Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Electrophysiology wikipedia , lookup
Single-unit recording wikipedia , lookup
Caridoid escape reaction wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Optogenetics wikipedia , lookup
Dendritic spine wikipedia , lookup
Brain-derived neurotrophic factor wikipedia , lookup
Neuroanatomy wikipedia , lookup
Biological neuron model wikipedia , lookup
Axon guidance wikipedia , lookup
Apical dendrite wikipedia , lookup
Development of the nervous system wikipedia , lookup
Long-term potentiation wikipedia , lookup
Nervous system network models wikipedia , lookup
Pre-Bötzinger complex wikipedia , lookup
End-plate potential wikipedia , lookup
De novo protein synthesis theory of memory formation wikipedia , lookup
Nonsynaptic plasticity wikipedia , lookup
Synaptic gating wikipedia , lookup
Activity-dependent plasticity wikipedia , lookup
Signal transduction wikipedia , lookup
Endocannabinoid system wikipedia , lookup
Long-term depression wikipedia , lookup
Stimulus (physiology) wikipedia , lookup
NMDA receptor wikipedia , lookup
Neuromuscular junction wikipedia , lookup
Neurotransmitter wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Molecular neuroscience wikipedia , lookup
Synaptogenesis wikipedia , lookup
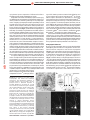
© 2001 Nature Publishing Group http://neurosci.nature.com © 2001 Nature Publishing Group http://neurosci.nature.com review Molecular heterogeneity of central synapses: afferent and target regulation Ann Marie Craig and Hélène Boudin Department of Anatomy and Neurobiology, Washington University School of Medicine, 660 S. Euclid, Campus Box 8108, 958 McDonnell Sciences Building, St. Louis, Missouri 63110, USA Correspondence should be addressed to A.M.C. ([email protected]) Electrophysiological recordings show a functional spectrum even within a single class of synapse, with individual synapses ranging widely in fundamental properties, including release probability, unitary response and effects of previous stimulation on subsequent response. Molecular and cellular biological approaches have shown a corresponding diversity in the complement of ion channels, receptors, scaffolds and signal transducing proteins that make up individual synapses. Indeed, we believe that each individual synapse is unique, a function of presynaptic cell type, postsynaptic cell type, environment, developmental stage and history of activity. We review here the molecular diversity of glutamatergic and GABAergic synapses in the mammalian brain in the context of potential cell biological mechanisms that may explain how individual cells develop and maintain such a mosaic of synaptic connections. The mammalian brain possesses a wonderful diversity and richness of synaptic connections. Machinery for regulated neurotransmitter release and response must be present at all synapses. However, from the earliest morphological studies classifying brain synapses into Gray’s type I and type II (ref. 1), it has been clear that all synapses are not equivalent. Synaptic composition is initially determined and continuously modified by ongoing bidirectional communication between presynaptic and postsynaptic partners. Nevertheless, individual features of synapses can sometimes result from a dominating influence, and it seems useful to consider major classes of cellular influences on synaptic composition (Fig. 1). One pattern of synaptic protein localization is ubiquitous expression and distribution (Fig. 1a). A few molecules, particularly some that are involved in synaptic vesicle fusion, are expressed by all neurons and distributed to all synapses. An alternative is cell-specific expression (Fig. 1b), in which molecular components of synapses are selectively transcribed and/or translated only in certain subsets of neurons. In the simplest form of heterogeneity, these components are targeted to all synapses made by the expressing neurons. Another pattern reflects the influence of presynaptic terminals on postsynaptic development. For example, glutamatergic versus GABAergic neurotransmitter phenotype is a feature of presynaptic neuron development that occurs independent of target contact. Thus, the precise localization of glutamate receptors in dendrites opposite glutamatergic terminals and GABA receptors in dendrites opposite GABAergic terminals must require some form of anterograde communication (Fig. 1c). Influences from the presynaptic cell determine not only the chemical nature of the receptive field but often also the complement of specific receptor subtypes and associated signaling proteins (Fig. 1d). Postsynaptic events can also influence presynaptic development (Fig. 1e). The target cell can strongly influence the molecular composition and release properties of individual nature neuroscience • volume 4 no 6 • june 2001 presynaptic sites of a given axon, a phenomenon that clearly requires retrograde signaling. Distribution of some proteins is regulated by cellular domain (Fig. 1f). Some aspects of synaptic composition are determined by synapse location on the cell, and this in turn can be determined intrinsic features of the neuron. Thus certain domains of axons or dendrites are permissive or instructive for specific classes or features of synapses. Finally, heterogeneity also occurs within one synapse type (Fig. 1g). Activity dynamically regulates the density of receptors and other signaling components at postsynaptic sites. Such regulation of AMPA-type receptors has been a focus of study as a mechanism underlying long-term changes in synaptic efficacy. Surprisingly, the molecular composition and release and response properties made by an individual axon onto an individual dendrite are still quite heterogeneous (Fig. 1h). Thus, some element of randomness, perhaps in addition to geometrical influences, is required to explain such heterogeneity even among autapses of an isolated neuron. Ubiquitous expression and distribution Surprisingly few proteins are common to all synapses. Indeed, we can think of no postsynaptic or cleft components that fit this category, although much of the machinery of synaptic vesicle fusion is conserved among synapse types. The v-SNARE VAMP-2/synaptobrevin II, t-SNARES syntaxin 1A and 1B and SNAP-25, calcium sensor synaptotagmin I, and regulatory proteins rab3A, SV2A,B, Munc 13-1 and Munc18-1 are widely expressed and function in all forebrain terminals2 (Fig. 2). However, even these core regulators of vesicle fusion are not ubiquitous. First, different isoforms of many of these proteins are expressed in other brain regions. VAMP-1 and synatotagmin II are expressed instead of VAMP-2 and synaptotagmin I in many parts of the brainstem and spinal cord3,4, and ribbon synapses in the retina contain syntaxin 3 instead of syntaxin 1 (ref. 5). Sec569 © 2001 Nature Publishing Group http://neurosci.nature.com review cell a Pyramidal (glutamatergic) Interneuron (GABAergic) Dendrite Axon Presynaptic protein Postsynaptic protein © 2001 Nature Publishing Group http://neurosci.nature.com Glu-R: Glutamate receptor gr.III-mGluR: group III metabotropic glutamate receptor GABA-R: GABA receptor Ubiquitous expression and distribution (presynaptic protein) b Cell specific expression: presynaptic protein c Glu-R GABA-R Cell specific expression: postsynaptic protein d Presynaptic influences postsynaptic: transmitter specificity (postsynaptic protein) e gr.III-mGluR Glu-R Presynaptic influences postsynaptic: glutamatergic synapses (postsynaptic protein) f GABA-R Regulation by cellular domain (postsynaptic protein) Postsynaptic influences presynaptic (presynaptic protein) g Glu-R h Heterogeneity within one synapse type: activity regulation (postsynaptic protein) Autaptic heterogeneity (post- and presynaptic proteins) ond, related isoforms of several of these proteins are also expressed in forebrain, although they may be functionally nonredundant. Hippocampal pyramidal neurons express several synaptotagmin isoforms including high levels of synaptotagmin III (ref. 3), but none can rescue the fast component of calciumdependent release that is abolished by deletion of synaptotagmin I (ref. 6). Presynaptic cytomatrix organizing proteins have not been characterized as fully; current evidence suggests they are also widely expressed, but not ubiquitous. For example, bassoon and piccolo are present at all forebrain glutamatergic and GABAergic synapses but not at the neuromuscular junction7. Cell-specific expression The simplest mechanism underlying synaptic heterogeneity is cell-type specific expression of a protein, and targeting of that protein to all presynaptic or postsynaptic sites made by a cell 570 Fig. 1. Cellular mechanisms for the generation of synaptic heterogeneity. Model cell pairs are drawn to show how the molecular composition of synapses might be influenced by the presynaptic and postsynaptic partners. The presence of a protein at a