Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
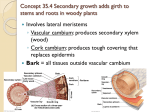
LECTURE PRESENTATIONS For CAMPBELL BIOLOGY, NINTH EDITION Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson Chapter 36 Resource Acquisition and Transport in Vascular Plants Lectures by Erin Barley Kathleen Fitzpatrick © 2011 Pearson Education, Inc. • The success of plants depends on their ability to gather and conserve resources from their environment • The transport of materials is central to the integrated functioning of the whole plant © 2011 Pearson Education, Inc. Concept 36.1: Adaptations for acquiring resources were key steps in the evolution of vascular plants • The algal ancestors of land plants absorbed water, minerals, and CO2 directly from the surrounding water • Early nonvascular land plants lived in shallow water and had aerial shoots • Natural selection favored taller plants with flat appendages, multicellular branching roots, and efficient transport © 2011 Pearson Education, Inc. • The evolution of xylem and phloem in land plants made possible the long-distance transport of water, minerals, and products of photosynthesis • Xylem transports water and minerals from roots to shoots • Phloem transports photosynthetic products from sources to sinks © 2011 Pearson Education, Inc. Figure 36.2-3 CO2 H2O O2 Light Sugar O2 H2O and minerals CO2 • Adaptations in each species represent compromises between enhancing photosynthesis and minimizing water loss © 2011 Pearson Education, Inc. Shoot Architecture and Light Capture • Stems serve as conduits for water and nutrients and as supporting structures for leaves • There is generally a positive correlation between water availability and leaf size • Phyllotaxy, the arrangement of leaves on a stem, is specific to each species • Leaf orientation affects light absorption • In low-light conditions, horizontal leaves capture more sunlight • In sunny conditions, vertical leaves are less damaged by sun and allow light to reach lower leaves © 2011 Pearson Education, Inc. Figure 36.4 Ground area covered by plant Plant A Leaf area 40% of ground area (leaf area index 0.4) Plant B Leaf area 80% of ground area (leaf area index 0.8) • Shoot height and branching pattern also affect light capture • There is a trade-off between growing tall and branching © 2011 Pearson Education, Inc. Root Architecture and Acquisition of Water and Minerals • Soil is a resource mined by the root system • Taproot systems anchor plants and are characteristic of gymnosperms and eudicots • Root growth can adjust to local conditions – For example, roots branch more in a pocket of high nitrate than low nitrate • Roots are less competitive with other roots from the same plant than with roots from different plants © 2011 Pearson Education, Inc. • Roots and the hyphae of soil fungi form mutualistic associations called mycorrhizae • Mutualisms with fungi helped plants colonize land • Mycorrhizal fungi increase the surface area for absorbing water and minerals, especially phosphate © 2011 Pearson Education, Inc. Figure 36.5 Roots Fungus Concept 36.2: Different mechanisms transport substances over short or long distances • There are two major pathways through plants – The apoplast – The symplast © 2011 Pearson Education, Inc. The Apoplast and Symplast: Transport Continuums • The apoplast consists of everything external to the plasma membrane • It includes cell walls, extracellular spaces, and the interior of vessel elements and tracheids • The symplast consists of the cytosol of the living cells in a plant, as well as the plasmodesmata © 2011 Pearson Education, Inc. • Three transport routes for water and solutes are – The apoplastic route, through cell walls and extracellular spaces – The symplastic route, through the cytosol – The transmembrane route, across cell walls © 2011 Pearson Education, Inc. Figure 36.6 Cell wall Apoplastic route Cytosol Symplastic route Transmembrane route Key Plasmodesma Plasma membrane Apoplast Symplast Short-Distance Transport of Solutes Across Plasma Membranes • Plasma membrane permeability controls shortdistance movement of substances • Both active and passive transport occur in plants • In plants, membrane potential is established through pumping H by proton pumps • In animals, membrane potential is established through pumping Na by sodium-potassium pumps © 2011 Pearson Education, Inc. Figure 36.7 CYTOPLASM ATP EXTRACELLULAR FLUID + H+ Hydrogen ion + + H+ H+ H+ H+ H+ Proton pump H+ H+ + H+ + + H + + H+ H+ H+ H+ H+ + H+ + Sucrose + (neutral solute) K+ K+ + + H+ H+ + + + + Nitrate H+ H+ Potassium ion