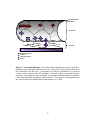
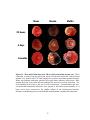
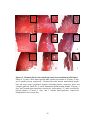
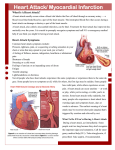
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Heart failure wikipedia , lookup
Cardiac contractility modulation wikipedia , lookup
Electrocardiography wikipedia , lookup
Mitral insufficiency wikipedia , lookup
Jatene procedure wikipedia , lookup
Hypertrophic cardiomyopathy wikipedia , lookup
Remote ischemic conditioning wikipedia , lookup
Quantium Medical Cardiac Output wikipedia , lookup
Echocardiography wikipedia , lookup
Coronary artery disease wikipedia , lookup
Ventricular fibrillation wikipedia , lookup
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
Up-regulation of HO-1 Attenuates Left Ventricular Remodeling Post Myocardial Infarction in Rats by Rebecca E. Tee A thesis submitted to the Department of Physiology in conformity with the requirements for the degree of Master of Science Queen’s University Kingston, Ontario, Canada September, 2007 Copyright © Rebecca Elizabeth Tee, 2007 Abstract Background/Objective: Reperfusion injury is a serious consequence of blood flow reestablishment after myocardial infarction (MI) mediated by reactive oxygen species and neutrophilic cellular damage. Following MI, the left ventricle (LV) undergoes remodeling characterized by progressive wall thinning and cavity dilatation. HemeOxygenase-1 (HO-1) dependent decrease in oxidative stress may attenuate injury in part by inhibiting transcription factor NFκB-mediated inflammation. Hypothesis: I hypothesized that upregulation of HO-1 by hemin administration confers acute and chronic cardioprotection against I/R injury in rats and attenuates LV remodeling post-MI. I proposed the HO-1-dependent decrease in oxidative stress attenuates post-ischemic myocardial injury in part by inhibiting NFκB-mediated inflammation. Methods: Six week old male Wistar rats were randomly assigned to sham, vehicle, or hemin-treated groups. Vehicle and hemin were administered intraperitoneally once daily for 3 consecutive days prior to left anterior descending (LAD) coronary artery occlusion. Administration resumed 48 hours post-operatively and continued once every 3 days. Infarct size was determined by H&E histological analysis and fibrosis was quantified by Masson’s Trichrome staining. Transthoracic echocardiography was used to assess LV parameters and wall motion. Results: Hemin increased HO-1 expression, decreased infarct size and fibrosis, and attenuated LV remodeling in the short-term (4 days postinfarction). The decrease in infarct size and area of fibrosis in the hemin group was accompanied by a decrease in NFκB activity. No significant difference in infarct size and area of fibrosis between hemin and vehicle-treated groups was observed at 3 months. ii LV diameter and cardiac function did not differ significantly between the two groups at 3 months despite an attenuation of anterior wall thinning in the hemin group. Conclusion: HO-1 upregulation by hemin administration conferred acute cardioprotection and attenuated LV remodeling, possibly by inhibiting NFκB-mediated inflammation. However, chronic treatment with hemin did not prevent long-term post-infarction LV remodeling. It is possible that cardioprotection afforded by HO-1 upregulation is strong enough to curtail inflammation post-reperfusion and prevent LV remodeling acutely, but is not robust enough to protect the myocardium to the same degree in the long-term. Future research should focus on optimal HO-1 upregulation to attenuate long-term LV remodeling due to reperfusion injury. iii Statement of Co-Authorship The ideas and hypotheses presented in this thesis are the result of a collaboration between Dr. Luis G. Melo, Dr. Kofoworola Ogunyankin, and Rebecca Tee. All in vivo and in vitro experiments were performed by Rebecca Tee. All echocardiography studies and analysis were performed by Rebecca Tee. All protocols used in this thesis were developed by Rebecca Tee with the help of Keith Brunt and Dr. Xiaoli Liu under the guidance of Dr. Melo and Dr. Ogunyankin. iv Acknowledgements First and foremost, I would like to thank my parents and sister who provided me with meaningful advice and endless support. My father taught me at an early age that nothing worth doing was easy and, if something was easy, everyone would be doing it. This piece of wisdom kept me motivated during many long nights in the lab alone. I would like to thank Dr. Luis Melo for his vision and guidance throughout this project. I am very grateful to Dr. Kofoworola Ogunyankin for his expertise in echocardiography and his patience when teaching me proper echocardiography techniques. I have learned more during my graduate studies than I could have imagined. Thank you for being patient with my steep learning curve. Many individual people have helped me along the way. I would like to thank Dr. Steven Pang and the Department of Anatomy and Cell Biology for their willingness to help me and provide me with equipment whenever necessary. I would also like to thank the Echocardiography Lab at Kingston General Hospital for being instrumental to my echocardiography studies. Many, many thanks also go to Dr. Xiaoli Liu for teaching me small animal surgical skills and histology techniques. I would like to make special mention of Keith Brunt who answered innumerable questions of mine and took much time to instruct me and help me troubleshoot. A great deal of what I have learned has come from you. Finally, thank you to all my friends who made my experience at Queens memorable. v Table of Contents Abstract………...……………………………………………………………………….....ii Statement of Co-Authorship……………………………………………………………...iv Acknowledgements………………………………………………………………………..v Table of Contents………………………………………………………………………....vi List of Tables …………………………………………………………………………….ix List of Figures.………………………………………………………………………….....x List of Abbreviations…………………………………………………………………….xii Chapter 1: Introduction…………………………………………………………………………..........1 1.1 Preamble………………………………………………………………………1 1.2 Functional Consequences of Metabolism……………………………………..2 1.2.1 Metabolism and Mechanical Function…………………………….2 1.2.2 Organelle Remodeling and Ion Handling…………………………3 1.2.3 Ischemia-induced Cytokine Production…………………………...4 1.3 Myocardial Reperfusion.………………………………………………………7 1.3.1 Reperfusion Injury………………………………………………...7 1.3.2 Metabolism and Mechanical Function Following Reperfusion…...7 1.3.3 Ion Handling and Ca2+ Overload During Reperfusion……………9 1.3.4 Contracture During Reperfusion………………………………….9 1.4 Reperfusion-induced myocardial inflammation……………………………..10 1.4.1 Neutrophil Infiltration and Injury………………………………..11 1.4.2 Mechanisms of Neutrophil-induced Injury………………………13 1.4.3 Reactive Oxygen Species………………………………………...13 1.4.4 Consequences of ROS Activity………………………………….17 1.4.5 Other Mechanisms of Neutrophil-mediated Injury………………18 1.4.5.1 Neutrophil-derived Proteolytic Enzymes………………...18 1.4.5.2 Arachidonic Acid Metabolites…………………………...18 1.5 Inflammatory Cytokines and Chemokines…………………………………...19 1.5.1 Inflammatory Cytokines…………………………………………19 1.5.2 Chemokines………………………………………………………20 1.5.3 Regulation of Cytokine Gene Expression During I/R Injury…….21 1.6 Wound Healing………………………………………………………………23 1.6.1 Scar Formation…………………………………………………...23 1.7 Ventricular Remodeling……………………………………………………...25 1.7.1 Infarct Expansion………………………………………………...25 1.7.2 Compensatory Hypertrophy and Ventricular Dilatation…………27 1.8 Echocardiography……………………………………………………………30 1.9 Heme-oxygenase System…………………………………………………….34 1.9.1 Cardioprotective Effects of Hemin and HO-1…………………...37 1.10 Rationale and Hypotheses………………………………………………..37 1.10.1 Rationale…………………………………………………………37 1.10.2 Hypotheses……………………………………………………….40 Chapter 2: Materials and Methods……………………………………………………….41 vi 2.1 Animals………………………………………………………………………41 2.2 Hemin Administration……………………………………………………….41 2.3 Left Anterior Descending (LAD) Coronary Artery Ligation………………..41 2.4 Transthoracic Echocardiography…………………………………………….42 2.5 Tissue Harvest……………………………………………………………….44 2.6 Isolation of Cytoplasmic and Nuclear Proteins……………………………...45 2.7 Western Blot Analysis……………………………………………………….45 2.8 NFκB Binding Activity Assay……………………………………………….46 2.9 Preparation of Histological Specimens………………………………………47 2.10 Morphometric Analysis…………………………………………………….48 2.11 Statistical Analysis………………………………………………………….48 Chapter 3: Results………………………………………………………………………..50 3.1 The Effect of Hemin Treatment in the Setting of Myocardial I/R Injury……50 3.1.1 Effect of Hemin Administration on HO-1 Expression…………….50 3.1.2 Effect of Ischemia on HO-1 Expression in Hemin and Vehicletreated Animals ………………………………………………………….50 3.1.3 Effect of Hemin Treatment on Ischemic and Reperfused Myocardium……………………………………………………………...53 3.2 Quantification of Myocardial Infarct Size…………………………………...53 3.2.1 Myocardial Area at Risk for Infarction…………………………….53 3.2.2 Myocardial Infarct Size…………………………………………….56 3.2.3 Quantification of Myocardial Fibrosis……………………………..61 3.3 Quantification of LV Structure and Function by Echocardiography………...65 3.3.1 LV Wall and Chamber Dimensions………………………………..65 3.3.2 Echocardiography Assessment of Cardiac Function………………72 3.3.2a Mid Level Ejection Fraction, Fractional Shortening and LV Mass……………………………………………………………...72 3.3.2b Apex Level Ejection Fraction, Fractional Shortening and LV Mass………………………………………………………….75 3.4 Quantification of NFκB Binding Post-reperfusion…………………………..81 3.5 Effect of Hemin Treatment on Hematocrit…………………………………..81 Chapter 4: Discussion……………………………………………………………………84 4.1 Major Findings……………………………………………………………….84 4.2 Histological Findings………………………………………………………...85 4.2.1 Myocardial Infarctions……………………………………………..85 4.2.2 Myocardial Fibrosis………………………………………………..87 4.3 Echo Findings………………………………………………………………..88 4.4 HO-1 Upregulation and Preservation of Coronary Vascular Homeostasis….92 4.5 Effect of HO-1 Upregulation of Post-infarction Left Ventricular Function…92 4.6 Comparison of Short and Long Term Studies……………………………….94 4.7 Changes in Cardiac Mass…………………………………………………….95 4.8 Role of NFκB in I/R Injury…………………………………………………..96 4.9 Hematocrit and Cardiac Function Post-I/R Injury…………………………...98 4.10 Potential Therapeutic Application of HO-1 Upregulation in Myocardial Infarction and Reperfusion………………………………………………………99 4.11 Study Consideration.………………………………………………………101 vii 4.12 Study Limitations………………………………………………………….102 4.12.1 Sample Size……………………………………………………...102 4.12.2 Use of Volatile Anaesthetic……………………………………..103 4.12.3 Echocardiography Technique…………………………………...103 4.13 Conclusions and Future Directions………………………………………..104 References………………………………………………………………………………106 viii List of Tables Table 1A. Left ventricle wall and chamber dimensions in sham-operated groups….….66 Table 1B. Left ventricle wall and chamber dimensions in hemin-treated groups…...….67 Table 1C. Left ventricle wall and chamber dimensions in buffer-treated groups…...….68 Table 2. Percent change in left ventricle dimensions from baseline echo………………70 Table 3. Percent change in functional parameters from baseline echo………………….79 Table 4. Percent change in left ventricle mass…………………………………………..80 ix List of Figures Figure 1. Consequences of ischemia and reperfusion…………………………………….8 Figure 2. Neutrophil infiltration…………………………………………………………12 Figure 3. Reactive oxygen species………………………………………………………16 Figure 4. NFκB pathway………………………………………………………………...22 Figure 5. Left ventricle remodeling post-infarction……………………………………..29 Figure 6. Sections of the left ventricle…………………………………………………..32 Figure 7. M-mode echocardiograms…………………………………………………….33 Figure 8. The heme oxygenase-1 pathway. …………………………………………….36 Figure 9. Left anterior descending coronary artery occlusion…………………………..39 Figure 10. HO-1 western blot…………………………………………………………...51 Figure 11. HO-1 expression in ischemic and non-ischemic myocardium………………52 Figure 12. HO-1 expression in reperfused and non-reperfused myocardium…………...54 Figure 13. Area at risk for infarction……………………………………………………55 Figure 14. Myocardial infarction after I/R in vehicle and hemin-treated rats…………..57 Figure 15. Hematoxylin & eosin stained left ventricle myocardium post-I/R injury…...59 Figure 16. Infarct size relative to area at risk……………………………………………60 Figure 17. Myocardial fibrosis…………………………………………………………..62 Figure 18. Area of fibrosis relative to the area at risk…………………………………...64 Figure 19. Mid level functional echo parameters…………………………….................73 Figure 20. Mid LV mass to body weight………………………………………………..74 Figure 21. Apex level functional echo parameters……………………………………...76 x Figure 22. Apex LV mass to body weight………………………………………………77 Figure 23. NFκB-DNA binding activity after I/R…………………………………….....82 Figure 24. Hematocrit at cardiac harvest………………………………………………..83 xi List of Abbreviations ANOVA = analysis of variance ASE = American Society of Echocardiography BR = bilirubin BV = biliverdin BVr = biliverdin reductase CAD = coronary artery disease cGMP = cyclic guanylyl monophosphate C5a = complement factor 5a CHF = congestive heart failure CO = carbon monoxide CO = cardiac output EPO = erythropoietin H 2 O 2 = hydrogen peroxide HO = heme oxygenase HO-1 = heme oxygenase-1 HSP-32 = heat shock protein-32 ICAM = intercellular adhesion molecule IκB = inhibitory protein of NFκB IL = interleukin I/R = ischemia reperfusion IVS = interventricular septum IVSdA = interventricular septum during diastole, apex level xii IVSdB = interventricular septum during diastole, base level IVSdM = interventricular septum during diastole, mid level IVSsA = interventricular septum during systole, apex level IVSsB = interventricular septum during systole, base level IVSsM = interventricular septum during systole, mid level LAD = left anterior descending (coronary artery) LV = left ventricle LVD = left ventricle diameter LVDdA = left ventricle diameter during diastole, apex level LVDdB = left ventricle diameter during diastole, base level LVDdM = left ventricle diameter during diastole, mid level LVDsA = left ventricle diameter during systole, apex level LVDsB = left ventricle diameter during systole, base level LVDsM = left ventricle diameter during systole, mid level MCP-1 = monocyte chemoattractant protein 1 MI = myocardial infarction MMP = matrix metalloproteinase NAD = nicotinamide adenine dinucleotide NADPH = nicotinamide adenine dinucleotide phosphate, reduced form NFκB = nuclear factor kappa B NO = nitric oxide OS = oxidative stress PAF = platelet activating factor xiii PECAM = platelet/endothelial cellular adhesion molecule PSLX = parasternal long axis PSSX = parasternal short axis PW = posterior wall PWdA = posterior wall during diastole, apex level PWdB = posterior wall during diastole, base level PWdM = posterior wall during diastole, mid level PWsA = posterior wall during systole, apex level PWsB = posterior wall during systole, base level PWsM = posterior wall during systole, mid level ROS = reactive oxygen species RT = room temperature SEM = standard error of the mean SOD = superoxide dismutase TGF-β = transforming growth factor-beta TIMP = tissue inhibitor of matrix metalloproteinase TNFα = tumour necrosis factor alpha VCAM = vascular cellular adhesion molecule VEGF = vascular endothelial growth factor xiv Chapter 1: Introduction 1.1 Preamble Myocardial infarction and reperfusion injury is a major cause of death in developed countries (Liu et al., 1997). An estimated 7.9 million Americans suffered myocardial infarctions in 2004 (Heart Disease and Stroke Statistics 2007 Update, American Heart Association). Coronary artery disease (CAD) frequently progresses to an occlusion of a major coronary artery, thereby restricting blood flow to the myocardium and compromising metabolism. Early reperfusion is essential to prevent cell death after coronary artery occlusion (Di Napoli et al., 2002). Paradoxically, sudden reoxygenation of ischemic myocardium may cause further tissue injury due to excess production of reactive oxygen species (ROS). The oxygen paradox during reperfusion describes the contradictory need to deliver oxygen and the resultant molecular reduction of oxygen to form free radicals that are involved in macromolecule oxidation, membrane dysfunction, apoptosis, and damaged calcium-sequestering ability (reviewed by Hoffman et al., 2004). These cell-damaging events are amplified by the hyper-activation of neutrophils, which promote the formation of pro-inflammatory mediators and oxygen radicals (Hoffman et al., 2004) as well as endothelial dysfunction. Myocardial necrosis and apoptosis result in increased mechanical loading of the surviving myocardium, causing changes in ventricular architecture involving the infarct and peri-infarct zones as well as the remote non-infarcted tissue. The resultant increase in load triggers a cascade of biochemical intracellular signaling processes that initiates and subsequently modulates reparative changes, which include dilatation, hypertrophy and the formation of a discrete collagen 1 scar (St John Sutton and Sharpe, 2000). It is clear that substantial alterations in gene expression occur in order to confer such profound changes within the cells of the remodeling myocardium (Stanton et al., 2000). Coordinated changes in expression of more than 400 genes related to cellular and extracellular architecture are consistent with the remodeling process in the post-ischemic heart (Stanton et al., 2000). Myocardial ischemia/reperfusion (I/R) injury is a cascade of events that requires an understanding of cellular and molecular events to explain the pathogenesis and subsequent functional consequences of ventricular remodeling. 1.2 Functional Consequences of Myocardial Ischemia 1.2.1 Metabolism and Mechanical Function Myocardial infarction is most commonly caused by a ruptured atherosclerotic lesion within a major coronary artery, wherein the vessel is occluded and blood flow to the myocardium is severely restricted. Within the first 10-15 seconds after the occlusion, the tissue partial oxygen pressure decreases, mitochondrial respiration is inhibited, and the myocytes begin to use anaerobic glycolysis as the principal source of new highenergy phosphate (Reimer et al., 1983). The delivery of glucose is markedly reduced and myocardial glycogen stores must provide the substrate for anaerobic glycolysis. In prolonged ischemia, lactate accumulates in the tissue because it cannot be further metabolized in the absence of oxygen nor can it be cleared by the systemic circulation. Eventually, glycolysis is inhibited by the accumulation of its own products, leading to further reduction of glucose metabolism in the absence of adequate ATP production 2 (Vanoverschelde et al., 1994). Protons, Na+, and Ca2+ continue to accumulate, leading to an acidotic microenvironment and transmembrane ionic imbalances. Meanwhile, the reactions that continue to require high energy phosphate (ATP) during ischemia include the myosin ATPase of the myofibrils, Na+/K+ ATPase, the Ca2+ ATPases of the sarcolemma and sarcoplasmic reticulum, adenylyl cyclase, fatty acidCoA synthetase, and other ATPases of the myocytes (Jennings and Reimer, 1991). The demand for ATP rapidly exceeds the supply. Electrical conduction continues, however, resulting in unproductive and/or sustained myocyte contractions (excitation contraction uncoupling), further requiring ATP for Ca2+ and Na+/K+ ATPases. Effective contractions terminate shortly thereafter. The metabolic changes occurring with ischemia thus include cessation of aerobic metabolism, onset of anaerobic glycolysis, and accumulation of glycolytic products (Jennings and Reimer, 1991). The degree of metabolic disturbance is proportional to the severity of ischemia. The metabolic recovery of the myocardium is dependent on rapid reestablishment of blood flow. In the absence of timely reperfusion, myocytes undergo irreversible damage, leading to cell death. 1.2.2 Organelle Remodeling and Ion Handling Prolonged (>30 minutes) ischemia causes biologically irreversible injury (Jennings and Reimer, 1991). Ultrastructural changes include diffuse mitochondrial swelling, the exhaustion of glycogen stores, marked peripheral aggregation of nuclear chromatin, and the appearance of discontinuities in the plasmalemma of the sarcolemma 3 (Jennings and Reimer, 1991). One of the hallmarks of myocyte death is sarcolemmal disruption (Jennings et al., 1983). Once breaks in the sarcolemma develop, myocytes become osmotically fragile, lose low molecular weight molecules, and accumulate extracellular electrolytes such as Na+ and Ca2+ (Jennings et al., 1990). Ca2+-induced activation of proteases causes alterations in contractile proteins, decreased sensitivity to Ca2+, and sustained impairment of contractility despite the elevated cytosolic Ca2+ (Steenbergen et al., 1987; Buja, 2005). A decreased Ca2+ sensitivity via Ca2+ overload and ischemia-induced conformational and functional changes in the channels cause contractile deficiencies. Furthermore, the increased cytosolic Ca2+ load may trigger mitochondrial release of cytochrome c and subsequently activate apoptotic caspases leading to cell death (Xu et al., 2001). Mitochondrial function is impaired during ischemia. Ischemia-induced depletion of ATP and subsequent cellular Ca2+ overload can lead to the opening of large permeability transition pores (MTP) in the mitochondria (Halestrap et al., 1998). The mitochondria not only lose small ions via these nonspecific pores, and thereby the ability to separate charges and to maintain membrane polarization, but also lose essential substrates like NAD/NADH (Vetterlein et al., 2003). 1.2.3 Ischemia-induced Cytokine Production Cytokine production and inflammation play crucial roles in determining the outcome of post-myocardial infarction. While it is widely accepted that reperfusion of 4 the ischemic tissue enables the inflammatory reaction, it is important to consider that the inflammatory reactions begin during the ischemic period (Neumann et al., 1995). Cytokines and chemokines are important pleiotropic mediators of inflammation (Neumann et al., 1995; Deten et al., 2002; Sharma and Das, 1997; Frangogiannis and Entman, 2005; Nian et al., 2004). As effectors of the immune system and the inflammatory response, cytokines modulate gene expression and transcription factor activity, and promote chemotaxis of inflammatory cells (Sun et al., 2004; Ono et al., 1998; Mercurio et al., 1997; Chandrasekar et al., 2001; Nian et al., 2004). Hypoxia and membrane disruption are likely the initial triggers for the upregulation of cytokines like TNF-α, IL-1β, and IL-6 through multiple cell signaling pathways (Deten et al., 2002). Cytokines such as TNF-α and IL-6 are rapidly released in the central ischemic zone but are also found in high concentration in the border zone (Gwechenberger et al., 1999). Ischemic stress is a powerful trigger for cytokine production, but direct myocardial mechanical stretch, which is maximal in the infarct and peri-infarct zone, is also a powerful stimulus for cytokine production (Nian et al., 2004). If the infarct size is small, the cytokine upregulation may promptly return to baseline. Deten and colleagues (2002) found that IL-1β and IL-6 mRNA were strongly induced in the myocardium after permanent coronary artery occlusion in rats. This induction persisted over the first 12 hours after infarction and rapidly declined to control levels thereafter (Deten et al., 2002). In contrast, if the infarction size is large, robust cytokine expression may be sustained chronically or may exhibit a second wave of upregulation, in keeping with the process of ventricular remodeling (Nian et al., 2004). 