Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Protein domain wikipedia , lookup
Immunoprecipitation wikipedia , lookup
List of types of proteins wikipedia , lookup
Nuclear magnetic resonance spectroscopy of proteins wikipedia , lookup
Western blot wikipedia , lookup
Structural alignment wikipedia , lookup
Circular dichroism wikipedia , lookup
Protein structure prediction wikipedia , lookup
Protein purification wikipedia , lookup
G protein–coupled receptor wikipedia , lookup
ATP-binding cassette transporter wikipedia , lookup
Protein–protein interaction wikipedia , lookup
Intrinsically disordered proteins wikipedia , lookup
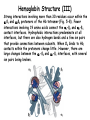
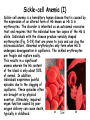
Chap. 5B Protein Function Topics • Reversible Binding of a Protein to a Ligand: Oxygen-binding Proteins • The heme group • Myoglobin • Hemoglobin • Sickle-cell Anemia Fig. 5-19. Normal (a) and sickle-cell anemia (b) erythrocytes. Erythrocytes, Hb, and Oxygen Transport Nearly all of the O2 carried by whole blood in animals is bound and transported by hemoglobin (Hb) in erythrocytes (red blood cells). Erythrocytes lack a nucleus and organelles such as mitochondria and have a half-life in the bloodstream of 120 days. Their main function is to carry Hb, which is dissolved in the cytosol at a very high concentration of ~34% by weight. Erythrocyte Hb is about 96% saturated with O2 in the lungs. In venous blood, erythrocyte Hb is 64% saturated. Thus about a third of the O2 carried by erythrocytes is released to peripheral tissues during one pass through the circulation. The binding of O2 to Hb occurs with less affinity than to Mb. Thus, Hb is more suited for delivery of O2 to tissues. Furthermore, ligands such as H+ and CO2, which are in relatively higher concentrations in the peripheral circulation, improve the release of O2 from Hb and its delivery to peripheral tissues. Hemoglobin Structure (I) Hb (Mr 64,500) is a tetrameric protein that is roughly spherical and has a diameter of about 5.5 nm. Each subunit contains a bound heme group that has the same structure as the heme of myoglobin. There are two types of chains in adult Hb, two chains (141 residues each), and two ß chains (146 residues each). Although fewer than half of the amino acid residues in the two chains are identical, the structures of the and ß chains are nearly superimposable. Furthermore their structures are very similar to that of myoglobin (Fig. 5-6), which has a more dissimilar amino acid sequence. Note that the helix naming conventions are the same for the Mb and Hb chains, except that the subunit lacks the short D helix. Like Mb, the heme-binding pocket is made up of the E and F helices in the and ß chains of Hb. Hemoglobin Structure (II) Despite the fact that the amino acid sequences of the three polypeptides are identical at only 27 positions (Fig. 5-7), the structures of the myoglobin and Hb chains are nearly identical. This illustrates how similar amino acids can form similar secondary and tertiary structures. Hemoglobin Structure (III) Strong interactions involving more than 30 residues occur within the 1ß1 and 2ß2 protomers of the Hb tetramer (Fig. 5-8). Fewer interactions involving 19 amino acids connect the 1-ß2 and 2-ß1 contact interfaces. Hydrophobic interactions predominate at all interfaces, but there are also hydrogen bonds and a few ion pairs that provide connections between subunits. When O2 binds to Hb, contacts within the protomers change little. However, there are large changes between the 1-ß2 and 2-ß1 interfaces, with several ion pairs being broken. Structural Changes in Hb on O2 Binding (I) X-ray diffraction studies revealed that Hb transitions between two major conformational states depending on whether oxygen is present or not. These are the R state (relaxed) and T state (tense). Although oxygen binds to Hb in either structural state, O2 binding stabilizes and favors the R state. In the absence of O2, the T state is more stable and is the predominant conformation. The two states are also referred to as oxy- and deoxyhemoglobin, respectively. The terms tense and relaxed were originally coined because the T state is stabilized by a greater number of ion pairs at the 1-ß2 and 2-ß1 interfaces than are present in the R state. Some of these T state interactions are shown in Fig. 5-9 a, and more are shown in Fig. 5-9 b (next slide). Structural Changes in Hb on O2 Binding (II) One important contact occurring between the 1-ß2 and 2-ß1 interfaces in the T state is the His HC3 to Lys C5 -carboxyl group/-amino group interaction (Fig. 5-9 a&b). His HC3 is the Cterminal residue in the