synapse is indicated by a red or blue circle (presynaptic proteins), or a red or blue square (postsynaptic proteins). Expression of a protein is indicated by color fill of the soma. (a) Ubiquitous expression and distribution. (b) Cell-specific expression. (c) Presynaptic terminal influences postsynaptic: transmitter specificity. (d) Presynaptic terminal influences postsynaptic: glutamatergic synapses. (e) Postsynaptic influences presynaptic. (f) Regulation by cellular domain. (g) Heterogeneity within one synapse type: activity regulation. (h) Autaptic heterogeneity. (Fig. 1b). On the presynaptic side, neurotransmitter synthetic enzymes and vesicular and plasma membrane transporters that determine the chemical nature of a synapse generally fall into this category. The GABA synthetic enzyme glutamic acid decarboxylase (GAD65 and GAD67), the vesicular inhibitory amino acid transporter responsible for loading GABA into vesicles (VIAAT/VGAT), and the major plasma membrane transporter responsible for reuptake of GABA (GAT-1) are found at symmetric but not asymmetric synapses8–11. In situ hybridization studies indicate this is due primarily to expression by GABAergic but not glutamatergic neurons9,12,13. Direct evidence for targeting of GAD to all presynaptic terminals of expressing cells has been shown by colocalization of GAD and synaptophysin immunofluorescence puncta in isolated neuron cultures14. Glutamatergic phenotype seems to be mainly conferred by expression of the vesicular glutamate transporter BNPI (refs. 15, 16). Exogenous expression of BNPI in GABAergic neurons resulted in release of both glutamate and GABA. Thus, expression of a small number of genes or even a single gene can determine presynaptic transmitter phenotype. There are many instances of cell-type-specific expression of postsynaptic components, resulting in differential signaling from a single axon to individual targets expressing a different complement of receptors or synaptic signaling molecules. Selective gene expression in pyramidal neurons versus interneurons of the hippocampus is thought to underlie major differences in properties of synaptic transmission and plasticity at glutamatergic synapses onto these two types of target cell. Whereas CA1 pyramidal neurons undergo homosynaptic long-term potentiation (LTP) in response to high-frequency stimulation of CA3 afferents, interneurons exhibit long-term depression (LTD) of the activated and non-activated synapses with the same stimulation method17. A major contributing factor to the absence of LTP in the interneurons may be the absence of the calcium-activated kinase CaMKIIα. CaMKIIα is highly expressed by pyramidal neurons, concentrated at spiny synapses, and required for LTP in pyramidal neurons18, but it is not expressed by interneurons19. Activated CaMKII enhances synaptic delivery of GluR1 (ref. 20) and potentiates AMPA receptor function by direct phosphorylation of GluR1 (refs. 21, 22). Other differences that likely contribute to differential signaling include higher levels of AMPA receptor, the postsynaptic scaffolding protein GKAP, and the activated Rho target citron, and lower levels or an absence of the calcium-activated phosphatase calcineurin, the synaptic Ras GTPase activating protein SynGAP, and the actin- and NMDA- receptorbinding protein α-actinin-2 in hippocampal interneurons compared with pyramidal neurons 19,23–25 . The mechanisms of localization of common components may also differ between cell types, as revealed by the greater detergent extractability and dependence on actin of synaptic AMPA receptor clusters on pyramidal neurons versus on interneurons26. nature neuroscience • volume 4 no 6 • june 2001 © 2001 Nature Publishing Group http://neurosci.nature.com review © 2001 Nature Publishing Group http://neurosci.nature.com Fig. 2. Molecular components of glutamate synapses and GABA synapses. Components are color coded according to function. Red, neurotransmitter receptors; pink, enzymes; blue, scaffolds; turquoise, cell adhesion proteins; green, vesicle fusion machinery. Gray lines, protein–protein interactions. Synaptic components not discussed in this review are not shown. Synaptobrevin II Syntaxin 1A,B SNAP-25 Synaptotagmin I Rab3A SV2A,B Munc13-1 Munc18-1 Glutamate BNPI Piccolo Bassoon Glutamate PRESYNAPTIC N-cadherin PICK1 Narp AMPAR Piccolo GABA VIAAT/VGAT Bassoon GABA mGluR7 N-cadherin Glutamate GAD Neurexin GABA NMDAR Neuroligin mGluR1 Cadherin GAT-1 NSF Cadherin GABA AR Thus, selective gene expression is a way SynGAP Stargazin PSD-95 GRIP that intrinsic features of a differentiated GRIP ABP Citron POSTSYNAPTIC neuron influence the molecular composiActinin GKAP Gephyrin Homer CaMKII tion of synaptic connections. Such intringephyrin Shank Calcineurin F-actin sic influences extend also to controlling GABA SYNAPSE synaptic morphology. Excitatory synapses GLUTAMATE SYNAPSE on hippocampal pyramidal neurons occur primarily on dendritic spines, whereas excitatory synapses on the interneuron dendrites are on shafts. tional pairing of presynaptic and postsynaptic elements: GABAA This morphological difference may be determined at least parreceptors are concentrated along with glutamate receptors at glutially by differential expression of actin regulatory proteins, and tamatergic mossy fiber synapses onto cerebellar granule cells and has major consequences for postsynaptic calcium dynamics27. are thought to be activated by GABA spillover from nearby synapses39. Group I metabotropic glutamate receptors are conCell-type regulation of postsynaptic architecture may be a widespread phenomenon; for example, inhibitory Renshaw cells show centrated at GABAergic postsynaptic specializations in monkey larger and more complex glycinergic postsynaptic specializations pallidus40; glycine receptors are concentrated adjacent to acetylthan neighboring motoneurons28. choline receptors at cholinergic synapses on chick ciliary ganglion neurons and can be activated by non-vesicular glycine release41. Presynaptic glutamic acid decarboxylase, which conPresynaptic influences: transmitter specificity verts glutamate to GABA and is generally specific for GABAerAn obvious and ubiquitous type of afferent-specific differentiagic terminals, is surprisingly required for development of the tion is the chemical matching of postsynaptic receptor with transpostsynaptic glutamate receptor field at the Drosophila glutamitter released (Fig. 1c). Immunoelectron microscopy shows that matergic neuromuscular junction42. These observations of misreceptors cluster only at a subset of postsynaptic sites, GABAA and glycine receptors generally at symmetric synapses29,30 and AMPA matched and unconventional appositions raise the idea that synapse development may occur by initial formation of non-speand NMDA glutamate receptors at asymmetric synapses31,32. Doucific appositions followed by selective maintenance of physioble-label immunofluorescence of hippocampal neurons in lowlogically appropriate appositions and destabilization of density culture shows that GABA receptors cluster selectively inappropriate appositions36. This model would imply an active opposite GABAergic terminals, whereas AMPA receptors cluster 33 opposite glutamatergic terminals . Thus, mechanisms must exist role of both presynaptic and postsynaptic partners. An attractive idea is that activity, by means of release of transfor targeting GABA versus glutamate receptors to separate postmitter and activation of the appropriate receptor, may be a key synaptic sites, often a few microns apart on a single dendrite. signal for synaptogenesis or synapse stabilization. Indeed, for Because neurons develop glutamatergic versus GABAergic glycinergic synapses, receptor activation is required for receptor phenotypes in the absence of target contact, anterograde signals stabilization at synapses 43,44. However, although one study must be involved in matching receptor to transmitter type. The role of presynaptic innervation was directly demonstrated by culreported that activation of AMPA receptors is required for mainture of purified motoneurons alone or with GABAergic innertenance of dendritic spines45, all studies of receptor distributions vation: isolated motoneurons lacked synaptic GABAA receptor by immunocytochemistry, calcium imaging or functional mapping report that postsynaptic clusters of GABAA and NMDAclusters, but contact with GABAergic axons locally induced synaptic GABAA receptor clustering34. However, experiments in and AMPA-type glutamate receptors form normally opposite appropriate terminals under conditions of chronic receptor other systems implicate retrograde signals: the targets of sympablockade33,36,46–48. This does not rule out a role for competitive thetic neurons direct cholinergic versus noradrenergic neuro35 transmitter selection , and even in hippocampal neurons a activity in selective synapse elimination, but the