5 Several independent groups including Frangogiannis et al. (1998) have confirmed the presence of mast cells in the infarcted myocardium of different species. Mast cells are regulated by their local micro-environment and, in the heart, are involved in complement activation and the release of proinflammatory and vasoactive mediators (Patella et al., 1995). Initial studies by Frangogiannis et al. (1998) established the presence of preformed TNF-α in cardiac mast cells in normal myocardium in both control hearts and areas of ischemic and reperfused dog heart. Complement factor C5a is known to induce degranulation in cardiac mast cells and is present exclusively in the area surrounding the myocardial injury before initiation of reperfusion (Frangogiannis et al., 1998; Patella et al., 1995; Rossen et al., 1994). TNF-α binds the cell surface receptors and triggers intracellular signaling cascades that can result in activation of transcription factors such as NF-κB or death domain effectors such as caspases (Sun et al., 2004). It is possible that the degranulation of cardiac mast cells and the release of preformed TNF-α represent the stimulus for the initiation of a cytokine cascade before the reestablishment of coronary blood flow. Cytokines have the unique ability to self-amplify their biological effects through a positive feedback loop targeting the transcription factor NF-κB (Irwin et al., 1999). A strong amplification of the cytokine response, however, generally requires the recruitment of inflammatory cells- including leukocytes, macrophages and mast cells- to the site of injury, requiring perfusion. This concept is a cornerstone of reperfusion injury and will be discussed further in the setting of reperfusion. 6 1.3 Myocardial Reperfusion 1.3.1 Reperfusion Injury Several investigators believe that reperfusion of ischemic tissue simply accelerates the injury caused during the ischemic period and that “reperfusion injury” does not truly exist (Zweier et al., 1987; Zahger et al., 1995; Vetterlein et al., 2003). However, morphologic observations show that reperfusion of severely ischemic myocardium causes structural d.erangements exceeding the amount of damage seen with ischemia alone, indicating that reperfusion is a truly pathologic phenomenon (Vetterlein et al., 2003). Moreover, it is well established that reperfusion of ischemic myocardium with oxygenated blood generates a “respiratory burst” (Figure 1), characterized by high consumption of oxygen in association with metabolic disturbances, biochemical changes, and clinical manifestations triggered largely by neutrophil-mediated inflammation and the production of oxygen free radicals (Duilio et al., 2001; Goldhaber and Weiss 1992). 1.3.2 Metabolism and Mechanical Function Following Reperfusion The functional recovery following reperfusion is mainly determined by the extent of accumulation of metabolic end products during ischemia and the substrate availability at reperfusion (Depre et al., 1999). When glycolysis is stimulated in the reperfused myocardium, the cytosolic accumulation of Ca2+ decreases (Jeremy et al., 1992). However, elevated circulating levels of free fatty acids, due to lipolysis caused by fasting and stress hormone release, inhibit glucose utilization during ischemia and reperfusion (Hendrickson et al., 1997). As a result, high glycolytic rates during reperfusion and low 7 O2 , High energy phosphates ATP H+ Lactate pH Sarcolemma disruption, mitochondrial injury, cytoskeletal damage K+ Na2+ Ischemic contracture, Cl- Ca2+ Cell death osmolar load, cell swelling Reperfusion Respiratory burst Free radical generation •Oxidative stress •Protein denaturation •Lipid peroxidation Pro-inflammatory mediator release Neutrophil Accumulation Cytokine Elaboration Protease Release •Chemotaxis •Amplification •ECM degradation •Myocyte slippage “No re-flow” Figure 1. Consequences of ischemia and reperfusion. Inadequate oxygen supply resulting in anaerobic glycolysis produces an accumulation of waste products and intracellular acidosis that weaken cytoskeletal structure and damage organelles. Altered ion distribution and dysfunctional organelles increase the osmolar load and promote cell swelling as well as encourage ischemic contracture, all of which can lead to necrotic cell death or programmed cell death by apoptosis. Reperfusion induces a respiratory burst with a multi-component deleterious cascade, at which point cells still viable after the period of ischemia are subjected to conditions which could potentially induce necrosis or apoptosis. 8 oxidation rates can result in substantial uncoupling of glycolysis from glucose oxidation (Lopaschuk and Stanley, 1995). This uncoupling contributes to the production of protons and to ischemic reperfusion injury and a decrease in cardiac efficiency (Liu et al., 1996). 1.3.3 Ion Handling and Ca2+ Overload during Reperfusion One of the primary causes of functional impairment in post-ischemic myocardium is cellular Ca2+ overload (Depre et al., 1999). Substantial evidence exists to support the idea that the Na+/Ca2+ exchanger serves as the major physiological route for Ca2+ entry during reperfusion (Bagchi et al., 1997). During ischemia, increased cellular acidosis favours the exchange of intracellular H+ for Na+. Sudden normalization of tissue pH upon reperfusion favours hypercontracture and contributes to further Ca2+ overload (Bond et al., 1991; Schafer et al., 2000). Grinwald and Brosnahan (1987) demonstrated that the Na+ pump is inhibited during hypoxia and sustained during reperfusion, and this system could account for the Na+ load and massive Ca2+ influx via Na+/Ca2+ exchange. Rapid H+ extrusion and Na+/H+ exchange activation cause a net influx of Na+ and, depending on its ability to remove the excess load of Na+, it may in turn activate the Na+/Ca2+ exchange mechanism in the “reverse mode”, enhancing the pre-existing Ca2+ overload (Piper et al., 1998). 1.3.4 Contracture during Reperfusion Contracture refers to a sustained shortening and stiffening of the myocardium that develops during ischemia as a result of insufficient ATP to dissociate myosin crossbridges from actin filaments (Kingsley et al., 1991; Piper et al., 2003). Hypercontracture 9 occurs during the first few minutes of reperfusion because of sudden ATP availability (“re-energization”) in the presence of abnormally high intracellular Ca2+; myocytes respond to re-energization by developing an extreme distortion of their architecture resulting from development of excessive contractile force (Ruiz-Meana et al., 1999; Garcia-Dorado et al., 1997; Ladilov et al., 1997; Ganote 1983). Hypercontracture is associated with sarcolemmal disruption and enzyme release which cause dramatic changes in cytosolic composition, in turn propagated to adjacent cells through gap junctions (Ruiz-Meana et al., 1999). Moreover, increased mural tension due to contracture further constricts small vessels within the area of ischemia, making any residual oxygen and glucose delivery impossible. This is sometimes called a loss of “perfusability”. 1.4 Reperfusion-induced Myocardial Inflammation Acute myocardial ischemia-reperfusion is associated with an intense inflammatory reaction that plays a part in both acute myocardial injury as well as repair of the myocardium (Nossuli et al., 2000). Reactive oxygen species and neutrophils are key players in reperfusion injury, interacting with each other at multiple sites, as well as with other elements such as the complement system, endothelial cells, lymphocytes, macrophages and myocytes (Litt et al., 1989; Liu et al., 1997; Amsterdam et al., 1995; Kaminski et al., 2002). Principal components of the myocardial inflammation response include cell-mediated inflammation, humoral inflammatory response, the role of 10 inflammatory cytokines and transcription factor NFκB, endothelial and vascular dysfunction, and the initiation of wound healing. 1.4.1 Neutrophil Infiltration and Injury One of the earliest events of reperfusion involves neutrophil trapping in the microvasculature, whereby neutrophils are entrapped in the coronary microcirculation prior to their involvement in inflammation (Frangogiannis et al., 2002). This is commonly referred to as the “no-reflow” phenomenon and can result in a delayed, progressive decrease in blood flow. Cellular adhesion molecules promote neutrophil adhesion to endothelial cells, lining the capillaries. These adhesion molecules include selectins, β2-integrins and the immunoglobulin family, including ICAM-1, VCAM-1 and platelet-endothelial cell adhesion molecule-1 (PECAM-1) (Jordan et al., 1999). For neutrophils, firm adhesion requires activation of the β2-integrins and results in binding to one of the intercellular adhesion molecules on the surfaces of endothelial cells (Frangogiannis et al., 2002). After the inflammatory stimulus is initiated, leukocytes roll along the post-capillary venules (but not arterioles or small arteries) at a slower velocity than that of flowing blood (Frangogiannis et al., 2002). This occurs as a multi-step process (Figure 2). The slower movement enables a secure contact, mediated by ICAM1, between the vessel endothelium and the leukocyte. Dreyer and colleagues (1989) reported peak chemotactic activity released into the cardiac lymph within the first hour of reperfusion. This sharp rise in chemotaxis could explain the accelerated neutrophil accumulation, especially as the reduction in neutrophil 11 Basement membrane Endothelium Blood Flow Vessel Lumen Interstitium Neutrophil rolling, firm adhesion and transmigration P-selectin glycoprotein 1 P-selectin Leukocyte β2 integrin ICAM-1 Figure 2. Neutrophil infiltration. The cell-mediated inflammatory response post-MI is dependent upon neutrophil-endothelial cell (EC) interaction to allow extravasation from the bloodstream into the tissue. Neutrophils roll along the endothelium at a reduced velocity and are tethered to the EC mediated by selectins, leading to neutrophil integrin activation. Firm adhesion of the neutrophil via neutrophil integrins binding to members of the immunoglobulin superfamily expressed on EC is necessary for transmigration into the extravascular tissue (adapted from Frangogiannis et al., 2002) 12 accumulation shares a close temporal relationship with a decrease in chemotactic activity over the same period (Dreyer et al., 1989; Dreyer et al., 1991). Resident mast cells in the reperfused area release prepackaged proinflammatory mediators and growth factors. Stem cell factor, secreted by a subset of macrophages in the reperfused area, may also be an important factor for mast cell mobilization (Nian et al., 2004). These inflammatory cells accumulate by three days post-reperfusion and are capable of further attracting mononuclear cells to the site of injury. 1.4.2 Mechanisms of Neutrophil-Induced Injury Myocardial injury is greatly enhanced if neutrophils emigrate to the extravascular tissue or adhere directly to myocytes. As with neutrophil-endothelial interactions, adherence is low without activation of either cell type; however, when both the neutrophils and myocytes are stimulated by C5a and/or cytokines, adhesion is markedly increased (Smith et al., 1991; Entman et al., 1991; Amsterdam et al., 1995). The mechanisms involved in tissue injury mediated by neutrophils include the generation of reactive oxygen species, neutrophil-derived proteases and arachidonic acid metabolites, and the elaboration of the cytokine cascade. 1.4.3 Reactive Oxygen Species Reactive oxygen species (ROS) are atoms or compounds of oxygen with hydrogen and nitrogen with unpaired electrons in their outer orbit (Frangogiannis et al., 2002). These byproducts of oxygen metabolism, which include the superoxide anion (•O2-), hydrogen peroxide (H2O2), and the hydroxyl radical (•OH), are highly reactive 13 molecules capable of damaging tissue during I/R injury (Myers et al., 1985; Chambers et al., 1985). The cellular sources of ROS within the heart include myocytes, endothelial cells and neutrophils (Myers et al., 1985; Zweier et al., 1988, Becker et al., 1999). Within myocytes, ROS are produced by mitochondrial electron transport, NADPH oxidase, and xanthine dehydrogenase/xanthine oxidase (Myers et al., 1985; Becker et al., 1999; Chambers et al., 1985; Zorov et al., 2000). Xanthine oxidase and nitric oxide synthase (NOS) are sources of ROS production in the endothelial cell (Liu et al., 1997; Chambers et al., 1985). Neutrophils are the primary source of ROS, especially superoxide anions, generated by activity of NADPH oxidase after prolonged ischemia (Duilio et al., 2001). The production of ROS by neutrophils stimulated by proinflammatory mediators exemplifies a respiratory burst, characterized by high metabolic activity and increased consumption of oxygen. Superoxide is formed by the transfer of an electron to molecular oxygen. Superoxide dismutase (SOD) catalyzes the formation of H2O2 from •O2- (Chambers et al., 1985). Contrary to •O2-, H2O2 is a moderately strong oxidant and a highly stable small molecule that crosses membranes freely as its diffusion properties are similar to water (Antunes and Cadenas, 2000; Hool, 2006). This makes H2O2 a significant pro-oxidant species and signaling molecule. H2O2 can participate in one-electron reactions with metal ions (Fenton reaction, see Figure 3a) and can react with •O2- to generate the very toxic hydroxyl radical (•OH-) through the Haber-Weiss reaction (Chambers et al., 1985; Hool, 2006; see Figure 3b). Other reactive species are derived from nitric oxide: NO 14 reacts with many molecules including oxygen, thiol groups and metals or with superoxide to produce peroxynitrite (ONOO-) (Liu et al., 1997; Lalu et al., 2002; see Figure 3c). 15 A. Fenton Reaction Mn+ + H2O2 B. Mn+1 + OH• Haber-Weiss Reaction O2•- + H2O2 C. + OH- O2 + OH• + OH- Peroxynitrite Formation NO• + O2•- ONOO- Figure 3. Reactive oxygen species. Superoxide is generated in abundance during reperfusion. SOD converts •O2- to H2O2. H2O2 can then participate in one-electron reactions with metal ions, iron in particular, to produce a hydroxyl radical through the Fenton reaction (Panel A). H2O2 can also react with •O2- to generate a through the HaberWeiss reaction (Panel B). At the same time that •O2- is generated, the myocardium produces a burst of NO by inducible NOS. NO and •O2- react rapidly to form ONOO-. 16 1.4.4 Consequences of ROS Activity Direct consequences of ROS production include DNA nicking, lipid peroxidation, degradation of interstitial matrix molecules, as well as oxidation of amino acids and sulfhydryl groups within proteins, denaturing ion channels, receptors, enzymes and transport proteins (Fantone and Ward, 1982; Xu et al., 1997; Ambrosio et al., 1991). Proteins modified by ROS are resistant to proteolytic degradation by proteosome and this leads to a cytosolic accumulation of modified proteins (Bulteau et al., 2001). Lipid peroxidation renders the sarcolemmal and intracellular membranes weak, compromising cellular integrity. ROS alter nucleic acid and gene function, especially mitochondrial DNA (mtDNA), as it is in direct proximity to the site of myocyte ROS formation and has limited repair activity (Ide et al., 2001; Zorov et al., 2000). In addition, endothelial damage by ROS and neutrophil adhesion attenuates the release of endothelium-derived factors with anti-neutrophil properties, such as basal release of NO, and attenuates endothelium-dependent vascular relaxation, which increases endothelial permeability and neutrophil transmigration (Zhao et al., 2000). Endothelial dysfunction may also lead to increased expression of adhesion molecules and production of inflammatory cytokines (Kharbanda et al., 2001). Under physiological conditions, the toxic effects of ROS can be prevented by endogenous antioxidant enzymes SOD, catalase, glutathione peroxidase and the nonenzymatic antioxidants including vitamin E, ubiquinol, ascorbate, and reduced glutathione (Haramaki et al., 1998; Tsutsui, 2006; Hool, 2006). Myocardial recovery 17 from reperfusion injury depends partly upon its endogenous anti-oxidant reserve prior to ischemia as well as the extent of the oxidative stress. 1.4.5 Other Mechanisms of Neutrophil-mediated Injury 1.4.5.1 Neutrophil-derived Proteolytic Enzymes Activated neutrophils also release other cytotoxic metabolites including serine proteases (Murohara et al., 1995). The serine proteases that have been implicated in cardiac tissue injury are human leukocyte elastase, cathepsin G, and PR3 as they are stored in an active form within leukocyte granules (Murohara et al., 1995; Pruefer et al., 2002; Weiss, 1989; Owen and Campbell, 1999). These proteases share a broad and overlapping spectrum of activity: extracellular matrix proteolysis of elastin, fibronectin, laminin and collagen; vascular endothelial and basement membrane injury; activation of TNFα and Il-1β pro-forms; degradation of plasma proteins (complement components and immunoglobulins); enhance neutrophil release of the chemotactic platelet activating factor (PAF) and leukotriene B4 (Buerke et al.,1994; Zimmerman and Granger, 1990; Camussi et al., 1988; Camussi et al., 1989; Owen and Campbell, 1999). 1.4.5.2 Arachidonic Acid Metabolites Stimulation of phospholipase A2 after neutrophil activation mobilizes membrane lipids and results in generation of 5-lipoxygenase products (i.e., leukotriene B4) and the phospholipid PAF (Hansen, 1995). The elevation of intracellular Ca2+ during ischemia may also activate phospholipase C to stimulate arachidonic acid metabolism, which generates ROS as by-products (Kuehl et al., 1980). Leukotriene B4 and PAF are potent 18 stimulants of neutrophil chemotaxis, adhesion to endothelial cells, oxidative metabolism and degranulation and may serve to amplify neutrophil-mediated tissue injury and vascular permeability (Hansen, 1995). 1.5 Inflammatory Cytokines and Chemokines 1.5.1 Inflammatory Cytokines Interleukin 1β, acting as the “first line” cytokine stimulates chemotaxis of neutrophils but may also induce apoptosis by acting in concert with free radicals (Rampart and Williams, 1988; Arnstall et al., 1999; Kaminski et al., 2002). IL-1β is a powerful stimulus for increased IL-6 production (VanDamme et al., 1987). It has been reported that IL-6 is rapidly induced after reperfusion and induces myocyte ICAM-1 expression which may cause lethal myocyte injury due to neutrophil adhesion and transmigration (Kukielka et al., 1995). Cardiac release of IL-6, TNF-α and IL-8 during the first few minutes of reperfusion has been confirmed by various investigators (Nossuli et al., 2000; Neumann et al., 1995; Kukielka et al., 1995). The release of these cytokines is likely induced and sustained by paracrine release of TNF-α from resident leukocytes and macrophages (Neumann et al., 1995; Frangogiannis et al., 1998). Macrophages resident in cardiac tissue are considered to be a major source of IL-1β and TNF-α early during reperfusion (Herskowitz et al., 1995; Chensue et al., 1988). Cytokine amplification takes place in part through direct recruitment of more inflammatory cells to the site of injury (Nian et al., 2004). In addition, free radicals activate redox-sensitive transcription factors, including nuclear factor-κB (NFκB), and trigger the expression of 19 IL-1ß and TNF-α, that are themselves potent inducers of NF-κB activation (Chandrasekar et al., 2001). While IL-1ß, TNF-α, and IL-6 are predominantly pro-inflammatory in the reperfusion model, IL-10 serves to limit the burst of inflammatory cytokines (Yang et al., 2000). Based on the ability of exogenously administered IL-10 to suppress the expression of proinflammatory cytokines and adhesion molecules and the effect of exogenous IL-10 in limiting the neutrophil infiltration, Yang and colleagues (2000) hypothesized that the primary actions of endogenous IL-10 are related to the modulation of the neutrophil activation and their tissue infiltration. Transforming growth factor-β1 (TGF-β1) has also demonstrated a cardioprotective effect in the setting of I/R by decreasing circulating TNF-α levels (Lefer et al., 1990). 1.5.2 Chemokines Chemokines are a family of low molecular weight proteins capable of stimulating neutrophil chemotaxis as well as modulating leukocyte adhesion, activation and gene expression, mitogenesis, and apoptosis (Frangogiannis and Entman, 2005). In the canine model of myocardial infarction, IL-8 upregulation markedly increased adhesion of neutrophils to isolated canine cardiac myocytes suggesting a potential role in neutrophilmediated injury (Kukielka et al., 1995; Frangogiannis and Entman, 2005). Monocyte chemoattractant protein-1 (MCP-1) is capable of modulating fibroblast phenotype and action by altering collagen expression and regulating matrix metalloproteinase synthesis as well as attracting neutrophils (Gharaee-Kermani et al., 1996). Furthermore, MCP-1 20 has been implicated in the pathogenesis of ischemic cardiomyopathy whereby prolonged chemokine-driven inflammatory response is triggered in the absence of cellular necrosis and may play a significant role in the progression of fibrosis (Frangogiannis and Entman, 2005; Dewald et al., 2005). 1.5.3 Regulation of Cytokine Gene Expression during I/R injury Free radicals activate redox-sensitive transcription factors, including nuclear factor-κB, and trigger the expression of IL-1β and TNF-α and other inflammatory mediators (Chandrasekar et al., 2001; Fan et al., 2002). Increased NFκB/DNA binding activity has been confirmed in the I/R rat model by multiple investigators (Chandrasekar et al., 2001; Li et al., 1999). The transcription factor is normally present in the cytosol in its inactive form, bound by inhibitory proteins IκB. The phosphorylation and degradation of IκB proteins are key steps in NFκB activation, translocation to the nucleus, and stimulation of gene expression (Li et al., 2001; McDonald et al., 1997; see Figure 4). The cytokines involved in ischemia and reperfusion injury, including IL-1β, TNFα, IL-6 and IL-8, feature κB or κB-like binding motifs in their 5’ regulatory region (McDonald et al., 1997). Reperfusion after myocardial ischemia in a rat model initiates a biphasic increase in both NFκB protein levels and DNA-binding activity possibly corresponding to a primary activation by ROS and a secondary activation by proinflammatory cytokines produced by the first activation (Chandrasekar and Freeman, 1997). 21 UV radiation Hypoxia/anoxia Cytokines (IL-1, TNF-α) Reactive Oxygen Species Virus Bacterial Antigens IκB kinases P IκB proteins NFκB Iκκ complex Ubiquitination, degradation Translocation to nucleus NFkB responsive gene Regulation of gene expression -cytokines -cell adhesion molecules -matrix metalloproteinases -inducible NOS Figure 4. NFκB pathway. Inhibitory proteins (IκB) retain the inactive transcription factor in the cytoplasm until a stimulus triggers the phosphorylation and degradation of IκB. NFκB then translocates to the nucleus and regulates the expression of many inflammatory-related genes. NFκB activation is bi-phasic, peaking after 15 min and 3h reperfusion, possibly corresponding to a primary activation by ROS and a secondary activation by pro-inflammatory cytokines produced by the first activation. 