Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
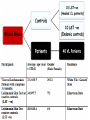
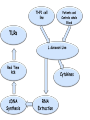
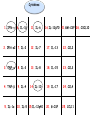
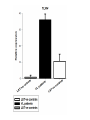
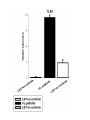
L.Donovani influenced cytokines and TLRs expression among Sudanese visceral leishmaniasis patients By: Dr.DinaTag Elsir Symposium on: Advances in Parasitology “Education and Research in Parasitology in the service of Mankind” Background Leishmaniasis remains a serious health problem. The outcome of Leishmania infection depends on the early innate response that directs the subsequent adaptive immune response to the infecting Leishmania parasite. The early innate immune response by the host, including macrophage - parasite interaction, are crucial in determining the persistence or clearance of the parasites (Bosque, et al 2000). Susceptibility to visceral leishmaniasis correlates with the presence of a Th2 response (Nyl´en S. and Sacks D. 2007). Toll-like receptors (TLRs) are hallmarks of cellular receptors that recognize pathogen-associated molecules and participate in innate responses to infections (Kaye and Aebischer 2011). We use in this study a whole blood model of stimulation to mimic the real infection scene and also a THP-1 cells model to detect the influence of the macrophage cell alone without the presence of the rest of other immune cells. We compare some of the outcome of these experiments in each model to detect if there is a similarity in stimulation and infection of these cells of both models which can give us a guide for forthcoming experiments and can help in understanding of the interplay between Leishmania and its hosts that leads to resistance or susceptibility. We measure the expressions of TLR 2, TLR4, and TLR9 and we measure the production of some cytokines in whole blood samples of visceral leishmaniasis patients and in the human macrophage cell line THP1 following stimulation promastigotes. with live L. donovani • Early events during Leishmania infections are a crucial phase that determines the persistence of the parasite or successful cure by the host immune response. The fact that the host cells of Leishmania parasites are the macrophage lineages cells suggest an important role of these cells in the pathogenesis of leishmaniasis. • Macrophages are known to recognize microbes via their toll like receptors which are currently the focus of the analysis of the innate immune responses. Materials and Methods The study was approved by the ethics committee of the Institute of Endemic Diseases, University of Khartoum. A consent forms were obtained from all participants before their enrolment. THP1 cell line Patients and Controls whole Blood TLRs L.donovani Live Real Time PCR cDNA Synthesis Cytokines RNA Extraction β Actin, TLR2, TLR4 and TLR9 oligonucleotides primers: Actin β TLR2 TLR4 TLR9 FW 5’-CTG TGG CAT CCA CGA AAC TA-3’ RV 5’-AGT ACT TGC GCT CAG GAG GA-3’ FW 5’-CGA TAT GCT AAA CAC AAT GAC -3’ RV 5’- CAA ATG ACG GTA CAT CCA CGT -3’ FW 5’CAGACATCAAGGCGCAT-3’ RV 5’TTCTTCACCTGCTCCACG-3’ FW RV 5’- GGG TTG GAA GAT GCT AGA AGA -3’ 5’- CGA GCA GGG GAG GGT CAG ACC -3’ Cytokines: 1. IFN-γ 6. IL-1β 11. IL-6 16.IL-12p70 2. IFN-α2 7. IL-2 12. IL-7 17. IL-13 22. CCL2 3. TNF-α 8. IL-3 13. IL-8 18. IL-15 23. CCL3 4. TNF-β 9. IL-4 14. IL-10 19. IL-17 24. CCL4 20. G-CSF 25. CCL11 5. IL-1α 10. IL-5 15.IL-12p40 21.GM-CSF 26. CXCL10 Results & Disscusion Cytokines • G-CSF stimulates the survival, proliferation, differentiation, and function of neutrophil precursors and mature neutrophils. This cytokine action could highlight the role of neutrophil in first hours post infection possibly the protection against Leishmania is granulocyte mediated with other several mediators. G-CSF can be used as an antileishmanial treatment since its correlation with the protection mode found in LST -ve and LST+ve groups. These intracellular parasites can use granulocytes as “Trojan horses” to invade their definitive host cells, the macrophages (van Zandbergen et al 2004). • IL-1 production was altered in VL patients. It has been shown that IL-1α administration in the susceptible BALB/c mice resulted in increased Th1 and strikingly decreased Th2 cytokine production. (Von Stebut . et al, 2003), IL-1 has been shown to affect the pathogenicity of leishmaniasis by generating an inflammatory response in afflicted tissues and by modulating adaptive T cellmediated immune responses, which act to limit parasite dissemination (Von Stebut . et al, 2003, Kostka. et al 2006). • Increased GM-CSF production following infection allows enhanced support of myelopoiesis. It was reported previously the addition of GM-CSF to human monocytes invitro increases their leishmanicidal effects (Dumas, et al 2003). Also when mice infected with L. donovani were treated with murine GM-CSF, they showed increased leishmanicidal activity, as well as leukocytosis and myelomonocytic accumulation in infected viscera (Singal, and Singh. 