Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
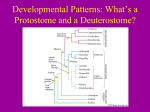
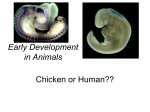
Frog & Sea Urchin Development BIOL212 1 Spring 2012 Frog & Sea Urchin Development Lab Objectives: • • • • • • • • • • • • • • • • Be able to recognize the various stages of animal development Compare and contrast the structural components of the blastula and gastrula embryo. Describe the purpose of gastrulation. Compare the process of gastrulation in the sea urchin and the frog. Describe the processes of fertilization and cleavage division in the sea urchin. Observe formation of the fertilization membrane as it expands to surround the sea urchin. Describe how the amount and distribution of cytoplasm within a fertilized egg influence the patterns of cleavage. Describe how cleavage occurs in an echinoderm and a frog. Compare the differences in development with and without the presence of a yolk Describe the structure of a typical blastula. Relate the structure of the frog blastula to the establishment of the dorso-ventral and antero-posterior axes of the embryo Describe the process of neurulation. Relate the development of the brain and the spinal column to the development of the neural tube and neural folds. Know differences related to external and internal early development Some species have one or more larval stages Relate commonalities to the Unity of Life Introduction: All living things must be able to reproduce and develop. In animals, gametes produced by the process of meiosis unite during fertilization to form a single diploid cell, the zygote. The processes of cell division, cell movement, cellular differentiation, and morphogenesis result in the development of a multicellular embryo that will grow to form an adult. The hereditary material within fertilized eggs, or zygotes, of animal organisms holds the key to both structure and function. DNA guides, through time, the structural and functional development of a single cell into an embryo and its morphogenesis into an adult. Even aging is a result of genetic programming. Biologists are continuing to understand more of the genetic control of development. Advanced technologies as well as molecular and recombinant DNA techniques have made it possible to discover more and more about the genomes of organisms and to map the entire genomes of some. Common patterns of gene function are beginning to emerge, suggesting that some day, perhaps, we will be able to explain the mysteries of animal development. Once again DNA exhibits its programming functions—DNA holds the secrets not only for “what” it produces, but also for “how” and “when” it produces its products. While Animal development is a complex process of sequential activation of genes, which in turn code for proteins that allow for the growth and differentiation of the organism, the early stages of animal development are, or at least appear, very similar across species. We have INTERCONNECTEDNESS REDUCE – REUSE – RECYCLE Frog & Sea Urchin Development BIOL212 2 Spring 2012 already seen the difference in Protostomes and Deuterostomes depending on whether the blastopore becomes the mouth or the anus. The presence or absence of a yolk imposes necessary differences on the common theme. You will examine prepared slides as well as live animal examples (if available) to get a sense of the changes that occur during early development. During this laboratory you will have the opportunity to explore the development of sea urchin and frog embryos. Advance Preparation: Review your notes on animal development from lecture and Sections 46.3 (p. 1005 – 1008), 4.5 (p. 1011 – 1015) and Chap. 47 (p. 1021 – 1044) Structures and terms to know: gametes, ovum, sperm, fertilization, zygote, cleavage, blastula, blastomeres, blastocoel, blastopore, mesenchyme cells, archenteron, gastrula, invagination, gastrulation, filopodia, animal pole, vegetal pole, dorsal, ventral, notochord, neural fold, neural plate, neural crest, neural tube, coelom, yolk sac, yolk stalk, somites, organogenesis, morphogenesis, holoblastic and meroblastic cleavage, totipotent, determination, differentiation, convergent extension, mouth, anus, fate map, ectoderm, endoderm, mesoderm, extra-embryonic membranes (amnion, chorion, yolk sac, allantois) polyspermy, vitelline membrane, Neurulation, homeobox Laboratory Procedures: You will find microscope slides of the stages of development of frogs and sea urchins and (hopefully) some live specimens to examine. For each organism you view under the microscope, you should draw and label the specimen. When you label the drawings, be sure to include all the structures or cells that you can identify on the specimen and the total magnification you used. To assist in tracking the stages, there will also be models available and attachments to these instructions. I. Slides 1. Prepared slides Microscope Slides: Frog [Order Anura (Merrem 1820)] and Sea Urchin [Strongylocentrotus sp. (Brandt, 1835)] Developmental Stages (see overhead: there may be a more detailed list.) 2. Live specimens Check overhead – if we have any live specimens, this and instructions will be announced! (If not available at this lab period, we will try to have them at the next lab period.) Part A: Fertilization and Early Development in Sea Urchins. Understanding how DNA regulates cell division, differentiation, and morphogenesis begins with observing the developing embryo. Sea urchins have long been the developmental biologist’s favorite organism of study because they are relatively simple to obtain and culture in the laboratory. The cells of the developing sea urchin are also fairly transparent, providing us with a limited ability to “look inside” the embryo. Early development of the sea urchin is under the genetic control not only of the zygote’s DNA but of messenger RNA (mRNA) stored in the egg during its development. These messages include maternal mRNAs synthesized from maternal DNA prior to the meiotic events of oogenesis—mRNAs made from DNA that may not be included in the egg itself. Thus, the story of development begins before fertilization. The unfertilized sea urchin egg is surrounded by a vitelline membrane that lies just above the surface of the cell’s plasma membrane. Within the cytoplasm, yolk granules (sea urchin eggs are microlecithal—they have very little yolk) can be observed. In addition to other cytoplasmic determinants and stored mRNAs, small cortical granules, composed of proteins and mucopolysaccharides, lie just beneath the plasma INTERCONNECTEDNESS REDUCE – REUSE – RECYCLE Frog & Sea Urchin Development BIOL212 3 Spring 2012 membrane (in the outer rim or cortex of the egg). When a single sperm enters the egg plasma membrane, the membrane potential quickly changes as a wave of depolarization spreads from the site of sperm entry. This reaction is often referred to as a “fast block to polyspermy,” since no additional sperm can gain entry following the change in membrane potential. Release of Ca2~ ions from the egg’s endoplasmic reticulum, in response to G protein, causes the cortical granules in the egg’s cortex to fuse with the plasma membrane. The cortical granules discharge their contents into the space between the plasma membrane and the vitelline membranes. The excess mucopolysaccharide now present in the perivitelline space lowers the water potential of that area, and water flows in. This causes the perivitelline space to increase in diameter, making it appear as if the vitelline membrane is lifting off the surface of the zygote. Addition of proteins to the vitelline membrane hardens (or “tans”) it as it is transformed into a fertilization membrane. Formation of the fertilization membrane (often called the “slow block to polyspermy”) offers additional protection against multiple sperm entry. Hundreds of sperm can usually be observed still attached to the old vitelline membrane, now the fertilization membrane. These will be removed by the action of enzymes released from the cortical granules. Following fertilization, the stored mRNAs are responsible for directing protein synthesis during the early stages of development—cleavage, blastula formation, and gastrulation. In addition to the mRNA and protein already present in the fertilized egg, newly synthesized protein products are responsible for establishing regional differences in the egg cytoplasm. These proteins or cytoplasmic determinants are responsible for establishing the cytoarchitecture of the unfertilized egg, reorganizing the cytoplasmic elements in response to fertilization, controlling the direction of the first cleavage divisions, and establishing the axis of the embryo. In this exercise, you will have the opportunity to observe the earliest stages of sea urchin development— beginning with fertilization. You will also have available later-stage embryos for observation of gastrulation and formation of pluteus larvae. Prepared Slides--Embryological Stages (and structures) Draw and label each of the slides indicated below, and answer the following questions. Sperm (sperm cells) a) How does the size of the sperm cells compare with the size of the unfertilized egg cells (be as specific as possible: indicate the magnification of the scope that you had to use.) Egg, unfertilized (egg cells) b) Is this cell haploid or diploid? Egg, fertilized (zygote, fertilization membrane) c) Is this cell haploid or diploid? d) How does the appearance of this cell compare with the appearance of the unfertilized egg cell? e) What structure causes the change in appearance? Part B: Cleavage In order for a fertilized egg or zygote to become a multicellular organism, the zygote must divide by mitosis. During early development, this process of division is known as cleavage. The fertilized egg is not a uniform sphere. Differential concentrations of cytoplasm and yolk (if present) can affect the cleavage process. The