ß subunits. His HC3 of the ß subunits also forms an interaction between its side-chain and Asp FG1 within the ß subunits. As shown in the next slide, the His HC3 to Lys C5 -carboxyl group/-amino group interaction is broken on binding of O2. Structural Changes in Hb on O2 Binding (III) The binding of O2 to a Hb subunit in the T state triggers a change in conformation to the R state. When the entire protein undergoes this transition, the structures of the individual subunits change little. However, the aß protomers slide past each other and rotate, narrowing the pocket between the ß subunits (Fig. 5-10). A number of contacts that stabilize the T state are broken and new ones are formed. The His HC3 to Lys C5 contact is one of the ones that is broken in the T R transition. As shown in Fig. 5-10, the His HC3 residues at the C-termini of the ß subunits rotate in the R state towards the center of the molecule where they are no longer involved in ionic interactions to the subunits. The size of the pocket between the ß chains also narrows as as a result of the T R transition. Structural Changes in Hb on O2 Binding (IV) The structural changes that occur at the 1-ß2 and 2-ß1 interfaces on O2 binding are ultimately triggered by movement of the proximal histidines, His F8, of the four subunits when O2 binds to the heme groups (Fig. 5-11). In the T state, the porphyrin is slightly puckered, causing the heme iron to protrude on the side where the proximal histidine is located. The binding of O2 causes the heme to assume a more planar conformation, shifting the position of the proximal His and the attached F helix in the R state. These movements lead to changes in the ion pairs at the 1-ß2 and 2-ß1 interfaces. The Hb O2 Binding Curve Hb must bind O2 efficiently in the lungs, where pO2 is about 13.3 kPa, and release O2 in the tissues, where pO2 is about 4 kPa. Myoglobin, or any protein that binds O2 with a hyperbolic binding curve, would be ill-suited to this function. For example, a protein such as myoglobin that binds O2 with high affinity would bind it efficiently in the lungs, but would not release much of it in the tissues. On the other hand if the protein bound O2 with sufficiently low affinity to release it in the tissues, it would not pick up much O2 in the lungs. Hb solves the problem by undergoing a structural transition from a low-affinity T state to a high-affinity R state as more O2 molecules are bound. As a result, Hb has a hybrid S-shaped, or sigmoid, binding curve for O2 (Fig. 512). A sigmoid curve can be viewed as a hybrid curve reflecting the transition between low- and high-affinity structural states in Hb on O2 binding. Sigmoidal ligand binding curves are indicative of cooperative binding of a ligand to a protein. In cooperative binding, the binding of one ligand to a protein alters its binding affinity for subsequent ligands. Cooperative Ligand Binding to Allosteric Proteins (I) Proteins such as Hb, in which structural transitions occur due to ligand binding and affect ligand-binding affinity, are called allosteric (Greek for “other shape”) proteins. The ligands themselves are broadly called structural modulators. Modulators for allosteric proteins can be either inhibitors or activators. When the normal ligand and modulator are identical as they are with Hb (i.e., O2), the modulator is termed homotropic. If the modulator is a molecule other than the normal ligand, the interaction is heterotropic. Some proteins can have two or more modulators, and therefore can have both homotropic and heterotropic interactions. As a result of allosteric transitions in the packing of Hb subunits, Hb binds O2 cooperatively. Cooperative binding confers a much more sensitive response to ligand concentration. It is important to the function of many multisubunit proteins. The principle of allostery extends readily to regulatory enzymes, as covered in Chap. 6. Cooperative Ligand Binding to Allosteric Proteins (II) Cooperative conformational changes generally depend on variations in the structural stability of different parts of a protein. The binding sites of an allosteric protein typically consist of stable segments in proximity to relatively unstable segments, with the later capable of frequent changes in conformation or intrinsic disorder (Fig. 5-13). When a ligand binds, the moving parts of the protein’s binding site may be stabilized in a particular conformation, affecting the conformation of adjacent polypeptide subunits. The conformational changes that occur as the ligand binds can convert the protein from a low- to a highaffinity state, which is a form of induced fit. The degree