Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
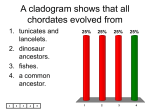
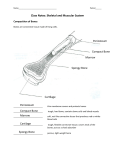
4/15/2009 The Chordates: Characteristics Structural Plan The name Chordata comes from the notochord, rodlike, semirigid tissue enclosed in a sheath. The chordates have five basic characteristics: Chapter 23 Chordates n n n n n 1) dorsal, tubular nerve cord overlying 2) a supportive notochord 3) pharyngeal slits 4) an endostyle for filter feeding 5) a postanal tail for propulsion Copyright © The McGrawHill Companies, Inc. Permission required for reproduction or display. Shared Features Classification of the Chordates Invertebrates n bilateral symmetry, anterioposterior axis, coelom, tubewithinatube body plan, metamerism and cephalization. Characteristics common within the group n radial cleavage, anus derived from the blastopore, mouth derived from a secondary opening, and a coelom formed by fusion of enterocoelous pouches Phylogenetic tree of the chordates, suggesting probable origin and relationships. Bulging and thinning of lines indicated the relative abundance in numbers of species of each group through geologic time as indicated by the fossil record. Most people recognize the general traditional groups; but cladists no longer use Agnatha and Reptilia. Reptiles are paraphyletic because they do not contain all of the descendents. n n n Reptiles, birds and mammals compose a monophyletic clade called Amniota. By cladistics, reptiles can only be grouped as amniotes that are not birds or mammals. There are no positive features that group only reptiles to the exclusion of birds and mammals. For the same reasons, agnathans (hagfishes and lampreys) are paraphyletic because their most common recent ancestor is also an ancestor of all remaining vertebrates. Cladogram of living members of the Phylum Chordata. 1 4/15/2009 Traditional classification makes certain distinctions Protochordata (or Acraniata) are separated from the Vertebrata (or Craniata) that have a skull. Vertebrates may be divided into Agnatha (jawless) and Gnathostomata (having jaws). n Gnathostomata is subdivided into Pisces with fins and Tetrapoda, usually with two pair of limbs. Five Chordate HallmarksNotochord This feature as well as the other three is always found at some embryonic stage of all chordates. The notochord is the first part of the endoskeleton to appear in the embryo. It serves as an axis for muscle attachment; it can bend without shortening and permits undulation. Vertebrates are also divided into Amniota, having an amnion, and Anamniota lacking an amnion. Many of these groupings are paraphyletic; alternative monophyletic taxa are suggested. Some cladistic classifications exclude Myxini (hagfishes) from the group Vertebrata Notochord In protochordates and jawless vertebrates, the notochord persists throughout life. In vertebrates, a series of cartilaginous or bony vertebrae form from mesenchymal cells derived from blocks of mesodermal cells lateral to the notochord. In most vertebrates, the notochord is entirely displaced by vertebrae although it remains persistant as the intervertebral discs. Pharyngeal Pouches and Slits Pharyngeal slits lead from the pharyngeal cavity to the outside. They form by the inpocketing of the outside ectoderm and the evagination of the pharynx endoderm. In aquatic chordates, the two pockets break through to form the pharyngeal slit. In amniotes these pockets may not break through and only grooves are formed. Dorsal Tubular Nerve Cord In most invertebrate phyla, the nerve cord is ventral to the alimentary canal and solid. In chordates, the single cord is dorsal to the alimentary canal and is tubular. The anterior end enlarges to form the brain. The cord is produced by the infolding of ectodermal cells on the dorsal side of the body. Pharyngeal Pouches and Slits In tetrapods, the pharyngeal pouches give rise to a variety of structures, including the Eustachian tube, middle ear cavity, tonsils and parathyroid glands. The perforated pharynx functions as a filter feeding apparatus in protochordates. Pharyngeal pouches or slits are not unique to chordates; hemichordates also have pharyngeal slits. Fishes added a capillary network with gas permeable walls; this network evolved into gills. 