upper portion of the egg, usually richest in cytoplasm, is known as the animal pole, and the lower portion of the egg, containing more yolk, as the vegetal pole. The first plane of cleavage is vertical, bisecting both the animal and vegetal poles. Depending upon the amount of yolk in the egg, the planes of cleavage may pass all the way through the zygote (holoblastic cleavage, typical of cells with small to medium amounts of INTERCONNECTEDNESS REDUCE – REUSE – RECYCLE Frog & Sea Urchin Development BIOL212 4 Spring 2012 yolk; sea urchin and frog) or through only a part of the zygote (meroblastic cleavage, typical of cells with large amounts of yolk; chicken). A second cleavage division typically occurs at a right angle to the first, producing four cells. The third cleavage division cuts horizontally to form eight cells, four on the top and four on the bottom. The cells produced during these cleavage divisions are known as blastomeres. If the blastomeres in the top “tier” lie directly above those in the bottom tier, the pattern of cleavage is said to be radial, a pattern characteristic of echinoderms and chordates (deuterostomes). In the sea urchin, the fourth cleavage results in the formation of 16 blastomeres of three different sizes. Eight medium-size mesomeres are the product of the division of the four blastomeres in the animal hemisphere. The lower four blastomeres (vegetal hemisphere) produce four large macromeres and four small micromeres. As a consequence of this cleavage pattern, cytoplasmic determinants are distributed in an unequal manner, laying the groundwork for future development. Prepared Slides--Embryological Stages (and structures) Draw and label each of the slides indicated below, and answer the following questions. Early Cleavage (morula) f) How many cells make up the morula of the embryo at this stage? g) How does the size of this morula compare with the size of the fertilized egg cell? h) What is the effect of cleavage on cell size? On embryo size? Late Cleavage (morula) i) How many cells make up the morula of the embryo at this stage? j) How does the size of this morula compare with the size of the morula at early cleavage? k) Is the mass of a four-cell or an eight-cell embryo any larger than the mass of the zygote? Part C: Formation of the Blastula Repeated cleavages will result in formation of a hollow ball of cells called a blastula. The cavity inside the blastula is called the blastocoel. Even as early as this blastula stage, groups or layers of cells are already destined to become particular organs or organ systems; these layers of cells are known as presumptive germ layers. The major germ layers and their derivatives are listed in Table 1. Table 1: Germ Layers and Their Derivatives Germ Layer Ectoderm Mesoderm Endoderm Derivative Brain, spinal cord, and neural crest cells; Skin Skeleton, circulatory system, excretory system, and parts of organs belonging to other systems; Notochord and spinal disks in some organisms; ganglia Gut and associated out-pocketings In the sea urchin, blastomeres that will give rise to cells of germ layers are already laid out at the 16-cell stage. A fate map can be assigned. Mesomeres will give rise to ectodermal structures including the cilia that develop on the blastula’s surface. Macromeres will give rise to endodermal structures. Micromeres will be responsible for formation of the body cavity, many internal organs, and the skeletal elements (spicules) of the embryo. Lab Prepared Slides--Embryological Stages (and structures) Draw and label each of the slides indicated below, and answer the following questions. Blastulae (Label blastocoel, blastomeres) l) How does the size of the blastula compare with the size of the morula? m) How has the position of the cells change? INTERCONNECTEDNESS REDUCE – REUSE – RECYCLE Frog & Sea Urchin Development BIOL212 5 Spring 2012 n) Why does the center appear to be lighter? Locate a later blastula stage. The cells of the blastula are now so small that it will be difficult to distinguish individual cells. Again, the outer rim appears to be dark. At this time, the cells of the blastula are covered by cilia that allow the blastula to spin and move. Draw your observations. o) Why? Part D: Gastrulation As a blastula, most of the cells of the embryo (including those destined to become cells of internal organs and tissues) are on the outside of the hollow ball, and it is obvious that some of the sheets of cells must move, or migrate, to the inside of the blastula. This process of cell movement is called gastrulation. In some organisms, such as the starfish and sea urchin, gastrulation is accomplished invagination, or simply by buckling or pushing inward, forming a depression or blastopore. Endoderm and mesoderm reach the inside of the embryo in this manner. In other organisms, such as the frog or the chick, cells migrate to the interior by way of the blastopore, the portion of the embryo that will