of cooperativity in ligand binding to an allosteric protein can be calculated using the Hill equation (not covered). Models for Cooperative O2 Binding to Hb (I) Although much is known about the structures of the T and R states of Hb, the mechanism by which this structural transition occurs on sequential ligand binding still is unsolved. Two principle models for cooperative binding of O2 to Hb (and ligand binding to any allosteric protein) are widely used to explain structural transitions. In the Monod, Wyman, and Changeux (MWC, or concerted) model, all subunits of Hb undergo the transition from one conformation to the other simultaneously (Fig. 5-15a). No tetramer has individual subunits in different conformations, and the two conformations are in equilibrium. The ligand can bind to either conformation, but binds each with different affinity. Successive binding of ligand molecules to the low-affinity conformation (which is more stable in the absence of ligand) makes the transition to the high-affinity conformation more likely. Models for Cooperative O2 Binding to Hb (II) In the second model, known as the sequential model proposed by Koshland (Fig. 5-15b), ligand binding can induce a change of conformation in an individual subunit. A conformational change in one subunit makes a similar change in a adjacent subunit, as well as the binding of a second ligand molecule, more likely. There are more potential intermediate states in the sequential model than in the concerted model. The two models are not mutually exclusive: the concerted model may be viewed as the all-or-none limiting case of the sequential model. Transport of H+ and CO2 by Hb (I) Hb binds to and transports about 40% of the total H+ and 15% to 20% of the CO2 formed in peripheral tissues to the lungs and kidneys. The remainder of the H+ is absorbed by the plasma’s bicarbonate buffer system. The remainder of the CO2 is transported as dissolved HCO3- and CO2. [Note that the solubility of CO2 in the blood is increased by the carbonic anhydrase reaction (CO2 + H2O H+ + HCO3-) which occurs in erythrocytes.] The binding of H+ and CO2 to Hb decreases the affinity of Hb for O2, favoring the release of O2 to the tissues where the concentrations of these components are relatively high. Conversely, in the capillaries of the lung, as CO2 is excreted and the blood pH consequently rises, the affinity of Hb for O2 increases and the protein binds more O2 for transport to the peripheral tissues. The effect of pH and CO2 concentration on the binding and release of O2 by Hb is known as the Bohr effect. The effect of pH on Hb O2 binding curves is shown in Fig. 5-16. Transport of H+ and CO2 by Hb (II) H+ binds to several amino acid side-chains in Hb. His HC3 at the C-termini of the ß chains makes a major contribution to H+ binding. When His HC3 is protonated, it forms a salt bridge to Asp FG1 that helps stabilize deoxyhemoglobin in the T state. The ion pair stabilizes the protonated form of His HC3, giving this residue an abnormally high pKa in the T state. The proton is released from His HC3 in the lungs (pH 7.6) when the binding of O2 to Hb drives the conformation to the R state. Carbon dioxide binds as a carbamate group to the -amino groups at the N-terminal ends of each globin chain, forming carbaminohemoglobin (see below). This reaction produces H+, contributing to the Bohr effect. The bound carbamates also form additional salt bridges that help to stabilize the T state and promote the release of O2. When Hb reaches the lungs, the high O2 concentration promotes binding of O2 and the release of bound CO2. BPG Regulation of O2 Binding to Hb (I) 2,3-bisphosphoglycerate (BPG), is a negative heterotropic modulator of O2 binding to Hb. BPG is important in physiological adaptation to the lower pO2 values that are present at high elevations. As discussed below, BPG binds to and favors the formation of deoxyhemoglobin (the T state). This facilitates O2 delivery to tissues. Synthesis of BPB in erythrocytes increases when an individual moves to higher elevation. BPG Regulation of O2 Binding to Hb (II) As shown in Fig. 5-17, the BPG concentration in normal human blood is about 5 mM at sea level and about 8 mM at high elevations (e.g., 4,500 m). Thus, at sea level, Hb is nearly saturated with O2 in the lungs, but is just over 60% saturated in the tissues, so the amount of O2 released in the tissues is about 38% of the maximum carried in the blood. At high elevations, O2 delivery declines about one-fourth to 30% of maximum in the absence of increased BPG production. When BPG synthesis increases after time spent at high elevation, the