Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
E. O. Wilson wikipedia , lookup
Human genetic variation wikipedia , lookup
Origin of language wikipedia , lookup
Darwinian literary studies wikipedia , lookup
Multiregional origin of modern humans wikipedia , lookup
Human nature wikipedia , lookup
Homo floresiensis wikipedia , lookup
Sociobiology wikipedia , lookup
Archaic human admixture with modern humans wikipedia , lookup
The Descent of Man, and Selection in Relation to Sex wikipedia , lookup
Before the Dawn (book) wikipedia , lookup
Homo erectus wikipedia , lookup
History of anthropometry wikipedia , lookup
Origins of society wikipedia , lookup
Homo naledi wikipedia , lookup
Homo heidelbergensis wikipedia , lookup
Evolutionary origin of religions wikipedia , lookup
Discovery of human antiquity wikipedia , lookup
Human evolutionary genetics wikipedia , lookup
Recent African origin of modern humans wikipedia , lookup
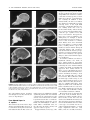
THE ANATOMICAL RECORD (PART B: NEW ANAT.) 276B:19 –26, 2004 FEATURE ARTICLE What Happened in the Origin of Human Consciousness? IAN TATTERSALL* At some point in its evolutionary history, our species Homo sapiens ceased to be a nonlinguistic, nonsymbolic organism, living in the world as presented to it by Nature, and instead began to exist in a world that it reconstructs in its own mind. Most scientists since Darwin have been content to explain this extraordinary transformation in human consciousness by the operation of natural selection. However, the human fossil and archaeological records indicate that modern human symbolic consciousness is not the culmination of the long trend that natural selection would predict. Instead, it shows that major change in the human past has been episodic and rare and that, as far as can be determined from the archaeological record, the passage from nonsymbolic to symbolic cognition is a recent event as well as an unprecedented one. So recent, indeed, that it significantly postdates the acquisition of modern human anatomy as expressed in skeletal structure. It, thus, appears most likely that the biological (neural) capacity underwriting the radically new behavioral mode arose as an incidental exaptation in the same process that produced the new skeletal structure of Homo sapiens, but that it lay unexpressed until it was “discovered” by means of a cultural innovation, plausibly the invention of language. As in the case of the modern anatomical structure, it appears that the new capacity was initially expressed in Africa and that its various behavioral potentials were sequentially discovered in a drawn-out process that is continuing today. An “accidental” origin of the human capacity helps understand why so many human behaviors have proven self-destructive and contradictory, a feature of our species that reductionist, selection-based scenarios are hard-put to explain. Anat Rec (Part B: New Anat) 276B:19 –26, 2004. © 2004 Wiley-Liss, Inc. KEY WORDS: human evolution; consciousness; symbolic cognition; exaptation; Neanderthals; intelligence; evolution INTRODUCTION There is undoubtedly something special about us humans, Homo sapiens. We are language-using, symbolically reasoning beings, whose relationship to the rest of the living world is, so far as we know, totally unlike that of any of the millions of other living species with whom we share our planet. In some elusive though all too real way, Dr. Tattersall is Curator in the Division of Anthropology at the American Museum of Natural History in New York City. A version of this article was originally written for publication in German in: T. Wabbel, editor. 2004. “Am Anfang war (k)ein Gott - Über die menschliche Existenz und andere Absurditäten.” Dusseldorf: PATMOS Publishers. *Correspondence to: Ian Tattersall, Division of Anthropology, American Museum of Natural History, Central Park West at 79th Street, New York, NY 10024. E-mail: [email protected] DOI 10.1002/ar.b.10041 Published online in Wiley InterScience (www.interscience.wiley.com). © 2004 Wiley-Liss, Inc. we stand apart from the rest of Nature, seeking to explain it, and worse, to manipulate and change it. Yet there can be no doubt that our origins lie firmly within the natural world. We are, in other words, directly descended from an ancestor that was neither linguistic nor rational (or irrational!) in the way in which we are. We started well on the other side of the narrow but deep gulf that now separates us from even the closest of our living relatives. HOW DID WE GET HERE? How could this extraordinary result have come about? By what remarkable means did our forebears contrive to cross that almost bottomless gulf? Most evolutionary biologists today seem to be content to put their faith in natural selection as the explanation. Common wisdom in evolutionary biology tells us that, over the eons all living creatures, ourselves included, have been finely honed by selection to an exquisite fit with their environments: that, in every generation, those individuals best adapted to