2 4/15/2009 Endostyle or Thyroid Gland Recently, the endostyle was recognized as a shared chordate character. The endostyle or its derivative, the thyroid gland, is found in all chordates. Some cells in the endostyle secrete iodinated proteins homologous with the iodinatedhormonesecreting thyroid gland of adult lampreys and the remainder of vertebrates. Subphylum Urochordata: Tunicata Diversity There are about 3000 species of tunicates identified. They occur in all seas and at all depths. Most are sessile as adults although a few are freeliving. The tunic is the tough, nonliving test that surrounds them and contains cellulose. In most species, only the larval form bears all the chordate hallmarks; adults lose many of these characters. Postanal Tail The postanal tail, plus musculature, provided motility for larval tunicates and Amphioxus to swim. This was increased in fishes but became smaller or vestigial in later lineages. Form and Function of Ascidians They are called sea squirts because they discharge a jet of water when disturbed. Most are attached when adults, and they are among the most abundant intertidal animals. Colonial and solitary ascidians have their own test; compound forms share a common test. In some compound ascidians, each has its own incurrent siphon but share the excurrent siphon. The mantle lines the tunic. The incurrent siphon marks the anterior; the excurrent siphon marks the dorsal side. Form and Function Form and Function Water entering the incurrent siphon passes through a ciliated pharynx with an elaborate basketwork. Feeding depends on the formation of a mucous net that is secreted by the endostyle. Cilia on gill bars of the pharynx pull the mucus into a sheet; particles trapped in the sheet are worked into a rope and carried back to the esophagus and stomach. The terminal part of the gut, referred to as the intestine, actually is not homologous with the intestine of other chordates, but rather with the hepatic cecum of Amphioxus. The heart drives blood first in one direction, then in reverse. The nervous system has one nerve ganglion and a plexus of nerves on the dorsal side of the pharynx. 3 4/15/2009 Form and Function Sea squirts are hermaphroditic with a single ovary and a single testes; fertilization occurs in the water. Adult sea squirts only have one of the five chordate features: pharyngeal slits. The tadpole larvae, however, have all five chordate characteristics. The larva does not feed, but swims awhile before attaching and developing into a sessile adult. Form and Function Amphioxus has the four distinctive characteristics of chordates in simple form. Water enters mouth driven by cilia in the buccal cavity. Water passes through pharyngeal slits where food is trapped in mucus. Food particles separated from the mucus are passed into a hepatic cecum where they are phagocytized. Filtered water leaves the body by an atriopore. Reproduction Sexes are separate. Sex cells are set free in the atrial cavity and pass out the atriopore where fertilization occurs outside. Cleavage is holoblastic and a gastrula forms by invagination. The larvae soon hatch and gradually become the shape of adults. Subphylum Cephalochordata Diversity Lancelets are slender, laterally flattened, translucent animals about 57 cm long. They live in sandy bottoms of coastal waters Originally labeled in the genus Amphioxus, they are by priority now in the genus Branchiostoma. About 25 species of amphioxus are described; four occur in North American coastal waters. Form and Function The closed circulatory system is complex but lacks a heart. Blood is pumped by peristaltic contractions in the ventral aorta, passes upward through branchial arteries in the pharyngeal bars to paired dorsal aortas. Their blood moves by microcirculation through tissues and returns to the ventral aorta. Blood lacks erythrocytes and hemoglobin and mainly transports nutrients. A hollow nerve cord lies above the notochord. Pairs of spinal nerve roots emerge at each trunk segment. Sense organs are simple, unpaired bipolar receptors located in various parts of the body. Basic Plan Amphioxus possesses features that suggest the vertebrate plan. A cecum is a diverticulum resembling the vertebrate pancreas that secretes digestive enzymes. n The trunk muscles suggest future vertebrate patterns. n They possess the basic circulatory plan that advanced chordates elaborate. n Many zoologists consider amphioxus a living descendant of ancestors that gave rise to both cephalochordates and vertebrates. This would make them the sister group of the vertebrates. 4 4/15/2009 Subphylum Vertebrata (Craniata) Adaptations That Guided Vertebrate Evolution The earliest vertebrates were substantially larger than the protochordates. The earliest vertebrates were also considerably more active than the protochordates. The earliest vertebrates where characterized by increased speed and mobility resulting from modifications of the skeleton and muscles. The higher activity level and size of vertebrates also requires structures specialized in the location, capture, and digestion of food and adaptations designed to support a high metabolic rate. Vertebrate Skeletal Systems Information on this topic drawn heavily from Chapter 29 The vertebrate endoskeleton is composed of bone and cartilage; it protects and supports, is the major reservoir