22 1.6 Wound Healing and Scar Formation After reperfusion and the induction of the acute inflammatory phase, removal of injured cells by leukocytes and macrophages is followed by proliferation, migration, and activation of mesenchymal cells, and deposition of extracellular matrix proteins (Frangogiannis et al., 2005). Successful cardiac recovery depends upon controlling inflammation and establishing a reparative scar in the infarct zone to maintain ventricle wall integrity and cardiac function. Infarct healing can be divided into 3 overlapping phases: the inflammatory phase, the proliferative phase, and the maturation phase (Frangogiannis et al., 2005). 1.6.1 Scar Formation The inflammatory phase is largely dominated by the infiltration of neutrophils and mononuclear cells that phagocytose dead cells and matrix debris and produce growth factors, inducing fibroblast migration, proliferation, and activation (Dobaczewski et al., 2006). In the early stages of wound healing, extravasation of plasma proteins results in the formation of a complex fibrillar provisional matrix composed primarily of fibrin and fibronectin (Dobaczewski et al., 2006). myofibroblast progenitors and During the proliferative phase, circulating normal interstitial fibroblasts transform into myofibroblasts that are hypersynthetic, less migratory and possess contractile properties (Serini and Gabbiani, 1999; Powell et al., 1999; Freed et al., 2005). Platelet-derived growth factor (PDGF), stem cell factor (SCF) and TGF-β in particular are the primary stimuli for the transformation of myofibroblasts (Desmouliere et al., 1993). 23 Peak proliferation of myofibroblasts occurs at 4 days post-MI, predominantly in the border zone (Virag and Murry, 2003). As repair proceeds, the myofibroblasts deposit a network of collagen, specifically types I and III, and the provisional matrix is resorbed (Virag and Murry, 2003; Sun et al., 2000). The peak of type III collagen production (as early as 3 days post-infarct) is followed by a lower and slower developing peak of type I collagen, generating tensile strength and stiffness to the healing wound (Cleutjens et al., 1999; Blankesteijn et al., 2001; Dobaczewski et al., 2006). During the maturation phase, myofibroblasts form highly organized arrays in the infarct area, parallel to the epi- and endocardium, and an organized assembly of fibers in the form of scar tissue becomes evident by 2-3 weeks post-MI (Blankesteijn et al., 2001). Contraction of scar tissue during wound healing is well documented and is due, in part, to the contractile features of myofibroblasts (Tomasek et al., 2002). Formation of new blood vessels is critical for supplying the healing infarcted myocardium with oxygen and nutrients necessary to sustain metabolism (Lu et al., 2004). VEGF is critical to the development of collateral vessels and its production is significantly increased during ischemia (Nordlie et al., 2006). Immediately after reperfusion, mast cells release TNF-α which upregulates interferon-inducible protein 10 (IP-10), a known angiostatic chemokine, inhibiting neovascularization (Frangogiannis et al., 2001). After 24 hours, however, TGF-β-mediated IP-10 downregulation and VEGF upregulation shift the balance toward angiogenesis (Nian et al., 2004). 24 1.7 Ventricular Remodeling Ventricular remodeling after myocardial infarction describes alterations in the ventricle structure that include gross morphologic, histological and molecular changes in both the infarcted and non-infarcted myocardium. The process of remodeling involves thinning of the myocardial wall, enlargement of the ventricular cavity and hypertrophy of myocytes in the remote region. Animal experiments have defined myocardial infarct size as a major determinant of the severity of left ventricular dilatation (Gaudron et al., 1993), and left ventricular dilatation and dysfunction are large predictors of post-MI prognosis. Post-infarction remodeling has been arbitrarily divided into an early phase (within 72 hours) and a late phase (beyond 72 hours) (St. John Sutton and Sharpe, 2000). Early remodeling includes extracellular matrix remodeling and infarct expansion leading to wall thinning. Late remodeling includes ventricular dilatation, compensatory myocyte hypertrophy, and altered myocardial geometry. 1.7.1 Infarct Expansion Infarct expansion was originally defined using post-mortem examinations of patients dying of acute phase infarction whose left ventricle exhibited regional thinning and elongation within the scar (Hutchins and Bulkley, 1978). Infarct expansion has been defined as acute dilatation and thinning of the area of infarction not explained by additional myocardial necrosis (Hutchins and Bulkley, 1978). However, infarct expansion does not occur within all infarcts; it occurs within large, transmural anterior and antero-lateral infarcts. Most importantly, it is associated with increased mortality and 25 may be important in the development of aneurysms (McKay et al., 1986). Prevention of infarct expansion attenuates a decrease in ejection fraction and preserves ventricular function (Kelley et al.,1999). Before and during the period of resorption of necrotic tissue but before there is extensive deposition of collagen and an increase in the tensile strength, the infarcted region can thin and elongate (Pfeffer and Braunwald, 1990). This involves degradation of the extracellular scaffolding, allowing side-by-side slippage of muscle bundles to occur, resulting in a reduction of the number of myocytes across the infarcted region (Weisman et al., 1988). Central to expansion is the degradation of the inter-myocyte collagen struts by serine proteases and the activation of matrix metalloproteinases (MMPs) released from neutrophils (Cleutjens et al., 1995; Deten et al., 2002). Tissue inhibitors of MMPs (TIMPs) confine collagenolytic activity to the region of injury and control matrix degradation. Synthesis of MMPs and TIMPs is regulated by locally generated growth factors including TGF-β1 (Sun et al., 2000). Infarct expansion reflects disequilibrium between MMPs and TIMPs (Cleutjens et al., 1995). Infarct expansion leads to functional alterations due to changes in wall stress. Infarctions involving the apex are particularly vulnerable to remodeling. Given the normal ellipsoid shape of the ventricle, the apex is the most highly curved as well as the thinnest region and has the lowest wall tension (Pfeffer, 1995). Any expansion or wall thinning at the apex is exposed to greater distortion forces. Wherever the wall thinning occurs, it is exposed to the same tension per unit muscle thickness as the rest of the 26 myocardium but increased mechanical stress. This phenomenon can further perpetuate wall thinning. Transthoracic short-axis two-dimensional echocardiography at the papillary muscle level is used to determine anterior and posterior segment lengths to confirm the presence of expansion (Pfeffer and Braunwald, 1990). 1.7.2 Compensatory Hypertrophy and Ventricular Dilatation In addition to changes that occur in the infarct zone, progressive changes in the non-infarcted regions of the left ventricle contribute to the overall process of chamber enlargement (Pfeffer and Braunwald, 1990). Myocyte hypertrophy of remaining viable myocardium is initiated by neurohormonal activation, myocardial stretch, the activation of the local tissue renin-angiotensin-system (RAS), and paracrine/autocrine factors (Anversa et al., 1985; St John Sutton and Sharpe, 2000). Increased myocyte shortening and increased heart rate from sympathetic stimulation result in hyperkinesis of the noninfarcted myocardium and temporary circulatory compensation (St John Sutton and Sharpe, 2000). However, the cellular hypertrophy exhibited in the remote area is often insufficient to counteract the loss of myocyte contractility within the infarct territory. The end result is slow developing ventricular dilatation. The enlargement of the LV cavity can lead to restoration of stroke volume without concomitant improvement in ejection fraction (Pfeffer, 1995). Stroke volume can thus be maintained initially by the dilated ventricle cavity (with or without compensatory hypertrophy) with less energy expenditure and less muscle exertion. The disadvantage of this initial compensation is that the architectural changes that create 27 greater loading conditions on the viable myocardium promote further enlargement as well as hypertrophy and dysfunction (Pfeffer, 1995). Considering Laplace’s Law, whereby tension is a function of pressure and radius, dilatation increases both diastolic and systolic wall stress. Although early infarct expansion is associated with immediate ventricle enlargement, cavity dilatation continues for months thereafter, depending on the severity of the infarct. Increased cavity volume, together with insufficient compensatory hypertrophy, results in loading conditions promoting further enlargement and dysfunction (Pfeffer and Braunwald, 1990; Figure 5). 28 Figure 5. Left ventricle remodeling post-infarction. Infarct expansion results from the degradation of the extracellular structure, allowing side-by-side slippage of muscle bundles to occur. Infarct expansion and wall thinning promote ventricle distortion forces initially compensated by chamber dilatation. These architectural changes create greater loading conditions on the viable myocardium thereby promoting further dilatation as well as hypertrophy and dysfunction. 29 1.8 Echocardiography Ultrasound imaging encompasses a wide range of imaging modes and techniques that utilize the interaction of sound waves with living tissues to produce an image of the tissues (Coatney, 2001). The development of high definition echocardiographic imaging systems with high frequency transducers has permitted imaging of the heart in small animal models of myocardial infarction to evaluate systolic and diastolic function, infarct size and left ventricular remodeling (Nozawa et al., 2006). The advantages of using transthoracic echocardiography compared with other image acquisition systems include its versatility, safety, and non-invasiveness. Two-dimensional mode imaging involves a gray scale cross-sectional image of the tissue while M-mode displays the image over time, indicating the tissue depth on the Y-axis and time on the X-axis. Of particular importance is the high temporal resolution of M-mode imaging. Information that can be obtained from an M-mode echocardiogram includes LV wall thickness and chamber dimensions at various time points throughout the cardiac cycle, but commonly at end systole and diastole (Coatney, 2001). End diastolic and end systolic volumes as well as LV mass can be measured from direct LV chamber dimensions. Electrocardiogram (ECG) leads are attached to the animal to monitor electrical activity. With heart rate monitored, quantification of stroke volume, cardiac output and percent ejection fraction are possible. Serial echo studies permit the investigator to assess and follow cardiac function and phenotype longitudinally as left ventricular remodeling develops over time. 30 Standardization of measurements in echocardiography had been inconsistent and less successful compared with other imaging techniques and, consequently, echocardiographic measurements were sometimes perceived as less reliable (Lang et al., 2005). The American Society of Echocardiography (ASE), working together with the European Association of Echocardiography, has critically reviewed literature and updated recommendations for quantifying cardiac chamber dimensions (Lang et al., 2005). The ASE established the “leading edge convention” wherein dimensions are measured from the leading edge of echocardiographic borders, resulting in the inclusion of endocardial echo from the interventricular septum and posterior wall and the exclusion of endocardial echo from the left ventricle internal diameter (Deague et al., 1999). Meticulous attention is required to obtain correct orientation of imaging planes with regard to internal landmarks (Schiller et al., 1989). In general, a standardized long axis view should maximize the size of the left ventricular cavity, whereas a standardized short-axis view should minimize it within the guidelines defining each particular view (Schiller et al., 1989). A precise endocardial definition is central to accurate quantification (Schiller et al., 1989). Two-dimensionally oriented M-mode images, perpendicular from the longitudinal axis slightly above the papillary muscle level (see Figure 6), are widely employed in clinical practice and epidemiological studies (Foppa et al., 2005; see Figure 7). As a research tool, noninvasive ultrasound imaging represents not only a refinement in technique, but also a significant advancement in the ability to evaluate and quantify target organ structure and function in rodent species (Coatney, 2001). 31 Base Mid Papillary Muscle Apex Figure 6. Sections of the left ventricle. The left ventricle is divided into base, mid, and apex sections to facilitate echocardiography studies. Mid sections, at the level of the papillary muscle, are frequently used in short- and long-axis echo studies because they are easier to image with clear endocardial borders. 32 A B Figure 7. M-mode echocardiograms. M-mode echo studies are often guided by 2D images in order to ensure correct placement of the echo cursor. This visual aid enables the investigator to distinguish base, mid, or apical sections of the left ventricle. By modifying the placement and angle of the echo probe on the subject’s chest, the investigator can obtain a parasternal long-axis (PSLX; Panel A) image or a parasternal short-axis (PSSX; Panel B) image. 33 1.9 Heme-Oxygenase System The heme-heme oxygenase system has recently been recognized to possess important regulatory properties tightly involved in both physiological and pathophysiological processes including cytoprotection, apoptosis, and inflammation (Wagener et al., 2003; Maines, 1997; Wagener et al., 2003). The heme molecule promotes mostly biological oxidation processes and is vital to cellular and whole body homeostasis (Ryter and Tyrrell, 2000). Heme is involved as a carrier in oxygen transport and storage (myoglobin and hemoglobin), mitochondrial respiration (cytochromes within the electron transport chain), drug metabolism (cytochrome P450 family), endogenous anti-oxidants (catalases and peroxidases), and many others (Maines, 1992; Wagener et al., 2003). Besides its function as a prosthetic moiety in heme proteins, heme itself may influence the expression of many genes including down-regulating its own production and upregulating heme metabolism (Wagener et al., 2003b). Not only does heme affect the expression of globin, cytochromes, myeloperoxidase, and heme oxygenase-1, it also modulates cellular differentiation and proliferation. However, the pleiotropic functions of hemin are not exclusively pro-survival. It may promote deleterious iron-dependent reactions leading to ROS generation, membrane lipid peroxidation, and organelle and cellular membrane disruption (Vercellotti et al., 1994; Jeney et al., 2002; Ryter and Tyrrell, 2000). An increased cellular “free heme pool” by exogenous heme administration, increased heme biosynthesis, or increased hemoprotein degradation results in oxidative stress and can potentially cause cellular injury if endogenous antioxidant activity is insufficient. In summary, heme exerts a dual role: in small amounts it 34 acts by itself or as the functional group of heme proteins and provides diverse and indispensable cellular functions, whereas in excessive amounts, free heme can cause severe tissue damage (Wagener et al., 2001; Ryter and Tyrrell, 2000). The heme molecule can be degraded by both enzymatic and chemical mechanisms, both utilizing molecular oxygen and requiring a reducing agent (Maines, 1997). Heme oxygenase is the rate-limiting enzyme in the catabolism of heme, which yields equimolar amounts of biliverdin, free iron (Fe2+), and carbon monoxide (Maulik et al., 1996; Csonka et al., 1999; Hangaishi et al., 2000; Figure 8). subsequently converted to bilirubin by biliverdin reductase. Biliverdin is Three isoforms of the enzyme have been identified to date: inducible HO-1, constitutively-expressed HO-2, and recently discovered HO-3. HO-1 has garnered a lot of attention because accumulating experimental evidence has shown that the byproducts of heme degradation exert cytoprotective properties in numerous in vitro and in vivo cellular and tissue models of injury (Figure 8). Although heme is the major inducer of HO-1, other stimuli also upregulate HO-1 activity including oxidative stress, hypoxia, hyperoxia, cytokines, heat shock, hydrogen peroxide, ultraviolet radiation, shear stress, peroxynitrite and others (Clark et al., 2000; Maines, 1997) (Figure 8). HO-1 induction involves many signal cascades including protein kinase C, cGMP-dependent protein kinase and cAMP-dependent protein kinase (Terry et al., 1999; Immenschuh et al., 1998; Maines, 1997). Moreover, the HO-1 gene 35 Heme Stimuli -Hypoxia, hyperoxia -Ischemia -Oxidative stress -Cytokines -Shear stress -NO -Prostaglandins, endotoxins Source -Endogenous biosynthesis -Exogenous administration Heme Oxygenase O2 NADPH H2 O NADP+ CO Biliverdin Biliverdin Reductase Bilirubin Fe2+ Oxidant Species Ferritin •Vasodilator •Anti-inflammatory •Anti-apoptotic •Anti-thrombotic •Anti-oxidant •Anti-inflammatory •Anti-oxidant •Iron homeostasis Figure 8. The heme oxygenase-1 pathway. HO-1 catalyzes the degradation of heme to yield equimolar CO, biliverdin and Fe2+. Biliverdin is subsequently converted to bilirubin by biliverdin reductase. The production of Fe2+ induces an upregulation of ferritin which sequesters the free iron. 36 has regulatory elements in the promoter region including DNA-binding sites for redoxsensitive transcription factors AP-2 and NFκB (Lavrovsky et al., 1994). 1.9.1 Cardioprotective Effects of Hemin and HO-1 The exogenous administration of hemin and subsequent upregulation of HO-1 confer potent resistance to stress and cellular injury by mediating oxidative stress and acute inflammation (Clark et al., 2000; Weber et al., 2001). There is considerable interest in the cardioprotective potential of HO-1 activity in the setting of myocardial infarction and ischemia/reperfusion injury. Hangaishi et al. (2000) observed the upregulation of cardiac HO-1 expression induced by hemin injection decreased infarct size 2 hours after reperfusion. Melo et al. (2002) reported a pre-emptive gene therapy strategy for myocardial protection from ischemia/reperfusion injury using adenoassociated virus to deliver HO-1 to the heart. In an allograft transplantation model, transgenic mice overexpressing HO-1 systemically sustained improved allograft outcomes with inhibition of the rejection response (Araujo et al., 2003), implicating antiinflammatory, anti-oxidant and anti-apoptotic properties. Most recently, Liu et al. (2006) showed complete recovery of function and normalization of ventricular dimensions in intramyocardial HO-1-delivered rats 1 year after MI. 1.10 Rationale and Hypotheses 1.10.1 Rationale The introduction of genetic manipulation into molecular medicine represents an advancement in the treatment of cardiovascular diseases. Controlling post-infarction 37 tissue inflammation through regulation of gene expression is an attractive means of limiting organ damage. Evaluating viability and contractile function in cultured cardiac myocytes is a good model for distinguishing between the simulated ischemia and reperfusion phases. However, in vitro studies frequently do not simulate the severity of reperfusion injury observed in vivo due to the absence of endothelial and inflammatory cells, the absence of inflammatory mediators, the lack of sufficient muscle for hypercontracture damage, and fewer sources of ROS (Vanden Hoek et al., 1996). Surgically induced myocardial infarct in rats is a widely accepted in vivo model of myocardial injury (Moises et al., 2000). The major advantage of this model is the possibility of keeping infarcted animals alive for extended periods of time, thereby allowing investigation of healing and recovery of the infarcted myocardium (Moises et al., 2000). Furthermore, left anterior descending coronary artery occlusions are especially useful to study because the left ventricle territory it supplies is large, compromising the anterior lateral wall (see Figure 9). Serial echocardiographic studies, including baseline and post-op studies for each animal, permit the investigation of left ventricular structure and function and abnormalities therein. 38 Figure 9. Left anterior descending coronary artery occlusion. The LAD coronary artery supplies most of the left ventricle territory and occlusions therein create large antero-lateral apical to mid-level infarcts making it very useful to study. 39 1.10.2 Hypotheses The purpose of this study was to investigate the effects of HO-1 upregulation in vivo in myocardial tissue recovering from I/R injury. I hypothesized that exogenous upregulation of HO-1 by hemin administration confers acute and chronic cardioprotection against I/R injury in rats and attenuates post-infarction left ventricular remodeling. I proposed the HO-1-dependent decrease in oxidative stress attenuates post-ischemic myocardial injury in part by inhibiting NFκB-mediated inflammation. Using histological methods, I quantified infarct size and left ventricle fibrosis in hemin- and buffer-treated animals subjected to ligation and release of the LAD coronary artery. Echocardiography was used to examine left ventricle wall and chamber dimensions as well as cardiac function. Transcription factor NFκB binding activity was measured. This study confirmed the hypothesis that HO-1 upregulation reduces acute injury after myocardial infarction at 24 hours and 4 days post-op. However, treatment with hemin did not reduce reperfusion injury nor prevent long term LV remodeling 3 months after infarction. 40 Chapter 2: Materials and Methods 2.1 Animals All animal handling and procedures were carried out according to the Canadian Council of Animal Care (CCAC) guidelines and approved by the University Animal Care Committee at Queen’s University. Six week old male Wistar rats (180-200 g) from Charles River, Canada were randomly assigned to sham, buffer, or hemin-treated groups. The animals were kept on a 12:12 hour light: dark cycle at an ambient temperature of 2123°C and 60% humidity. Food and water were provided ad libitum. 2.2 Hemin Administration Hemin (Sigma Aldrich, Oakville, ON) was dissolved in buffer consisting of 0.1N NaOH and 0.01M KPO4 monobasic to a concentration of 75 μM and administered at a concentration of 50 μMol/kg body weight. Buffer and hemin were administered intraperitoneally once daily for 3 consecutive days prior to left anterior descending (LAD) coronary artery occlusion. Intraperitoneal administration was resumed 48 hours post-operatively and continued once every 3 days. Sham animals were injected with vehicle solution. 