K+ K+ K+ Ion channel (c) H+ and cotransport of ions + H+ + + H+ H+ H+ H+NO3 cotransporter H+ (b) H+ and cotransport of neutral solutes H+ H+ H+/sucrose cotransporter K+ H+ + H+ H+ (a) H+ and membrane potential K+ + + (d) Ion channels Figure 36.7a CYTOPLASM ATP EXTRACELLULAR FLUID + + + H+ H+ Hydrogen ion H+ H+ H+ H+ Proton pump + + (a) H+ and membrane potential H+ H+ • Plant cells use the energy of H gradients to cotransport other solutes by active transport © 2011 Pearson Education, Inc. Figure 36.7b H+ H+ + + + H+ H+ H+ H+ H+ H+ H+/sucrose cotransporter H+ H+ + H+ + Sucrose + (neutral solute) (b) H+ and cotransport of neutral solutes Figure 36.7c H+ H+ + + + H+ H+ H+ H+ H+ + + H+ H+NO3 cotransporter + (c) H+ and cotransport of ions H+ Nitrate H+ H+ • Plant cell membranes have ion channels that allow only certain ions to pass © 2011 Pearson Education, Inc. Figure 36.7d K+ K+ K+ + + + Potassium ion K+ K+ K+ Ion channel (d) Ion channels K+ + + Short-Distance Transport of Water Across Plasma Membranes • To survive, plants must balance water uptake and loss • Osmosis determines the net uptake or water loss by a cell and is affected by solute concentration and pressure © 2011 Pearson Education, Inc. • Water potential is a measurement that combines the effects of solute concentration and pressure • Water potential determines the direction of movement of water • Water flows from regions of higher water potential to regions of lower water potential • Potential refers to water’s capacity to perform work © 2011 Pearson Education, Inc. • Water potential is abbreviated as Ψ and measured in a unit of pressure called the megapascal (MPa) • Ψ = 0 MPa for pure water at sea level and at room temperature © 2011 Pearson Education, Inc. How Solutes and Pressure Affect Water Potential • Both pressure and solute concentration affect water potential • This is expressed by the water potential equation: Ψ ΨS ΨP • The solute potential (ΨS) of a solution is directly proportional to its molarity • Solute potential is also called osmotic potential © 2011 Pearson Education, Inc. • Pressure potential (ΨP) is the physical pressure on a solution • Turgor pressure is the pressure exerted by the plasma membrane against the cell wall, and the cell wall against the protoplast • The protoplast is the living part of the cell, which also includes the plasma membrane © 2011 Pearson Education, Inc. • Consider a U-shaped tube where the two arms are separated by a membrane permeable only to water • Water moves in the direction from higher water potential to lower water potential © 2011 Pearson Education, Inc. Figure 36.8 Solutes have a negative effect on by binding water molecules. Positive pressure has a positive effect on by pushing water. Solutes and positive pressure have opposing effects on water movement. Negative pressure (tension) has a negative effect on by pulling water. Pure water at equilibrium Pure water at equilibrium Pure water at equilibrium Pure water at equilibrium H2O H2O H2O H2O Adding solutes to the right arm makes lower there, resulting in net movement of water to the right arm: Applying positive pressure to the right arm makes higher there, resulting in net movement of water to the left arm: Positive pressure In this example, the effect of adding solutes is offset by positive pressure, resulting in no net movement of water: Positive pressure Applying negative pressure to the right arm makes lower there, resulting in net movement of water to the right arm: Negative pressure Pure water Membrane H2O Solutes Solutes H2O H2O H2O Water Movement Across Plant Cell Membranes • Water potential affects uptake and loss of water by plant cells • If a flaccid cell is placed in an environment with a higher solute concentration, the cell will lose water and undergo plasmolysis • Plasmolysis occurs when the protoplast shrinks and pulls away from the cell wall © 2011 Pearson Education, Inc. Figure 36.9 Initial flaccid cell: 0.4 M sucrose solution: Plasmolyzed cell at osmotic equilibrium with its surroundings P 0 S 0.9 0.9 MPa P 0 S 0.9 0.9 MPa (a) Initial conditions: cellular environmental P 0 S 0.7 0.7 MPa Pure water: P 0 S 0 0 MPa Turgid cell at osmotic equilibrium with its surroundings P 0.7 S 0.7 0 MPa (b) Initial conditions: cellular environmental • If a flaccid cell is placed in a solution with a lower solute concentration, the cell will gain water and become turgid • Turgor loss in plants causes wilting, which can be reversed when the plant is watered © 2011 Pearson Education, Inc. Aquaporins: Facilitating Diffusion of Water • Aquaporins are transport proteins in the