evidence indicates that activity per se is not a basic requirement for synaptosimple anterograde signaling model does not account for some genesis. The formation of some morphological synapses in vivo recent observations36. Hippocampal neurons grown in isolation in the absence of Munc18-1, a protein essential for synaptic vesion a small dot of permissive substrate form functional autapscle fusion, also supports this idea49. es37,38. However, in addition to the appropriately matched connections, these isolated neurons also form mismatched Potential molecular families that may serve as signals for appositions of receptor and transmitter 36 . By triple-label synaptogenesis include neurexins and neuroligins50,51, cadimmunocytochemistry, isolated glutamatergic pyramidal neuherins52,53, cadherin-related neuronal receptors54, and ephrins rons were shown to form clusters of NMDA glutamate receptor and Eph receptors55,56. One can imagine how binding of presyopposite some terminals, as expected, but also to form clusters naptic and postsynaptic partners could lead to costabilization of of GABAA receptor opposite different terminals of the same protein scaffolds or activation of signaling pathways thus triggering common aspects of synapse development, or triggering axon36. The specific aggregation of GABAA receptors opposite development of specialized synaptic properties if the participatthese glutamatergic terminals could not be accounted for by raning molecules exhibit selective expression patterns. All of these dom apposition. In vivo, some synapse types also use unconven- nature neuroscience • volume 4 no 6 • june 2001 571 © 2001 Nature Publishing Group http://neurosci.nature.com review © 2001 Nature Publishing Group http://neurosci.nature.com Fig. 3. Mechanisms of postsynaptic protein targeting and potential sites of afferent-specific influence (*). Potential trafficking routes are diagrammed for different glutamate receptor subunits (white and black triangles) and different GABA receptor subunits (white and black squares). This figure is modified from ref. 130. required for postsynaptic clustering of both glycine and GABAA receptors68,69. Motoneurons participate postsynaptically in pure glycinergic, pure GABAergic, and mixed glycinergic and GABAergic synapses, raising the interesting question of how receptor and gephyrin distribution is regulated34. protein families have been localized to subsets of synapses and/or bind to synaptic components and have multiple family members that could mediate selective interactions. Neuroligins presented on the surface of a fibroblast can induce presynaptic specializations in glutamatergic axons51; it is not yet known whether this ability extends to other classes of axons. N-cadherin shows an interesting localization to both GABAergic and glutamatergic synapses early in development and a restriction to glutamatergic synapses later in development in hippocampal cultures57. Studies are no doubt underway to determine the roles of these protein families in synapse matching. Another protein that may be involved in such a mechanism at a specific type of synapse from hippocampal pyramidal neurons to interneurons is Narp (neuronal activity-regulated pentraxin). Narp self-associates, clusters AMPA receptors when expressed on the same or an opposing cell, and is selectively localized to pyramidal-interneuron synapses and to pyramidal axons but not GABAergic axons in culture58. Based on these properties, it is tempting to speculate that Narp secreted by pyramidal cell axons and by interneuron dendrites may be stabilized at these points of contact and induce the high levels of AMPA receptor at such synapses. Within the postsynaptic cell, most scaffolding proteins and some signal transducing enzymes are also concentrated at specific chemical synapse types. The PSD-95 family, stargazin, PICK1, GKAP and Shank scaffolding proteins and the enzymes CaMKIIα and SynGAP are concentrated opposite glutamatergic but not GABAergic terminals24,59–63. One exception to this idea is the AMPA receptor binding protein GRIP, which has been found at glutamatergic and at GABAergic synapses64,65. The transmitter-specific localization of most of these postsynaptic density proteins is presumably controlled by the same transsynaptic signals as for the glutamate receptors. Furthermore, local aggregation and regulation of these scaffolding proteins is thought to contribute to the selective localization of the receptors. Synaptic targeting of AMPA receptors requires stargazin59, and stabilization of AMPA receptors at synapses requires GRIP or ABP66. The major inhibitory scaffolding protein gephyrin binds glycine receptors and colocalizes with both glycine and GABAA receptors but not glutamate receptors67. Furthermore, gephyrin is 572 Presynaptic influences: glutamatergic synapses Individual neurons can establish a distinct complement of glutamate receptor subtypes opposite different glutamatergic afferents (Fig. 1d). Such selective targeting can occur between dendrites or between parts of a single dendrite. A classic example is the composition and properties of mossy fiber versus commissural/associational synapses on hippocampal CA3 pyramidal neurons. Whereas commissural/associational-CA3 synapses exhibit LTP that is dependent on activation of postsynaptic NMDA receptors, LTP at mossy fiber-CA3 synapses has a presynaptic locus and is independent of NMDA receptor activity70. Furthermore, the NMDA receptor-mediated conductance is less at mossy fiber-CA3 synapses than at commissural/associationalCA3 synapses71. The basis for this difference can be accounted for by differences in postsynaptic levels of NMDA receptor protein observed with the sensitive method of antigen retrieval immunohistochemistry72,73. NR1 and NR2A subunits are of low abundance at synapses postsynaptic to mossy fibers, and NR2B almost undetectable, whereas all are abundant at synapses postsynaptic to commissural/associational fibers. These differences could be visualized by light microscopy due to the laminar distribution of inputs. We suspect that such heterogeneity in postsynaptic glutamate receptor content may be widespread, but perhaps not so readily visualized in regions where inputs are not so highly segregated. Indeed, single hippocampal interneurons can express functional GluR2-containing AMPA receptors opposite inputs from CA3 neurons and GluR2-lacking AMPA receptors opposite some inputs from mossy fibers74; the sites of input have not been mapped but may well be intermingled. Two general cellular mechanisms could underlie such selective postsynaptic targeting; local receptor clustering could be triggered by specific innervation during a limited developmental stage, or be continuously dependent on an anterograde signal. The developmental stage of the postsynaptic neuron can be a limiting factor in synaptogenesis. Neurons from different embryonic stages grown in culture for identical periods have different synaptogenic potential75,76, as do neurons from a single embryonic stage grown in culture for different periods77. Postsynaptic protein expression restricted to the time of contact by one input could lead to a localized response that is then propagated by other means. However, this is not the only possible mechanism for establishing differential synaptic composition, and indeed evidence indicates that some glutamatergic synapses initially develop with a similar composition and later diverge. Specifically, in mature Purkinje cells, delta-type glutamate receptors are present at parallel fiber but not climbing fiber synapses, even though these receptors are present at both glutamatergic synapse types earlier in development78. Moreover, a simpler possible mechanism than differential responsiveness on the part of the postsynaptic neuron is a differential signal from the presynaptic afferent. Such a signal could be either differential patterned activnature neuroscience • volume 4 no 6 • june 2001 © 2001 Nature Publishing Group http://neurosci.nature.com © 2001 Nature Publishing Group http://neurosci.nature.com review ity by the two inputs or expression of different cell surface or secreted molecular signals independent of activity. Regardless of the nature of the afferent-specific inducing signal, establishment and maintenance of differential receptor localization presents an interesting challenge for the postsynaptic neuron. Targeting of postsynaptic receptors presumably involves synthesis in the soma and sorting into vesicles from the Golgi network, vesicle transport, vesicle fusion with the plasma membrane, and anchoring and possibly trapping of the receptors in the plasma membrane (Fig. 3). Questions abound regarding all of these steps. For example, are glutamate versus GABA receptors, and even different glutamate receptor types, transported in common or separate vesicles? Which molecular motors79 transport each class of vesicle? A