prevailing circumstances have reproduced most successfully and, thus, have preferentially passed on their favorable heritable characteristics to the next one. To members of a brainy and egotistical species, it has been self-evident that our metabolically expensive large brains and expansive intelligence are a Good Thing. And it follows from this, does it not, that these human properties must have been consistently favored by natural selection over the millennia, finally culminating in the splendidly burnished cognition we so admire in ourselves today? Charles Darwin certainly thought so. To him, the constant pressure of natural selection over a vast span of time was the unambiguous explanation for modern human intelligence (Darwin, 1871). As far as he was concerned, our vaunted modern cognitive 20 THE ANATOMICAL RECORD (PART B: NEW ANAT.) capacities essentially represent the culmination of a longstanding trend in which smarter individuals have out-reproduced dumber ones. And so eminently reasonable does this scenario appear that few scientists have seen fit to question it. Vociferous objections have been heard, of course, from those with religious preconceptions; but those objections are grounded in a form of knowledge that lies entirely outside the purview of science. Within science, and most particularly within reductionist science, it has been more or less taken for granted that increasing intelligence was more or less inevitable once human precursors had taken up such intellectually demanding activities as stone tool making. Yet the putative agent of all this, natural selection, is not a creative force: it cannot be, because it can only promote novelties that are already in existence. In Nature, form has to precede function, if only because without form there can be no function. And natural selection cannot by itself conjure up anything new, no matter how advantageous the possession of a particular novelty might potentially be. There is no evident reason why human symbolic intelligence should have been an exception to this rule. Unlike his colleague, Darwin’s coinventor of the notion of evolution by natural selection, Alfred Russel Wallace, was evidently perturbed by considerations of this kind. While in all other ways even more Darwinian than Darwin himself on the subject of natural selection, Wallace could simply not see how selection could have brought about the extraordinary modern human cognitive faculty, with all of its ramifications of faith, superstition, and insight in addition to its ratiocinative aspects (Tattersall, 1998a). For Wallace, something other than natural selection had to be invoked in the mysterious acquisition of what Alex Marshack (1985) has termed “the human capacity.” In Victorian England, the guileless Wallace, too innocent to suspect fraud in others, turned to Spiritualism for an explanation; today it is necessary for scientists to look elsewhere. Where? Well, in Darwin’s and Wallace’s time, the human fossil and archaeological records barely existed. Now, however, they constitute a fulsome archive of the anatomies and inferred behaviors of our long-vanished kin, and they provide the obvious point of entry into this question. If we can understand the patterns of acquisition of notable anatomical and behavioral novelties among our forebears, perhaps we can begin to hazard some guesses about how we became the remarkable creatures that we are. And maybe we can begin to perceive whether Homo sapiens is indeed the burnished result of fine-tuning by natural selection over the eons (and thus that our evolution was by implication a more or less linear, directional affair), or whether we are alternatively the untidy product of an altogether more haphazard process. THE HUMAN STORY As far as is currently known, the hominid lineage separated from the one In Nature, form has to precede function, if only because without form there can be no function. leading to its closest great ape relative at around 7– 8 million years (myr) ago, probably in response to climatic changes in Africa that converted a relatively consistent forest cover throughout the continent into one into which woodland and grassland increasingly intruded, fragmenting the forest. The exact picture of events is cloudy in the first few million years of hominid existence, but one thing is clear: there are many ways to be a hominid, and the ancient hominids were busy exploring them all as new hominid species with varying characteristics emerged and went out to do battle in their own ways in the unstable ecological theatre. This set a pattern whereby multiple hominid species have typically occupied the world at any one point in time (see Figure 1). By some time over approximately 4 myr ago, however, a fairly standardized hominid morphology had emerged. Hominids of this period were FEATURE ARTICLE rather short-statured (around three to four feet tall), with relatively long arms and short legs and long, somewhat curved extremities. The pelvis was widely flaring, but the thorax tapered upward and the