2005). • GM-CSF can activate macrophages to produce high levels of proinflammatory cytokines such as interleukin-1β (IL-1β), IL-6, and IL-18 and various chemokines including CCL5, CCL4, CCL3, CCL2, which enhance parasite killing. In this study, they were found to be very high and non discriminating between the study groups which in accordance with production of GM-CSF in all the three clinical groups. • The development of cell-mediated immune responses capable of controlling Leishmania infection and resolving disease is critically dependent on interferon gamma (IFNG), secreted primarily by activated T cells and NK cells in response to IL-12 (Ghosh et al 2008). Since the protective role is provided to LST ve group as well as LST+ve controls, the non discriminating production of IL-2 and IFN-γ between the VL patients and LST ve group suggesting factors other than TH1 may be genetic factors and it has been demonstrated previously the differences in genetic susceptibility to VL in the Sudanese population (Mohamed et al 2003). • IL-10 is known to play a regulatory role in Leishmaniasis. A previous report showed that few hours following exposure to L. amazonensis, PBMCs from naive volunteers presented a predominant Th2 response. Such response is supported by a high production of IL13 and IL-10 production. However, despite of this cytokine milieu, IFNG was produced (Rogers and Titus, 2004). • Dendritic cells can induce in vivo development of Th2 cells in the absence of IL-4 is in agreement with our result suggesting that IL-4 is not required for the initiation of Th2 response and other cytokines such as IL-13 that influence the development of Th2 response (Sacks and Anderson, 2004). • Moreover our data showed an increase in coproduction of IL-10 and IFNG levels in patients and controls, which reflected the mixed T cell subsets pattern in our study groups, and this is in consistent with many previous reports (Louzir et al 1998, Bosque et al 2000, Belkaid et al 2001, Khalil et al 2005,Costa et al 2012). • A number of earlier studies have implicated IL-10 in the immune suppression associated with Leishmania infection, arguing either for Th2 response (Ghalib et al 1995) or regulatory T cells (Belkaid et al 2002) as the main source of immune-suppressive IL-10. • CXCL10 chemokine is capable of recruiting and activating NK cells, which are IFNG producers, (Vester et al 1999). In this study the production of CXCL10 is clearly associated with the pathogenicity of VL and can be a biomarker for disease progression. • The correlation of this chemokine with other tropical diseases had been reported, CXCL10 were found tightly associated with fatal cerebral malaria (Wilson. et al, 2011) and Tuberculosis (Hassan. et al, 2011). CXCL10 and other proinflammatory factors in patients with active skin and mucosal lesions suggest a possible involvement of these molecules in disease outcome. • Measurement of cytokines concentration in THP1 human macrophages produced IFNG concentration more than all other clinical groups, it has been reported previously the auto activation of macrophages (Schindler, et al 2001). • A few studies reported production of IFNG by human and murine macrophage after Leishmania infection (Schindler, et al 2001, Bogdan and Schleicher. 2006). • TNF has no significant difference macrophages and the clinical groups between while IL-10, IL-1β and IL-6 were less than other groups. These findings suggest that the presence of other cells in the blood modulates the chemokine and cytokine responses of macrophages to Leishmania infections. Real Time PCR • A significant increase in expression of TLR4, a receptor known to bind to the proteoglycolipid complex ligand on Leishmania parasite that, favoring a Th1 response and increasing the production of inducible nitric oxide synthase (Kropf et al 2004,Chandra et al 2008). • Similarly, the same VL samples showed a significant increase in the expression of TLR9, a known Th1 inducer leading to protective immunity. TLR9 activation in NK cells is known to induce IFNG production and enhance the clearance of the parasite (Abou Fakher et al 2009) • TLR2 is known to bind to lipophosphoglycan (LPG) on Leishmania parasites leading to production of IFNG and TNF proinflammatory cytokines production Interestingly, LST-ve controls (55,56). showed similar TLR expression profiles as the VL patients. The fact that those individuals tested negative with Leishmanin skin test (LST-ve) suggests their susceptibility to clinical VL. • Despite the high production of IL-10 and IFNG in LST+ve controls expression of these TLRs were negatively correlated with the production of these cytokines which reported also by Srivastava et al 2013. • The significant expression of TLR4 and TLR9 in whole blood samples of VL patients compared with stimulated THP1 cell line suggests the participation of other cells eg: NK, DC, and neutrophils in VL patients responses. Conclusions: • Live Leishmania promastigotes increased the expression of TLR4 and TLR9 in peripheral blood samples of VL patients leading to the induction of aTh1 and Th2 or T reg mixed cytokine response. TLR9 seems to be recognising L.donovani DNA. • On the other hand, our data support the hypothesis that the TLRs activation seems to be the major mechanism associated with active disease. • In this context also the protective function of IFNG was demonstrated among the LST+ve controls. • Macrophage infection reveals a varied immune response distinct of that of whole blood infection scenario. • The increasing expression of TLR2 in THP1 cell line induced IFNG-γ production more than in whole blood samples, which suggesting the presence of other factors in whole blood samples that affect the IFNG production. • Despite infection and stimulation with one parasite strain within these study groups, different immune responses were elicited.