molecular linkage mediated by the PDZ domain proteins mLin2, mLin7 and mLin10 connects NMDA receptors and KIF17, a novel dendritic (+) end-directed microtubule motor80. It is not yet known whether these KIF17-driven vesicles also carry other neurotransmitter receptors. Immunogold localization revealed segregation of GluR4 and mGluR1α to synapses and to intracellular membranes of basal dendrites relative to apical dendrites in fusiform cells of the dorsal cochlear nucleus81, suggesting the possibility of selective transport to different dendrites. Additional such molecular and anatomical studies combined with live imaging of dendritic vesicles following the GFP-based visualization of polarized vesicle transport82 may serve to define classes and routes of dendritic vesicle trafficking. An equally challenging question is to determine the sites of fusion of receptor-bearing vesicles with the plasma membrane and the behavior of receptors once in the plasma membrane. It seems unlikely that vesicle fusion occurs within the postsynaptic density, but whether it is localized near the appropriate synapse or occurs over the entire dendrite surface is not known. Trapping and anchoring in the membrane, and endocytosis, further sorting, and reinsertion or degradation may also have major roles in selective receptor localization. Any of these processes could be influenced by a local afferent-derived signal. NMDA receptors are maintained at synapses independent of F-actin26, possibly by anchora Fig. 4. Selective localization of mGluR7a receptors to presynaptic sites formed onto GABAergic interneurons. (a) Immunogold localization of mGluR7a receptors in the CA3 area of the hippocampus shows a differential expression of mGluR7a depending on the target cell. A single nerve terminal (T) contains two types of synapses: the one heavily labeled for mGluR7 is facing a dendritic shaft (D), characteristic of GABAergic interneurons, whereas the other one lacking any detectable mGluR7a immunoreactivity is facing a dendritic spine (S), characteristic of pyramidal cells. Scale bar, 0.2 µm. (b) Target-signal-dependent presynaptic localization of mGluR7 as evidenced in the hippocampus. (c) The target-specific localization of mGluR7a onto GABAergic interneurons is also observed in cultured hippocampal neurons. Cultured hippocampal neurons at 18 days in vitro were immunolabeled for mGluR7a and glutamic acid decarboxylase (GAD), as a marker for GABAergic cells (*). mGluR7a immunoreactivity is selectively more intense in axon boutons innervating GABAergic interneurons (arrows) than pyramidal cells (arrowheads). Scale bar, 50 µm. Panels (a) and (b) provided by P. Somogyi and colleagues89; published with permission from Nature, copyright 1996. nature neuroscience • volume 4 no 6 • june 2001 ing to PDZ domain proteins such as the PSD-95 family83, and do not undergo rapid cycles of endo/exocytosis84. In contrast, AMPA receptor maintenance is dependent on F-actin26 and involves rapid recycling of surface and intracellular receptors84,85. Examples of changes in anchoring or endo/exocytosis exist for activity regulation of individual synapses (see below), but not yet for afferent cell-type specific regulation. Yet another possible form of afferent-specific response could be through regulation of local protein synthesis. However, this would only seem possible for those few components with moderately abundant levels of dendritic mRNA86, such as CaMKIIα, the NMDA receptor NR1 subunit87, and the glycine receptor alpha subunits88. Similar cellular trafficking events as those described above and outlined in Fig. 3 in the context of differential targeting of postsynaptic glutamate receptors are also presumed to underlie the transmitter-specific clustering of glutamate versus GABA receptors (previous section) as well as the target-specific presynaptic clustering of glutamate receptors presented in the next section. Postsynaptic influences Not only can two postsynaptic sites on a single dendrite exhibit heterogeneity, but two boutons of a single axon can have a different molecular composition and functional properties (Fig. 1e). Such a phenomenon must involve target-specific retrograde signals. One of the most striking examples of presynaptic heterogeneity is the selective localization of mGluR7a and other group III metabotropic glutamate receptors 89–91 (Fig. 4). mGluR7a density in a single hippocampal CA3 axon is ∼10-fold higher at terminals opposite a subset of interneurons compared with terminals opposite CA1 pyramidal cells89. This target-specific selective localization is reflected in functional properties of transmission. Glutamate release from a single CA3 cell was inhibited by group III mGluR antagonists at synapses onto interneurons but not onto CA1 cells92. The selective localization of mGluR7a is reproduced in dissociated neuron cultures b c 573 © 2001 Nature Publishing Group http://neurosci.nature.com review © 2001 Nature Publishing Group http://neurosci.nature.com a b (Fig. 4), a system that may be amenable to mechanistic analysis. Several other instances of differential target-dependent signaling by a single axon have been reported, although in most cases the molecular basis for this difference is not known. For example, high-frequency stimulation of single neocortical neurons leads to a facilitating response in some target cells but a depressing response in others93,94, and BDNF-induced potentiation of transmission from single glutamatergic hippocampal neurons is also target-cell specific95. The locus of these differences in signaling is thought to be presynaptic in all of these cases, suggesting that postsynaptic contact during development results in differences in molecular composition and thus release properties, and possibly also structural characteristics, of these different terminals of single axons. It may be common for particular synaptic properties to result only from interdependent afferent and target-derived signals. For instance, hippocampal dentate granule–CA3 contacts are characterized by large zinc-rich mossy fiber boutons onto branched postsynaptic spines96. The same axons form simple bouton contacts with interneurons, and the same dendrites form simple spine contacts with other pyramidal cells, suggesting that joint anterograde and retrograde signaling is involved in determining the complex synaptic form. In this case, postsynaptic domainspecific factors may also have a role, because these complex synapses form on the proximal dendrites of the CA3 neurons, forming the stratum lucidum. Regulation by cellular domain Intrinsic features of neuronal geometry can specify synapse type and molecular composition (Fig. 1f). Certain subcellular domains can be permissive for specific synapse types and even selective for particular receptors. A prime example of such a selective cellular domain is the axon initial segment. This region of hippocampal pyramidal cells is permissive (or perhaps instructive) for formation of GABAergic but not glutamatergic postsynaptic sites8. Furthermore, GABAergic synapses on the axon initial segment have a higher density of the GABAA receptor α2 subunit 574 Fig. 5. Model for the trafficking of glutamate AMPA receptors at postsynaptic sites. (a) Both AMPA and NMDA receptor activation induce a dynamin-dependent AMPA receptor endocytosis into intracellular compartments without affecting NMDA receptor distribution. The PDZ domain-containing protein PICK1 might participate in this process, whereas GRIP might be primarily involved in the postsynaptic stabilization of AMPA receptors. NMDA receptor activation leads to a complete recycling of AMPA receptors to the cell surface, whereas upon AMPA receptor activation, a fraction of internalized AMPA receptors is degraded. The intracellular pathway of AMPA receptor trafficking is depicted in red when induced by AMPA receptor activation and in blue when induced by NMDA receptor activation. (b) Hypothesis of the involvement of AMPA receptor cycling in LTP and LTD. Under basal conditions, exocytosis and endocytosis occur at the same rate. Specific stimuli, by decoupling exo- and endocytosis, could induce a net gain (LTP) or loss (LTD) of membrane-associated synaptic AMPA receptors. than do GABA synapses on dendrites of the same cell, whereas the α1 subunit is more uniformly targeted to both somatodendritic and axon initial segment GABAergic synapses97. This selective targeting of the α2 subunit may be in part induced by the specific innervation of the axon initial segment by the axo-axonic class of GABAergic neuron. However, the postsynaptic neuron seems to have an active role in the formation of GABAergic synapses on this domain, as the GABAA but not glutamate receptor clusters form on this domain even in pyramidal neurons grown in the absence of GABAergic input (A. Rao and A.M.C., unpublished data). GABAergic synapses also form preferentially on somata