for calcium and phosphorus, and is the origin of blood cells. Except in jawless vertebrates, the notochord is surrounded or replaced by backbone during development. Bone Bone is living tissue with significant deposits of calcium salts in an extracellular matrix. Bone is nearly as strong as cast iron, yet only onethird as heavy. Bone is always laid down in a “replacement area” formed by some connective tissue. Most bone develops from cartilage and is called endochondral or replacement bone. Skeletal Modifications Most vertebrates possess both an exoskeleton and endoskeleton of cartilage or bone. The endoskeleton permits almost unlimited body size with much great economy of building materials. The endoskeleton forms excellent jointed scaffolding for the attachment of segmented muscles. Cartilage Cartilage is a major skeletal element of jawless fishes and elasmobranchs; the cartilage is a derived feature since their Paleozoic ancestors had bony skeletons. Hyaline cartilage is the basic form of cartilage made of cartilage cells surrounded by firm complex protein gel interlaced with a meshwork of collagenous fibers. Blood vessels are missing in cartilage; this is the reason many sport injuries heal poorly. Hyaline cartilage also makes up articulating surfaces of bone joints of adult vertebrates, as well as supporting tracheal, laryngeal and bronchial rings. Bone Embryonic cartilage is eroded, leaving honeycomb spaces that are invaded by bone forming cells. Boneforming cells deposit calcium salts around the strandlike remnants of the cartilage. Intramembranous bone develops from sheets of embryonic cells, mainly the face and cranium. Cancellous, or spongy, bone has an open, interlacing framework of bony tissue. Some bones proceed to add additional salts to become compact bone. 5 4/15/2009 Microscopic Bone Structure Microscopic Structure of Bone Compact bone has a calcified bone matrix arranged in concentric rings. The rings contain cavities (lacunae) filled with bone cells (osteocytes). Lacunae are interconnected with canaliculi passages that distribute nutrients. The whole cylindrical structure is an osteon or haversian system. Fig. 29.7b Osteoclasts slowly resorb bone while osteoblasts deposit additional bone. These simultaneous processes allow the growth of a rigid structure without any weakening. Parathyroid hormone stimulates bone resorption; calcitonin inhibits bone resorption. Both hormones, together with a derivative of vitamin D, maintain constant blood Ca ++ levels. Bone growth also responds to usage; astronauts living without gravity suffer bone loss. Fig. 29.7c Effect of Body Size on Bone Stress Crosssectiontovolumeratio Consider one animal twice as long, wide and tall as a second animal. n n n The larger animal would have 8X the volume and 8X the weight. However, the crosssectional area of bones, tendons and muscles would be 4X greater. Therefore, 8X the weight would have to be carried by 4X the strength. Evolution of Vertebrate Skeletal Systems The endoskeleton probably as composed initially of cartilage and later gave rise to bone. Endoskeleton of living hagfishes, lampreys, sharks and their kin, and in some “bony” fishes, like sturgeons, is mostly composed of cartilage. The structural strength of bone is superior to cartilage, making it ideal for muscle attachment in areas of high mechanical stress. Perhaps bone evolved, in part, as a means of mineral regulation (since P and Ca ++ are used for many physiological processes and are in particularly high demand in organisms with high metabolic rates). Effect of Body Size on Bone Stress Mammalian bone is uniform per crosssectional area and this places an upper limit on size. Bone shape does not change much; mammals adapted by shifting posture and alignment. Mechanical advantage can be gained by aligning weight with ground reaction forces, as is the case with an upright horse; beyond the size of a horse, not much advantage can be gained. Elephants and large dinosaurs had thick and robust bones but this decreases running speed. Evolution of Vertebrate Skeletal Systems Some of the most primitive fishese, including Ostracoderms and placoderms were partly covered in a bony, dermal armor (this armor is modified in later fishes as scales). Many of the bones encasing the brain of advanced vertebrates develop from tissue that originates from the dermis. Most vertebrates are further protected with keratinized structures derived from the epidermis, such as reptilian scales, hair, feathers, claws, and horns. 