shoulders were narrow. The result was that, although these creatures were certainly upright bipeds when on the ground, they retained many features that would have facilitated climbing in the trees. This was not a transitional adaptation, but rather was a notably stable one that remained essentially unaltered for a couple of million years, even as new hominid species entered and departed the evolutionary stage. It was presumably in the opening period of hominid existence that our precursors acquired a taste for animal protein. The new woodland environment that our remote ancestors first exploited, with scattered trees interspersed by grassy spaces, would have presented both new dangers and new opportunities to these unusual creatures. When on the ground, these slow and essentially defenseless primates were more exposed to large predators than their arboreal ancestors had been; and there does exist good evidence that the early hominids were indeed the prey of both terrestrial and avian predators (e.g., Brain, 1981; Berger and Clarke, 1995). At the same time, though, the hominids would have had the enhanced opportunity, at least from time to time, to supplement a largely fruit-based diet not only with grassland tubers but also with the remains of dead herbivores. Some chimpanzee populations are known to hunt small mammals both cooperatively and with some regularity (e.g., Goodall, 1986), and there is no reason to think that ancestral hominids might not have done the same. What is more, the hominids would also have been able to vary their diet with scavenged remains with greater regularity (something in which chimpanzees are not apparently interested; see Stanford, 2003). What these possibilities actually meant in the lives of these smallbrained, large-faced creatures is more difficult to say, however; and there is no evidence in this early period that any hominid had cognitive or communicative abilities that exceeded those of today’s apes. And this, of course, is Figure 1. One possible scheme of relationships among species assigned to Hominidae, showing the consistent tendency to multiple branching throughout the family’s history that reflects the ongoing exploration of the multiple ways there are to be hominid. It is evidently an unusual situation for our species Homo sapiens to be the lone hominid extant. 姝 Ian Tattersall. 22 THE ANATOMICAL RECORD (PART B: NEW ANAT.) Figure 2. Median sagittal sections of various crania of extant hominoids and fossil hominids. a: Chimpanzee; b: Sterkfontein 5; c: Bodo; d: Kabwe; e: Petralona; f: Sima de los Huesos Skull 5; g: Guattari; h: extant Homo sapiens. Note particularly the contrast in cranial and facial form between the Guattari (Homo neanderthalensis) and Homo sapiens crania. From Bookstein et al. (1999), with permission. why paleoanthropologists nowadays increasingly refer to these ancient relatives as “bipedal apes.” From Bipedal Apes to H. sapiens The archaic body form of these bipedal apes notwithstanding, however, it is somewhere among this early hominid radiation that the first stone tool maker emerged, establishing a highly significant pattern that was evidently common in our evolutionary past: the arrival of new behavioral or technological innovations has not tended to coincide with the appearance of new kinds of hominid (Figure 1). This actually makes considerable sense, for the only place in which a novelty can appear is within a species. At approximately 2.5 myr ago, a hu- FEATURE ARTICLE man precursor (whose parents did not do this) spontaneously began to strike sharp flakes from small river cobbles: a feat that no living ape has yet been induced to replicate, even with intensive coaching. In addition to their manual skills, the first tool makers also displayed considerable foresight, carrying stones of the right kind around for considerable distances before making them into tools as needed. Here finally, without doubt, were creatures that had advanced beyond the ape cognitive level, and whose new behavior had opened a huge vista of new possibilities— enabling them, for example, to detach whole limbs from carcasses and to retreat to safer locations to consume them. What’s more, these hominids also used cobbles to pound on limb bones, cracking them to expose the nutritious marrow within. Among all competing hunters and scavengers, only hyenas had the massive jaw apparatus to be interested in this particular resource and, thus, to be a special danger to their early hominid competitors at kill sites. It would be a million years before a significant advance was made on those initial small but razor-sharp flakes that had ushered in humanity’s technological career. And meanwhile a wholly new kind of hominid had appeared. Now usually known as Homo ergaster, this human precursor was distinctly different from the more archaic hominids in having a body that was generally like our own in most features. But it still had a smallish brain (up to half as large as our own, compared with its ancestors’ one third), and a face that jutted