by an activity-independent mechanism98. Another intriguing example of how geometry influences synaptic composition and function is the evidence for gradients of increasing synaptic size and current amplitude along a single dendrite with increasing distance from the soma. In hippocampal pyramidal neurons, synaptic conductance increases with distance from the soma, thus counterbalancing dendritic filtering effects and resulting in location-independent responses at the soma99. The data are consistent with a mechanism involving a gradient in synapse size. For inhibitory glycinergic synapses, somatodendritic gradients in synapse size, or immunofluorescence cluster size of glycine receptors100 or gephyrin28, has been observed, potentially also to compensate for greater electrotonic attenuation of synaptic responses with distance. Exactly how proximo-distal position is sensed to establish such a gradient presents an interesting puzzle. Another study101 reported that postsynaptic responsiveness varies inversely with the density of synaptic input onto hippocampal neurons in culture, perhaps a mechanism to adjust the dynamic range of the spike-firing decision. Heterogeneity within one synapse Considering only one class of synapse (a single presynaptic cell type making synapses onto a single postsynaptic cell type), a surprising heterogeneity of features still exists (Fig. 1g). This diversity may be in large part determined by the activity of individual synapses. One aspect of this heterogeneity that has received much recent attention is the variability in postsynaptic AMPA type glutamate receptor content in hippocampal pyramidal neurons. Quantitative electron microscopy studies of CA3 Schaeffer collateral synapses onto CA1 neurons found 19% of synapses strongly immunoreactive, 67% moderately immunoreactive and 17% immunonegative for AMPA receptors23. Immunonegative synapses were exclusively the smaller synapses31. Because every Schaeffer collateral synapse onto CA1 pyramidal cell spines contains immunoreactive NMDA receptors31,32, a subset of these synapses nature neuroscience • volume 4 no 6 • june 2001 © 2001 Nature Publishing Group http://neurosci.nature.com © 2001 Nature Publishing Group http://neurosci.nature.com review express only NMDA receptors and could correspond to the anatomical substrate for the electrophysiologically defined ‘silent synapses.’ These observations suggest that the targeting mechanisms and the regulation of cell surface expression of these two types of glutamate receptors follow different rules. Indeed, recent studies converge to indicate that, unlike NMDA receptors, AMPA receptors are highly mobile in a short time scale and that these dynamic properties are relevant for the observed heterogeneity in AMPA receptor synapse composition and could have an important part in mediating different forms of synaptic plasticity. Based on imaging and biochemical approaches, several reports indicate that all AMPA receptors subunits can be rapidly removed from and inserted into the postsynaptic membrane (Fig. 5). The selective labeling of cell-surface AMPA receptors by antibodies against extracellular epitopes or cell surface biotinylation assays in cultured hippocampal neurons showed a rapid constitutive internalization of AMPA receptors but not NMDA receptors under basal conditions84,85,102. Intriguingly, blocking clathrin-mediated internalization of AMPA receptors affected neither the total number of AMPA receptors expressed at the cell surface nor AMPA mEPSCs84,102,103 (but see ref. 104). Ehlers84 directly showed that internalization and reinsertion of the internalized receptors to the plasma membrane was achieved at the same rate, and that an increase in endocytosis induced by picrotoxin directly correlated with an increase in receptor recycling to the plasma membrane. These results suggest that there is an unknown mechanism tightly coupling the constitutive endocytosis and insertion of AMPA receptors to maintain a stable amount of synaptic AMPA receptor. This mechanism seems to involve interactions of AMPA receptor subunits with the PDZ domain proteins GRIP or ABP105 and interactions of GluR2 with the chaperone protein NSF106. The rates of AMPA receptor endocytosis and exocytosis can be uncoupled by synaptic activity, depending on the extent and balance of AMPA and NMDA receptor stimulation, to effect a net change in surface AMPA receptor levels. Treatment of cultured neurons with high levels of glutamate produces a massive internalization of AMPA receptors, again without affecting NMDA receptor distribution107. Treatment with either AMPA or NMDA induces AMPA receptor endocytosis at a similar rate84. However, the mechanisms, the intracellular sorting and the subsequent fate of internalized AMPA receptors are different upon activation of each receptor type. NMDA induced only a transient decrease in the total level of surface AMPA receptors, whereas AMPA treatment induced late endosomal targeting and lysosomal degradation of a fraction of internalized AMPA receptors84. This results in a loss of membrane-associated AMPA receptors even after a long time of recovery84,85, which parallels a decrease in the frequency of mEPSCs107. The hypothesis that activity-dependent redistribution of AMPA receptors to and from the postsynaptic plasma membrane is important in NMDA receptor-dependent LTP and LTD has been strengthened over the past few years. Tetanic stimulation generating LTP induced the appearance of electrophysiologically tagged GluR1 at synapses in hippocampal slices108. This synaptic recruitment of GluR1 observed upon LTP expression is dependent on a PDZ domain-containing protein because this effect required an intact C-terminus of GluR1 (ref. 20). Several studies report that LTD involves net endocytosis of synaptic AMPA receptors102,109, possibly through protein kinase C phosphorylation of the receptor, binding to the PDZ domain protein PICK1, and inhibition of binding to GRIP and ABP110–112. Trafficking of different AMPA receptor subunits can also occur independently, as evidenced by another form of plasticity in cerebellar nature neuroscience • volume 4 no 6 • june 2001 stellate cells involving a rapid activity-dependent switch in the subunit composition of synaptic AMPA receptors113. Unlike AMPA receptors, short-term manipulation of the level of activity does not change the subcellular distribution of NMDA receptors, but chronic blockade in hippocampal cultures induces a large increase in synaptic NMDA receptor immunoreactivity114 and NMDA receptor-mediated excitotoxicity115. This effect is mediated by redistribution of existing receptors to the synapse by a mechanism requiring activation of cAMP-dependent protein kinase115. AMPA receptors also exhibit long-term homeostatic regulation of synaptic density, in most studies apparently independently of NMDA receptors114,116,117 (but see ref. 118). The localization of synaptic signaling proteins can also be rapidly modified in response to changes in synaptic activity. The Ca2+/calmodulin-dependent protein kinase II (CaMKII) is a signaling protein involved in synaptic plasticity: CaMKIIα mutant mice show impaired hippocampal LTP and behavioral defects in spatial learning and memory119. Live imaging studies of a GFPtagged CaMKII expressed in cultured hippocampal neurons showed that NMDA receptor stimulation, through Ca2+ influx, led to a rapid redistribution of CaMKII from dendritic cytosol to postsynaptic sites120. This synaptic translocation required binding to calmodulin, and the stabilization at synapses for several minutes required the autophosphorylation of the kinase120–122. The transient localization of CaMKII at synapses could modulate the function of various synaptic proteins, such as AMPA receptors, by a direct phosphorylation21,22 or by phosphorylation of intermediate molecules involved in GluR1 synaptic delivery during LTP20. Heterogeneity of postsynaptic receptor composition also exists at inhibitory synapses. Cerebellar granule cells receive GABAergic input only from Golgi cells and thus represent an anatomically homogeneous class of synapses. All Golgi synapses were found to contain similar levels of GABAA receptor β2/3 subunits, but to be highly heterogeneous with regard to their α subunit content. Quantitative electron microscopy indicated that three populations of synapses were found according to their α subunit content: one expressing only or mostly α1 subunit, one expressing only or mostly α6 subunit and one expressing both subunits at similar levels39,123. This variability in α1/α6 subunit ratio could account for the existence of fast and slow decays of spontaneous IPSCs observed in granule cells124. The insertion of different α subunits might be regulated by the activity of pre- and postsynaptic elements to adapt the postsynaptic composition of GABAA receptors to changing needs of the system. The basis for synaptic