eventually contribute to the development of the anus. Mesoderm, endoderm, and chordamesoderm (notochord material) migrate to the inside. With the change in position of the germ layers, the blastocoel of echinoderms (starfish and sea urchin) and chordates (frog) is eventually obliterated and a new cavity, the archenteron, is formed within the gastrula. The archenteron is the primitive gut of the embryo. Note that during sea urchin development, secondary mesenchyme cells help to “pull” the endoderm of the newly forming archenteron (gut) toward the opposite side of the embryo, where it will fuse with the outer layer, or ectoderm, to form the mouth. (Hence the term “deuterostome” or second mouth, the anus forming at the blastopore or first opening.) Prepared Slides--Embryological Stages (and structures) Draw and label each of the slides indicated below, and answer the following questions. Gastrulae (Label gastrula, archenteron, endoderm, ectoderm, blastocoel) p) What is the fate of the pore that has formed by the invagination of gastrulation? q) What is the fate of the archenteron? r) What is the fate of the bastocoel? Pluteus Larvae (archenteron, mouth, anus) s) What is the symmetry of this larvae? t) How does this compare to the symmetry of the adult? Part E: Neurulation in Vertebrates During this stage of development, a strip of neural ectoderm on the outside of the dorsal surface of the embryo turns upward to form a neural tube. The underlying mesoderm and notochord tissue induce formation of the neural tube. The folds of tissue forming the tube are the neural folds and the groove between them is the neural groove. Eventually the anterior end of the neural tube will expand to form the brain; the spinal cord will develop posterior to the brain. Aggregations of mesoderm (mesodermal somites) behind the brain and alongside the spinal cord will form vertebrae (back bones) that protect and enclose the spinal cord. Mesodermal somites also give rise to dorsal skeletal muscles and to the dermis of the skin. The presence of somites is an indication of the segmented nature of vertebrate embryos. Biologists have recently shown that a series of genes control the development of segmentation in all segmented embryos, whether in fruit flies, mice, or in humans. These genes, called homeotic genes, often work in a cascade with other genes that control the basic head-to-tail and anteriorto-posterior architecture of the embryo. INTERCONNECTEDNESS REDUCE – REUSE – RECYCLE Frog & Sea Urchin Development BIOL212 6 Spring 2012 Within each homeotic gene, a special sequence or homeobox of 180 base pairs codes for a protein that is 60 amino acids long. This is a DNA regulatory protein that can “turn on” other genes involved in segmentation and segment identity (whether wings, antennae, legs, and so forth, are attached). The same homeobox is found in all homeotic genes of segmented organisms—its base sequence has been conserved throughout evolution. The gut will tubulate during this later stage of development, and the ventral unsegmented mesoderm will split to form a mesodermally lined coelom. Procedure 1. Examine and draw a cross section of a frog late neurula. Identify and label the neural tube, notochord, gut cavity, epidermis, and mesoderm lining the coelom. u) What happens to the shape of the embryo after the sides of the neural tube close? NO PRELAB (Besides the text and lecture note reading and define morula.) Post lab: Use your observations from today to answer the following questions. These questions should be completed on a separate piece of paper (not in your lab notebook) and turned into your instructor at the start of the next lab period unless another date is indicated. 1.) Explain why you would not expect a sea urchin's egg to have a large yolk. 2.) Explain why the zygote and early blastosphere (morula) are about the same size. 3.) Does the sea urchin unfertilized egg have an animal pole and vegetal pole? 4.) How can you tell the animal from the vegetal pole? 5.) Fill in the following table to summarize the major events of early development. Formation of the Blastula Early Gastrula Late Gastrula Neurula Processes occurring Type of Structure(s) formed Characteristics of structure Significance of stage 6.) Compare and contrast major developmental events in the sea urchin and frog by completing the following table. INTERCONNECTEDNESS REDUCE – REUSE – RECYCLE Frog & Sea Urchin Development BIOL212 7 Spring 2012 Sea Urchin Frog Type of egg Type/pattern of cleavage Distinguishing characteristics of blastula How gastrulation occurs Events of neurulation Distinguishing characteristics of later development 7.) What are cytoplasmic determinants? How do they affect early development, including cleavage of the zygote? 8.) How can maternal DNA affect the development of an embryo if a particular maternal allele is not included in the egg produced as a result of meiosis? INTERCONNECTEDNESS REDUCE – REUSE – RECYCLE