noticeably (e.g., Walker and Leakey, 1993; Figure 2). Still, it was a radically new kind of hominid, whose body structure finally emancipated it from the forest edges and woodlands to which its forerunners had been confined. Hard on the heels of achieving this new body form, hominids had expanded out of Africa and into the farthest reaches of Asia. Yet it was not until several hundred thousand years (kyr) after their appearance that these new-model hominids started to make a new type of tool. This was the socalled “Acheulean handaxe,” a much larger implement than the early flakes, and one carefully formed by FEATURE ARTICLE flaking on both sides to a symmetrical tear-drop shape. For the first time, hominids were making tools to a “mental template” held in their minds before manufacture started, rather than simply going for an attribute: a sharp cutting edge, independent of the actual shape of the tool. This, too, must have represented a cognitive leap forward; yet we have no inkling of what difference this made in the wider existences of the tool makers, and it is pretty certain that, no matter how like yourself such creatures might have looked at a distance, if you met one you would have discovered little in common intellectually. It is, for example, highly unlikely that such creatures spoke, although it’s possible that they had quite sophisticated vocal/gestural communication (e.g., Corballis, 2002). At 1.5 myr ago, then, and despite having acquired both an anatomy (adapted for a striding gait out on the open savanna) and a behavior (stone tool making) that clearly had profound long-term consequences, hominids were still well integrated with the rest of nature: despite their peculiarities, they had certainly not crossed the bottomless gulf of which I spoke earlier. Indeed, this crossing was to be a very much later development. For, over the next million years or so, successive hominid species merely tended to do what their forebears had done, if perhaps a little better. By approximately half a million years ago, though, hominids were on the scene that had brains up into the range of modern humans, although still well below the modern average. And by not long after this time, we start to find the earliest evidence of the building of artificial shelters and of the domestication of fire in hearths (Klein, 1999). We also find, miraculously preserved at the German site of Schoeningen, the first finely crafted wooden throwing spears, made of a material that normally fails to be preserved (Thieme, 1997). These javelin-like spears, two meters and more in length, suggest that the hunting techniques and abilities of their makers were a good bit more sophisticated than one would have guessed from the stone tool record alone; for it is only in the THE ANATOMICAL RECORD (PART B: NEW ANAT.) 23 Figure 3. Comparison of a composite Neanderthal skeleton (left) with that of a Homo sapiens of similar stature (approximately 5 feet 6 inches; 167 cm). Note the multiple differences throughout the skeleton and particularly in the proportions of the pelvis and rib cage. Courtesy of Ken Mowbray. period between approximately 300 and 200 kyr ago that we find a radically new approach to the manufacture of stone tools. This was the socalled “prepared-core” technique, in which a stone “core” was carefully shaped with multiple strikes until a single blow would detach a flake that needed little modification into a finished tool (e.g., Schick and Toth, 1993). This was a reflection, one could guess, of yet another cognitive advance; but what that advance suggested of its possessor’s subjective experience of the world it is impossible to say. All we can hazard is that, throughout this period, hominids’ relationship to the world around them was becoming more complex; but these advances nonetheless seem to be signaling a refinement of what had gone before, rather than a radical departure from earlier ways of doing business. 24 THE ANATOMICAL RECORD (PART B: NEW ANAT.) Neanderthals and Cro-Magnons Perhaps the most sophisticated practitioners of the prepared-core stone tool-making technique were the Neanderthals, a group of hominids—Homo neanderthalensis—who left a remarkable record of themselves in Europe and western Asia in the period between approximately 200 and 30 kyr ago. The Neanderthals had brains that were fully as large as our own but that were housed in very different-looking skulls (see Tattersall and Schwartz, 1998, 2000), which were situated atop bodies that, while basically similar to ours, were also characteristically different from those of Homo sapiens, as is made abundantly evident by the first restoration of a composite Neanderthal skeleton, recently completed by G.J. Sawyer and Blaine Maley at the American Museum of Natural History (Figure 3). A recent study (Bailey, 2002) has also emphasized the significant dental differences that separate Homo neanderthalensis from Homo sapiens and, indeed, from all other fossil hominids examined. The last surviving descendants of an endemic European