2.3 Left Anterior Descending (LAD) Coronary Artery Ligation Animals were anaesthetized using 60 mg/kg sodium pentobarbital given intraperitoneally and additional doses were given as needed to maintain surgical plane. Animals were intubated and ventilated using a rodent ventilator (model 683, Harvard Apparatus; Montreal, QC) with 100% oxygen throughout the procedure. Rats were 41 ventilated at a tidal volume of 1.25 ml/100 g body weight (to a maximum tidal volume of 5 ml) and rate of 72 breaths per minute. Left horizontal thoracotomy was performed in the second intercostal space. The pectoral muscles were separated by blunt dissection and intercostal muscles were cut to expose the heart in the thoracic cavity. The second and third ribs were retracted and the pericardium was torn to expose the left ventricle wall. Polypropylene suture thread (6.0 Ethicon; Johnson and Johnson, Markham, ON) was pulled through the left ventricular myocardium, beneath the left auricle. A small (1mm diameter) piece of cotton was placed within the ligature knot to prevent crushing of the vessel. The ligature was tied tightly for 30 minutes and subsequently loosened, allowing reperfusion of the left anterior descending coronary artery. Sham animals were subjected to thoracotomy and ligature placement without vessel occlusion. The ligature was left in place, the cotton removed from the knot, and the thoracotomy was closed using 3.0 Ethicon suture thread. Phosphate buffered saline (PBS) (Sigma) was administered intraperitoneally at the onset of ischemia and during recovery to maintain hydration and restore any potential blood loss. Once extubated, buprenorphine (0.3 mg/ml) was administered subcutaneously at 0.05 mg/kg post-op and again 8 hours later for pain control. 2.4 Transthoracic echocardiography Echocardiography was performed on all treatment groups 1 day pre-operatively and 4 days post-operatively using a Phillips HDI-5000 Ultrasound System with a P 12.5MHz transducer at 2 cm depth. For some groups, an additional echo study was performed at 3 months. Measurements of LV chamber and wall dimensions were made 42 according to the leading edge guidelines set by the American Society of Echocardiography. For image acquisition, the animals were shaved and sedated with halothane (Sigma) by nose cone inhalation. Animals were placed on a ramp with a fixed heating pad and linen cloth to maintain body temperature and were positioned such that a comparable left lateral decubitus position was achieved. Three electrocardiograph lead wires were inserted sub-cutaneously on the left arm, left leg, and right arm in order to monitor heart rate. Two-dimensional and M-mode images from the parasternal long and short axes were taken from the basal (at the tip of the mitral valve leaflets), mid (at the papillary muscle), and apical (distal to the papillary muscle) levels and recorded on an optical disk for later review. Image analysis and measurements were performed blind to the treatment group. Echocardiography studies were reviewed and the best frames within an animal study were selected. Three consecutive contraction cycles were measured within a single frame at each level of the heart, in both parasternal long and short axes. Endocardial borders were traced to calculate the interventricular septum, cavity diameter, and posterior wall thickness during diastole and systole. Values were recorded in Microsoft Excel and imported to Microsoft Access for data organization and queries, and formula derivation to calculate left ventricle mass, fractional shortening, and ejection fraction. LV mass was calculated using the ASE formula (1.04 x [LVd + PWd + AWd)3- LVd3] where 1.04 is the specific gravity of myocardial tissue. LVd is LV diameter during diastole and PWd and AWd are end diastolic posterior wall and anterior wall (or interventricular septum) thicknesses respectively. Fractional shortening (FS) was calculated by (LVD-LVs)/LVd 43 x 100, where LVs is end-systolic diameter. Ejection fraction (EF) is calculated by (LV diastolic volume- LV systolic volume)/ LV diastolic volume. 2.5 Tissue harvest Hearts were harvested at 24 hours, 4 days and 3 months post-op. The animals were anaesthetized with sodium pentobarbital (60 mg/kg) by intraperitoneal injection and intubated. The sternum was cut mid-sagitally and the thoracic cage was retracted. The right carotid artery was bluntly dissected and cannulated with 1 mm plastic tubing with suture thread to secure the cannula in place. The thymus was removed to expose the aortic arch and heart. Suture thread was placed around the aortic arch between the right brachiocephalic and the left common carotid artery. A second suture was placed around the right subclavian branch. Arterial blood was collected in a 3 ml syringe and then centrifuged at 3200 rcf for 10 minutes at 4°C. Serum was collected and stored at -80°C. Capillary tubes were filled with arterial blood and spun for 6 minutes in a micro hematocrit centrifuge (International Equipment Company; Fisher Scientific, Ottawa, ON) to determine the blood hematocrit by micro-capillary reader (IEC). Next, the heart was perfused through the carotid cannula with cold PBS + KCl (0.01M) to arrest the heart in diastole. The LAD suture thread was tied tightly and Evan’s blue dye (1 mg/ml PBS; Sigma) was infused via the carotid cannula. Evan’s blue dye perfused and stained the area of the LV not at risk for infarction while the myocardium distal to the LAD ligature was not perfused, clearly marking the ventricular area at risk. Hearts were quickly excised and washed with cold PBS. 44 2.6 Isolation of Cytoplasmic and Nuclear Proteins Infarcted and non-infarcted tissues were quickly dissected from the harvested heart and immediately immersed in liquid nitrogen. Tissues were pulverized in liquid N2 using a mortar and pestle and stored at -80°C until use. Cytosolic and nuclear fractions were obtained from each sample. Samples were first homogenized with ice-cold hypotonic buffer (10mM HEPES, 10mM KCl, 0.1mM EDTA, 0.1mM EGTA, 1mM DTT, protease inhibitor, phosphatase inhibitor; 500 μl/100 mg dry weight) using a Polytron PT-2100, carried out on ice, to lyse the cell membrane. Homogenate was centrifuged at 2000 rpm for 1 minute (4°C) and the supernatant was collected and incubated on ice for 20 minutes. 10% Igepal was added (50 μl/100 mg) to the supernatant, vortexed for 30 seconds, and centrifuged at 10,000 rpm (1min, 4°C). The cytosolic fraction was collected and stored at -80°C. The pellet was rinsed once in hypotonic buffer, repelleted by centrifugation and the supernatant was discarded. The pellet was then resuspended in ice-cold hypertonic buffer (20mM HEPES, 0.4M NaCl, 1mM EDTA, 1mM EGTA, 1mM DTT, protease inhibitor, phosphatase inhibitor; 100 μl/100 mg original tissue weight), homogenized using a Dounce homogenizer and incubated for 40 minutes on ice with frequent mixing. The nuclear homogenate was then centrifuged for 15 minutes at 14,000 rpm. This supernatant was collected as nuclear extract and stored at -80°C. 2.7 Western Blot Analysis Total protein concentration was determined for each sample by using the Bradford assay with a spectrophotometer (DU® 530; Beckman Coulter, Mississauga, ON, 45 Canada) at 530 ηm. Protein samples (25 μg/μl) were mixed with 4x Laemmli buffer with β-mercaptoethanol and were electrophoresed on a 12% poly-acrylamide gel using a Bio Rad system (Bio Rad Laboratories, Mississauga, ON, Canada). Transfer onto PVDF membrane (0.45 μM; Immobilon-P, Millipore, Billierica, MA, USA) was carried out using a Bio Rad semi-dry apparatus. Membranes were blocked with 4% non-fat dry milk and probed for human HO-1 primary antibody (dilution 1:5000; SPA 895, Stressgen, Victoria, BC). After the membranes were washed 3x 15 minutes with TBST, they were incubated in 1:5000 horseradish peroxidase-conjugated anti-rabbit secondary antibody (#7074, Cell Signaling; Danvers, Massachusetts, USA). Following 3 x 15 minutes washes, membranes were exposed using enhanced chemiluminescence according to the manufacturer (Pierce; Fisher Scientific, Ottawa, ON) and an AlphaEaseTM gel documentation system (Fluorchem 8900; Alpha Innotech Corp., San Leandro, CA, USA) for visualization of the protein. Blot images were analyzed for spot densitometry using Image J software. Membranes were stripped using a 0.2M glycine, pH 2.6 stripping buffer solution for 1 hour with rocking and were re-probed for β-actin (1:5000) for normalization of in vivo western blots. 2.8 NFκB Binding Activity Assay Transcription factor activity was quantified using the NoShiftTM II NFκB Transcription Factor Assay kit (Novagen; VWR CANLAB; Mississauga, ON). Briefly, binding solution and nuclear samples were distributed to sample wells in a 96-well plate and were incubated for 20 minutes at RT. Binding reagent was added and the plate was incubated for another 20 minutes at RT. Digestion reaction buffer was added and the 46 plate was incubated at 37°C for 20 minutes. Hybridization reaction buffer was added to corresponding wells of the NoShift II Capture Microplate. Samples were then transferred to the microplate and incubated for 45 minutes at RT. Samples were washed four times with wash buffer and aspirated. Detection reagent was added to each well and the microplate was incubated for 30 minutes at RT. The detection reagent was then aspirated, and the wells were washed four times. Chemiluminescent substrate was added and the microplate was incubated for 30 minutes at 37°C. Chemiluminescence was measured with luminometer using a plate reader (Fluostar Optima, BMG Biotech) and results were expressed as relative light units (RLU) per μg of protein. 2.9 Preparation of Histological Specimens Warmed agarose gel (2%; Sigma) was poured over the excised heart to temporarily fix it in place. Left ventricles were cut into 3-4 sections, approximately 3-4 mm thick, and were photographed with a digital camera fixed to a dissecting microscope for morphometric analysis of the risk and non-risk areas. Sections were washed in cold PBS. Samples were immersed in 2,3,5-triphenyltetrazolium chloride (TTC) solution (1 mg/ml PBS; Sigma) and incubated for 12 minutes at 37°C. Samples were photographed for morphometric analysis of the area of infarction. Sections were placed in 10% formalin (Accustain, Sigma) and stored at 4°C overnight and then stored in 70% ethanol at 4°C until paraffin embedding. 47 2.10 Morphometric Analysis Ventricular sections from the apex, mid, and basal segments of the heart were embedded in paraffin wax (TissuePrep, Fisher Scientific), cut into 5 μm thick with a “820” Spencer Microtome (American Optical Corporation; Colorado, USA), and placed on 3-(aminopropyl)triethoxysilane-coated histology glass slides. Sections were stained with hematoxylin and eosin (H&E) and Masson’s Trichrome (Accustain, Sigma). To image the entire heart section, slides were scanned using a high resolution Epson scanner. Adobe Photoshop 5.5 was used to quantify the area of left ventricular infarction using multiple (3-4) coronal sections of each animal specimen. The area of infarction, ventricle cavity and the entire left ventricle were measured using the lasso tool and were expressed in relative pixels. This lasso method was also used to quantify the area at risk of infarction, delineated by the Evan’s Blue infusion, relative to the entire ventricle once photographed with the dissecting microscope. The area of infarction was then expressed relative to the area at risk in order to normalize any variation in infarct size. The same method was used to measure the area of fibrosis relative to the area at risk of infarction. 2.11 Statistical Analysis Statistical analysis was performed by unpaired t-test or 1-way ANOVA with P value set at <0.05 and post-hoc Bonferroni multiple comparison tests using GraphPad Prism 4.0 (GraphPad Software, Inc.; San Diego, California). To determine the statistical significance in area at risk, infarct size and collagen fraction between hemin- and buffertreated animals, unpaired t-test was performed between the two groups at each time point (24 hours, 4 days, 3 months). To determine the statistical significance in NFκB-DNA 48 binding activity, unpaired t-test was also performed between the two groups at each time point. Echocardiographic parameters were compared between sham, hemin- and buffertreated groups using 1-way ANOVA followed by Bonferroni multiple comparison test. Comparisons were made between the groups at each of the time points (pre-op, 4 days, 3 months). 49 Chapter 3: Results 3.1 The Effect of Hemin Treatment in the Setting of Myocardial I/R Injury 3.1.1 Effect of Hemin Administration on HO-1 Expression The effect of hemin administration on myocardial HO-1 protein expression is shown in Figure 10. HO-1 protein expression was increased 24 hours and 4 days post I/R in the hemin-treated animals compared to vehicle after daily intraperitoneal injection of hemin for 3 days prior to and 2 days after I/R. These results confirm that hemin administration upregulates myocardial HO-1 expression in the setting of I/R. By day 4, the HO-1 protein levels were approximately two-fold higher in the hemin-treated animals compared to the vehicle-treated animals. 3.1.2 Effect of Ischemia on HO-1 Expression in Hemin and Vehicle-treated Animals Since ischemia per se may induce HO-1 expression, I compared expression of HO-1 protein in the ischemic and non-ischemic region of the hemin and vehicle-treated animals. Twenty-four hours after hemin or vehicle treatments, HO-1 protein levels were comparable in the ischemic and non-ischemic myocardium of the vehicle-treated animals (Figure 11) indicating that I/R had no direct effect on endogenous HO-1 expression. In contrast, HO-1 protein expression was markedly increased in the ischemic and nonischemic myocardium in the hemin-treated animals (Figure 11). 50 A Spot Density (arbitrary units) 300 200 100 4d H 4d B H B 24 h 24 h 0 B4d H4d B C HO-1 +ve control B24h H24h β-actin reprobe Figure 10. HO-1 western blot. A) Densitometric analysis of HO-1 expression in control, buffer- and hemin-treated left ventricle myocardial tissue at 24 hours and 4 days. HO-1 expression in hemin-treated animals was greater at 24 hour and 4 days after I/R (n=1). B) Representative HO-1 western blot showing HO-1 protein expression at 24 hours and 4 days in both hemin- and buffer-treated animals. C) β-actin reprobing of the same blot reveals equal loading. 51 A Spot Density (arbitrary units) 500 400 300 200 100 B24h . I. N Is ch . N Is ch . . I. 0 H24h B HO-1 +ve control C I NI B24h I NI H24h β-actin reprobe Figure 11. HO-1 expression in ischemic and non-ischemic myocardium. A) Densitometric analysis of HO-1 protein expression in ischemic and non-ischemic tissue in both buffer- and hemin-treated animals at 24 hours. No difference was noted between ischemic and non-ischemic tissue in the buffer treated sample. In contrast, HO-1 expression was markedly increased in both the ischemic and non-ischemic myocardium of hemin-treated animals. B) Representative western blot showing HO-1 expression in the ischemic and non-ischemic regions of the myocardium in vehicle and hemin-treated animals 24 hours after I/R and C) β-actin reprobing of the same blot as in (B). 52 3.1.3 Effect of Hemin Treatment on Ischemic and Reperfused Myocardium Since reperfusion by itself generates ROS and may induce HO-1 protein expression, the isolated effects of myocardial reperfusion and ischemia on HO-1 expression were determined. The results are shown in Figure 12. Although, hemin was shown to induce HO-1 expression in ischemic and non-ischemic myocardium (Figure 11), reperfusion had an additive effect on HO-1 expression in both ischemic and nonischemic regions of the myocardium after reperfusion. This indicates that reperfusion by itself compounds the induction of HO-1 by hemin administration. 3.2 Quantification of Myocardial Infarct Size 3.2.1 Myocardial Area at Risk for Infarction A prerequisite for the assessment of myocardial injury is that the area at risk of I/R injury is comparable between the study groups. Thus, area at risk for infarction was calculated to assess infarct size accuracy and to normalize for infarct size discrepancies between animals. The area of risk for the various hemin and vehicle-treated groups was calculated by measuring the area of the LV unstained by Evan’s blue dye, expressing it as a percent of the entire LV. The percent of the LV area at risk for infarction was not significantly different between the hemin and buffer-treated animals. However, the mean area at risk for the hemin-treated animals tended to be greater than the vehicle group at every time point. Interestingly, there was a time-dependent trend towards smaller areas of risk in both groups at later time points, possibly because of thinning of the infarcted wall and concurrent hypertrophy of the remote non-infarcted myocardium. 53 A Spot Density (arbitrary units) 400 300 200 100 ch ic on -Is ch em Reperfused N Is N Is on -Is ch em ic ch 0 Non-reperfused B HO-1 +ve control I NI Reperfused heart I NI Non-reperfused C β-actin reprobe Figure 12. HO-1 expression in reperfused and non-reperfused myocardium. A) Densitometric analysis of HO-1 protein expression in hemin-treated animals. Ischemic reperfused myocardium expressed greater HO-1 than non-ischemic myocardium and ischemic non-reperfused myocardium (n=1). B) Representative western blot and C) βactin reprobe to confirm equal protein loading. 54 % Left Ventricle Area 80 60 40 20 B uf fe r 3m 3m in em H B uf fe r 4d 4d in em H r2 4h uf fe B H em in 24 h 0 Figure 13. Area at risk for infarction. Percentage of left ventricle area at risk for infarction after LAD occlusion in hemin- and buffer-treated animals at 24 hours, 4 days, and 3 months. Though no significant differences were observed among the groups, hemin-treated animals showed a general trend of greater mean areas at risk compared with buffer-treated animals at corresponding time points. (24 hours, hemin, n=7, buffer, n=10; 4 days, hemin, n=6, buffer, n=6; 3 months, hemin, n=8, buffer, n=4; P<0.05) 55 3.2.2 Myocardial Infarct Size Figure 14 depicts representative thick ventricular sections from the infarcted or sham-operated LV from each treatment group. Evan’s blue staining represents myocardial tissue not at risk for infarction, while red or pink staining signifies viable tissue and white staining represents dead myocardium. The buffer 24 hour representative image reveals a very large, transmural infarct with minimal viable myocardium within the ventricle’s infarct zone while the representative buffer 4 day image reveals a massive circumferential infarct. The buffer 3 month sample shows considerable myocardial injury and wall thinning with no visible myocardial salvage within the infarct zone. It was observed upon organ harvest that the lateral ventricle wall was fixed to the chest well with scar tissue and was difficult to dissect; it was technically challenging to maintain the shape of the ventricle once sliced and the true nature of the paper-thin infarct zone is somewhat lost. The hemin samples, on the other hand, display discrete bands of injury within the left ventricle wall and confirm the presence of viable tissue in the subendocardium and subepicardium. Differences in the shades of viable red tissue are due to lighting differences from the dissecting microscope upon taking the photographs. 56 Sham Hemin Buffer 24 hours 4 days 3 months Figure 14. Myocardial infarction after I/R in vehicle and hemin-treated rats. Thick ventricular sections from the apical-mid region of infarcted hemin and vehicle-treated animals were stained with TTC and compared to sections from sham-operated animals. White area indicates infarction, whereas red or pink tissue indicates viable tissue. Blue staining from the Evan’s blue infusion indicates the area not at risk for infarction. The area excluding Evan’s blue is the area at risk for infarction. Circumferential and nearcircumferential transmural infarctions were present in the buffer-treated animals at 24 hours and 4 days compared to the smaller infarcts in the hemin-treated animals. Extensive wall thinning was seen in the buffer-treated animal 3 months after infarction. 57 H&E histologically stained samples of LV myocardium are shown in Figure 15. Panels A, D, and G show sham-operated H&E stained myocardium at 24 hours, 4 days, and 3 months post-op, respectively, and the maintenance of myocardial integrity is clear. Panel B illustrates a hemin-treated animal 24 hours after reperfusion and the beginning of the injury development, manifested by tissue edema and disorganization, is clear. However, the injury shown in the buffer-treated 24 hour animal (Panel C) appears far greater in area and injury intensity. Panels E and F compare a hemin-treated 4 day animal with a buffer-treated 4 day animal, respectively, and the difference in the area of injury is evident. Panels H and I show hemin 3 month and buffer 3 month images side by each. Wall thinning is apparent in both images, however, myocardial salvage is noticeable in the hemin 3 month animal (Panel H) and the integrity of the remaining myocardium is comparable to the sham-operated 3 month animal (Panel G). In contrast, extreme wall thinning and transmural scar formation are evident in the buffer 3 month animal. Infarct size was normalized to the area at risk within each treatment group. Once this was taken into account, significant differences in infarct size (expressed as a fraction of the LV) were observed between the hemin- and buffer-treated groups at 24 hours (0.211 ± 0.001 vs. 0.435 ± 0.003, respectively) and 4 days (0.344 ± 0.004 vs. 0.682 ± 0.004, respectively) post-MI (Figure 16). Surprisingly, no significant difference was seen between the treatment groups at 3 months, although the mean infarct size of the buffertreated animals showed a trend towards larger infarcts (0.507 ± 0.091) compared to the hemin-treated animals (0.459 ± 0.063). 58 A B C D E F G H I Figure 15. Hematoxylin & eosin stained left ventricle myocardium post-I/R injury. Panels A, D, and G show sham-operated H&E stained myocardium at 24 hours, 4 days, and 3 months post-op, respectively. Hematoxylin stains nuclear material dark purpleblue and eosin stains cytoplasmic material (including proteins, connective tissue and collagen) pink-red. Panels B, E, and H demonstrate hemin-treated animals 24 hours, 4 days, and 3 months post-reperfusion, respectively, while panels C, F, and I reveal buffertreated animals 24 hours, 4 days, and 3 months post-reperfusion, respectively. (Magnification 10x; inserts 20x) 59 0.7 0.6 * 0.5 0.4 0.3 0.2 0.1 3m B uf fe r 3m in 4d H em B uf fe r 4d H em in B uf fe r H em in 24 h 0.0 24 h Infarct Size: Area at Risk Ratio * 0.8 Figure 16. Infarct size relative to area at risk. Infarct size was normalized to area at risk to account for differences in area at risk between animals. Infarct size was significantly greater in the buffer-treated animals compared to the hemin-treated animals at 24 hours and 4 days after I/R (P<0.05). No significant difference was seen between buffer and hemin-treated groups at 3 months, although the mean ratio of the buffertreated animals tended to be greater (0.507 ± 0.091) than that of the hemin-treated animals (0.459 ± 0.063). (24 hours, hemin, n=7, buffer, n=10; 4 days, hemin, n=6, buffer, n=6; 3 months, hemin, n=7, buffer, n=4) 60 3.2.3 Quantification of Myocardial Fibrosis Myocardial fibrosis in the paraffin-embedded sections stained with Masson’s Trichrome is shown in Figure 17. Masson’s Trichrome staining represents the presence of collagen by blue staining, while white staining reveals dead tissue and red staining reveals viable tissue. The hemin 4 day representative image reveals a typical hemin infarct whereby there is a longitudinal band of injury and a limited development of fibrosis beyond the peri-vascular ring of collagen. The buffer 4 day image reveals considerable peri-vascular fibrosis and the beginning of a transmural collagen network wherein fibrosis present at 4 days is evident as widespread, diffuse blue specks within the infarct zone. The 3 month hemin representative image displays a mature scar wherein collagen has completely replaced the infarcted myocardium. However, the subendocardium has been salvaged (nearly half the width of the ventricle’s free wall) and its architecture has been maintained. The 3 month buffer image displays a transmural infarct that has been replaced entirely with collagen and no viable myocardium remains. The difference in staining intensity between the 4 day and the 3 month samples likely resulted simply from different experimental conditions or variation in the length of staining. 