heterogeneity within one class of synapse can also originate from some variability at presynaptic sites. Measures of the release probability of neurotransmitter at Schaeffer collateral/commissural fibers making synapses onto CA1 neurons indicated that among these glutamatergic synapses, two distinct classes of synapses with a sixfold difference in the release probability are resolved125. Synapses with low release probability contribute over half of the transmission and are more sensitive to drugs enhancing transmitter release. This nonuniformity in release probability could rely on structural factors, such as the size of the release site, the number of available vesicles and possible differences in the Ca2+ channel density126. Autaptic heterogeneity Even in the case of autaptic synapses formed in isolated neurons, much evidence suggests that all synapses are not equivalent (Fig. 1h). For instance, there is a wide variability in the level of immunostaining for several synaptic vesicle-associated proteins, 575 © 2001 Nature Publishing Group http://neurosci.nature.com © 2001 Nature Publishing Group http://neurosci.nature.com review such as synaptophysin, synapsin I, and SV2, which correlates with the extent of FM1-43 labeling at synapses127. Thus, differential expression of synaptic vesicle-associated proteins can occur at different boutons of the same neuron. The existence of two classes of synapses with different release probability described above was also demonstrated in single cultured hippocampal neurons128. Similarly, the existence of a heterogeneity in the presynaptic maturation of synaptic vesicles in glutamatergic hippocampal neurons was recently demonstrated by the generation of null mutant mice for Munc 13-1, a brain-specific presynaptic phorbol ester receptor 129. Recordings performed on cultures from Munc 13-1-deficient mice showed that most synapses of any given single excitatory neuron did not release neurotransmitter, whereas transmitter release from a small subpopulation of synapses of the same cell was not affected and was therefore independent of Munc 13-1. It is not yet clear whether these differences in release properties are related to synapse size, nor whether they are somehow determined by geometrical considerations such as the distance between the release site and the soma or whether they result from a more random process. Postsynaptic receptor composition also varies widely among autaptic sites of isolated neurons, as described above, resulting in both matched appositions of terminals with the appropriate receptors and mismatched appositions with chemically inappropriate receptors36. In this case, sites of different receptor composition have been observed intermingled along a single dendrite. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. Conclusions The molecular composition and functional properties of individual synapses, even among limited afferent and target cell types, are surprisingly heterogeneous. Synaptic diversity results from selective cellular expression and selective subcellular targeting of synaptic components regulated by combinations of afferentderived and target-derived signals. Synaptic activity is one major afferent-derived signal. Additional intrinsic factors such as cellular geometry and perhaps some element of randomness are also required to explain the diversity even among autapses made by a single neuron. Synaptic variation is manifest as differences in release and response characteristics, ultrastructural features, and molecular composition; future studies combining all three types of assays will be important to understand the properties of individual synapses. ACKNOWLEDGEMENTS We thank A. Rao, K. Tovar and R. Wong for comments. Research in the authors’ laboratory is supported by NIH grants NS33184, NS39286, NS34448 and the Pew Charitable Trust. 21. 22. 23. 24. 25. 26. 27. 28. 29. RECEIVED 28 FEBRUARY; ACCEPTED 9 APRIL 2001 30. 1. Gray, E. G. Axo-somatic and axo-dendritic synapses of the cerebral cortex: an electron microscope study. J. Anat. 93, 420–433 (1959). 2. Sudhof, T. C. The synaptic vesicle cycle: a cascade of protein–protein interactions. Nature 375, 645–653 (1995). 3. Ullrich, B. et al. Functional properties of multiple synaptotagmins in brain. Neuron 13, 1281–1291 (1994). 4. Trimble, W. S., Gray, T. S., Elferink, L. A., Wilson, M. C. & Scheller, R. H. Distinct patterns of expression of two VAMP genes within the rat brain. J. Neurosci. 10, 1380–1387 (1990). 5. Morgans, C. W., Brandstatter, J. H., Kellerman, J., Betz, H. & Wassle, H. A SNARE complex containing syntaxin 3 is present in ribbon synapses of the retina. J. Neurosci. 16, 6713–6721 (1996). 6. Geppert, M. et al. Synaptotagmin I: a major Ca2+ sensor for transmitter release at a central synapse. Cell 79, 717–727 (1994). 7. Fenster, S. D. et al. Piccolo, a presynaptic zinc finger protein structurally related to bassoon. Neuron 25, 203–214 (2000). 8. Ribak, C. E. & Roberts, R. C. GABAergic synapses in the brain identified with 576 31. 32. 33. 34. 35. antisera to GABA and its synthesizing enzyme, glutamate decarboxylase. J. Electron Microsc. Tech. 15, 34–48 (1990). Minelli, A., Brecha, N. C., Karschin, C., DeBiasi, S. & Conti, F. GAT-1, a highaffinity GABA plasma membrane transporter, is localized to neurons and astroglia in the cerebral cortex. J. Neurosci. 15, 7734–7746 (1995). Dumoulin, A. et al. Presence of the vesicular inhibitory amino acid transporter in GABAergic and glycinergic synaptic terminal boutons. J. Cell Sci. 112, 811–823 (1999). Chaudhry, F. A. et al. The vesicular GABA transporter, VGAT, localizes to synaptic vesicles in sets of glycinergic as well as GABAergic neurons. J. Neurosci. 18, 9733–9750 (1998). Esclapez, M., Tillakaratne, N. J., Kaufman, D. L., Tobin, A. J. & Houser, C. R. Comparative localization of two forms of glutamic acid decarboxylase and their mRNAs in rat brain supports the concept of functional differences between the forms. J. Neurosci. 14, 1834–1855 (1994). Sagne, C. et al. Cloning of a functional vesicular GABA and glycine transporter by screening of genome databases. FEBS Lett. 417, 177–183 (1997). Crump, F. T., Fremeau, R. T. & Craig, A. M. Localization of the brain-specific high-affinity l-proline transporter in cultured hippocampal neurons: molecular heterogeneity of synaptic terminals. Mol. Cell. Neurosci. 13, 25–39 (1999). Takamori, S., Rhee, J. S., Rosenmund, C. & Jahn, R. Identification of a vesicular glutamate transporter that defines a glutamatergic phenotype in neurons. Nature 407, 189–194 (2000). Bellocchio, E. E., Reimer, R. J., Fremeau, R. T. & Edwards, R. H. Uptake of glutamate into synaptic vesicles by an inorganic phosphate transporter. Science 289, 957–960 (2000). McMahon, L. L. & Kauer, J. A. Hippocampal interneurons express a novel form of synaptic plasticity. Neuron 18, 295–305 (1997). Kennedy, M. B. Signal transduction molecules at the glutamatergic postsynaptic membrane. Brain Res. Brain Res. Rev. 26, 243–257 (1998). Sik, A., Hajos, N., Gulacsi, A., Mody, I. & Freund, T. F. The absence of a major Ca2+ signaling pathway in GABAergic neurons of the hippocampus. Proc. Natl. Acad. Sci. USA 95, 3245–3250 (1998). Hayashi, Y. et al. Driving AMPA receptors into synapses by LTP and CaMKII: requirement for GluR1 and PDZ domain interaction. Science 287, 2262–2267 (2000). Barria, A., Muller, D., Derkach, V., Griffith, L. C. & Soderling, T. R. Regulatory phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term potentiation. Science 276, 2042–2045 (1997). Lee, H. K., Barbarosie, M., Kameyama, K., Bear, M. F. & Huganir, R. L. Regulation of distinct AMPA receptor phosphorylation sites during bidirectional synaptic plasticity. Nature 405, 955–959 (2000). Nusser, Z. et al. Cell type and pathway dependence of synaptic AMPA receptor number and variability in the hippocampus. Neuron 21, 545–559 (1998). Rao, A., Kim, E., Sheng, M. & Craig, A. M. Heterogeneity in the molecular composition of excitatory postsynaptic sites during development of hippocampal neurons in culture. J. Neurosci. 18, 1217–1229 (1998). Zhang, W., Vazquez, L., Apperson, M. & Kennedy, M. B. Citron binds to PSD95 at glutamatergic synapses on inhibitory neurons in the hippocampus. J. Neurosci. 19, 96–108 (1999). Allison, D. W., Gelfand, V. I., Spector, I. & Craig, A. M. Role of actin in anchoring postsynaptic receptors in cultured hippocampal neurons: differential attachment of NMDA versus AMPA receptors. J. Neurosci. 18, 2423–2436 (1998). Yuste, R., Majewska, A. & Holthoff, K. From form to function: calcium compartmentalization in dendritic spines. Nat. Neurosci. 3, 653–659 (2000). Alvarez, F. J., Dewey, D. E., Harrington, D. A. & Fyffe, R. E. Cell-type specific organization of glycine receptor clusters in the mammalian spinal cord. J. Comp. Neurol. 