radiation of hominids, the Neanderthals were abruptly replaced by invading Homo sapiens over the period between approximately 40 and 30 kyr ago. The ultimate origin of the invaders, known as the Cro-Magnons, probably lay somewhere in the African continent, in the time between approximately 200 and 150 kyr ago. It is well established that biological novelties are likely to become fixed only in very small populations, and it seems very plausible that the anatomically distinctive Homo sapiens was born out of an episode during which the ancestral human population was reduced to very small numbers, perhaps even in the low thousands, as a result of environmental stress during the climatic vagaries of the Ice Ages. This episode has been most firmly inferred on the basis of comparative studies of modern human mitochondrial DNA (mtDNA), which is substantially less variable among modern humans than it is, for example, among the far less numerous and more geographically restricted modern chimpanzees (Gagneux et al., 1999. Kaessman et al., 2001). What is more, human mtDNA variation is greatest today in the African continent, suggesting that diversification has been continuing there longer than elsewhere in the world (Cann et al., 1987). The logical conclusion, referred to commonly as the “Out of Africa” hypothesis, thus places the origin of all modern humans in the African continent. Indeed, mtDNA lineages from other continents appear to be descended from lineages that originated in Africa, in which continent the fossil record also hints that human origins lie (see Stringer and McKie, 1996). However, the number of mtDNA sequence differences between Neanderthals and modern humans is significantly higher than the number of differences among humans—so much so that the earliest common ancestor between Neanderthals and Cro-Mag- There is little reason to believe that the Neanderthals possessed symbolic reasoning abilities, or articulate language, as the Cro-Magnons so clearly did. non man would appear to predate considerably Cro-Magnon’s appearance in Europe and other sites where the H. neanderthalensis fossil record exists (Krings et al., 1997). On the basis of this analysis, Neanderthals are close evolutionary cousins of modern man at best, and not likely at all to be our direct evolutionary ancestors (Paäbo, 1999). The contrast between the lives of the Neanderthals and the Cro-Magnons could not be more dramatic (White, 1989; Tattersall, 1995). Although the Neanderthals made beautiful stone tools, at least occasionally practiced the simple burial of the dead, protected disadvantaged members of their society, and managed to survive in very harsh environmental conditions, their lives (at least before contact with the Cro-Magnons) were FEATURE ARTICLE bereft of symbolic activities. On the other hand, the Cro-Magnons established themselves in Europe with the full panoply of modern behaviors at their disposal, making implements out of soft materials such as bone and antler as well as out of stone. Well over 30 kyr ago, they were already painting powerful animal images on the walls of caves. They made notations (perhaps including lunar calendars) as well as delicate engravings on bone and stone plaques; and some of the most elegant carvings ever made date from this time. The Cro-Magnons made music on bone flutes with subtle sound capabilities, and doubtless sang and danced as well. They buried their dead in elaborately decorated clothing, and with a plethora of grave goods. By soon after 30 kyr ago, eyed bone needles announce the arrival of couture, and at the same time, ceramic technology was invented, clay figurines being baked in simple but highly effective kilns. The list could go on and on, but it should already be evident that the Cro-Magnons were us, not only in their anatomy but in all their behavioral complexities as well (Tattersall, 1998b). They had crossed the gulf. The same cannot be said for the Neanderthals, who could best be described as an improved version of their forebears. There is little reason to believe that the Neanderthals possessed symbolic reasoning abilities, or articulate language, as the Cro-Magnons so clearly did. THE ARRIVAL OF MODERN HUMAN COGNITION Perhaps surprisingly at first thought, this limitation also apparently applied to the earliest anatomically modern Homo sapiens. The fossil record of the emergence of Homo sapiens is less than ideal, but it is clear that the earliest members of our species who were skeletally exactly like us had not yet traversed the behavioral threshold whose crossing so dramatically distinguishes us today. This fact is most clearly illustrated by the Homo sapiens who lived in Israel some 100 kyr ago. For these people were culturally indistinguishable from the Neanderthals with whom they somehow partitioned the Levantine environment for upward of 50 kyr. Modern human be- FEATURE ARTICLE haviors came on the scene substantially after the distinctive modern human anatomy. So what was going on? Recall, as I have already mentioned, that