61 A B C D Figure 17. Myocardial fibrosis. Panel A displays a representative hemin 4 day image with minimal fibrosis and maintenance of the subepicardium and endocardium. The development of fibrosis is more noticeable in Panel B, a buffer-treated animal at 4 days, particularly peri-vascularly. Note the large size of the infarct with very little viable myocardium. Panel C, a hemin 3 month image, shows significant fibrosis despite considerable myocardial salvage. Panel D shows a 3 month buffer-treated animal’s nearcompletely fibrotic left ventricular free wall, including diffuse fibrosis of the papillary muscle. 62 The quantification of fibrosis requires some clarification. Figure 18, which shows the fibrotic left ventricular area normalized for the area at risk at 4 days and 3 months, should be interpreted as follows: at 4 days, the results represent the area of the LV at risk that has shown evidence of the development of fibrosis, not complete replacement of the myocardial architecture (myocytes, endothelial cells) by collagen. The process of collagen deposition requires the homing and trans-differentiation of fibroblasts into myofibroblasts, proliferation of myofibroblasts (which peaks at 4 days) and synthesis/elaboration of collagen (Virag and Murray, 2003). By this definition, fibrosis observed at 4 days has only just commenced. It would be incorrect to interpret the data at 4 days in Figure 18 as the area of the LV that has been replaced with collagen entirely as it biologically impossible to have a mature scar of collagen at 4 days post-I/R injury. The results at 3 months, however, can be interpreted as the ratio of the LV which has been replaced by collagen normalized to the area at risk. Figure 17 clearly showed the differences in the qualitative nature of acute and long-term fibrosis remodeling. The area of myocardium displaying evidence of collagen deposition was significantly greater in the buffer group at 4 days compared to the hemin group (0.382 ± 0.005 vs. 0.802 ± 0.007, P< 0.05; Figure 18). However, no significant difference in fibrosis was seen between the two groups at 3 months, although the hemin-treated group exhibited a trend towards greater mean area of fibrosis (0.869 ± 0.121) than the buffertreated group (0.784 ± 0.081). This may be due to the small sample size and the variability between samples. 63 Fibrosis: Area at Risk 1.2 1.0 * 0.8 0.6 0.4 0.2 m B uf fe r3 3m H em in fe r4 uf B H em in 4d d 0.0 Figure 18. Area of fibrosis relative to the area at risk. Post-MI fibrosis in the left ventricle between hemin and vehicle-treated groups at 4 days and 3 months. A significant difference (P<0.05) was observed between hemin- and buffer-treated animals at 4 days. No significant difference in fibrosis between the two groups was seen at 3 months. (4 days, hemin, n=10, buffer n=8; 3 months, hemin, n=7, buffer, n=4). 64 3.3 Quantification of LV Structure and Function by Echocardiography 3.3.1 LV Wall and Chamber Dimensions The time course of echocardiographic measurements is summarized in Table 1. Because of the anatomy of the LAD and the position of the ligature, the infarcts are predominantly in the mid-papillary and apical region of the LV; the most striking differences between hemin and vehicle animals are seen in those regions. At 4 days, wall thickness was significantly reduced in the buffer-treated group compared to the hemintreated group (Table 1). Significant changes in posterior wall thickness were observed at the apex only in the buffer group. This is likely due to chamber remodeling resulting from the expansion and thinning of the infarct. At 3 months after I/R, hemin and buffer-treated animals showed interventricular septal wall thinning and chamber dilatation at the apical, mid, and base levels. Posterior wall thinning at 3 months was seen in the apex in the buffer group and the apex and mid sections in the hemin group. Anterior wall thinning at the base level in both treatment groups may result from distorted forces exerted upon the interventricular myocardial wall resulting from the infarct expansion in the apical and mid levels and subsequent dysfunctional wall motion rather than infarction at the base level. This wall thinning at the base also may explain the observed chamber dilatation at the base level. It is important to highlight the fact that some LV parameters from the buffertreated groups displayed a significant difference from the hemin-treated group as well as 65 PARAMETER SHAM Wall Dimensions Pre-op 4 Days Post-op 3 Months Post-op IVSdA IVSsA IVSdM IVSsM IVSdB IVSsB 0.127 ± 0.002 0.169 ± 0.002 0.138 ± 0.003 0.172 ± 0.006 0.121 ± 0.001 0.161 ± 0.006 0.122 ± 0.006 0.168 ± 0.001 0.141 ± 0.001 0.179 ± 0.003 0.146 ± 0.004 0.168 ± 0.006 0.155 ± 0.003 0.198 ± 0.007 0.158 ± 0.003 0.216 ± 0.003 0.161 ± 0.001 0.204 ± 0.003 PWdA PWsA PWdM PWsM PWdB PWsB 0.146 ± 0.009 0.197 ± 0.004 0.139 ± 0.004 0.208 ± 0.008 0.134 ± 0.006 0.189 ± 0.014 0.146 ± 0.005 0.199 ± 0.008 0.148 ± 0.006 0.204 ± 0.010 0.147 ± 0.008 0.195 ± 0.012 0.180 ± 0.011 0.230 ± 0.005 0.168 ± 0.005 0.247 ± 0.001 0.174 ± 0.004 0.228 ± 0.007 0.596 ± 0.060 0.364 ± 0.059 0.631 ± 0.059 0.401 ± 0.052 0.664 ± 0.020 0.436 ± 0.022 0.682 ± 0.034 0.404 ± 0.024 0.816 ± 0.022 0.514 ± 0.044 0.791 ± 0.029 0.497 ± 0.014 Chamber Dimensions LVDdA LVDsA LVDdM LVDsM LVDdB LVDsB 0.558 ± 0.055 0.357 ± 0.061 0.603 ± 0.010 0.338 ± 0.015 0.649 ± 0.018 0.418 ± 0.015 Table 1A. Left ventricle wall and chamber dimensions in sham-operated groups. Pre-op, 4 day, and 3 month echo measurements for sham groups (measured in cm ± SEM). (Sham, pre-op, n=3; 4 days, n=3; 3 months, n=3) 66 PARAMETER HEMIN Wall Dimensions Pre-op 4 Days Post-op 3 Months Post-op IVSdA IVSsA IVSdM IVSsM IVSdB IVSsB 0.128 ± 0.003 0.165 ± 0.003 0.132 ± 0.003 0.167 ± 0.003 0.140 ± 0.006 0.172 ± 0.006 0.108 ± 0.004 0.131 ± 0.006 * 0.124 ± 0.003 0.149 ± 0.006 0.127 ± 0.003 * 0.152 ± 0.005 0.106 ± 0.007 * 0.141 ± 0.008 * 0.128 ± 0.008 0.161 ± 0.009 * 0.147 ± 0.003 0.182 ± 0.004 * PWdA PWsA PWdM PWsM PWdB PWsB 0.140 ± 0.003 0.192 ± 0.005 0.149 ± 0.004 0.204 ± 0.005 0.154 ± 0.005 0.209 ± 0.006 0.147 ± 0.004 0.190 ± 0.006 0.144 ± 0.003 0.198 ± 0.006 0.144 ± 0.004 0.200 ± 0.006 0.148 ± 0.004 * 0.178 ± 0.005 * 0.155 ± 0.009 0.206 ± 0.010 * 0.167 ± 0.002 0.217 ± 0.004 0.590 ± 0.019 0.416 ± 0.020 0.648 ± 0.017 0.445 ± 0.018 0.669 ± 0.017 0.480 ± 0.017 0.849 ± 0.036 * 0.658 ± 0.047 * 0.956 ± 0.034 * 0.745 ± 0.038 * 0.933 ± 0.039 0.696 ± 0.037 * Chamber Dimensions LVDdA LVDsA LVDdM LVDsM LVDdB LVDsB 0.529 ± 0.018 0.303 ± 0.016 0.589 ± 0.017 0.365 ± 0.015 0.599 ± 0.017 0.360 ± 0.003 Table 1B. Left ventricle wall and chamber dimensions in hemin-treated groups. Pre-op, 4 day, and 3 month echo measurements for hemin groups (measured in cm ± SEM). Significant difference (P<0.05) from sham is denoted by (*). (Hemin, pre-op, n=15; 4 days, n=15; 3 months, n=7) 67 PARAMETER BUFFER Wall Dimensions Pre-op 4 Days Post-op 3 Months Post-op IVSdA IVSsA IVSdM IVSsM IVSdB IVSsB 0.132 ± 0.003 0.174 ± 0.005 0.138 ± 0.004 0.172 ± 0.005 0.135 ± 0.004 0.172 ± 0.003 0.096 ± 0.004 * 0.114 ± 0.005 * 0.105 ± 0.004 *^ 0.122 ± 0.006 *^ 0.115 ± 0.004 *^ 0.146 ± 0.005 0.095 ± 0.005 * 0.105 ± 0.006 *^ 0.096 ± 0.005 *^ 0.117 ± 0.010 *^ 0.142 ± 0.006 * 0.180 ± 0.001 * PWdA PWsA PWdM PWsM PWdB PWsB 0.147 ± 0.004 0.204 ± 0.006 0.152 ± 0.004 0.206 ± 0.008 0.154 ± 0.004 0.200 ± 0.009 0.129 ± 0.005^ 0.163 ± 0.010 0.139 ± 0.005 0.182 ± 0.007 0.133 ± 0.004 0.196 ± 0.006 0.131 ± 0.008 * 0.164 ± 0.004 * 0.151 ± 0.010 0.205 ± 0.011 0.169 ± 0.003 0.224 ± 0.003 0.661 ± 0.022 0.494 ± 0.026 0.725 ± 0.015^ 0.551 ± 0.021 *^ 0.715 ± 0.023 0.518 ± 0.029 0.864 ± 0.026 * 0.680 ± 0.032 * 1.010 ± 0.041 * 0.791 ± 0.044 * 0.944 ± 0.034 0.711 ± 0.047 * Chamber Dimensions LVDdA LVDsA LVDdM LVDsM LVDdB LVDsB 0.552 ± 0.018 0.324 ± 0.014 0.615 ± 0.012 0.377 ± 0.020 0.637 ± 0.010 0.408 ± 0.016 Table 1C. Left ventricle wall and chamber dimensions in buffer-treated groups. Pre-op, 4 day, and 3 month echo measurements for buffer groups (measured in cm ± SEM). Significant difference (P<0.05) from sham is denoted by (*) while significant difference (P<0.05) between hemin and buffer groups is denoted by (^). (Buffer, pre-op, n=11; 4 days, n=11; 3 months, buffer, n=4). 68 from the sham-operated group. At 4 days after I/R, significantly reduced interventricular septal wall thickness, more pronounced at the mid level, was seen in the buffer group compared to hemin. The upregulation of HO-1 by hemin likely decreased ROS and prevented expansion to some degree at the mid-level, resulting in a significant difference from sham in the hemin group but also protecting the infarct zone from further expansion as seen in the buffer group. This also explains the significant difference in LV chamber dilatation seen at 4 days between the two treatment groups at the mid level, as infarct expansion and wall thinning beget chamber dilatation. Similarly, at 3 months after infarction, a significant difference in interventricular septal wall thinning was seen between hemin and buffer groups, where the IVS wall thinning was attenuated at the apex and mid level LV in the hemin group compared with the buffer group. This is likely due to the prevention of infarct expansion to some degree in the hemin group relative to the buffer group. The percent change of each echocardiographic parameter relative to its baseline is shown in Table 2. At 4 days post-I/R, the hemin-treated group showed considerably less interventricular septal wall thinning than the buffer-treated animals, at all levels of the LV, although this was more pronounced in the apex and mid sections. Similarly, chamber dilatation was more than doubled in the buffer-treated group at both the apex and mid sections, with comparable changes occurring at the base level. The increased posterior wall thickness observed at the apex section of the hemin-treated group may be due to regional compensatory hypertrophy of the remote myocardium. The posterior wall 69 PARAMETER % Change at 4 days % Change at 3 months Wall Dimensions Sham Hemin Buffer Sham Hemin Buffer IVSdA IVSsA IVSdM IVSsM IVSdB IVSsB -3.58 ± 3.9 -0.95 ± 1.5 2.09 ± 2.2 4.41± 5.3 20.2 ± 2.2 9.01 ± 1.8 -15.6 ± 3.1 -20.1 ± 3.6* -5.19 ± 2.9 -10.5 ± 3.9 -7.4 ± 3.2* -11.0 ± 3.0 -27.2 ± 3.0*^ -36.6 ± 2.4*^ -23.2 ± 4.3*^ -31.5 ± 3.2*^ -13.3 ± 3.7* -25.5 ± 5.2*^ 24.5 ± 0.8 16.7 ± 4.8 14.1 ± 1.7 23.4 ± 6.1 32.6 ± 0.8 27.0 ± 6.7 -7.41 ± 6.9* -12.2 ± 6.1* -0.97 ± 7.6* 0.06 ± 6.3 9.7 ± 2.9* 9.31 ± 3.3 -28.3 ± 6.4* -41.7 ± 5.8*^ -30.3 ± 2.2* -32.8 ± 6.6*^ 9.04 ± 5.3* 3.93 ± 4.7* PWdA PWsA PWdM PWsM PWdB PWsB 0.58 ± 9.5 0.91 ± 4.3 6.17 ± 5.7 -1.02 ± 7.4 9.93 ± 9.7 3.52 ± 3.1 5.63 ± 3.2 -0.73 ± 3.2 -5.7 ± 3.0 -2.3 ± 2.7 -4.3 ± 3.2 -5.9 ± 4.8 -13.8 ± 4.3^ -26.7 ± 3.8*^ -9.7 ± 3.7 -11.6 ± 7.5 -14.5 ± 3.9 -3.6 ± 5.7 23.2 ± 5.7 16.9 ± 4.7 20.4 ± 1.2 19.5 ± 4.5 29.6 ± 6.0 21.2 ± 5.0 5.32 ± 3.8 -8.2 ± 5.1* 6.4 ± 7.4 2.39 ± 7.9 12.5 ± 3.9 7.4 ± 4.4 -10.3 ± 4.3* -22.5 ± 4.8* 0.56 ± 5.1 -14.5 ± 3.3 12.1 ± 6.1 6.5 ± 8.7 12.3 ± 3.4 36.5 ± 6.7 11.0 ± 3.9 25.1 ± 8.8 14.4 ± 2.9 35.7 ± 8.3 22.7 ± 8.6 56.4 ± 9.3* 20.4 ± 3.1 60.3 ± 12.5 14.6 ± 3.8 34.9 ± 9.1 23.3 ± 6.3 18.2 ± 14.6 35.3 ± 4.6 52.2 ± 10.7 21.8 ± 8.8 17.8 ± 2.3 58.7 ± 13.4 96.5 ± 17.7* 61.0 ± 13.3 106 ± 28.0 52.9 ± 8.8 83.8 ± 17.7 57.2 ± 5.3 137 ± 14.3* 62.4 ± 3.4 148 ± 24.8 48.9 ± 6.7 83.3 ± 9.4 Chamber Dimensions LVDdA LVDsA LVDdM LVDsM LVDdB LVDsB 6.89 ± 4.9 3.5 ± 8.1 4.42 ± 8.4 18.6 ± 13.1 2.7 ± 5.9 4.69 ± 6.9 Table 2. Percent change in left ventricle dimensions from baseline echo. Results are expressed as percent change from mean pre-op echo dimension to mean 4 day and 3 month echo dimension within each treatment group. Significant difference from comparable sham time point is denoted by (*) (P<0.05), while significant difference between hemin and buffer groups within the same time point is denoted by (^) (P<0.05). (Percent change at 4 days, sham, n=3, hemin, n=15, buffer, n=11; 3 months, sham n=3, hemin, n=7, buffer, n=4) 70 experienced less percent change from baseline in both groups as it is the more remote area of the LV. Attenuation of wall thinning at 3 months in the hemin-treated group is observed at the apical and mid levels only, as there is little difference in percent change between the groups at the base level. At 3 months, chamber dilation during diastole and systole is relatively comparable between the hemin and buffer groups. The percent change in PW observed at the base level of both hemin- and buffer-treated groups is small but likely reflects the animals’ growth from 6 weeks of age to 4.5 months of age. Similar to the acute time-point, the small percent change observed at the apex of the PW in the hemintreated group at 3 months is difficult to reconcile and may be due to the intricacy of imaging the apical PW endocardial borders. Overall, at 4 days, less change was observed at every level of the anterior and posterior ventricle wall in the hemin group compared to the buffer group (except for a small increase in change at the base level of the posterior wall). Similarly, a stark trend of reduced change- nearly by half- in chamber dimension in the hemin group (save for the base level) was observed. At 3 months, changes in anterior and posterior wall thickness were abrogated at the apex and mid levels and comparable at the base in the hemin group compared to buffer. Changes in LV diameter in the hemin group were comparable to buffer in the apex and mid levels during diastole and were considerably reduced during systole, while changes in LV diameter at the base level were similar in both groups. 71 3.3.2 Echocardiography Assessment of Cardiac Function Cardiac function was assessed by calculating ejection fraction (EF), fractional shortening (FS) and LV mass using standard echocardiographic formulas. 3.3.2a Mid Level Ejection Fraction, Fractional Shortening and LV Mass The mid ventricle region of the LV showed the greatest degree of post-I/R change in EF and FS (Figure 19) in both the hemin and buffer-treated groups. Percent change relative to baseline results is shown in Table 2 for each group. Only in the 4-day buffertreated animals was EF significantly decreased relative to its pre-op value and relative to the 4 day hemin animals. At 4 days after I/R, EF was preserved in the hemin-treated animals (Figure 19A). At 3 months, EF was nearly identical between the buffer and hemin groups (0.523 ± 0.026 and 0.526 ± 0.029, respectively). Mid level FS (Figure 19B) showed a trend similar to EF. FS at 4 days in the buffer-treated animals was significantly decreased relative to the sham and hemin groups at the same time point. The difference in FS between the hemin and buffer groups at 3 months was almost indistinguishable (21.90 ± 1.57% and 22.26 ± 1.58%, respectively). No significant differences in LV mass: body weight (Figure 20) were observed among the three study groups at the mid level of the left ventricle at 3 months. 72 A 1.00 0.75 EF *^ * * 0.50 0.25 eo P p re -o p S 4d H 4d B 4d S 3m H 3m B 3m B Pr H S Pr eop 0.00 B 50 FS (%) 40 *^ 30 * * 20 10 S 3m B 3m H 3m S 4d B H 4d 4d S Pr e H -o p Pr e B - op P re -o p 0 Figure 19. Mid level functional echo parameters. Comparison of A) ejection fraction and B) fractional shortening at mid level ventricle among all treatment groups, pre-op and 4 days and 3 months post-MI. Significant difference from sham is denoted by (*, P<0.05), while significant difference between hemin and buffer groups is denoted by (^, P<0.05). (Pre-op, sham, n=3, hemin, n=15, buffer, n=11; 4 days, sham, n=3, hemin, n=15, buffer, n=11; 3 months, sham n=3, hemin, n=7, buffer, n=4) 73 0.004 0.003 0.002 0.001 3m B 3m H 3m S B Pr eop Pr H Pr S eop 0.000 eop LV Mass to Weight Ratio 0.005 Figure 20. Mid LV mass to body weight. Left ventricle mass normalized for body weight at mid level (measured along the parasternal long axis) among all treatment groups, pre-op and 3 months post-MI. No significant difference was observed. (Pre-op, sham, n=3, hemin, n=7, buffer, n=4; 3 months, sham n=3, hemin, n=7, buffer, n=4) 74 3.3.2b Apex Level Ejection Fraction, Fractional Shortening and LV Mass Significant difference in apical EF from sham was observed in both hemin and buffer groups at 3 months and the values are similar (0.5386 ± 0.04 and 0.5100 ± 0.03, respectively; Figure 21A). Though no significant difference was observed at the 4 day time-point among all treatment groups, the mean EF in the hemin group is greater than the mean EF in the buffer group (0.6387 ± 0.032 and 0.5608 ± 0.057, respectively). The trend observed in EF is also seen in the FS results (Figure 21B). Significant difference from sham was observed in both hemin and buffer groups at 3 months and the values are similar (23.07 ± 2.25% and 21.40 ± 1.66%, respectively). Though no significant difference was noted among treatment groups at 4 days (likely due to considerable standard error of the mean), the mean FS of the hemin group is greater than the mean FS in the buffer group (29.70 ± 2.14 and 25.38 ± 2.92, respectively). No significant differences in LV mass: body weight (Figure 22) were observed among the three study groups at the apex level of the left ventricle at 3 months. 75 A 1.00 EF 0.75 * * 0.50 0.25 S Pr e H -o p Pr e B - op P re -o p S 4d H 4d B 4d S 3m H 3m B 3m 0.00 B 50 FS (%) 40 * 30 * * 20 10 S 3m B 3m H 3m S 4d B H 4d 4d S Pr e H -o p Pr e B - op P re -o p 0 Figure 21. Apex level functional echo parameters. Comparison of A) ejection fraction and B) fractional shortening at the level of the apex (measured along the parasternal long axis) among all treatment groups, pre-op and 4 days and 3 months post-MI. Significant difference from sham is denoted by (*, P<0.05), while significant difference between hemin and buffer groups is denoted by (#, P<0.05). (Pre-op, sham, n=3, hemin, n=15, buffer, n=11; 4 days, sham, n=3, hemin, n=15, buffer, n=11; 3 months, sham n=3, hemin, n=7, buffer, n=4) 76 0.004 0.003 0.002 0.001 3m B 3m H 3m S H Pr e S Pr eop B Pr eop 0.000 -o p LV Mass to Weight Ratio 0.005 Figure 22. Apex LV mass to body weight. Left ventricle mass normalized for body weight at the level of the apex (measured along the parasternal long axis) among all treatment groups, pre-op and 3 months post-MI. No significant difference was observed. (Pre-op, sham, n=3, hemin, n=7, buffer, n=4; 3 months, sham n=3, hemin, n=7, buffer, n=4) 77 The percent change of each derived parameter from its baseline measurement is shown in Table 3. The decrease in EF observed at the apex and mid levels in the buffer group at 4 days is attenuated in the 4 day hemin group at the apex and mid levels but not at the level of the base. A similar trend is observed in FS at 4 days. While there was no observed statistically significant difference in EF and FS between the hemin and buffer 3 month groups, there is a trend whereby the decline in function in the hemin group is attenuated by approximately 5-15% compared to the change observed in the buffer 3 month group. The percent change in EF and FS within the sham group is marginal at the apex and base, but there was an unexpected yet modest decrease in EF and FS at the mid level. This could be due to the anaesthesia used during the echo procedure. LV mass at 3 months decreased in a similar fashion among the sham, hemin, and buffer groups (Table 4). There was no significant difference among the groups and the decrease is representative of the animals’ growth from 6 weeks of age to 4.5 months, thereby decreasing the ration of LV mass to body weight. 78 % Change at 4 days LV Function SHAM HEMIN BUFFER % Change at 3 months SHAM HEMIN BUFFER EF Apex 5.1 ± 3.8 -20.9 ± 4.3 -32.4 ± 7.4* 8.9 ± 8.0 -30.9 ± 6.5* -39.8 ± 3.6* EF Mid -9.4 ± 3.1 -10.7 ± 3.7 -29.9 ± 4.7^ -9.5 ± 3.9 -28.7 ± 5.3 -38.4 ± 3.6* EF Base -1.5 ± 3.7 -15.4 ± 4.7 -12.2 ± 5.7 -20.2 ± 6.2 -25.3 ± 3.4* FS Apex 8.2 ± 6.4 -30.1 ± 5.7* -42.2 ± 6.98* 15.0 ± 14.6 -41.6 ± 7.4* -54.7 ± 3.9* FS Mid -16.3 ± 4.9 -15.5 ± 5.2 -39.4 ± 6.1^ -12.9 ± 3.4 -38.2 ± 6.5 -53.3 ± 4.6* FS Base -2.5 ± 5.9 -22.4 ± 6.6 -24.2 ± 6.6 -28.7 ± 9.0 -36.1 ± 3.2* 3.9 ± 2.9 6.8 ± 5.0 Table 3. Percent change in functional parameters from baseline echo. Results are expressed as percent change from mean pre-op echo values to mean 4 day and 3 month echo values within each treatment group. (Percent change at 4 days, sham, n=3, hemin, n=15, buffer, n=11; Percent change at 3 months, sham, n=3, hemin, n=7, buffer, n=4; P<0.05) 79 % Change at 3 months Ventricle Mass SHAM HEMIN BUFFER LV Mass:Wt Apex -49.1 ± 4.0 -39.8 ± 2.9 -46.6 ± 4.3 LV Mass:Wt Mid -45.4 ± 3.9 -34.0 ± 3.7 -39.5 ± 3.6 LV Mass:Wt Base -42.9 ± 5.9 -25.2 ± 4.0 -26.4 ± 2.9 Table 4. Percent change in left ventricle mass. Results are expressed as a ratio of LV mass normalized to body weight. No significance was observed at 3 months. (Percent change at 3 months, sham n=3, hemin, n=7, buffer, n=4) 80 3.4 Quantification of NFκB-DNA Binding Post-reperfusion NFκB-DNA binding activity was significantly attenuated in the 4 day hemin group compared to buffer (Figure 23). While not statistically significant, the NFκB activity (represented by relative light units) in the buffer 24 hour group is notably greater than the NFκB activity observed in the hemin 24 hour group. Together, this suggests that HO-1 upregulation abrogates the transcription factor’s translocation to the nucleus and/or activity despite redox stimulation from ROS due to reperfusion. 3.5 Effect of Hemin Treatment on Hematocrit Hematocrit was measured at cardiac harvest at all three time points within all three treatment groups, though no significant differences were observed (Figure 24). In fact, very little variation in the mean hematocrit was seen between sham, hemin, and buffer groups within each time group, though the mean hematocrit results are slightly lower at the 24 hour time point. 