cell membrane that allow the passage of water • These affect the rate of water movement across the membrane © 2011 Pearson Education, Inc. Long-Distance Transport: The Role of Bulk Flow • Efficient long distance transport of fluid requires bulk flow, the movement of a fluid driven by pressure • Water and solutes move together through tracheids and vessel elements of xylem, and sieve-tube elements of phloem • Efficient movement is possible because mature tracheids and vessel elements have no cytoplasm, and sieve-tube elements have few organelles in their cytoplasm © 2011 Pearson Education, Inc. Concept 36.3: Transpiration drives the transport of water and minerals from roots to shoots via the xylem • Plants can move a large volume of water from their roots to shoots © 2011 Pearson Education, Inc. Absorption of Water and Minerals by Root Cells • Most water and mineral absorption occurs near root tips, where root hairs are located and the epidermis is permeable to water • Root hairs account for much of the surface area of roots • After soil solution enters the roots, the extensive surface area of cortical cell membranes enhances uptake of water and selected minerals © 2011 Pearson Education, Inc. Animation: Transport in Roots Right-click slide / select “Play” © 2011 Pearson Education, Inc. • The concentration of essential minerals is greater in the roots than soil because of active transport © 2011 Pearson Education, Inc. Transport of Water and Minerals into the Xylem • The endodermis is the innermost layer of cells in the root cortex • It surrounds the vascular cylinder and is the last checkpoint for selective passage of minerals from the cortex into the vascular tissue © 2011 Pearson Education, Inc. • Water can cross the cortex via the symplast or apoplast • The waxy Casparian strip of the endodermal wall blocks apoplastic transfer of minerals from the cortex to the vascular cylinder • Water and minerals in the apoplast must cross the plasma membrane of an endodermal cell to enter the vascular cylinder © 2011 Pearson Education, Inc. Figure 36.10 Casparian strip Pathway along Endodermal cell apoplast Pathway through symplast Plasma membrane Casparian strip Apoplastic route Symplastic route Vessels (xylem) Root hair Epidermis Endodermis Cortex Vascular cylinder (stele) • The endodermis regulates and transports needed minerals from the soil into the xylem • Water and minerals move from the protoplasts of endodermal cells into their cell walls • Diffusion and active transport are involved in this movement from symplast to apoplast • Water and minerals now enter the tracheids and vessel elements © 2011 Pearson Education, Inc. Bulk Flow Transport via the Xylem • Xylem sap, water and dissolved minerals, is transported from roots to leaves by bulk flow • The transport of xylem sap involves transpiration, the evaporation of water from a plant’s surface • Transpired water is replaced as water travels up from the roots • Is sap pushed up from the roots, or pulled up by the leaves? © 2011 Pearson Education, Inc. Pushing Xylem Sap: Root Pressure • At night root cells continue pumping mineral ions into the xylem of the vascular cylinder, lowering the water potential • Water flows in from the root cortex, generating root pressure • Root pressure sometimes results in guttation, the exudation of water droplets on tips or edges of leaves © 2011 Pearson Education, Inc. Figure 36.11 • Positive root pressure is relatively weak and is a minor mechanism of xylem bulk flow © 2011 Pearson Education, Inc. Pulling Xylem Sap: The Cohesion-Tension Hypothesis • According to the cohesion-tension hypothesis, transpiration and water cohesion pull water from shoots to roots • Xylem sap is normally under negative pressure, or tension © 2011 Pearson Education, Inc. Transpirational Pull • Water vapor in the airspaces of a leaf diffuses down its water potential gradient and exits the leaf via stomata • As water evaporates, the air-water interface retreats further into the mesophyll cell walls • The surface tension of water creates a negative pressure potential © 2011 Pearson Education, Inc. • This negative pressure pulls water in the xylem into the leaf • The transpirational pull on xylem sap is transmitted from leaves to roots © 2011 Pearson Education, Inc. Figure 36.12 Cuticle Xylem Upper epidermis Mesophyll Air space Microfibrils in cell wall of mesophyll cell Lower epidermis Cuticle Stoma Microfibril Water Air-water (cross section) film interface Figure 36.13 Xylem sap Outside air 100.0 MPa Mesophyll cells Stoma Leaf (air spaces) 7.0 MPa Trunk xylem 0.8 MPa Water