affinity of Hb for O2 decreases, so approximately 37% of what can be carried is again delivered to the tissues. Note that Hb binds very tightly to O2 in the absence of BPG (curve at left). BPG Regulation of O2 Binding to Hb (III) One molecule of BPG binds to Hb in the cavity between the ß subunits that is present in the T state (Fig. 5-18a). This cavity is lined with positively charged amino acid R groups that interact with the negatively charged groups of BPG. The binding site is absent in the R state, precluding BPG binding (Fig. 5-18b). BPG lowers Hb’s affinity for O2 by stabilizing the T state. Thus, BPG favors the release of O2 from Hb and increases its delivery to peripheral tissues. Interestingly, BPG does not bind to fetal Hb which has a subunit composition of 22. (Note that the subunit is expressed instead of the ß subunit during the last two trimesters of fetal life.) The subunit lacks the basic residues that are needed for BPG binding to Hb. For this reason, fetal Hb has a higher affinity for O2 than maternal Hb, and transfer of O2 from the maternal to the fetal circulation in the placenta is facilitated. Carbon Monoxide Poisoning Carbon monoxide (CO) is a colorless, odorless gas that is responsible for more than half of the annual poisoning deaths worldwide. CO has a 250-fold greater affinity for Hb than O2, and exposure to CO reduces the oxygen-carrying capacity of the blood. Symptoms of CO poisoning depend on the percent of total Hb that is bound to the gas. At 10% bound, symptoms are rarely observed. At 20% to 30%, the individual will experience a severe headache accompanied by nausea, dizziness, confusion, and visual disturbances. At CO Hb levels of 30% to 50%, neurological symptoms become more severe, and at levels near 50%, the individual loses consciousness and can fall into a coma. Respiratory failure may follow. Death normally occurs rapidly when CO Hb levels rise above 60%. CO is a component of tobacco smoke, and chain-smokers can have CO Hb levels of 15%. Thus, smoking predisposes individuals to the effects of CO poisoning. In addition, the fetus of a pregnant woman is highly susceptible because fetal Hb has a higher affinity for CO than maternal Hb. As CO binds to one or two subunits of a Hb tetramer, the affinity of the remaining subunits for O2 is increased substantially. Thus a Hb tetramer with two bound CO molecules can efficiently bind O2 in the lungs; however it releases O2 very inefficiently in the peripheral tissues. Poisoning is further exacerbated by the binding of CO to cytochromes of the mitochondrial electron transport chain. Sickle-cell Anemia (I) Sickle-cell anemia is a hereditary human disease that is caused by the expression of an altered form of Hb known as Hb S in erythrocytes. The disorder is inherited as an autosomal recessive trait and requires that the individual have two copies of the Hb S allele. Individuals with the disease produce variably shaped erythrocytes (Fig. 5-19) that are prone to lysis and can clog the microvasculature. Abnormal erythrocytes only form when Hb S undergoes deoxygenation in capillaries. The sickled erythrocytes are fragile and rupture easily. This results in a significant anemia wherein the Hb content of the blood is only about 50% of normal. In addition, individuals experience painful episodes due to the clogging of capillaries. These episodes often are brought on by physical exertion. Ultimately, impaired organ function caused by poor oxygen delivery can cause death, typically in childhood. Sickle-cell Anemia (II) The abnormal structure of sickle-cell erythrocytes is caused by subtle differences in the conformation of sickle-cell Hb (Hb S) (Fig. 5-20). Namely, the substitution of Val for Glu at position 6 of the ß chains creates a sticky hydrophobic contact point at this site which is located on the outer surface of the Hb S molecules. These sticky spots cause deoxyhemoglobin S molecules to associate abnormally with each other, forming long fibrous aggregates (Fig. 5-20 b). These fibers distort the shape of erythrocytes that contain them. Individuals who inherit only one Hb S allele, and are thus heterozygous, experience a milder medical condition known as sickle-cell trait. Only about 1% of their erythrocytes become sickled on deoxygenation. These individuals may live completely normal lives if they avoid vigorous exercise and other stresses on the circulatory system. The Hb S allele is most prevalent in people of African descent. Heterozygotes who express a single copy of the Hb S allele have some protection against malaria. For this reason, the allele is prevalent in this population group.