6 4/15/2009 Bony Integumentary Structures Muscular Movement Most animal movement relies on a single fundamental mechanism: contractile proteins. Contractile machinery is composed of ultrafine fibrils: fine filaments, striated fibrils or tubular fibrils. All are arranged to contract when powered by ATP. The most important protein contractile system is composed of actin and myosin. The actomyosin system is almost universal and is found from protozoa to vertebrates. Striated Muscle Structure Striated muscle is named for the periodic bands visible under the light microscope. Each cell or fiber is a multinucleated tube with many myofibrils packed together. The cell membrane folds in to form the sarcolemma. A myofibril contains thick filaments of protein myosin and thin filaments of protein actin. The sarcomere extends between successive Z lines (dense structure that holds thin filaments together). Repeated, highintensity, short exercises cause synthesis of additional actin and myosin. Endurance exercise develops more mitochondria, myoglobin and capillaries. Thin Filaments Thin filaments are composed of three different proteins. The backbone is a double strand of actin twisted into a double helix. Surrounding the actin filament are two strands of tropomyosin that lie in the grooves of actin. Each tropomyosin is a double helix. The third protein of the thin filament is troponin, a complex of three globular proteins. Troponin is a calciumdependent switch that acts as the control point in contraction. Thick Filaments Each thick filament is made of myosin molecules packed together in a bundle. Each myosin molecule has two polypeptide chains, each having a clubshaped head. The double heads of each myosin molecule face outward from the center of the filament. The heads act as molecular cross bridges that interact with the thin filaments in contraction. Sliding Filament Hypothesis of Muscle Contraction According to this model, the thick and thin filaments link together by molecular cross bridges. They then act as levers to pull the filaments past each other. The cross bridges on the thick filaments snap rapidly backandforth, attaching and releasing from special receptor sites on the thin filaments. This ratchet action draws the Z lines together. 7 4/15/2009 Sliding Filament Hypothesis of Muscle Contraction All sarcomere units shorten together as the muscle contracts. Relaxation is passive; when the cross bridges between thick and thin filaments release, the sarcomeres are free to lengthen. This requires some force, usually supplied by antagonistic muscles or by gravity. See figure 2918 for description of attachpullrelease cycle Fast Fibers One type of fast fiber lacks efficient blood supply and high density of mitochondria and myoglobin; they are pale in color and, function anaerobically and fatigue rapidly. n White meat of chicken Another kind of fast fiber has the extensive blood supply, mitochondria and myoglobin and functions aerobically and can sustain exercise for long periods of time. n Geese, dogs and ungulates limb muscles The cat family has running muscles made up almost entirely of fast fibers that operate anaerobically. n Such muscles rapidly build up an oxygen debt; cheetahs must rest 3040 minutes after a chase. Chordate Physiology Vertebrates have modifications to the digestive, respiratory, circulatory, and excretory systems required to meet an increased metabolic demand. The perforated pharynx evolved as a filterfeeding device in early chordates. Water with suspended food particles was drawn through the pharynx by ciliary action and trapped by mucous secreted by the endostyle. In the larger, predatory vertebrates, the pharynx was modified into a muscular apparatus that pumped water through the pharynx. With the origin of highly vascularized gills, the function of the pharynx shifted to primarily gas exchange. Muscle Performance Fast and Slow Fibers Skeletal muscles have several types of fibers. Slow fibers are specialized for slow, sustained contractions without fatigue (e.g. posture). Slow fibers constitute red muscles because they contain a rich blood supply, a high density of mitochondria, and abundant stored myoglobin oxygen reserves. Two kinds of fast fibers provide fast, powerful contractions. Importance of Tendons in Energy Storage During walking and running, kinetic energy is stored from step to step. A kangaroo also uses the recoil of energy in tendons to bound along; therefore each movement does not have to rely on alternate muscle contractions and relaxations. Elastic storage occurs in legs of grasshoppers and fleas, wing hinges of flying insects, hinge ligaments of bivalve molluscs, and the dorsal ligament that supports the head of hoofed mammals. Other systems Changes in the gut, including a shift from movement of food by ciliary action to muscular action and additon of accessory digestive glands, the liver and pancrease were necessary to manage the increased amount of food ingested. A ventral threechambered heart consisting of a sinus venosus, atrium, and ventricle, and erythrocytes with hemoglobin enhanced transporation of nutrients, gases, and other substances. Protochordates have no distinct kidneys, but vertebrates possess paired, glomerular kidneys that remove metabolic waste products and regulated body fluids and ions. More detailed descriptions of these systems will follow in discussion of remaining classes of vertebrates. 