379, 150–170 (1997). Nusser, Z., Roberts, J. D., Baude, A., Richards, J. G. & Somogyi, P. Relative densities of synaptic and extrasynaptic GABAA receptors on cerebellar granule cells as determined by a quantitative immunogold method. J. Neurosci. 15, 2948–2960 (1995). Triller, A., Cluzeaud, F., Pfeiffer, F., Betz, H. & Korn, H. Distribution of glycine receptors at central synapses: an immunoelectron microscopy study. J. Cell Biol. 101, 683–688 (1985). Takumi, Y., Ramirez-Leon, V., Laake, P., Rinvik, E. & Ottersen, O. P. Different modes of expression of AMPA and NMDA receptors in hippocampal synapses. Nat. Neurosci. 2, 618–624 (1999). Racca, C., Stephenson, F. A., Streit, P., Roberts, J. D. & Somogyi, P. NMDA receptor content of synapses in stratum radiatum of the hippocampal CA1 area. J. Neurosci. 20, 2512–2522 (2000). Craig, A. M., Blackstone, C. D., Huganir, R. L. & Banker, G. Selective clustering of glutamate and gamma-aminobutyric acid receptors opposite terminals releasing the corresponding neurotransmitters. Proc. Natl. Acad. Sci. USA 91, 12373–12377 (1994). Levi, S., Chesnoy-Marchais, D., Sieghart, W. & Triller, A. Synaptic control of glycine and GABA(A) receptors and gephyrin expression in cultured motoneurons. J. Neurosci. 19, 7434–7449 (1999). Francis, N. J. & Landis, S. C. Cellular and molecular determinants of sympathetic neuron development. Annu. Rev. Neurosci. 22, 541–566 (1999). nature neuroscience • volume 4 no 6 • june 2001 © 2001 Nature Publishing Group http://neurosci.nature.com © 2001 Nature Publishing Group http://neurosci.nature.com review 36. Rao, A., Cha, E. M. & Craig, A. M. Mismatched appositions of presynaptic and postsynaptic components in isolated hippocampal neurons. J. Neurosci. 20, 8344–8353 (2000). 37. Bekkers, J. M. & Stevens, C. F. Excitatory and inhibitory autaptic currents in isolated hippocampal neurons maintained in cell culture. Proc. Natl. Acad. Sci. USA 88, 7834–7838 (1991). 38. Segal, M. M. Epileptiform activity in microcultures containing one excitatory hippocampal neuron. J. Neurophysiol. 65, 761–770 (1991). 39. Nusser, Z., Sieghart, W., Stephenson, F. A. & Somogyi, P. The alpha 6 subunit of the GABAA receptor is concentrated in both inhibitory and excitatory synapses on cerebellar granule cells. J. Neurosci. 16, 103–114 (1996). 40. Hanson, J. E. & Smith, Y. Group I metabotropic glutamate receptors at GABAergic synapses in monkeys. J. Neurosci. 19, 6488–6496 (1999). 41. Tsen, G. et al. Receptors with opposing functions are in postsynaptic microdomains under one presynaptic terminal. Nat. Neurosci. 3, 126–132 (2000). 42. Featherstone, D. E. et al. Presynaptic glutamic acid decarboxylase is required for induction of the postsynaptic receptor field at a glutamatergic synapse. Neuron 27, 71–84 (2000). 43. Levi, S., Vannier, C. & Triller, A. Strychnine-sensitive stabilization of postsynaptic glycine receptor clusters. J. Cell Sci. 111, 335–345 (1998). 44. Kirsch, J. & Betz, H. Glycine-receptor activation is required for receptor clustering in spinal neurons. Nature 392, 717–720 (1998). 45. McKinney, R. A., Capogna, M., Durr, R., Gahwiler, B. H. & Thompson, S. M. Miniature synaptic events maintain dendritic spines via AMPA receptor activation. Nat. Neurosci. 2, 44–49 (1999). 46. Verderio, C., Coco, S., Fumagalli, G. & Matteoli, M. Spatial changes in calcium signaling during the establishment of neuronal polarity and synaptogenesis. J. Cell Biol. 126, 1527–1536 (1994). 47. Mammen, A. L., Huganir, R. L. & O’Brien, R. J. Redistribution and stabilization of cell surface glutamate receptors during synapse formation. J. Neurosci. 17, 7351–7358 (1997). 48. Cottrell, J. R., Dube, G. R., Egles, C. & Liu, G. Distribution, density, and clustering of functional glutamate receptors before and after synaptogenesis in hippocampal neurons. J. Neurophysiol. 84, 1573–1587 (2000). 49. Verhage, M. et al. Synaptic assembly of the brain in the absence of neurotransmitter secretion. Science 287, 864–869 (2000). 50. Missler, M., Fernandez-Chacon, R. & Sudhof, T. C. The making of neurexins. J. Neurochem. 71, 1339–1347 (1998). 51. Scheiffele, P., Fan, J., Choih, J., Fetter, R. & Serafini, T. Neuroligin expressed in nonneuronal cells triggers presynaptic development in contacting axons. Cell 101, 657–669 (2000). 52. Colman, D. R. Neurites, synapses, and cadherins reconciled. Mol. Cell. Neurosci. 10, 1–6 (1997). 53. Uchida, N., Honjo, Y., Johnson, K. R., Wheelock, M. J. & Takeichi, M. The catenin/cadherin adhesion system is localized in synaptic junctions bordering transmitter release zones. J. Cell Biol. 135, 767–779 (1996). 54. Kohmura, N. et al. Diversity revealed by a novel family of cadherins expressed in neurons at a synaptic complex. Neuron 20, 1137–1151 (1998). 55. Buchert, M. et al. The junction-associated protein AF-6 interacts and clusters with specific Eph receptor tyrosine kinases at specialized sites of cell–cell contact in the brain. J. Cell Biol. 144, 361–371 (1999). 56. Torres, R. et al. PDZ proteins bind, cluster, and synaptically colocalize with Eph receptors and their ephrin ligands. Neuron 21, 1453–1463 (1998). 57. Benson, D. L. & Tanaka, H. N-cadherin redistribution during synaptogenesis in hippocampal neurons. J. Neurosci. 18, 6892–6904 (1998). 58. O’Brien, R. J. et al. Synaptic clustering of AMPA receptors by the extracellular immediate- early gene product Narp. Neuron 23, 309–323 (1999). 59. Chen, L. et al. Stargazing regulates synaptic targeting of AMPA receptors by two distinct mechanisms. Nature 408, 936–943 (2000). 60. Xia, J., Zhang, X., Staudinger, J. & Huganir, R. L. Clustering of AMPA receptors by the synaptic PDZ domain-containing protein PICK1. Neuron 22, 179–187 (1999). 61. Naisbitt, S. et al. Shank, a novel family of postsynaptic density proteins that binds to the NMDA receptor/PSD-95/GKAP complex and cortactin. Neuron 23, 569–582 (1999). 62. Liu, X. B. & Jones, E. G. Localization of alpha type II calcium calmodulindependent protein kinase at glutamatergic but not gamma-aminobutyric acid (GABAergic) synapses in thalamus and cerebral cortex. Proc. Natl. Acad. Sci. USA 93, 7332–7336 (1996). 63. Kim, J. H., Liao, D., Lau, L. F. & Huganir, R. L. SynGAP: a synaptic RasGAP that associates with the PSD-95/SAP90 protein family. Neuron 20, 683–691 (1998). 64. Wyszynski, M. et al. Association of AMPA receptors with a subset of glutamate receptor-interacting protein in vivo. J. Neurosci. 19, 6528–6537 (1999). 65. Dong, H. et al. Characterization of the glutamate receptor-interacting proteins GRIP1 and GRIP2. J. Neurosci. 19, 6930–6941 (1999). 66. Osten, P. et al. The AMPA receptor GluR2 C terminus can mediate a reversible, ATP-dependent interaction with NSF and alpha- and beta-SNAPs. Neuron 21, 99–110 (1998). 67. Sassoe-Pognetto, M. & Fritschy, J. M. Mini-review: gephyrin, a major postsynaptic protein of GABAergic synapses. Eur. J. Neurosci. 12, 2205–2210 (2000). 68. Feng, G. et al. Dual requirement for gephyrin in glycine receptor clustering nature neuroscience • volume 4 no 6 • june 2001 and molybdoenzyme activity. Science 282, 1321–1324 (1998). 69. Kneussel, M. et al. Loss of postsynaptic GABA(A) receptor clustering in gephyrin-deficient mice. J. Neurosci. 19, 9289–9297 (1999). 70. Nicoll, R. A. & Malenka, R. C. Contrasting properties of two forms of longterm potentiation in the hippocampus. Nature 377, 115–118 (1995). 71. Jonas, P., Major, G. & Sakmann, B. Quantal components of unitary EPSCs at the mossy fibre synapse on CA3 pyramidal cells of rat hippocampus. J. Physiol. (Lond.) 472, 615–663 (1993). 72. Watanabe, M. et al. Selective scarcity of NMDA receptor channel subunits in the stratum lucidum (mossy fibre-recipient layer) of the mouse hippocampal CA3 subfield. Eur. J. Neurosci. 10, 478–487 (1998). 73. Fritschy, J. M., Weinmann, O., Wenzel, A. & Benke, D. Synapse-specific localization of NMDA and GABA(A) receptor subunits revealed by antigenretrieval immunohistochemistry. J. Comp. Neurol. 390, 194–210 (1998). 74. Toth, K. & McBain, C. J. Afferent-specific innervation of two distinct AMPA receptor subtypes on single hippocampal interneurons. Nat. Neurosci. 1, 572–578 (1998). 75. Vicario-Abejon, C., Collin, C., McKay, R. D. & Segal, M. Neurotrophins induce formation of functional excitatory and inhibitory synapses between cultured hippocampal neurons. J. Neurosci. 18, 7256–7271 (1998). 76. Mohrmann, R., Werner, M., Hatt, H. & Gottmann, K. Target-specific factors regulate the formation of glutamatergic transmitter release sites in cultured neocortical neurons. J. Neurosci. 19, 10004–10013 (1999). 77. Fletcher, T. L., De Camilli, P. & Banker, G. Synaptogenesis in hippocampal cultures: evidence indicating that axons and dendrites become competent to form synapses at different stages of neuronal development. J. Neurosci. 14, 6695–6706 (1994). 78. Zhao, H. M., Wenthold, R. J. & Petralia, R. S. Glutamate receptor targeting to synaptic populations on Purkinje cells is developmentally regulated. J. Neurosci. 18, 5517–5528 (1998). 