CJ, Quirk K, McKernan RM, Wingrove PB, Whiting PJ, Kemp JA. A novel allosteric modulatory site on the GABAA receptor beta subunit. (1994) Neuron. 12(4):775-82. Walaas SI, Greengard P. (1984) DARPP-32, a dopamine- and adenosine 3':5'monophosphate-regulated phosphoprotein enriched in dopamine-innervated brain regions. I. Regional and cellular distribution in the rat brain. J Neurosci. 4(1):84-98. Walaas SI, Greengard P. (1991) Protein phosphorylation and neuronal function. Pharmacol Rev. 43(3):299-349. Waldvogel HJ, Kubota Y, Trevallyan SC, Kawaguchi Y, Fritschy JM, Mohler H, Faull RL. (1997) The morphological and chemical characteristics of striatal neurons immunoreactive for the alpha1-subunit of the GABA(A) receptor in the rat. Neuroscience. 80(3):775-92. Wall MJ, Usowicz MM. (1997) Development of action potential-dependent and independent spontaneous GABAA receptor-mediated currents in granule cells of postnatal rat cerebellum. Eur J Neurosci. 9(3):533-48. Wei W, Zhang N, Peng Z, Houser CR, Mody I. (2003) Perisynaptic localization of delta subunit-containing GABA(A) receptors and their activation by GABA spillover in the mouse dentate gyrus. J Neurosci. 23(33):10650-61. Wilson CJ, Chang HT, Kitai ST. (1990) Firing patterns and synaptic potentials of identified giant aspiny interneurons in the rat neostriatum. J Neurosci. 10(2):508-19. Wilson CJ, Groves PM. (1980) Fine structure and synaptic connections of the common spiny neuron of the rat neostriatum: a study employing intracellular inject of horseradish peroxidase. J Comp Neurol. 194(3):599-615. Yan Z, Flores-Hernandez J, Surmeier DJ. (2001) Coordinated expression of muscarinic receptor messenger RNAs in striatal medium spiny neurons. Neuroscience. 103(4):101724. 197 Yan Z, Surmeier DJ. (1997) D5 dopamine receptors enhance Zn2+-sensitive GABA(A) currents in striatal cholinergic interneurons through a PKA/PP1 cascade. Neuron. 19(5):1115-26. Yasumoto S, Tanaka E, Hattori G, Maeda H, Higashi H. (2002) Direct and indirect actions of dopamine on the membrane potential in medium spiny neurons of the mouse neostriatum. J Neurophysiol. 87(3):1234-43. Yung KK, Smith AD, Levey AI, Bolam JP. (1996) Synaptic connections between spiny neurons of the direct and indirect pathways in the neostriatum of the rat: evidence from dopamine receptor and neuropeptide immunostaining. Eur J Neurosci. 8(5):861-9. Zhang XF, Hu XT, White FJ. (1998) Whole-cell plasticity in cocaine withdrawal: reduced sodium currents in nucleus accumbens neurons. J Neurosci. 18(1):488-98. Zhou FM, Wilson CJ, Dani JA. (2002) Cholinergic interneuron characteristics and nicotinic properties in the striatum. J Neurobiol. 53(4):590-605. 198