the only place a novelty can arise is within a species. The distinctive modern human anatomy, with its tall, slender body and its delicate face tucked beneath a high, globular skull, was probably acquired in a small, isolated hominid population in Africa at some time over 150 kyr ago. When conditions were right, this population then expanded to other areas of the Old World, although obviously this expansion cannot in early phases be attributed to the acquisition of modern human cognition. However, it nonetheless seems most likely that the developmental reorganization expressed in the new skeletal structure also involved some changes in the association areas of the brain. Exactly what those innovations were is a matter of debate, made all the more complicated by the fact that the human brain is a remarkably plastic organ capable of “rewiring” itself both in early development and later in life, depending on the function demanded of it. And the brain of the first anatomical Homo sapiens must certainly have been “back-compatible,” because if a new cognitive potential was acquired with the new anatomy, this potential does not appear to have been exploited for many thousands of years, archaic patterns of behavior persisting more or less unchanged for a very long time. Strange though this might seem, it actually reflects a general pattern in evolutionary history. I have already noted that, in evolution, form must precede function and that innovation itself cannot be driven by natural selection. Any novelty has to arise spontaneously as an exaptation, a structure existing independently of any new function for which it might later be co-opted. Only once such a structure has been established may it assume a new role in the life of its possessor and thus become an adaptation to that function. One of the most famous examples of this phenomenon is the feathers of birds. These new structures were used for insulation by ancestral birds for millions of years before they eventually made flight possible. THE ANATOMICAL RECORD (PART B: NEW ANAT.) 25 Perhaps, then, it is reasonable to infer that the neural substrate for our vaunted modern cognition first arose as a byproduct of the physical reorganization that gave birth to our anatomically distinctive species. This new substrate then lay fallow for a long period of time before its potential was “discovered” by its possessors. How this discovery was made is anybody’s guess, although it must have been through a cultural invention. The most plausible candidate for such an invention is language, for this activity is almost synonymous with the symbolic reasoning that marks us off from even our closest relatives in nature. Language involves forming intangible symbols in the mind, and it allows us to combine those symbols in new ways. And it also allows us to pose the “what if” questions that allow us to relate to the world around us in new and unprecedented ways. It is this It is this leap to symbolic manipulation in the mind that most truly marks us off from other forms of life on Earth. leap to symbolic manipulation in the mind that most truly marks us off from other forms of life on Earth; and the ability to do this is evidently something that arose rather abruptly, as a byproduct of something else, rather than through a process of gradual fine-tuning over the generations. Our intelligence is an emergent quality, the result of a chance combination of factors, rather than a product of Nature’s engineering. Yes, as Darwin perceived, our vaunted mental capacities are indeed the eventual outcome of a long (and rather untidy) process of neural accretion over the eons; but, as Wallace equally saw, this extraordinary acquisition was not— could not have been— driven by natural selection. Homo sapiens as we know it today is not the outcome of a long and inexorable process of perfection through natural selection, much as we often like to think of ourselves in that way. Where did modern human cognition emerge? Almost certainly in Africa, like modern human anatomy. For it is in this continent that we find the first glimmerings of “modern” behaviors. From Blombos Cave, near the continent’s southern tip, comes the first indisputably symbolic object, a geometrically engraved ochre plaque almost 80,000 years old (Henshilwood et al., 2002). From the same time range in Africa come the earliest pierced shells, evidence of (symbolic) body ornamentation. There is also early documentation of such typically human activities as long-distance trade in valued materials and flintmining (McBrearty and Brooks, 2000). There is nothing in the early African archaeological record as impressive as that of the Cro-Magnons in Europe; but this latter record is much later, and there is no reason to expect that the multifarious dimensions of the underlying human capacity should have been discovered all at once. Indeed, it is scarcely credible that they could have been. For the human capacity is a generalized one, with boundless—and often contradictory—possibilities (Tattersall, 