79. Goldstein, L. S. & Yang, Z. Microtubule-based transport systems in neurons: the roles of kinesins and dyneins. Annu. Rev. Neurosci. 23, 39–71 (2000). 80. Setou, M., Nakagawa, T., Seog, D. H. & Hirokawa, N. Kinesin superfamily motor protein KIF17 and mLin-10 in NMDA receptor-containing vesicle transport. Science 288, 1796–1802 (2000). 81. Rubio, M. E. & Wenthold, R. J. Differential distribution of intracellular glutamate receptors in dendrites. J. Neurosci. 19, 5549–5562 (1999). 82. Burack, M. A., Silverman, M. A. & Banker, G. The role of selective transport in neuronal protein sorting. Neuron 26, 465–472 (2000). 83. Kornau, H. C., Seeburg, P. H. & Kennedy, M. B. Interaction of ion channels and receptors with PDZ domain proteins. Curr. Opin. Neurobiol. 7, 368–373 (1997). 84. Ehlers, M. D. Reinsertion or degradation of AMPA receptors determined by activity-dependent endocytic sorting. Neuron 28, 511–525 (2000). 85. Lin, J. W. et al. Distinct molecular mechanisms and divergent endocytotic pathways of AMPA receptor internalization. Nat. Neurosci. 3, 1282–1290 (2000). 86. Steward, O. Targeting of mRNAs to subsynaptic microdomains in dendrites. Curr. Opin. Neurobiol. 5, 55–61 (1995). 87. Benson, D. L. Dendritic compartmentation of NMDA receptor mRNA in cultured hippocampal neurons. Neuroreport 8, 823–828 (1997). 88. Racca, C., Gardiol, A. & Triller, A. Dendritic and postsynaptic localizations of glycine receptor alpha subunit mRNAs. J. Neurosci. 17, 1691–1700 (1997). 89. Shigemoto, R. et al. Target-cell-specific concentration of a metabotropic glutamate receptor in the presynaptic active zone. Nature 381, 523–525 (1996). 90. Shigemoto, R. et al. Differential presynaptic localization of metabotropic glutamate receptor subtypes in the rat hippocampus. J. Neurosci. 17, 7503–7522 (1997). 91. Brandstatter, J. H., Koulen, P., Kuhn, R., van der Putten, H. & Wassle, H. Compartmental localization of a metabotropic glutamate receptor (mGluR7): two different active sites at a retinal synapse. J. Neurosci. 16, 4749–4756 (1996). 92. Scanziani, M., Gahwiler, B. H. & Charpak, S. Target cell-specific modulation of transmitter release at terminals from a single axon. Proc. Natl. Acad. Sci. USA 95, 12004–12009 (1998). 93. Markram, H., Wang, Y. & Tsodyks, M. Differential signaling via the same axon of neocortical pyramidal neurons. Proc. Natl. Acad. Sci. USA 95, 5323–5328 (1998). 94. Reyes, A. et al. Target-cell-specific facilitation and depression in neocortical circuits. Nat. Neurosci. 1, 279–285 (1998). 95. Schinder, A. F., Berninger, B. & Poo, M. Postsynaptic target specificity of neurotrophin-induced presynaptic potentiation. Neuron 25, 151–163 (2000). 96. Amaral, D. G. & Dent, J. A. Development of the mossy fibers of the dentate gyrus: I. A light and electron microscopic study of the mossy fibers and their expansions. J. Comp. Neurol. 195, 51–86 (1981). 97. Nusser, Z., Sieghart, W., Benke, D., Fritschy, J. M. & Somogyi, P. Differential synaptic localization of two major gamma-aminobutyric acid type A receptor alpha subunits on hippocampal pyramidal cells. Proc. Natl. Acad. Sci. USA 93, 11939–11944 (1996). 98. Benson, D. L. & Cohen, P. A. Activity-independent segregation of excitatory and inhibitory synaptic terminals in cultured hippocampal neurons. J. Neurosci. 16, 6424–6432 (1996). 99. Magee, J. C. & Cook, E. P. Somatic EPSP amplitude is independent of synapse location in hippocampal pyramidal neurons. Nat. Neurosci. 3, 895–903 (2000). 577 © 2001 Nature Publishing Group http://neurosci.nature.com © 2001 Nature Publishing Group http://neurosci.nature.com review 100. Triller, A., Seitanidou, T., Franksson, O. & Korn, H. Size and shape of glycine receptor clusters in a central neuron exhibit a somato-dendritic gradient. New Biol. 2, 637–641 (1990). 101. Liu, G. & Tsien, R. W. Properties of synaptic transmission at single hippocampal synaptic boutons. Nature 375, 404–408 (1995). 102. Man, Y. H. et al. Regulation of AMPA receptor-mediated synaptic transmission by clathrin-dependent receptor internalization. Neuron 25, 649–662 (2000). 103. Wang, Y. T. & Linden, D. J. Expression of cerebellar long-term depression requires postsynaptic clathrin-mediated endocytosis. Neuron 25, 635–647 (2000). 104. Luscher, C. et al. Role of AMPA receptor cycling in synaptic transmission and plasticity. Neuron 24, 649–658 (1999). 105. Osten, P. et al. Mutagenesis reveals a role for ABP/GRIP binding to GluR2 in synaptic surface accumulation of the AMPA receptor. Neuron 27, 313–325 (2000). 106. Noel, J. et al. Surface expression of AMPA receptors in hippocampal neurons is regulated by an NSF-dependent mechanism. Neuron 23, 365–376 (1999). 107. Lissin, D. V., Carroll, R. C., Nicoll, R. A., Malenka, R. C. & von Zastrow, M. Rapid, activation-induced redistribution of ionotropic glutamate receptors in cultured hippocampal neurons. J. Neurosci. 19, 1263–1272 (1999) (erratum, J. Neurosci. 19, 3275, 1999). 108. Shi, S. H. et al. Rapid spine delivery and redistribution of AMPA receptors after synaptic NMDA receptor activation. Science 284, 1811–1816 (1999). 109. Carroll, R. C., Lissin, D. V., von Zastrow, M., Nicoll, R. A. & Malenka, R. C. Rapid redistribution of glutamate receptors contributes to long-term depression in hippocampal cultures. Nat. Neurosci. 2, 454–460 (1999). 110. Matsuda, S., Launey, T., Mikawa, S. & Hirai, H. Disruption of AMPA receptor GluR2 clusters following long-term depression induction in cerebellar Purkinje neurons. EMBO J. 19, 2765–2774 (2000). 111. Xia, J., Chung, H. J., Wihler, C., Huganir, R. L. & Linden, D. J. Cerebellar long-term depression requires PKC-regulated interactions between GluR2/3 and PDZ domain-containing proteins. Neuron 28, 499–510 (2000). 112. Daw, M. I. et al. PDZ proteins interacting with C-terminal GluR2/3 are involved in a PKC-dependent regulation of AMPA receptors at hippocampal synapses. Neuron 28, 873–886 (2000). 113. Liu, S. Q. & Cull-Candy, S. G. Synaptic activity at calcium-permeable AMPA receptors induces a switch in receptor subtype. Nature 405, 454–458 (2000). 114. Rao, A. & Craig, A. M. Activity regulates the synaptic localization of the NMDA receptor in hippocampal neurons. Neuron 19, 801–812 (1997). 115. Crump, F. T., Dillman, K. & Craig, A. M. cAMP-dependent protein kinase 578 116. 117. 118. 119. 120. 121. 122. 123. 124. 125. 126. 127. 128. 129. 130. mediates activity-regulated synaptic targeting of NMDA receptors. J. Neurosci. (in press). Lissin, D. V. et al. Activity differentially regulates the surface expression of synaptic AMPA and NMDA glutamate receptors. Proc. Natl. Acad. Sci. USA 95, 7097–7102 (1998). Liao, D., Zhang, X., O’Brien, R., Ehlers, M. D. & Huganir, R. L. Regulation of morphological postsynaptic silent synapses in developing hippocampal neurons. Nat. Neurosci. 2, 37–43 (1999). Watt, A. J., van Rossum, M. C., MacLeod, K. M., Nelson, S. B. & Turrigiano, G. G. Activity coregulates quantal AMPA and NMDA currents at neocortical synapses. Neuron 26, 659–670 (2000). Elgersma, Y. & Silva, A. J. Molecular mechanisms of synaptic plasticity and memory. Curr. Opin. Neurobiol. 9, 209–213 (1999). Shen, K. & Meyer, T. Dynamic control of CaMKII translocation and localization in hippocampal neurons by NMDA receptor stimulation. Science 284, 162–166 (1999). Strack, S., Choi, S., Lovinger, D. M. & Colbran, R. J. Translocation of autophosphorylated calcium/calmodulin-dependent protein kinase II to the postsynaptic density. J. Biol. Chem. 272, 13467–13470 (1997). Yoshimura, Y. & Yamauchi, T. Phosphorylation-dependent reversible association of Ca2+/calmodulin-dependent protein kinase II with the postsynaptic densities. J. Biol. Chem. 272, 26354–26359 (1997). Nusser, Z., Sieghart, W. & Somogyi, P. Segregation of different GABAA receptors to synaptic and extrasynaptic membranes of cerebellar granule cells. J. Neurosci. 18, 1693–1703 (1998). Puia, G., Costa, E. & Vicini, S. Functional diversity of GABA-activated Clcurrents in Purkinje versus granule neurons in rat cerebellar slices. Neuron 12, 117–126 (1994). Hessler, N. A., Shirke, A. M. & Malinow, R. The probability of transmitter release at a mammalian central synapse. Nature 366, 569–572 (1993). Schikorski, T. & Stevens, C. F. Quantitative ultrastructural analysis of hippocampal excitatory synapses. J. Neurosci. 17, 5858–5867 (1997). Staple, J. K., Osen-Sand, A., Benfenati, F., Pich, E. M. & Catsicas, S. Molecular and functional diversity at synapses of individual neurons in vitro. Eur. J. Neurosci. 9, 721–731 (1997). Rosenmund, C., Clements, J. D. & Westbrook, G. L. Nonuniform probability of glutamate release at a hippocampal synapse. Science 262, 754–757 (1993). Augustin, I., Rosenmund, C., Sudhof, T. C. & Brose, N. Munc13-1 is essential for fusion competence of glutamatergic synaptic vesicles. Nature 400, 457–461 (1999). Craig, A. M. & Banker, G. Neuronal polarity. Annu. Rev. Neurosci. 17, 267–310 (1994). nature neuroscience • volume 4 no 6 • june 2001