potential gradient Leaf (cell walls) 1.0 MPa Water molecule Transpiration Atmosphere Xylem cells Adhesion by hydrogen bonding Cell wall Cohesion and adhesion in the xylem Cohesion by hydrogen bonding Water molecule Root hair Trunk xylem 0.6 MPa Soil 0.3 MPa Soil particle Water uptake from soil Water Figure 36.13a Water molecule Root hair Soil particle Water uptake from soil Water Figure 36.13b Xylem cells Adhesion by hydrogen bonding Cell wall Cohesion and adhesion in the xylem Cohesion by hydrogen bonding Figure 36.13c Xylem sap Mesophyll cells Stoma Water molecule Atmosphere Transpiration Adhesion and Cohesion in the Ascent of Xylem Sap • Water molecules are attracted to cellulose in xylem cell walls through adhesion • Adhesion of water molecules to xylem cell walls helps offset the force of gravity © 2011 Pearson Education, Inc. Animation: Water Transport Right-click slide / select “Play” © 2011 Pearson Education, Inc. Animation: Transpiration Right-click slide / select “Play” © 2011 Pearson Education, Inc. • Water molecules are attracted to each other through cohesion • Cohesion makes it possible to pull a column of xylem sap • Thick secondary walls prevent vessel elements and tracheids from collapsing under negative pressure • Drought stress or freezing can cause cavitation, the formation of a water vapor pocket by a break in the chain of water molecules © 2011 Pearson Education, Inc. Xylem Sap Ascent by Bulk Flow: A Review • The movement of xylem sap against gravity is maintained by the transpiration-cohesion-tension mechanism • Bulk flow is driven by a water potential difference at opposite ends of xylem tissue • Bulk flow is driven by evaporation and does not require energy from the plant; like photosynthesis it is solar powered © 2011 Pearson Education, Inc. • Bulk flow differs from diffusion – It is driven by differences in pressure potential, not solute potential – It occurs in hollow dead cells, not across the membranes of living cells – It moves the entire solution, not just water or solutes – It is much faster © 2011 Pearson Education, Inc. Concept 36.4: The rate of transpiration is regulated by stomata • Leaves generally have broad surface areas and high surface-to-volume ratios • These characteristics increase photosynthesis and increase water loss through stomata • Guard cells help balance water conservation with gas exchange for photosynthesis © 2011 Pearson Education, Inc. Figure 36.14 Stomata: Major Pathways for Water Loss • About 95% of the water a plant loses escapes through stomata • Each stoma is flanked by a pair of guard cells, which control the diameter of the stoma by changing shape • Stomatal density is under genetic and environmental control © 2011 Pearson Education, Inc. Mechanisms of Stomatal Opening and Closing • Changes in turgor pressure open and close stomata – When turgid, guard cells bow outward and the pore between them opens – When flaccid, guard cells become less bowed and the pore closes © 2011 Pearson Education, Inc. Figure 36.15 Guard cells turgid/ Stoma open Guard cells flaccid/ Stoma closed Radially oriented cellulose microfibrils Cell wall Vacuole Guard cell (a) Changes in guard cell shape and stomatal opening and closing (surface view) H2O K H2O H2O H2O H2O H2O H2O H2O H2O H2O (b) Role of potassium in stomatal opening and closing Figure 36.15a Guard cells turgid/ Stoma open Radially oriented cellulose microfibrils Guard cells flaccid/ Stoma closed Cell wall Vacuole Guard cell (a) Changes in guard cell shape and stomatal opening and closing (surface view) • This results primarily from the reversible uptake and loss of potassium ions (K) by the guard cells © 2011 Pearson Education, Inc. Figure 36.15b Guard cells turgid/ Stoma open H 2O K Guard cells flaccid/ Stoma closed H 2O H 2O H 2O H 2O H 2O H 2O H2O H 2O H 2O (b) Role of potassium in stomatal opening and closing Stimuli for Stomatal Opening and Closing • Generally, stomata open during the day and close at night to minimize water loss • Stomatal opening at dawn is triggered by – Light – CO2 depletion – An internal “clock” in guard cells • All eukaryotic organisms have internal clocks; circadian rhythms are 24-hour cycles © 2011 Pearson Education, Inc. • Drought, high temperature, and wind can cause stomata to close during the daytime • The hormone abscisic acid is produced in response to water deficiency and causes the closure of stomata © 2011 Pearson Education, Inc. Effects of Transpiration on Wilting and Leaf Temperature • Plants lose a large amount of water by transpiration • If the lost water is not replaced by sufficient transport of