81 Relative Light Units (RLU/μ g) 1.5 * 1.0 0.5 4d B 4d H 24 h B H 24 h 0.0 Figure 23. NFκB-DNA binding activity after I/R. NFκB activity post-reperfusion was significantly elevated in the buffer-treated animals at 4 days post-MI (P<0.05). While not statistically different, the mean NFκB activity observed in the buffer 24 hour group is greater than the mean observed in the hemin 24 hour group. (24 hours, hemin, n=3, buffer, n=3; 4 days, hemin, n=4, buffer, n=4) 82 Hematocrit (%) 60 40 20 S2 4h H 24 h B 24 h S4 d H 4d B 4d S3 m H 3m B 3m 0 Figure 24. Hematocrit at cardiac harvest. No significant difference was observed among the groups at each time point. However, the mean hematocrit value for each treatment group is slightly lower at 24 hours compared with the other time points. (24 hours, sham, n=4, hemin, n=3, buffer, n=5; 4 days, sham, n=3, hemin, n=6, buffer, n=5; 3 months, sham, n=3, hemin, n=6, buffer, n=5) 83 Chapter 4: Discussion 4.1 Major Findings This thesis tested the hypothesis that HO-1 upregulation via hemin administration attenuates acute and chronic I/R-induced myocardial injury and subsequent postmyocardial infarction left ventricular remodeling. The results showed that treatment with hemin prior to I/R significantly reduced acute myocardial injury, leading to reduced infarction size and improved post-MI recovery at 4 days. However, chronic hemin treatment did not prevent long term left ventricular remodeling, despite its acute cardioprotective effects. The reasons for this discrepancy have not been elucidated in the present study, but could possibly be attributed, at least in part, to the reduced sample size of the 3 month study as well as biological differences in infarct size. Alternatively, HO-1 upregulation may not be maintained by chronic hemin administration such that it continues to exert cardioprotective properties. The acute protective effect of hemin treatment was associated with reduced myocardial fibrosis and was accompanied by a decrease in DNA binding activity of the redox-sensitive transcription factor NFκB, suggesting that inhibiting this pro-inflammatory transcription factor may underlie the mechanism by which hemin reduces acute myocardial injury. However, the intermediate signalling pathways leading to inhibition of NFκB by hemin are not known. The injection of hemin was associated with induction of HO-1 protein expression and with reduced myocardial injury, suggesting that the enzyme confers cardioprotection via the byproducts of heme catabolism, CO, biliverdin, and iron. However, the isolated effects of the byproducts on myocardial protection were not assessed in this thesis. 84 4.2 Histological Findings 4.2.1 Myocardial Infarctions Because of inter-animal variability in risk area after ligation of the LAD, infarct size was calculated as a fraction of the area at risk for all animals. The predominant acute histopathological finding is decreased infarct size in the hemin-treated animals at 24 hours and 4 days after I/R compared with buffer-treated animals. H&E staining in the hemin-treated animals revealed reduced area of infarction and the maintenance of myocardial subendocardium within the area at risk compared to buffer-treated animals. Generally, infarction in hemin-treated animals looked like a discrete longitudinal band of injury within the cross-section of the left ventricle with a distinct infarct border. Furthermore, histology revealed non-infarcted, viable subendocardium with architecture similar to sham-operated animals in nearly all 24 hour and 4 day hemin-treated animals. Notwithstanding biological differences in coronary artery distribution and architecture, this may be a genuine pattern of injury among HO-1-upregulated animals, although proving a direct association between HO-1 levels and this pattern of myocardial injury is beyond the scope of this thesis. Buffer-treated animals at the same time points experienced infarcts that appeared more diffuse and involved all layers of the myocardium, including the subendocardium and the subepicardium, with borders that extend beyond the anterolateral wall of the left ventricle. Circumferential and nearcircumferential infarcts were observed only in buffer-treated animals. The reason for the observed reduced infarct size within the 24 hour and 4 day hemin-treated animals is likely multifactorial. 85 The HO-1 byproducts decrease the inflammatory reaction upon reperfusion by decreasing endothelial dysfunction and by enhancing the anti-oxidant reserve, leading to increased tolerability of the myocytes to the pro-inflammatory and pro-oxidant conditions of the infarct environment. It is likely these protective functions of HO-1 together are responsible for decreased neutrophil infiltration and decreased ROS generation, curtailing the cytokine/chemokine elaboration and decreasing the release of proteolytic enzymes. This in turn limits the cellular injury and decreases myocyte and endothelial cell apoptosis, as evidenced by a decreased area of infarction and the maintenance of myocardial integrity within the subendocardium. Residual myocardial viability after infarction is important not only for contraction and cardiac performance but the thickness of the surviving myocardium is also inversely associated with infarct expansion within the infarct zone (Pirolo et al., 1986). While a certain degree of necrosis is unavoidable after prolonged ischemia, Clark et al. (2000) reported that rat hearts displaying high levels of HO-1 protein expression showed considerably reduced infarct size and preserved mitochondrial integrity after ischemia reperfusion. Vulapalli et al. (2002) reported an antiapoptotic activity of specific HO-1 over-expression in protecting cardiomyocytes from I/R-induced apoptosis in vivo, and Tang et al. (2004) found that Bcl-2, an anti-apoptotic protein, is upregulated in myocardium treated with a hypoxia-sensitive hHO-1 plasmid compared with myocardium treated with saline. The precise mechanism responsible for the observed reduction in infarct size among the hemin-treated animals in unknown but is likely a combination of the enzyme’s pleiotropic activities. The 3 month hemin and buffer groups, on the other hand, had comparable mean areas of infarction. There was greater variability in the appearance of the infarcts within 86 the hemin group, unlike the acute time points. This may be attributable, at least in part, to technical variability as well as biological variation between the animals. In addition, it is also possible the animals did not adapt to the initial I/R insult in the same manner which may have been exaggerated over 3 months. Hemodynamic changes including blood volume, arterial blood pressure, systemic vascular resistance and pulmonary pressure may have changed secondary to left ventricular remodeling, which would impair basal LV function and hemodynamics and would be more prevalent at 3 months after reperfusion. 4.2.2 Myocardial Fibrosis The difference in the development of fibrosis at 4 days post-MI between heminand buffer-treated animals was significant. The mean infarct size in the buffer 4 day group was significantly greater and is therefore expected to be followed by extensive reparative fibrosis to preserve structural integrity of the infarcted tissue (Sun et al., 2000). In contrast, there was no statistically significant difference between the two groups at 3 months. Fibrosis developed beyond the infarct zone and in remote regions in both groups with web-like spindles of collagen deposited throughout the ventricle unlike the concentrated areas of fibrosis in the acute study. Both 3 month treatment groups experienced significant infarcts and left ventricular remodeling (including chamber dilatation) as well as reduced cardiac performance, suggesting the animals were progressing to heart failure. It is now recognized that accumulation of collagen occurs in the interstitial space of the hypertrophied and failing heart, a process termed “reactive 87 interstitial fibrosis” (Weber and Brilla, 1991; Sabbah et al., 1995). The exact mechanism that promotes the accumulation of fibrosis in the remote areas of the myocardium is not fully understood, but has been attributed, in part, to enhanced activity of the reninangiotensin system (Weber and Brilla, 1991). Cleutjens et al. (1995) demonstrated that myofibroblasts are responsible for type I and III collagen synthesis in the repairing myocardium; however, myofibroblasts are not found within interstitial fibrosis at noninfarcted remote sites after MI (Sun et al., 2000). This suggests that some other mechanism, dissimilar to reparative fibrosis, is responsible for the collagen accumulation in remote myocardium, perhaps increased TGF-β1 production by interstitial macrophages (Sun et al., 2000). The development of fibrosis beyond the infarct zone may be a compensatory attempt to strengthen the myocardial wall post-I/R injury or may simply be a response to chronically elevated TGF-β1 levels. This possibility may explain why some of the values of fibrosis relative to the area at risk were greater than 1.0. It is also noteworthy that this reactive interstitial fibrosis is maladaptive in that the expansion of the interstitial space by the deposition of collagen results in decreased capillary distance and increased oxygen diffusion distance, which could lead to chronic hypoxia and adversely influence the function and viability of the collagen-encircled myocyte (Sabbah et al., 1995). 4.3 Echo Findings In agreement with the histological findings, the echocardiographic analysis in LV wall and chamber dimensions revealed reduced wall thinning and decreased enlargement 88 of the LV cavity in the hemin group relative to the buffer group at 4 days post-MI. Changes in wall thickness were localized to the anterior LV wall in the hemin group, whereas the buffer group showed significant thinning of the posterior wall compared to the sham and hemin groups. The anterior wall thinning observed in the 4 day hemin group at the base level is unexpected, particularly in light of the absence of significant thinning at the mid level. There is a possibility this is a technical error in which the placement of the 2D echo image cursor along the theoretical border between the base and mid levels of the left ventricle was too close to the mid section but mistaken as base level instead. This is likely the reason since there is no other remodeling of the LV at the base level in the hemin 4 day group. Although it may seem surprising to see significant LV remodeling at all 3 levels of the ventricle in the buffer group just 4 days post-MI, similar findings have been reported by others in animals with large infarcts. For example, Weisman and colleagues (1985) showed significant wall thinning and increased regional radius of curvature in adjacent and remote regions of the heart within just 3 days after infarction. At 3 months, the degree of anterior wall thinning was still attenuated in the hemin-treated animals compared to the buffer-treated animals despite comparable infarct sizes. However, in spite of the attenuated wall thinning in the hemin-treated animals, LV cavity enlargement did not differ significantly from the buffer-treated groups. Even though significant attenuation in anterior wall thinning at the mid and apical levels was observed in the hemin group, the mean infarct size was still large (> 40%), suggesting that the presence of cavity enlargement may be an indication of blood volume congestion in an attempt to maintain adequate CO in the face of reduced EF (from 75% pre-op to 89 50% at 3 months). If the injury to the myocardium is substantial enough to affect pump function, then it is plausible there is a compensatory increase in renin production in an attempt to normalize blood volume and cardiac output. This increased volume exerts an outward pressure on the ventricle walls in all directions, thereby explaining the cavity dilatation despite attenuated anterior wall thinning. Thus, global myocardial remodeling after infarction leads to a condition of overload on the surviving myocardium that consists primarily of a volume component though a pressure component may be operative as well (Olivetti et al., 1990). Congestion within the circulatory system significant enough to effect cavity dilatation would occur over the long term and would indeed be seen at 3 months post-MI in rats. Interestingly, wall thinning and chamber dilatation were seen at all levels of the LV at 3 months in both groups. It is surprising to observe anterior wall thinning at the base of the ventricle (occurring in both treatment groups) since the area at risk for infarction is primarily the mid and apical sections. However, both groups experienced infarcts large enough to induce significant thinning at the anterior and posterior apical wall as well as the anterior mid region. When disproportionate wall thinning and chamber dilatation occur in the infarct region, they are accompanied by a distortion in shape of the entire heart including the normal remote myocardium (Weisman et al., 1985). In both 3 month treatment groups, the mean chamber diameter changed very little from the level of the apex to the level of the base, indicating both groups had adopted a sac-like cardiac phenotype. The degree of ventricular remodeling was great enough that the normal ellipsoid shape of the ventricle was lost. Upon volume overload, increased end-diastolic pressure causes an increase in diastolic wall stress. Chamber enlargement 90 accommodates the volume overload and returns diastolic pressure toward normal at first, but chamber enlargement also results in increased systolic wall stress (McKay et al., 1986). The anterior wall could have been subjected to forces and/or mechanical stretch from blood volume congestion and aneurysm formation at the apex and mid level, culminating in an apparent reduction in wall thickness at the base, despite a lack of infarction at that level. Furthermore, wall thinning and architectural distortion at the base could potentially lead to the cavity dilatation observed at the base level of both treatment groups. The significantly reduced posterior wall thickness at the mid level of the 3 month hemin group is surprising, especially because it is nearly identical to the mean observed in the buffer group (0.206 ± 0.10 cm vs 0.205 ± 0.011 cm, respectively). In both 3 month treatment groups, the mid level posterior wall measurements are nearly identical to the measurements at the base. This could indicate simply that the mid posterior wall did not grow over 3 months in the groups that underwent LAD ligature compared with shamoperated group. It is unlikely that either treatment group experienced infarctions so large as to induce wall thinning at the mid level posterior wall; however this possibility can not be totally discounted. It is more probable, though, that the absence of increased posterior wall thickness is due instead to considerable architectural changes at the apex and anterior mid section, generating distorting forces and potentially subjecting the mid posterior wall to mechanical stretch mistakenly interpreted as wall thinning due to infarction. 91 4.4 HO-1 Upregulation and Preservation of Coronary Vascular Homeostasis A principal component in preventing left ventricular remodeling is myocardial salvage and the prevention of wall thinning and infarct expansion. As previously mentioned, HO-1 upregulation reduces infarct size through multiple mechanisms, one of which is likely a decrease in myocyte apoptosis which could explain the preservation of the subendocardium revealed by H&E as well as the attenuated wall thinning observed in the 4 day hemin group. One of the primary goals of reperfusion is to achieve infarctrelated artery patency which also influences the progression of ventricular remodeling. It is critical that the remaining viable myocardium has unobstructed blood flow in order to restore cellular ATP, re-establish cellular homeostasis and reverse the injury induced by reperfusion. It is possible that some of the protection afforded by HO-1 upregulation occurs through decreased neutrophil adhesion to the coronary microcirculation and decreased occurrence of “neutrophil plugging”, thereby allowing prompt reperfusion to the ischemic area. The production of CO through heme metabolism may ameliorate I/Rinduced endothelial dysfunction and reduce coronary vascular tone and possibly prevent the no-reflow phenomenon. 4.5 Effect of HO-1 Upregulation of Post-Infarction Left Ventricular Function In agreement with the hypothesis that HO-1 upregulated animals exhibit attenuated ventricular remodeling, post-infarction ejection fraction and fractional shortening were maintained in the 4 day hemin-treated animals while the buffer-treated 92 group revealed significantly reduced EF and FS compared to sham and hemin groups. The preservation of left ventricle structural integrity in the 4 day hemin-treated group likely enables the ventricles to maintain proper pump function. Although LV wall thinning was attenuated in the 3 month hemin-treated group compared to the buffer group, the mid level ejection fraction and fractional shortening at this time point were nearly identical in both groups. This may be an indication that the degree to which the wall thinning was attenuated was not sufficient to prevent volume congestion and subsequent pump dysfunction, evidenced by the significantly reduced EF and FS in the hemin group at 3 months compared to sham. Acute compensatory responses serve to maintain pump function, including distension of the viable myocardium and the operation of the Frank-Starling mechanism, and augmentation of chronotropic and inotropic receptor stimulation (Pfeffer and Braunwald, 1990). It is possible that the hemin-treated animals involved in the long-term study had sufficient acute compensatory mechanisms to maintain normal cardiac function at 4 days but not once they reached the 3 month time point. Moreover, it is important to consider that post-infarction left ventricular remodeling in rats takes several months to complete. Apical level EF and FS followed the same pattern observed at mid-level with the exception that apical level EF was not significantly reduced at 4 days in the buffer-treated group compared to sham. The mean EF was, however, less than both the sham and hemin-treated groups. This may simply reflect varying degrees of infarction of the apical region in this group, which could be due to differences in the architecture of the coronary microvasculature in this region. 93 4.6 Comparison of Short and Long Term Studies While it was surprising at first to observe the unmistakeable prevention of acute LV remodeling in the hemin group compared with the significant remodeling that took place by 3 months, it should not be understated that the long term remodeling in the hemin group was attenuated compared to the buffer group and may be biologically significant. The percent change from baseline measurements in the hemin 3 month group was noticeably decreased at the apex and mid level anterior and posterior walls. Moreover, the expansion of the LV in the hemin group at the apex and mid levels was reduced by 30% during systole compared to buffer. Coupled with the attenuation of wall thinning at the apex and mid levels compared to buffer, this could indicate some preservation of systolic function compared to buffer. The 4 day time point is quite early within the remodeling process when it is not yet complete. It is possible that HO-1 upregulation prevents remodeling in the short term, before the insidious onset of architectural changes, or minimizes and/or delays the process. It is also feasible that long term hemin administration does not upregulate HO-1 as expected. It is possible that long term hemin administration actually causes some degree of toxicity rather than protecting tissues by repeated exposure of oxidative stress; this could likely be the problem since hemin administration post-MI induces additional oxidative stress independent of the oxidative stress due to the generation and activity of ROS upon reperfusion. Alternatively, it is possible that continued hemin administration does upregulate HO-1, but the window of opportunity for cardioprotection expires beyond 4 days. 94 Failing heart muscle generally exhibits distinct changes in intracellular Ca2+ handling including impaired removal of cytosolic Ca2+; reduced Ca2+ loading of the cardiac sarcoplasmic reticulum (SR) with downregulation of SR Ca2+-ATPase 2 (SERCA2); and defects in SR Ca2+ release accompanied by impairment of cardiac relaxation and systolic function (Marx et al., 2000; Morgan et al., 1990; Nordlie et al., 2006). It is possible that increased HO-1 expression and a codependent activation of soluble guanylyl-cyclase could lead to an increase in cGMP levels and to a decrease in calcium overload (Maines, 1997), all of which involves components of the calcium handling system including SERCA2a and phospholamban. Indeed, SERCA2a gene transfer has proven to normalize LV volumes and improve cardiac function in rats with induced heart failure, while ablated phospholamban resulted in complete prevention of the heart failure phenotype in a mouse model of dilated cardiomyopathy, along with improved cardiac SR Ca2+ storage and release (del Monte et al., 2001; Minamisawa et al., 1999). Considering the evidence provided above, one might have anticipated that the hemin 3 month group would have exhibited preserved cardiac performance, at least to some degree. The fact that it didn’t could strengthen the hypothesis that the upregulation of HO-1 was not as robust in the 3 month hemin group as it was observed in the 4 day hemin group. 4.7 Changes in Cardiac Mass There were no significant differences in LV mass post-infarction between the hemin and buffer-treated animals compared to sham at any level of the ventricle at 3 95 months. LV mass (expressed as a ratio to body weight) decreased markedly in all 3 experimental groups because the animals’ body weight increased far more over 3 months than did their heart mass. There were also no significant differences among the groups in the percent change from baseline within each group. If ventricular volume had been measured instead, there likely would have been significant increases in both hemin and buffer 3 month groups considering the LV chamber dilatation observed in both groups. Pfeffer et al. (1982) examined cardiac mass and volume at 2, 21, and 100 days after MI and noted a progressive decrease in the ratio of mass to volume, indicating that LV volume was increasing out of proportion to mass. This suggests that volume-overload hypertrophy, marked by sarcomere replication in series, fiber elongation and chamber enlargement, occurs perhaps in response to increased end-diastolic wall stress (McKay et al., 1986; Pfeffer et al., 1982). Considering there was significant cavity enlargement but no significantly increased cardiac mass in both 3 month groups in the current study, it is possible that both hemin and buffer 3 month groups experienced volume-overload as proposed by McKay et al. (1986). 4.8 Role of NFkB in I/R injury A significantly increased level of NFκB binding activity was observed in the 4 day buffer group compared with the hemin group at the same time. Although no statistical significance between the hemin and buffer groups was found at 24 hours postMI, the buffer group displayed a trend towards higher NFκB-DNA binding activity. Much of the reperfusion-induced inflammatory response is due to modified gene 96 expression involving NFκB activity (Brown et al., 2005; Mercurio and Manning, 1999). It is possible the smaller infarct size within the hemin-treated groups at 24 hours and 4 days is partly due to decreased apoptosis and reduced inflammatory response due to lower NFκB activation than seen in the buffer-treated group. NFκB has been implicated in the regulation of a myriad of pro-inflammatory and pro-apoptotic genes that participate in I/R injury such as intracellular adhesion and chemoattractant molecules as well as inflammatory cytokines (IL-1β, IL-6, IL-8 and TNF-α) (Brown et al., 2005). Brown et al. (2005) reported >70% reduction in infarct size in NFκB activation-blocked transgenic mice, clearly demonstrating the overall activity of NFκB in the heart after I/R leads to injury. The significant difference between the two groups at 4 days represents the second wave of NFκB activation in keeping with the second wave of cytokine expression within the remodeling phase. While the NFκB DNA binding activity at 24 hours was not statistically significant (this could be due in part to the small sample size), there is a trend towards greater NFκB activation among the buffer-treated animals. An argument could be made that the decreased NFκB activation in the hemin 24 hours group is biologically significant considering how central NFκB activation is to the propagation of inflammation in reperfusion injury. Since stimulation of NFκB by oxidative stress triggers the release of NFκB from IκB proteins and results in its translocation to the nucleus to bind to DNA, one of the mechanisms by which increased HO-1 expression attenuates NFκB binding activity may occur by increasing antioxidant capacity even prior to the I/R and subsequently reducing 97 activation of NFκB. NFκB activation is dependent on the phosphorylation and degradation of IκB-α (Henkel et al., 1993); it is possible that increased HO-1 activity via its by-products interferes with one or several essential components of the signal transduction cascade involved in activation of NFκB (Rossi et al., 1997). Moreover, Anderson et al. (1994) reported that tyrosine kinase activity is a common requirement for NFκB activation and that tyrosine kinase activity is increased by oxidants (ROS upon reperfusion, for example) and decreased by antioxidants (bilirubin, for example). 