1998b). In this sense, there must have been a threshold effect in the acquisition of fully modern human cognition, just as there must be in the spread within populations of genetic innovations (Schwartz, 1999). Certainly, the archaeological record before 50 – 80 kyr ago contains isolated, sporadic expressions, such as the 250-kyr-old Berekhat Ram “figurine” from Israel, that one can—if one wishes—interpret as evidence of symbolic cognition in some form (see Marshack, 1997). But are such objects reliable evidence of a fully formed human sensibility? No; for even if they do indeed represent stirrings of the human symbolic capacity, they remain irrelevant to the greater human condition as long as they were the products of eccentric individuals, rather than reflections of the larger societies in which those individuals lived. And there is no evidence that any early objects of this kind embodied wider social values. The long and elaborate process of discovering the human capacity had evi- 26 THE ANATOMICAL RECORD (PART B: NEW ANAT.) dently begun in Africa by around 80 kyr ago (and possibly well before that). And the Cro-Magnons, with their art, music, ornamentation, and all their other evident cognitive and social complexities, had very clearly crossed the critical threshold, however, one might wish to define it. But the moment of transformation still eludes us and may well do so almost indefinitely. For the process of becoming human is ongoing; it is one that is still unfolding today, as technological and artistic innovation, highly sporadic before the emergence of the human capacity, becomes routine in daily human experience. LITERATURE CITED Bailey SE. 2002. A closer look at Neanderthal postcanine dental morphology: The mandibular dentition. Anat Rec (New Anat) 269:148 –156. Berger LR, Clarke RJ. 1995. Eagle involvement in accumulation of the Taung Child fauna. J Hum Evol 29:275–299. Bookstein F, Schafer K, Prossinger H, Seidler H, Fieder M, Stringer C, Weber GW, Arsuaga JL, Slice DE, Rohlf FJ, Recheis W, Jara M, Marcus LF. 1999. Comparing frontal cranial profiles in archaic and modern Homo by morphometric analysis. Anat Rec (New Anat) 257:217–224. Brain CK. 1981. The hunters or the hunted? Chicago IL: Chicago University Press. Cann R, Stoneking M, Wilson A. 1987. Mitochondrial DNA and human evolution. Nature 325:31–36. Corballis M. 2002. From hand to mouth. Princeton, NJ: Princeton University Press. Darwin C. 1871. The descent of man. London: John Murray. Gagneux P, Wills C, Gerloff U, et al. 1999. Mitochondrial sequences show diverse evolutionary histories of African hominoids. Proc Natl Acad Sci U S A 96:5077– 5082. Goodall J. 1986. The chimpanzees of Gombe: Patterns of behavior. Cambridge, MA: Harvard University Press. Henshilwood CS, d’Errico F, Yates R, et al. 2003. Emergence of modern human behavior: Middle Stone Age engravings from South Africa. Science 295:1278 – 1280. Kaessmann H, Wiebe V, Weiss G, Paäbo, S. 2001. Great ape DNA sequences reveal a reduced diversity and an expansion in humans. Nat Genet 27:155–156. Klein R. 1999. The human career. 2nd ed. Chicago: Chicago University Press. Krings M, Stone A, Schmitz RW, Krainitzki H, Stoneking M, Paäbo S. 1997. Neandertal DNA sequences and the origin of modern humans. Cell 90:19 –30. Marshack A. 1985. Hierarchical evolution of the human capacity. New York: American Museum of Natural History. Marshack A. 1997. The Berekhat Ram figurine: A late Acheulian carving from the Middle East. Antiquity 71:327–337. McBrearty S, Brooks A. 2000. The revolution that wasn’t: A new interpretation of the origin of modern human behavior. J Hum Evol 39:453–563. FEATURE ARTICLE Paäbo S. 1999. Human evolution. Trends Cell Biol 9:M13–M16. Schick KD, Toth N. 1993. Making silent stones speak: Human evolution and the dawn of technology. New York: Simon & Schuster. Schwartz JH. 1999. Sudden origins: Fossils, genes and the emergence of species. New York: John Wiley & Sons. Stanford C. 2003. Upright: The evolutionary key to becoming human. Boston: Houghton Mifflin. Stringer C, McKie R. 1996. African exodus: The origins of modern humanity. New York: Henry Holt. Tattersall I. 1995. The last Neanderthal: The rise, success and mysterious extinction of our closest human relative. New York: Macmillan. Tattersall I. 1998a. The origin of the human capacity. New York: American Museum of Natural History. Tattersall I. 1998b. Becoming human: Evolution and human uniqueness. New York: Harcourt Brace. Tattersall I. 2002. Adaptation: The unifying myth of biological anthropology. Teaching Anthropology: SACC Notes 9:9 –11, 39. Tattersall I, Schwartz J. 1998. Morphology, paleoanthropology, and Neanderthals. Anat Rec (New Anat) 253:113–117. Tattersall I, Schwartz JH. 2000. Extinct humans. Boulder, CO: Westview Press. Thieme H. 1997. Lower Palaeolithic hunting spears from Germany. Nature 385: 807–810. Walker A, Leakey R. 1993. The Nariokotome Homo erectus skeleton. Cambridge, MA: Harvard University Press. White R. 1989. Visual thinking in the Ice Age. Sci Am 260:92–99.