water, the plant will lose water and wilt • Transpiration also results in evaporative cooling, which can lower the temperature of a leaf and prevent denaturation of various enzymes involved in photosynthesis and other metabolic processes © 2011 Pearson Education, Inc. Adaptations That Reduce Evaporative Water Loss • Xerophytes are plants adapted to arid climates © 2011 Pearson Education, Inc. Figure 36.16 Ocotillo (leafless) Oleander leaf cross section Cuticle Upper epidermal tissue 100 m Ocotillo after heavy rain Oleander flowers Trichomes Crypt Stoma (“hairs”) Ocotillo leaves Old man cactus Lower epidermal tissue • Some desert plants complete their life cycle during the rainy season • Others have leaf modifications that reduce the rate of transpiration • Some plants use a specialized form of photosynthesis called crassulacean acid metabolism (CAM) where stomatal gas exchange occurs at night © 2011 Pearson Education, Inc. Concept 36.5: Sugars are transported from sources to sinks via the phloem • The products of photosynthesis are transported through phloem by the process of translocation © 2011 Pearson Education, Inc. Movement from Sugar Sources to Sugar Sinks • In angiosperms, sieve-tube elements are the conduits for translocation • Phloem sap is an aqueous solution that is high in sucrose • It travels from a sugar source to a sugar sink • A sugar source is an organ that is a net producer of sugar, such as mature leaves • A sugar sink is an organ that is a net consumer or storer of sugar, such as a tuber or bulb © 2011 Pearson Education, Inc. • A storage organ can be both a sugar sink in summer and sugar source in winter • Sugar must be loaded into sieve-tube elements before being exposed to sinks • Depending on the species, sugar may move by symplastic or both symplastic and apoplastic pathways • Companion cells enhance solute movement between the apoplast and symplast © 2011 Pearson Education, Inc. Figure 36.17 Key Apoplast Symplast Mesophyll cell Companion (transfer) cell Cell walls (apoplast) Plasma membrane High H concentration H Proton pump Sieve-tube element Cotransporter S Plasmodesmata ATP BundleMesophyll cell sheath cell (a) Phloem parenchyma cell H Low H concentration (b) H Sucrose S • In many plants, phloem loading requires active transport • Proton pumping and cotransport of sucrose and H+ enable the cells to accumulate sucrose • At the sink, sugar molecules diffuse from the phloem to sink tissues and are followed by water © 2011 Pearson Education, Inc. Bulk Flow by Positive Pressure: The Mechanism of Translocation in Angiosperms • Phloem sap moves through a sieve tube by bulk flow driven by positive pressure called pressure flow © 2011 Pearson Education, Inc. Animation: Translocation of Phloem Sap in Summer Right-click slide / select “Play” © 2011 Pearson Education, Inc. Animation: Translocation of Phloem Sap in Spring Right-click slide / select “Play” © 2011 Pearson Education, Inc. Figure 36.18 Sieve Source cell tube (leaf) (phloem) Vessel (xylem) H2O 1 1 Loading of sugar Sucrose H2O Bulk flow by negative pressure Bulk flow by positive pressure 2 2 Uptake of water 3 Unloading of sugar Sink cell (storage root) 4 Water recycled 3 4 H2O Sucrose • The pressure flow hypothesis explains why phloem sap always flows from source to sink • Experiments have built a strong case for pressure flow as the mechanism of translocation in angiosperms • Self-thinning is the dropping of sugar sinks such as flowers, seeds, or fruits © 2011 Pearson Education, Inc. Figure 36.19 EXPERIMENT 25 m Sievetube element Sap droplet Aphid feeding Stylet Sap droplet Stylet in sieve-tube Separated stylet exuding sap element Concept 36.6: The symplast is highly dynamic • The symplast is a living tissue and is responsible for dynamic changes in plant transport processes © 2011 Pearson Education, Inc. Changes in Plasmodesmata • Plasmodesmata can change in permeability in response to turgor pressure, cytoplasmic calcium levels, or cytoplasmic pH • Plant viruses can cause plasmodesmata to dilate so viral RNA can pass between cells © 2011 Pearson Education, Inc. Figure 36.20 Plasmodesma Virus particles Cell wall 100 nm Phloem: An Information Superhighway • Phloem is a “superhighway” for systemic transport of macromolecules and viruses • Systemic communication helps integrate functions of the whole plant © 2011 Pearson Education, Inc. Electrical Signaling in the Phloem • The phloem allows for rapid electrical communication between widely separated organs – For example, rapid leaf movements in the sensitive plant (Mimosa pudica) © 2011 Pearson Education, Inc.