4.9 Hematocrit and Cardiac Function Post-I/R Injury Erythropoetin (EPO), a cytokine produced by the adult kidney, is a well known hematopoietic factor and increased HO-1 expression has recently been linked to EPO treatment in hemodialysis patients (Moon et al., 2003; Calo et al., 2006). A benefit of increasing the oxygen-carrying capacity of blood by increasing the red blood cell mass (hematocrit) by blood transfusion has been reported in elderly patients with acute MI (Wu et al., 2001). The advantage of increasing oxygen-carrying capacity in a state of oxidative stress is obvious. However, in this study, acute and chronic upregulation of HO-1 by hemin did not result in significant differences in hematocrit at cardiac harvest among any of the groups. It is possible the upregulation of HO-1 by hemin was not robust enough to mediate changes in EPO expression. Conversely, EPO expression may not be amplified or influenced by increased HO-1 activity in the setting of I/R injury. Although EPO’s antiapoptotic effects strongly point to a central role for HO-1 as a 98 mediator, the literature reveals a very limited understanding of the interaction between HO-1 and EPO and the pathways they influence (Calo et al., 2006). 4.10 Potential Therapeutic Application of HO-1 Up-regulation in Myocardial Infarction This project demonstrated a beneficial effect of hemin pre-treatment on myocardial reperfusion injury via HO-1 upregulation in the short term and a mild attenuation of injury in the long term. The trend of current and future research continues to focus on methods of translating the protective properties of the endogenous HO-1 stress response into therapeutic strategies for the treatment of human disease (Ryter et al., 2006). For example, experimentation with natural antioxidants and pharmaceuticals has raised the possibility that HO-1 induction in tissue could be stimulated by agents selected for their capacity to invoke the response without eliciting tissue damage (Ryter et al., 2006). Some investigators believe hemin in particular is contra-indicated during a period of tissue vulnerability. Balla et al. (1991) showed that hemin rapidly intercalates into cultured endothelial cells resulting in their marked hypersusceptibility to subsequent oxidant-mediated cytolysis by H2O2 or adherent polymorphonuclear leukocytes. These authors further described the dichotomous effects of heme exposure on endothelial cells: heme amplifies oxidant-mediated cytotoxicity during brief (2 h) exposure, yet markedly protects oxidant-exposed target cells if provided for longer durations (Balla et al., 1992). Because free iron is a potent pro-oxidant, increasing its tissue concentration via HO-1 upregulation during a reperfusion-induced respiratory burst is not clinically feasible. 99 Furthermore, it takes time for HO-1-upregulation to result in subsequent ferritin upregulation and iron sequestration. For this reason, the administration of CO and biliverdin and/or bilirubin, rather than the induction of HO-1, may prove to be a safer and more practical means of conferring cytoprotection in a clinical setting (Kirkby and Adin, 2006). Patients stratified at high risk for MI could be considered for pre-emptive therapy involving the upregulation of HO-1 directly or administration of HO-1 byproducts. As previously mentioned, carbon monoxide releasing molecules (CORMs) provide a safer, alternative method of CO delivery than does direct CO inhalation which could induce tissue hypoxia. Indeed, the administration of CORMs just before reperfusion has proven to cause a reduction in infarct size similar to the effect afforded by early or late preconditioning (Guo et al., 2004). The administration of CORMs and albumin-bound bilirubin at the time of reperfusion in clinical practice might be an attractive method of limiting reperfusion damage and enhancing cellular antioxidant defense. Another promising application of HO-1 upregulation is the development of hypoxia-regulated vectors for delivery of HO-1. Because myocardial ischemia oftens occurs silently with multiple episodes in the clinical setting, prompt treatment is required in spite of the absence of symptoms. Pachori et al. (2006) provided the first evidence of the therapeutic efficacy of pre-emptive adeno-associated virus-HO-1 delivery for the prevention against recurrent ischemic injury. Furthermore, our laboratory has already shown the feasibility of AAV-based HO-1 gene delivery for prevention of LV remodeling after single and recurrent episodes of I/R (Liu et al., 2006; Melo et al., 2002). It should be recognized, however, that the systemic effects of chronic HO-1 upregulation have not yet been fully elucidated. In fact, constitutive overexpression of 100 HO-1 may lead to cellular toxicity that may in turn lead to kernicterus and tissue hypoxia caused by the catalytic byproducts of heme degradation (Platt and Nath, 1998). Tang et al. (2004) developed a hypoxia-sensitive vigilant human HO-1 plasmid in mice that provided cardioprotection through decreased apoptosis and fibrosis in the setting of myocardial ischemia. The next step in the evolution of this endogenous cellular defense system fit for short term protection may be to construct an adeno-associated viral plasmid to make a viral vigilant long-term cardioprotective system (Tang et al., 2004). 4.11 Study Consideration It is important to appreciate that coronary artery ligation in rats is not akin to the complex development of coronary artery disease leading to myocardial infarction requiring interventional reperfusion in the clinical setting. However, investigating postI/R histological and LV functional changes in animals provides insight to the changes that occur from the onset of ischemia to reperfusion that lead to infarct expansion, left ventricle chamber dilatation, ventricle function depression, and decreased cardiac output, all of which may be likened to dilated cardiomyopathy and CHF. Pfeffer et al. (1979) observed a wide range of physiological outcomes in the rat infarction model: normal baseline ventricular filling pressures and cardiac output were observed in rat infarctions constituting less than 47% of the left ventricle while more extensive infarcts were associated with overt depression of cardiac performance manifested by elevated left ventricle filling pressures and reduced mean arterial pressure and cardiac output. They concluded that the rat model of coronary artery ligation provides an entire range of postinfarction function from an absence of detectable impairment to gross failure (Pfeffer et 101 al., 1979). It is therefore important to recognize that recovery from ischemia reperfusion injury after a large infarction (i.e. >40% of the left ventricle) in humans may not be as successful as is often noted among rats and likely involves more complicated, multivariate pathologies. In fact, ischemic heart disease is the most common cause of CHF among human patients and often begins after acute myocardial infarction; moreover, it is associated with a twofold increase in in-hospital morbidity and a fourfold increase in in-hospital one year mortality (O’Connor et al., 1997). Although conclusions drawn from animal models in basic science research may not necessarily be directly applied to the clinical setting, the endeavour to reduce reperfusion injury and salvage viable myocardium post-MI through experimentation is warranted and should remain a priority. 4.12 Study Limitations There are several technical and design limitations to consider within the current study. 4.12.1 Sample size Sample sizes upwards of twenty to thirty for each treatment group would have been impossible to achieve due to the volume of experiments and time required for each animal. This project focused on whole animal experiments to organ function studies and molecular studies, all of which incurred a huge potential for biologic variation at any point from the LAD ligation surgery to organ harvest. Hence, one must be careful about 102 drawing robust conclusions from the modest sample size in the long-term (3 month) study. Surprisingly, three of the seven animals within the buffer-treated group did not in fact experience a myocardial infarct in the LAD coronary artery (confirmed by histology) and were excluded. An intrinsic limitation to this study was the inability to confirm with certainty the production of a myocardial infarct while keeping the animal alive. 4.12.2 Use of volatile anaesthetic Halothane was used to sedate the animals for the echo procedures. This volatile anaesthetic was selected for its rapid induction of sedation and relative ease to administer. However, volatile anaesthetics are extremely fast-acting and no gauge was available to monitor the amount given to each animal. Two animals died not from coronary artery ligation nor reperfusion injury but from halothane overdose due to unpredictable responses to the drug. Furthermore, the effects of halothane on cardiac function were not addressed. It would have been more prudent to use a ventilator equipped to deliver measurable concentrations of anaesthesia known not to depress cardiac function. 4.12.3 Echocardiography technique The animals’ chests were shaved using rodent hair clippers and commercially available disposable razors. In order to obtain clear 2-D and M-mode images, the animals needed to be shaved very closely and required applying pressure to the chest wall with each stroke of the razor. Because this was done while the animals were sedated, it was challenging to accomplish good skin preparation without imposing pressure on the 103 heart. It may prove advantageous instead to use dilapitory cream for hair removal in preparation for any future echocardiography studies in the lab. Imaging of the rat’s apex is technically challenging. This is in part due to the size of the rat heart’s apex and due to its position in the chest. During human echocardiography studies, a sonographer can ask patients to reposition or modify their decubital position or take deep breaths to better image the apex during inspiration. It is not possible to do this in immobilized animals. For this reason, mid-papillary level images are often used for experimental studies. 4.13 Conclusions and Future Directions The results of this project provide support to the general hypothesis that HO-1 upregulation via intraperitoneal administration of hemin in rats confers a cardioprotective effect after I/R injury in the short term, as evidenced through histological and echocardiographic examination of hemin-treated animals compared with buffer-treated and sham-operated animals. The cardioprotective effect was accompanied by the induction of HO-1 and decreased NFκB activation; however this study provides no direct evidence that the by-products of heme catabolism are responsible for the acute cardioprotective effects of hemin. In the long term, chronic treatment with hemin did not prevent post-infarction LV remodeling, though it was mildly attenuated. These findings are limited by the small sample size. Future research should focus on optimizing HO-1 upregulation and its effect on left ventricular remodeling months after I/R injury once 104 remodeling is complete. The development of an ideal strategy for long term HO-1 upregulation (i.e. long term hemin dose and schedule or direct cardiac injection with a hypoxia-sensitive HO-1 plasmid) will help establish whether or not HO-1 manipulation yields therapeutic value in the setting of reperfusion injury, LV remodeling, and congestive heart failure. 105 References Ambrosio G, Flaherty JT, Duilio C, Tritto I, Santoro G, Elia PP, Condorelli M and Chiariello M (1991). Oxygen radicals generated at reflow induce peroxidation of membrane lipids in reperfused hearts. J Clin Invest. 87, 2056-2066. American Heart Association (2007). Heart disease and stroke statistics- 2007 update. Dallas, Texas: American Heart Association. Amsterdam EA, Stahl GL, Pan HL, Rendig SV, Fletcher MP and Longhurst JC (1995). Limitation of reperfusion injury by a monoclonal antibody to C5a during myocardial infarction in pigs. Am J Physiol Heart Circ Physiol 268, H448-H457. Anderson MT, Staal FJT, Gitler C, Herzenberg LA and Herzenber LA (1994). Separation of oxidant-initiated and redox-regulated steps in the NF-κB signal transduction pathway. Proc Natl Acad Sci USA 91, 11527-11531. Antunes F and Cadenas E (2000). Estimation of H2O2 gradients across biomembranes. FEBS Lett. 475, 121-126. Anversa P, Loud AV, Levicky V and Guideri G (1985). Left ventricular failure induced by myocardial infarction. Am J Physiol Heart Circ Physiol. 248, H876-H882. Araujo JA, Meng L, Tward AD, Hancock WW, Zhai Y, Lee A, Ishikawa K, Iyer S, Buelow R, Busutill RW, Shih DM, Lusis AJ and Kupiec-Weglinski JW (2003). Systemic rather than local heme oxygenase-1 overexpression improves cardiac allograft outcomes in a new transgenic mouse. Journ Immuno. 171, 1572-1580. Arnstall MA, Sawyer DB, Fukazawa R and Kelly RA (1999). Cytokine-mediated apoptosis in cardiac myocytes: the role of inducible nitric oxide synthase induction and peroxynitrite generation. Circ Res. 85, 829-840. Bagchi D, Wetscher GJ, Bagchi M, Hinder PR, Perdikis G, Stohs SJ, Hinder RA and Das DK (1997). Interrelationship between cellular calcium homeostasis and free radical generation in myocardial reperfusion injury. Chemico-Biological Interactions 104, 6585. Balla G, Jacob HS, Balla J, Rosenberg M, Nath K, Apple F, Eaton JW and Vercellotti GM (1992). Ferritin: a cytoprotective antioxidant stratagem of endothelium. J Biol Chem. 267, 18148-18153. Balla G, Vercellotti GM, Muller-Eberhard U, Eaton J and Jacob HS (1991). Exposure of endothelial cells to free heme potentiates damage mediated by granulocytes and toxic oxygen species. Lab Invest. 64, 648-655. 106 Becker LB, vanden Hoek TL, Shao Z, Li C and Schumacker PT (1999). Generation of superoxide in cardiomyocytes during ischemia before reperfusion. Am J Physiol Heart Circ Physiol. 277, H2240-H2246. Blankesteijn WM, Creemers E, Lutgens E, Cleutjens JPM, Daemen MJAP and Smits JFM (2001). Dynamics of wound healing following myocardial infarction: observations in geneticall altered mice. Acta Physiol Scand. 173, 75-82. Bond JM, Herman B, Lemasters JJ (1991). Protection by acidotic pH against anoxia/reoxygenation injury to rat neonatal cardiac myocytes. Biochem Biophys Res Commun. 179, 798-803 Brown M, McGuinness M, Wright T, Ren X, Wang Y, Boivin GP, Hahn H, Feldman AM and Jones WK (2005). Cardiac-specific blockade of NF-κB in cardiac pathophysiology: differences between acute and chronic stimuli in vivo. Am J Physiol Heart Circ Physiol. 289, H466-H476. Buerke M, Weyrich AS and Lefer AM (1994). Isolated cardiac myocytes are sensitized by hypoxia-reoxygenation to neutrophil related mediators. Am J Physiol. 266, H128-136. Buja LM (2005). Myocardial ischemia and reperfusion injury. Cardiovascular Path 14, 170-175. Bulteau AL, Lundberg KC, Humphries KM, Sadek HA, Szweda PA, Friguet B and Szweda LI (2001). Oxidative metabolism and inactivation of the proteasome during coronary artery occlusion/reperfusion. J Biol Chem. 276, 30057-30063. Calo LA, Davis PA, Piccoli A and Pessina AC (2006). A role for heme oxygenase-1 in the antioxidant and antiapoptotic effects of erythropoietin: the start of a good news/bad news story? Nephr Physiol. 103, 107-111. Camussi G, Tetta C, Bussolino F and Baglioni C (1988). Synthesis and release of platelet activating factor or α1-antichymotrypsin and is stimulated by proteinases. J Exp Med. 168, 1293-1306. Camussi G, Tetta C, Bussolino F and Baglioni C (1989). Tumour necrosis factor stimulates human neutrophils to release leukotriene B4 and platelet activating factor. Eur J Biochem. 182, 661-666. Chambers DE, Parks DA, Patterson G, Roy R, McCord JM, Yoshida S, Parmley LF and Downey JM (1985). Xanthine oxidase as a source of free radical damage in myocardial ischemia. J Mol Cell Cardiol. 17, 145-152. Chandrasekar B and Freeman GL (1997) Induction of nuclear factor κB and activation protein-1 in postischemic myocardium. FEBS Lett. 401, 30-34. 107 Chandrasekar B, Smith J and Freeman G (2001). Ischemia-reperfusionof rat myocardium activates nuclear factor-κB and induces neutrophil infiltration via lipopolysaccharideinduced CXC chemokine. Circulation 103, 2296-2302. Chensue SW, Remick DG, Shmyr-Forsch C, Beals TF and Kunkel SL (1988). Immunohistochemical demonstration of cytoplasmic and membrane-associated tumour necrosis factor in murine macrophages. Am J Path. 133, 564-572. Clark JE, Foresti R, Sarathchandra P, Kaur H, Green CJ and Motterlini R (2000). Heme oxygenase-1-derived bilirubin ameliorates postischemic myocardial dysfunction. Am J Physiol Heart Circ Physiol. 278, H643-H651. Cleutjens JPM, Kandala JC, Guarda E, Guntaka RV and Weber KT (1995). Regulation of collagen degradation in the rat myocardium after infarction. J Mol Cell Cardiol. 27, 1281-1292. Cleutjens JPM, Blankesteijn WM, Daemen MJAP and Smits JFM (1999). The infracted myocardium: simply dead tissue or a lively target for therapeutic interventions. Cardiovasc Res. 44, 232-241. Coatney RW (2001). Ultrasound imaging: principles and applications in rodent research. ILAR Journal 42, 233-247. Csonka C, Varga E, Kovacs P, Ferdinandy P, Blasig IE, Szilvassy Z and Tosaki A (1999). Heme oxygenase and cardiac function in ischemic/reperfused rat hearts. Free Rad Biol Med. 27, 119-126. Deague JA, Wilson CM, Grigg LE and Harrap SB (1999). Discrepancies between echocardiographic measurements of left ventricular mass in a healthy population. Clin Sci. 97, 377-383. Del Monte F, Williams E, Lebeche D, Schmidt U, Rosenzweig A, Gwathmey JK, Lewandowski ED and Hajjar RJ (2001). Improvement in survival and cardiac metabolism after gene transfer sarcoplasmic reticulum Ca2+-ATPase in a rat model of heart failure. Circulation 104, 1424-1429. Depre C, Vanoverschelde JJ and Taegtmeyer H (1999). Circulation 99, 578-588. Glucose for the heart. Desmouliere A, Geinoz A, Gabbiani F and Gabbiani G (1993). Transforming growth factor-b1 induces a-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J Cell Biol. 122, 103-111. Deten A, Volz HC, Briest W and Zimer H (2002). Cardiac cytokine expression is upregulated in the acute phase after myocardial infarction: experimental studies in rats. Cardiovascular Res. 55, 329-340. 108 Dewald O, Zymek P, Winkelmann K, Koerting A, Ren G, Abou-Kamis T, Michael LH, Rollins BJ, Entman ML and Frangogiannis NG (2005). CCL2/monocyte chemoattractant protein-1 regulates inflammatory responses critical to healing myocardial infarcts. Circ Res. 96, 881-889. Di Napoli P, Taccardi A, De Caterina R and Barsotti A (2002). Pathophysiology of ischemia-reperfusion injury: experimental data. Ital Heart J. 3 (Suppl 4), 24S-28S. Dobaczewski M, Bujak M, Zymek P, Ren G, Entman ML and Frangogiannis NG (2006). Extracellular matrix remodeling in canine and mouse myocardial infarcts. Cell Tissue Res. 324, 475-488. Dreyer WJ, Smith CW, Michael LH, Rossen RD Hughes BJ, Entman ML and Anderson DC (1989). Canine neutrophil activation by cardiac lymph obtained during reperfusion of ischemic myocardium. Circ Res. 65, 1751-1762. Dreyer WJ, Michael LH, West MS, Smith CW, Rothlein R, Rossen RD, Anderson DC and Entman ML (1991). Neutrophil accumulation in ischemic canine myocardium. Insights into time course, distribution, and mechanism of localization during early reperfusion. Circulation 84, 400-411. Duilio C, Ambrosio G, Kuppumasmy P, Dipaula A, Becker LC and Zweier JL (2001). Neutrophils are primary source of O2 radicals during reperfusion after prolonged myocardial ischemia. Am J Physiol Heart Circ Physiol 280, H2649-H2657. Entman ML, Michael L, Rossen RD, Dreyer WJ, Anderson DC, Taylor AA and Smith CW (1991). Inflammation in the course of early myocardial ischemia. FASEB J. 5, 25292537. Fan H, Sun B, Gu Q, Lafond-Walker A, Cao S and Becker LC (2002). Oxygen radicals trigger activation of NF-κB and upregulation of ICAM-1 in reperfused canine heart. Am J Physiol Heart Circ Physiol. 282, H1778-H1786. Fantone JC and Ward PA (1982). Role of oxygen-derived free radicals and metabolites in leukocyte-dependent inflammatory reactions. Am J Path 107, 397-418 Foppa M, Duncan BB and Rohde LEP (2005). Echocardiography-based left ventricular mass estimation. How should we define hypertophy? Cardiovascular Ultrasound 3, 113. Frangogiannis NG, Lindsey ML, Michael LH, Youker KA, Bressler RB, Mendoza LH, Spengler RN, Smith CW and Entman ML (1998). Resident cardiac mast cells degranulate and release preformed TNFα, initiating the cytokine cascade in experimental canine myocardial ischemia/reperfusion. Circulation 98, 699-710. 109 Frangogiannis NG, Smith CW and Entman ML (2002). The inflammatory response in myocardial infarction. Cardiovasc Res. 53, 31-47. Frangogiannis NG, Mendoza LH, Lewallen M, Michael LH, Smith WC and Entman ML (2001). Induction and suppression of interferon-inducible protein (IP)-10 in reperfused myocardial infarcts may regulate angiogenesis. FASEB J doi:10.1096/fj.00-0745fje. Frangogiannis NG and Entman ML (2005). Chemokines in myocardial ischemia. Trends Cardiovasc Med. 15, 163-169. Critical role of endogenous thrombospondin-1 in preventing expansion of healing myocardial infarcts. Circulation 111, 2935-2042 Ganote CE (1983). Contraction band necrosis and irreversible myocardial injury. J Mol Cell Cardiol. 15, 67-73. Garcia-Dorado D, Inserte J, Ruiz-Meana M, Gonzalez MA, Solares J, Julia M, Barrabes JA and Soler-Soler J (1997). Gap junction uncoupler heptanol prevents cell-to-cell progression of hypecontracture and limits necrosis during myocardial reperfusion. Circulation 96, 3579-3586. Gaudron P, Eilles C, Kugler I and Ertl G (1993). Progressive left ventricular dysfunction and remodeling after myocardial infarction. Potential mechanisms and early predictors. Circulation 87, 755-763. Gharaee-Kermani M, Denholm EM and Phan SH (1996). Costimulation of fibroblast collagen and transforming growth factor-β1 gene expression by monocyte chemoattractant protein-1 via specific receptors. Journ Biol Chem. 271, 17779- 17784. Goldhaber JI and Weiss JN (1992). Oxygen free radicals and cardiac reperfusion abnormalities. Hypertension 20, 118-127. Grinwald PM and Brosnahan C (1987). Sodium imbalance as a cause of calcium overload in post-hypoxic reoxygenation injury. J Mol Cell Cardiol. 19, 487-495. Guo Y, Stein AB, Wu W, Tan W, Zhu X, Li Q, Dawn B, Motterlini R and Bolli R (2004). Administration of a CO-releasing molecule at the time of reperfusion reduces infarct size in vivo. Am J Physiol Heart Circ Physiol. 