8 4/15/2009 New Head, Brain, and Sensory Systems When vertebrate ancestors shifted from filter feeding to active predation, new sensory, motor, and integrative controls became essential for location and capture of larger prey. The anterior end of the nerve cord became enlarged as a tripartite brain (forebrain, midbrain, and hindbrain) and was protected by a cartilaginous or bony cranium. Paired special sense organs such as eyes evolved, along with paired inner ears designed for equilibrium and sound reception. Many other receptors also evolved: mechanoreceptors, chemoreceptors, electroreceptors, and olfactory receptors. Search for the Vertebrate Ancestral Stock Fossil invertebrate chordates are rare and known primarily from two fossil beds: the wellknown middle Cambrian Burgess Shales of Canada and the recently discovered early Cambrian fossil beds of Chengjiang and Haikou, China. The mix of vertebrate and protochordate characters in Haikouella suggests evolution of “vertebrate” soft characters before the evolution of an endoskeleton. Garstang’s Hypothesis of Chordate Larval Evolution The chordates have pursued two paths in their early evolution. One path led to the sedentary urochordates; the other to active, mobile cephalochordates and vertebrates. At the time of its discovery in 1869, the tadpolelike larva of tunicates was considered a descendant of an ancient freeswimming chordate ancestor. In 1928, Walter Garstang of England, however, suggested that the chordate ancestral stock was derived by retaining into adulthood the larval form of sessile tunicatelike animals. Ammocoete Larvae of Lampreys as a Model of the Primitive Vertebrate Body Plan Lampreys have a larval stage called the ammocoete that closely resembles the amphioxus. Ammocoete larvae were originally considered cephalochordate adults. The ammocoete larval mouth resembles the amphioxus but draws water in by muscular pumping. The endostyle, mucus, body muscles, notochord and circulatory system closely resemble amphioxus. Garstang’s Hypothesis Garstang termed this paedomorphosis, the evolutionary retention of larval traits in an adult body. Paedomorphosis occurs in some amphibians. His suggestion that evolution may occur at larval stages matches embryological evidence. Recently, the idea that the chordate ancestor was free swimming, based on phylogenies generated from molecular data has been revived. Basic Vertebrate Body Plan In contrast to amphioxus, ammocoetes have a two chambered heart, a threepart brain, a median nostril, auditory vesicles, a thyroid and pituitary gland. More extensive pharyngeal filaments serve in respiration. The ammocoete has a true liver, gallbladder and pancreatic tissue. In total, the ammocoete larva has the most primitive condition of this set of vertebrate structures. 9 4/15/2009 Earliest Vertebrates: Jawless Ostracoderms Found in the late Cambrian deposits in the United States, Bolivia and Australia, they were small, heavily armored, jawless, and lacked paired fins. Ostracoderms Although they possess many typically vertebrate characters, such as the Wshaped myomeres, a heart, cranium, and fin rays, they lack evidence of mineralized tissues The earliest Ostracoderms were armored with bone in their dermis and lacked paired fins that later fishes used for stability. The ostratocderms are not considered to be a natural evolutionary assemblage, but a convenience for describing several groups of heavily armored extinct jawless fishes, such as the heterostracans. Early Jawed Vertebrates All living and extinct jawed vertebrates are called gnathostomes in contrast to agnathans. Living agnathans, the lampreys and hagfishes, are often called cyclostomes. n Agnathans, defined by the absence of jaws, may be paraphyletic. Gnathostomes constitute a monophyletic group Placoderms appeared in the early Devonian and were heavily armored; some were large. Acanthodians contemporary with placoderms may have given rise to bony fishes. Evolution of Jaws Evidence indicates that jaws arose by modification of the first two cartilaginous gill arches. Both gill arches and jaws form from upper and lower bars that bend forward and are hinged. n Both are derived from neural crest cells rather than from mesodermal tissue as are most bones. n The jaw musculature is homologous to the musculature that originally supported gills. n The mandibular arch may have first become enlarged to assist gill ventilation, perhaps to meet the increasing metabolic demands of early vertebrates. n 10