286, H1649-H1653. Gwechenberger M, Mendoza LH, Youker KA, Frangogiannis NG, Smith CW, Michael LH and Entman ML (1999). Cardiac myocytes produce interleukin-6 in culture and in viable border zone of reperfused infarctions. Circulation 99, 546-551. Halestrap AP, Kerr PM, Javadov S and Woodfield KY (1998). Elucidating the molecular mechanism of the permeability transition pore and its role in reperfusion injury of the heart. Biochim Biophys Acta 1366, 79-94. 110 Hangaishi M, Ishizaka N, Aizawa T, Kurihara Y, Taguchi J, Nagai R, Kimura S and Ohno M (2000). Induction of heme oxygenase-1 can act protectively against cardiac ischemia/reperfusion in vivo. Biochem Biophys Res Commun. 279, 582-588. Hansen PR (1995). Role of neutrophils in myocardial ischemia and reperfusion. Circulation 91, 1872-1885. Haramaki N, Stewart DB, Aggarwal S, Ikeda H, Reznick AZ and Packer L (1998). Networking antioxidants in the isolated rat heart are selectively depleted by ischemiareperfusion. Free Rad Biol Med. 25, 329-339. Hendrickson SC, St Louis JD, Lowe JE and Abdel-Aleem S (1997). Free fatty acid metabolism during myocardial ischemia and reperfusion. Mol Cell Biochem. 166, 85-94. Henkel T, Machleidt T, Alkalay I, Kronke M, Ben-Neriah Y and Baeuerle PA (1993). Rapid proteolysis of IκBα is necessary for activation of transcription factor NF-κB. Nature 365, 182-185. Herskowitz A, Choi S, Ansari AA and Wesselingh S (1995). Cytokine mRNA expression in post-ischemic/reperfused myocardium. Am J Path. 146, 419-428 Hoffman JW Jr, Gilbert TB, Poston RS, Silldorff EP (2004). Myocardial reperfusion injury: etiology, mechanisms and therapies. J Extra Corpor Technol. 36, 391-411. Hool LC (2006). Reactive oxygen species in cardiac signaling: from mitochondria to plasma membrane ion channels. Clin Exp Pharm Physiol. 33, 146-151. Hutchins GM and Bulkley BH (1978). Infarct expansion versus extension: two different complications of acute myocardial infarction. Am J Cardiol. 41, 1127-1132 Ide T, Tsutsui H, Hayashidani S, Kang D, Suematsu N, Nakamura K, Utsumi H, Hamasaki N and Takeshita A (2001). Mitochondrial DHA damage and dysfunction associated with oxidative stress in failing hearts after myocardial infarction. Circ Res. 88, 529-535. Immenschuh S, Hinke V, Ohlmann A, Gifhorn-Katz S, Katz N, Jungermann K and Kietzmann T (1998). Transcriptional activation of the heme oxygenase-1 gene by cGMP via a cAMP response element/activator protein-1 element in primary cultures of rat hepatocytes. Biochem J. 334, 141-146. Irwin M, Mak S, Mann D, Qu R, Dawood F, Wen WH, Penninger J and Liu P (1999). Tissue expression and immunolocalization of tumour necrosis factor-alpha in post infarction-dysfunctional myocardium. Circulation 99, 1492-1498. Jeney V, Balla J, Yachie A, Varga Z, Vercellotti GM, Eaton JW and Balla G (2002). Pro-oxidant and cytotoxic effects of circulating heme. Blood 100, 879- 887. 111 Jennings RB and Reimer KA (1991). The cell biology of acute myocardial ischemia. Annu Rev Med. 42, 225-246. Jennings RB, Steenbergen C, Kinney RB, Hill ML and Reimer KA (1983). Comparison of the effect of ischemia and anoxia on the sarcolemma of the dog heart. Eur Heart J. 4 (Suppl H), 123-137. Jennings RB, Murry CE, Steenbergen C Jr, Reimer KA (1990). Development of cell injury in sustained ischemia. Circulation 82 (Suppl 3), II2-II12. Jeremy RW, Koretsune Y, Marban E and Becker LC (1992). Relation between glycolysis and calcium homeostasis in postischemic myocardium. Circ. Res. 70, 11801190. Jordan JE, Zhao ZQ and Vinten-Johansen J (1999). The role of neutrophils in myocardial ischemia-reperfusion injury. Cardiovasc Res. 43, 860-878. Kaminski KA, Bonda TA, Korecki J and Musial WJ (2002). Oxidative stress and neutrophil activation- the two keystones of ischemia/reperfusion injury. Intern J Cardiol. 86, 41-59. Kelley ST, Malekan R, Gorman JH, Jackson BM, Gorman RC, Suzuki Y, Plappert T, Bogen DK, St John Sutton MG and Edmunds LH (1999). Restraining infarct expansion preserves left ventricular geometry and function after acute anteroapical infarction. Circulation 99, 135-142. Kharbanda RK, Peters M, Walton B, Kattenhorn M, Mullen M, Klein N, Vallance P, Deanfield J and MacAllister R (2001). Ischemic preconditioning prevents endothelial injury and systemic neutrophil activation during ischemia-reperfusion in humans in vivo. Circulation 103, 1624-1630. Kirkby KA and Adin CA (2006). Products of heme oxygenase and their potential therapeutic applications. Am J Physiol Renal Physiol. 290, 563-571. Kingsley PB, Sako EY, Yang MQ, Zimmer SD, Ugurbil K, Foker JE and From AHL (1991). Ischemic contracture begins when anaerobic glycolysis stops: a 31P-NMR study of isolated rat hearts. Am J Physiol Heart Circ Physiol. 261(30), H469-H478. Kuehl EA, Humes JL, and Ham EA (1980). Inflammation: the role of peroxidase-derived products. Adv Prostaglandin Thromboxane Res. 6, 77-86. Kukielka GL, Smith CW, Manning AM, Youker KA, Michael LH, and Entman ML (1995). Induction of interleukin-6 synthesis in the myocardium. Potential role in postreperfusion inflammatory injury. Circulation 92, 1866-1875. 112 Ladilov YV, Siegmund B and Piper HM (1997). Simulated ischemia increases the susceptibility of rat cardiomyocytes to hypercontracture. Circ Res. 80, 69-75. Lalu MM, Wang W and Schulz R (2002). Peroxynitrite in myocardial ischemiareperfusion injury. Heart Fail Rev. 7, 359-369. Lang RM, Bierig M, Devereux RB, Flachskampf FA, Foster E, Pellikka PA, Picard MH, Roman MJ, Seward J, Shanewise JS, Solomon SD, Spencer KT, St John Sutton M and Stewart WJ (2005). Recommendations for chamber quantification: a report from the american society of echocardiography’s guidelines and standards committee and the chamber quantification writing group, developed in conjunction with the european association of echocardiography, a branch of the european society of cardiology. J Am Soc Echocardiogr. 18, 1440-1463. Lavrovsky Y, Schwartzman ML, Levere RD, Kappas A and Abraham NG (1994). Identification of binding sites for transcription factors NF-κB and AP-2 in the promoter region of the human heme oxygenase 1 gene. Proc Natl Acad Sci USA. 91, 5987-5991. Lefer AM, Tsao P, Aoki N and Palladino MJ (1990). Mediation of cardioprotection by TGFβ. Science 249, 61-64. Li C, Browder W and Kao RL (1999). Early activation of transcription factor NF-κB during ischemia in perfused rat heart. Am J Physiol Heart Circ Physiol 276, H543-H552. Li C, Kao RL, Ha T, Kelley J, Browder IW and Williams DL (2001). Early activation of IKKβ during in vivo myocardial ischemia. Am J Physiol Heart Circ Physiol 280, H1264H1271. Litt MR, Jeremy RW, Weisman HF, Winkelstein JA and Becker LC (1989). Neutrophil depletion limited to reperfusion reduces myocardial infarct size after 90 minutes of ischemia. Evidence for neutrophil-mediated reperfusion injury. Circulation 80, 18161827. Liu B, el Alaoui-Talibi Z, Clanachan AS, Schulz R and Lopaschuk GD (1996). Uncoupling of contractile function from mitochondrial TCA cycle activity and MVO2 during reperfusion of ischemic hearts. Am J Physiol Heart Circ Physiol. 270, 72-80. Liu P, Hock C, Nagele R, Wong P (1997). Formation of nitric oxide, superoxide, and peroxynitrite in myocardial ischemia-reperfusion in rats. Am J Physiol. 272 (Heart Circ Physiol 41), H2327-H2336. Liu X, Pachori AS, Ward CA, Davis JP, Gnecchi M, Kong D, Zhang L, Murduck J, Yet S, Perrella MA, Pratt RE, Dzau VJ and Melo LG (2006). Heme oxygenase-1 (HO-1) inhibits postmyocardial infarct remodeling and restores ventricular function. FASEB J. 20, 207-216. 113 Lopaschuk GD and Stanley WC (1997). Glucose metabolism in the ischemic heart. Circulation 96, 313-315. Lu Li, Zhang JQ, Ramires FJ and Sun Y (2004). Molecular and cellular events at the site of myocardial infarction: from the perspective of rebuilding myocardial tissue. Biochem Biophys Res Commun. 320, 907-913. Maines MD (1992). Heme-oxygenase: clinical applications and functions. Boca Raton, FL: CRC Press. Maines MD (1997). The heme oxygenase system: a regulator of second messenger gases. Annu Rev Pharmacol Toxicol. 37, 517-554. Marx SO, Reiken S, Hisamatsu Y, Jayaraman T, Burkhoff D and Rosemblit N (2000). PKA phosphorylation dissociates FKBP12.6 from the calcium release channel (ryanodine receptor): defective regulation in failing hearts. Cell 101, 365-376. Maulik N, Shamra HS and Das DK (1996). Induction of the haem oxygenase gene expression during the reperfusion of ischemic rat myocardium. J Mol Cell Cardiol. 28, 1261-1270. McKay RG, Pfeffer MA, Pasternak RC, Markis JE, Come PC, Nakao S, Alderman JD, Ferguson JJ, Safian RD and Grossman W (1986). Left ventricular remodeling after myocardial infarction: a corollary to infarct expansion. Circulation 74, 693-702. McDonald PP, Bald A and Cassatella MA (1997). Activation of the NF-κB pathway by inflammatory stimuli in human neutrophils. Blood 89, 3421-3433. Melo LG, Agrawal R, Zhang L, Rezvani M, Mangi AA, Ehsan A, Griese DP, Dell’Acqua G, Mann MJ, Oyama J, Yet S, Layne MD, Perrella MA and Dzau V (2002). Gene therapy strategy for long-term myocardial protection using adeno-associated virusmediated delivery of heme oxygenase gene. Circulation 105, 602-607. Mercurio F and Manning AM (1999). response. Oncogene 18, 6163-6171. NF-κB as a primary regulator of the stress Mercurio F, Zhu H, Murray BW, Shevchenko A, Bennett BL, wu Li J, Young DB, Barbosa M, Mann M, Manning A and Rao A (1997). IKK-1 and IKK-2: cytokineactivated IkappaB kinases essential for NFκB activation. Science 278, 860-866. Meyers ML, Bolli R, Lekich RF, Hartley CJ and Roberts R (1985). Enhancement of recovery of myocardial function by oxygen free-radical scavengers after reversible regional ischemia. Circulation 72, 915-921. Minamisawa S, Hoshijima M, Chu G, Ward CA, Frank K, Gu Y, Martone ME, Wang Y, Ross J, Kranias EG, Giles WR and Chien KR (1999). Chronic phospholamban– 114 sarcoplasmic reticulum calcium ATPase interaction is the critical calcium cycling defect in dilated cardiomyopathy. Cell 99, 313-322. Moises VA, Ferreira RL, Nozawa E, Kanashiro RM, Campos O, de Andrade JL, Carvalho ACC and Tucci PJF (2000). Structural and functional characteristics of rat hearts with and without myocardial infarct. Initial experience with Doppler echocardiography. Arq Bras Cardiol. 75, 131-136. Moon C, Krawczyk M, Ahn D, Ahmet I, Paik D, Lakatta EG and Talan MI (2003). Erythropoietin reduces myocardial infarction and left ventricular functional decline after coronary artery ligation in rats. Proc Natl Acad Sci USA 100,11612-11617. Morgan JP, Erny RE, Allen PD, Grossman W and Gwathmey JK (1990). Abnormal intracellular calcium handling, a major cause of systolic and diastolic dysfunction in ventricular myocardium from patients with heart failure. Circulation 81 (Suppl 2), III2132. Murohara T, Guo J and Lefer AM (1995). Cardioprotection by a novel recombinant serine protease inhibitor in myocardial ischemia and reperfusion injury. J Pharm Exp Ther. 274, 1246-1253. Neumann F, Ott I, Gawaz M, Richardt G, Holzapfel H, Jochum M and Schomig A (1995). Cardiac release of cytokines and inflammatory responses in acute myocardial infarction. Cicrulation 92, 748-755. Nian M, Lee P, Khaper N and Liu P (2004). Inflammatory cytokines and postmyocardial infarction remodeling. Circ Res. 94, 1543-1553. Nordlie MA, Wold LE, Simkhovich BZ, Sesti C and Kloner RA (2006). Molecular aspects of ischemic heart disease: ischemia/reperfusion-induced genetic changes and potential applications of gene and RNA interference therapy. J Cardiovasc Pharmacol Therapeut. II(I), 17-30. Nossuli TO, Lakshminarayanan V, Baumgarten G, Taffet GE, Ballantyne CM, Michael LH and Entman ML (2000). A chronic mouse model of myocardial ischemiareperfusion: essential in cytokine studies. Am J Physiol Heart Circ Physiol 278, H1049H1055. Nozawa E, Kanashiro RM, Murad N, Carvalho ACC, Cravo SLD, Campos O, Tucci TJF and Moises VA (2006). Performance of two-dimensional Doppler echocardiography for the assessment of infarct size and left ventricular function in rats. Braz J Med Biol Res. 39, 687-695. O’Connor C, Hathaway WR, Bates ER, Leimberger J, Sigmon KN, Kereiakes DJ, George BS, Samaha JK, Abbottsmith JW, Candela RJ, Topol EJ and Califf RM (1997). Clinical characteristics and long-term outcome of patients in whom congestive heart 115 failure develops after thrombolytic therapy for acute myocardial infarction: development of a predictive model. Am Heart J. 133, 663-673. Ono K, Matsumori A, Shioi T, Furukawa Y and Sasayama S (1998). Cytokine gene expression after myocardial infarction in rat hearts: possible implication in left ventricular remodeling. Circulation 98, 149-156. Olivetti G, Capasso JM, Sonnenblick EH and Anversa P (1990). Side-to-side slippage of myocytes participates in ventricular wall remodeling acutely after myocardial infarction in rats. Circ Res. 67, 23-34. Owen CA and Campbell EJ (1999). The cell biology of leukocyte-mediated proteolysis. J Leukoc Biol. 65, 137-150. Pachori AS, Melo LG, Zhang L, Solomon SD and Dzau VJ (2006). Chronic recurrent myocardial ischemic injury is significantly attenuated by pre-emptive adeno-associated virus heme oxygenase-1 gene delivery. J Am Coll Cardiol. 47, 635-643. Patella V, Marino I, Lamparter B, Arbustini E, Adt M and Marone G (1995). Human heart mast cells: isolation, purification, ultrastructure and immunologic characterization. Journal of Immunology 154, 2855-2865. Pfeffer MA, Pfeffer JM, Fishbein MC, Fletcher PJ, Spadaro J, Kloner RA and Braunwald E (1979). Myocardial infarct size and ventricular function in rats. Circ Res. 44, 503-512. Pfeffer JM, Pfeffer MA, Mirsky I, Steinberg CR and Braunwald E (1982). Progressive ventricular dilatation and diastolic wall stress in rats with myocardial infarction and failure. Circulation 66 (Suppl), II-66. Pfeffer MA (1995). Left ventricular remodeling after acute myocardial infarction. Annu Rev Med. 46, 455-466. Pfeffer MA and Braunwald E (1990). Ventricular remodeling after myocardial infarction. Experimental observations and clinical implications. Circulation 81, 11611172. Piper HM, Garcia-Dorado D and Ovize M (1998). A fresh look at reperfusion injury. Cardiovasc Res. 38, 291-300. Piper HM, Meuter K and Schafer C (2003). Cellular mechanisms of ischemiareperfusion injury. Ann Thorac Surg. 75, S644-S648. Pirolo JS, Hutchins GM and Moore GW (1986). Infarct expansion: pathologic analysis of 204 patients with a single myocardial infarct. J Am Coll Cardiol. 7, 349-354. 116 Platt JL and Nath KA (1998). Heme oxygenase: protective gene or Trojan horse? Nat Med. 4, 1364-1365. Powell DW, Mifflin RC, Valentich JD, Crowe SE, Saada JI and West AB (1999). Myofibroblasts: paracrine cells important in health and disease. Am J Physiol. 277, C1C9. Pruefer D, Buerke U, Khalil M, Dahm M, Darius H, Oelert H and Buerke M (2002). Cardioprotective effects of the serine protease inhibitor aprotinin after regional ischemia and reperfusion on the beating heart. J Thorac Cardiovasc Surg. 124, 942-949. Rampart M and Wiliams TJ (1988). Evidence that neutrophil accumulation induced by interleukin-1 requires both local protein biosynthesis and neutrophil CD18 antigen expression in vivo. Br J Pharmacol. 94, 1143-1148. Reimer KA, Jennings RB and Tatum AH (1983). Pathobiology of acute myocardial ischemia: metabolic, functional and ultrastructural studies. Am J Cardiol. 52, 71A-81A. Rossen RD, Michael LH, Hawkins HK, Youker K, Dreyer WJ, Baughn RE and Entman ML (1994). Cardiolipin-protein comnplexes and initiation of complement activation after coronary artery occlusion. Circ. Res. 75, 546-555. Rossi A, Elia G and Santoro MG (1997). Inhibition of nuclear factor-κB by prostaglandin A1: an effect associated with heat shock factor transcription activation. Proc Natl Acad Sci USA 94, 746-750. Ruiz-Meana M, Garcia-Dorado D, Hofstaetter B, Piper HM and Soler-Soler J (1999). Propagation of cardiomyocyte hypercontracture by passage of Na+ through gap junctions. Circ Res. 85, 280-287. Ryter SW, Alam J, Choi AMK (2006). Heme oxygenase-1/carbon monoxide: from basic science to therapeutic applications. Physiol Rev. 86, 583-650. Sabbah HN, Sharov VG, Lesch M and Goldstein S (1995). Progression of heart failure: a role for interstitial fibrosis. Mol Cell Biochem. 147, 29-34. Schafer C, Ladilov YV, Siegmund B, Piper HM (2000). Importance of bicarbonate transport for protection of cardiomyoctes against reoxygenation injury. Am J Physiol Heart Circ Physiol. 278, H1457-H1463 Schiller NB, Shah PM, Crawford M, DeMaria A, Devereux R, Feigenbaum H, Gutgesell H, Reichek N, Sahn D, Schnittger I, Silverman NH and Tajik AJ (1989). Recommendations for quantitation of the left ventricle by two-dimensional echocardiography. J Am Soc Echocardiogr. 2, 358-367. 117 Serini G and Gabbiani D (1999). Mechanisms of myofibroblast activity and phenotypic modulation. Exp Cell Res. 250, 273-283. Sharma HS and Das DK (1997). Role of cytokines in myocardial ischemia and reperfusion. Mediators of Inflammation 6, 175-183. Smith CW, Entman ML, Lane CL, Beaudet AL, Ty TI, Youker K, Hawkins HK and Anderson DC (1991). Adherence of neutrophils to canine cardiac myocytes in vitro is dependent on intercellular adhesion molecule-1. J Clin Invest. 88, 1216-1223. Stanton LW, Garrard LJ, Damm D, Garrick BL, Lam A, Kapoun AM, Zheng Q, Protter AA, Schreiner GF and White RT (2000). Circ Res. 86, 939-945. Steenbergen C, Murphy E, Levy L and London RE (1987). Elevation in cytosolic free calcium concentration early in myocardial ischemia in perfused rat heart. Circ Res. 60, 700-707. St John Sutton MG and Sharpe N (2000). Left ventricular remodeling after myocardial infarction: pathophysiology and therapy. Circulation. 101, 2981-2988. Sun M, Dawood F, Wen W, Chen M, Dixon I, Kirshenbaum LA and Liu PP (2004). Excessive tumor necrosis factor activation after infarction contributes to susceptibility of myocardial rupture and left ventricular dysfunction. Circulation 110, 3221-3228. Sun Y, Zhang JQ, Zhang J and Lamparter S (2000). Cardiac remodeling by fibrous tissue after infarction in rats. J Lab Clin Med. 135, 316-323. TangYL, Tang Y, Zhang YC, Qian K, Shen L and Phillips MI (2004). Protection from ischemic heart injury by a heme oxygenase-1 plasmid system. Hypertension 43, 746751. Terry CM, Clikeman JA, Hoidal JR and Callahan KS (1999). TNFα and IL-1α induce heme oxygenase-1 via protein kinase C, Ca2+, and phospholipase A2 in endothelial cells. Am J Physiol Heart Circ Physiol. 276, H1493-H1501. Tomasek J, Gabbiani G, Hinz B, Chaponnier C and Brown RA (2002). Myofibroblasts and mechno-regulation of connective tissue remodeling. Nat Rev Mol Cell Biol. 3, 349363. Tsutsui H (2006). Mitochondrial oxidative stress and heart failure. Int Med. 45, 809-813 VanDamme J, Opdenakker G, Simpson RJ, Rubira MR, Cayphas S, Vink A, Billiau A and VanSnick J (1987). Identification of the human 26-kD protein, interferon beta 2 (IFN-beta 2), as a B cell hybridoma/plasmacytoma growth factor induced by interleukin 1 and tumor necrosis factor. J Exp Med. 165, 914-919 118 Vanden Hoek TL, Shao Z, Li C, Zak R, Schumacker PT and Becker LC (1996). Reperfusion injury in cardiac myocytes after simulated ischemia. Am J Physiol Heart Circ Physiol. 270, H1334-H1341. Vanoverschelde JL, Janier MF and Bergmann SR (1994). The relative importance of myocardial energy metabolism compared with ischemic contracture in the determination of ischemic injury in isolated perfused rabbit hearts. Circ Res. 74, 817-828. Vercellotti GM, Balla G, Balla J, Nath K, Eaton JW and Jacob HS (1994). Heme and the vasculature: an oxidative hazard that induces antioxidant defenses in the endothelium. Artif Cells Blood Substit Immobil Biotechnol. 22, 207-213. Vetterlein F, Schrader C, Volkmann R, Neckel M, Ochs M, Schmidt G and Hellige G (2003). Extent of damage in ischemic, non-reperfused, and reperfused myocardium of anaesthetized rats. Am J Physiol Heart Circ Physiol. 285, 755-765. Virag JI and Murry CE (2003). Myofibroblast and endothelial cell proliferation during murine myocardial infarct repair. Am J Path. 163, 2433-2440. Vulapalli SR, Chen Z, Chua BHL, Wang T and Liang C (2002). Cardioselective overexpression of HO-1 prevents I/R-induced cardiac dysfunction and apoptosis. Am J Physiol Heart Circ Physiol. 283, H688-H694. Wagener F, Eggert A, Boerman OC, Oyen WJG, Verhofstad A, Abraham NG, Adema G, van Kooyk Y, de Witte T and Figdor CG (2001). Heme is a potent inducer of inflammation in mice and is counteracted by heme oxygenase. Blood 98, 1802-1811. Wagener F, van Beurden HE, von den Hoff JW, Adema GJ and Figdor CG (2003). The heme-heme oxygenase system: a molecular switch in wound healing. Blood 102, 521528. Wagener F, Volk H, Willis D, Abraham NG, Soares MP, Adema GJ and Figdor CG (2003). Different faces of the heme-heme oxygenase system in inflammation. Pharm Rev. 55, 551-571. Weber KT and Brilla CG (1991). Pathological hypertrophy and cardiac interstitium. Fibrosis and renin-angiotensin-aldosterone system. Circulation 83, 1849-1865. Weber TP, Meissner A, Boknik P, Hartlage MG, Mollhoff T, Van Aken H and Rolf N (2001). Hemin, inducer of heme oxygenase-1, improves functional recovery from myocardial stunning in conscious dogs. J Cardiothorac Vasc Anaes. 15, 422-427. Weisman HF, Bush DE, Mannisi JA and Bulkley BH (1985). Global cardiac remodeling after acute myocardial infarction: a study in the rat model. J Am Coll Cardiol. 5, 13551362. 119 Weisman HF, Bush DE, Mannisi JA, Weisfeldt ML and Healy B (1988). Cellular mechanisms of myocardial infarct expansion. Circulation 78, 186-201. Weiss SJ (1989). Tissue destruction by neutrophils. N Eng J Med. 320, 365-376. Wu WC, Rathore SS, Wang Y, Radford MJ and Krumholz HM (2001). Blood transfusions in elderly patients with acute myocardial infarction. N Engl J Med. 345, 1230-1236. Xu KY, Zweier JL and Becker LC (1997). Hydroxyl radical inhibits sarcoplasmic reticulum Ca2+/ATPase function by direct attack on the ATP binding site. Circ Res. 80, 76-81. Xu M, Wang Y, Hirai K, Ayub A and Ashraf M (2001). Calcium preconditioning inhibits mitochondrial permeability transition and apoptosis. Am J Physiol Heart Circ Physiol. 280, H899-H908. Yang Z, Zingarelli B and Szabo C (2000). Crucial role of endogenous interleukin-10 production in myocardial ischemia/reperfusion injury. Circulation 101, 1019-1026. Zahger D, Yano J, Chaux A, Fishbein MC and Ganz W (1995). Absence of lethal reperfusion injury after 3 hours of reperfusion. A study in a single-canine-heart model of ischemia-reperfusion. Circulation 91, 2989-2994. Zhao Z, Nakamura M, Wang N, Velez DA, Hewan-Lowe KO, Guyton RA and VintenJohansen J (2000). Dynamin progression of contractile and endothelial dysfunction and infarct extension in the late phase of reperfusion. J Surg Res. 94, 133-144. Zimmerman BJ and Granger DN (1990). Reperfusion-induced leukocyte infiltration: role of elastase. Am J Physiol. 259, H390-H394. Zorov DB, Filburn CR, Klotz L, Zweier JL and Sollott SJ (2000). Reactive oxygen species (ROS)- induced ROS release: a new phenomenon accompanying induction of the mitochondrial permeability transition in cardiac myocytes. J Exp Med. 192, 1001-1014. Zweier JL, Flaherty JT and Weisfeldt ML (1987). Direct measurement of free radical generation following reperfusion of ischemic myocardium. Proc. Natl. Acad. Sci. 84, 1404-1407. Zweier JL, Kuppusamy J and Lutty GA (1988). Measurement of endothelial cell free radical generation: evidence for a central mechanism of free radical injury in postischemic tissues. Proc Natl Acad Sci USA 85, 4046-4050. 120